产生具有新型颜色的种子的茄子的制作方法
本发明涉及产生具有新型种子颜色的种子的新型茄子植物(茄子(solanummelongenal.))。本发明还涉及这种植物的种子和后代,以及用于得到这种植物的繁殖材料。此外,本发明涉及植物、来源于这种植物的种子和繁殖材料作为种质在育种项目中的用途。
发明背景
茄子植物(eggplant或aubergine)(茄子(solanummelongenal.))是茄科(茄科(solanaceae))的植物,并且属于茄属(solanum)。其与番茄和马铃薯紧密相关。茄子结出具有相同名称的果实,所述果实通常用于烹饪。所述植物的不同品种产生不同尺寸、形状和颜色的果实。茄子是一种重要的食物农作物,在保护栽培和在旷野中在所有条件和气候下种植遍布全世界。
茄子果实因其令人愉悦的苦味和松软的口感而受欢迎。尽管果皮光滑并且颜色上可能经由各程度的紫色色素沉着从白色、黄色、绿色至几乎黑色,果肉是白色的或乳白色的并且口感松软。包含在果肉中的是以圆锥形式排列的种子。尽管处在种子发育早期的茄子种子具有白色或乳白色,但是在可商售的果实阶段(收获茄子果实售卖给人消费的阶段),茄子种子具有棕色的颜色。在该阶段,茄子果实中的种子尚未完全成熟。仅当茄子果实处在消费者认为过熟的阶段时,种子达到成熟阶段,在该阶段,它们的棕色更深。茄子种子的棕色是由种子最外层,即含有棕色色素的种皮(也称为外种皮(testa))导致的。
在多种物种中,种皮包含深色(棕色到黑色)色素。在拟南芥(arabidopsisthaliana)中已对种皮着色进行了充分的研究。在拟南芥种子中,在种子发育晚期阶段观察到种皮色素沉着。然而,称为原花色素(pa)或缩合丹宁的色素的实际合成始于胚发育的早期阶段(在受精后1-2天)。这些黄酮类开始作为无色化合物在内皮(外皮的最内细胞层)的液泡中积聚,在种子干燥过程中被氧化,并且使成熟种子呈现棕色。已知有一些拟南芥种皮色素沉积突变体。在这些称为透明外种皮(transparenttesta)的突变体中,拟南芥种皮呈现白色至淡黄色的颜色。多种transparenttesta基因编码黄酮类生物合成途径中的酶,而另一些编码参与该途径的一些点的调节酶。黄酮类途径中的大部分基因是单拷贝基因。在多种透明外种皮突变体中,植物的其他部分(例如,花和果实)的颜色受到影响,原因在于除了在种皮中具有被破坏的原花色素积聚之外,它们在其他植物部分或整个植物中的黄酮醇和/或花色素苷生成也被破坏。
由于处在可商售的果实阶段的茄子果实的棕色种子在白色或乳白色的果肉中突出,并且使得茄子果实看上去对消费者吸引性较小,因此使果实产生颜色较浅的种子是优选的性状。尤其是如果该较浅的种子颜色独立于果皮的颜色时。
技术实现要素:
因此,本发明的一个目的是提供一种产生具有新颜色的种子、特别是具有较浅颜色的种子的新型茄子植物。
在产生本发明的研究中,培育了一种新型茄子植物,发现所述茄子植物产生具有新颜色的种子,特别是具有本文进一步定义的较浅颜色的种子。发现具有这种较浅颜色的种子的产生与茄子基因组内一种qtl的纯合存在相关,所述qtl如同存在于由代表性样品以保藏号42054保藏在ncimb的种子生长的植物中一样,并且可从由代表性样品以保藏号42054保藏在ncimb的种子生长的植物得到。发现在由保藏的种子生长的植物的基因组中,标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15(完整的序列数据提供在表1中)各自与引起本发明的新型种子颜色的qtl连锁。尽管这些标志物中的任一种或这些标志物的任意组合可以用于鉴定引起本发明新型种子颜色的性状的qtl,由于标志物seqidno:11和seqidno:13在统计检验中具有最高的lod得分,因此这两个标志物是优选的。
通过使用标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15的组合,标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11和seqidno:13的组合,标志物seqidno:1、seqidno:3、seqidno:5和seqidno:7的组合,标志物seqidno:9、seqidno:11、seqidno:13和seqidno:15的组合,标志物seqidno:9、seqidno:11和seqidno:13的组合,或标志物seqidno:11和seqidno:13的组合,可以确定引起本发明的新型种子颜色性状的qtl在植物基因组中的存在情况。
因此,qtl存在于所保藏的材料的基因组中,并且因此该材料是能够用于使所述新型种子颜色性状基因渐渗到其他茄子植物中的qtl的来源。此类植物可以作为起点用于培育其他具有该新型种子颜色的品种。任何包含本发明的qtl的茄子植物都是本发明的植物,这与所述qtl的来源无关。
“基因渐渗”用在本文中旨在意指性状的引入,藉由杂交和在所述性状变为可见的或可以选择qtl的一代中选择,使引起所述性状的qtl基因渐渗到不携带所述性状的植物中而引入。所述基因渐渗由多个标志物诸如片段内标志物(in-fragmentmarker)的可用性而促进。
在保藏的种子中,qtl与分子标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15中的每一种连锁。这些标志物还可以与杂交中用作亲本的任一个或两个茄子植物中包含的qtl连锁,从而将新型种子颜色性状转移到其他植物,但是所提及的标志物中至少一种的存在不是必要的,只要存在引起所述性状的qtl即可。由于所述qtl是编码该新型种子颜色性状的遗传信息,因此该新型种子颜色表型的存在是本发明的qtl存在的直接指示。当作为该表型基础的qtl存在于植物中、而标志物不再存在于植物中时,本发明具有如本文所述的新型种子颜色性状的植物仍然是本发明的植物。标志物有时是性状的遗传原因,但不总是性状的遗传原因。标志物可以位于引起所述性状的基因中或者与引起所述性状的基因遗传连锁。它们经常作为工具用于跟踪性状的遗传。在育种过程中,在保藏的种子中与遗传决定子连锁的分子标志物由此可以用于辅助新型种子颜色性状转移到其他植物。熟练的育种者将理解该新型种子颜色性状转移到茄子植物中可以通过目测检查、分光光度法或图像分析、或通过监测和育种以确定本文所述的分子标志物的存在(即,标志物辅助的选择)、或二者进行监测。所述标志物定位在特定的基因组区域进一步允许相连的序列在育种中的应用以及开发另外连锁的遗传标志物。因此,强调的是,可以使用遗传绘图在所鉴定的区域内的任意分子标志物来实施本发明,条件是所述标志物在杂交的亲本之间是多态性的,从而将新型种子颜色性状转移到其他的植物。
本发明由此提供包含qtl的茄子植物(solanummelongenal.),所述qtl在纯合存在时使所述茄子植物产生具有本文定义的新型颜色的种子,其中所述qtl与由保藏号为ncimb42507的种子长成的植物的基因组中存在的qtl相同或相似,并且在其中与选自下组的至少一种标志物连锁:seqidno:1,seqidno:3,seqidno:5,seqidno:7,seqidno:9,seqidno:11,seqidno:13和seqidno:15,并且其中所述qtl优选纯合存在。所述qtl是如同存在于由代表性样品以保藏号ncimb42507保藏在ncimb的种子长成的茄子植物的基因组中一样,并且可从由代表性样品以保藏号ncimb42507保藏在ncimb的种子长成的茄子植物获得。
在该情形中,“茄子植物”旨在包括茄子(solanummelongenal.)。
本发明的性状为隐性性状,所述性状为在所产生的种子的种皮中棕色色素的减少或完全不存在,导致具有较浅颜色的种子。本发明的qtl由此必须以纯合状态存在于本发明的茄子植物中,从而使得本发明的性状在所述植物产生的种子中显现。
种皮是种子的外保护层,并且来源于胚珠的珠被。种皮由此是二倍体母体起源的。种子的颜色因此由产生所述种子的植物(杂交中接受花粉的母本植物)的基因型决定。
本发明还涉及可通过将第一茄子植物与第二茄子植物杂交得到的本发明的茄子植物,其中所述植物中至少一种包含如其代表性种子以保藏号ncimb42507保藏在ncimb的茄子植物或其携带所述qtl的后代植物中所包含的qtl,并且通过目测检查,优选在f2中目测检测本发明的性状,和/或选择本文所述的分子标志物的存在。
通过将不携带本发明的qtl的野生型植物与关于本发明的qtl纯合或杂合的植物杂交,并且选择所需要的表型,优选在杂交的f2中选择所需要的表型,和/或针对本文所述的分子标志物的存在进行选择,优选在杂交的f1和/或f2中选择,可以将本发明的性状引入到野生型茄子植物中。f2通过f1自交得到。
可以引入本发明的性状的茄子植物可以是任何类型、任何果实形式或果实颜色的茄子植物。
由纯合地携带本发明的qtl的植物产生的种子的新型种子颜色定义为所述种子与由不是纯合地包含所述qtl的植物产生的种子的颜色相比在成熟的种子阶段具有的较浅颜色。所述种子的种皮具有减少量的棕色色素,所述棕色色素存在于不是纯合地包含所述qtl的植物产生的成熟种子阶段的种子的种皮中。特别地,所述种子的种皮具有如此低量的通常存在于野生型种子的种皮中的棕色色素,使得所述本发明的植物的种子的种皮中的棕色颜色不能肉眼检测到。更特别地,所述种皮完全缺少通常存在于野生型种子的种皮中的棕色色素。
本发明纯合携带遗传决定子的植物由此可以由其种子的新型颜色鉴定。由本发明纯合携带本发明的qtl的植物产生的种子和杂合携带或完全不携带所述遗传决定子的同基因植物的种子之间的颜色差异容易由熟练的茄子种植者观察到。当比较由纯合携带本发明的qtl的本发明植物产生的种子和杂合携带或完全不携带所述遗传决定子的同基因植物的种子的颜色时,所有种子必须处在相同的发育阶段。可以在可商售的果实阶段,或优选地在成熟种子阶段,评估种子的颜色差异。尽管由纯合携带本发明的qtl的本发明植物产生的种子和杂合携带或完全不携带所述遗传决定子的同基因植物的种子之间的颜色差异在可商售的果实阶段已经是清楚的,所述颜色差异在成熟种子阶段最清楚。
术语“成熟种子”对熟练的茄子种植者是清楚的,并且特别意指完全生理发育的种子的种子阶段。成熟种子处在完全生理发育的茄子果实中存在的种子阶段。所述完全生理发育的茄子果实超过了可商售的果实阶段——收获茄子果实售卖给人消费的阶段——并且将被消费者认为是过熟的。熟练的茄子种植者知晓茄子果实何时处在可商售的果实阶段以及茄子果实何时完全生理发育。
本发明纯合携带所述遗传决定子的植物的成熟种子的颜色比杂合携带或完全不携带所述遗传决定子的同基因植物的成熟种子的颜色浅得多。与杂合携带或完全不携带所述遗传决定子的同基因植物相比,纯合携带所述遗传决定子的植物的成熟种子较浅的颜色在紧接着果实收获后的新鲜种子中最明显可见。
术语“新鲜种子”也是技术人员所清楚的,并且特别意指紧接着收获后在茄子果实中存在的种子或新近从果实中采集且仍然湿的种子。换言之,新鲜种子是那些还没干的种子。
尽管不携带本发明的qtl的新鲜成熟的野生型茄子种子的颜色可以是所谓的中等棕色、沙棕色、古铜色或铜色,本发明纯合地携带所述遗传决定子的植物的新鲜成熟种子的颜色可以表示为灰白色、乳白色、米黄色、非常浅的黄色或浅灰黄色。
当种子变干后,野生型种子和本发明的植物的种子的颜色都改变。然而,同样当完全变干时,本发明的植物的种子的颜色明显可与干燥的野生型种子的颜色区分开并且比干燥的野生型种子的颜色浅。不携带本发明的qtl的干燥野生型茄子种子的颜色可能被称为棕色、沙棕色、中等棕色、古铜色或铜色,而纯合地携带所述遗传决定子的本发明的植物的干燥成熟种子的颜色可以表示为米黄色、浅黄色、淡黄色、米色或土黄色。当比较种子颜色时,明显的是这必须在相同阶段进行,即新鲜种子与新鲜种子比较,干燥种子与干燥种子比较,以及处在成熟种子阶段的种子与处在成熟种子阶段的种子比较,和处在可商售的果实阶段的种子与处在可商售的果实阶段的种子比较。
本发明的新型种子颜色由此可以定义为由本发明的植物产生的新鲜种子与由不是纯合地包含所述qtl的植物产生的种子相比在成熟种子阶段具有的更浅的颜色,和/或定义为由所述植物产生的干燥种子与由不是纯合地包含所述qtl的植物产生的种子相比在成熟种子阶段具有的更浅的颜色。
本发明的种子的新型种子颜色表型是由种皮(也称为外种皮,其透出在种皮下子叶的颜色)中棕色色素的减少或不存在导致。
本发明的种子和野生型种子的着色也可以使用rhs比色表(theroyalhorticulturalsociety,london,uk)确定。不携带本发明的qtl的新鲜成熟野生型种子的颜色在其最深的部分类似于颜色165b,并且在其最浅的部分类似于165c,而本发明的新鲜成熟种子的颜色在其最浅部分类似于颜色161d,并且在其最深部分类似于161c(表2)。干燥成熟野生型种子的颜色类似于颜色162a,而本发明的干燥成熟种子的颜色类似于颜色164b(表2)。
虽然植物育种者和种植者常用rhs比色表来确定植物颜色,但是明显的是,也可以使用其他比色表或系统来确定颜色。例如,也可以使用munsell比色系统以rgb色码指定颜色,或者可以使用比色计或图像分析确定颜色。技术人员知晓怎样使用这些不同的颜色系统以及不同颜色系统之间的色码转换。关于这些rhs颜色的rgb、cielab和cielch值在下表中列出。
广泛用于测量(例如使用比色计或图像分析)颜色的色度是cielab色度。所述色度包括3个数据变量:l*,a*和b*。l*表示0-100度的亮度,其中0为黑色,100为白色。变量a*和b*表示红、绿、蓝和黄色的量:a*值表示从绿(负值)到红(正值)的颜色变化,而b*表示从黄(正值)到蓝(负值)的颜色变化。两种样品之间颜色的差异可以以l*和/或a*、和/或b*的变化表示。
如技术人员所清楚的,颜色参数依用来定量颜色的方法而变化。仍然地,当比较野生型新鲜成熟种子与本发明的新鲜成熟种子的具体颜色定量方法(使用rhs比色表的颜色估测、比色法或图像分析)的l*(10°/d65)评分时,与不携带本发明的qtl的野生型种子相比,本发明的种子的l*评分始终更高。此外,当比较野生型干燥成熟种子与本发明的干燥成熟种子的具体颜色定量方法(使用rhs比色表的颜色估测、比色法或图像分析)的l*(10°/d65)评分时,与不携带本发明的qtl的野生型种子相比,本发明的种子的l*评分始终更高。更高的l*评分表示更浅的颜色。
在一个实施方案中,本发明涉及纯合包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用rhs比色表并将rhs色码转换成相对应的cielab色度对所述植物的干燥成熟种子确定的l*(10°/d65)评分至少为47,50,54,58,59,65,70,80,90(按照优选递增的顺序)。使用rhs比色表对所述植物的干燥成熟种子确定的l*(10°/d65)评分适当地不高于99。
在一个实施方案中,本发明涉及纯合地包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用rhs比色表并将rhs色码转换成相对应的cielab色度对所述植物的新鲜成熟种子确定的l*(10°/d65)评分至少为49,52,57,63,67,70,80,90(按照优选递增的顺序)。使用rhs比色表对所述植物的新鲜成熟种子确定的l*(10°/d65)评分适当地不高于99。
在一个实施方案中,本发明涉及纯合地包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用比色计对所述植物的干燥成熟种子确定的l*(10°/d65)评分至少为61.5,62,63,65,70,80,90(按照优选递增的顺序)。使用比色计对所述植物的干燥成熟种子确定的l*(10°/d65)评分适当地不高于99。
在一个实施方案中,本发明涉及纯合包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用比色计对所述植物的新鲜成熟种子确定的l*(10°/d65)评分至少为85,87,89,90(按照优选递增的顺序)。使用比色计对所述植物的新鲜成熟种子确定的l*(10°/d65)评分适当地不高于99。
在一个实施方案中,本发明涉及纯合包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用图像分析(例如,如实施例3所述)对所述植物的干燥成熟种子确定的l*(10°/d65)评分至少为55,58,60,62,65,70,80,90(按照优选递增的顺序)。使用比色计对所述植物的干燥成熟种子确定的l*(10°/d65)评分适当地不高于99。
在一个实施方案中,本发明涉及纯合包含本发明的qtl的茄子(茄子(solanummelongena))植物,其中使用图像分析(例如,如实施例3所述)对所述植物的新鲜成熟种子确定的l*(10°/d65)评分至少为62,64,66,68,70,80,90(按照优选递增的顺序)。使用比色计对所述植物的新鲜成熟种子确定的l*(10°/d65)评分适当地不高于99。
因此,当用于本文时,术语“新型种子颜色”旨在指灰白色、乳白色、米白色、非常浅的黄色或浅灰黄色和/或在其最浅部分显示出rhs色码的颜色161d和在其最深部分显示出rhs色码的161c的新鲜成熟种子的种子颜色,以及米黄色、浅黄色、淡黄色、米色或土黄色和/或显示出rhs色码的颜色164b的干燥成熟种子的种子颜色。
本发明还涉及本发明的茄子植物的种子和适于有性繁殖的植物其他部分。所述植物部分可以选自:小孢子、花粉、子房、胚珠、胚囊和卵细胞。
另外,本发明还涉及本发明的茄子植物适于营养繁殖的部分,例如组织培养物、切下部分、根、茎、细胞和原生质体。组织培养物可由叶、花粉、胚、子叶、下胚轴、分生细胞、根、花药、花、种子或茎长成。
本发明还涉及包含qtl的本发明茄子植物的种子,所述qtl在纯合存在时生成产生具有新型颜色的种子的茄子植物。在这样的种子中,所述qtl可以纯合或杂合存在,但是优选纯合存在。
本发明还涉及能够长成本发明的茄子植物的种子,其中所述胡椒属种子的基因组包含qtl,当所述qtl纯合存在于由所述种子长成的植物中时,所述qtl引起新型种子颜色的性状。在所述种子中,所述qtl可以纯合或杂合存在,但是优选纯合存在。
本发明还涉及杂交种子和产生杂交种子的方法,所述方法包括将第一亲本植物与第二亲本植物杂交,并且收获所得到的杂交种子。所述杂交种子可以杂合地或纯合地包含本发明的qtl。为了使所有的杂交种子都纯合携带本发明的性状以使由所述杂交种子长成的植物产生具有新型颜色的种子,两个亲本都需要对于引起新型种子颜色的性状的qtl是纯合的。由此,两个亲本都携带本发明的qtl。对于其他性状,它们不需要是一致的。
除了茄子植物的种子之外,本发明还涵盖源自包含qtl的茄子植物的后代,所述qtl在纯合存在时生成产生具有新型颜色的种子的茄子植物。所述后代可以由本发明的植物或其后代植物的有性繁殖或营养繁殖而产生。所述后代携带如在本发明的植物中存在的和如同在由代表性样品以保藏号ncimb42507保藏于ncimb的种子长成的植物中存在一样的qtl。所述qtl可以纯合和杂合地存在于后代中,优选以第一选择(即,纯合)存在。除此之外,植物可以在除种子颜色以外的一个或多个特征方面进行修饰。此类另外的修饰例如通过杂交和选择、诱变或通过用转基因转化而实现。
当用在本文中时,词语“后代”旨在意指来自任意茄子植物与本发明携带本发明的qtl(作为新型种子颜色性状的基础)的植物杂交的后代或第一代和所有后续的子代。本发明的后代包括任意与本发明携带引起本发明的新型种子颜色性状的qtl的植物的杂交的子代。此类后代例如可通过将第一茄子植物与第二茄子植物杂交而得到,其中所述植物之一由代表性样品以保藏号ncimb42507保藏的种子长成,此类后代也可以是携带如同存在于ncimb42507中一样的本发明qtl的任意其他茄子植物的后代。
此外,本发明还涵盖本发明的茄子植物的后代或由源自本发明的植物的种子长成的茄子植物的后代,其中所述植物的后代包含所述qtl,并且所述qtl可以以杂合状态存在,但是所述qtl优选以纯合状态存在。
源自本发明的茄子植物或本发明的茄子植物的茄子种子的繁殖材料也包括在本发明内,其中所述繁殖材料包含所述qtl,所述qtl在纯合存在时引起新型种子颜色,并且所述qtl优选以纯合状态存在。
本发明还涉及能够长成本发明的茄子植物的繁殖材料,其中所述繁殖材料包含所述qtl,所述qtl在纯合存在时引起该新型种子颜色,并且所述qtl优选以纯合状态存在。
例如,所述源自本发明的茄子植物的繁殖材料以及能够长成本发明的植物的繁殖材料选自:愈伤组织,小孢子,花粉,子房,胚珠,胚,胚囊,卵细胞,切下部分,根,茎,细胞,原生质体,叶,子叶,胚轴,分生细胞,根,根尖,小孢子,花药,花,种子和茎,或其部分或组织培养物。
本发明还涉及本发明的茄子植物的细胞,所述细胞包含在纯合状态下赋予所述新型种子颜色性状的qtl,其中所述qtl如同存在于由代表性样品以保藏号ncimb42507保藏在ncimb的种子长成的茄子植物的基因组中一样。所述茄子植物可通过将茄子植物与第二茄子植物、特别是由以保藏号ncimb42507保藏的种子长成的茄子植物杂交并选择(优选在f2中)具有本发明的新型种子颜色性状的茄子植物而得到。因此,所述细胞包含的遗传信息与由代表性样品以保藏号42507保藏于ncimb的种子长成的茄子植物中编码所述新型种子颜色性状的遗传信息、更具体本文所述的qtl基本相同、优选完全相同的遗传信息。优选地,本发明的细胞是植物的一部分或植物部分的一部分,但是所述细胞也可以以分离的形式存在。
在一个实施方案中,本发明涉及ncimb保藏号为ncimb42507的种子用于将赋予本发明的新型种子颜色性状的本发明qtl转移到另一株茄子植物中的用途。本发明还涉及具有与保藏的种子相同的qtl的茄子植物用于将本发明的qtl转移到另一株茄子植物中的用途。
在另一个实施方案中,本发明涉及携带本发明的qtl的茄子植物作为农作物的用途。
本发明还涉及携带本发明的qtl的茄子植物作为种子来源的用途。
在另一个实施方案中,本发明涉及携带本发明的qtl的茄子植物作为繁殖材料的来源的用途。
此外,本发明涉及携带本发明的qtl的茄子植物用于消费的用途。
在另一个实施方案中,本发明涉及携带本发明的qtl的茄子植物用于将所述qtl给予茄子植物的用途。
在另一个实施方案中,本发明涉及茄子植物、特别是由ncimb保藏号为ncimb42507的种子长成的茄子植物作为本发明的qtl的接受体的用途。
本发明还涉及由产生具有新型种子颜色的种子的果实并且包含本文定义的qtl的本发明茄子植物收获的茄子果实或其部分。自然地,这也涉及由所述茄子果实制成的任何食品或加工食品。
本发明还涉及由包含杂合状态的本文定义的qtl的本发明茄子植物收获的茄子果实或其部分。自然地,这也涉及由所述茄子果实制成的任何食品或加工食品。尽管新型种子颜色性状在此类植物中并不显现,但是它们仍然是qtl的来源,并且例如可以用于育种。
加工的茄子果实还可以包含在另一种食品中,诸如酱料、茄子酱(babaghanoush)、咖哩粉、馅饼、茄盒(moussaka)、汤或干燥的或新鲜的面团产品,如馄饨、饺子、粗通心粉(cannelloni)等。此类食品通常将被预先包装,目的在于在超市销售。由此,本发明还涉及由本发明的茄子植物收获的茄子果实或其部分在制备食品(特别是酱料、沙拉、馅饼、汤和面团)中的用途。此外,以加工的形式,所述茄子果实仍然包含本发明的qtl。
本发明的茄子植物还可以用作育种项目中的种质用于培育包含引起新型种子颜色的qtl的其他茄子植物。本发明也涵盖这种用途。
此外,本发明涉及核酸或其部分,所述核酸或其部分任选地以分离形式存在,所述核酸或其部分在以纯合状态存在时在本文定义的茄子植物中引起新型种子颜色,基于hirakawa等,draftgenomesequenceofeggplant(solanummelongenal.):therepresentativesolanumspeciesindigenoustotheoldworld,2014,dnaresearch21,649-660中提供的基因组序列,所述核酸源自连锁群11,并且在其上与选自下组的至少一种分子标志物连锁:seqidno:1,seqidno:3,seqidno:5,seqidno:7,seqidno:9,seqidno:11,seqidno:13和seqidno:15和/或存在于ncimb保藏号为ncimb42507的种子中。特别地,所述核酸与下述连锁:标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11和seqidno:13的组合,更特别地,标志物seqidno:9、seqidno:11和seqidno:13的组合,最特别的是标志物seqidno:11和seqidno:13的组合或标志物seqidno:1、seqidno:3、seqidno:5和seqidno:7的组合。本领域技术人员将能够从本发明的茄子植物的基因组分离引起本发明的种子颜色性状的核酸或其部分,并且利用其产生与所述qtl和本发明的性状连锁的新分子标志物。
本发明还涉及分子标志物在鉴定本发明的茄子植物中的qtl或培育具有新型种子颜色的茄子植物中的用途,其中所述标志物选自:seqidno:1,seqidno:3,seqidno:5,seqidno:7,seqidno:9,seqidno:11,seqidno:13和seqidno:15。
本发明进一步涉及所述分子标志物用于鉴定或培育与引起该新型种子颜色的qtl连锁的其他标志物的用途。
为了确定本发明的qtl在种子或植物的基因组中的存在,需要至少一种分子标志物,也可以使用seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15所述的分子标志物的任意组合。使用选自下组的至少一组分子标志物:seqidno:1加上seqidno:2,seqidno:3加上seqidno:4,seqidno:5加上seqidno:6,seqidno:7加上seqidno:8,seqidno:9加上seqidno:10,seqidno:11加上seqidno:12,seqidno:13加上seqidno:14和seqidno:15加上seqidno:16,可以对在本发明的种子颜色性状方面不一致的植物的群体或集合进行基因分型。
本发明的新型种子颜色qtl在植物中的纯合存在也可以通过检查由所述植物产生的成熟种子的颜色而从表型上确定。可以对任意植物或植物群体从表型上确定本发明的新型种子颜色性状的存在。在基因渐渗过程中,从表型上确定本发明的新型种子颜色性状的存在在其中所述性状在基因渐渗过程中分离(优选在f2中)的任意一代中信息性最强。
在一方面,本发明涉及用于产生具有生成带有新型颜色的种子的性状的茄子植物的方法,所述方法包括:
a)将包含qtl的植物与另一种植物杂交,其中所述qtl在纯合存在时产生新型种子颜色;
b)在f1和/或f2代中选择具有该新型种子颜色的植物;
c)任选地进行另一轮或另外多轮的自交和/或杂交和/或与优选的亲本的回交,并且随后选择包含/显示出本发明的性状的植物。
选择具有生成带有新型颜色的种子的性状的植物可以在分子水平上使用与本文所述的性状连锁的分子标志物在f1或任意其他代中进行和/或在表型上在所述性状分离的f2或任意其他代中进行。
在本申请的情形中,词语“性状”是指植物表型。特别地,词语“性状”是指本发明的性状,更特别地是指产生具有新型颜色的种子的茄子植物的性状。术语“qtl”(即,“数量性状遗传位点”)用于指植物基因组中以纯合状态存在时引起本发明的新型种子颜色性状的遗传信息。当植物的基因组包含引起本发明的性状的qtl时,由所述植物产生的种子具有本发明的新型种子颜色。因此,所述植物具有本发明的qtl。
清楚的是,提供本发明的性状的亲本不必是直接由保藏的种子长成的植物。亲本也可以是来自保藏的种子的后代植物(例如,通过自交或杂交得到)或来自通过其他方式鉴定为具有本发明的性状的种子的后代植物。
本发明另外提供在具有产生带有新型颜色的种子的性状的茄子植物中引入另一种需要的性状的方法,所述方法包括:
a)将具有该新型种子颜色性状的茄子植物(其代表性的种子以保藏号ncimb42507保藏)与包含需要的性状的第二茄子植物杂交,以产生f1代;
b)在f2代中选择包含所述新型种子颜色性状和所述需要的性状的植物;
c)将所选的f2代植物与任一亲本杂交,以产生回交后代;
d)选择包含所述需要的性状和所述新型种子颜色性状的回交后代;和
e)任选地连续重复步骤c)和d)一次或多次,从而产生包含所述需要的性状和所述新型种子颜色性状的选择的第四代或更高代的回交后代。本发明包括通过该方法产生的茄子植物。
在一个实施方案中,具有该新型种子颜色性状的植物的选择通过使用seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15.所述的标志物中任一种或任意组合在f1或任意其他代中进行。在另一方面,本发明的性状的选择起始于杂交的f2或备选地起始于回交的f2。f2中植物的选择也可以通过使用直接或间接检测作为所述性状的基础的qtl的一种或多种所述标志物从表型上进行。表型选择可以适当地通过使用rhs比色表作为参照以确定种子颜色和/或通过茄子种子的比色测量和/或图像分析确定颜色谱而进行。
在一个实施方案中,对具有所述新型种子颜色性状的植物的选择起始于f3或以后的代。
在一个实施方案中,包含所述qtl的植物是近交系、杂交种、加倍单倍体或分离群体的植物。
在一个实施方案中,本发明的植物,即包含本发明的qtl的植物,是农艺学优良的茄子植物。
在本发明的情形中,农艺学优良的茄子植物是具有这样的基因型的植物:作为人干预的定向杂交和选择的结果,所述基因型引起允许生产者收获有商业意义的产物的可区分且合乎需要的农艺性状的累积。
在育种携带本发明的qtl的新茄子植物的过程中,可以独立于本发明的qtl而在所述茄子植物中引入合乎需要的农艺性状。用于本文时,“合乎需要的性状”包括,但不限于例如提高的产量、果实形状、果实尺寸、果实颜色、种子尺寸、植物活力、植物高度以及对一种或多种疾病或引起疾病的生物体的抗性。这些合乎需要的性状中的任一个可以与本发明的qtl组合。
在另一个实施方案中,本发明的农艺优良的茄子植物是近交系或杂交种。
用于本文时,近交系植物是三轮或更多轮自交或回交产生的植物群体的植物;或者所述植物是双单倍体。例如,近交系可以是用于产生商业杂交种的亲本品系。
用于本文时,杂交种植物是具有不同基因型的两种不同植物之间杂交产生的植物。更特别地,杂交种植物是两种不同近交系植物之间杂交产生的,例如,此类杂交种植物可以是商业f1杂交种品种的植物。
在一个实施方案中,包含本发明的qtl的植物是f1杂交种品种。
本发明还提供用于产生具有新型种子颜色性状的茄子植物的方法,所述方法通过使用加倍单倍体产生技术产生具有所述性状的加倍单倍体系进行。
此外,本发明涉及可以长成具有新型种子颜色性状的植物的杂交种种子和用于产生所述杂交种种子的方法,所述方法包括将第一亲本植物与第二亲本植物杂交,并收获所产生的杂交种种子,其中所述第一亲本植物和/或所述第二亲本植物是所要求保护的植物。
在一个实施方案中,本发明涉及用于产生具有新型种子颜色性状的杂交种茄子植物的方法,所述方法包括将第一亲本茄子植物与第二亲本茄子植物杂交,并且收获所产生的具有新型种子颜色性状的第一亲本植物和第二亲本植物的杂交种种子,并且使所述杂交种种子长成具有所述新型种子颜色性状的杂交种植物。
本发明还涉及用于产生具有新型种子颜色性状的茄子植物的方法,所述方法通过使用纯合地包含本发明的qtl的种子长成所述茄子植物而进行。所述种子适当地是代表性样品以保藏号ncimb42507保藏在ncimb的种子。
本发明还涉及种子生产方法,所述方法包括从代表性样品以保藏号ncimb42507保藏于ncimb的种子长成茄子植物,允许所述植物产生种子,并且收获这些种子。这些种子的产生适当地通过杂交或自交进行。
在一个实施方案中,本发明涉及通过使用组织培养物生成具有产生带有新型颜色的种子的性状的茄子植物的方法。
此外,本发明涉及通过使用营养繁殖生成具有产生带有新型颜色的种子的性状的茄子植物的方法。
在一个实施方案中,本发明涉及用于生成具有产生带有新型颜色的种子的性状的茄子植物的方法,所述方法通过使用遗传修饰的方法使引起所述性状的qtl基因渐渗到所述茄子植物中而进行。遗传修饰包括使用来自非可杂交物种的基因或合成的基因进行的转基因修饰或转基因,以及使用来自农作物本身或来自性相容供体植物的编码(农业)性状的天然基因进行的同源转基因修饰(cisgenicmodification)或同源转基因(cisgenesis)。
在一个实施方案中,获得遗传信息、特别是所述qtl的来源由从所保藏的种子长成的植物或其有性繁殖子代或营养繁殖子代形成。
本发明还涉及用于培育具有产生新型颜色的种子的性状的茄子植物的育种方法,其中使用包含引起所述性状的qtl的种质。所述种质由生物体的全部遗传特征组成,并且按照本发明,至少包括本发明的新型种子颜色性状。包含所述qtl的植物并且代表所述种质的代表性种子以保藏号ncimb42507保藏在ncimb。
在另一个实施方案中,本发明涉及用于生成具有产生带有新型颜色的种子的性状的茄子植物的方法,其中包含赋予所述性状的qtl的植物的后代或繁殖材料用作使所述性状基因渐渗到另一株茄子植物中的来源。包含所述qtl的植物的代表性种子以保藏号ncimb42507保藏在ncimb。
此外,本发明涉及产生具有本发明的新型种子颜色性状的茄子植物的新型种子颜色基因,其中所述新型种子颜色基因如同存在于代表性样品以保藏号ncimb42507保藏于ncimb的植物的基因组中一样。熟练的育种者知晓怎样利用此类植物作为该新型种子颜色基因的来源使所述新型种子颜色基因基因渐渗到植物中。
本发明还涉及生成具有产生带有新型种子颜色的种子的性状的茄子植物的qtl用于产生具有新型种子颜色性状的植物(特别是具有该新型种子颜色性状的茄子植物)的用途,所述qtl如同存在于代表性样品以保藏号ncimb42507保藏的植物的基因组中一样。
按照其另一方面,本发明涉及产生具有新型种子颜色的种子的非天然存在的植物,所述新型种子颜色是植物基因组中存在qtl的结果,所述qtl如同存在于代表性样品以保藏号42507保藏的植物的基因组中一样。所述非天然存在的植物特别是突变植物。
术语“核酸”用于包含引起本发明的性状的遗传信息的大分子、dna或rna分子。当植物表现出本发明的表型性状时,其基因组包含引起所述性状的核酸。由此所述植物具有本发明的核酸。在本发明中,所述核酸是如同存在于代表性样品以保藏号ncimb42507保藏的植物的基因组中一样的qtl的一部分。
本发明优选提供具有新型种子颜色性状的茄子植物,所述植物可通过本文所述的方法和/或技术人员熟悉的方法中的任一种得到。
当不存在分子标志物时,或者在qtl与标志物之间已经发生重组使得所述标志物不再预测所述qtl在植物基因组中的存在的情形中,可以通过等位性试验确定遗传决定子与本发明的qtl的等价性。为了进行等位性试验,使对于本发明的已知qtl是纯合的材料,即试验植物,与对于待检测的遗传决定子是纯合的材料杂交。后一种植物称为供体植物。待检测的供体植物应该针对待检测的决定子是纯合的或应该针对待检测的决定子制成纯合的。技术人员知晓怎样得到针对待检测的决定子是纯合的植物。当在供体植物和试验植物之间杂交的f2中没有观察到与本发明的基因座相关的表型分离时,已证明了供体植物的决定子和试验植物的已知基因座是等价的。应该观察到的表型是新型种子颜色,特别是本文定义的较浅的种子颜色。试验植物可以是,但不限于,由以保藏号42054保藏于ncimb的种子长成的植物。
当多于一种基因可能负责某种性状,并且进行等位性试验来确定等价性时,进行试验的技术人员必须确保所有相关的基因纯合存在,以使试验正确进行。
保藏信息
具有生成产生带有新型种子颜色的茄子植物的qtl的茄子植物的种子于2016年1月5日保藏在ncimbltd.(fergusonbuilding,craibstoneestate,bucksburn,aberdeen,ab219ya,uk),ncimb保藏号为ncimb42507。该保藏的种子纯合包含所述qtl。由这些种子长成的植物产生具有本文定义的新型种子颜色的种子。
保藏的种子不满足获得植物品种保护所需要的dus标准,并且因此不能被视为植物品种。
标志物
标志物信息
表1
分子snp标志物
在保藏号为ncimb42507的种子的基因组中,标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15的snp序列与赋予茄子植物以产生具有新型种子颜色的种子的性状的本发明的qtl连锁。seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14和seqidno:16的序列分别表示分子snp标志物seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15的野生型茄子等位基因。
与本发明的性状连锁的标志物与野生型之间差异的核苷酸用下划线和粗体标出。
标志物等位基因seqidno:1与标志物等位基因seqidno:2的差异不在于核苷酸差异,而在于单个碱基对缺失:存在于标志物seqidno:2的位置47上的a在seqidno:1中缺失。标志物等位基因seqidno:3与标志物等位基因seqidno:4的差异在于位置101的snp,其中seqidno:3在位置101具有g而seqidno:4在位置101具有a。标志物等位基因seqidno:5与标志物等位基因seqidno:6的差异在于位置77的snp,其中seqidno:5在位置77具有t而seqidno:6在位置77具有c。标志物等位基因seqidno:7与标志物等位基因seqidno:8的差异在于位置101的snp,其中seqidno:7在位置101具有a而seqidno:8在位置101具有g。标志物等位基因seqidno:9与标志物等位基因seqidno:10的差异在于位置101的snp,其中seqidno:9在位置101具有t而seqidno:10在位置101具有g。标志物等位基因seqidno:11与标志物等位基因seqidno:12的差异在于位置101的snp,其中seqidno:11在位置101具有t而seqidno:12在位置101具有c。标志物等位基因seqidno:13与标志物等位基因seqidno:14的差异在于位置101的snp,其中seqidno:13在位置101具有c而seqidno:14在位置101具有g。标志物等位基因seqidno:15与标志物等位基因seqidno:16的差异在于位置92的snp,其中seqidno:15在位置92具有t而seqidno:16在位置92具有c。
这些标志物序列中的snp可以用作分子标志物来检测本发明的qtl的存在。这些snp是实际的标志物。当使用snp作为标志物时,围绕所述snp的序列不需要与此处提供的序列长度相同。例如,snp可以用于检测本发明的qtl在不包含本发明的qtl的茄子植物与包含本发明的qtl的茄子植物之间杂交的后代中的存在,所述包含本发明的qtl的茄子植物可以是由代表性样品以ncimb保藏号42507保藏于ncimb的种子长成的植物。
附图说明
将在下述实施例中进一步举例说明本发明,所述实施例绝不意欲限制本发明的范围。在所述实施例中,参考下述附图:
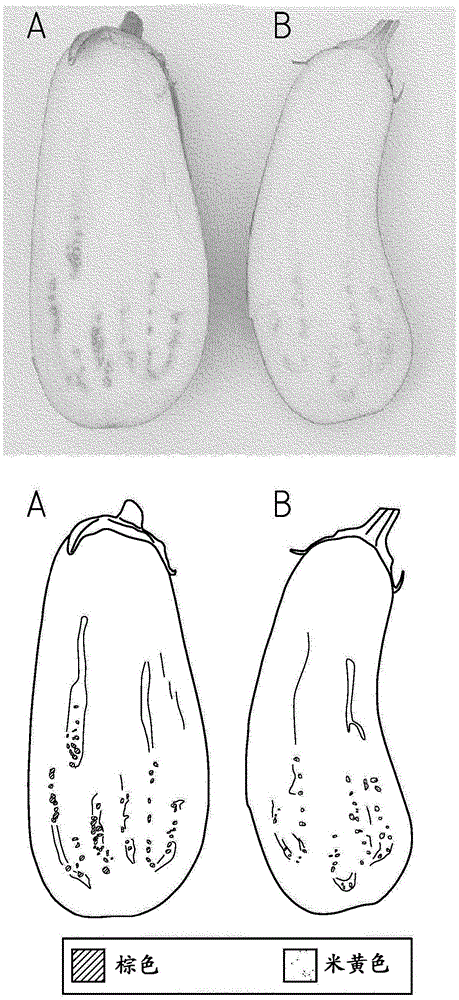
图1:处于可商售阶段的本发明的茄子果实(b)和处于可商售阶段的野生型茄子果实(a)的横切面,显示内部的种子。
图2:处于可商售阶段的本发明的茄子果实(b)和处于可商售阶段的野生型茄子果实(a)的横切面的放大图,显示内部的种子。
图3:干燥成熟的本发明的茄子种子(a)和干燥成熟的野生型茄子种子(b)。
图4:分开的野生型成熟茄子种子,显示(a)种皮,(b)完好的半个种子,和(c)半个种子的胚加胚乳。分开的成熟的本发明的茄子种子显示(a)种皮,(b)完好的半个种子,和(c)半个种子的胚加胚乳。
实施例
实施例1
本发明的植物的培育
以下述方式获得本发明的茄子植物:在用f1杂交种茄子植物起始产生的加倍单倍体茄子植物的选择中,令人惊讶地发现41株植物中有2株产生具有本发明的新型颜色的种子。这是令人惊讶的,因为用作生成该加倍单倍体群体的基础的f1杂交种植物和由对f1杂交种植物自花授粉产生的f2植物以及后几代都没有产生具有本发明新型颜色的种子。使本发明初始培育的植物cb-2与野生型植物杂交。继续几轮自交、杂交和选择产生包含纯合状态的本发明qtl的茄子的种子。
将这些种子于2016年1月5日保藏于ncimbltd(fergusonbuilding,craibstoneestate,bucksburn,aberdeenab219ya,uk),保藏号为ncimb42507。该保藏的种子包含纯合状态的所述qtl。
实施例2
本发明的植物的表型表征
纯合携带所述遗传决定子的本发明的植物可以通过其种子的新型颜色进行鉴定。
纯合携带本发明的qtl的本发明植物的成熟种子的颜色要比杂合携带或完全不携带本发明的qtl的同基因植物的种子的颜色浅得多(图1和2)。与杂合携带或完全不携带本发明的qtl的同基因植物相比,纯合携带所述遗传决定子的植物的成熟种子的较浅的颜色在紧接着果实收获后的新鲜成熟种子中最明显可见。成熟种子是存在于完全生理发育的茄子果实中的那些种子。新鲜种子是紧接着收获后在茄子果实中存在的那些种子或新近从果实中采集且仍然湿的那些种子。不携带本发明的qtl的新鲜成熟的野生型茄子种子的颜色可以被称为中等棕色、沙棕色、古铜色或铜色,而本发明纯合携带所述遗传决定子的植物的新鲜成熟种子的颜色可以表示为灰白色、乳白色、米黄色、非常浅的黄色或浅灰黄色。
当种子变干后,野生型种子和本发明的植物的种子的颜色都改变。然而,在完全变干时,本发明的植物的成熟种子的颜色也明显可与成熟的野生型种子的颜色区分开并且比成熟的野生型种子的颜色浅(图3)。不携带本发明的qtl的干燥成熟野生型茄子种子的颜色可以被称为棕色、沙棕色、中等棕色、古铜色或铜色,而纯合携带所述遗传决定子的本发明的植物的干燥成熟种子的颜色可以表示为米黄色、浅黄色、淡黄色、米色或土黄色。
当剥掉表现出新型种子颜色表型的植物的成熟种子和野生型对照种子的种皮时,可清楚看出本发明种子的新型种子颜色表型是由于种皮(也称为外种皮,其透出在种皮下子叶的颜色)中棕色色素的减少或不存在导致(图4)。
本发明的种子和野生型种子的着色可以使用rhs比色表(theroyalhorticulturalsociety,london,uk)评分。不携带本发明的qtl的新鲜成熟野生型种子的颜色在其最深的部分评分为165b,并且在其最浅的部分评分为165c,而本发明的新鲜成熟种子的颜色在其最浅部分评分为161d,并且在其最深部分评分为161c(表2)。干燥成熟野生型种子的颜色评分为162a,而本发明的干燥成熟种子的颜色评分为164b(表2)。
表2
新鲜和干燥的野生型茄子种子和纯合携带本发明qtl的本发明茄子的种子的rhs颜色估测。还将rhs色码转换成cielabl*a*b*色度值(d65照明和10度的观察者角度)。
环境条件,包括光源、物体尺寸、颜色背景和视觉或照明的角度可能影响着色的物体怎样呈现给人观察者。为了克服这种主观性,已经开发了颜色测量系统和仪器用来就描述颜色的变量定量颜色并表示颜色。除了如上文所述用rhs比色表估测种子颜色之外,还使用两种实验方法:比色法/分光光度法和图像分析。
在荷兰德利尔使用具有3mm测量孔的cm700d型konicaminolta手持比色计,利用cielabl*a*b*色彩空间、d65照明和10度观察者角度,对成熟茄子种子的颜色进行评分。在使用之前,按照供应商的使用说明校正仪器。对于比色计测量,将约10粒种子堆在白色背景上,尽可能用种子覆盖所述背景。成熟的野生型种子和本发明的种子尺寸相似。对每份样品比色计评分四次。每份样品的平均值显示在表3中。
表3
通过konicaminolta比色计cm700d,利用cielabl*a*b*色彩空间、d65照明和10度的观察者角度确定新鲜和干燥的野生型茄子种子与纯合携带本发明qtl的本发明茄子的种子的平均颜色。
使用具有带有圆形b+w偏振滤光片的nikonaf-s35mmf/1.8gdx35mm镜头的nikond7000照相机,在暗室中以标准化设置进行拍摄。标准化的照相机设置使用带有偏振滤光片的日光荧光灯(4x36瓦,5400k,cri98,40khz)。灯头与样品平台成45度角。在拍照之前,打开灯,并允许预热至少30分钟。将照相机安装在台子上,镜头朝下并位于样品平台上方。对每份样品拍摄约10粒彼此明显分开的成熟种子。
使用imagejmacro(1.48u)和x-ritecolorcheckerpassport色卡进行照片的颜色校正。使用cellprofilerpipeline进行图像分析并产生校正的rgb值。然后使用色彩校正算法将校正的rgb颜色值转换为cielabl*a*b*颜色值(d65照明和10度的观察者角度)。
表4
通过图像分析确定的新鲜和干燥的野生型茄子种子和纯合携带本发明qtl的本发明茄子的种子以校正的rgb值和校正的ciel*a*b*值(10°/d65)表示的平均颜色。
cielab色度广泛用于测量颜色并显示颜色的差异。色度包括3个数据变量:l*,a*和b*。l*表示0-100度的亮度,其中0为黑色,100为白色。变量a*和b*表示红、绿、蓝和黄色的量:a*值表示从绿(负值)到红(正值)的颜色变化,而b*表示从黄(正值)到蓝(负值)的颜色变化。两种样品之间颜色的差异可以以l*和/或a*、和/或b*的变化表示。
从表2-4中不同l*a*b*评分清楚可见,颜色参数因用于定量颜色的方法而变化。同样地,当比较野生型新鲜成熟种子与本发明的新鲜成熟种子的具体颜色定量方法(使用rhs颜色表的颜色估测、比色法或图像分析)的l*(10°/d65)评分时,与不携带本发明的qtl的野生型种子相比,本发明的种子的l*评分始终更高。比较野生型干燥成熟种子与本发明的干燥成熟种子的具体颜色定量方法(使用rhs颜色表的颜色估测、比色法或图像分析)的l*(10°/d65)评分时也是一样的,即与不携带本发明的qtl的野生型种子相比,本发明的种子的l*评分始终更高。
实施例3
对本发明性状的qtl分析
为了将本发明的性状作图并鉴定本发明的性状的qtl,使纯合携带遗传决定子的加倍单倍体(dh)系cb-2与产生正常颜色的种子并且不携带本发明的遗传决定子的dh系bb-15杂交。由该杂交产生的f1植物用于产生gh群体,所述gh群体由371个dh系组成。使这371个dh植物生长,并对它们产生的种子的颜色进行表型分型。
对来自gh群体的所有dh植物用两种基因型评分:aa用于纯合亲本cb-2,bb用于纯合亲本bb-15。滤除非多态性标志物、具有强高遗漏值以及强失真的标志物并进行标志物相校正后,在joinmap4.0软件(kyazmab.v.,wageningen)中进行剩余91个snp标志物的遗传图谱构建。首先利用最大可能性绘图方法来估测标志物在连锁群中的次序。这之后使用由最大可能性绘图得到的标志物起点顺序通过回归绘图(regressionmapping)来预测标志物在连锁群中的位置。利用haldane绘图功能将各标志物之间的重组频率转换成各标志物之间的遗传距离(单位为centimorgan,cm)。连锁群编号按照draftgenomesequenceofeggplant(solanummelongenal.)(hirakawa等,draftgenomesequenceofeggplant(solanummelongenal.):therepresentativesolanumspeciesindigenoustotheoldworld,2014,dnaresearch21,649-660)进行。利用内参图谱位置调整标志物在图谱上的方向。
对于本发明的性状构建柱状图,以观察数据分布:本发明的性状具有两个评分的相等分布:新型种子颜色和正常种子颜色。
在gh群体中进行本发明的性状的qtl绘图。使用r统计软件,利用逐步回归法进行qtl绘图。对于连锁群11检测到关于本发明的性状的一个主要的qtl,解释方差为71%。16个标志物绘制在连锁群11上,连锁群11具有63.4cm的总长度。在连锁群11上36.7cm处的标志物seqidno:13鉴定为峰值标志物。峰值标志物的lod评分为193.6,解释方差为71%,叠加效应为-0.48。上侧翼标志物为在34.4cm处的seqidno:9,下侧翼标志物为在40.8cm处的seqidno:15。另外的标志物为在34.4cm处的seqidno:7、seqidno:5、seqidno:3和seqidno:1以及在36.7cm处的标志物seqidno:11。
实施例4
将本发明的性状转移到其他茄子
使纯合携带本发明qtl的本发明茄子植物(供体植物;由保藏物ncimb42507的种子长成)与一些不同的野生型茄子植物杂交,所述野生型茄子植物不携带本发明的性状,而在类型、果实形式和果实颜色方面不同。如果本发明的茄子植物用作杂交中的母本,野生型植物用作花粉供体,则由该杂交产生的f1种子具有本发明的新型种子颜色。然而,如果选择以野生型植物作为杂交中的母本,并且本发明的植物作为花粉供体,则由该杂交产生的f1种子具有正常的种子颜色。由此,种子的颜色由产生所述种子的植物(杂交中的母本植物)的基因型决定。
不管本发明的茄子植物在与野生型植物的杂交中是用作母本还是用作花粉供体,由产生的f1种子长成的植物具有与野生型植物相同的表型,即它们产生正常颜色的种子,这表明所述性状是隐性的。然而,通过分子标志物能够在所有f1植物中检测本发明的性状以杂合状态的存在。用于这一目的的单核苷酸多态性(snp)标志物以seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13和seqidno:15显示在表1中。
在f2代中,本发明的性状以与单基因隐性遗传相对应的方式分离(表5)。
表5
在来自本发明的植物与野生型茄子植物(具有正常着色的种子)杂交的f2群体中本发明性状的分离。
χ2检验证实具有野生型亲本表型(正常种子颜色)和供体亲本表型(新型种子颜色)的f2植物观察到的数量符合性状以单基因隐性方式分离所预测的方式,即3:1(wt亲本表型:供体亲本表型)。χ2概率值高于0.05。因此,从表5的分离数据可以推出本发明的qtl作为单基因隐性性状起作用。
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!