与CD3结合的抗体的制作方法
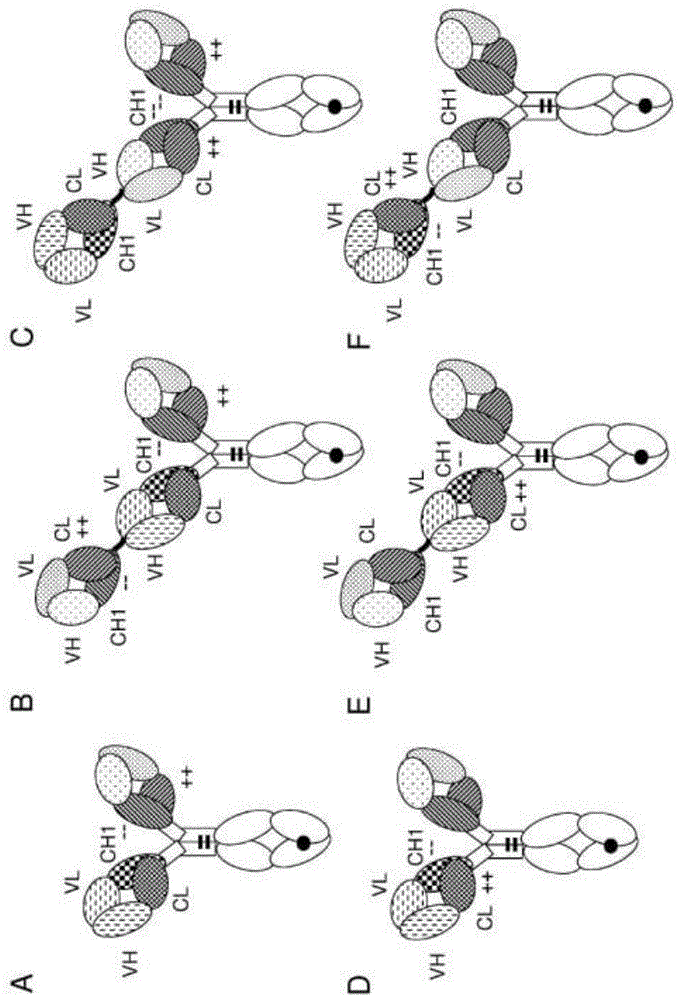
本发明总体上涉及与cd3结合的抗体,包括例如用于激活t细胞的多特异性抗体。此外,本发明涉及编码此类抗体的多核苷酸以及包含此类多核苷酸的载体和宿主细胞。本发明进一步涉及用于生产所述抗体的方法以及在疾病的治疗中使用所述抗体的方法。
背景技术:
:cd3(分化抗原簇3)是由四个亚基,cd3γ链、cd3δ链和两个cd3ε链组成的蛋白质复合物。cd3与t细胞受体和ζ链缔合,以在t淋巴细胞中生成激活信号。cd3已广泛用作药物靶点。靶向cd3的单克隆抗体已被用作自身免疫性疾病(例如i型糖尿病)或治疗移植排斥的免疫抑制剂疗法。cd3抗体莫罗单抗-cd3(okt3)是第一种获准用于临床的单克隆抗体,于1985年获得批准。cd3抗体的最新应用呈双特异性抗体的形式,其一方面与cd3结合,另一方面与肿瘤细胞抗原结合。此类抗体同时与两个靶点的结合将促使靶细胞与t细胞之间发生临时相互作用,引起任何细胞毒性t细胞激活并且随后使靶细胞裂解。出于治疗目的,抗体必须满足的一个重要要求是在体外(用于储存药物)和体内(向患者施用后)均具有足够高的稳定性。类似于天冬酰胺脱酰胺的修饰是重组抗体的典型降解方式,可同时影响体外稳定性和体内生物学功能。鉴于抗体、特别是双特异性抗体对t细胞激活具有巨大的治疗潜力,因此需要具有优化特性的cd3抗体。技术实现要素:本发明提供了与cd3结合的抗体,包括多特异性(例如双特异性)抗体,这些抗体对例如天冬酰胺脱酰胺降解具有抗性,因此特别稳定,满足治疗目的的要求。这些(多特异性)抗体进一步结合了良好的功效和可生产性以及低毒性和有利的药代动力学特性。如本文中所示,本发明提供的与cd3结合的抗体(包括多特异性抗体)在ph7.4于37℃储存2周后与cd3的结合活性相对于在ph6于-80℃储存2周后的结合活性保留超过约90%,所述结合活性通过表面等离子体共振(spr)测得。在一个方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3。在一个方面,vh包含与seqidno:7的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列,并且/或者vl包含与seqidno:11的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。在进一步方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含seqidno:7的vh序列和seqidno:11的vl序列。在一个方面,第一抗原结合结构域为fab分子。在一个方面,抗体包含由第一亚基和第二亚基组成的fc结构域。在一个方面,抗体包含与第二抗原结合的第二抗原结合结构域和任选的第三抗原结合结构域。在一个方面,其中第二抗原结合结构域和/或在存在时的第三抗原结合结构域为fab分子。在一个方面,第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1,特别是可变结构域vl和vh彼此替换。在一个方面,第二抗原结合结构域和在存在时的第三抗原结合结构域为常规的fab分子。在一个方面,第二抗原结合结构域和在存在时的第三抗原结合结构域为fab分子,其中在恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号);并且在恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号),且213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在一个方面,第一抗原结合结构域与第二抗原结合结构域彼此融合,任选地经由肽连接基融合。在一个方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,并且(i)第二抗原结合结构域在fab重链的所述c末端与第一抗原结合结构域的fab重链的所述n末端融合,或者(ii)第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合。在一个方面,第一抗原结合结构域、第二抗原结合结构域和在存在时的第三抗原结合结构域各自为fab分子,并且抗体包含由第一亚基和第二亚基组成的fc结构域;并且其中(i)第二抗原结合结构域在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合,并且第一抗原结合结构域在fab重链的c末端与fc结构域的第一亚基的所述n末端融合,或者(ii)第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合,并且第二抗原结合结构域在fab重链的c末端与fc结构域的第一亚基的n末端融合;并且第三抗原结合结构域在存在时在fab重链的c末端与fc结构域的第二亚基的n末端融合。在一个方面,fc结构域是igg,特别是igg1,fc结构域。在一个方面,fc结构域是人fc结构域。在一个方面,fc包含促进fc结构域的第一亚基和第二亚基进行缔合的修饰。在一个方面,fc结构域包含一个或多个氨基酸置换,所述一个或多个氨基酸置换减少与fc受体的结合和/或降低效应子功能。在一个方面,第二抗原是靶细胞抗原,特别是肿瘤细胞抗原。在一个方面,第二抗原是tyrp-1。在一个方面,第二抗原结合结构域和在存在时的第三抗原结合结构域包含:vh,其包含seqidno:15的hcdr1、seqidno:16的hcdr2和seqidno:17的hcdr3;以及vl,其包含seqidno:19的lcdr1、seqidno:20的lcdr2和seqidno:21的lcdr3。在一个方面,第二抗原结合结构域和在存在时的所述第三抗原结合结构域包含:vh,其包含与seqidno:18的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列;和/或vl,其包含与seqidno:22的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。在一个方面,第二抗原是egfrviii。在一个方面,第二抗原结合结构域和在存在时的第三抗原结合结构域包含:vh,其包含seqidno:85的hcdr1、seqidno:86的hcdr2和seqidno:87的hcdr3;以及vl,其包含seqidno:89的lcdr1、seqidno:90的lcdr2和seqidno:91的lcdr3。在一个方面,第二抗原结合结构域和在存在时的所述第三抗原结合结构域包含:vh,其包含与seqidno:88的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列;和/或vl,其包含与seqidno:92的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。根据本发明的进一步方面,提供了一种编码本发明的抗体的分离的多核苷酸以及包含本发明的分离的多核苷酸的宿主细胞。在另一方面,提供了一种生产与cd3结合的抗体的方法,该方法包括以下步骤:(a)在适于表达抗体的条件下培养根据本发明的宿主细胞;以及任选地(b)回收抗体。本发明还涵盖通过本发明的方法生产的与cd3结合的抗体。本发明进一步提供了一种药物组合物,该药物组合物包含本发明的抗体以及药用载体。本发明还涵盖使用本发明的抗体和药物组合物的方法。在一个方面,本发明提供了用作药物的根据本发明所述的抗体或药物组合物。在一个方面,提供了用于治疗疾病的根据本发明所述的抗体或药物组合物。在一个具体方面,疾病为癌症。还提供了根据本发明所述的抗体或药物组合物在制备药物中的用途、根据本发明所述的抗体或药物组合物在制备用于治疗疾病(特别是癌症)的药物中的用途。本发明还提供了一种治疗个体疾病的方法,该方法包括向所述个体施用有效量的根据本发明所述的抗体或药物组合物。附图说明图1.本发明的(多特异性)抗体的示例性构型。(a,d)“1+1crossmab”分子的图示。(b,e)“2+1iggcrossfab”分子的图示,其包含替代性顺序的crossfab和fab组分(“反向”)。(c,f)“2+1iggcrossfab”分子的图示。(g,k)“1+1iggcrossfab”分子的图示,其包含替代性顺序的crossfab和fab组分(“反向”)。(h,l)“1+1iggcrossfab”分子的图示。(i,m)“2+1iggcrossfab”分子的图示,其包含两个crossfab。(j,n)“2+1iggcrossfab”分子的图示,其包含两个crossfab以及替代性顺序的crossfab和fab组分(“反向”)。(o,s)“fab-crossfab”分子的图示。(p,t)“crossfab-fab”分子的图示。(q,u)“(fab)2-crossfab”分子的图示。(r,v)“crossfab-(fab)2”分子的图示。(w,y)“fab-(crossfab)2”分子的图示。(x,z)“(crossfab)2-fab”分子的图示。黑点:fc结构域中促进异源二聚化的任选修饰。++、--:任选引入ch1和cl结构域中的具有相反电荷的氨基酸。crossfab分子被描绘为包含vh区和vl区的交换,但是在其中在ch1和cl结构域中不引入荷电改性的方面,可替代性地包含ch1和cl结构域的交换。图2.原始和优化后的cd3结合剂(cd3orig和cd3opt)与重组cd3的相对结合活性,所述活性通过spr在无应激条件、在40℃和ph6下储存14天后或在37℃和ph7.4下储存14天后测得(igg形式)。图3.原始和优化后的cd3结合剂(cd3orig和cd3opt)与jurkatnfat细胞的结合,所述结合通过流式细胞术测得(igg形式)。利用荧光标记的抗人fc特异性二抗检测与jurkatnfat细胞结合的抗体。图4.实例3中使用的cd3激活测定的示意图。图5.原始和优化后的cd3结合剂(cd3orig和cd3opt)的jurkatnfat激活作用(igg形式)。将jurkatnfat报告细胞与表达抗pglala的cho(cho-pglala)细胞在cd3orig或cd3optiggpglala存在下或以cd3optiggwt作为阴性对照一起孵育。在24h后,通过测量发光来定量分析cd3激活。图6.在实例中制备的t细胞双特异性抗体(tcb)分子的示意图。所有检测的tcb抗体分子均作为包含荷电改性的“2+1iggcrossfab,反向”制得(cd3结合剂中的vh/vl交换,靶抗原结合剂中的荷电改性,ee=147e,213e;rk=123r,124k)。图7.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)的tyrp1tcb与重组cd3的相对结合活性,所述活性通过spr在无应激条件、在40℃和ph6下储存14天后或在37℃和ph7.4下储存14天后测得。图8.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)或相应的tyrp1igg的tyrp1tcb与重组tyrp1的相对结合活性,所述活性通过spr在无应激条件、在40℃和ph6下储存14天后或在37℃和ph7.4下储存14天后测得。图9.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)的tyrp1tcb与jurkatnfat细胞的结合,所述结合通过流式细胞术测得。利用荧光标记的抗人fc特异性二抗检测与jurkatnfat细胞结合的tcb。图10.包含原始和优化后的cd3结合剂的tyrp1tcb的jurkatnfat激活作用。将jurkatnfat报告细胞与黑素瘤细胞系m150543在tyrp1tcbcd3orig或tyrp1tcbcd3opt存在下一起孵育。在24h后,通过测量发光来定量分析在tcb的存在下的cd3激活。图11.包含原始和优化后的cd3结合剂的tyrp1tcb的肿瘤细胞杀伤和t细胞激活作用。利用24h(a,g,m)和48h(b,h,n)后释放的ldh,测定用tyrp1tcbcd3orig和tyrp1tcbcd3opt处理后,来自三个不同健康供体(a-f:供体1,g-l:供体2,m-r:供体3)的pbmc对黑素瘤细胞系m150543的杀伤作用。平行地,利用流式细胞术测量cd25(c,e,i,k,o,q)和cd69(d,f,j,l,p,r)对pbmc内cd8(e,f,k,l,q,r)和cd4(c,d,i,j,o,p)t细胞的上调,作为48h后t细胞激活的标志物。图12.egfrviiiiggpglala的特异性结合。利用流式细胞术检测cho-egfrviii(a)、egfrviii阳性dk-mg(b)和表达egfrwt的mkn-45(c)上egfrviiiiggpglala抗体与egfrviii(不存在与egfrwt的交叉反应性)的特异性结合。包含西妥昔单抗作为egfrwt表达的阳性对照。图13.egfrviiiiggpglala的carj激活作用。将表达抗pglalacar的jurkatnfat报告细胞与表达egfrviii的dk-mg细胞一起孵育,并且采用egfrviiiiggpglala抗体或dp47iggpglala作为阴性对照。通过测量22h后的发光,对jurkatnfat细胞的激活进行定量分析。图14.egfrviiiiggpglala及相应的tcb与egfrviii的结合。利用流式细胞术测量作为iggpglala和转化为tcb的egfrviii结合剂与cho-egfrviii(a)和mkn-45(b)细胞的特异性结合。图15.egfrviiitcb的jurkatnfat激活租用。测定jurkatnfat激活,将其作为cd3在egfrviii阳性dk-mg细胞存在下与egfrviiitcb的接合的制作物。包含dp47tcb作为阴性对照。图16.egfrviiitcb的肿瘤细胞裂解作用。在与新鲜分离的pbmc以及egfrviii阳性dk-mg细胞(a,b)或egfrwt阳性mkn-45细胞(c,d)共培养24h(a,c)或48h(b,d)后,测定egfrviiitcb所诱导的特异性肿瘤细胞裂解。图17.egfrviiitcb的t细胞激活作用。在与新鲜分离的pbmc以及egfrviii阳性dk-mg细胞(a,b,e,f)或egfrwt阳性mkn-45细胞(c,d,g,h)共培养后,测定egfrviiitcb所诱导的t细胞激活,其中使用cd4t细胞(a-d)或cd8t细胞(e-h)上的激活标志物cd25(a,c,e,g)或cd69(b,d,f,h)。图18.egfrviiitcb的细胞因子释放作用。在与新鲜分离的pbmc以及egfrviii阳性dk-mg细胞(a-c)或egfrwt阳性mkn-45细胞(d-f)共培养后,测定egfrviiitcb所诱导的ifnγ(a,d)、tnfα(b,e)和颗粒酶b(c,f)的释放。图19.亲和力成熟的egfrviiiiggpglala的特异性结合。在u87mg-egfrviii细胞(a)和egfrwt阳性细胞系mkn-45(b)上,比较亲和力成熟的egfrviii抗体和亲代egfrviii结合剂与egfrviii的特异性结合。图20.egfrviiitcb的jurkatnfat激活作用。测定jurkatnfat激活,将其作为cd3在egfrviii阳性dk-mg细胞(a)、u87mg-egfrviii细胞(b)和mkn-45细胞(c)存在下与egfrviiitcb的接合的标志物。包含dp47tcb作为阴性对照。图21.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)的egfrviiitcb与重组cd3的相对结合活性,所述活性通过spr在无应激条件、在40℃和ph6下储存14天后或在37℃和ph7.4下储存14天后测得。图22.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)的egfrviiitcb与重组egfrviii的相对结合活性,所述活性通过spr在无应激条件、在40℃和ph6下储存14天后或在37℃和ph7.4下储存14天后测得。图23.包含原始和优化后的cd3结合剂(cd3orig或cd3opt)的egfrviiitcb与jurkatnfat细胞的结合,所述结合通过流式细胞术测得。利用荧光标记的抗人fc特异性二抗检测与jurkatnfat细胞结合的tcb。图24.包含p063.056或p056.021egfrviii结合剂的egfrviiitcb与u87mg-egfrviii细胞的结合,所述结合通过流式细胞术测得。利用荧光标记的抗人fc特异性二抗检测与u87mg-egfrviii细胞结合的tcb。图25.egfrviiitcb的肿瘤细胞裂解和t细胞激活作用。在与新鲜分离的pbmc和u87mg-egfrviii细胞共培养24h(a,c)或48h(b,d)后,测定egfrviiitcb所诱导的特异性肿瘤细胞裂解(a,b)和t细胞激活(c,d)。包含dp47tcb作为阴性对照。图26.比较egfrviiitcb2+1形式和1+1形式的jurkatnfat激活作用。测定jurkatnfat激活,将其作为在egfrviii阳性u87mg-egfrviii细胞存在下,cd3与呈2+1反向形式和呈1+1头到尾形式的egfrviiitcb的接合的标志物。图27.比较egfrviiitcb2+1形式和1+1形式的肿瘤细胞裂解和t细胞激活作用。在与新鲜分离的pbmc和u87mg-egfrviii细胞共培养24h(a,c)或48h(b,d)后,测定呈2+1反向形式和呈1+1头到尾形式的egfrviiitcb所诱导的特异性肿瘤细胞裂解(a,b)和t细胞激活(c,d)。图28.egfrviiitcb的t细胞激活和增殖作用。在与分离自健康供体的u87mg-egfrviii和pbmc共培养后,测定egfrviiitcb所诱导的cd4t细胞(a,b)和cd8t细胞(c,d)的t细胞增殖(a,c)和t细胞激活。图29.egfrviiitcb的肿瘤细胞裂解、t细胞激活和细胞因子释放作用。在与u87mg-egfrviii细胞和pbmc共培养后,测定egfrviiitcb所诱导的肿瘤细胞裂解(a,b)、t细胞激活(c,d)以及ifnγ和tnfα(e,f)的释放作用。在处理24h和48h后,测量肿瘤细胞裂解作用;在48h后测量t细胞激活和细胞因子释放作用。图30.tyrp-1tcb的肿瘤细胞裂解、t细胞激活和细胞因子释放作用。在与源自患者的黑素瘤细胞系m150543和pbmc共培养后,tyrp-1tcb所诱导的肿瘤细胞裂解(a,b)、t细胞激活(c,d)以及ifnγ和tnfα(e,f)的释放作用。在处理24h和48h后,测量肿瘤细胞裂解作用;在48h后测量t细胞激活和细胞因子释放作用。图31.tyrp-1tcb的体内功效。在人源化nsg小鼠中,皮下注射igr-1人黑素瘤细胞系,以在黑素瘤皮下异种移植模型中研究肿瘤生长抑制作用。与媒介物组相比,在tyrp-1tcb组(68%tgi,p=0.0058*)中观察到显著的肿瘤生长抑制(tgi)。图32.egfrviiitcb的体内功效。在人源化nsg小鼠中,皮下注射u87-huegfrviii人胶质母细胞瘤细胞系,以在胶质母细胞瘤皮下异种移植模型中研究肿瘤生长抑制作用。在egfrviiitcb组中观察到显著的肿瘤控制作用,所有小鼠均达到完全缓解。具体实施方式i.定义除非在下文中另外定义,否则本文使用的术语通常如本领域中所使用的。如本文所用,关于抗原结合结构域的术语“第一”、“第二”或“第三”等用于在存在多于一个类型的部分时便于区分。除非明确说明,否则这些术语的使用并非旨在赋予这些部分特定的次序或取向。术语“抗cd3抗体”和“结合cd3的抗体”是指这样的抗体,其能够以足够的亲和力结合cd3,使得所述抗体可用作靶向cd3的诊断和/或治疗剂。在一个方面,例如通过表面等离子体共振(spr)所测得的,抗cd3抗体与不相关的非cd3蛋白的结合程度小于抗体与cd3结合程度的约10%。在某些方面,与cd3结合的抗体具有≤1μm、≤500nm、≤200nm或≤100nm的解离常数(kd)。当抗体具有例如通过spr测得的1μm或更小的kd时,该抗体被认为与cd3“特异性结合”。在某些方面,抗cd3抗体结合cd3的表位,所述表位在来自不同物种的cd3中是保守的。本文的术语“抗体”以最广泛的含义使用,并且包括各种抗体结构,包括但不限于单克隆抗体、多克隆抗体、多特异性抗体(例如,双特异性抗体)和抗体片段,只要它们表现出所需的抗原结合活性即可。“抗体片段”是指完整抗体以外的分子,其包含完整抗体的一部分,该部分结合完整抗体所结合的抗原。抗体片段的实例包括但不限于fv、fab、fab'、fab'-sh、f(ab')2、双体抗体、线性抗体、单链抗体分子(例如,scfv和scfab)、单结构域抗体以及由抗体片段形成的多特异性抗体。关于某些抗体片段的综述,请参见hollinger和hudson,naturebiotechnology23:1126-1136(2005)。术语“全长抗体”、“完整抗体”和“整个抗体”在本文中可互换使用以指代具有与天然抗体结构基本上相似的结构的抗体。如本文所用的术语“单克隆抗体”是指从基本上同质的抗体群体获得的抗体,即除了可能的变异抗体外,该群体中包括的各个抗体是相同的和/或结合相同的表位,所述可能的变异抗体例如含有天然存在的突变或是在单克隆抗体制备物的生产过程中产生的,此类变体通常以微量存在。与通常包括针对不同决定簇(表位)的不同抗体的多克隆抗体制剂相反,单克隆抗体制剂中的每种单克隆抗体针对抗原上的单一决定簇。因此,修饰语“单克隆”表示抗体的特征是从基本上同质的抗体群体获得的,并且不应解释为需要通过任何特定方法产生抗体。例如,单克隆抗体可通过多种技术制备,包括但不限于杂交瘤方法、重组dna方法、噬菌体展示方法,以及利用含有全部或部分人免疫球蛋白基因座的转基因动物的方法,在本文中描述了用于制备单克隆抗体的此类方法和其他示例性方法。“分离的”抗体为已从其自然环境的组分中分离的抗体。在一些方面,通过例如电泳(例如,sds-page、等电聚焦(isoelectricfocusing,ief)、毛细管电泳)或色谱(例如,离子交换或反相hplc、亲和色谱、体积排阻色谱)方法测定,将抗体纯化至大于95%或99%的纯度。关于评定抗体纯度的方法的综述,请参见例如flatman等人,j.chromatogr.b848:79-87(2007)。在一些方面,本发明提供的抗体为分离的抗体。术语“嵌合”抗体是指这样的抗体,在所述抗体中重链和/或轻链的一部分来源于特定来源或物种,而重链和/或轻链的其余部分来源于不同的来源或物种。“人源化”抗体是指这样的嵌合抗体,其包含来自非人cdr的氨基酸残基和来自人fr的氨基酸残基。在某些方面,人源化抗体将基本上包含所有的至少一个、通常两个可变结构域,其中所有或基本上所有cdr对应于非人抗体的cdr,并且所有或基本上所有的fr对应于人抗体的fr。此类可变结构域在本文中称为“人源化可变区”。人源化抗体任选地可以包含来源于人抗体的抗体恒定区的至少一部分。在一些方面,人源化抗体中的一些fr残基被来自非人抗体(例如,cdr残基所来源于的抗体)的相应残基置换,例如以恢复或改善抗体特异性或亲和力。抗体(例如非人抗体)的“人源化形式”是指已经历人源化的抗体。“人抗体”是这样的抗体,该抗体具有的氨基酸序列对应于由人或人细胞产生的抗体的氨基酸序列,或来源于利用人抗体库或其他人抗体编码序列的非人源的抗体的氨基酸序列。人抗体的该定义特别地排除了包含非人抗原结合残基的人源化抗体。在某些方面,人抗体来源于非人转基因哺乳动物,例如小鼠、大鼠或兔。在某些方面,人抗体来源于杂交瘤细胞系。在本文中从人抗体文库分离出的抗体或抗体片段也被认为是人抗体或人抗体片段。术语“抗原结合结构域”是指抗体的一部分,该部分包含与抗原的部分或全部结合并且互补的区域。抗原结合结构域可以由例如一个或多个抗体可变结构域(也称为抗体可变区)提供。在优选的方面,抗原结合结构域包含抗体轻链可变结构域(vl)和抗体重链可变结构域(vh)。术语“可变区”或“可变结构域”是指抗体重链或轻链的参与抗体与抗原结合的结构域。天然抗体的重链和轻链的可变结构域(分别为vh和vl)通常具有相似的结构,其中每个结构域包含四个保守框架区(fr)和互补决定区(cdr)。参见例如:kindt等人,kubyimmunology,第6版,w.h.freeman&co.,第91页(2007)。单个vh或vl结构域可足以赋予抗原结合特异性。此外,结合特定抗原的抗体可分别使用来自结合该抗原的抗体的vh或vl结构域来进行分离,以筛选互补vl或vh结构域的文库。参见例如:portolano等人,j.immunol.150:880-887(1993);clarkson等人,nature352:624-628(1991)。如本文结合可变区序列所用,“kabat编号”是指由kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991)提出的编号系统。如本文所用,重链和轻链的所有恒定区和结构域的氨基酸位置是根据kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991)中描述的kabat编号系统编号的,在本文中被称为“根据kabat编号”或“kabat编号”。具体地,将kabat编号系统(参见kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991)的第647-660页)用于κ和λ同种型的轻链恒定结构域cl,并且将kabat的eu索引编号系统(参见第661-723页)用于重链恒定结构域(ch1、铰链、ch2和ch3),这在本文中通过在此情况下称为“根据kabateu索引编号”或“kabateu索引编号”而进一步分类。如本文所用的术语“高变区”或“hvr”是指抗体可变结构域的在序列中高变并确定抗原结合特异性的各个区域,例如“互补决定区”(“cdr”)。通常,抗体包含六个cdr;三个在vh中(hcdr1,hcdr2,hcdr3),三个在vl中(lcdr1,lcdr2,lcdr3)。本文中的示例性cdr包括:(a)在氨基酸残基26-32(l1)、50-52(l2)、91-96(l3)、26-32(h1)、53-55(h2)和96-101(h3)处发生的高可变环(chothia和lesk,j.mol.biol.196:901-917(1987));(b)存在于氨基酸残基24-34(l1)、50-56(l2)、89-97(l3)、31-35b(h1)、50-65(h2)和95-102(h3)处的cdr(kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md(1991));以及(c)存在于氨基酸残基27c-36(l1)、46-55(l2)、89-96(l3)、30-35b(h1)、47-58(h2)和93-101(h3)处的抗原接触点(maccallum等人,j.mol.biol.262:732-745(1996))。除非另有说明,否则cdr根据kabat等人所述的方法(同上)确定。本领域技术人员将理解,也可以根据chothia(同上)、mccallum(同上)所述的方法或任何其他在科学上接受的命名系统来确定cdr名称。“框架”或“fr”是指除互补决定区(cdr)之外的可变结构域残基。可变结构域的fr通常由以下四个fr结构域组成:fr1、fr2、fr3和fr4。因此,hvr和fr序列通常在vh(或vl)中以以下次序出现:fr1-hcdr1(lcdr1)-fr2-hcdr2(lcdr2)-fr3-hcdr3(lcdr3)-fr4。除非另外指明,否则可变结构域中的cdr残基和其他残基(例如,fr残基)在本文中根据kabat等人,出处同上编号。出于本文目的的“受体人框架”是这样的框架,其包含来源于如下所定义的人免疫球蛋白框架或人共有框架的轻链可变结构域(vl)框架或重链可变结构域(vh)框架的氨基酸序列。“来源于”人免疫球蛋白框架或人共有框架的受体人框架可包含与所述人免疫球蛋白框架或人共有框架相同的氨基酸序列,或者其可以包含氨基酸序列变化。在一些方面,氨基酸变化的数量为10个或更少、9个或更少、8个或更少、7个或更少、6个或更少、5个或更少、4个或更少、3个或更少,或2个或更少。在一些方面,vl受体人框架在序列中与vl人免疫球蛋白框架序列或人共有框架序列相同。“人共有框架”是这样的框架,其表示在人免疫球蛋白vl或vh框架序列的选择中最常存在的氨基酸残基。一般而言,人免疫球蛋白vl或vh序列的选择来自于可变结构域序列的亚组。一般而言,序列的亚组是如kabat等人,sequencesofproteinsofimmunologicalinterest,第五版,nihpublication91-3242,bethesdamd(1991),第1-3卷中所述的亚组。本文的术语“免疫球蛋白分子”是本文指具有天然存在的抗体的结构的蛋白质。例如,igg类免疫球蛋白是约150,000道尔顿的异四聚体糖蛋白,其由通过二硫键键合的两条轻链和两条重链组成。从n末端到c末端,每条重链具有可变结构域(vh)(也称为可变重链结构域或重链可变区),接着是三个恒定结构域(ch1、ch2和ch3)(也称为重链恒定区)。类似地,从n末端到c末端,每条轻链具有可变结构域(vl)(也称为可变轻链结构域或轻链可变区),接着是一个恒定轻链(cl)结构域(也称为轻链恒定区)。免疫球蛋白的重链可分为以下五种类型中的一种:称为α(iga)、δ(igd)、ε(ige)、γ(igg)或μ(igm),它们中的一些可进一步分为亚型,例如γ1(igg1)、γ2(igg2)、γ3(igg3)、γ4(igg4)、α1(iga1)和α2(iga2)。免疫球蛋白的轻链可以基于其恒定结构域的氨基酸序列而被分配为以下两种类型中的一种:称为卡帕(κ)和拉姆达(λ)。免疫球蛋白基本上由通过免疫球蛋白铰链区连接的两个fab分子和一个fc结构域组成。抗体或免疫球蛋白的“类”是指其重链具有的恒定结构域或恒定区的类型。存在五大类抗体:iga、igd、ige、igg和igm,并且它们中的一些可以进一步分为亚类(同种型),例如igg1、igg2、igg3、igg4、iga1,以及iga2。对应于不同类别的免疫球蛋白的重链恒定结构域分别称为α、δ、ε、γ和μ。“fab分子”是指由免疫球蛋白的重链的vh和ch1结构域(“fab重链”)和轻链的vl和cl结构域(“fab轻链”)组成的蛋白质。“交叉(crossover)”fab分子(也称为“crossfab”)是指这样的fab分子,其中fab重链和轻链的可变结构域或恒定结构域被交换(即彼此替换),即交叉fab分子包含由轻链可变结构域vl和重链恒定结构域1ch1组成的肽链(vl-ch1,在n末端至c末端方向上),以及由重链可变结构域vh和轻链恒定结构域cl组成的肽链(vh-cl,在n末端至c末端方向上)。为清楚起见,在fab轻链和fab重链的可变结构域被交换的交叉fab分子中,包含重链恒定结构域1ch1的肽链在本文中被称为(交叉)fab分子的“重链”。相反地,在fab轻链和fab重链的恒定结构域被交换的交叉fab分子中,包含重链可变结构域vh的肽链在本文中被称为(交叉)fab分子的“重链”。与此相比之下,“常规”fab分子是指处于天然形式的fab分子,即包含由重链可变结构域和恒定结构域组成的重链(vh-ch1,在n末端至c末端方向上),以及由轻链可变结构域和恒定结构域组成的轻链(vl-cl,在n末端至c末端方向上)。本文的术语“fc结构域”或“fc区”用于定义免疫球蛋白重链的c末端区,该c末端区含有恒定区的至少一部分。该术语包括天然序列fc区和变体fc区。在一个方面,人igg重链fc区从cys226或从pro230延伸至重链的羧基末端。然而,由宿主细胞产生的抗体可以经历对来自重链的c末端的一个或多个,特别是一个或两个氨基酸的翻译后切割。因此,由宿主细胞通过表达编码全长重链的特定核酸分子产生的抗体可以包括全长重链,或者所述抗体可以包括全长重链的切割变体。这可能是重链的最后两个c末端氨基酸为甘氨酸(g446)和赖氨酸(k447,根据kabateu索引)的情况。因此,fc区的c末端赖氨酸(lys447)或c末端甘氨酸(gly446)和赖氨酸(lys447)可以存在或可以不存在。如果没有另外指明,则包含fc区(或如本文所定义的fc结构域的亚基)的重链的氨基酸序列在本文中被表示为没有c末端甘氨酸-赖氨酸二肽。在一个方面,包括如本文所指定的fc区(亚基)的重链包含在根据本发明的抗体中,该重链包含另外的c末端甘氨酸-赖氨酸二肽(g446和k447,根据kabateu索引编号)。在一个方面,包括如本文所指定的fc区(亚基)的重链包含在根据本发明的抗体中,该重链包含另外的c末端甘氨酸残基(g446,根据kabateu索引编号)。除非本文另外指明,否则fc区或恒定区中氨基酸残基的编号为根据eu编号系统(也称为eu索引),如在kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md,1991(同样参见上文)中所描述的。如本文所用的fc结构域的“亚基”是指形成二聚fc结构域的两种多肽中的一种,即包含免疫球蛋白重链的c末端恒定区的多肽,该多肽能够稳定自缔合。例如,iggfc结构域的亚基包含iggch2和iggch3恒定结构域。所谓“融合”意指组分(例如fab分子和fc结构域亚基)直接地或经由一个或多个肽连接基,通过肽键连接。术语“多特异性”意指抗体能够与至少两种不同的抗原决定簇特异性结合。多特异性抗体可以是例如双特异性抗体。通常,双特异性抗体包含两个抗原结合位点,每个抗原结合位点特异于不同的抗原决定簇。在某些方面,多特异性(例如双特异性)抗体能够同时结合两个抗原决定簇,特别是在两种不同细胞上表达的两种抗原决定簇。如本文所用的术语“价”表示在抗原结合分子中存在指定数目的抗原结合位点。因此,术语“与抗原单价结合”表示在抗原结合分子中存在对抗原具有特异性的一个(并且不超过一个)抗原结合位点。“抗原结合位点”是指提供与抗原的相互作用的抗原结合分子的位点,即一个或多个氨基酸残基。例如,抗体的抗原结合位点包含来自互补决定区(complementaritydeterminingregion,cdr)的氨基酸残基。天然免疫球蛋白分子通常具有两个抗原结合位点,fab分子通常具有单个抗原结合位点。如本文所用,术语“抗原决定簇”或“抗原”是指多肽大分子上的位点(例如一段连续的氨基酸或由非连续氨基酸的不同区域组成的构象构型),抗原结合结构域与所述位点结合,从而形成抗原结合结构域-抗原复合物。有用的抗原决定簇可以在例如肿瘤细胞的表面上、病毒感染细胞的表面上、其他患病细胞的表面上、免疫细胞的表面上、血清中的游离物和/或细胞外基质(ecm)中找到。在一个优选的方面,抗原是人蛋白质。除非另有说明,否则“cd3”是指来自任何脊椎动物来源的任何天然cd3,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人)、非人灵长类动物(例如食蟹猴)和啮齿动物(例如小鼠和大鼠)。该术语包括“全长”的未加工cd3,以及通过细胞中加工产生的任何形式的cd3。该术语还涵盖cd3的天然存在变体,例如剪接变体或等位基因变体。在一个方面,cd3是人cd3,特别是人cd3的ε亚基(cd3ε)。人cd3ε的氨基酸序列示出于seqidno:112(不含信号肽)中。另外参见uniprot(www.uniprot.org)登录号p07766(第189版)或ncbi(www.ncbi.nlm.nih.gov/)refseqnp_000724.1。在另一方面,cd3是食蟹猴(macacafascicularis)cd3,特别是食蟹猴cd3ε。食蟹猴cd3ε的氨基酸序列示出于seqidno:113(不含信号肽)中。另外参见ncbigenbank编号bab71849.1。在某些方面,本发明的抗体与cd3的表位结合,该表位在不同物种的cd3抗原(特别是人和食蟹猴cd3)中是保守的。在优选的方面,抗体与人cd3结合。如本文所用的“靶细胞抗原”是指存在于靶细胞表面上的抗原决定簇,该靶细胞为例如肿瘤中的细胞诸如癌细胞或肿瘤基质的细胞(在那种情况下,称为“肿瘤细胞抗原”)。优选地,靶细胞抗原并非cd3,并且/或者与cd3在不同细胞上表达。在一个优选的方面,靶细胞抗原是tyrp-1,特别是人tyrp-1。在另一个优选的方面,靶细胞抗原是egfrviii,特别是人egfrviii。“tyrp1”或“tyrp-1”代表酪氨酸相关蛋白1,它是一种参与黑色素合成的酶。成熟形式的tyrp1最初也称为gp75,是一种75kda的跨膜糖蛋白。人tyrp1的序列示出于seqidno:114(不含信号肽)中。还参见uniprot登录号p17643(第185版)。除非另有说明,否则如本文所用的“tyrp1”是指来自任何脊椎动物来源的任何天然tyrp1,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人)、非人灵长类动物(例如食蟹猴)和啮齿动物(例如小鼠和大鼠)。该术语包括“全长”的未加工tyrp1,以及通过细胞中加工产生的任何形式的tyrp1。该术语还涵盖tyrp1的天然存在变体,例如剪接变体或等位基因变体。在一个方面,tyrp1是人tyrp1。“egfrviii”代表表皮生长因子受体变体iii,它是egfr的一种突变体,由外显子2-7的框内缺失形成,导致缺失267个氨基酸,在连接点处发生甘氨酸置换。人egfrviii的序列示出于seqidno:115(不含信号肽)中。野生型人egfr的序列示出于seqidno:116(不含信号肽)中。还参见uniprot登录号p00533(第258版)。除非另有说明,否则如本文所用的“egfrviii”是指来自任何脊椎动物来源的任何天然egfrviii,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人)、非人灵长类动物(例如食蟹猴)和啮齿动物(例如小鼠和大鼠)。该术语涵盖“全长”未加工的egfrviii(但不包括野生型egfr)以及在细胞中加工产生的任何形式的egfrviii(例如不含信号肽的egfrviii)。在一个方面,egfrviii是人egfrviii。“亲和力”是指分子(例如,抗体)的单个结合位点与其结合配偶体(例如,抗原)之间的非共价相互作用的总和的强度。除非另有说明,否则如本文所用的“结合亲和力”是指内在结合亲和力,其反映了结合对的成员(例如抗体和抗原)之间的1:1相互作用。分子x对其配偶体y的亲和力一般可由解离常数(kd)表示。亲和力可以通过本领域已知的完善确立的方法测量,包括本文所述的那些方法。用于测量亲和力的优选的方法是表面等离子体共振(spr)。“亲和力成熟的”抗体是指在一个或多个互补决定区(cdr)中具有一个或多个改变的抗体,与不具有此类改变的亲本抗体相比,此类改变导致了抗体对抗原的亲和力的改善。“降低的结合”,例如降低的与fc受体的结合,是指对相应相互作用的亲和力降低,如例如通过spr所测量的。为清楚起见,该术语还包括将亲和力降低至零(或低于分析方法的检测极限),即完全消除相互作用。相反地,“增加的结合”是指对相应相互作用的结合亲和力增加。如本文所用,“t细胞激活”是指t淋巴细胞、特别是细胞毒性t淋巴细胞的一种或多种细胞应答,这些细胞应答选自:增殖、分化、细胞因子分泌、细胞毒性效应分子释放、细胞毒性活性和激活标志物的表达。用于测量t细胞激活的合适的测定方法是本领域已知的并且如本文所述。“促进fc结构域的第一亚基和第二亚基缔合的修饰”是对肽骨架的操纵或对fc结构域亚基的翻译后修饰,该操纵或翻译后修饰减少或阻止包含fc结构域亚基的多肽与相同多肽缔合形成同型二聚体。如本文所用的促进缔合的修饰优选地包括对期望缔合的两个fc结构域亚基(即fc结构域的第一亚基和第二亚基)中的每一者进行的单独修饰,其中修饰彼此互补以促进两个fc结构域亚基的缔合。例如,促进缔合的修饰可以改变fc结构域亚基中的一个或两个亚基的结构或电荷,以便分别使它们的缔合为在空间上或静电上有利。因此,在包含第一fc结构域亚基的多肽与包含第二fc结构域亚基的多肽之间发生(杂)二聚化,该(杂)二聚化可能在以下意义上是不同的:与所述亚基中的每个亚基融合的其他组分(例如抗原结合结构域)是不同的。在一些方面,促进fc结构域的第一亚基和第二亚基进行缔合的修饰包含fc结构域中的氨基酸突变,具体地包含氨基酸置换。在一些优选的方面,促进fc结构域的第一亚基和第二亚基进行缔合的修饰包含fc结构域的两个亚基中的每一个的单独的氨基酸突变,具体地包含氨基酸置换。术语“效应子功能”是指可归因于抗体的fc区、随着抗体同种型的变化而变化的那些生物活性。抗体效应子功能的示例包括:c1q结合和补体依赖性细胞毒性(cdc)、fc受体结合、抗体依赖性细胞介导的细胞毒性(adcc)、抗体依赖性细胞吞噬作用(adcp)、细胞因子分泌、免疫复合物介导的抗原呈递细胞的抗原摄取、下调细胞表面受体(例如b细胞受体),以及b细胞激活。“激活fc受体”是这样的fc受体,该fc受体在由抗体的fc结构域接合后引发信号传导事件,该信号传导事件刺激携带受体的细胞执行效应子功能。人激活fc受体包括fcγriiia(cd16a)、fcγri(cd64)、fcγriia(cd32)和fcαri(cd89)。抗体依赖性细胞介导的细胞毒性(adcc)是导致免疫效应细胞裂解抗体包被的靶细胞的免疫机制。靶细胞是与包含fc区的抗体或其衍生物特异性结合的细胞,该特异性结合通常是通过fc区的n末端的蛋白质部分。如本文所用,术语“降低的adcc”被定义为在靶细胞周围的培养基中给定抗体浓度下在给定时间内通过上面定义的adcc机制裂解的靶细胞数量减少,和/或在靶细胞周围的培养基中通过adcc机制在给定时间内实现对给定数量的靶细胞的裂解所必需的抗体浓度增加。adcc降低是相对于使用相同的标准生产、纯化、配制和储存方法(该等方法为本领域技术人员已知的),由相同类型的宿主细胞产生但尚未被工程化的相同抗体介导的adcc。例如,由fc结构域中包含降低adcc的氨基酸置换的抗体介导的adcc的降低是相对于由fc结构域中没有该氨基酸置换的相同抗体介导的adcc。用于测量adcc的合适测定法是本领域中熟知的(参见例如pct公开号wo2006/082515或pct公开号wo2012/130831)。如本文所用,术语“工程化、工程化的、工程改造”被认为包括对肽骨架的任何操纵,或对天然存在的或重组的多肽或其片段的翻译后修饰。工程改造包括对氨基酸序列、糖基化模式或单独氨基酸的侧链基团的修饰,以及这些方法的组合。如本文所用的术语“氨基酸突变”表示涵盖氨基酸取代、缺失、插入和修饰。可进行置换、缺失、插入和修饰的任何组合以获得最终构建体,前提条件是最终构建体具有所需特征,例如减少与fc受体的结合或增加与另一种肽的缔合。氨基酸序列缺失和插入包括氨基酸的氨基末端和/或羧基末端缺失和插入。优选的氨基酸突变是氨基酸取代。出于改变例如fc区的结合特征的目的,非保守氨基酸置换,即将一种氨基酸替换为具有不同结构和/或化学性质的另一种氨基酸,是特别优选的。氨基酸取代包括用非天然存在的氨基酸或用二十种标准氨基酸的天然存在的氨基酸衍生物(例如4-羟基脯氨酸、3-甲基组氨酸、鸟氨酸、高丝氨酸、5-羟基赖氨酸)进行替代。可以使用本领域熟知的遗传或化学方法来产生氨基酸突变。遗传方法可包括定点诱变、pcr、基因合成等。设想通过除基因工程之外的方法(诸如化学修饰)改变氨基酸侧链基团的方法也是有用的。本文可使用各种名称来指示相同的氨基酸突变。例如,fc结构域的329位的脯氨酸被置换为甘氨酸,可以表示为329g、g329、g329、p329g或pro329gly。相对于参照多肽序列的“氨基酸序列一致性百分比(%)”被定义为在比对候选序列与参考多肽序列并引入空位(如果必要的话)以实现最大的序列一致性百分比之后,并且在不考虑将任何保守取代作为序列一致性的组成部分的情况下,候选序列中的氨基酸残基与参考多肽序列中的氨基酸残基相同的百分比。用于确定氨基酸序列一致性百分比的比对可以通过本领域技术范围内的各种方式实现,例如使用公众可获得的计算机软件,诸如blast、blast-2、clustalw、megalign(dnastar)软件或fasta程序包。本领域技术人员可确定用于比对序列的适当参数,包括在所比较的序列的全长上实现最大比对所需的任何算法。替代性地,可使用序列比较计算机程序align-2生成一致性百分比值。align-2序列比较计算机程序由genentech,inc.编写,并且源代码已经与用户文档一起提交到u.s.copyrightoffice,washingtond.c.,20559,在那里以美国版权登记号txu510087注册,并且如wo2001/007611所述。除非另外指明,否则出于本文的目的,用blosum50判断矩阵,使用fasta包第36.3.8c版或更高版本的ggsearch程序生成氨基酸序列一致性%的值。fasta程序包由w.r.pearson和d.j.lipman(“improvedtoolsforbiologicalsequenceanalysis”,pnas85(1988)2444-2448),pnas(“effectiveproteinsequencecomparison”meth.enzymol.266(1996)227-258)以及pearson等人(genomics46(1997)24-36)创作,并且可从www.fasta.bioch.virginia.edu/fasta_www2/fasta_down.shtml或www.ebi.ac.uk/tools/sss/fasta公开获得。替代性地,可以使用可在fasta.bioch.virginia.edu/fasta_www2/index.cgi处访问的公共服务器来比较序列,使用ggsearch(全局蛋白质:蛋白质)程序和默认选项(blosum50;开放:-10;ext:-2;ktup=2)来确保执行全局而非局部比对。在输出的比对标头(alignmentheader)中给出氨基酸一致性百分比。术语“多核苷酸”或“核酸分子”包括任何包含核苷酸聚合物的化合物和/或物质。每个核苷酸由碱基组成,特别是嘌呤或嘧啶碱基(即胞嘧啶(c)、鸟嘌呤(g)、腺嘌呤(a)、胸腺嘧啶(t)或尿嘧啶(u))、糖(即脱氧核糖或核糖)和磷酸基团。通常,核酸分子通过碱基序列进行描述,由此所述碱基代表核酸分子的一级结构(线性结构)。碱基序列通常表示为从5'至3'。在本文中,术语核酸分子涵盖脱氧核糖核酸(dna)(包括例如互补dna(cdna)和基因组dna)、核糖核酸(rna)(特别是信使rna(mrna))、dna或rna的合成形式,以及包含这些分子中的两种或更多种的混合聚合物。核酸分子可以是线性的或环状的。此外,术语核酸分子包括有义链和反义链,以及单链和双链形式。此外,本文所描述的核酸分子可含有天然存在的或非天然存在的核苷酸。非天然存在的核苷酸的示例包括具有衍生化的糖或磷酸主链键或经化学修饰的残基的经修饰的核苷酸碱基。核酸分子还涵盖适合作为用于本发明的抗体的体外和/或体内(例如,在宿主或患者体内)直接表达的载体的dna和rna分子。此类dna(例如cdna)或rna(例如mrna)载体可以是未修饰的或经修饰的。例如,可以对mrna进行化学修饰以增强rna载体的稳定性和/或编码分子的表达,使得可以将mrna注射到受试者体内以生成体内抗体(参见例如stadler等人(2017)naturemedicine23:815-817或ep2101823b1)。“分离的”核酸分子是指已从其天然环境的组分中分开的核酸分子。分离的核酸分子包括这样的核酸分子,其包含在通常含有所述核酸分子的细胞中,但所述核酸分子存在于染色体外或与其天然染色体位置不同的染色体位置处。“编码抗体的分离的多核苷酸(或核酸)”是指编码抗体重链和轻链(或其片段)的一种或多种多核苷酸分子,包括在单个载体或单独载体中的此类一种或多种多核苷酸分子,以及存在于宿主细胞中的一个或多个位置处的此类一种或多种多核苷酸分子。如本文所用,术语“载体”是指能够载运与其相链接的另一核酸的核酸分子。该术语包括作为自我复制核酸结构的载体,以及整合入其已被引入的宿主细胞的基因组中的载体。某些载体能够指导与其可操作链接的核酸的表达。此类载体在本文中称为“表达载体”。术语“宿主细胞”“宿主细胞系”和“宿主细胞培养物”可互换使用,并且是指外源核酸已被引入其中的细胞,包括此类细胞的子代。宿主细胞包括“转化体”和“转化细胞”,其包括原代转化细胞和来源于所述原代转化细胞的子代,不考虑传代次数。子代可能不与亲本细胞的核酸内容物完全一致,而是可能含有突变。本文包括如在原始转化细胞中筛选或选择的具有相同功能或生物活性的突变子代。宿主细胞是可用于生成本发明的抗体的任何类型的细胞系统。宿主细胞包括培养的细胞,例如哺乳动物的培养细胞,诸如仅举几个示例hek细胞、cho细胞、bhk细胞、ns0细胞、sp2/0细胞、yo骨髓瘤细胞、p3x63小鼠骨髓瘤细胞、per细胞、per.c6细胞或杂交瘤细胞、酵母细胞、昆虫细胞和植物细胞,还包括转基因动物、转基因植物或培养的植物或动物组织中包含的细胞。在一个方面,本发明的宿主细胞是真核细胞,特别是哺乳动物细胞。在一个方面,宿主细胞并非人体内的细胞。术语“药物组合物”或“药物制剂”是指处于允许包含在其中的活性成分的生物活性有效的形式,并且不含对于将被施用组合物的受试者具有不可接受的毒性的另外组分的制剂。“药用载体”是指药物组合物或制剂中除活性成分外的对受试者无毒的成分。药用载体包括但不限于缓冲剂、赋形剂、稳定剂,或防腐剂。如本文所用,“治疗(treatment)”(及其语法变体,诸如“治疗(treat)”或“治疗(treating)”)是指试图改变所治疗个体的疾病的自然病程,并且可以执行以用于预防或在临床病理学过程中执行的临床干预。治疗的期望效果包括但不限于预防疾病的发生或复发、减轻症状、削弱疾病的任何直接或间接病理学后果、预防转移、降低疾病进展的速率、改善或减轻疾病状态,以及缓解或改善预后。在一些方面,本发明的抗体用于延迟疾病的发展或减缓疾病的进展。“个体”或“受试者”是哺乳动物。哺乳动物包括但不限于驯养的动物(例如牛、绵羊、猫、犬和马)、灵长类动物(例如人和非人灵长类动物,诸如猴)、兔以及啮齿类动物(例如小鼠和大鼠)。在某些方面,个体或受试者是人。药剂(例如药物组合物)的“有效量”是指在必要的剂量和时间段下有效实现所需治疗或预防结果的量。术语“包装插页”用于指治疗产品的商业包装中通常包括的说明书,其含有涉及此类治疗产品的使用的有关适应症、用法、剂量、施用、联合疗法、禁忌症和/或警告的信息。ii.组合物和方法本发明提供了结合cd3的抗体,包括结合cd3和第二抗原的多特异性抗体。这些抗体表现出优异的稳定性,并且结合了用于治疗应用的其他有利的特性,例如,在功效和安全性、药代动力学以及可生产性方面。本发明的抗体可用于例如治疗疾病(诸如癌症)。a.抗cd3抗体在一个方面,本发明提供了与cd3结合的抗体。在一个方面,提供了与cd3结合的分离的抗体。在一个方面,本发明提供了与cd3特异性结合的抗体。在某些方面,抗cd3抗体在ph7.4于37℃储存2周后与cd3的结合活性相对于在ph6于-80℃储存2周后的结合活性保留超过约90%,所述结合活性通过表面等离子体共振(spr)测得。在一个方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3。在一个方面,抗体是人源化抗体。在一个方面,抗原结合结构域是人源化抗原结合结构域(即,人源化抗体的抗原结合结构域)。在一个方面,vh和/或vl是人源化可变区。在一个方面,vh和/或vl包含受体人框架,例如人免疫球蛋白框架或人共有框架。在一个方面,vh包含seqidno:7的重链可变区序列的一个或多个重链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vh包含与seqidno:7的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含与seqidno:7的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vh包含与seqidno:7的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vh序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与cd3结合的能力。在某些方面,在seqidno:7的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vh包含seqidno:7的氨基酸序列。任选地,vh包含seqidno:7的氨基酸序列,包括该序列的翻译后修饰。在一个方面,vl包含seqidno:11的轻链可变区序列的一个或多个轻链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vl包含与seqidno:11的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vl包含与seqidno:11的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vl包含与seqidno:11的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vl序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与cd3结合的能力。在某些方面,在seqidno:11的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vl包含seqidno:11的氨基酸序列。任选地,vl包含seqidno:11的氨基酸序列,包括该序列的翻译后修饰。在一个方面,vh包含与seqidno:7的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列,并且vl包含与seqidno:11的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含seqidno:7的氨基酸序列,并且vl包含seqidno:11的氨基酸序列。在进一步方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含:vh,其包含seqidno:7的氨基酸序列;以及vl,其包含seqidno:11的氨基酸序列。在进一步方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含seqidno:7的vh序列和seqidno:11的vl序列。在另一方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含:vh,其包含seqidno:7的vh的重链cdr序列;以及vl,其包含seqidno:11的vl的轻链cdr序列。在进一步方面,第一抗原结合结构域包含seqidno:7的vh的hcdr1、hcdr2和hcdr3氨基酸序列以及seqidno:11的vl的lcdr1、lcdr2和lcdr3氨基酸序列。在一个方面,vh包含seqidno:7的vh的重链cdr序列以及与seqidno:7的vh的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vh包含seqidno:7的vh的重链cdr序列以及与seqidno:7的vh的框架序列具有至少95%的序列同一性的框架。在另一方面,vh包含seqidno:7的vh的重链cdr序列以及与seqidno:7的vh的框架序列具有至少98%的序列同一性的框架。在一个方面,vl包含seqidno:11的vl的轻链cdr序列以及与seqidno:11的vl的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vl包含seqidno:11的vl的轻链cdr序列以及与seqidno:11的vl的框架序列具有至少95%的序列同一性的框架。在另一方面,vl包含seqidno:11的vl的轻链cdr序列以及与seqidno:11的vl的框架序列具有至少98%的序列同一性的框架。在一个方面,本发明提供了一种与cd3结合的抗体,其中该抗体包含第一抗原结合结构域,该第一抗原结合结构域包含如上文所提供的方面中任一项所述的vh序列以及如上文所提供的方面中任一项所述的vl序列。在一个方面,抗体包含人恒定区。在一个方面,抗体是包含人恒定区的免疫球蛋白分子,特别是包含人ch1、ch2、ch3和/或cl结构域的igg类免疫球蛋白分子。人恒定结构域的示例性序列在seqidno:120和seqidno:121(分别为人κ和λcl结构域)和seqidno:122(人igg1重链恒定结构域ch1-ch2-ch3)中给出。在一个方面,抗体包含轻链恒定区,该轻链恒定区包含与seqidno:120或seqidno:121的氨基酸序列、特别是seqidno:120的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。在一个方面,抗体包含重链恒定区,该重链恒定区包含与seqidno:122的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。特别地,重链恒定区可包含如本文所述的fc结构域中的氨基酸突变。在一个方面,第一抗原结合结构域包含人恒定区。在一个方面,第一抗原结合部分为包含人恒定区、特别是人ch1和/或cl结构域的fab分子。在一个方面,第一抗原结合结构域包含轻链恒定区,该轻链恒定区包含与seqidno:120或seqidno:121的氨基酸序列、特别是seqidno:120的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。具体地,轻链恒定区可以包含如本文所述处于“荷电改性”下的氨基酸突变和/或如果在交叉fab分子中则可以包含对一个或多个(特别是两个)n末端氨基酸的缺失或取代。在一些方面,第一抗原结合结构域包含重链恒定区,该重链恒定区包含与seqidno:122的氨基酸序列中包含的ch1结构域序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。具体地,重链恒定区(特别是ch1结构域)可以包含如本文所述处于“荷电改性”下的氨基酸突变。在一个方面,抗体为单克隆抗体。在一个方面,抗体是igg,特别是igg1抗体。在一个方面,抗体为全长抗体。在另一方面,抗体为选自fv分子、scfv分子、fab分子和f(ab')2分子;特别是fab分子的组的抗体片段。在另一方面,抗体片段为双体抗体、三体抗体或四体抗体。在一个方面,第一抗原结合结构域为fab分子。在一个优选的方面,第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1,特别是可变结构域vl和vh彼此替换(即,第一抗原结合结构域是交叉fab分子)。在进一步方面,根据上述方面中任一项所述的抗体可单独或组合地并入如下文第ii.a.1.-8.节所述的特征中的任一者。在一个优选的方面,抗体包含fc结构域,特别是iggfc结构域,更特别是igg1fc结构域。在一个方面,fc结构域是人fc结构域。在一个方面,fc结构域是人igg1fc结构域。fc结构域由第一亚基和第二亚基组成,并且可单独或组合地并入如下文所述的关于fc结构域变体(第ii.a.8.节)所述的特征中的任一者。在另一个优选的方面,抗体包含与第二抗原结合的第二抗原结合结构域和任选的第三抗原结合结构域(即,抗体为多特异性抗体),如下文(第ii.a.7.节)进一步所述。1.抗体片段在某些方面,本文提供的抗体为抗体片段。在一个方面,抗体片段为fab、fab'、fab'-sh或f(ab')2分子,特别是如本文所述的fab分子。“fab'分子”与fab分子的不同之处在于fab'分子在ch1结构域的羧基末端添加了一些残基,这些残基包括来自抗体铰链区的一个或多个半胱氨酸。fab'-sh是这样的fab'分子,其恒定结构域的一个或多个半胱氨酸残基具有游离硫醇基。胃蛋白酶处理产生f(ab')2分子,该分子具有两个抗原结合位点(两个fab分子)和fc区的一部分。在另一方面,抗体片段为双体抗体、三体抗体或四体抗体。“双体抗体”是具有两个抗原结合位点的抗体片段,其可以是二价或双特异性的。参见例如:ep404,097;wo1993/01161;hudson等人,nat.med.9:129-134(2003);和hollinger等人,proc.natl.acad.sci.usa90:6444-6448(1993)。hudson等人在《自然医学》(nat.med.)9:129-134(2003)中还描述了三体抗体和四体抗体。在进一步方面,抗体片段为单链fab分子。“单链fab分子”或“scfab”是由抗体重链可变结构域(vh)、抗体重链恒定结构域1(ch1)、抗体轻链可变结构域(vl)、抗体轻链恒定结构域(cl)和连接基组成的多肽,其中所述抗体结构域和所述连接基在n末端至c末端方向上具有以下顺序中的一种:a)vh-ch1-连接基-vl-cl,b)vl-cl-连接基-vh-ch1,c)vh-cl-连接基-vl-ch1,或d)vl-ch1-连接基-vh-cl。特别地,所述连接基为包含至少30个氨基酸、优选在32个与50个之间的氨基酸的多肽。所述单链fab分子经由cl结构域与ch1结构域之间的天然二硫键而稳定化。此外,这些单链fab分子可以通过经由插入半胱氨酸残基(例如根据kabat编号的可变重链中的44位和可变轻链中的100位)生成链间二硫键,而进一步稳定化。在另一方面,抗体片段为单链可变片段(scfv)。“单链可变片段”或“scfv”为抗体的重链(vh)和轻链(vl)的可变结构域的融合蛋白,其通过连接基连接。特别地,连接基为包含10个至25个氨基酸的短多肽,并且通常富含甘氨酸以获得柔性,以及富含丝氨酸或苏氨酸以获得溶解性,并且可以将vh的n末端与vl的c末端连接,或反之亦然。尽管去除了恒定区并且引入了连接基,但是该蛋白质仍然保留了原始抗体的特异性。关于scfv片段的综述,参见例如plückthun在theharmacologyofmonoclonalantibodies,第113卷,rosenburg和moore编著(springer-verlag,newyork,第269-315页(1994))中所述;还可参见wo93/16185;以及美国专利号5,571,894和5,587,458。在另一方面,抗体片段为单结构域抗体。“单结构域抗体”是包含抗体的全部或部分重链可变结构域或全部或部分轻链可变结构域的抗体片段。在某些方面,单结构域抗体是人单结构域抗体(domantis,inc.,waltham,ma;参见例如美国专利号6,248,516b1)。抗体片段可以通过各种技术制备,包括但不限于完整抗体的蛋白水解消化以及由重组宿主细胞(例如,大肠杆菌)产生,如本文所述。2.人源化抗体在某些方面,本文提供的抗体为人源化抗体。通常,非人抗体经过人源化以降低对人的免疫原性,同时保留亲代非人抗体的特异性和亲和力。一般而言,人源化抗体包含一个或多个可变结构域,其中cdr(或其部分)源自非人抗体,并且fr(或其部分)源自人抗体序列。人源化抗体任选地还将包含人恒定区的至少一部分。在一些方面,人源化抗体中的一些fr残基被来自非人抗体(例如,cdr残基所来源于的抗体)的相应残基置换,例如以恢复或改善抗体特异性或亲和力。人源化抗体及其制备方法在例如almagro和fransson,front.biosci.13:1619-1633(2008)中综述,并且进一步描述于例如riechmann等人,nature332:323-329(1988);queen等人,proc.nat’lacad.sci.usa86:10029-10033(1989);美国专利号5,821,337、7,527,791、6,982,321和7,087,409;kashmiri等人,methods36:25-34(2005)(描述了特异性决定区(sdr)移植);padlan,mol.immunol.28:489-498(1991)(描述了“表面再塑”);dall’acqua等人,methods36:43-60(2005)(描述了“fr改组”);以及osbourn等人,methods36:61-68(2005)和klimka等人,br.j.cancer,83:252-260(2000)(描述了用于fr改组的“指导选择”方法)中。可用于人源化的人框架区包括但不限于:使用“最佳匹配”方法选择的框架区(参见例如sims等人,j.immunol.151:2296(1993));来源于具有轻链或重链可变区的特定子组的人抗体的共有序列的框架区(参见例如carter等人,proc.natl.acad.sci.usa,89:4285(1992);以及presta等人,j.immunol.,151:2623(1993));人成熟(体细胞突变)框架区或人类种系框架区(参见例如almagro和fransson,front.biosci.13:1619-1633(2008));以及来源于筛选fr文库的框架区(参见例如baca等人,j.biol.chem.272:10678-10684(1997)和rosok等人,j.biol.chem.271:22611-22618(1996))。3.糖基化变体在某些方面,改变本文提供的抗体以增加或降低抗体糖基化的程度。可通过改变氨基酸序列,使得产生或去除一个或多个糖基化位点,从而实现便利地添加或去除抗体的糖基化位点。在抗体包含fc区的情况下,可以改变与其连接的寡糖。由哺乳动物细胞产生的天然抗体通常包含具有支链的双触角寡糖,所述双触角寡糖通常通过n-连接附接于fc区的ch2结构域的asn297。参见,例如,wright等人tibtech15:26-32(1997)。寡糖可包括各种碳水化合物,例如,甘露糖、n-乙酰基葡糖胺(glcnac)、半乳糖和唾液酸,以及附接于双触角寡糖结构的“主干”中的glcnac的岩藻糖。在一些方面,可以对本发明的抗体中的寡糖进行修饰,以便产生具有某些改善的特性的抗体变体。在一个方面,提供具有非岩藻糖基化寡糖的抗体变体,即一种缺少连接(直接或间接)至fc区的岩藻糖的寡糖结构。此类非岩藻糖基化寡糖(也称为“去岩藻糖基化”寡糖)特别是在双角寡糖结构的茎中缺少与第一glcnac连接的岩藻糖残基的n-连接的寡糖。在一个方面,提供的抗体变体与天然或亲代抗体相比,在fc区的非岩藻糖基化寡糖比例有所增加。例如,非岩藻糖基化寡糖的比例可以为至少约20%、至少约40%、至少约约60%、至少约80%或甚至约100%(即,不存在岩藻糖基化寡糖)。非岩藻糖基化寡糖的百分比是缺少岩藻糖残基的寡糖相对于连接至asn297的所有寡糖的总和的(平均)量(例如,复合物、杂交和高甘露糖结构),其通过maldi-tof质谱法测得,如wo2006/082515中所述。asn297是指位于fc区中约297位的天冬酰胺残基(fc区残基的eu编号);但是,由于抗体中的序列变化微小,因此asn297也可以位于位置297的上游或下游大约±3个氨基酸,即,在位置294与300之间。在fc区具有增加的非岩藻糖基化寡糖比例的此类抗体可具有改善的fcγriiia受体结合和/或改善的效应子功能,特别是改善的adcc功能。参见例如us2003/0157108和us2004/0093621。能够产生具有减少的岩藻糖基化的抗体的细胞系的实例包括:缺乏蛋白质岩藻糖基化的lec13cho细胞(ripka等人,arch.biochem.biophys.249:533-545(1986);us2003/0157108;以及wo2004/056312,尤其是实例11);以及敲除细胞系,诸如α-1,6-岩藻糖基转移酶基因fut8敲除cho细胞(参见例如yamane-ohnuki等人,biotech.bioeng.,87:614-622(2004);kanda,y.等人,biotechnol.bioeng.,94(4):680-688(2006);和wo2003/085107);或具有减少或消除gdp-岩藻糖合成活性的细胞或转运蛋白的细胞(参见例如us2004259150、us2005031613、us2004132140、us2004110282)。在进一步方面,提供了包含两分型寡糖的抗体变体,例如,其中连接至抗体的fc区的双角寡糖被glcnac两分。如上所述,此类抗体变体可具有减少的岩藻糖基化和/或改善的adcc功能。此类抗体变体的实例描述于例如以下文献中:umana等人,natbiotechnol17,176-180(1999);ferrara等人,biotechnbioeng93,851-861(2006);wo99/54342;wo2004/065540;wo2003/011878。还提供了在附接于fc区的寡糖中具有至少一个半乳糖残基的抗体变体。此类抗体变体可具有改善的cdc功能。此类抗体变体描述于例如wo1997/30087、wo1998/58964和wo1999/22764中。4.经半胱氨酸工程化改造的抗体变体在某些方面,可期望产生经半胱氨酸工程化改造的抗体,例如thiomabtm抗体,其中抗体的一个或多个残基被半胱氨酸残基置换。在优选的方面,置换的残基存在于抗体的可接近位点。通过用半胱氨酸置换那些残基,反应性硫醇基团由此定位于抗体的可接近位点,并且可用于将抗体与其他部分,诸如药物部分或连接基-药物部分缀合,以产生免疫缀合物,如本文进一步所述。可按照例如美国专利号7,521,541、8,30,930、7,855,275、9,000,130或wo2016040856所述生成经半胱氨酸工程化改造的抗体。5.抗体衍生物在某些方面,可进一步修饰本文提供的抗体以使其含有本领域已知且易于获得的另外的非蛋白质部分。适合于抗体衍生化的部分包括但不限于水溶性聚合物。水溶性聚合物的非限制性示例包括但不限于聚乙二醇(peg)、乙二醇/丙二醇的共聚物、羧甲基纤维素、葡聚糖、聚乙烯醇、聚乙烯吡咯烷酮、聚-1,3-二氧戊环、聚-1,3,6-三噁烷、乙烯/马来酸酐共聚物、聚氨基酸(均聚物或随机共聚物)和葡聚糖或聚(n-乙烯吡咯烷酮)聚乙二醇、丙二醇均聚物、聚环氧丙烷/环氧乙烷共聚物、聚氧乙烯化多元醇(例如甘油)、聚乙烯醇以及它们的混合物。由于其在水中的稳定性,聚乙二醇丙醛在制造中可具有优势。聚合物可具有任何分子量,并且可以具有支链或不具有支链。附接至抗体的聚合物的数目可变,并且如果附接了多于一个聚合物,那么它们可以为相同或不同的分子。通常,可基于以下考虑因素确定用于衍生化的聚合物的数目和/或类型,包括但不限于抗体待改善的特定特性或功能、抗体衍生物是否将用于限定条件下的疗法等。6.免疫缀合物本发明还提供了免疫缀合物,其包含与一种或多种治疗剂缀合的如本文所述的抗cd3抗体,所述一种或多种治疗剂为诸如细胞毒性剂、化疗剂、药物、生长抑制剂、毒素(例如细菌、真菌、植物或动物来源的蛋白质毒素、酶活性毒素,或它们的片段)或放射性同位素。在一个方面,免疫缀合物是抗体药物偶联物(adc),其中抗体与上文所述的治疗剂中的一种或多种缀合。抗体通常使用连接基与治疗剂中的一种或多种连接。adc技术概述如pharmacolreview68:3-19(2016)所示,其中包括治疗剂和药物以及连接基的实例。在另一方面,免疫缀合物包含与酶活性毒素或其片段缀合的本发明的抗体,这些酶活性毒素或其片段包括但不限于:白喉毒素a链、白喉毒素的非结合活性活性片段、外毒素a链(来自铜绿假单胞菌)、蓖麻毒蛋白a链、相思豆毒素a链、modeccina链、α-八叠球菌、油桐蛋白质、石竹素蛋白质、phytolacaamericana蛋白质(papi、papii和pap-s)、苦瓜抑制剂、毒蛋白、巴豆毒蛋白、sapaonariaofficinalis抑制剂、白树毒素、mitogellin、局限曲菌素、酚霉素、伊诺霉素和单端孢霉烯。在另一方面,免疫缀合物包含本发明的抗体,该抗体与放射性原子缀合以形成放射性缀合物。各种放射性同位素可用于生产放射性缀合物。实例包括at211、i131、i125、y90、re186、re188、sm153、bi212、p32、pb212以及lu的放射性同位素。当放射性缀合物用于检测时,它可以包含用于闪烁摄影研究的放射性原子,例如tc99m或i123,或用于核磁共振(nmr)成像(也称为磁共振成像,mri)的自旋标记,诸如i123、i131、in111、f19、c13、n15、o17、钆、锰或铁。可使用多种双功能蛋白偶联剂诸如n-琥珀酰亚胺基-3-(2-吡啶基二硫代)丙酸酯(spdp)、4-(n-马来酰亚胺基甲基)环己烷-1-羧酸琥珀酰亚胺酯(smcc)、亚氨基硫杂环戊烷(it)、亚氨基酯的双官能衍生物(诸如己二酸二甲酯盐酸盐)、活性酯(诸如辛二酸二琥珀酰亚胺基酯)、醛(诸如戊二醛)、双叠氮基化合物(诸如双(对叠氮基苯甲酰基)己二胺)、双重氮衍生物(诸如双-(对重氮苯甲酰基)-乙二胺)、二异氰酸酯(诸如甲苯2,6-二异氰酸酯)和双活性氟化合物(诸如1,5-二氟-2,4-二硝基苯)制备抗体与细胞毒性剂的缀合物。例如,可以如vitetta等人,《科学》(science)238:1098(1987)中所述制备蓖麻毒蛋白免疫毒素。碳-14标记的1-异硫氰基苄基-3-甲基二亚乙基三胺五乙酸(mx-dtpa)为一种示例性螯合剂,用于将放射性核苷酸缀合至抗体。参见wo94/11026。连接基可以是便于在细胞中释放细胞毒性药物的“可切割连接基”。例如,可使用酸不稳定的连接基、肽酶敏感的连接基、光不稳定的连接基、二甲基连接基或含二硫键的连接基(chari等人,cancerres.52:127-131(1992);美国专利号5,208,020)。本文的免疫缀合物或adc明确考虑但不限于用交联试剂制备的此类缀合物,其中包括但不限于可商购获得(例如,购自piercebiotechnology,inc.,rockford,il.,u.s.a)的bmps、emcs、gmbs、hbvs、lc-smcc、mbs、mpbh、sbap、sia、siab、smcc、smpb、smph、磺基-emcs、磺基-gmbs、磺基-kmus、磺基-mbs、磺基-siab、磺基-smcc和磺基-smpb以及svsb(琥珀酰亚胺基-(4-乙烯基砜)苯甲酸酯)。7.多特异性抗体在某些方面,本文提供的抗体为多特异性抗体,特别是双特异性抗体。多特异性抗体是对至少两种不同的抗原决定簇(例如,两种不同的蛋白质或同一蛋白质上的两个不同的表位)具有结合特异性的单克隆抗体。在某些方面,多特异性抗体具有三种或更多种结合特异性。在某些方面,结合特异性之一针对cd3,并且另一特异性针对任何其他抗原。在某些方面,多特异性抗体可与cd3的两个(或更多个)不同表位结合。多特异性(例如,双特异性)抗体还可用于将细胞毒性剂或细胞定位至表达cd3的细胞。多特异性抗体可制备呈全长抗体或抗体片段。用于制备多特异性抗体的技术包括但不限于具有不同特异性的两个免疫球蛋白重链-轻链对的重组共表达(参见milstein和cuello,nature305:537(1983))和“杵臼结构”工程化(参见例如美国专利号5,731,168和atwell等人,j.mol.biol.270:26(1997))。多特异性抗体还可通过以下方法进行制备:用于制备抗体fc-异源二聚分子的工程化静电转向效应(参见例如wo2009/089004);交联两种或更多种抗体或片段(参见例如:美国专利号4,676,980;以及brennan等人,science,229:81(1985));使用亮氨酸拉链产生双特异性抗体(参见例如:kostelny等人,j.immunol.,148(5):1547-1553(1992);以及wo2011/034605);使用常见的轻链技术来规避轻链错配问题(参见例如wo98/50431);使用“双体抗体”技术制备双特异性抗体片段(参见例如hollinger等人,proc.natl.acad.sci.usa,90:6444-6448(1993));以及使用单链fv(sfv)二聚体(参见例如gruber等人,j.immunol.,152:5368(1994));以及按照例如tutt等人(j.immunol.147:60(1991))所述的方法制备三特异性抗体。本文还包括具有三个或更多个抗原结合位点的工程化改造的抗体,包括例如“章鱼(octopus)抗体”或dvd-ig(参见例如wo2001/77342和wo2008/024715)。具有三个或更多个抗原结合位点的多特异性抗体的其他实例可参见wo2010/115589、wo2010/112193、wo2010/136172、wo2010/145792和wo2013/026831。多特异性抗体或其抗原结合片段还包括“双重作用fab”或“daf”,其包含与cd3以及另一种不同抗原或cd3的两个不同表位结合的抗原结合位点(参见例如us2008/0069820和wo2015/095539)。还可以提供呈不对称形式的多特异性抗体,其在一个或多个具有相同抗原特异性的结合臂中具有结构域交叉(所谓“crossmab”技术),即通过交换vh/vl结构域(参见例如wo2009/080252和wo2015/150447)、ch1/cl结构域(参见例如wo2009/080253)或完整的fab臂(参见例如wo2009/080251、wo2016/016299;也参见:schaefer等人,pnas,108(2011)1187-1191和klein等人,mabs8(2016)1010-20)。不对称的fab臂也可以通过在结构域界面中引入带电荷或不带电荷的氨基酸突变来进行工程化改造,以引导正确的fab配对。参见例如wo2016/172485。多特异性抗体的各种其他分子形式是本领域已知的,并且包括在本文中(参见例如spiess等人,molimmunol67(2015)95-106)。还包括在本文中的一种特定类型的多特异性抗体是设计为同时与靶细胞(例如肿瘤细胞)上的表面抗原以及t细胞受体(tcr)复合物的激活非变体组分(诸如cd3)结合的双特异性抗体,其用于重新定向t细胞以杀死靶细胞。因此,在优选的方面,本文提供的抗体为多特异性抗体,特别是双特异性抗体,其中结合特异性之一针对的是cd3,并且另一种特异性针对的是靶细胞抗原。可用于此目的的双特异性抗体形式的实例包括但不限于:所谓“bite”(双特异性t细胞衔接子)分子,其中两个scfv分子通过柔性连接基融合(参见例如wo2004/106381、wo2005/061547、wo2007/042261以及wo2008/119567,nagorsen和expcellres317,1255-1260(2011));双体抗体(holliger等人,proteng9,299-305(1996))及其衍生物,诸如串联双体抗体(“tandab”;kipriyanov等人,jmolbiol293,41-56(1999));“dart”(双亲和性重新靶向)分子,其基于双体抗体形式,但以c末端二硫键为特征以用于额外的稳定化(johnson等人,jmolbiol399,436-449(2010));以及所谓的三功能抗体,其为完整的杂交小鼠/大鼠igg分子(如seimetz等人所综述:cancertreatrev36,458-467(2010))。本文所包含的特定的t细胞双特异性抗体形式描述于以下文献中:wo2013/026833;wo2013/026839;wo2016/020309;bacac等人,oncoimmunology5(8)(2016)e1203498。下文描述了本发明的多特异性抗体的优选方面。在一个方面,本发明提供了一种与cd3结合的抗体,该抗体包含与cd3结合的第一抗原结合结构域(如本文所述)并且包含与第二抗原结合的第二抗原结合结构域和任选的第三抗原结合结构域。根据本发明的优选方面,抗体中包含的抗原结合结构域为fab分子(即,抗原结合结构域由重链和轻链组成,该重链和轻链各自包含可变结构域和恒定结构域)。在一个方面,第一抗原结合结构域、第二抗原结合结构域和/或在存在时的第三抗原结合结构域为fab分子。在一个方面,所述fab分子是人fab分子。在一个优选的方面,所述fab分子经过人源化。在又一方面,所述fab分子包含人重链恒定结构域和人轻链恒定结构域。优选的,抗原结合结构域中的至少一个是交叉fab分子。此类修饰减少了来自不同fab分子的重链和轻链的错配,从而提高了重组生产中本发明的(多特异性)抗体的产率和纯度。在可用于本发明的(多特异性)抗体的优选的交叉fab分子中,fab轻链和fab重链的可变结构域(分别为vl和vh)交换。然而,即使进行该结构域交换,由于错配的重链与轻链之间的所谓bencejones型相互作用,(多特异性)抗体的制备可能包含某些副产物(参见schaefer等人,pnas,108(2011)11187-11191)。为进一步减少来自不同fab分子的重链和轻链的错配并由此提高所需(多特异性)抗体的纯度和产率,可以在结合第一抗原(cd3)的fab分子或与第二抗原(例如靶细胞抗原,诸如tyrp-1或egfrviii)结合的一种或多种fab分子中的任一者的ch1和cl结构域的特定氨基酸位置处引入带相反电荷的荷电氨基酸,如本文进一步描述的。在(多特异性)抗体中包含的一种或多种常规fab分子中(诸如在图1a-c、图g-j中所示)或在(多特异性)抗体中包含的一种或多种vh/vl交叉fab分子中(诸如在图1d-f、图k-n中所示)(而不是在两者中)进行荷电改性。在优选的方面,在(多特异性)抗体中包含的一种或多种常规fab分子中进行荷电改性(其在优选的方面,与第二抗原(例如靶细胞抗原,诸如tyrp-1或egfrviii)结合)。在根据本发明所述的一个优选的方面,(多特异性)抗体能够同时与第一抗原(即,cd3)和第二抗原(例如靶抗原,诸如tyrp-1或egfrviii)结合。在一个方面,(多特异性)抗体能够通过同时与cd3和靶细胞抗原结合而使t细胞与靶细胞交联。在一个甚至更优选的方面,此类同时结合导致靶细胞裂解,该靶细胞特别是表达靶细胞抗原(例如tyrp-1或egfrviii)的肿瘤细胞。在一个方面,此类同时结合导致t细胞的激活。在其他方面,此类同时结合导致t淋巴细胞、特别是细胞毒性t淋巴细胞的细胞应答,该细胞应答选自下组:增殖、分化、细胞因子分泌、细胞毒性效应分子释放、细胞毒性活性和激活标志物的表达。在一个方面,(多特异性)抗体与cd3结合而不同时与靶细胞抗原结合,不引起t细胞激活。在一个方面,(多特异性)抗体能够将t细胞的细胞毒性活性重新导向靶细胞。在一个优选的方面,所述重新导向与mhc介导的靶细胞的肽抗原呈递和/或t细胞的特异性无关。优选地,根据本发明的方面中任一项所述的t细胞是细胞毒性t细胞。在一些方面,t细胞是cd4+或cd8+细胞,特别是cd8+t细胞。a)第一抗原结合结构域本发明的(多特异性)抗体包含至少一个与cd3结合的抗原结合结构域(第一抗原结合结构域)。在优选的方面,cd3是人cd3(seqidno:112)或食蟹猴cd3(seqidno:113),更特别是人cd3。在一个方面,第一抗原结合结构域对人cd3和食蟹猴cd3具有交叉反应性(即,特异性结合)。在一些方面,cd3为cd3的ε亚基(cd3ε)。在一个优选的方面,(多特异性)抗体包含不多于一个与cd3结合的抗原结合结构域。在一个方面,(多特异性)抗体提供了与cd3的单价结合。在一个方面,与cd3结合的抗原结合结构域为选自fv分子、scfv分子、fab分子和f(ab')2分子的组的抗体片段。在一个优选的方面,与cd3结合的抗原结合结构域为fab分子。在优选的方面,与cd3结合的抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域ch1和cl彼此交换/替换的fab分子。在此类方面,与第二抗原(例如,靶细胞抗原,诸如tyrp-1或egfrviii)结合的一个或多个抗原结合结构域优选地是常规fab分子。在存在多于一个与(多特异性)抗体中包含的第二抗原结合的抗原结合结构域、特别是fab分子的方面,与cd3结合的抗原结合结构域优选地是交叉fab分子,并且与第二抗原结合的抗原结合结构域是常规fab分子。在另一方面,与cd3结合的抗原结合结构域是常规fab分子。在此类方面,与第二抗原(例如靶细胞抗原,诸如tyrp-1或egfrviii)结合的一个或多个抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域ch1和cl彼此交换/替换的fab分子。在存在多于一个与(多特异性)抗体中包含的cd3结合的抗原结合结构域、特别是fab分子的方面,与第二抗原结合的抗原结合结构域优选地是交叉fab分子,并且与cd3结合的抗原结合结构域是常规fab分子。在优选的方面,第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1,特别是可变结构域vl和vh彼此替换(即,根据此类方面,第一抗原结合结构域是交叉fab分子,其中fab轻链和fab重链的可变结构域或恒定结构域发生交换)。在一个此类方面,第二抗原结合结构域(以及第三抗原结合结构域,如有)是常规fab分子。在一个方面,(多特异性)抗体中存在不超过一个与cd3结合的抗原结合结构域(即,抗体提供了与cd3的单价结合)。b)第二抗原结合结构域(和第三抗原结合结构域)在某些方面,本发明的(多特异性)抗体包含至少一个与第二抗原结合的抗原结合结构域,特别是fab分子。第二抗原优选地并非cd3,即不同于cd3。在一个方面,第二抗原是在与cd3不同的细胞上表达的抗原(例如,在不同于t细胞的细胞上表达)。在一个方面,第二抗原是靶细胞抗原,特别是肿瘤细胞抗原。在一个具体方面,第二抗原是tyrp-1。在另一个具体的方面,第二抗原是egfrviii。第二抗原结合结构域能够将(多特异性)抗体导向靶位点,例如导向表达第二抗原的特定类型的肿瘤细胞。在一个方面,与第二抗原结合的抗原结合结构域为选自fv分子、scfv分子、fab分子和f(ab')2分子的组的抗体片段。在一个优选的方面,与第二抗原结合的抗原结合结构域为fab分子。在某些方面,(多特异性)抗体包含两个与第二抗原结合的抗原结合结构域,特别是fab分子。在一个优选的此类方面,这些抗原结合结构域中的每一个与相同的抗原决定簇结合。在一个甚至更优选的方面,这些抗原结合结构域中的全部均相同,即,它们具有相同的分子形式(例如,常规fab分子或交叉fab分子)并且包含相同的氨基酸序列,这些氨基酸序列包括如本文所述的ch1结构域和cl结构域中的相同的氨基酸置换(如有)。在一个方面,(多特异性)抗体包含不超过两个与第二抗原结合的抗原结合结构域,特别是fab分子。在优选的方面,与第二抗原结合的一个或多个抗原结合结构域是常规fab分子。在此类方面,与cd3结合的一个或多个抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域ch1和cl彼此交换/替换的fab分子。在另一方面,与第二抗原结合的一个或多个抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域ch1和cl彼此交换/替换的fab分子。在此类方面,与cd3结合的一个或多个抗原结合结构域是常规fab分子。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含人恒定区。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)是包含人恒定区、特别是人ch1和/或cl结构域的fab分子。人恒定结构域的示例性序列在seqidno:120和seqidno:121(分别为人κ和λcl结构域)和seqidno:122(人igg1重链恒定结构域ch1-ch2-ch3)中给出。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含轻链恒定区,该轻链恒定区包含与seqidno:120或seqidno:121的氨基酸序列、特别是seqidno:120的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。具体地,轻链恒定区可以包含如本文所述处于“荷电改性”下的氨基酸突变和/或如果在交叉fab分子中则可以包含对一个或多个(特别是两个)n末端氨基酸的缺失或取代。在一些方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含重链恒定区,该重链恒定区包含与seqidno:122的氨基酸序列中包含的ch1结构域序列至少约95%、96%、97%、98%、99%或100%相同的氨基酸序列。具体地,重链恒定区(特别是ch1结构域)可以包含如本文所述处于“荷电改性”下的氨基酸突变。tyrp-1在优选的方面,第二抗原是tyrp-1,特别是人tyrp-1(seqidno:114)。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:重链可变区(vh),其包含seqidno:15的重链互补决定区(hcdr)1、seqidno:16的hcdr2和seqidno:17的hcdr3;以及轻链可变区(vl),其包含seqidno:19的轻链互补决定区(lcdr)1、seqidno:20的lcdr2和seqidno:21的lcdr3。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)为(来源于)人源化抗体。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)为人源化抗原结合结构域(即人源化抗体的抗原结合结构域)。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh和/或vl为人源化可变区。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh和/或vl包含受体人框架,例如人免疫球蛋白框架或人共有框架。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含seqidno:18的一个或多个重链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vh包含与seqidno:18的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含与seqidno:18的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vh包含与seqidno:18的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vh序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与tyrp-1结合的能力。在某些方面,在seqidno:18的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vh包含seqidno:18的氨基酸序列。任选地,vh包含seqidno:18的氨基酸序列,包括该序列的翻译后修饰。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含seqidno:22的一个或多个轻链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vl包含与seqidno:22的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vl包含与seqidno:22的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vl包含与seqidno:22的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vl序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与tyrp-1结合的能力。在某些方面,在seqidno:22的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vl包含seqidno:22的氨基酸序列。任选地,vl包含seqidno:22的氨基酸序列,包括该序列的翻译后修饰。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含与seqidno:18的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列,并且第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含与seqidno:22的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含seqidno:18的氨基酸序列,并且vl包含seqidno:22的氨基酸序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh,其包含seqidno:18的序列;以及vl,其包含seqidno:22的序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含seqidno:18的vh序列以及seqidno:22的vl序列。在另一方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh,其包含seqidno:18的vh的重链cdr序列;以及vl,其包含seqidno:22的vl的轻链cdr序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含seqidno:18的vh的hcdr1、hcdr2和hcdr3氨基酸序列以及seqidno:22的vl的lcdr1、lcdr2和lcdr3氨基酸序列。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含seqidno:18的vh的重链cdr序列以及与seqidno:18的vh的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vh包含seqidno:18的vh的重链cdr序列以及与seqidno:18的vh的框架序列具有至少95%的序列同一性的框架。在另一方面,vh包含seqidno:18的vh的重链cdr序列以及与seqidno:18的vh的框架序列具有至少98%的序列同一性的框架。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含seqidno:22的vl的轻链cdr序列以及与seqidno:22的vl的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vl包含seqidno:22的vl的轻链cdr序列以及与seqidno:22的vl的框架序列具有至少95%的序列同一性的框架。在另一方面,vl包含seqidno:22的vl的轻链cdr序列以及与seqidno:22的vl的框架序列具有至少98%的序列同一性的框架。egfrviii在优选的方面,第二抗原是egfrviii,特别是人egfrviii(seqidno:115)。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:重链可变区(vh),其包含seqidno:85的重链互补决定区(hcdr)1、seqidno:86的hcdr2和seqidno:87的hcdr3;以及轻链可变区(vl),其包含seqidno:89的轻链互补决定区(lcdr)1、seqidno:90的lcdr2和seqidno:91的lcdr3。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)为(来源于)人源化抗体。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)为人源化抗原结合结构域(即人源化抗体的抗原结合结构域)。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh和/或vl为人源化可变区。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh和/或vl包含受体人框架,例如人免疫球蛋白框架或人共有框架。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含seqidno:88的一个或多个重链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vh包含与seqidno:88的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含与seqidno:88的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vh包含与seqidno:88的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vh序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与egfrviii结合的能力。在某些方面,在seqidno:88的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vh包含seqidno:88的氨基酸序列。任选地,vh包含seqidno:88的氨基酸序列,包括该序列的翻译后修饰。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含seqidno:92的一个或多个轻链框架序列(即fr1、fr2、fr3和/或fr4序列)。在一个方面,vl包含与seqidno:92的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vl包含与seqidno:92的氨基酸序列至少约95%相同的氨基酸序列。在一个方面,vl包含与seqidno:92的氨基酸序列至少约98%相同的氨基酸序列。在某些方面,具有至少95%、96%、97%、98%或99%的同一性的vl序列包含相对于参比序列的置换(例如,保守置换)、插入或缺失,但包含该序列的抗体保留与egfrviii结合的能力。在某些方面,在seqidno:92的氨基酸序列中,总共有1至10的氨基酸被置换、插入和/或缺失。在某些方面,置换、插入或缺失发生于cdr之外的区域(即,在fr中)。在一个方面,vl包含seqidno:92的氨基酸序列。任选地,vl包含seqidno:92的氨基酸序列,包括该序列的翻译后修饰。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含与seqidno:88的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列,并且第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含与seqidno:92的氨基酸序列至少约95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,vh包含seqidno:88的氨基酸序列,并且vl包含seqidno:92的氨基酸序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh,其包含seqidno:88的序列;以及vl,其包含seqidno:92的序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含seqidno:88的vh序列以及seqidno:92的vl序列。在另一方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh,其包含seqidno:88的vh的重链cdr序列;以及vl,其包含seqidno:92的vl的轻链cdr序列。在进一步方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含seqidno:88的vh的hcdr1、hcdr2和hcdr3氨基酸序列以及seqidno:92的vl的lcdr1、lcdr2和lcdr3氨基酸序列。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vh包含seqidno:88的vh的重链cdr序列以及与seqidno:88的vh的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vh包含seqidno:88的vh的重链cdr序列以及与seqidno:88的vh的框架序列具有至少95%的序列同一性的框架。在另一方面,vh包含seqidno:88的vh的重链cdr序列以及与seqidno:88的vh的框架序列具有至少98%的序列同一性的框架。在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的vl包含seqidno:92的vl的轻链cdr序列以及与seqidno:92的vl的框架序列具有至少95%、96%、97%、98%或99%的序列同一性的框架。在一个方面,vl包含seqidno:92的vl的轻链cdr序列以及与seqidno:92的vl的框架序列具有至少95%的序列同一性的框架。在另一方面,vl包含seqidno:92的vl的轻链cdr序列以及与seqidno:92的vl的框架序列具有至少98%的序列同一性的框架。在另一方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh序列,其如上面这一节提供的有关egfrviii的任一方面所述;以及vl序列,其如上面这一节提供的有关egfrviii的任一方面所述;但是基于以下序列(按行排序),而不是seqidno:85(hcdr1)、seqidno:86(hcdr2)、seqidno:87(hcdr3)、seqidno:88(vh)、seqidno:89(lcdr1)、seqidno:90(lcdr2)、seqidno:91(lcdr3)和seqidno:92(vl):在一个方面,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)包含:vh序列,其如上面这一节提供的任一方面所述;以及vl序列,其如上面这一节提供的任一方面所述。抗tyrp-1和抗egfrviii抗体本发明还提供了与tyrp-1结合的抗体,该抗体包含:vh序列,如上面这一节提供的关于tyrp-1的方面中的任一项所述;以及vl序列,如上面这一节提供的关于tyrp-1的方面中的任一项所述(例如,与tyrp-1结合的抗体,其包含:重链可变区(vh),其包含seqidno:15的重链互补决定区(hcdr)1、seqidno:16的hcdr2和seqidno:17的hcdr3;以及轻链可变区(vl),其包含seqidno:19的轻链互补决定区(lcdr)1、seqidno:20的lcdr2和seqidno:21的lcdr3;或者与tyrp-1结合的抗体包含:vh,其包含seqidno:18的序列;以及vl,其包含seqidno:22的序列)。本发明还提供了与egfrviii结合的抗体,该抗体包含:vh序列,如上面这一节提供的关于egfrviii的方面中的任一项所述;以及vl序列,如上面这一节提供的关于egfrviii的方面中的任一项所述(例如,与egfrviii结合的抗体,其包含:重链可变区(vh),其包含seqidno:85的重链互补决定区(hcdr)1、seqidno:86的hcdr2和seqidno:87的hcdr3;以及轻链可变区(vl),其包含seqidno:89的轻链互补决定区(lcdr)1、seqidno:90的lcdr2和seqidno:91的lcdr3;或者与tyrp-1结合的抗体,其包含:vh,其包含seqidno:88的序列;以及vl,其包含seqidno:92的序列)。在进一步方面,根据上述方面中任一项所述的与tyrp-1或egfrviii结合的抗体可单独或组合地并入如本文所述的关于与cd3结合的抗体的特征中的任一者(除非明确是指抗cd3抗体,诸如结合序列)。c)荷电改性本发明的(多特异性)抗体可以在其中所包含的fab分子中包含氨基酸置换,这些氨基酸置换特别有效地减少轻链与不匹配重链的错配(bence-jones型副产物),该错配可发生在基于fab的多特异性抗体的产生中,这些多特异性抗体在其结合臂中的一个(或在分子包含多于两个抗原结合fab分子的情况下为多个)结合臂中具有vh/vl交换(同样参见pct公开号wo2015/150447,特别是其中的实例,其全文通过引用并入本文)。所需的(多特异性)抗体与不期望的副产物,特别是在多特异性抗体的结合臂中的一个结合臂中具有vh/vl结构域交换的多特异性抗体中出现的bencejones型副产物的比率,可以通过在ch1和cl结构域中的特定氨基酸位置处引入带相反电荷的荷电氨基酸(有时在本文中称为“荷电改性”)来提高。因此,在一些方面,其中(多特异性)抗体的第一抗原结合结构域和第二抗原结合结构域(以及在存在时的第三抗原结合结构域)均为fab分子,并且在抗原结合结构域中的一者(特别是第一抗原结合结构域)中,fab轻链和fab重链的可变结构域vl和vh彼此替换,i)在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被带正电荷的氨基酸置换(根据kabat编号),并且其中在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被带负电荷的氨基酸置换(根据kabateu索引编号);或者ii)在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被带正电荷的氨基酸置换(根据kabat编号),并且其中在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被带负电荷的氨基酸置换(根据kabateu索引编号)。(多特异性)抗体不包含i)和ii)中提到的两种修饰。具有vh/vl交换的抗原结合结构域的恒定结构域cl和ch1彼此不相互替换(即保持不变)。在一个更具体的方面,i)在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号);或者ii)在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在一个此类方面,在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在进一步方面,在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在一个优选的方面,在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号);并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在一个更优选的方面,在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)置换(根据kabat编号);并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号)。在一个甚至更优选的方面,在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被精氨酸(r)置换(根据kabat编号);并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号)。在优选的方面,如果根据以上方面所述的氨基酸置换在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl和恒定结构域ch1中进行,第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl为κ同种型的。替代性地,根据以上方面所述的氨基酸置换可以在第一抗原结合结构域的恒定结构域cl和恒定结构域ch1中进行,而不是在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl和恒定结构域ch1中进行。在优选的此类方面,第一抗原结合结构域的恒定结构域cl为κ同种型的。因此,在一个方面,在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸或213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在进一步方面,在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在又一方面,在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号);并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。在一个方面,在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)置换(根据kabat编号);并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号)。在另一方面,在第一抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被精氨酸(r)置换(根据kabat编号);并且在第一抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号)。在一个优选的方面,本发明的(多特异性)抗体包含(a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;以及(b)与第二抗原结合的第二抗原结合结构域和任选的第三抗原结合结构域;其中在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域cl中,124位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号)(在一个优选的方面,被赖氨酸(k)或精氨酸(r)独立地置换),123位的氨基酸被赖氨酸(k)、精氨酸(r)或组氨酸(h)独立地置换(根据kabat编号)(在一个优选的方面,被赖氨酸(k)或精氨酸(r)独立地置换);并且在第二抗原结合结构域(以及在存在时的第三抗原结合结构域)的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)或天冬氨酸(d)独立地置换(根据kabateu索引编号)。d)多特异性抗体形式根据本发明所述的(多特异性)抗体可具有各种构型。示例性配置在图1中描述。在优选的方面,(多特异性)抗体中包含的抗原结合结构域为fab分子。在此类方面,第一抗原结合结构域、第二抗原结合结构域、第三抗原结合结构域等在本文中可分别称为第一fab分子、第二fab分子、第三fab分子等。在一个方面,(多特异性)抗体的第一抗原结合结构域与第二抗原结合结构域彼此融合,任选地经由肽连接基融合。在优选的方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子。在一个此类方面,第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合。在另一个此类方面,第二抗原结合结构域在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合。在其中(i)第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合或者(ii)第二抗原结合结构域在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合的方面,另外地,第一抗原结合结构域的fab轻链与第二抗原结合结构域的fab轻链可彼此融合,任选地经由肽连接基融合。可使用具有能够与第二抗原(例如靶细胞抗原,诸如tyrp-1或egfrviii,如图1a、d、g、h、k、l所示)特异性结合的单个抗原结合结构域(诸如fab分子)的(多特异性)抗体,特别是预期在结合高亲和力抗原结合结构域结合后第二抗原发生内化的情况下。在此类情况下,对第二抗原具有特异性的多于一个抗原结合结构域的存在可以增强第二抗原的内化,从而降低其可用性。但是,在其他情况下,具有包含对第二抗原(例如靶细胞抗原,参见如图1b、1c、1e、1f、1i、1j、1m或1n所示的实例)具有特异性的两个或更多个抗原结合结构域(诸如fab分子)的(多特异性)抗体将是有利的,例如有利于优化靶向靶位点或实现靶细胞抗原的交联。因此,在优选的方面,根据本发明所述的(多特异性)抗体包含第三抗原结合结构域。在一个方面,第三抗原结合结构域与第二抗原(例如靶细胞抗原,诸如tyrp-1或egfrviii)结合。在一个方面,第三抗原结合结构域为fab分子。在一个方面,第三抗原结构域与第二抗原结合结构域相同。在一些方面,第三抗原结合结构域和第二抗原结合结构域各自为fab分子,并且第三抗原结合结构域与第二抗原结合结构域相同。因此,在这些方面,第二抗原结合结构域和第三抗原结合结构域包含相同的重链和轻链氨基酸序列,并且具有相同的结构域排列(即,常规结构域或交叉结构域)。此外,在这些方面,第三抗原结合结构域与第二抗原结合结构域包含相同的氨基酸置换(如有)。例如,本文描述为“荷电改性”的氨基酸置换将在第二抗原结合结构域和第三抗原结合结构域中的每个的恒定结构域cl和恒定结构域ch1中进行。替代性地,所述氨基酸置换可以在第一抗原结合结构域的恒定结构域cl和恒定结构域ch1中进行(其在优选的方面也是fab分子),而不是在第二抗原结合结构域和第三抗原结合结构域的恒定结构域cl和恒定结构域ch1中进行。像第二抗原结合结构域一样,第三抗原结合结构域优选地是常规fab分子。但是,还考虑到其中第二抗原结合结构域和第三抗原结合结构域是交叉fab分子(并且第一抗原结合结构域是常规fab分子)的方面。因此,在优选的方面,第二抗原结合结构域和第三抗原结合结构域各自是常规fab分子,并且第一抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域cl和ch1彼此交换/替换的fab分子。在其他方面,第二抗原结合结构域和第三抗原结合结构域各自是交叉fab分子,并且第一抗原结合结构域是常规fab分子。如果存在第三抗原结合结构域,在一个优选的方面,第一抗原结构域与cd3结合,并且第二抗原结合结构域和第三抗原结合结构域与第二抗原(特别是靶细胞抗原,诸如tyrp-1或egfrviii)结合。在优选的方面,本发明的(多特异性)抗体包含由第一亚基和第二亚基组成的fc结构域。fc结构域的第一亚基和第二亚基能够稳定缔合。根据本发明所述的(多特异性)抗体可具有不同的构型,即第一抗原结合结构域、第二抗原结合结构域(与任选的第三抗原结合结构域)可彼此融合并且与fc结构域以不同的方式融合。组分可以直接彼此融合,或优选地通过一种或多种合适的肽连接基融合。当fab分子与fc结构域的亚基的n末端融合时,该融合通常是经由免疫球蛋白铰链区。在一些方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,并且第一抗原结合结构域在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合。在此类方面,第二抗原结合结构域可以在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合,或与fc结构域的亚基中的另一个的n末端融合。在优选的此类方面,第二抗原结合结构域是常规fab分子,并且第一抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域cl和ch1彼此交换/替换的fab分子。在其他此类方面,第二抗原结合结构域是交叉fab分子,并且第一抗原结合结构域是常规fab分子。在一个方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,第一抗原结合结构域在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合,并且第二抗原结合结构域在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合。在一个具体方面,(多特异性)抗体基本上由第一fab分子和第二fab分子、由第一亚基和第二亚基组成的fc结构域和任选的一个或多个肽连接基组成,其中第二fab分子在fab重链的c末端与第一fab分子的fab重链的n末端融合,并且第一fab分子在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合。图1g和1k中示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子)。任选地,第一fab分子的fab轻链和第二fab分子的fab轻链可另外地彼此融合。在另一方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,并且第一抗原结合结构域和第二抗原结合结构域各自在fab重链的c末端与fc结构域的亚基中的一个的n末端融合。在一个具体方面,(多特异性)抗体基本上由第一fab分子和第二fab分子、由第一亚基和第二亚基组成的fc结构域和任选的一个或多个肽连接基组成,其中第一fab分子和第二fab分子各自在fab重链的c末端与fc结构域的亚基中的一个的n末端融合。图1a和1d中示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域是常规fab分子)。第一fab分子和第二fab分子可以直接或通过肽连接基与fc结构域融合。在一个优选的方面,第一fab分子和第二fab分子各自通过免疫球蛋白铰链区与fc结构域融合。在一个具体方面,免疫球蛋白铰链区是人igg1铰链区,特别是其中fc结构域为igg1fc结构域。在一些方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,并且第二抗原结合结构域在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合。在此类方面,第一抗原结合结构域可以在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合或(如上文所述)与fc结构域的亚基中的另一个的n末端融合。在优选的此类方面,所述第二抗原结合结构域是常规fab分子,并且第一抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域cl和ch1彼此交换/替换的fab分子。在其他此类方面,所述第二抗原结合结构域是交叉fab分子,并且第一抗原结合结构域是常规fab分子。在一个方面,第一抗原结合结构域和第二抗原结合结构域各自为fab分子,第二抗原结合结构域在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合,并且第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合。在一个具体方面,(多特异性)抗体基本上由第一fab分子和第二fab分子、由第一亚基和第二亚基组成的fc结构域和任选的一个或多个肽连接基组成,其中第一fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合,并且第二fab分子在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合。图1h和1l中示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域是常规fab分子)。任选地,第一fab分子的fab轻链和第二fab分子的fab轻链可另外地彼此融合。在一些方面,第三抗原结合结构域,特别是第三fab分子,在fab重链的c末端与fc结构域的第一亚基或第二亚基的n末端融合。在优选的此类方面,第二抗原结合结构域和第三抗原结合结构域各自为常规fab分子,并且第一抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域cl和ch1彼此交换/替换的fab分子。在其他此类方面,所述第二抗原结合结构域和第三抗原结合结构域各自为交叉fab分子,并且第一抗原结合结构域是常规fab分子。在一个优选的此类方面,第一抗原结合结构域和第三抗原结合结构域在fab重链的c末端与fc结构域的亚基中的一个的n末端融合,并且第二抗原结合结构域在fab重链的c末端与第一fab分子的fab重链的n末端融合。在一个具体方面,(多特异性)抗体基本上由第一fab分子、第二fab分子和第三fab分子、由第一亚基和第二亚基组成的fc结构域和任选的一个或多个肽连接基组成,其中第二fab分子在fab重链的c末端与第一fab分子的fab重链的n末端融合,并且第一fab分子在fab重链的c末端与fc结构域的第一亚基的n末端融合,并且其中第三fab分子在fab重链的c末端与fc结构域的第二亚基的n末端融合。图1b和1e示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域和第三抗原结合结构域是常规fab分子),并且图1j和1n也示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是常规fab分子,并且第二抗原结合结构域和第三抗原结合结构域是vh/vl交叉fab分子)。第一fab分子和第三fab分子可以直接或通过肽连接基与fc结构域融合。在一个优选的方面,第一fab分子和第三fab分子各自通过免疫球蛋白铰链区与fc结构域融合。在一个具体方面,免疫球蛋白铰链区是人igg1铰链区,特别是其中fc结构域为igg1fc结构域。任选地,第一fab分子的fab轻链和第二fab分子的fab轻链可另外地彼此融合。在另一个此类方面,第二抗原结合结构域和第三抗原结合结构域各自在fab重链的c末端与fc结构域的亚基中的一个的n末端融合,并且第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合。在一个具体方面,(多特异性)抗体基本上由第一fab分子、第二fab分子和第三fab分子、由第一亚基和第二亚基组成的fc结构域和任选的一个或多个肽连接基组成,其中第一fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合,并且第二fab分子在fab重链的c末端与fc结构域的第一亚基的n末端融合,并且其中第三fab分子在fab重链的c末端与fc结构域的第二亚基的n末端融合。图1c和1f示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域和第三抗原结合结构域是常规fab分子),并且图1i和1m也示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是常规fab分子,并且第二抗原结合结构域和第三抗原结合结构域是vh/vl交叉fab分子)。第二fab分子和第三fab分子可以直接或通过肽连接基与fc结构域融合。在一个优选的方面,第二fab分子和第三fab分子各自通过免疫球蛋白铰链区与fc结构域融合。在一个具体方面,免疫球蛋白铰链区是人igg1铰链区,特别是其中fc结构域为igg1fc结构域。任选地,第一fab分子的fab轻链和第二fab分子的fab轻链可另外地彼此融合。在其中fab分子在fab重链的c末端通过免疫球蛋白铰链区与fc结构域的亚基中的每一个的n末端融合的(多特异性)抗体的构型中,两个fab分子、铰链区和fc结构域基本上形成免疫球蛋白分子。在一个优选的方面,免疫球蛋白分子是igg类免疫球蛋白。在一个甚至更优选的方面,免疫球蛋白是igg1亚类免疫球蛋白。在另一方面,免疫球蛋白是igg4亚类免疫球蛋白。在又一个优选的方面,免疫球蛋白是人免疫球蛋白。在其他方面,免疫球蛋白是嵌合免疫球蛋白或人源化免疫球蛋白。在一个方面,免疫球蛋白包含人恒定区,特别是人fc区。在本发明的(多特异性)抗体的一部分中,第一fab分子的fab轻链与第二fab分子的fab轻链彼此融合,任选地经由肽连接基融合。根据第一fab分子和第二fab分子的构型不同,第一fab分子的fab轻链可以在其c末端与第二fab分子的fab轻链的n末端融合,或者第二fab分子的fab轻链可以在其c末端与第一fab分子的fab轻链的n末端融合。第一fab分子和第二fab分子的fab轻链的融合进一步减少了不匹配的fab重链和轻链的错配,并且还减少了表达本发明的(多特异性)抗体中的一部分所需的质粒数量。抗原结合结构域可以直接或通过肽连接基与fc结构域融合,该肽连接基包含一个或多个氨基酸,通常为约2-20个氨基酸。肽连接基是本领域中已知的并在本文中描述的。合适的非免疫原性肽连接基包括例如(g4s)n、(sg4)n、(g4s)n或g4(sg4)n肽连接基。“n”通常为1至10的整数,通常为2至4。在一个方面,所述肽连接基的长度为至少5个氨基酸;在一个方面,长度为5至100个氨基酸;在进一步方面,长度为10至50个氨基酸。在一个方面,所述肽连接基是(gxs)n或(gxs)ngm,其中g=甘氨酸,s=丝氨酸,并且(x=3、n=3、4、5或6,并且m=0、1、2或3)或(x=4、n=2、3、4或5并且m=0、1、2或3);在一个方面,x=4并且n=2或3;在进一步方面,x=4并且n=2。在一个方面,所述肽连接基是(g4s)2。一个特别适合使第一fab分子与第二fab分子的fab轻链彼此融合的肽连接基是(g4s)2。一种适于连接第一fab片段和第二fab片段的fab重链的示例性肽连接基包含序列(d)-(g4s)2(seqidno:118和seqidno:119)。另一种合适的此类连接基包含序列(g4s)4。另外,连接基可包含免疫球蛋白铰链区(的一部分)。特别地,在fab分子与fc结构域亚基的n末端融合的情况下,可以在具有或没有另外的肽连接基的情况下经由免疫球蛋白铰链区或其一部分进行融合。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(即第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与fc结构域亚基共享羧基末端肽键(vl(1)-ch1(1)-ch2-ch3(-ch4));以及这样的多肽,在该多肽中,第二fab分子的fab重链与fc结构域亚基共享羧基末端肽键(vh(2)-ch1(2)-ch2-ch3(-ch4))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,多肽例如通过二硫键共价连接。在某些方面,根据本发明所述的(多特异性)抗体包含包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(即第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链恒定区继而与fc结构域亚基共享羧基末端肽键(vh(1)-cl(1)-ch2-ch3(-ch4));以及这样的多肽,在所述多肽中第二fab分子的fab重链与fc结构域亚基共享羧基末端肽键(vh(2)-ch1(2)-ch2-ch3(-ch4))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,多肽例如通过二硫键共价连接。在一些方面,(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与fc结构域亚基共享羧基末端肽键(vl(1)-ch1(1)-vh(2)-ch1(2)-ch2-ch3(-ch4))。在其他方面,(多特异性)抗体包含这样的多肽,在该多肽中,第二fab分子的fab重链与第一fab分子的fab轻链可变区共享羧基末端肽键,该fab轻链可变区继而与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该第一fab分子的fab重链恒定区继而与fc结构域亚基共享羧基末端肽键(vh(2)-ch1(2)-vl(1)-ch1(1)-ch2-ch3(-ch4))。在这些方面中的一些方面,(多特异性)抗体进一步包含第一fab分子的交叉fab轻链多肽,其中第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1)),并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在这些方面中的其他方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键,该fab轻链恒定区继而与第二fab分子的fab轻链多肽共享羧基末端肽键(vh(1)-cl(1)-vl(2)-cl(2));或者在适当的情况下包含这样的多肽,在该多肽中,第二fab分子的fab轻链多肽与第一fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vl(2)-cl(2)-vh(1)-cl(1))。根据这些方面所述的(多特异性)抗体,可进一步包含:(i)fc结构域亚基多肽(ch2-ch3(-ch4)),或(ii)多肽,其中第三fab分子的fab重链与fc结构域亚基共享羧基末端肽键(vh(3)-ch1(3)-ch2-ch3(-ch4))并且与第三fab分子的fab轻链多肽共享羧基末端肽键(vl(3)-cl(3))。在某些方面,多肽例如通过二硫键共价连接。在一些方面,(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链恒定区继而与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与fc结构域亚基共享羧基末端肽键(vh(1)-cl(1)-vh(2)-ch1(2)-ch2-ch3(-ch4))。在其他方面,(多特异性)抗体包含这样的多肽,在该多肽中,第二fab分子的fab重链与第一fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该第一fab分子的fab轻链恒定区继而与fc结构域亚基共享羧基末端肽键(vh(2)-ch1(2)-vh(1)-cl(1)-ch2-ch3(-ch4))。在这些方面中的一些方面,(多特异性)抗体进一步包含第一fab分子的交叉fab轻链多肽,其中第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1)),并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在这些方面中的其他方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键,该fab重链恒定区与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(1)-ch1(1)-vl(2)-cl(2));或者在适当的情况下包含这样的多肽,在该多肽中,第二fab分子的fab轻链多肽与第一fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vl(2)-cl(2)-vh(1)-cl(1))。根据这些方面所述的(多特异性)抗体,可进一步包含:(i)fc结构域亚基多肽(ch2-ch3(-ch4)),或(ii)多肽,其中第三fab分子的fab重链与fc结构域亚基共享羧基末端肽键(vh(3)-ch1(3)-ch2-ch3(-ch4))并且与第三fab分子的fab轻链多肽共享羧基末端肽键(vl(3)-cl(3))。在某些方面,多肽例如通过二硫键共价连接。在某些方面,(多特异性)抗体不包含fc结构域。在优选的此类方面,所述第二抗原结合结构域以及在存在时的第三抗原结合结构域各自为常规fab分子,并且第一抗原结合结构域是如本文所述的交叉fab分子,即其中fab重链和轻链的可变结构域vh和vl或恒定结构域cl和ch1彼此交换/替换的fab分子。在其他此类方面,所述第二抗原结合结构域以及在存在时的第三抗原结合结构域各自为交叉fab分子,并且第一抗原结合结构域是常规fab分子。在一个此类方面,(多特异性)抗体基本上由第一抗原结合结构域和第二抗原结合结构域和任选的一个或多个肽连接基组成,其中第一抗原结合结构域和第二抗原结合结构域均为fab分子,并且第二抗原结合结构域在fab重链的c末端与第一抗原结合结构域的fab重链的n末端融合。图1o和1s示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子并且第二抗原结合结构域是常规fab分子)。在另一个此类方面,(多特异性)抗体基本上由第一抗原结合结构域和第二抗原结合结构域和任选的一个或多个肽连接基组成,其中第一抗原结合结构域和第二抗原结合结构域均为fab分子,并且第一抗原结合结构域在fab重链的c末端与第二抗原结合结构域的fab重链的n末端融合。图1p和1t中示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域是常规fab分子)。在一些方面,第二fab分子在fab重链的c末端与第一fab分子的fab重链的n末端融合,并且(多特异性)抗体进一步包含第三抗原结合结构域,特别是第三fab分子,其中所述第三fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合。在某些此类方面,(多特异性)抗体基本上由第一fab分子、第二fab分子和第三fab分子和任选的一个或多个肽连接基组成,其中第二fab分子在fab重链的c末端与第一fab分子的fab重链的n末端融合,并且第三fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合。图1q和1u示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域和第三抗原结合结构域各自为常规fab分子),或者图1x和1z也示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是常规fab分子,并且第二抗原结合结构域和第三抗原结合结构域各自为vh/vl交叉fab分子)。在一些方面,第一fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合,并且(多特异性)抗体进一步包含第三抗原结合结构域,特别是第三fab分子,其中所述第三fab分子在fab重链的n末端与第二fab分子的fab重链的c末端融合。在某些此类方面,(多特异性)抗体基本上由第一fab分子、第二fab分子和第三fab分子和任选的一个或多个肽连接基组成,其中第一fab分子在fab重链的c末端与第二fab分子的fab重链的n末端融合,并且第三fab分子在fab重链的n末端与第二fab分子的fab重链的c末端融合。图1r和1v示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是vh/vl交叉fab分子,并且第二抗原结合结构域和第三抗原结合结构域各自为常规fab分子),或者图1w和1y也示意性地描绘了此类构型(在这些实例中,第一抗原结合结构域是常规fab分子,并且第二抗原结合结构域和第三抗原结合结构域各自为vh/vl交叉fab分子)。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第二fab分子的fab重链与第一fab分子的fab轻链可变区共享羧基末端肽键,该fab轻链可变区继而与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换)(vh(2)-ch1(2)-vl(1)-ch1(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与第二fab分子的fab重链共享羧基末端肽键(vl(1)-ch1(1)-vh(2)-ch1(2))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第二fab分子的fab重链与第一fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区继而与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换)(vh(2)-ch1(2)-vh(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链恒定区继而与第二fab分子的fab重链共享羧基末端肽键(vh(1)-cl(1)-vh(2)-ch1(2))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第三fab分子的fab重链与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与第一fab分子的fab轻链可变区共享羧基末端肽键,该fab轻链可变区继而与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换)(vh(3)-ch1(3)-vh(2)-ch1(2)-vl(1)-ch1(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在一些方面,(多特异性)抗体进一步包含第三fab分子的fab轻链多肽(vl(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第三fab分子的fab重链与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与第一fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区继而与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换)(vh(3)-ch1(3)-vh(2)-ch1(2)-vh(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在一些方面,(多特异性)抗体进一步包含第三fab分子的fab轻链多肽(vl(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与第三fab分子的fab重链共享羧基末端肽键(vl(1)-ch1(1)-vh(2)-ch1(2)-vh(3)-ch1(3))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在一些方面,(多特异性)抗体进一步包含第三fab分子的fab轻链多肽(vl(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab重链可变区与第一fab分子的fab轻链恒定区共享羧基末端肽键(即,第一fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链恒定区继而与第二fab分子的fab重链共享羧基末端肽键,该第二fab分子的fab重链继而与第三fab分子的fab重链共享羧基末端肽键(vh(1)-cl(1)-vh(2)-ch1(2)-vh(3)-ch1(3))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第一fab分子的fab轻链可变区与第一fab分子的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1))并且与第二fab分子的fab轻链多肽共享羧基末端肽键(vl(2)-cl(2))。在一些方面,(多特异性)抗体进一步包含第三fab分子的fab轻链多肽(vl(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab重链与第二fab分子的fab轻链可变区共享羧基末端肽键,该fab轻链可变区继而与第二fab分子的fab重链恒定区共享羧基末端肽键(即,第二fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与第三fab分子的fab轻链可变区共享羧基末端肽键,该第三fab分子的fab轻链可变区继而与第三fab分子的fab重链恒定区共享羧基末端肽键(即,第三fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换)(vh(1)-ch1(1)-vl(2)-ch1(2)-vl(3)-ch1(3))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第二fab分子的fab重链可变区与第二fab分子的fab轻链可变区共享羧基末端肽键(vh(2)-cl(2)),并且与第一fab分子的fab轻链多肽共享羧基末端肽键(vl(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第三fab分子的fab重链可变区与第三fab分子的fab轻链可变区共享羧基末端肽键(vh(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第一fab分子的fab重链与第二fab分子的fab重链可变区共享羧基末端肽键,该fab重链可变区继而与第二fab分子的fab轻链可变区共享羧基末端肽键(即,第二fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链可变区继而与第三fab分子的fab重链可变区共享羧基末端肽键,该第三fab分子的fab重链可变区继而与第三fab分子的fab轻链可变区共享羧基末端肽键(即,第三fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换)(vh(1)-ch1(1)-vh(2)-cl(2)-vh(3)-cl(3))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第二fab分子的fab轻链可变区与第二fab分子的fab重链恒定区共享羧基末端肽键(vl(2)-ch1(2)),并且与第一fab分子的fab轻链多肽共享羧基末端肽键(vl(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第三fab分子的fab轻链可变区与第三fab分子的fab重链恒定区共享羧基末端肽键(vl(3)-ch1(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第三fab分子的fab轻链可变区与第三fab分子的fab重链恒定区共享羧基末端肽键(即,第三fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该fab重链恒定区继而与第二fab分子的fab轻链可变区共享羧基末端肽键,该第二fab分子的fab轻链可变区继而与第二fab分子的fab重链恒定区共享羧基末端肽键(即,第二fab分子包含交叉fab重链,其中重链可变区被轻链可变区替换),该第二fab分子的fab重链恒定区继而与第一fab分子的fab重链共享羧基末端肽键(vl(3)-ch1(3)-vl(2)-ch1(2)-vh(1)-ch1(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第二fab分子的fab重链可变区与第二fab分子的fab轻链可变区共享羧基末端肽键(vh(2)-cl(2)),并且与第一fab分子的fab轻链多肽共享羧基末端肽键(vl(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第三fab分子的fab重链可变区与第三fab分子的fab轻链可变区共享羧基末端肽键(vh(3)-cl(3))。在某些方面,根据本发明所述的(多特异性)抗体包含这样的多肽,在该多肽中,第三fab分子的fab重链可变区与第三fab分子的fab轻链可变区共享羧基末端肽键(即,第三fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该fab轻链可变区继而与第二fab分子的fab重链可变区共享羧基末端肽键,该第二fab分子的fab重链可变区继而与第二fab分子的fab轻链可变区共享羧基末端肽键(即,第二fab分子包含交叉fab重链,其中重链恒定区被轻链恒定区替换),该第二fab分子的fab轻链可变区继而与第一fab分子的fab重链共享羧基末端肽键(vh(3)-cl(3)-vh(2)-cl(2)-vh(1)-ch1(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第二fab分子的fab轻链可变区与第二fab分子的fab重链恒定区共享羧基末端肽键(vl(2)-ch1(2)),并且与第一fab分子的fab轻链多肽共享羧基末端肽键(vl(1)-cl(1))。在一些方面,(多特异性)抗体进一步包含这样的多肽,在该多肽中,第三fab分子的fab轻链可变区与第三fab分子的fab重链恒定区共享羧基末端肽键(vl(3)-ch1(3))。在一个方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;以及;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域,其中第二抗原结合结构域是(常规)fab分子;c)由第一亚基和第二亚基组成的fc结构域;其中(i)在a)中的第一抗原结合结构域在fab重链的c末端与在b)中的第二抗原结合结构域的fab重链的n末端融合,并且在b)中的第二抗原结合结构域在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合,或者(ii)在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;以及;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子;并且c)由第一亚基和第二亚基组成的fc结构域;其中(i)在a)中的第一抗原结合结构域在fab重链的c末端与在b)中的第二抗原结合结构域的fab重链的n末端融合,并且在b)中的第二抗原结合结构域和在b)中的第三抗原结合结构域各自在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合;或者(ii)在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域各自在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在另一方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh或恒定结构域cl和ch1彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;以及;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域,其中第二抗原结合结构域是(常规)fab分子;c)由第一亚基和第二亚基组成的fc结构域;其中(i)在a)中的第一抗原结合结构域和在b)中的第二抗原结合结构域各自在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在根据本发明所述的(多特异性)抗体的所有不同构型中,如本文所述的氨基酸置换(“荷电改性”)在存在时可以在第二抗原结合结构域/fab分子和(如果存在)第三抗原结合结构域/fab分子的ch1和cl结构域中,或者在第一抗原结合结构域/fab分子的ch1和cl结构域中。优选地,它们在第二抗原结合结构域/fab分子和(如果存在)第三抗原结合结构域/fab分子的ch1和cl结构域中。根据本发明的概念,如果如本文所述的氨基酸置换在第二抗原结合结构域/fab分子(以及在存在时的第三抗原结合结构域/fab分子)中进行,则在第一抗原结合结构域/fab分子中不进行此类氨基酸置换。相反,如果如本文所述的氨基酸置换在第一抗原结合结构域/fab分子中进行,则在第二抗原结合结构域/fab分子(以及在存在时的第三抗原结合结构域/fab分子)中不进行此类氨基酸置换。氨基酸置换优选地在包含fab分子的(多特异性)抗体中进行,在该fab分子中,fab轻链和fab重链的可变结构域vl和vh1彼此替换。在根据本发明所述的(多特异性)抗体的优选的方面,特别是其中如本文所述的氨基酸置换在第二抗原结合结构域/fab分子(以及在存在时的第三抗原结合结构域/fab分子)中进行的情况下,第二fab分子(以及在存在时的第三fab分子)的恒定结构域cl为κ同种型的。在根据本发明所述的(多特异性)抗体的其他方面,特别是其中如本文所述的氨基酸置换在第一抗原结合结构域/fab分子中进行的情况下,第一抗原结合结构域的恒定结构域cl/fab分子为κ同种型的。在一些方面,第二抗原结合结构域/fab分子(以及在存在时的第三抗原结合结构域/fab分子)和第一抗原结合结构域的恒定结构域cl/fab分子的恒定结构域cl为κ同种型的。在一个方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域,其中第二抗原结合结构域是(常规)fab分子;c)由第一亚基和第二亚基组成的fc结构域;其中在b)中的第二抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中(i)在a)中的第一抗原结合结构域在fab重链的c末端与在b)中的第二抗原结合结构域的fab重链的n末端融合,并且在b)中的第二抗原结合结构域在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合,或者(ii)在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子;并且c)由第一亚基和第二亚基组成的fc结构域;其中在b)中的第二抗原结合结构域和b)中的第三抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域和b)中的第三抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中(i)在a)中的第一抗原结合结构域在fab重链的c末端与在b)中的第二抗原结合结构域的fab重链的n末端融合,并且在b)中的第二抗原结合结构域和在b)中的第三抗原结合结构域各自在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合;或者(ii)在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域各自在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在另一方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;b)与第二抗原(特别是靶细胞抗原,更特别是tyrp-1或egfrviii)结合的第二抗原结合结构域,其中第二抗原结合结构域是(常规)fab分子;c)由第一亚基和第二亚基组成的fc结构域;其中在b)中的第二抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中在a)中的第一抗原结合结构域和b)中的第二抗原结合结构域在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。根据以上方面中的任一方面,(多特异性)抗体的组分(例如fab分子、fc结构域)可以直接或通过各种连接基融合,这些连接基特别是包含一个或多个氨基酸(通常约2-20个氨基酸)的肽连接基,如本文所述或本领域中已知。合适的非免疫原性肽连接基包括例如(g4s)n、(sg4)n、(g4s)n或g4(sg4)n肽连接基,其中n通常为1至10(通常为2至4)的整数。在一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)结合cd3的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;b)与tyrp-1结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子并且包含:重链可变区(vh),其包含seqidno:15的重链互补决定区(hcdr)1、seqidno:16的hcdr2和seqidno:17的hcdr3;以及轻链可变区(vl),其包含seqidno:19的轻链互补决定区(lcdr)1、seqidno:20的lcdr2和seqidno:21的lcdr3;c)由第一亚基和第二亚基组成的fc结构域;其中其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中进一步地,在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域彼此在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在又一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换并且包含:重链可变区,其包含seqidno:7的氨基酸序列;以及轻链可变区,其包含seqidno:11的氨基酸序列;b)与tyrp-1结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子并且包含:重链可变区,其包含seqidno:18的氨基酸序列;以及轻链可变区,其包含seqidno:22的氨基酸序列;c)由第一亚基和第二亚基组成的fc结构域;其中其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中进一步地,在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域彼此在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)结合cd3的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换,并且包含:重链可变区(vh),其包含seqidno:2的重链互补决定区(hcdr)1、seqidno:3的hcdr2和seqidno:5的hcdr3;以及轻链可变区(vl),其包含seqidno:8的轻链互补决定区(lcdr)1、seqidno:9的lcdr2和seqidno:10的lcdr3;b)与egfrviii结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子并且包含:重链可变区(vh),其包含seqidno:85的重链互补决定区(hcdr)1、seqidno:86的hcdr2和seqidno:87的hcdr3;以及轻链可变区(vl),其包含seqidno:89的轻链互补决定区(lcdr)1、seqidno:90的lcdr2和seqidno:91的lcdr3;c)由第一亚基和第二亚基组成的fc结构域;其中其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中进一步地,在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域彼此在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在又一个优选的方面,本发明提供了一种(多特异性)抗体,该抗体包含:a)与cd3结合的第一抗原结合结构域,其中第一抗原结合结构域为fab分子,其中fab轻链和fab重链的可变结构域vl和vh彼此替换并且包含:重链可变区,其包含seqidno:7的氨基酸序列;以及轻链可变区,其包含seqidno:11的氨基酸序列;b)与egfrviii结合的第二抗原结合结构域和第三抗原结合结构域,其中第二抗原结合结构域和第三抗原结合结构域各自为(常规)fab分子并且包含:重链可变区,其包含seqidno:88的氨基酸序列;以及轻链可变区,其包含seqidno:92的氨基酸序列;c)由第一亚基和第二亚基组成的fc结构域;其中其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域cl中,124位的氨基酸被赖氨酸(k)置换(根据kabat编号),并且123位的氨基酸被赖氨酸(k)或精氨酸(r)置换(根据kabat编号)(最优选地被精氨酸(r)置换),并且其中在b)中的第二抗原结合结构域和第三抗原结合结构域的恒定结构域ch1中,147位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号),并且213位的氨基酸被谷氨酸(e)置换(根据kabateu索引编号);并且其中进一步地,在b)中的第二抗原结合结构域在fab重链的c末端与在a)中的第一抗原结合结构域的fab重链的n末端融合,并且在a)中的第一抗原结合结构域和在b)中的第三抗原结合结构域彼此在fab重链的c末端与在c)中的fc结构域的亚基中的一个的n末端融合。在一个方面,根据本发明的这些方面,在fc结构域的第一亚基中,366位的苏氨酸残基被替换为色氨酸残基(t366w),并且在fc结构域的第二亚基中,407位的酪氨酸残基被替换为缬氨酸残基(y407v),并且任选地,366位的苏氨酸残基被替换为丝氨酸残基(t366s)并且368位的亮氨酸残基被替换为丙氨酸残基(l368a)(根据kabateu索引编号)。在进一步方面,根据本发明的这些方面,在fc结构域的第一亚基中,另外地354位的丝氨酸残基被替换为半胱氨酸残基(s354c)或者356位的谷氨酸残基被替换为半胱氨酸残基(e356c)(特别是,354位的丝氨酸残基被替换为半胱氨酸残基),并且在fc结构域的第二亚基中,另外地349位的酪氨酸残基被替换为半胱氨酸残基(y349c)(根据kabateu索引编号)。在根据本发明的这些方面中的又一方面,在fc结构域的第一亚基和第二亚基中的每个中,234位的亮氨酸残基被替换为丙氨酸残基(l234a),235位的亮氨酸残基被替换为丙氨酸残基(l235a),并且329位的脯氨酸残基被替换为甘氨酸残基(p329g)(根据kabateu索引编号)。在根据本发明的这些方面中的又一方面,fc结构域是人igg1fc结构域。在一个优选的具体方面,(多特异性)抗体包含:多肽,其包含与seqidno:23的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽,其包含与seqidno:24的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽(特别是两个多肽),其包含与seqidno:25的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;以及多肽,其包含与seqidno:27的序列至少95%、96%、97%、98%或99%相同的氨基酸序列。在另一优选的具体方面,(多特异性)抗体包含:多肽,其包含seqidno:23的氨基酸序列;多肽,其包含seqidno:24的氨基酸序列;多肽(特别是两个多肽),其包含seqidno:25的氨基酸序列;以及多肽,其包含seqidno:27的氨基酸序列。在一个方面,本发明提供了一种与cd3和tyrp-1结合的(多特异性)抗体,该抗体包含:多肽,其包含与seqidno:23的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽,其包含与seqidno:24的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽(特别是两个多肽),其包含与seqidno:25的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;以及多肽,其包含与seqidno:27的序列至少95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,本发明提供了一种与cd3和tyrp-1结合的(多特异性)抗体,该抗体包含:多肽,其包含seqidno:23的氨基酸序列;多肽,其包含seqidno:24的氨基酸序列;多肽(特别是两个多肽),其包含seqidno:25的氨基酸序列;以及多肽,其包含seqidno:27的氨基酸序列。在另一优选的具体方面,(多特异性)抗体包含:多肽,其包含与seqidno:109的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽,其包含与seqidno:110的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽(特别是两个多肽),其包含与seqidno:111的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;以及多肽,其包含与seqidno:27的序列至少95%、96%、97%、98%或99%相同的氨基酸序列。在另一优选的具体方面,(多特异性)抗体包含:多肽,其包含seqidno:109的氨基酸序列;多肽,其包含seqidno:110的氨基酸序列;多肽(特别是两个多肽),其包含seqidno:111的氨基酸序列;以及多肽,其包含seqidno:27的氨基酸序列。在一个方面,本发明提供了一种与cd3和egfrviii结合的(多特异性)抗体,该抗体包含:多肽,其包含与seqidno:109的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽,其包含与seqidno:110的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;多肽(特别是两个多肽),其包含与seqidno:111的序列至少95%、96%、97%、98%或99%相同的氨基酸序列;以及多肽,其包含与seqidno:27的序列至少95%、96%、97%、98%或99%相同的氨基酸序列。在一个方面,本发明提供了一种与cd3和egfrviii结合的(多特异性)抗体,该抗体包含:多肽,其包含seqidno:109的氨基酸序列;多肽,其包含seqidno:110的氨基酸序列;多肽(特别是两个多肽),其包含seqidno:111的氨基酸序列;以及多肽,其包含seqidno:27的氨基酸序列。8.fc结构域变体在优选的方面,本发明的(多特异性)抗体包含由第一亚基和第二亚基组成的fc结构域。(多特异性)抗体的fc结构域由包含免疫球蛋白分子的重链结构域的一对多肽链组成。例如,免疫球蛋白g(igg)分子的fc结构域是二聚体,该二聚体的每个亚基包含ch2和ch3igg重链恒定结构域。fc结构域的两个亚基能够彼此稳定缔合。在一个方面,本发明的(多特异性)抗体包含不超过一个fc结构域。在一个方面,(多特异性)抗体的fc结构域是iggfc结构域。在一个优选的方面,fc结构域是igg1fc结构域。在另一方面,fc结构域是igg4fc结构域。在一个更具体的方面,fc结构域是以下igg4fc结构域,其包含s228位的氨基酸置换(kabateu索引编号),特别是氨基酸置换s228p。该氨基酸置换减少igg4抗体的体内fab臂交换(参见stubenrauch等人,drugmetabolismanddisposition38,84-91(2010))。在又一个优选的方面,fc结构域是人fc结构域。在一个甚至更优选的方面,fc结构域是人igg1fc结构域。人igg1fc的示例性序列在seqidno:117中给出。a)促进异源二聚化的fc结构域修饰根据本发明所述的(多特异性)抗体包含不同的抗原结合结构域,这些抗原结合结构域可以与fc结构域的两个亚基中的一个或另一个融合,因此fc结构域的两个亚基通常包含在两个不相同的多肽链中。这些多肽的重组共表达和随后的二聚化导致了两种多肽的几种可能的组合。为了在重组生产中提高(多特异性)抗体的产率和纯度,在(多特异性)抗体的fc结构域中引入促进所需多肽的缔合的修饰将是有利的。因此,在优选的方面,根据本发明所述的(多特异性)抗体的fc结构域包含促进fc结构域的第一亚基和第二亚基进行缔合的修饰。人iggfc结构域的两个亚基之间最广泛的蛋白质间相互作用位点在fc结构域的ch3结构域中。因此,在一个方面,所述修饰在fc结构域的ch3结构域中。存在几种对fc结构域的ch3结构域进行修饰以实施异源二聚化的方法,这些方法例如在wo96/27011、wo98/050431、ep1870459、wo2007/110205、wo2007/147901、wo2009/089004、wo2010/129304、wo2011/90754、wo2011/143545、wo2012058768、wo2013157954、wo2013096291中详细描述。通常,在所有此类方法中,fc结构域的第一亚基的ch3结构域和fc结构域的第二亚基的ch3结构域均以互补的方式经过工程化改造,使得每个ch3结构域(或包含其的重链)可以不再与其自身同源二聚化,而是被迫与互补工程化改造的其他ch3结构域异源二聚化(使得第一和第二ch3结构域异源二聚化并且在两个第一或两个第二ch3结构域之间不形成同源二聚体)。这些用于实现改善的重链异源二聚化的不同方法被认为是与(多特异性)抗体中的重链-轻链修饰(例如在一个结合臂中进行vh和vl交换/替换并且在ch1/cl界面中引入带有相反电荷的荷电氨基酸的置换)组合的不同替代方案,所述不同替代方案减少重链/轻链错配和bencejones型副产物。在一个具体方面,所述促进fc结构域的第一亚基和第二亚基缔合的修饰是所谓的“杵臼结构”修饰,该修饰包含在fc结构域的两个亚基中的一个亚基中的“杵”修饰以及在fc结构域的两个亚基中的另一个亚基中的“臼”修饰。杵臼结构技术描述于例如us5,731,168;us7,695,936;ridgway等人,proteng9,617-621(1996)和carter,jimmunolmeth248,7-15(2001)中。通常,该方法涉及在第一多肽的界面处引入凸起(“杵”)并在第二多肽的界面中引入相应的空腔(“臼”),使得所述凸起可以定位在所述空腔中,以便促进异二聚体的形成并阻碍同源二聚体的形成。凸起是通过用较大侧链(例如酪氨酸或色氨酸)取代来自第一多肽的界面的小氨基酸侧链而构建的。具有与凸起相同或相似大小的补偿空腔是通过用较小的氨基酸侧链(例如丙氨酸或苏氨酸)取代大氨基酸侧链而在第二多肽的界面中创建的。因此,在一个优选的方面,在(多特异性)抗体的fc结构域的第一亚基的ch3结构域中,氨基酸残基被替换为具有较大侧链体积的氨基酸残基,从而在第一亚基的ch3结构域内生成突起,该突起可定位在第二亚基的ch3结构域内的空腔中;并且在fc结构域的第二亚基的ch3结构域中,氨基酸残基被替换为具有较小侧链体积的氨基酸残基,从而在第二亚基的ch3结构域内生成空腔,第一亚基的ch3结构域内的突起可定位在该空腔内。优选地,所述具有较大侧链体积的氨基酸残基选自由精氨酸(r)、苯丙氨酸(f)、酪氨酸(y)和色氨酸(w)组成的组。优选地,所述具有较小侧链体积的氨基酸残基选自由丙氨酸(a)、丝氨酸(s)、苏氨酸(t)和缬氨酸(v)组成的组。凸起和空腔可以通过改变编码多肽的核酸来制备,例如通过位点特异性诱变或通过肽合成。在一个具体方面,在fc结构域的第一亚基(“杵”亚基)(的ch3结构域中),366位的苏氨酸残基被替换为色氨酸残基(t366w),并且在fc结构域的第二亚基(“臼”亚基)(的ch3结构域)中,407位的酪氨酸残基被替换为缬氨酸残基(y407v)。在一个方面,在fc结构域的第二亚基中,另外地366位的苏氨酸残基被替换为丝氨酸残基(t366s)并且368位的亮氨酸残基被替换为丙氨酸残基(l368a)(根据kabateu索引编号)。在又一方面,在fc结构域的第一亚基中,另外地354位的丝氨酸残基被替换为半胱氨酸残基(s354c)或者356位的谷氨酸残基被替换为半胱氨酸残基(e356c)(特别是354位的丝氨酸残基被替换为半胱氨酸残基);并且在fc结构域的第二亚基中,另外地349位的酪氨酸残基被替换为半胱氨酸残基(y349c)(根据kabateu索引编号)。引入这两个半胱氨酸残基导致在fc结构域的两个亚基之间形成二硫桥,从而进一步稳定所述二聚体(carter,jimmunolmethods248,7-15(2001))。在一个优选的方面,fc结构域的第一亚基包含氨基酸置换s354c和t366w,并且fc结构域的第二亚基包含氨基酸置换y349c、t366s、l368a和y407v(根据kabateu索引编号)。在一个优选的方面,与cd3结合的抗原结合结构域与fc结构域的第一亚基(包含“杵”修饰)融合(任选地经由与第二抗原融合的第二抗原结合结构域融合,和/或经由肽连接基融合)。不希望受理论的束缚,结合cd3的抗原结合结构域与fc结构域的包含杵的亚基的融合将(进一步)最大程度减少包含与cd3结合的两个抗原结合结构域的抗体的生成(两个包含杵的多肽的空间位阻)。用于实施异源二聚化的其他ch3修饰技术被设想为根据本发明的替代方案,并且描述于例如wo96/27011、wo98/050431、ep1870459、wo2007/110205、wo2007/147901、wo2009/089004、wo2010/129304、wo2011/90754、wo2011/143545、wo2012/058768、wo2013/157954、wo2013/096291中。在一个方面,替代性地使用ep1870459中描述的异源二聚化方法。该方法基于在fc结构域的两个亚基之间的ch3/ch3结构域界面中的特定氨基酸位置处引入带有相反电荷的荷电氨基酸。本发明的(多特异性)抗体的一个特定方面是在(fc结构域的)两个ch3结构域中的一个中的氨基酸突变r409d和k370e,以及在fc结构域的ch3结构域中的另一个中的氨基酸突变d399k和e357k(根据kabateu索引编号)。在另一方面,本发明的(多特异性)抗体包含在fc结构域的第一亚基的ch3结构域中的氨基酸突变t366w以及在fc结构域的第二亚基的ch3结构域中的氨基酸突变t366s、l368a、y407v,并且另外包含在fc结构域的第一亚基的ch3结构域中的氨基酸突变r409d和k370e以及在fc结构域的第二亚基的ch3结构域中的氨基酸突变d399k和e357k(根据kabateu索引编号)。在另一方面,本发明的(多特异性)抗体包含在fc结构域的第一亚基的ch3结构域中的氨基酸突变s354c、t366w以及在fc结构域的第二亚基的ch3结构域中的氨基酸突变y349c、t366s、l368a、y407v,或者所述(多特异性)抗体包含在fc结构域的第一亚基的ch3结构域中的氨基酸突变y349c、t366w以及在fc结构域的第二亚基的ch3结构域中的氨基酸突变s354c、t366s、l368a、y407v,并且另外包含在fc结构域的第一亚基的ch3结构域中的氨基酸突变r409d和k370e以及在fc结构域的第二亚基的ch3结构域中的氨基酸突变d399k和e357k(根据kabateu索引编号)。在一个方面,替代性地使用wo2013/157953中描述的异源二聚化方法。在一个方面,第一ch3结构域包含氨基酸突变t366k,并且第二ch3结构域包含氨基酸突变l351d(根据kabateu索引编号)。在进一步方面,第一ch3结构域包含进一步的氨基酸突变l351k。在进一步方面,第二ch3结构域进一步包含选自y349e、y349d和l368e的氨基酸突变(特别是l368e)(根据kabateu索引编号)。在一个方面,替代性地使用wo2012/058768中描述的异源二聚化方法。在一个方面,第一ch3结构域包含氨基酸突变l351y、y407a,并且第二ch3结构域包含氨基酸突变t366a、k409f。在进一步方面,第二ch3结构域包含进一步的t411、d399、s400、f405、n390或k392位的氨基酸突变,例如选自:a)t411n、t411r、t411q、t411k、t411d、t411e或t411w;b)d399r、d399w、d399y或d399k;c)s400e、s400d、s400r或s400k;d)f405i、f405m、f405t、f405s、f405v或f405w;e)n390r、n390k或n390d;f)k392v、k392m、k392r、k392l、k392f或k392e(根据kabateu索引编号)。在进一步方面,第一ch3结构域包含氨基酸突变l351y、y407a,并且第二ch3结构域包含氨基酸突变t366v、k409f。在进一步方面,第一ch3结构域包含氨基酸突变y407a,并且第二ch3结构域包含氨基酸突变t366a、k409f。在进一步方面,第二ch3结构域进一步包含氨基酸突变k392e、t411e、d399r和s400r(根据kabateu索引编号)。在一个方面,替代性地使用wo2011/143545中描述的异源二聚化方法,例如在选自由368和409组成的组的位置处进行氨基酸修饰(根据kabateu索引编号)。在一个方面,替代性地使用wo2011/090762中描述的异源二聚化方法,该异源二聚化方法也使用上述杵臼结构技术。在一个方面,第一ch3结构域包含氨基酸突变t366w,并且第二ch3结构域包含氨基酸突变y407a。在一个方面,第一ch3结构域包含氨基酸突变t366y,并且第二ch3结构域包含氨基酸突变y407t(根据kabateu索引编号)。在一个方面,(多特异性)抗体或其fc结构域属于igg2亚类,并且替代性地使用在wo2010/129304中描述的异源二聚化方法。在另一方面,促进fc结构域的第一亚基和第二亚基缔合的修饰包括介导静电转向效应的修饰,例如如在pct公开wo2009/089004中所述。通常,该方法涉及用带电荷的氨基酸残基取代两个fc结构域亚基的界面处的一个或多个氨基酸残基,使得同源二聚体形成变得在静电上不利,但异源二聚化在静电上有利。在一个此类方面,第一ch3结构域包含带负电荷的氨基酸与k392或n392的氨基酸置换(例如谷氨酸(e)或天冬氨酸(d),特别是k392d或n392d),并且第二ch3结构域包含带正电荷的氨基酸与d399、e356、d356或e357的氨基酸置换(例如赖氨酸(k)或精氨酸(r),特别是d399k、e356k、d356k或e357k,并且更特别是d399k和e356k)。在进一步方面,第一ch3结构域进一步包含带负电荷的氨基酸与k409或r409的氨基酸置换(例如谷氨酸(e)或天冬氨酸(d),特别是k409d或r409d)。在进一步方面,第一ch3结构域进一步或替代性地包含带负电荷的氨基酸与k439和/或k370的氨基酸置换(例如谷氨酸(e)或天冬氨酸(d))(根据kabateu索引进行所有编号)。在又一方面,替代性地使用wo2007/147901中描述的异源二聚化方法。在一个方面,第一ch3结构域包含氨基酸突变k253e、d282k和k322d,并且第二ch3结构域包含氨基酸突变d239k、e240k和k292d(根据kabateu索引编号)。在又一方面,可替代性地使用wo2007/110205中描述的异源二聚化方法。在一个方面,fc结构域的第一亚基包含氨基酸置换k392d和k409d,并且fc结构域的第二亚基包含氨基酸置换d356k和d399k(根据kabateu索引编号)。b)降低fc受体结合和/或效应子功能的fc结构域修饰fc结构域赋予(多特异性)抗体有利的药代动力学性质,包括有助于靶组织中的良好积聚的长血清半衰期和有利的组织-血液分配比。然而,与此同时,可能导致(多特异性)抗体不期望地靶向表达fc受体的细胞,而不是优选的抗原携带细胞。此外,fc受体信号传导通路的共激活可导致细胞因子释放,该细胞因子释放与t细胞激活性质与(多特异性)抗体的长半衰期组合,在全身施用后导致对细胞因子受体的过度激活和严重的副作用。由于(例如)nk细胞可能破坏t细胞,因此激活t细胞以外的(携带fc受体)免疫细胞甚至可能降低(多特异性)抗体的功效。因此,在优选的方面,与天然igg1fc结构域相比,根据本发明所述的(多特异性)抗体的fc结构域表现出降低的对fc受体的结合亲和力和/或降低的效应子功能。在一个此类方面,fc结构域(或包含所述fc结构域的(多特异性)抗体)表现出与天然igg1结构域(或包含天然igg1fc结构域的(多特异性)抗体)相比小于50%、特别是小于20%、更特别是小于10%并且最特别是小于5%的与fc受体的结合亲和力,和/或与天然igg1fc结构域(或包含天然igg1fc结构域的(多特异性)抗体)相比小于50%、特别是小于20%、更特别是小于10%并且最特别是小于5%的效应子功能。在一个方面,fc结构域(或包含所述fc结构域的(多特异性)抗体)基本上不结合fc受体和/或诱导效应子功能。在一个优选的方面,fc受体是fcγ受体。在一个方面,fc受体是人fc受体。在一个方面,fc受体是激活fc受体。在一个具体方面,fc受体是激活人fcγ受体,更具体地是人fcγriiia、fcγri或fcγriia,最具体地是人fcγriiia。在一个方面,效应子功能是选自cdc、adcc、adcp和细胞因子分泌的组中的一种或多种效应子功能。在一个优选的方面,效应子功能是adcc。在一个方面,与天然igg1fc结构域相比,fc结构域对新生fc受体(fcrn)表现出基本相似的结合亲和力。当fc结构域(或包含所述fc结构域的(多特异性)抗体)表现出天然igg1fc结构域(或包含天然igg1fc结构域的(多特异性)抗体)对fcrn的结合亲和力的大于约70%、特别是大于约80%、更特别是大于约90%时,实现了基本上相似的与fcrn的结合。在某些方面,fc结构域经过工程化改造,以与非工程化改造的fc结构域相比具有降低的对fc受体的结合亲和力和/或降低的效应子功能。在优选的方面,(多特异性)抗体的fc结构域包含降低fc结构域对fc受体的结合亲和力和/或效应子功能的一个或多个氨基酸突变。典型地,相同的一个或多个氨基酸突变存在于fc结构域的两个亚基中的每一个中。在一个方面,氨基酸突变降低fc结构域对fc受体的结合亲和力。在一个方面,氨基酸突变将fc结构域对fc受体的结合亲和力降低至少2倍、至少5倍或至少10倍。在存在多于一个降低fc结构域对fc受体的结合亲和力的氨基酸突变的方面,这些氨基酸突变的组合可以将fc结构域对fc受体的结合亲和力降低至少10倍、至少20倍,或甚至至少50倍。在一个方面,与包含非工程化改造的fc结构域的(多特异性)抗体,包含工程化改造的fc结构域的(多特异性)抗体表现出对fc受体的结合亲和力的小于20%、特别是小于10%、更特别是小于5%。在一个优选的方面,fc受体是fcγ受体。在一些方面,fc受体是人fc受体。在一些方面,fc受体是激活fc受体。在一个具体方面,fc受体是激活人fcγ受体,更具体地是人fcγriiia、fcγri或fcγriia,最具体地是人fcγriiia。优选地,与这些受体中的每一个的结合力降低。在一些方面,对互补组分的结合亲和力,对c1q的特异性结合亲和力也降低。在一个方面,对新生儿fc受体(fcrn)的结合亲和力不降低。当fc结构域(或包含所述fc结构域的(多特异性)抗体)表现出fc结构域的非工程化改造形式(或包含fc结构域的所述非工程化改造形式的(多特异性)抗体)对fcrn的结合亲和力的大于约70%时,实现了基本上类似的与fcrn的结合,即实现了fc结构域对所述受体的结合亲和力的保留。fc结构域或本发明的包含所述fc结构域的(多特异性)抗体可表现出这种亲和力的大于约80%以及甚至大于约90%。在某些方面,(多特异性)抗体的fc结构域经过工程化改造,以相比于非工程化改造的fc结构域具有降低的效应子功能。降低的效应子功能可包括但不限于以下项中的一种或多种:降低的补体依赖性细胞毒性(cdc)、降低的抗体依赖性细胞介导的细胞毒性(adcc)、降低的抗体依赖性细胞吞噬作用(adcp)、减少的细胞因子分泌、减少的免疫复合物介导的抗原呈递细胞对抗原的摄取、减少的与nk细胞的结合、减少的与巨噬细胞的结合、减少的与单核细胞的结合、减少的与多形核细胞的结合、减少的直接信号传导诱导的细胞凋亡、减少的靶结合抗体的交联、减少的树突细胞成熟,或减少的t细胞致敏。在一个方面,降低的效应子功能是选自降低的cdc、降低的adcc、降低的adcp和减少的细胞因子分泌的组中的一种或多种降低的效应子功能。在一个优选的方面,降低的效应子功能是降低的adcc。在一个方面,降低的adcc是由非工程化改造的fc结构域(或包含所述非工程化改造的fc结构域的(多特异性)抗体)诱导的adcc的小于20%。在一个方面,降低fc结构域对fc受体的结合亲和力和/或效应子功能的氨基酸突变是氨基酸置换。在一个方面,fc结构域包含在选自e233、l234、l235、n297、p331和p329的组的位置处的氨基酸置换(根据kabateu索引编号)。在一个更具体的方面,fc结构域包含在选自l234、l235和p329的组的位置处的氨基酸置换(根据kabateu索引编号)。在一些方面,fc结构域包含氨基酸置换l234a和l235a(根据kabateu索引编号)。在一个此类方面,fc结构域是igg1fc结构域,特别是人igg1fc结构域。在一个方面,fc结构域在p329位处包含氨基酸取代。在一个更具体的方面,氨基酸置换是p329a或p329g,特别是p329g(根据kabateu索引编号)。在一个方面,fc结构域包含p329位的氨基酸置换以及进一步的在选自e233、l234、l235、n297和p331的组的位置处的氨基酸置换(根据kabateu索引编号)。在一个更具体的方面,进一步的氨基酸置换是e233p、l234a、l235a、l235e、n297a、n297d或p331s。在优选的方面,fc结构域包含在p329、l234和l235位的氨基酸置换(根据kabateu索引编号)。在更优选的方面,fc结构域包含氨基酸突变l234a、l235a和p329g(“p329glala”、“pglala”或“lalapg”)。具体地,在优选的方面,fc结构域的每个亚基包含氨基酸置换l234a、l235a和p329g(kabateu索引编号),即在fc结构域的第一亚基和第二亚基中的每个中,234位的亮氨酸残基被替换为丙氨酸残基(l234a)、235位的亮氨酸残基被替换为丙氨酸残基(l235a)并且329位的脯氨酸残基被替换为甘氨酸残基(p329g)(根据kabateu索引编号)。在一个此类方面,fc结构域是igg1fc结构域,特别是人igg1fc结构域。氨基酸取代的“p329glala”组合几乎完全消除了人igg1fc结构域的fcγ受体(以及补体)结合,如在pct公开号wo2012/130831中所述,该pct公开的全部内容以引用方式并入本文。wo2012/130831还描述了制备此类突变fc结构域的方法和确定其性质(诸如fc受体结合或效应子功能)的方法。与igg1抗体相比,igg4抗体表现出降低的对fc受体的结合亲和力和降低的效应子功能。因此,在一些方面,本发明的(多特异性)抗体的fc结构域是igg4fc结构域,特别是人igg4fc结构域。在一个方面,igg4fc结构域包含s228位的氨基酸置换,具体地包含氨基酸置换s228p(根据kabateu索引编号)。为了进一步降低其对fc受体的结合亲和力和/或其效应子功能,在一个方面,igg4fc结构域包含在l235位的氨基酸置换,具体地包含氨基酸置换l235e(根据kabateu索引编号)。在另一方面,igg4fc结构域包含在p329位的氨基酸置换,具体地,包含氨基酸置换p329g(根据kabateu索引编号)。在一个优选的方面,igg4fc结构域包含在s228、l235和p329位的氨基酸置换,具体地包含氨基酸置换s228p、l235e和p329g(根据kabateu索引编号)。这种igg4fc结构域突变体以及它们的fcγ受体结合性质描述于pct公开号wo2012/130831中,该pct公开的全部内容以引用方式并入本文。在一个优选的方面,与天然igg1fc结构域相比表现出降低的对fc受体的结合亲和力和/或降低的效应子功能的fc结构域是包含氨基酸置换l234a、l235a以及任选的p329g的人igg1fc结构域或包含氨基酸置换s228p、l235e以及任选的p329g的人igg4fc结构域(根据kabateu索引编号)。在某些方面,消除了fc结构域的n糖基化。在一个此类方面,fc结构域包含在n297位的氨基酸突变,特别是用丙氨酸(n297a)或天冬氨酸(n297d)替换天冬酰胺的氨基酸置换(根据kabateu索引编号)。除了上文和pct公开号wo2012/130831中所述的fc结构域外,具有减少的fc受体结合和/或降低的效应子功能的fc结构域还包括具有对fc结构域残基238、265、269、270、297、327和329中的一者或多者的取代的那些fc结构域(美国专利号6,737,056)(根据kabat的eu索引编号)。此类fc突变体包括在两个或更多个第265、269、270、297和327位氨基酸处具有置换的fc突变体,包括所谓的“dana”fc突变体,其残基265和297被置换为丙氨酸(美国专利号7,332,581)。可以使用本领域熟知的遗传或化学方法,通过氨基酸缺失、取代、插入或修饰来制备突变fc结构域。遗传方法可包括对编码dna序列的位点特异性诱变、pcr、基因合成等。可以例如通过测序来验证正确的核苷酸变化。与fc受体的结合可以例如通过elisa或通过表面等离子体共振(spr)使用标准仪器(诸如biacore仪器(gehealthcare))容易地确定,并且fc受体诸如可以通过重组表达获得。替代性地,可以使用已知表达特定fc受体的细胞系(诸如表达fcγiiia受体的人nk细胞)来评估fc结构域或包含fc结构域的(多特异性)抗体对fc受体的结合亲和力。fc结构域或包含所述fc结构域的(多特异性)抗体的效应子功能可通过本领域已知的方法测量。用于评定目标分子的adcc活性的体外测定的示例描述于美国专利号5,500,362;hellstrom等人,procnatlacadsciusa83,7059-7063(1986)和hellstrom等人,procnatlacadsciusa82,1499-1502(1985);美国专利号5,821,337;bruggemann等人,jexpmed166,1351-1361(1987)中。替代性地,可采用非放射性测定(参见例如,用于流式细胞术的actitm非放射性细胞毒性测定(celltechnology,inc.mountainview,ca);以及cytotox非放射性细胞毒性测定(promega,madison,wi))。用于此类测定的有用效应细胞包括外周血单核细胞(pbmc)和自然杀伤(nk)细胞。替代性地或另外地,可例如在诸如在clynes等人,procnatlacadsciusa95,652-656(1998)中公开的动物模型中体内评估目标分子的adcc活性。在一些方面,fc结构域与补体组分(具体地为c1q)的结合减少。因此,在一些方面,其中fc结构域经过工程化改造以具有降低的效应子功能,所述降低的效应子功能包括降低的cdc。可以进行c1q结合测定以确定fc结构域或包含该fc结构域的(多特异性)抗体是否能够结合c1q并因此具有cdc活性。参见例如wo2006/029879和wo2005/100402中的c1q和c3c结合elisa。为了评定补体激活,可以执行cdc测定(参见例如gazzano-santoro等人,jimmunolmethods202,163(1996);cragg等人,blood101,1045-1052(2003);以及cragg和glennie,blood103,2738-2743(2004))。fcrn结合和体内清除/半衰期测定也可以使用本领域已知的方法执行(参见例如petkova,s.b.等人,int’l.immunol.18(12):1759-1769(2006);wo2013/120929)。b.多核苷酸本发明进一步提高了编码本发明的抗体的分离的多核苷酸。所述分离的多核苷酸可以是单个多核苷酸或多个多核苷酸。编码本发明的(多特异性)抗体的多核苷酸可以表达为编码完整抗体的单个多核苷酸,或表达为共表达的多个(例如,两个或更多个)多核苷酸。由共表达的多核苷酸编码的多肽可以经由例如二硫键或其他方式缔合以形成功能性抗体。例如,抗体的轻链部分可以由来自包含抗体的重链的抗体部分的单独的多核苷酸编码。当共表达时,重链多肽将与轻链多肽缔合以形成免疫抗体。在另一个实例中,包含两个fc结构域亚基中的一个亚基和任选的一种或多种fab分子(的一部分)的抗体部分可由来自包含所述两个fc结构域亚基中的另一个亚基和任选的fab分子的(一部分)的抗体部分的单独多核苷酸编码。当共表达时,fc结构域亚基将缔合以形成fc结构域。在一些方面,分离的多核苷酸编码如本文所述的根据本发明的完整抗体分子。在其他方面,分离的多核苷酸编码如本文所述的包含在根据本发明的抗体中的多肽。在某些方面,多核苷酸或核酸是dna。在其他方面,本发明的多核苷酸是rna,例如以信使rna(mrna)的形式。本发明的rna可以是单链或双链的。c.重组方法本发明的抗体可以例如通过固态肽合成(例如merrifield固相合成)或重组生产获得。对于重组生产,将例如如上所述的编码抗体的一种或多种多核苷酸分离并且插入至一个或多个载体中以用于在宿主细胞中进一步克隆和/或表达。此类多核苷酸可使用常规方法容易地分离并且测序。在一个方面,提供了包含本发明的多核苷酸(即单个多核苷酸或多个多核苷酸)的载体,特别是表达载体。可以使用本领域技术人员熟知的方法来构建含有抗体的编码序列以及适当转录/翻译控制信号的表达载体。这些方法包括体外重组dna技术、合成技术和体内重组/遗传重组。参见例如以下文献所述的技术:maniatis等人,molecularcloning:alaboratorymanual,coldspringharborlaboratory,n.y.(1989);和ausubel等人,currentprotocolsinmolecularbiology,greenepublishingassociatesandwileyinterscience,n.y(1989)。表达载体可以是质粒、病毒的一部分,或者可以是核酸片段。表达载体包括表达盒,编码抗体的多核苷酸(即编码区)与启动子和/或其他转录或翻译控制元件可操作缔合地克隆至所述表达盒中。如本文所用,“编码区”是核酸的一部分,该部分由翻译成氨基酸的密码子组成。尽管“终止密码子”(tag、tga或taa)未被翻译成氨基酸,但其(如果存在的话)可被认为是编码区的一部分,而任何侧翼序列,例如启动子、核糖体结合位点、转录终止子、内含子、5'和3'非翻译区等不是编码区的一部分。两个或更多个编码区可存在于单个多核苷酸构建体中(例如在单个载体上),或在单独的多核苷酸构建体中(例如在单独的(不同的)载体上)。此外,任何载体可含有单一编码区,或可包含两个或更多个编码区,例如本发明的载体可以编码一种或多种多肽,该一种或多种多肽在翻译后或翻译时通过蛋白水解切割分离成最终的蛋白质。此外,本发明的载体、多核苷酸或核酸可编码异源编码区,该异源编码区与编码本发明的抗体的多核苷酸或其变体或衍生物融合或不融合。异源编码区包括但不限于特化元件或基序,诸如分泌信号肽或异源功能结构域。可操作缔合是当基因产物(例如多肽)的编码区以某种方式与一个或多个调控序列缔合,以使基因产物的表达处于调控序列的影响或控制下。如果启动子功能的诱导导致编码所需基因产物的mrna的转录,并且如果两个dna片段之间的连接性质不干扰表达调控序列指导基因产物表达的能力或干扰待转录的基因模板的能力,则该两个dna片段(诸如多肽编码区和与其相关的启动子)是“可操作地缔合的”。因此,如果启动子能够影响该核酸的转录,则启动子区域将与编码多肽的核酸可操作地缔合。启动子可以是细胞特异性启动子,该细胞特异性启动子仅在预定细胞中指导dna的实质转录。除启动子外,其他转录控制元件,例如增强子、操纵子、阻遏物和转录终止信号,可以与多核苷酸可操作地缔合以指导细胞特异性转录。本文公开了合适的启动子和其他转录控制区。多种转录控制区是本领域技术人员已知的。这些转录控制区包括但不限于在脊椎动物细胞中起作用的转录控制区,诸如但不限于来自巨细胞病毒的启动子和增强子区段(例如立即早期启动子结合内含子-a)、猿猴病毒40(例如早期启动子)和逆转录病毒(诸如例如劳氏肉瘤病毒)。其他转录控制区包括来源于脊椎动物基因(诸如肌动蛋白、热休克蛋白、牛生长激素和兔β珠蛋白)的那些转录控制区,以及能够控制真核细胞中基因表达的其他序列。其他合适的转录控制区包括组织特异性启动子和增强子以及诱导型启动子(例如四环素可诱导启动子)。类似地,各种翻译控制元件是本领域普通技术人员已知的。这些翻译控制元件包括但不限于核糖体结合位点、翻译起始和终止密码子,以及来源于病毒系统的元件(特别是内部核糖体进入位点,或ires,也称为cite序列)。表达盒还可以包括其他特征,诸如复制起点,和/或染色体整合元件,诸如逆转录病毒长末端重复序列(ltr),或腺相关病毒(aav)反向末端重复序列(itr)。本发明的多核苷酸和核酸编码区可以与编码分泌肽或信号肽的附加编码区缔合,所述附加编码区指导由本发明的多核苷酸编码的多肽的分泌。例如,如果需要分泌抗体,可以将编码信号序列的dna置于编码本发明的抗体或其片段的核酸的上游。根据信号假设,由哺乳动物细胞分泌的蛋白质具有信号肽或分泌前导序列,一旦已经起始跨粗面内质网输出生长的蛋白质链,该信号肽或分泌前导序列就被从成熟蛋白质上切割下来。本领域普通技术人员知道由脊椎动物细胞分泌的多肽通常具有与所述多肽的n末端融合的信号肽,所述信号肽被从翻译的多肽上切割下来以产生所述多肽的分泌或“成熟”形式。在某些方面,使用天然信号肽(例如免疫球蛋白重链或轻链信号肽),或保留指导与其可操作地缔合的多肽分泌的能力的该序列的功能性衍生物。替代性地,可以使用异源哺乳动物信号肽或其功能衍生物。例如,野生型前导序列可以被人组织纤溶酶原激活剂(tpa)或小鼠β葡糖醛酸酶的前导序列取代。编码可用于促进后续纯化的短蛋白质序列(例如组氨酸标签)或帮助标记抗体的dna可包含在抗体(片段)编码多核苷酸的内部或末端。在进一步方面,提供了包含本发明的多核苷酸(即,单个多核苷酸或多个多核苷酸)的宿主细胞。在某些方面,提供了包含本发明的载体的宿主细胞。多核苷酸和载体可以单独或组合地渗入本文中分别关于多核苷酸和载体描述的任何特征。在一个此类方面,宿主细胞包含一种或多种载体(例如已经用一种或多种载体转化或转染),该一种或多种载体包含编码本发明的抗体(的一部分)的一种或多种多核苷酸。如本文所用,术语“宿主细胞”是指可以被工程化改造以生成本发明的抗体或其片段的任何种类的细胞系统。适于复制和支持抗体表达的宿主细胞是本领域中熟知的。此类细胞可以用特定的表达载体适当地转染或转导,并且可以生长大量含有载体的细胞以用于接种大规模发酵罐来获得足够量的抗体用于临床应用。合适的宿主细胞包括原核微生物,诸如大肠杆菌,或各种真核细胞,诸如中国仓鼠卵巢细胞(cho)、昆虫细胞等。例如,多肽可以在细菌中产生,特别是当不需要糖基化时。多肽在表达后可以在可溶性级分中从细菌细胞糊状物中分离,并可以进一步纯化。除原核生物外,诸如丝状真菌或酵母之类的真核微生物也是用于编码多肽的载体的合适克隆或表达宿主,包括这样的真菌和酵母菌株,该真菌和酵母菌株的糖基化途径已经被“人源化”,从而导致产生具有部分或完全人糖基化模式的多肽。参见gerngross,natbiotech22,1409-1414(2004)和li等人,natbiotech24,210-215(2006)。用于表达(糖基化)多肽的合适宿主细胞还来源于多细胞生物(无脊椎动物和脊椎动物)。无脊椎动物细胞的实例包括植物细胞和昆虫细胞。已经鉴定出了许多可以与昆虫细胞一起使用的杆状病毒株,特别是用于转染草地夜蛾(spodopterafrugiperda)细胞。植物细胞培养物也可用作宿主。参见例如美国专利号5,959,177、6,040,498、6,420,548、7,125,978和6,417,429(描述了用于在转基因植物中产生抗体的plantibodiestm技术)。脊椎动物细胞也可用作宿主。例如,适于在悬浮液中生长的哺乳动物细胞系可能是有用的。有用的哺乳动物宿主细胞系的其他实例为由sv40转化的猴肾cv1系(cos-7);人胚肾系(293或293t细胞,如例如在graham等人,jgenvirol36,59(1977)中所述)、幼仓鼠肾细胞(bhk)、小鼠塞尔托利氏细胞(tm4细胞,如例如在mather,biolreprod23,243-251(1980)中所述)、猴肾细胞(cv1)、非洲绿猴肾细胞(vero-76)、人宫颈癌细胞(hela)、犬肾细胞(mdck)、布法罗大鼠肝细胞(brl3a)、人肺细胞(w138)、人肝细胞(hepg2)、小鼠乳腺肿瘤细胞(mmt060562)、tri细胞(如例如在mather等人,annalsn.y.acadsci383,44-68(1982)中所述)、mrc5细胞,以及fs4细胞。其他有用的哺乳动物宿主细胞系包括中国仓鼠卵巢(cho)细胞,包括dhfr-cho细胞(urlaub等人,procnatlacadsciusa77,4216(1980));以及骨髓瘤细胞系,诸如yo、ns0、p3x63和sp2/0。关于适用于蛋白质生产的某些哺乳动物宿主细胞系的综述,请参见例如yazaki和wu,methodsinmolecularbiology,第248卷(b.k.c.lo编辑,humanapress,totowa,nj),第255-268页(2003)。宿主细胞包括培养细胞,例如仅举几个例子而言哺乳动物培养细胞、酵母细胞、昆虫细胞、细菌细胞和植物细胞,还包括转基因动物、转基因植物或培养植物或动物组织中所包含的细胞。在一个方面,宿主细胞是真核细胞,特别是哺乳动物细胞,诸如中国仓鼠卵巢(cho)细胞、人胚肾(hek)细胞或淋巴细胞(例如y0、ns0、sp20细胞)。在一个方面,宿主细胞并非人体内的细胞。用于在这些系统中表达外源基因的标准技术是本领域已知的。可以对表达包含抗原结合结构域的重链或轻链的多肽(诸如抗体)的细胞进行工程化改造,以便也表达另一条抗体链,使得表达的产物为具有重链和轻链的抗体。在一个方面,提供了一种生产根据本发明所述的抗体的方法,其中该方法包括在适于表达抗体的条件下培养包含编码如本文所提供的抗体的多核苷酸的宿主细胞,以及任选地从该宿主细胞(或宿主细胞培养基)回收抗体。本发明的(多特异性)抗体的组分可彼此遗传融合。(多特异性)抗体可设计为使得它的组分直接彼此融合或通过连接基序列间接融合。连接基的组成和长度可以根据本领域熟知的方法测定,并且可以测试连接基的功效。本文提供了(多特异性)抗体的不同组分之间的连接基序列的实例。如果需要的话,还可包括另外的序列(例如内肽酶识别序列)以掺入切割位点来分离融合的各个组分。如本文所述制备的抗体可通过本领域已知的技术纯化,所述技术为诸如高效液相色谱、离子交换色谱、凝胶电泳、亲和色谱、体积排阻色谱等。用于纯化特定蛋白质的实际条件将部分取决于诸如净电荷、疏水性、亲水性等因素,并且对于本领域技术人员而言将是显而易见的。对于亲和色谱纯化,可以使用与抗体结合的抗体、配体、受体或抗原。例如,对于本发明的抗体的亲和色谱纯化,可以使用具有蛋白a或蛋白g的基质。基本上如实例中所述,可使用顺序蛋白a或g亲和色谱和体积排阻色谱来分离抗体。抗体的纯度可以通过各种熟知的分析方法中的任何一种来确定,所述各种熟知-的分析方法包括凝胶电泳、高压液相色谱等。d.测定本文所提供的抗体的物理/化学性质和/或生物学活性可通过本领域中已知的各种测定法进行鉴定、筛选或表征。1.结合测定抗体与fc受体或靶抗原的结合(亲和力)可例如使用标准仪器诸如biacore仪器(gehealthcare)以及可通过重组表达获得的受体或靶蛋白,通过表面等离子体共振(spr)来确定。替代性地,抗体与不同受体或靶抗原的结合可使用表达特定受体或靶抗原的细胞系来评估,例如通过流式细胞术(facs)进行评估。下文描述了用于测量与cd3的结合活性的具体说明性和示例性方面。在一个方面,与cd3的结合活性通过以下spr进行测定:在biacoret200仪器(gehealthcare)上执行spr。使用标准的胺偶联化学方法,在4000–6000共振单位(ru)的表面密度下,将抗fab捕获抗体(gehealthcare,#28958325)固定到s系列传感器芯片cm5(gehealthcare)上。使用hbs-p+(10mmhepes,150mmnaclph7.4,0.05%表面活性剂p20)作为运行和稀释缓冲液。在约60s内,以5μl/min的流速注入浓度为2μg/ml(在20mmhis、140mmnacl中,ph6.0)的cd3抗体。所用的cd3抗原是cd3δ和cd3ε胞外结构域的异源二聚体,其融合至具有杵臼结构修饰和c末端avi标签的人fc结构域上(参见seqidno:28和seqidno:29)。在120s内注入浓度为10μg/ml的cd3抗原,并且在约120s内以5μl/min的流速监测解离。通过连续两次(每次约60s)注入10mm甘氨酸(ph2.1),使芯片表面再生。通过减去空白注入并通过减去从空白对照流通池获得的响应来校正本体折射率偏差。为实施评估,在注入结束后5秒采集结合响应。为了使结合信号标准化,将cd3结合除以抗fab响应(在固定化抗fab抗体上捕获cd3抗体后获得的信号(ru))。相对于不同处理后抗体与cd3的结合活性(也称为相对活性浓度(rac)),经过特定处理后的抗体与cd3的结合活性通过参考经过特定处理的抗体样品的结合活性与经过不同处理后相应的抗体样品的结合活性来计算。2.活性测定本发明的(多特异性)抗体的生物学活性可通过如实例中所述的各种测定方法进行测量。生物学活性可例如包括诱导t细胞的增殖、诱导t细胞中的信号传导、诱导t细胞中的激活标志物的表达、诱导t细胞的细胞因子分泌、诱导靶细胞诸如肿瘤细胞的裂解以及诱导肿瘤消退和/或存活率的改善。e.组合物、配方和施用途径在进一步方面,本发明提供了包含本文提供的抗体中任一者的药物组合物,其例如用于以下治疗方法中的任一者中。在一个方面,药物组合物包含根据本发明所述的抗体以及药用载体。在另一方面,药物组合物包含根据本发明所述的抗体以及至少一种另外的治疗剂,例如如下所述。进一步提供了一种以适于体内施用的形式产生本发明的抗体的方法,该方法包括(a)获得根据本发明的抗体,以及(b)用至少一种药用载体配制该抗体,由此配制抗体以用于体内施用。本发明的药物组合物包含溶解或分散在药用载体中的有效量的抗体。术语“药用”是指分子实体和组合物在所采用的剂量和浓度下通常对接受者无毒,即当视情况而定施用于动物(例如人)时不产生不利的、过敏的或其他不良反应。根据本公开内容,含有抗体以及任选的另外的活性成分的药物组合物的制备将对本领域技术人员而言是已知的,如由remington'spharmaceuticalsciences,第18版,mackprintingcompany,1990例示的,其通过引用并入本文。此外,对于动物(例如,人)施用,应理解制备物应满足fda生物标准办公室或其他国家/地区相应当局所要求的无菌性、产热原性、一般安全性和纯度标准。优选的组合物是冻干制剂或水溶液。如本文所使用的,“药用载体”包括任何和所有的溶剂、缓冲液、分散介质、包衣、表面活性剂、抗氧化剂、防腐剂(例如抗细菌剂、抗真菌剂)、等渗剂、吸收延迟剂、盐、防腐剂、抗氧化剂、蛋白质、药物、药物稳定剂、聚合物、凝胶、结合剂、赋形剂、崩解剂、润滑剂、甜味剂、调味剂、染料,类似物质以及它们的组合,如本领域普通技术人员应已知的(参见例如remington'spharmaceuticalsciences,第18版,mackprintingcompany,1990,第1289-1329页,其通过引用并入本文)。除了任何常规载体与活性成分不相容的情况之外,所述载体在药物组合物中的用途是可预期的。本发明的抗体(和任何另外的治疗剂)可以通过任何合适的方式施用,包括肠胃外、肺内和鼻内,并且如果需要的话用于局部治疗、病灶内施用。肠胃外输注包括肌内、静脉内、动脉内、腹膜内或皮下施用。投配可以通过任何合适的途径进行,例如通过注射,诸如静脉内或皮下注射,部分取决于施用是短暂的还是长期的。肠胃外组合物包括设计用于通过注射(例如,皮下、皮内、病灶内、静脉内、动脉内、肌内、鞘内或腹膜内注射)施用的那些组合物。对于注射,本发明的抗体可以在水溶液中配制,特别是在生理上相容的缓冲液(诸如hanks溶液、ringer溶液或生理盐水缓冲液)中配制。溶液可含有配制剂(formulatoryagent),诸如悬浮剂、稳定剂和/或分散剂。替代性地,抗体可以是粉末形式,用于在使用前用合适的媒介物(例如无菌无热原水)构建。通过根据需要将本发明的抗体以所需的量与下面列举的各种其他成分一起掺入适当的溶剂中来制备无菌可注射溶液。例如,无菌可以通过无菌过滤膜过滤而容易地实现。通常,通过将各种灭菌的活性成分掺入含有基础分散介质和/或其他成分的无菌媒介物中来制备分散体。在用于制备无菌可注射溶液、悬浮液或乳液的无菌粉末的情况下,优选的制备方法是真空干燥或冻干技术,所述真空干燥或冻干技术产生来自先前无菌过滤的液体介质的活性成分加上任何附加所需成分的粉末。如果需要的话,液体介质应适当缓冲,并且在注射之前应首先使用足够的盐水或葡萄糖来使液体稀释剂等渗。该组合物必须是在制造和贮存条件下稳定的,并且保存为抗诸如细菌和真菌等微生物的污染作用。应当理解,内毒素污染应以例如低于0.5ng/mg蛋白的安全水平保持最低。合适的药用载体包括但不限于:缓冲剂,例如磷酸盐、柠檬酸盐和其他有机酸;抗氧化剂,包括抗坏血酸和蛋氨酸;防腐剂(诸如十八烷基二甲基苄基氯化铵;氯化六烃季铵;苯扎氯铵;苄索氯铵;酚、丁醇或苄醇;对羟基苯甲酸烷基酯,诸如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3-戊醇;和间甲酚);低分子量(少于约10个残基)多肽;蛋白质,诸如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,诸如聚乙烯吡咯烷酮;氨基酸,诸如甘氨酸、谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖和其他碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂,诸如edta;糖类,诸如蔗糖、甘露醇、海藻糖或山梨糖醇;成盐抗衡离子,诸如钠;金属络合物(例如锌蛋白络合物);和/或非离子表面活性剂,诸如聚乙二醇(peg)。水性注射悬浮液可含有增加悬浮液粘度的化合物,诸如羧甲基纤维素钠、山梨糖醇、葡聚糖等。任选地,悬浮液还可含有合适的稳定剂或增大化合物溶解度的试剂,以允许制备高浓度溶液。另外,活性化合物的悬浮液可以制备成适当的油性注射悬浮液。合适的亲脂性溶剂或载体包括脂肪油,诸如芝麻油;或合成脂肪酸酯,诸如油酸乙酯或甘油三酯;或脂质体。活性成分可以包埋在例如通过凝聚技术或通过界面聚合而制备的微胶囊(例如分别为羟甲基纤维素或明胶微胶囊和聚(甲基丙烯酸甲酯)微胶囊)中;在胶体药物递送系统(例如,脂质体、白蛋白、微球、微乳液、纳米粒子和纳米胶囊)中;或在粗乳液中。此类技术在remington'spharmaceuticalsciences(第18版,mackprintingcompany,1990)中公开。可以制备缓释制备物。缓释制备物的合适示例包括含有多肽的固态疏水聚合物的半透性基质,所述基质是例如膜或微胶囊等成型制品的形式。在特定方面,可注射组合物的延长吸收可以通过在所述组合物中使用延迟吸收的试剂(例如单硬脂酸铝、明胶或它们的组合)来实现。除了先前描述的组合物之外,抗体还可以配制成贮库制剂。此类长效制剂可以通过植入(例如皮下或肌内植入)或通过肌内注射施用。因此,例如,抗体可以用合适的聚合或疏水材料配制(例如作为可接受油中的乳液)或用离子交换树脂配制,或配制为微溶的衍生物,例如配制为微溶盐。包含本发明的抗体的药物组合物可通过常规混合、溶解、乳化、包封、包埋或冻干的手段制备。药物组合物可以使用一种或多种生理上可接受的载体、稀释剂、赋形剂或助剂以常规方式配制,所述载体、稀释剂、赋形剂或助剂有助于将蛋白质加工成可以在药学上使用的制备物。适当的配方取决于所选择的施用途径。抗体可以配制成游离酸或碱、中性或盐形式的组合物。药用盐是基本上保留游离酸或游离碱的生物活性的盐。这些药用盐包括酸加成盐,例如与蛋白质组合物的游离氨基形成的酸加成盐,或与无机酸(诸如盐酸或磷酸)或有机酸(如乙酸、草酸、酒石酸或扁桃酸)形成的酸加成盐。用游离羧基形成的盐也可以衍生自无机碱,诸如氢氧化钠、氢氧化钾、氢氧化铵、氢氧化钙或氢氧化铁;或者有机碱,诸如异丙胺、三甲胺、组氨酸或普鲁卡因。与相应的游离碱形式相比,药用盐倾向于更易溶于水性和其他质子溶剂中。f.治疗方法和组合物本文提供的任何抗体可用于治疗方法。本发明的抗体可用作免疫治疗剂,例如用于治疗癌症。为了用于治疗方法中,本发明的抗体将以符合良好医学实践的方式配制、投配和施用。在这种情况下需要考虑的因素包括所治疗的特定疾患、所治疗的特定哺乳动物、个体患者的临床病症、疾患的原因、药剂的递送部位、施用方法、施用的时间安排,以及执业医师已知的其他因素。在一个方面,将本发明的抗体提供为用作药物。在其他方面,将本发明的抗体提供为用于治疗疾病。在某些方面,提供了用于治疗方法的本发明的抗体。在一个方面,本发明提供了本发明的抗体,其用于治疗有此需要的个体的疾病。在某些方面,本发明提供了一种用于治疗患有疾病的个体的方法中的抗体,该方法包括向个体施用有效量的抗体。在某些方面,待治疗的疾病是增殖性疾病。在一个优选的方面,疾病为癌症。在某些方面,该方法还包括向个体施用有效量的至少一种另外的治疗剂,例如如果待治疗的疾病是癌症,则使用抗癌剂。在进一步方面,本发明提供了本发明的抗体,其用于诱导靶细胞、特别是肿瘤细胞的裂解。在某些方面,本发明提供了用于在个体中诱导靶细胞、特别是肿瘤细胞的裂解的方法中的本发明的抗体,该方法包括向个体施用有效量的抗体以诱导靶细胞的裂解。根据上述方面中的任一方面的“个体”是哺乳动物,优选地是人。在进一步方面,本发明提供了本发明的抗体在制备或配制药物中的用途。在一个方面,药物用于治疗有此需要的个体的疾病。在进一步方面,药物用于治疗疾病的方法中,该方法包括向患有疾病的个体施用有效量的药物。在某些方面,待治疗的疾病是增殖性疾病。在一个优选的方面,疾病为癌症。在一个方面,该方法进一步包括向个体施用有效量的至少一种另外的治疗剂,例如如果待治疗的疾病是癌症,则使用抗癌剂。在进一步方面,药物用于诱导靶细胞、特别是肿瘤细胞的裂解。在进一步方面,药物用于在个体中诱导靶细胞、特别是肿瘤细胞的裂解的方法中的本发明的抗体,该方法包括向个体施用有效量的药物以诱导靶细胞的裂解。根据上述方面中的任一方面的“个体”可以是哺乳动物,优选地是人。在进一步方面,本发明提供了一种用于治疗疾病的方法。在一个方面,该方法包括向患有此类疾病的个体施用有效量的本发明的抗体。在一个方面,向所述个体施用组合物,该组合物包含药用形式的本发明的抗体。在某些方面,待治疗的疾病是增殖性疾病。在一个优选的方面,疾病为癌症。在某些方面,该方法还包括向个体施用有效量的至少一种另外的治疗剂,例如如果待治疗的疾病是癌症,则使用抗癌剂。根据上述方面中的任一方面的“个体”可以是哺乳动物,优选地是人。在进一步方面,本发明提供了一种用于诱导靶细胞、特别是肿瘤细胞的裂解的方法。在一个方面,该方法包括在t细胞、特别是细胞毒性t细胞的存在下使靶细胞与本发明的抗体接触。在进一步方面,提供了一种用于在个体中诱导靶细胞、特别是肿瘤细胞的裂解的方法。在一个此类方面,该方法包含向个体施用有效量的本发明的抗体以诱导靶细胞的裂解。在一个方面,“个体”是人。在某些方面,待治疗的疾病是增殖性疾患,特别是癌症。癌症的非限制性实例包括膀胱癌、脑癌、头颈癌、胰腺癌、肺癌、乳腺癌、卵巢癌、子宫癌、宫颈癌、子宫内膜癌、食道癌、结肠癌、结直肠癌、直肠癌、胃癌、前列腺癌、血癌、皮肤癌、鳞状细胞癌、骨癌,以及肾癌。可以使用本发明的抗体治疗的其他细胞增殖疾患包括但不限于位于以下部位中的肿瘤:腹部、骨骼、乳腺、消化系统、肝脏、胰腺、腹膜、内分泌腺(肾上腺、甲状旁腺、垂体、睾丸、卵巢、胸腺、甲状腺)、眼、头颈部、神经系统(中枢神经系统和外周神经系统)、淋巴系统、骨盆、皮肤、软组织、脾脏、胸部以及泌尿生殖系统。还包括癌前病症或病变和癌转移。在某些方面,癌症选自由以下项组成的组:肾癌、膀胱癌、皮肤癌、肺癌、结直肠癌、乳腺癌、脑癌、头颈癌和前列腺癌。在一个方面,特别是在其中抗体是与作为第二抗原的tyrp-1结合的多特异性抗体的情况下,癌症是表达tyrp-1的癌症。在一个方面,特别是在其中抗体是与作为第二抗原的tyrp-1结合的多特异性抗体的情况下,癌症是皮肤癌,特别是黑素瘤。在一个方面,特别是在其中抗体是与作为第二抗原的egfrviii结合的多特异性抗体的情况下,癌症是表达egfrviii的癌症。在一个方面,特别是在其中抗体是与作为第二抗原的egfrviii结合的多特异性抗体的情况下,癌症是脑癌,特别是胶质母细胞瘤。本领域技术人员容易地认识到,在许多情况下,抗体可能无法提供治愈,而仅可以提供部分益处。在一些方面,具有某些益处的生理变化也被视为具有治疗益处。因此,在一些方面,提供生理变化的抗体的量被认为是“有效量”。需要治疗的受试者、患者或个体通常是哺乳动物,更特别地是人。在一些方面,将有效量的本发明的抗体施用于个体以治疗疾病。为了预防或治疗疾病,本发明的抗体的适当剂量(当单独使用或与一种或多种附加治疗剂联合使用时)将取决于待治疗疾病的类型、施用途径、患者体重、抗体类型、疾病的严重程度和进程、抗体是出于预防还是治疗目的施用、先前或同时进行的治疗性干预、患者的临床病史和对抗体的应答以及主治医师的酌处权。在任何情况下,负责施用的执业者将针对个体受试者来确定组合物中活性成分的浓度和适当剂量。本文考虑了各种投配时间安排,包括但不限于在各个时间点处的单次或多次施用、推注施用,以及脉冲输注。抗体适当地一次或在一系列治疗中施用于患者。取决于疾病的类型和严重性,约1μg/kg至15mg/kg(例如0.1mg/kg-10mg/kg)的抗体可以是例如通过一次或多次单独施用或通过连续输注而施用于患者的初始候选剂量。取决于上述因素,一种典型的日剂量的范围可以为约1μg/kg至100mg/kg或更多。对于数天或更长时间的重复施用,取决于病症,治疗通常会持续直至发生所需的疾病症状抑制。抗体的一种示例性剂量的范围为约0.005mg/kg至约10mg/kg。在其他非限制性实例中,剂量还可包括每次施用约1微克/千克体重、约5微克/千克体重、约10微克/千克体重、约50微克/千克体重、约100微克/千克体重、约200微克/千克体重、约350微克/千克体重、约500微克/千克体重、约1毫克/千克体重、约5毫克/千克体重、约10毫克/千克体重、约50毫克/千克体重、约100毫克/千克体重、约200毫克/千克体重、约350毫克/千克体重、约500毫克/千克体重至约1000毫克/千克体重或更多,以及从其中推导出的任何范围。在从本文所列数字可推导出的范围的非限制性实例中,约5毫克/千克体重至约100毫克/千克体重、约5微克/千克体重至约500毫克千克体重等的范围可以基于上述数值施用。因此,可以向患者施用约0.5mg/kg、2.0mg/kg、5.0mg/kg或10mg/kg(或它们的任何组合)的一种或多种剂量。此类剂量可以间歇施用,例如每周或每三周施用(例如,使得患者接受约两次至约二十次,或例如约六次剂量的抗体)。可施用初始较高负荷剂量,然后施用一种或多种较低剂量。然而,其他剂量方案可能有用。该疗法的进展通过常规技术和测定而容易地监测。本发明的抗体将通常以有效实现预期目的的量使用。为用于治疗或预防病症,将本发明的抗体或其药物组合物以有效量施用或施加。对于全身施用,有效剂量可以最初根据体外测定(诸如细胞培养物测定)来进行估计。可以随后在动物模型中配制剂量,以实现包括如在细胞培养中测定的ic50循环浓度范围。此类信息可用于更准确地确定对人类的有用剂量。初始剂量也可以使用本领域公知的技术,根据体内数据(例如动物模型)来估计。可以单独地调节剂量的量和间隔,以提供足以维持治疗效果的抗体的血浆水平。通过注射施用的常用患者剂量的范围是约0.1至50mg/kg/天,通常约0.5至1mg/kg/天。通过每天施用多个剂量可以实现治疗有效的血浆水平。可以例如通过hplc来测量血浆中的水平。有效剂量的本发明的抗体通常将在不会引起显著毒性的情况下提供治疗益处。抗体的毒性和治疗功效可通过标准药学方法在细胞培养或实验动物中测定。细胞培养测定和动物研究可以用于测定ld50(致死群体的50%的剂量)和ed50(在群体的50%中治疗有效的剂量)。毒性和疗效之间的剂量比是治疗指数,所述治疗指数可以表示为比率ld50/ed50。表现出大治疗指数的抗体是优选的。在一个方面,根据本发明的抗体表现出高治疗指数。从细胞培养测定和动物研究获得的数据可用于配制适用于人类的一系列剂量。剂量优选在包括几乎没有毒性或没有毒性的ed50的循环浓度的范围内。剂量可以取决于多种因素而在该范围内变化,所述多种因素为例如所采用的剂型、所利用的施用途径、受试者的病症等。确切的配方、施用途径和剂量可以由个别医生根据患者的病症来选择(参见例如fingl等人,1975,在:thepharmacologicalbasisoftherapeutics,第1章,第1页中,该文献的全部内容以引用方式并入本文中)。用本发明的抗体治疗的患者的主治医师将知道如何以及何时由于毒性、器官功能障碍等终止、中断或调节施用。相反地,如果临床应答不充分(排除毒性),则主治医师也会知道将治疗调节到更高水平。在目标病症的管理中施用的剂量的大小将随着待治疗病症的严重程度、施用途径等而变化。例如,可以部分地通过标准预后评估方法来评估病症的严重性。此外,剂量和可能的剂量频率也将根据个体患者的年龄、体重和应答而变化。本发明的抗体可以与治疗中的一种或多种其他药剂组合施用。例如,本发明的抗体可与至少一种另外的治疗剂共同施用。术语“治疗剂”包括被施用以治疗需要这种治疗的个体的症状或疾病的任何药剂。此类另外的治疗剂可包含适合于所治疗的具体疾病的任何活性成分,优选地是具有不会彼此不利地影响的互补活性的活性成分。在某些方面,另外的治疗剂是免疫调节剂、细胞生长抑制剂、细胞粘附抑制剂、细胞毒性剂、细胞凋亡激活剂,或增加细胞对凋亡诱导剂的敏感性的试剂。在一个优选的方面,另外的治疗剂是抗癌剂,例如微管破坏剂、抗代谢物、拓扑异构酶抑制剂、dna嵌入剂、烷化剂、激素疗法、激酶抑制剂、受体拮抗剂、肿瘤细胞凋亡激活剂或抗血管生成剂。此类其他药剂适当地以对预期目的有效的量组合存在。此类其他药剂的有效量取决于所用抗体的量、病症或治疗的类型,以及上面讨论的其他因素。抗体通常以与本文所述相同的剂量和施用途径使用,或以本文所述剂量的约1%至99%使用,或以经验地/临床上确定为合适的任何剂量和任何途径使用。上述此类组合疗法包括联合施用(其中两种或更多种治疗剂包括在相同或不同的组合物中)和单独施用,在单独施用的情况下,本发明的抗体的施用可以在施用另外的治疗剂和/或佐剂之前、同时和/或之后进行。本发明的抗体也可以与放射疗法组合使用。g.制品在本发明的另一个方面中,提供了一种制品,其含有可用于治疗、预防和/或诊断上述病症的物质。该制品包括容器和在所述容器上或与所述容器相关的标签或包装插页(packageinsert)。合适的容器包括例如瓶子、小瓶、注射器、静脉内注射(iv)溶液袋等。所述容器可以由诸如玻璃或塑料等多种材料形成。所述容器容纳组合物,该组合物本身或与另一种组合物组合以有效治疗、预防和/或诊断病症,并且所述容器可具有无菌进入口(例如,所述容器可以是静脉内溶液袋或具有可由皮下注射针刺穿的塞子的小瓶)。组合物中的至少一种活性剂是本发明的抗体。标签或包装插页指示该组合物用于治疗所选择的病症。此外,制品可包括(a)第一容器,该第一容器中含有组合物,其中组合物包含本发明的抗体;以及(b)第二容器,该第二容器中含有组合物,其中组合物包含另外的细胞毒性剂或其他治疗剂。本发明这一方面的制品还可包含包装插页,该包装插页指示组合物可用于治疗特定病症。替代性地或另外地,制品还可包括第二容器(或第三容器),该第二容器(或第三容器)容器包括药用缓冲液,诸如抑菌性注射用水(bwfi)、磷酸盐缓冲盐水、林格氏溶液和葡萄糖溶液。制品还可以包括从商业和用户角度所需的其他物质,包括其他缓冲剂、稀释剂、过滤器、针头和注射器。h.用于诊断和检测的方法和组合物在某些方面,本文提供的任何抗体均可用于检测生物样品中是否存在相应的抗体靶点(例如cd3、tyrp-1、egfrviii)。如本文所用的术语“检测”涵盖定量或定性检测。在某些方面,生物样品包括细胞或组织,诸如前列腺组织。在一个方面,提供了用于诊断或检测方法中的根据本发明所述的抗体。在进一步方面,提供了一种检测生物样品中cd3、tyrp-1或egfrviii的存在的方法。在某些方面,该方法包括在允许本发明的抗体与cd3、tyrp-1或egfrviii结合的条件下使生物样品与该抗体接触,以及检测在抗体与cd3、tyrp-1或egfrviii之间是否形成了复合物。此类方法可以是体外或体内方法。在一个方面,本发明的抗体用于选择符合用结合cd3、tyrp-1和/或egfrviii的抗体进行治疗的受试者,例如其中cd3、tyrp-1和/或egfrviii是患者选择的生物标志物。可使用本发明的抗体诊断的示例性疾病包括癌症,特别是皮肤癌或脑癌。在某些方面,提供了根据本发明所述的抗体,其中抗体得到标记。标记包括但不限于直接检测的标记或部分(诸如荧光标记、发色标记、电子致密标记、化学发光标记,以及放射性标记),以及间接(例如通过酶促反应或分子相互作用)检测的部分(诸如酶或配体)。示例性标记包括但不限于放射性同位素32p、14c、125i、3h和131i;荧光团,诸如稀土螯合物或荧光素(fluorescein)及其衍生物、罗丹明及其衍生物、丹酰、伞形酮;荧光素酶(luciferase),例如萤火虫荧光素酶和细菌荧光素酶(美国专利no.4,737,456);荧光素(luciferin);2,3-二氢二氮杂萘二酮(2,3-dihydrophthalazinedione);辣根过氧化物酶(hrp);碱性磷酸酶;β-半乳糖苷酶;葡糖淀粉酶;溶菌酶;糖氧化酶,例如葡萄糖氧化酶、半乳糖氧化酶和葡萄糖-6-磷酸脱氢酶;杂环氧化酶,诸如尿酸氧化酶和黄嘌呤氧化酶;与采用过氧化氢来氧化染料前体的酶(诸如hrp、乳过氧化物酶,或微过氧化物酶)偶联;生物素/抗生物素蛋白;纺丝标记;噬菌体标记;稳定自由基等。iii.序列iv.实例以下是本发明的方法和组合物的实例。应当理解,在给出以上提供的一般描述的情况下,可以实践各种其他方面。实例1-优化后的cd3结合剂的生成从之前描述的(参见例如wo2014/131712,其通过引用并入本文)cd3结合剂(在本文中称为“cd3orig”并且分别包含seqidno:6和seqidno:11的vh和vl序列)开始,旨在通过去除重链cdr3的kabat位置97和100处的两个天冬酰胺脱酰胺序列基序,优化该结合剂的特性。为此,生成一个适于噬菌体展示的重链抗体库,该抗体库去除了kabat位置97和100处的天冬酰胺,并且随机分配了cdrh1、h2和h3,以便补偿通过亲和力成熟过程替换asn97和asn100所引起的亲和力的损失。通过与次要外壳蛋白p3融合,将该文库置于丝状噬菌体上(marks等人(1991)jmolbiol222,581-597),并且选择与重组cd3ε结合。在初步筛选中鉴定出10个候选克隆,其作为fab片段在重组抗原上表现出通过spr测得的可接受的结合(在大肠杆菌中产生)。但是,这些克隆中仅一个克隆在转化为igg形式后表现出与cd3表达细胞的可接受的结合活性,该结合活性通过流式细胞仪测得。进一步评估所选择的克隆,在本文中为“cd3opt”,其分别包含seqidno:7和seqidno:11的vh和vl序列,并且如下所述使其转化为双特异性形式。实例2-优化后的cd3结合剂与cd3的结合与重组cd3的结合利用表面等离子体共振(spr)测定优化后的cd3结合剂“cd3opt”和原始cd3结合剂“cd3orig”与重组cd3的结合,这两种结合剂均为在fc区中包含p329gl234al235a(“pglala”,根据eu编号)突变的人igg1的形式(seqidno:12和seqidno:14(cd3orig)和seqidno:13和seqidno:14(cd3opt))。为评估脱酰胺作用位点去除的效果及其对抗体稳定性的影响,在37℃或40℃温度应激下储存14天后,检测原始cd3结合剂和优化后的cd3结合剂与重组cd3的结合。利用储存于-80℃的样品作为参比。参比样品和在40℃经过应激的样品在20mmhis、140mmnacl(ph6.0)中,以及在37℃在pbs(ph7.4)中经过应激的样品,其浓度均为1.2-1.3mg/ml。在应激期(14天)结束后,将pbs中的样品透析回20mmhis、140mmnacl(ph6.0)中进行进一步分析。利用spr按照以下步骤测定样品的相对活性浓度(rac)。在biacoret200仪器(gehealthcare)上执行spr。使用标准的胺偶联化学方法,在4000-6000共振单位(ru)的表面密度结果,将抗fab捕获抗体(gehealthcare,#28958325)固定到s系列传感器芯片cm5(gehealthcare)上。使用hbs-p+(10mmhepes,150mmnaclph7.4,0.05%表面活性剂p20)作为运行和稀释缓冲液。在60s内,以5μl/min的流速注入浓度为2μg/ml的cd3抗体。在120s内注入浓度为10μg/ml的cd3抗原(见下文),并且在120s内以5μl/min的流速监测解离。通过连续两次(每次60s)注入10mm甘氨酸(ph2.1),使芯片表面再生。通过减去空白注入并通过减去从空白对照流通池获得的响应来校正本体折射率偏差。为实施评估,在注入结束后5秒采集结合响应。为了使结合信号标准化,将cd3结合除以抗fab响应(在固定化抗fab抗体上捕获cd3抗体后获得的信号(ru))。通过参考每个经温度应激的样品与相应的未经应激的样品,计算出相对活性浓度。所用的抗原是cd3δ和cd3ε胞外结构域的异源二聚体,其融合至具有杵臼结构修饰和c末端avi标签的人fc结构域上(参见seqidno:28和seqidno:29)。该实验的结果在图2中示出。从图中可以看出,与原始cd3结合剂cd3orig相比,优化后的cd3结合剂cd3opt在经过温度应激(在37℃和ph7.4下暴露2周)后表现出显著改善的与cd3的结合。这一结果证明,脱酰胺作用位点的去除取得成功,并且产生了具有优异的稳定性的抗体,其稳定性与体内半衰期以及中性ph抗体的配制相关。与jurkat细胞上cd3的结合利用facs测定优化后的cd3结合剂“cd3opt”和原始cd3结合剂“cd3orig”与人报告t细胞系jurkatnfat上cd3的结合,这两种结合剂均为在fc区中包含p329gl234al235a(“pglala”,根据eu编号)突变的人igg1的形式(seqidno:12和seqidno:14(cd3orig)以及seqidno:13和seqidno:14(cd3opt))。jurkat-nfat报告细胞(gloresponsejurkatnfat-re-luc2p;promega#cs176501)是一种包含nfat启动子的人急性淋巴白血病报告细胞系,其表达人cd3。将细胞以0.1-0.5mio细胞/ml的密度在rpmi1640、2g/l葡萄糖、2g/lnahco3、10%fcs、25mmhepes、2mml-谷氨酰胺、1xneaa、1x丙酮酸钠中培养。每当细胞传代时,添加最终浓度为200μg/ml潮霉素b。为了进行结合测定,收获jurkatnfat细胞,用pbs洗涤,然后重悬于facs缓冲液中。在96孔圆形底板中进行抗体染色。因此,在每个孔中接种100000至2000000个细胞。在400xg下将孔板离心4min,并且除去上清液。将检测抗体用facs稀释液稀释,然后在4℃,在30min内将20μl抗体溶液加入细胞中。为去除未结合的抗体,将细胞用facs缓冲液洗涤两次,然后加入稀释后的二抗(pe缀合的affinipuref(ab')2片段,山羊抗人iggfcg片段特异性;jacksonimmunoresearch#109-116-170)。在4℃孵育30min后,洗去未结合的二抗。在测量之前,将细胞重悬于200μlfacs缓冲液中,然后使用bdcantoii设备通过流式细胞术进行分析。如图3所示,优化后的cd3结合剂“cd3opt”和原始cd3结合剂“cd3orig”与jurkat细胞上cd3的结合均相当好。实例3–优化后的cd3结合剂的功能活性在jurkat报告细胞测定中检测优化后的cd3结合剂“cd3opt”的功能活性,并且与原始cd3结合剂“cd3orig”的活性进行比较。未检测igg的功能活性,将表达抗pglala的cho细胞与jurkatnfat报告细胞在浓度不断增加的cd3opt人igg1pglala或cd3orig人igg1pglala存在下一起孵育。在t细胞交联后,jurkatnfat报告细胞上cd3的激活诱导了荧光素酶的产生,并且可测量发光以将其作为激活标志物。包含cd3orig人igg1wt作为阴性对照,其无法与表达抗pglala的cho细胞结合,因此无法在jurkatnfat细胞上发生交联。该测定的示意图提供在图4中。表达抗pglala的cho细胞是经过工程化改造的以在其表面上表达抗体(具体地,结合人igg1fc(pglala)的抗体)的cho-k1细胞(参见wo2017/072210,其通过引用并入本文)。在包含5%fcs+1%glumax的dmem/f12培养基中培养这些细胞。jurkatnfat报告细胞如实例2所述。在cd3huigg1pglala与cho上表达的抗pglala与jurkat-nfat报告细胞上表达的cd3同时结合后,nfat启动子被激活,并且导致活性萤火虫荧光素酶的表达。发光信号的强度(通过添加荧光素酶底物获得)与cd3激活和信号传导的强度成正比。jurkat-nfat报告细胞在悬浮液中生长,并且以0.1-0.5mio细胞/ml、200μg/ml潮霉素的密度在rpmi1640、2g/l葡萄糖、2g/lnahco3、10%fcs、25mmhepes、2mml-谷氨酰胺、1xneaa、1x丙酮酸钠中培养。为了进行测定,收获cho细胞,并且使用vicell测定细胞活力。将30000个靶细胞/孔铺板到100μl培养基中的平底白壁96孔板(greinerbio-one#655098)中,并且将50μl/孔的稀释抗体或培养基(用作对照)添加至cho细胞中。随后,收获jurkat-nfat报告细胞,并且使用vicell评估细胞活力。将细胞以1.2mio细胞/ml的浓度重悬于不含潮霉素b的细胞培养基中,并且以60000个细胞/孔(50μl/孔)的密度加入cho细胞中,以获得2:1的最终效应子与靶标(e:t)之比,并且每孔最终体积为200μl。然后,将4μlglosensor(promega#e1291)加入每个孔中(最终体积的2%)。将细胞在加湿培养箱中于37℃孵育24h。在孵育时间结束时,使用tecanspark10m检测发光。如图5所示,优化后的cd3结合剂cd3opt与cd3orig在交联后在jurkatnfat细胞上具有相似的活性。实例4-包含优化后的cd3结合剂的t细胞双特异性抗体的生成tyrp1tcb利用在实例1中鉴定出的优化后的cd3结合剂(“cd3opt”,seqidno:7(vh)和seqidno:11(vl)),生成t细胞双特异性抗体(tcb),其靶向cd3和tyrp1(“tyrp1tcb”)。该tcb中包含的tyrp1结合剂通过tyrp1结合剂“ta99”的人源化生成(重链和轻链分别参见genbank条目axq57811和axq57813),并且包含分别如seqidno:18和seqidno:22所示的重链和轻链可变区序列。tcb分子的示意图提供于图6中,其完整序列在seqidno:23、seqidno:24、seqidno:25和seqidno:27中给出。还制备了具有原始cd3结合序列的类似分子(seqidno:23、seqidno:24、seqidno:25和seqidno:26)。双特异性分子通过瞬时转染hek293ebna细胞而生成。将细胞用相应的表达载体以1:2:1:1的比率(“载体重链(vh-ch1-vl-ch1-ch2-ch3)”:“载体轻链(vl-cl)”:“载体重链(vh-ch1-ch2-ch3)”:“载体轻链(vh-cl)”)进行转染。对细胞进行离心,然后用经过预热的cdcho培养基(thermofisher,#10743029)。将表达载体在cdcho培养基中混合,加入pei(聚乙烯亚胺,polysciences,#23966-1),将溶液涡旋混合,并且在室温下孵育10分钟。然后,将细胞(2mio/ml)与载体/pei溶液混合,将其转移至烧瓶中,并且置于振荡培养箱中,在5%co2的气氛下于37℃孵育3小时。孵育后,加入包含补充剂的excell培养基(总体积的80%)。转染后一天,加入补充剂(饲料,占总体积的12%)。7天后,通过离心和随后的过滤(0.2μm过滤器)收获细胞上清液。利用标准方法从过滤的细胞培养物上清液中纯化蛋白质。简言之,利用蛋白a亲和色谱法(mabselectsure,gehealthcare:平衡缓冲液:20mm柠檬酸钠,20mm磷酸钠,ph7.5;洗脱缓冲液:20mm柠檬酸钠,100mmnacl,100mm甘氨酸,ph3.0)从细胞培养上清液纯化含fc的蛋白质。在ph3.0下实现洗脱,随后立即中和样品的ph。利用离心(milliporeultra-15(#ufc903096))对蛋白质进行浓缩,并且利用体积排阻色谱(superdex200,gehealthcare)在20mm组氨酸、140mm氯化钠(ph6.0)中将聚集的蛋白质与单体蛋白质分离。通过测量280nm处的吸光度来测定纯化的蛋白质的浓度,其中根据pace等人(1995)(proteinscience4,2411-23)所述的方法,使用基于氨基酸序列计算得出的质量消光系数。在存在和不存在还原剂的情况下,使用labchipgxii(perkinelmer),通过ce-sds分析蛋白质的纯度和分子量(表1)。利用hplc色谱法在25℃测定聚集体含量,该系统使用分析型体积排阻色谱柱(tskgelg3000swxl或up-sw3000),并且在运行缓冲液(分别为25mmk2hpo4、125mmnacl、200mml-精氨酸盐酸盐(ph6.7)或200mmkh2po4、250mmkcl(ph6.2))中平衡(表2)。表1.tyrp1tcb的ce-sds分析(非还原型)。表2.tyrp1tcb的生产和纯化总结。实例5-包含优化后的cd3结合剂的t细胞双特异性抗体与cd3和tyrp1的结合与重组cd3的结合利用spr评估tyrp1tcb与重组cd3的结合,其中使用包含优化后的cd3结合序列(tyrp1tcbcd3opt)或原始cd3结合序列(tyrp1tcbcd3orig)的tcb。spr实验采用biacoret200,其使用hbs-ep作为运行缓冲液(0.01mhepesph7.4、0.15mnacl、0.05%(v/v)表面活性剂p20(gehealthcare))。将tyrp1tcb捕获到cm5传感器芯片表面上,该芯片包含固定化抗体,该抗体具体地结合人igg1fc(pglala)(参见wo2017/072210,其通过引用并入本文)。通过使用标准胺偶联试剂盒(gehealthcare),通过在ph5.0下直接固定约8700个共振单位(ru),使捕获抗体偶联至传感器芯片表面。在5nm下以10μl/min的流速捕获30s的tcb分子。人和食蟹猴抗原(见下文)的浓度为12.35-3000nm,以30μl/min的流速在240s内流过流通池。监测解离阶段240s,并且通过将从样品溶液切换为hbs-ep来触发。在每次循环后,在30s内一次注入10mm甘氨酸(ph2.0),使芯片表面再生。所用的抗原是人或食蟹猴cd3δ和cd3ε胞外结构域与包含杵臼结构修饰和c末端avi标签的人fc结构域融合的异源二聚体(参见seqidno:28和seqidno:29(人cd3)和seqidno:30和seqidno:31(食蟹猴cd3))。通过减去在参比流通池(无捕获的tcb)中获得的响应来校正本体折射率偏差。使用biaeval软件(gehealthcare),通过拟合为1:1langmuir结合,从动力学速率常数中得出亲和常数。对于tyrp1tcbcd3opt,测得的与人和食蟹猴cd3结合的kd值分别为50nm和20nm,并且与tyrp1tcbcd3orig(分别为50nm和40nm)的值相近。这表明在无应激条件下,包含cd3opt或cd3orig的两种tcb与重组cd3的结合均相当好。还在37℃或40℃的温度应激14天后评估tyrp1tcb与重组人cd3的结合,其中使用包含优化后的cd3结合序列或原始cd3结合序列的tcb。如上文实例2所述开展该实验,其中使用tcb代替igg分子。该实验的结果在图7中示出。从图7中可以看出,与包含原始cd3结合剂cd3orig的tcb相比,包含优化后的cd3结合剂cd3opt的tcb在经过应激(在37℃和ph7.4下暴露2周)后表现出显著改善的与cd3的结合。这一结果证实优化后的cd3结合剂(参见实例2)的改善的特性保持在tcb水平。与重组tyrp1的结合利用spr评估与重组tyrp1的结合,其中使用通过相应抗体的血纤维蛋白溶酶酶解所制备的tyrp1fab片段。spr实验采用biacoret200,其使用hbs-ep作为运行缓冲液(0.01mhepesph7.4、0.15mnacl、0.05%(v/v)表面活性剂p20(gehealthcare))。使用标准胺偶联试剂盒(gehealthcare),在ph5.0下将具体结合人igg1fc(pglala)的抗体(参见wo2017/072210,其通过引用并入本文)直接偶联在cm5传感器芯片上。以10μl/min的流速捕获抗原(见下文)30s。使tyrp1fab片段的3倍稀释系列样品以30μl/min的流速在180s内通过流通池,以记录缔合阶段。监测解离阶段180s或1200s,并且通过从样品溶液切换为hbs-ep来触发。在每次循环后,在30s内以30μl/min的流速一次注入10mm甘氨酸(ph2),使芯片表面再生。所用的抗原是与包含杵臼结构(和pglala)修饰以及c末端avi标签的人fc结构域的人、食蟹猴或小鼠tyrp1细胞外结构域(ecd)的单体融合体(参见seqidno:32和seqidno:35(人tyrp1)、seqidno:33和seqidno:35(食蟹猴tyrp1)或seqidno:34和seqidno:35(小鼠tyrp1))。通过减去在参比流通池(无捕获的抗原)中获得的响应来校正本体折射率偏差。使用biaeval软件(gehealthcare),通过拟合为1:1langmuir结合,从动力学速率常数中得出亲和常数(kd)。测得的与人、食蟹猴和小鼠tyrp1结合的kd值分别为130pm、180pm和530pm,并且与亲代ta99抗体的结果类似(分别为90pm、120pm和310pm)。还在37℃或40℃的温度应激14天后评估tyrp1tcb与重组tyrp1的结合,其中使用包含优化后的cd3结合序列或原始cd3结合序列的tcb。如上文有关与cd3的结合所述开展该实验,其中使用重组tyrp1(sinobiologicals)作为抗原。该实验的结果在图8中示出。结果证实,两种tcb(以及相应的igg形式的tyrp1结合剂)与人tyrp1的结合不受应激条件的影响。与jurkat细胞上cd3的结合如上文实例2中所述,利用facs测定包含优化后的cd3结合剂“cd3opt”或原始cd3结合剂“cd3orig”的tyrp1tcb与人报告t细胞系jurkatnfat上cd3的结合。如图9所示,包含优化后的cd3结合剂“cd3opt”的tcb与jurkat细胞上cd3的结合至少与包含原始cd3结合剂“cd3orig”的tcb相当。实例6-包含优化后的cd3结合剂的t细胞双特异性抗体的功能活性cd3激活在jurkatnfat报告细胞测定(参见实例3)中,在tyrp1阳性黑素瘤细胞m150543(原代黑素瘤细胞系,得自苏黎世大学皮肤细胞库)存在下,检测包含优化后的cd3结合剂cd3opt或原始cd3结合剂cd3orig的tyrp1tcb(实例4)。在tyrp1tcb与tyrp1阳性靶细胞和cd3抗原(在jurkat-nfat报告细胞上表达)同时结合后,nfat启动子被激活,并且导致活性萤火虫荧光素酶的表达。发光信号的强度(通过添加荧光素酶底物获得)与cd3激活和信号传导的强度成正比。如实例3中所述开展该测定,其中使用m150543代替表达抗pglala的cho细胞。如igg的结果所示(实例3),包含cd3opt或cd3orig的两种tcb在jurkatnfat报告细胞上具有相似的功能活性,并且以依赖于浓度的方式诱导cd3激活(图10)。靶细胞杀伤下一步,在肿瘤细胞杀伤测定中,通过将两种tcb分子与来自三个不同供体的新鲜分离的人pbmc与人黑素瘤细胞系m150543一起孵育,对其进行检测。通过24h和48h后的ldh释放来测定肿瘤细胞裂解。在48h后,通过两个细胞亚群中cd69和cd25的上调来分析cd4和cd8t细胞的激活。简言之,用胰蛋白酶/edta收获靶细胞,将其洗涤,并且使用平底96孔板以30000个细胞/孔的密度铺板。使细胞粘附过夜。外周血单核细胞(pbmc)通过对健康人供体的新鲜血液进行histopaque密度离心来制备。将新鲜血液用无菌pbs稀释,并且铺到histopaque梯度液(sigma#h8889)上。离心(450xg,30min,室温)后,弃去包含pbmc的中间相上方的血浆,并且将pbmc转移至新的falcon管中,随后用50mlpbs填充。离心混合物(400xg,10min,室温),弃去上清液,将pbmc颗粒用无菌pbs洗涤两次(离心步骤350xg,10分钟)。对所得的pbmc群体自动计数(vicell),并且在37℃和5%co2的细胞培养箱中储存于包含10%fcs和1%l-丙氨酰-l-谷氨酰胺的rpmi1640培养基(biochrom#k0302)中直至进一步使用(不长于24h)。为了进行杀伤测定,加入指定的浓度的抗体并且重复三次。将pbmc以10:1的最终效应子与靶标(e:t)之比加入靶细胞中。在37℃、5%co2下孵育24h后,通过凋亡/坏死细胞(ldh检测试剂盒,rocheappliedscience#11644793001)对释放到细胞上清液中的ldh进行定量分析,评估靶细胞杀伤作用。通过将靶细胞与1%tritonx-100一起孵育,实现最大程度的靶细胞裂解(=100%)。最小程度裂解(=0%)是指靶细胞与不含双特异性构建体的效应细胞一起孵育。利用流式细胞术评估tcb介导靶细胞的t细胞杀伤后cd8和cd4t细胞的激活,其中使用识别t细胞激活标志物cd25(晚期激活标志物)和cd69(早期激活标志物)的抗体。孵育48h后,将pbmc转移至圆底96孔板中,在350xg下离心5min,并且用facs缓冲液洗涤两次。根据供应商的说明对cd4apc(biolegend#300514)、cd8fitc(biolegend#344704)、cd25bv421(biolegend#302630)和cd69pe(biolegend#310906)进行表面染色。将细胞用150μl/孔的facs缓冲液洗涤两次,并且使用100μl/孔的固定缓冲液(bd#554655)在4℃固定15min。离心后,将样品重悬于200μl/孔的facs缓冲液中。利用bdfacsfortessa分析样品。在所有三种供体上,包含优化后的cd3结合剂或原始cd3结合剂的两种tcb以相似的方式诱导t细胞激活和肿瘤细胞裂解(图11)。48h后,所有三种供体的肿瘤细胞裂解ec50值汇总于表3中。表3.tyrp1tcb在48h时的肿瘤细胞杀伤ec50值总结。实例7–小鼠中t细胞双特异性抗体的pk研究向人fcrn转基因小鼠(line32,纯合)和fcrn基因敲除小鼠(jacksonlaboratory品系编号003982和014565)(n=3/品系/测试化合物)静脉推注1mg/kg的剂量后,研究包含不同cd3结合剂(cd3orig和cd3opt)的tyrp1tcb的药代动力学(pk)。从人fcrn转基因(tg)小鼠中连续采集微量血液样品长达672h(在给药后5min至672h,对于每只小鼠,采集9个样品),并且在fcrn基因敲除(ko)小鼠中连续采集微量血液样品长达96h(在给药后5min至96h,对于每只小鼠,采集8个样品)。制备血清,并且将其冷冻保存直至分析。使用e411(roche)仪器,在非glp条件下用对人ig/fabch1/κ结构域具有特异性的通用eclia方法对小鼠血清样品进行分析。使用标准非区室分析进行药代动力学评估。本研究的结果如表4中所示。结果表明,cdr的工程化改造未引起可能影响抗体清除的其他序列不利因素。cd3opt与cd3orig在血清半衰期方面同样出色,同时具有cdr稳定性增加的额外的益处。表4.hufcrntg小鼠和fcrnko小鼠的清除率数据(ml/天/kg;平均值和(cv))。实例8–包含优化后的cd3结合剂的另一种t细胞双特异性抗体的生成利用在实例1中鉴定出的优化后的cd3结合剂(“cd3opt”,seqidno:7(vh)和seqidno:11(vl)),生成t细胞双特异性抗体(tcb),其靶向cd3和egfrviii(“egfrviiitcb”)。该tcb(p063.056)中包含的egfrviii结合剂来自噬菌体展示,随后亲和力成熟(见下文),并且分别包含seqidno:88和seqidno:92中所示的重链可变区序列和轻链可变区序列。tcb分子的示意图提供于图6中,其完整序列在seqidno:109、seqidno:110、seqidno:111和seqidno:27中给出。还制备了具有原始cd3结合序列的类似分子(seqidno:109、seqidno:110、seqidno:111和seqidno:26)。通过瞬时转染hek293ebna细胞生成双特异性分子,并且如上文实例4所述的方法进行纯化和分析。此外,以类似的方式(对于igg重链和轻链,以1:1的比率用表达载体转染hekebna细胞),产生人igg1形式的源自噬菌体展示的egfrviii抗体,其使用如下文所述。如体积排阻色谱法所测定的,所有igg和tcb构建体以相当的质量得到纯化,单体含量超过95%。egfrviii抗体的选择egfrviii抗体源自噬菌体展示并且是亲和力成熟的。使用稳定表达egfrviii的cho细胞以及egfrviii阳性人胶质母细胞瘤细胞系dk-mg,测试对egfrviii(p056.021(seqidno:40和seqidno:44)、p056.052(seqidno:48和seqidno:52)、p047.019(seqidno:56和seqidno:60)、p057.012(seqidno:64和seqidno:68)、p057.011(seqidno:72和seqidno:76)、p056.027(seqidno:80和seqidno:84))表现出高亲和力结合和特异性的抗体与细胞表面上表达的egfrviii的结合。为证实特异性并且排除与野生型egfr(egfrwt)的交叉反应性,测试所选抗体与egfrwt阳性人肿瘤细胞系mkn-45的结合(图12)。包含西妥昔单抗作为用于与egfrwt结合的阳性对照,并且包含非靶向dp47igg作为阴性对照。所有选定的抗体具体地与egfrviii结合而与egfrwt无交叉反应,并且考虑对其进行进一步表征。下一步,将dk-mg细胞与表达抗pglala嵌合抗原受体(car)的jurkatnfat报告细胞一起孵育,通过测量发光来评估作为igg1pglala的这些egfrviii抗体(人igg1形式,在fc区中包含p329gl234al235a(“pglala”,根据eu编号)突变)功能活性(carj测定,参见pct申请号pct/ep2018/086038,其全文通过引用并入本文)。包含dp47igg1pglala作为阴性对照。所有测试的egfrviii抗体均诱导表达car的jurkatnfat报告细胞发生显著激活(图13)。除p047.019表现出最弱的结合和激活外,选择将测试的所有其他egfrviii抗体转化为tcb形式(以cdorig作为cd3结合剂)。比较转化为tcb形式的所选的egfrviii抗体与cho-egfrviii细胞的结合与相应igg的结合(图14),以证实转化为tcb形式对egfrviii抗体结合能力无影响。大多数测试的egfrviii克隆在转化为tcb形式后仍然保留与egfrviii的结合能力;仅克隆p057.011在tcb形式下与egfrviii的结合与相应的igg相比略有降低(表5)。表5.egfrviiiigg和tcb与cho-egfrviii(ec50)的结合。egfrviii克隆ec50igg(nm)ec50tcb(nm)p056.02116.513.0p056.02713.115.9p056.05218.219.5p057.0123.05.5p057.0115.312.8随后,在针对egfrviii阳性dk-mg细胞的jurkatnfat报告细胞测定中,对egfrviiitcb的功能活性进行测试(图15)。所有测试的egfrviiitcb均在jurkatnfat报告细胞测定中具有活性,其中p056.021最有效,其次是具有相似活性的p056.027、p056.052和p057.012,而p057.011的活性最低。接下来,在肿瘤细胞裂解测定中,使用与dk-mg或mkn-45细胞共培养的pbmc测试egfrviiitcb,以排除egfrviiitcb与egfrwt的交叉反应性(图16)。在该测定中,除肿瘤细胞裂解以外,还测量t细胞激活(图17)和细胞因子释放(图18)作为额外的读出。如之前的报告细胞测定所示,egfrviiitcbp056.021对egfrviii阳性具有最高的活性,而对egfrwt阳性细胞没有任何活性。egfrviiitcbp057.011对egfrwt细胞表现出非特异性活性,因此被排除在外。egfrviiitcbp056.027、p056.052和p057.012具有可比的活性。基于这些结果,选择egfrviii结合剂p056.021和p057.012进行另一轮亲和力成熟。无法由p057.012得出良好的结合剂(结果未显示)。利用spr测得的源自p056.021所选egfrviii结合剂对egfrviii的亲和力和特异性如表6所示。表6.利用spr测得的所选egfrviii结合剂对egfrviii的亲和力和特异性。还比较了亲和力成熟的egfrviii结合剂(p063.056(seqidno:88和seqidno:92)、p064.078(seqidno:96和seqidno:100)、p065.036(seqidno:104和seqidno:108))与亲代结合剂对u87mg-egfrviii和mkn-45细胞上的egfrviii的特异性结合(图19)。选择对egfrviii具有最佳亲和力和特异性的egfrviii结合剂p063.056,将其转化为tcb形式,以cd3orig或cd3opt作为cd3结合剂。在jurkatnfat报告细胞测定中,比较egfrviiitcbp063.056(包含cd3opt或cd3orig)与亲代egfrviiitcbp056.021在u87mg-egfrviii、dk-mg和mkn-45细胞上的功能活性(图20)。所有三种tcb仅在egfrviii阳性细胞存在下诱导jurkatnfat特异性激活。egfrviiitcbp063.056的活性略高于亲代egfrviiitcbp056.021。方法表面等离子体共振在biacoret200上利用表面等离子体共振测定egfrviii抗体与egfrviii的亲和力,其中使用hbs-ep作为运行缓冲液(0.01mhepesph7.4,0.15mnacl,0.005%(v/v)表面活性剂p20;gehealthcare)在25℃进行测定。用抗体(具体地结合固定在cm5芯片上的人igg1fc(pglala)(参见wo2017/072210,其通过引用并入本文))在30s内捕获25nm的抗egfrviiipglalaigg。egfrviii-ecdavihis抗原(见下文,实例9)的浓度为12.4-1000nm,以30μl/min的流速在200s内流过流通池。监测解离阶段300s,并且通过将从样品溶液切换为hbs-ep来触发。在每次循环后,在30s内两次注入10mm甘氨酸(ph2.0),使芯片表面再生。通过减去在参比流通池中获得的响应来校正本体折射率偏差。使用biaeval软件(gehealthcare),通过拟合为1:1langmuir结合,从动力学速率常数中得出亲和常数。在特异性测定中,用固定在cm5芯片上的抗his(pentahis,qiagen)在40s捕获100nm的egfrviii和egfrwtecd抗原。在60s内单次注入浓度为500nm的抗egfrviii抗体,然后用10mm甘氨酸ph2.0在60s内使其再生。观察到egfrviii结合的应答单位高于50。高于5个应答单位(ru)的应答被认为对egfrwt结合呈阳性,并且将应答低于5ru的igg归类为对egfrwt具有特异性。细胞系jurkat-nfat报告细胞(gloresponsejurkatnfat-re-luc2p;promega#cs176501)是一种包含nfat启动子的人急性淋巴白血病报告细胞系,其表达人cd3。将细胞以0.1-0.5mio细胞/ml的密度在rpmi1640、2g/l葡萄糖、2g/lnahco3、10%fcs、25mmhepes、1%glutamax、1xneaa、1x丙酮酸钠中培养。每当细胞传代时,添加最终浓度为200μg/ml潮霉素b。包含pglalacar的jurkatnfat细胞在内部生成。原始细胞系(jurkatnfat;signosis)是一种人急性淋巴白血病报告细胞系,其nfat启动子在经由人cd3的激活后导致荧光素酶表达。它们经过工程化改造以表达能够识别p293glala突变的嵌合抗原受体。培养时,该细胞悬于补充有10%fcs和1%谷氨酰胺的rpmi1640中,并且维持在0.4-1.5mio细胞/ml之间。cho-egfrviii细胞在内部生成。用egfrviii稳定地转导cho-k1细胞。在包含5%fcs、1%glutamax和6μg/ml嘌呤霉素的dmem/f12培养基中培养细胞。dk-mg(dsmz#acc277)是人胶质母细胞瘤细胞系。通过细胞分选富集表达egfrviii的dk-mg细胞。在rpmi1860、10%fcs和1%glutamax中培养细胞。u87mg-egfrviii(atcchtb-14)是人胶质母细胞瘤细胞系,其经过egfrviii稳定转导。在dmem、10%fcs和1%glutamax中培养细胞。mkn-45(dsmzacc409)是表达高水平egfrwt的人胃腺癌细胞。在包含2%fcs和1%glutamax的高级rpmi1640中培养细胞。利用流式细胞术测定靶向结合收获用于结合实验的细胞,用pbs洗涤,并且重悬于facs缓冲液中。在96孔圆形底板中进行抗体染色。收获细胞,计数,并且在每个孔中接种100000至200000个细胞。在400xg下将孔板离心4min,并且除去上清液。将检测抗体用facs稀释液稀释,然后在4℃,在30min内将20μl抗体溶液加入细胞中。为去除未结合的抗体,将细胞用facs缓冲液洗涤两次,然后加入稀释后的二抗pe缀合的affinipuref(ab')2片段,山羊抗人iggfcg片段特异性(jacksonimmunoresearch,#109-116-170或#109-116-098)。在4℃孵育30min后,洗去未结合的二抗。在测量之前,将细胞重悬于200μlfacs缓冲液中,使用bdcantoii或bdfacsfortessa通过流式细胞术进行分析。使用egfrviiipglalaigg进行carjnfat报告细胞测定使用carjnfat报告细胞测定来评估egfrviiipglalaigg诱导t细胞激活的效能。该测定的原理是将jurkat-nfat工程化改造的效应细胞与表达肿瘤抗原的癌细胞共培养。仅经由pglala突变和靶抗原egfrviii将igg与car同时结合时,nfat启动子才被激活,并且导致jurkat效应细胞中荧光素酶表达的增加。加入适当的底物后,活性萤火虫荧光素酶导致发光,这可以作为car介导的激活信号进行测量。简言之,收获靶细胞并且测定其活力。在测定开始前一天,将30000个靶细胞/孔铺板到100μl培养基中的平底白壁96孔板(greinerbio-one,#655098)中。第二天,除去培养基,并且向靶细胞中加入25μl/孔的稀释抗体或培养基(用于对照)。随后,收获jurkat-nfat报告细胞,并且使用vicell评估细胞活力。将细胞以1.5mio细胞/ml的浓度重悬于细胞培养基中,并且以75000个细胞/孔(50μl/孔)的密度加入肿瘤细胞中,以获得2.5:1的最终效应子与靶标(e:t)之比,并且每孔最终体积为75μl。然后将4μlglosensor(promega,#e1291)加入每个孔中(最终体积的2%)。将细胞在加湿培养箱中于37℃孵育24h。在孵育时间结束时,将平板调整至室温(约15min)。然后加入25μl/孔的one-glo荧光素酶(promega,#e6120),并且将孔板在暗处孵育15min,然后使用tecanspark检测发光。使用egfrviiitcb进行jurkatnfat报告细胞测定使用egfrviii阳性细胞和jurkat-nfat报告细胞,评估包含改善的cd3结合剂或原始cd3结合剂的egfrviiitcb诱导t细胞交联及随后的t细胞激活的能力。在egfrviiitcb与egfrviii阳性靶细胞和cd3抗原(在jurkat-nfat报告细胞上表达)同时结合后,nfat启动子被激活,并且导致活性萤火虫荧光素酶的表达。发光信号的强度(通过添加荧光素酶底物获得)与cd3激活和信号传导的强度成正比。为了进行测定,收获靶细胞,并且测定细胞活力。将30000个靶细胞/孔铺板到100μl培养基中的平底白壁96孔板(greinerbio-one,#655098)中,并且将50μl/孔的稀释抗体或培养基(用作对照)添加至靶细胞中。随后,收获jurkat-nfat报告细胞,并且使用vicell评估细胞活力。将细胞以1.2mio细胞/ml的浓度重悬于不含潮霉素b的细胞培养基中,并且以60000个细胞/孔(50μl/孔)的密度加入肿瘤细胞中,以获得2:1的最终效应子与靶标(e:t)之比,并且每孔最终体积为200μl。然后将4μlglosensor(promega,#e1291)加入每个孔中(最终体积的2%)。将细胞在加湿培养箱中于37℃孵育24h。在孵育时间结束时,使用tecanspark检测发光。t细胞介导的肿瘤细胞杀伤用胰蛋白酶/edta收获靶细胞,将其洗涤,并且使用平底96孔板以30000个细胞/孔的密度铺板。使细胞粘附过夜。外周血单核细胞(pbmc)通过对健康人供体的新鲜血液进行histopaque密度离心来制备。将新鲜血液用无菌pbs稀释,并且铺到histopaque梯度液(sigma,#h8889)上。离心(450xg,30min,室温)后,弃去包含pbmc的中间相上方的血浆,并且将pbmc转移至新的falcon管中,随后用50mlpbs填充。离心混合物(400xg,10min,室温),弃去上清液,将pbmc颗粒用无菌pbs洗涤两次(离心步骤350xg,10分钟)。对所得的pbmc群体自动计数(vicell),并且在37℃和5%co2的细胞培养箱中储存于包含10%fcs和1%glutamax的rpmi1640培养基中直至进一步使用(不长于24h)。为了进行杀伤测定,加入指定的浓度的抗体并且重复三次。将pbmc以10:1的最终效应子与靶标(e:t)之比加入靶细胞中。在37℃、5%co2下孵育24h后,通过凋亡/坏死细胞(ldh检测试剂盒,rocheappliedscience,#11644793001)对释放到细胞上清液中的ldh进行定量分析,评估靶细胞杀伤作用。通过将靶细胞与1%tritonx-100一起孵育,实现最大程度的靶细胞裂解(=100%)。最小程度裂解(=0%)是指靶细胞与不含双特异性构建体的效应细胞一起孵育。利用流式细胞术评估tcb介导靶细胞的t细胞杀伤后cd8和cd4t细胞的激活,其中使用识别t细胞激活标志物cd25(晚期激活标志物)和cd69(早期激活标志物)的抗体。孵育48h后,将pbmc转移至圆底96孔板中,在350xg下离心5min,并且用facs缓冲液洗涤两次。根据供应商的说明对cd4apc(biolegend,#300514)、cd8fitc(biolegend,#344704)、cd25bv421(biolegend,#302630)和cd69pe(biolegend,#310906)进行表面染色。将细胞用150μl/孔的facs缓冲液洗涤两次,并且使用100μl/孔的固定缓冲液(bd,#554655)在4℃固定15min。离心后,将样品重悬于200μl/孔的facs缓冲液中。利用bdfacsfortessa分析样品。根据制造商的说明,使用流式细胞术微珠阵列(cba),通过流式细胞术测量上清液中的细胞因子分泌,但并非使用50μl微珠和样品,而是仅使用25μl上清液和微珠。使用以下cba试剂盒(bdbiosciences):cba人干扰素γ(ifnγ)flex套装、cba人颗粒酶βflex套装和cba人tnfflex套装。使用bdfacscantoii或bdfacsfortessa测量样品,并且使用diva软件(bdbiosciences)进行分析。实例9–包含优化后的cd3结合剂的t细胞双特异性抗体与cd3和egfrviii的结合与重组cd3的结合利用spr评估egfrviiitcb与重组cd3的结合,其中使用包含优化后的cd3结合序列(egfrviiitcbcd3opt)或原始cd3结合序列(egfrviiitcbcd3orig)的tcb,如上文实例5中针对tyrp1tcb所述。使用标准胺偶联试剂盒(gehealthcare),通过在ph5.0下直接固定约5200个共振单位(ru),使捕获抗体偶联至传感器芯片表面,并且在20nm下以10μl/min的流速捕获30s的tcb分子。对于tyrp1tcbcd3opt,测得的与人和食蟹猴cd3结合的kd值分别为30nm和20nm,并且与tyrp1tcbcd3orig(分别为40nm和30nm)的值相近。这表明在无应激条件下,包含cd3opt或cd3orig的两种tcb与重组cd3的结合均相当好。还在37℃或40℃的温度应激14天后评估egfrviiitcb与重组人cd3的结合,其中使用包含优化后的cd3结合序列或原始cd3结合序列的tcb。如上文实例2所述开展该实验,其中使用tcb代替igg分子。该实验的结果在图21中示出。从图21中可以看出,与包含原始cd3结合剂cd3orig的tcb相比,包含优化后的cd3结合剂cd3opt的tcb在经过应激(在37℃和ph7.4下暴露2周)后表现出显著改善的与cd3的结合。这一结果再次证实,优化后的cd3结合剂(参见实例2)的改善的特性保持在tcb水平。与重组egfrviii的结合利用spr评估egfrviiitcb与重组egfrviii的结合。spr实验采用biacoret200,其使用hbs-ep作为运行缓冲液(0.01mhepesph7.4、0.15mnacl、0.05%(v/v)表面活性剂p20(gehealthcare))。使用标准胺偶联试剂盒(gehealthcare),在ph5.0下将抗fc抗体(gehealthcare)直接偶联在cm5传感器芯片上。以10μl/min的流速捕获egfrviiitcb(5nm)30s。使egfrviii抗原的3倍稀释系列样品以30μl/min的流速在200s下通过流通池,以记录缔合阶段。监测解离阶段300s,并且通过从样品溶液切换为hbs-ep来触发。在每次循环后,在30s内以20μl/min的流速一次注入3mmgcl2,使芯片表面再生。所用的抗原包含与c末端的avi标签和his标签融合的人egfrviii的细胞外结构域(egfrviii-ecdavihis;seqidno:36)。通过减去在参比流通池(无捕获的tcb)中获得的响应来校正本体折射率偏差。使用biaeval软件(gehealthcare),通过拟合为1:1langmuir结合,从动力学速率常数中得出亲和常数(kd)。利用动力学分析通过速率常数近似计算表观亲和常数kd,其中在该2:1相互作用下采用1:1结合拟合。对于包含cd3opt或cd3orig的egfrviiitcb,测得与人egfrviii结合的kd值(亲和力)为6nm。还在37℃或40℃的温度应激14天后评估egfrviiitcb与重组egfrviii的结合,其中使用包含优化后的cd3结合序列或原始cd3结合序列的tcb。如上文实例5所述开展该实验,其中使用egfrviii-ecdavihis作为抗原(见上文)。该实验的结果在图22中示出。结果证实,两种tcb与人egfrviii的结合不受应激条件的影响。与jurkat细胞上cd3的结合如上文实例2中所述,利用facs测定包含优化后的cd3结合剂“cd3opt”或原始cd3结合剂“cd3orig”的egfrviiitcb与人报告t细胞系jurkatnfat上cd3的结合。如图23所示,包含优化后的cd3结合剂“cd3opt”或原始cd3结合剂“cd3orig”的tcb与jurkat细胞上cd3的结合均相当好。与u87mg-egfrviii细胞上egfrviii的结合利用facs测定包含egfrviii结合剂p063.056(cd3opt或cd3orig)的egfrviiitcb或egfrviii克隆p056.021(包含cd3orig)与人胶质母细胞瘤细胞系u87mg-egfrviii上egfrviii的结合。还包含呈igg形式的egfrviii结合剂p063.056。如图24所示,所有三种tcb均以高亲和力与u87mg-egfrviii细胞上表达的egfrviii结合,并且结合不受转化为tcb形式的影响。实例10-包含优化后的cd3结合剂的t细胞双特异性抗体的功能活性cd3激活在jurkatnfat报告细胞测定中,在egfrviii阳性胶质母细胞瘤细胞dk-mg、u87mg-huegfrviii和egfrwt阳性mkn45细胞存在下(如上文实例8中所述),测试包含所选egfrviii结合剂(p063.056)以及优化后的cd3结合剂cd3opt或原始cd3结合剂cd3orig的egfrviiitcb。如igg的结果所示(实例3),包含cd3opt或cd3orig的两种tcb在jurkatnfat报告细胞上具有相似的功能活性,并且以依赖于浓度的方式诱导cd3激活(图20)。靶细胞杀伤在肿瘤细胞杀伤实验中,在胶质母细胞瘤细胞系u87mg-egfrviii和pbmc存在下(如上文实例8中所述),比较包含所选egfrviii结合剂(p063.056)以及优化后的cd3结合剂cd3opt或原始cd3结合剂cd3orig的egfrviiitcb与包含亲代egfrviii结合剂p056.021和cd3结合剂cd3orig的egfrviiitcb。与在jurkatnfat报告细胞中观察到的一样,在诱导肿瘤细胞裂解和cd4和cd8t细胞的激活方面,包含cd3opt或cd3orig的tcb具有相似的功能活性,该功能活性通过cd69上调测得(图25)。此外,比较包含egfrviii结合剂p063.056和优化后的cd3结合剂cd3opt的呈“2+1形式”的egfrviiitcb(如图6中所示)与包含相同的egfrviii和cd3结合剂的呈“1+1头到尾形式”的egfrviiitcb(示意性地示出于图1g中)的功能活性。在jurkatnfat报告细胞测定和肿瘤细胞杀伤测定中,使用如上文实例8中所述的胶质母细胞瘤细胞系u87mg-egfrviii测试这两种egfrviiitcb。呈2+1形式的egfrviiitcb在jurkatnfat报告细胞测定中测得的cd3激活(图26)以及在使用pbmc的杀伤测定中诱导肿瘤细胞杀伤和t细胞激活(图27)方面均具有优异的功能活性。实例11-包含优化后的cd3结合剂的t细胞双特异性抗体的功能表征egfrviiitcb的t细胞增殖和激活作用在u87mg-egfrviii细胞上的t细胞增殖测定中,比较包含所选egfr结合剂(p063.056)以及优化后的cd3结合剂cd3opt或原始cd3结合剂cd3orig的egfrviiitcb与包含亲代egfrviii结合剂p056.021和cd3结合剂cd3orig的egfrviiitcb的功能活性(图28)。所有三种tcb均诱导了cd4t细胞和cd8t细胞的显著增殖和激活。包含cd3opt的p063.056egfrviiitcb具有高于另外两种egfrviiitcb的活性。利用egfrviiitcb进行肿瘤细胞裂解接下来,在pbmc与dk-mg细胞共培养的肿瘤细胞裂解测定中,比较包含所选egfr结合剂(p063.056)和优化后的cd3结合剂cd3opt的egfrviiitcb与包含亲代egfrviii结合剂p056.021和cd3结合剂cd3orig的egfrviiitcb(图29)。在该测定中,除肿瘤细胞裂解以外,还测量t细胞激活和细胞因子释放作为额外的读出。如前所述,在肿瘤细胞裂解、t细胞激活以及ifnγ和tnfα的释放方面,包含cd3opt的p063.056egfrviiitcb的活性高于包含cd3orig的p056.021egfrviiitcb。利用tyrp1tcb进行t细胞激活和肿瘤细胞裂解通过将原代黑素瘤细胞系m150543与分离自健康供体的pbmc共培养,测试tyrp1tcb诱导细胞因子释放的功能特性。在处理24h和48h后,分析由t细胞经由tyrp1tcb介导的肿瘤细胞裂解(图30)。在处理48h后,分析释放至上清液中的ifnγ和tnfα以及cd4和cd8t细胞激活。tyrp1tcb能够在24h后诱导有效的肿瘤细胞裂解。这一作用伴随通过cd25的上调所确定的cd4和cd8t细胞的显著激活以及ifnγ和tnfα的大量释放。方法pbmc分离外周血单核细胞(pbmc)通过对健康人供体的新鲜血液进行histopaque密度离心来制备。将新鲜血液用无菌pbs稀释,并且铺到histopaque梯度液(sigma,#h8889)上。离心(450xg,30min,室温)后,弃去包含pbmc的中间相上方的血浆,并且将pbmc转移至新的falcon管中,随后用50mlpbs填充。离心混合物(400xg,10min,室温),弃去上清液,将pbmc颗粒用无菌pbs洗涤两次(离心步骤350xg,10分钟)。对所得的pbmc群体自动计数(vicell),并且在37℃和5%co2的细胞培养箱中储存于包含10%fcs和1%glutamax的rpmi1640培养基中直至进一步使用(不长于24h)或冷冻并且储存于液氮中直至进一步使用。在使用前一天,将冷冻的pbmc解冻,并且在37℃培养基中培养过夜。t细胞增殖简言之,收获靶细胞,计数,并且用pbs洗涤两次。将细胞以5mio细胞/ml的密度重悬于pbs中。在37℃,用最终浓度为5μm的细胞增殖染料efluor670(ebioscience,#65-0840-85)对细胞染色10min。为终止染色反应,将4倍体积的冷的完全细胞培养基加入细胞悬浮液中,并且在4℃孵育5min,然后用培养基洗涤三次。对标记的靶细胞计数,并且在rpmi1640、10%fcs和1%glutamax中调整为0.1mio细胞/ml。以10000个靶细胞/孔的密度接种到96孔板中。然后以指定的浓度添加处理液,最后向每个孔中加入分离自健康供体的100000个pbmc。将细胞在37℃孵育5天,然后收获pbmc,并且用cd3buv395(biolegend,#563548)、cd4pe(biolegend,#300508)、cd8apc(biolegend,#344722)、cd25pe/cy7(biolegend,#302612)进行染色。通过用流式细胞仪(facsfortessa,bdbioscience)测得的cd4t细胞和cd8t细胞稀释efluor670染料来确定增殖,并且通过测量cd25的上调来测定cd4和cd8t细胞的激活。t细胞介导的肿瘤细胞杀伤用胰蛋白酶/edta收获靶细胞,将其洗涤,并且使用平底96孔板以30000个细胞/孔的密度铺板。使细胞粘附过夜。为了进行杀伤测定,加入指定的浓度的抗体并且重复三次。将pbmc以10:1的最终效应子与靶标(e:t)之比加入靶细胞中。在37℃、5%co2下孵育24h后,通过凋亡/坏死细胞(ldh检测试剂盒,rocheappliedscience,#11644793001)对释放到细胞上清液中的ldh进行定量分析,评估靶细胞杀伤作用。通过将靶细胞与1%tritonx-100一起孵育,实现最大程度的靶细胞裂解(=100%)。最小程度裂解(=0%)是指靶细胞与不含双特异性构建体的效应细胞一起孵育。t细胞激活利用流式细胞术评估tcb介导靶细胞的t细胞杀伤后cd8和cd4t细胞的激活,其中使用识别t细胞激活标志物cd25(晚期激活标志物)和cd69(早期激活标志物)的抗体。孵育48h后,将pbmc转移至圆底96孔板中,在350xg下离心5min,并且用facs缓冲液洗涤两次。根据供应商的说明对cd4apc(biolegend,#300514)、cd8fitc(#344704,biolegend)、cd25bv421(biolegend,#302630)和cd69pe(biolegend,#310906)进行表面染色。将细胞用150μl/孔的facs缓冲液洗涤两次,并且使用100μl/孔的固定缓冲液(bd,#554655)在4℃固定15min。离心后,将样品重悬于200μl/孔的facs缓冲液中。利用bdfacsfortessa分析样品。细胞因子分泌根据制造商的说明,使用流式细胞术微珠阵列(cba),通过流式细胞术测量上清液中的细胞因子分泌,但并非使用50μl微珠和样品,而是仅使用25μl上清液和微珠。使用以下cba试剂盒(bdbiosciences):cba人干扰素γ(ifnγ)flex套装和cba人tnfflex套装。使用bdfacscantoii或bdfacsfortessa测量样品,并且使用diva软件(bdbiosciences)进行分析。实例13–包含优化后的cd3结合剂的t细胞双特异性抗体的体内功效在人肿瘤细胞系的异种移植小鼠模型(igr-1黑素瘤异种移植模型)中,测试tyrp1tcb(包含在实例1中鉴定出的优化后的cd3结合剂)的抗肿瘤功效。在包含10%fcs(sigma)的dmem培养基中培养igr-1细胞(人黑素瘤)。在5%co2下,在水饱和的气氛中于37℃下培养细胞。利用第6代进行移植。细胞活力为96.7%。使用1ml结核菌素注射器(bdbiosciences,德国),以每只动物2×106个细胞的量,将100μlrpmi细胞培养基(gibco)中的细胞皮下注射到小鼠胁腹。根据规定指南(gv-solas;felasa;tierschg),将完全人源化nsg雌性小鼠(rocheglycartag,瑞士)维持在不含特定病原体的条件下,日循环为12h光照/12h黑暗。实验研究方案经过地方政府的审查和批准(zh223/2017)。定期进行连续健康状态监测。在研究第0天,向小鼠皮下注射2×106个igr-1细胞,随机分组并且称重。在注射肿瘤细胞后二十天(肿瘤体积>200mm3),每周两次向小鼠静脉内注射10μg(0.5mg/kg)tyrp1tcb,并且持续五周。向所有小鼠静脉内注射200μl适当的溶液。给媒介物组中的小鼠注射组氨酸缓冲液,向实验组中的小鼠注射tyrp1tcb构建体。为了在每200μl中获得适量的抗体,如有必要,将储备液用组氨酸缓冲液稀释。每周用卡尺测量三次肿瘤大小,并且使用grahpadprism软件作图,体积单位为mm3+/-sem。使用jmp12软件进行统计分析。图31显示,与媒介物组相比,tyrp1tcb在抑制肿瘤生长方面具有显著的功效(68%tgi,p=0.0058*)。在人肿瘤细胞系的异种移植小鼠模型(u87-egfrviii胶质母细胞瘤异种移植模型)中,类似地测试egfrviiitcb(包含在实例1中鉴定出的优化后的cd3结合剂)的抗肿瘤功效。u87细胞(人胶质母细胞瘤)最初得自atcc(manassas,usa),并且被稳定转染以表达人egfrviii蛋白质(rocheglycartag,瑞士)。扩增后,将细胞存放在rocheglycart内部细胞库中。在包含10%fcs(sigma)和0.5μg/ml嘌呤霉素(invitrogen)的dmem培养基中培养u87-egfrviii细胞系。在5%co2下,在水饱和的气氛中于37℃下培养细胞。利用第8代进行移植。细胞活力为94.7%。使用1ml结核菌素注射器(bdbiosciences,德国),以每只动物5×105个细胞的量,将100μlrpmi细胞培养基(gibco)中的细胞皮下注射到小鼠胁腹。根据规定指南(gv-solas;felasa;tierschg),将完全人源化nsg雌性小鼠(rocheglycartag,瑞士)维持在不含特定病原体的条件下,日循环为12h光照/12h黑暗。实验研究方案经过地方政府的审查和批准(zh223/2017)。定期进行连续健康状态监测。在研究第0天,向小鼠皮下注射5×105个u87-egfrviii细胞,随机分组并且称重。在注射肿瘤细胞后两周(肿瘤体积>200mm3),每周两次向小鼠静脉内注射10μg(0.5mg/kg)egfrviiitcb,并且持续三周。向所有小鼠静脉内注射200μl适当的溶液。给媒介物组中的小鼠注射组氨酸缓冲液,向实验组中的小鼠注射egfrviiitcb构建体。为了在每200μl中获得适量的抗体,如有必要,将储备液用组氨酸缓冲液稀释。每周用卡尺测量三次肿瘤大小,并且使用grahpadprism软件作图,体积单位为mm3+/-sem。图32表明,egfrviiitcb在控制肿瘤生长方面具有显著的功效,所有小鼠均达到完全缓解。实例12–egfrviiitcb在小鼠中的pk研究在向人fcrn转基因(line32,纯合)小鼠和nod-scid小鼠静脉推注1mg/kg的剂量后,研究包含优化后的cd3结合剂(cd3opt)的egfrviiitcb的药代动力学(pk)。从人fcrn转基因(tg)小鼠和nod-scid小鼠中连续采集微量血液样品长达672h(在给药后5min至672h,对于每只小鼠,采集9个样品)。使用特异性酶联免疫吸附测定(elisa),在非glp条件下分析经过egfrviiitcb处理的小鼠血清样品。在链霉亲和素包被的微量滴定板(sa-mtp)上,用生物素化egfrviii抗原(huegfrviiihis生物素)捕获egfrviiitcb。利用洋地黄毒苷标记的抗人igg1fc(pglala)的单克隆抗体(参见实例3),然后添加抗洋地黄毒苷-pod第二检测抗体,对结合的egfrviiitcb进行检测。通过添加过氧化物酶底物(abts)生成信号。校准范围为2.35ng/ml至150ng/ml,其中2.5ng/ml为定量下限(lloq)。本研究的结果如表7中所示。对于测试的两种小鼠品系,egfrviiitcb的pk特征均在预期范围内。结果表明,cd3结合剂的cdr的工程化改造未引起可能影响抗体清除的其他序列不利因素。表7.hufcrntg小鼠和nod-scid小鼠的清除率数据(ml/天/kg;平均值)。***尽管为了清楚理解的目的先前已经通过说明和实例相当详细地描述了发明,但是所述说明和实例不应解释为限制本发明的范围。本文引用的所有专利和科学文献的公开内容的全部内容以引用方式明确地并入。序列表<110>豪夫迈·罗氏有限公司(f.hoffmann-larocheag)<120>与cd3结合的抗体<130>p35214<150>ep18214994.8<151>2018-12-21<160>122<170>patentin3.5版<210>1<211>5<212>prt<213>人工序列<220><223>cd3orighcdr1<400>1thrtyralametasn15<210>2<211>5<212>prt<213>人工序列<220><223>cd3opthcdr1<400>2sertyralametasn15<210>3<211>19<212>prt<213>人工序列<220><223>cd3orig/cd3opthcdr2<400>3argileargserlystyrasnasntyralathrtyrtyralaaspser151015vallysgly<210>4<211>14<212>prt<213>人工序列<220><223>cd3orighcdr3<400>4hisglyasnpheglyasnsertyrvalsertrpphealatyr1510<210>5<211>14<212>prt<213>人工序列<220><223>cd3opthcdr3<400>5histhrthrpheprosersertyrvalsertyrtyrglytyr1510<210>6<211>125<212>prt<213>人工序列<220><223>cd3origvh<400>6gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserser115120125<210>7<211>125<212>prt<213>人工序列<220><223>cd3optvh<400>7gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglypheglnphesersertyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisthrthrpheprosersertyrvalsertyrtyr100105110glytyrtrpglyglnglythrleuvalthrvalserser115120125<210>8<211>14<212>prt<213>人工序列<220><223>cd3orig/cd3optlcdr1<400>8glyserserthrglyalavalthrthrserasntyralaasn1510<210>9<211>7<212>prt<213>人工序列<220><223>cd3orig/cd3optlcdr2<400>9glythrasnlysargalapro15<210>10<211>9<212>prt<213>人工序列<220><223>cd3orig/cd3optlcdr3<400>10alaleutrptyrserasnleutrpval15<210>11<211>109<212>prt<213>人工序列<220><223>cd3orig/cd3optvl<400>11glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleu100105<210>12<211>453<212>prt<213>人工序列<220><223>cd3origigghc<400>12gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserseralaserthr115120125lysglyproservalpheproleualaproserserlysserthrser130135140glyglythralaalaleuglycysleuvallysasptyrpheproglu145150155160provalthrvalsertrpasnserglyalaleuthrserglyvalhis165170175thrpheproalavalleuglnserserglyleutyrserleuserser180185190valvalthrvalproserserserleuglythrglnthrtyrilecys195200205asnvalasnhislysproserasnthrlysvalasplyslysvalglu210215220prolyssercysasplysthrhisthrcysproprocysproalapro225230235240gluleuleuglyglyproservalpheleupheproprolysprolys245250255aspthrleumetileserargthrprogluvalthrcysvalvalval260265270aspvalserhisgluaspprogluvallyspheasntrptyrvalasp275280285glyvalgluvalhisasnalalysthrlysproargglugluglntyr290295300asnserthrtyrargvalvalservalleuthrvalleuhisglnasp305310315320trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu325330335proalaproileglulysthrileserlysalalysglyglnproarg340345350gluproglnvaltyrthrleuproproserargaspgluleuthrlys355360365asnglnvalserleuthrcysleuvallysglyphetyrproserasp370375380ilealavalglutrpgluserasnglyglnprogluasnasntyrlys385390395400thrthrproprovalleuaspseraspglyserphepheleutyrser405410415lysleuthrvalasplysserargtrpglnglnglyasnvalpheser420425430cysservalmethisglualaleuhisasnhistyrthrglnlysser435440445leuserleuserpro450<210>13<211>453<212>prt<213>人工序列<220><223>cd3optigghc<400>13gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglypheglnphesersertyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisthrthrpheprosersertyrvalsertyrtyr100105110glytyrtrpglyglnglythrleuvalthrvalserseralaserthr115120125lysglyproservalpheproleualaproserserlysserthrser130135140glyglythralaalaleuglycysleuvallysasptyrpheproglu145150155160provalthrvalsertrpasnserglyalaleuthrserglyvalhis165170175thrpheproalavalleuglnserserglyleutyrserleuserser180185190valvalthrvalproserserserleuglythrglnthrtyrilecys195200205asnvalasnhislysproserasnthrlysvalasplyslysvalglu210215220prolyssercysasplysthrhisthrcysproprocysproalapro225230235240glualaalaglyglyproservalpheleupheproprolysprolys245250255aspthrleumetileserargthrprogluvalthrcysvalvalval260265270aspvalserhisgluaspprogluvallyspheasntrptyrvalasp275280285glyvalgluvalhisasnalalysthrlysproargglugluglntyr290295300asnserthrtyrargvalvalservalleuthrvalleuhisglnasp305310315320trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu325330335glyalaproileglulysthrileserlysalalysglyglnproarg340345350gluproglnvaltyrthrleuproproserargaspgluleuthrlys355360365asnglnvalserleuthrcysleuvallysglyphetyrproserasp370375380ilealavalglutrpgluserasnglyglnprogluasnasntyrlys385390395400thrthrproprovalleuaspseraspglyserphepheleutyrser405410415lysleuthrvalasplysserargtrpglnglnglyasnvalpheser420425430cysservalmethisglualaleuhisasnhistyrthrglnlysser435440445leuserleuserpro450<210>14<211>216<212>prt<213>人工序列<220><223>cd3orig/cd3optigglc<400>14glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuargthrval100105110alaalaproservalpheilepheproproseraspgluglnleulys115120125serglythralaservalvalcysleuleuasnasnphetyrproarg130135140glualalysvalglntrplysvalaspasnalaleuglnserglyasn145150155160serglngluservalthrgluglnaspserlysaspserthrtyrser165170175leuserserthrleuthrleuserlysalaasptyrglulyshislys180185190valtyralacysgluvalthrhisglnglyleuserserprovalthr195200205lysserpheasnargglyglucys210215<210>15<211>5<212>prt<213>人工序列<220><223>tyrp1hcdr1<400>15asptyrpheleuhis15<210>16<211>17<212>prt<213>人工序列<220><223>tyrp1hcdr2<400>16trpileasnproaspasnglyasnthrvaltyralaglnlysphegln151015gly<210>17<211>12<212>prt<213>人工序列<220><223>tyrp1hcdr3<400>17argasptyrthrtyrglulysalaalaleuasptyr1510<210>18<211>121<212>prt<213>人工序列<220><223>tyrp1vh<400>18glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglypheasnilelysasptyr202530pheleuhistrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnproaspasnglyasnthrvaltyralaglnlysphe505560glnglyargvalthrmetthralaaspthrserthrserthrvaltyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095thrargargasptyrthrtyrglulysalaalaleuasptyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>19<211>11<212>prt<213>人工序列<220><223>tyrp1lcdr1<400>19argalaserglyasniletyrasntyrleuala1510<210>20<211>7<212>prt<213>人工序列<220><223>tyrp1lcdr2<400>20aspalalysthrleualaasp15<210>21<211>9<212>prt<213>人工序列<220><223>tyrp1lcdr3<400>21glnhisphetrpserleuprophethr15<210>22<211>107<212>prt<213>人工序列<220><223>tyrp1vl<400>22aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglyasniletyrasntyr202530leualatrptyrglnglnlysproglylysvalprolysleuleuile354045tyraspalalysthrleualaaspglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspvalalathrtyrtyrcysglnhisphetrpserleuprophe859095thrpheglyglnglythrlysleugluilelys100105<210>23<211>674<212>prt<213>人工序列<220><223>tyrp1vh-ch1(ee)-cd3orig/cd3optvl-ch1-fc(杵,pglala)<400>23glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglypheasnilelysasptyr202530pheleuhistrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnproaspasnglyasnthrvaltyralaglnlysphe505560glnglyargvalthrmetthralaaspthrserthrserthrvaltyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095thrargargasptyrthrtyrglulysalaalaleuasptyrtrpgly100105110glnglythrleuvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvalgluasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalaspglulysvalgluprolyssercys210215220aspglyglyglyglyserglyglyglyglyserglnalavalvalthr225230235240glngluproserleuthrvalserproglyglythrvalthrleuthr245250255cysglyserserthrglyalavalthrthrserasntyralaasntrp260265270valglnglulysproglyglnalapheargglyleuileglyglythr275280285asnlysargalaproglythrproalaargpheserglyserleuleu290295300glyglylysalaalaleuthrleuserglyalaglnprogluaspglu305310315320alaglutyrtyrcysalaleutrptyrserasnleutrpvalphegly325330335glyglythrlysleuthrvalleuserseralaserthrlysglypro340345350servalpheproleualaproserserlysserthrserglyglythr355360365alaalaleuglycysleuvallysasptyrpheprogluprovalthr370375380valsertrpasnserglyalaleuthrserglyvalhisthrphepro385390395400alavalleuglnserserglyleutyrserleuserservalvalthr405410415valproserserserleuglythrglnthrtyrilecysasnvalasn420425430hislysproserasnthrlysvalasplyslysvalgluprolysser435440445cysasplysthrhisthrcysproprocysproalaproglualaala450455460glyglyproservalpheleupheproprolysprolysaspthrleu465470475480metileserargthrprogluvalthrcysvalvalvalaspvalser485490495hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu500505510valhisasnalalysthrlysproargglugluglntyrasnserthr515520525tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn530535540glylysglutyrlyscyslysvalserasnlysalaleuglyalapro545550555560ileglulysthrileserlysalalysglyglnproarggluprogln565570575valtyrthrleuproprocysargaspgluleuthrlysasnglnval580585590serleutrpcysleuvallysglyphetyrproseraspilealaval595600605glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro610615620provalleuaspseraspglyserphepheleutyrserlysleuthr625630635640valasplysserargtrpglnglnglyasnvalphesercysserval645650655methisglualaleuhisasnhistyrthrglnlysserleuserleu660665670serpro<210>24<211>449<212>prt<213>人工序列<220><223>tyrp1vh-ch1(ee)-fc(臼,pglala)<400>24glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglypheasnilelysasptyr202530pheleuhistrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnproaspasnglyasnthrvaltyralaglnlysphe505560glnglyargvalthrmetthralaaspthrserthrserthrvaltyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095thrargargasptyrthrtyrglulysalaalaleuasptyrtrpgly100105110glnglythrleuvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvalgluasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalaspglulysvalgluprolyssercys210215220asplysthrhisthrcysproprocysproalaproglualaalagly225230235240glyproservalpheleupheproprolysprolysaspthrleumet245250255ileserargthrprogluvalthrcysvalvalvalaspvalserhis260265270gluaspprogluvallyspheasntrptyrvalaspglyvalgluval275280285hisasnalalysthrlysproargglugluglntyrasnserthrtyr290295300argvalvalservalleuthrvalleuhisglnasptrpleuasngly305310315320lysglutyrlyscyslysvalserasnlysalaleuglyalaproile325330335glulysthrileserlysalalysglyglnproarggluproglnval340345350cysthrleuproproserargaspgluleuthrlysasnglnvalser355360365leusercysalavallysglyphetyrproseraspilealavalglu370375380trpgluserasnglyglnprogluasnasntyrlysthrthrpropro385390395400valleuaspseraspglyserphepheleuvalserlysleuthrval405410415asplysserargtrpglnglnglyasnvalphesercysservalmet420425430hisglualaleuhisasnhistyrthrglnlysserleuserleuser435440445pro<210>25<211>214<212>prt<213>人工序列<220><223>tyrp1vl-cl(rk)<400>25aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglyasniletyrasntyr202530leualatrptyrglnglnlysproglylysvalprolysleuleuile354045tyraspalalysthrleualaaspglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspvalalathrtyrtyrcysglnhisphetrpserleuprophe859095thrpheglyglnglythrlysleugluilelysargthrvalalaala100105110proservalpheilepheproproserasparglysleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>26<211>232<212>prt<213>人工序列<220><223>cd3origvh-cl<400>26gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserseralaserval115120125alaalaproservalpheilepheproproseraspgluglnleulys130135140serglythralaservalvalcysleuleuasnasnphetyrproarg145150155160glualalysvalglntrplysvalaspasnalaleuglnserglyasn165170175serglngluservalthrgluglnaspserlysaspserthrtyrser180185190leuserserthrleuthrleuserlysalaasptyrglulyshislys195200205valtyralacysgluvalthrhisglnglyleuserserprovalthr210215220lysserpheasnargglyglucys225230<210>27<211>232<212>prt<213>人工序列<220><223>cd3optvh-cl<400>27gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglypheglnphesersertyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisthrthrpheprosersertyrvalsertyrtyr100105110glytyrtrpglyglnglythrleuvalthrvalserseralaserval115120125alaalaproservalpheilepheproproseraspgluglnleulys130135140serglythralaservalvalcysleuleuasnasnphetyrproarg145150155160glualalysvalglntrplysvalaspasnalaleuglnserglyasn165170175serglngluservalthrgluglnaspserlysaspserthrtyrser180185190leuserserthrleuthrleuserlysalaasptyrglulyshislys195200205valtyralacysgluvalthrhisglnglyleuserserprovalthr210215220lysserpheasnargglyglucys225230<210>28<211>360<212>prt<213>人工序列<220><223>人cd3ε柄-fc(杵)-avi<400>28glnaspglyasngluglumetglyglyilethrglnthrprotyrlys151015valserileserglythrthrvalileleuthrcysproglntyrpro202530glysergluileleutrpglnhisasnasplysasnileglyglyasp354045gluaspasplysasnileglyseraspgluasphisleuserleulys505560gluphesergluleugluglnserglytyrtyrvalcystyrproarg65707580glyserlysprogluaspalaasnphetyrleutyrleuargalaarg859095valsergluasncysvalaspgluglnleutyrpheglnglyglyser100105110prolysseralaasplysthrhisthrcysproprocysproalapro115120125gluleuleuglyglyproservalpheleupheproprolysprolys130135140aspthrleumetileserargthrprogluvalthrcysvalvalval145150155160aspvalserhisgluaspprogluvallyspheasntrptyrvalasp165170175glyvalgluvalhisasnalalysthrlysproargglugluglntyr180185190asnserthrtyrargvalvalservalleuthrvalleuhisglnasp195200205trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu210215220proalaproileglulysthrileserlysalalysglyglnproarg225230235240gluproglnvaltyrthrleuproprocysargaspgluleuthrlys245250255asnglnvalserleutrpcysleuvallysglyphetyrproserasp260265270ilealavalglutrpgluserasnglyglnprogluasnasntyrlys275280285thrthrproprovalleuaspseraspglyserphepheleutyrser290295300lysleuthrvalasplysserargtrpglnglnglyasnvalpheser305310315320cysservalmethisglualaleuhisasnhistyrthrglnlysser325330335leuserleuserproglylysserglyglyleuasnaspilepheglu340345350alaglnlysileglutrphisglu355360<210>29<211>325<212>prt<213>人工序列<220><223>人cd3δ柄-fc(臼)-avi<400>29phelysileproileglugluleugluaspargvalphevalasncys151015asnthrserilethrtrpvalgluglythrvalglythrleuleuser202530aspilethrargleuaspleuglylysargileleuaspproarggly354045iletyrargcysasnglythraspiletyrlysasplysgluserthr505560valglnvalhistyrargmetcysargsergluglnleutyrphegln65707580glyasplysthrhisthrcysproprocysproalaprogluleuleu859095glyglyproservalpheleupheproprolysprolysaspthrleu100105110metileserargthrprogluvalthrcysvalvalvalaspvalser115120125hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu130135140valhisasnalalysthrlysproargglugluglntyrasnserthr145150155160tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn165170175glylysglutyrlyscyslysvalserasnlysalaleuproalapro180185190ileglulysthrileserlysalalysglyglnproarggluprogln195200205valcysthrleuproproserargaspgluleuthrlysasnglnval210215220serleusercysalavallysglyphetyrproseraspilealaval225230235240glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro245250255provalleuaspseraspglyserphepheleuvalserlysleuthr260265270valasplysserargtrpglnglnglyasnvalphesercysserval275280285methisglualaleuhisasnhistyrthrglnlysserleuserleu290295300serproglylysserglyglyleuasnaspilepheglualaglnlys305310315320ileglutrphisglu325<210>30<211>351<212>prt<213>人工序列<220><223>食蟹猴cd3ε柄-fc(杵)-avi<400>30glnaspglyasngluglumetglyserilethrglnthrprotyrgln151015valserileserglythrthrvalileleuthrcysserglnhisleu202530glyserglualaglntrpglnhisasnglylysasnlysgluaspser354045glyaspargleupheleuproglupheserglumetgluglnsergly505560tyrtyrvalcystyrproargglyserasnprogluaspalaserhis65707580hisleutyrleulysalaargvalsergluasncysvalaspglugln859095leutyrpheglnglyglyserprolysseralaasplysthrhisthr100105110cysproprocysproalaprogluleuleuglyglyproservalphe115120125leupheproprolysprolysaspthrleumetileserargthrpro130135140gluvalthrcysvalvalvalaspvalserhisgluaspprogluval145150155160lyspheasntrptyrvalaspglyvalgluvalhisasnalalysthr165170175lysproargglugluglntyrasnserthrtyrargvalvalserval180185190leuthrvalleuhisglnasptrpleuasnglylysglutyrlyscys195200205lysvalserasnlysalaleuproalaproileglulysthrileser210215220lysalalysglyglnproarggluproglnvaltyrthrleupropro225230235240cysargaspgluleuthrlysasnglnvalserleutrpcysleuval245250255lysglyphetyrproseraspilealavalglutrpgluserasngly260265270glnprogluasnasntyrlysthrthrproprovalleuaspserasp275280285glyserphepheleutyrserlysleuthrvalasplysserargtrp290295300glnglnglyasnvalphesercysservalmethisglualaleuhis305310315320asnhistyrthrglnlysserleuserleuserproglylyssergly325330335glyleuasnaspilepheglualaglnlysileglutrphisglu340345350<210>31<211>334<212>prt<213>人工序列<220><223>食蟹猴cd3δ柄-fc(臼)-avi<400>31phelysileprovalglugluleugluaspargvalphevallyscys151015asnthrservalthrtrpvalgluglythrvalglythrleuleuthr202530asnasnthrargleuaspleuglylysargileleuaspproarggly354045iletyrargcysasnglythraspiletyrlysasplysgluserala505560valglnvalhistyrargmetserglnasncysvalaspgluglnleu65707580tyrpheglnglyglyserprolysseralaasplysthrhisthrcys859095proprocysproalaprogluleuleuglyglyproservalpheleu100105110pheproprolysprolysaspthrleumetileserargthrproglu115120125valthrcysvalvalvalaspvalserhisgluaspprogluvallys130135140pheasntrptyrvalaspglyvalgluvalhisasnalalysthrlys145150155160proargglugluglntyrasnserthrtyrargvalvalservalleu165170175thrvalleuhisglnasptrpleuasnglylysglutyrlyscyslys180185190valserasnlysalaleuproalaproileglulysthrileserlys195200205alalysglyglnproarggluproglnvalcysthrleuproproser210215220argaspgluleuthrlysasnglnvalserleusercysalavallys225230235240glyphetyrproseraspilealavalglutrpgluserasnglygln245250255progluasnasntyrlysthrthrproprovalleuaspseraspgly260265270serphepheleuvalserlysleuthrvalasplysserargtrpgln275280285glnglyasnvalphesercysservalmethisglualaleuhisasn290295300histyrthrglnlysserleuserleuserproglylysserglygly305310315320leuasnaspilepheglualaglnlysileglutrphisglu325330<210>32<211>699<212>prt<213>人工序列<220><223>人tyrp1ecd-fc(杵)-avi<400>32glnpheproargglncysalathrvalglualaleuargserglymet151015cyscysproaspleuserprovalserglyproglythraspargcys202530glyserserserglyargglyargcysglualavalthralaaspser354045argprohisserproglntyrprohisaspglyargaspaspargglu505560valtrpproleuargphepheasnargthrcyshiscysasnglyasn65707580pheserglyhisasncysglythrcysargproglytrpargglyala859095alacysaspglnargvalleuilevalargargasnleuleuaspleu100105110serlysgluglulysasnhisphevalargalaleuaspmetalalys115120125argthrthrhisproleuphevalilealathrargargsergluglu130135140ileleuglyproaspglyasnthrproglnphegluasnileserile145150155160tyrasntyrphevaltrpthrhistyrtyrservallyslysthrphe165170175leuglyvalglyglngluserpheglygluvalasppheserhisglu180185190glyproalapheleuthrtrphisargtyrhisleuleuargleuglu195200205lysaspmetglnglumetleuglngluproserpheserleuprotyr210215220trpasnphealathrglylysasnvalcysaspilecysthraspasp225230235240leumetglyserargserasnpheaspserthrleuileserproasn245250255servalpheserglntrpargvalvalcysaspserleugluasptyr260265270aspthrleuglythrleucysasnserthrgluaspglyproilearg275280285argasnproalaglyasnvalalaargprometvalglnargleupro290295300gluproglnaspvalalaglncysleugluvalglyleupheaspthr305310315320proprophetyrserasnserthrasnserpheargasnthrvalglu325330335glytyrseraspprothrglylystyraspproalavalargserleu340345350hisasnleualahisleupheleuasnglythrglyglyglnthrhis355360365leuserproasnaspproilephevalleuleuhisthrphethrasp370375380alavalpheaspglutrpleuargargtyrasnalaaspileserthr385390395400pheproleugluasnalaproileglyhisasnargglntyrasnmet405410415valprophetrpproprovalthrasnthrglumetphevalthrala420425430proaspasnleuglytyrthrtyrgluileglntrpproserargglu435440445pheservalprogluglyserasplysthrhisthrcysproprocys450455460proalaproglualaalaglyglyproservalpheleuphepropro465470475480lysprolysaspthrleumetileserargthrprogluvalthrcys485490495valvalvalaspvalserhisgluaspprogluvallyspheasntrp500505510tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu515520525gluglntyrasnserthrtyrargvalvalservalleuthrvalleu530535540hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn545550555560lysalaleuglyalaproileglulysthrileserlysalalysgly565570575glnproarggluproglnvaltyrthrleuproprocysargaspglu580585590leuthrlysasnglnvalserleutrpcysleuvallysglyphetyr595600605proseraspilealavalglutrpgluserasnglyglnprogluasn610615620asntyrlysthrthrproprovalleuaspseraspglyserphephe625630635640leutyrserlysleuthrvalasplysserargtrpglnglnglyasn645650655valphesercysservalmethisglualaleuhisasnhistyrthr660665670glnlysserleuserleuserproglylysserglyglyleuasnasp675680685ilepheglualaglnlysileglutrphisglu690695<210>33<211>698<212>prt<213>人工序列<220><223>食蟹猴tyrp1ecd-fc(杵)-avi<400>33glnpheproargglucysalathrvalglualaleuargserglymet151015cyscysproaspleuserprometserglyproglythraspargcys202530glyserserserglyargglyargcysglualavalthralaaspser354045argprohisserproargtyrprohisaspglyargaspaspargglu505560valtrpproleuargphepheasnargthrcyshiscysasnglyasn65707580pheserglyhisasncysglythrcysargproglytrpargglyala859095alacysaspglnargvalleuvalvalargargasnleuleuaspleu100105110serlysgluglulysasnhisphevalargalaleuaspmetalalys115120125argthrthrhisproleuphevalilealathrargargsergluglu130135140ileleuglyproaspglyasnthrproglnphegluasnileserile145150155160tyrasntyrphevaltrpthrhistyrtyrservallyslysthrphe165170175leuglyalaglyglngluserpheglygluvalasppheserhisglu180185190glyproalapheleuthrtrphisargtyrhisleuleuargleuglu195200205lysaspmetglnglumetleuglngluproserpheserleuprotyr210215220trpasnphealathrglylysasnvalcysaspilecysthraspasp225230235240leumetglyserargserasnpheaspserthrleuileserproasn245250255servalpheserglntrpargvalvalcysaspserleugluasptyr260265270aspthrleuglythrleucysasnserthrgluserglyproilearg275280285argasnproalaglyasnvalalaargprometvalglnargleupro290295300gluproglnaspvalalaglncysleugluvalglyleupheaspthr305310315320proprophetyrserasnserthrasnserpheargasnthrvalglu325330335glytyrseraspprothrglylystyraspproalavalargserleu340345350hisasnleualahisleupheleuasnglythrglyglyglnthrhis355360365leuserproasnaspproilephevalleuleuhisthrphethrasp370375380alavalpheaspglutrpleuargargtyrasnalaaspileserthr385390395400pheproleugluasnalaproileglyhisasnargglntyrasnmet405410415valprophetrpproprovalthrasnthrglumetphevalthrala420425430proaspasnleuglytyrthrtyrgluvalglntrpproserargglu435440445pheservalproglyserasplysthrhisthrcysproprocyspro450455460alaproglualaalaglyglyproservalpheleupheproprolys465470475480prolysaspthrleumetileserargthrprogluvalthrcysval485490495valvalaspvalserhisgluaspprogluvallyspheasntrptyr500505510valaspglyvalgluvalhisasnalalysthrlysproarggluglu515520525glntyrasnserthrtyrargvalvalservalleuthrvalleuhis530535540glnasptrpleuasnglylysglutyrlyscyslysvalserasnlys545550555560alaleuglyalaproileglulysthrileserlysalalysglygln565570575proarggluproglnvaltyrthrleuproprocysargaspgluleu580585590thrlysasnglnvalserleutrpcysleuvallysglyphetyrpro595600605seraspilealavalglutrpgluserasnglyglnprogluasnasn610615620tyrlysthrthrproprovalleuaspseraspglyserphepheleu625630635640tyrserlysleuthrvalasplysserargtrpglnglnglyasnval645650655phesercysservalmethisglualaleuhisasnhistyrthrgln660665670lysserleuserleuserproglylysserglyglyleuasnaspile675680685pheglualaglnlysileglutrphisglu690695<210>34<211>699<212>prt<213>人工序列<220><223>小鼠tyrp1ecd-fc(杵)-avi<400>34glnpheproargglucysalaasnileglualaleuargargglyval151015cyscysproaspleuleuproserserglyproglythraspprocys202530glyserserserglyargglyargcysvalalavalilealaaspser354045argprohisserarghistyrprohisaspglylysaspaspargglu505560alatrpproleuargphepheasnargthrcysglncysasnaspasn65707580pheserglyhisasncysglythrcysargproglytrpargglyala859095alacysasnglnlysileleuthrvalargargasnleuleuaspleu100105110serprogluglulysserhisphevalargalaleuaspmetalalys115120125argthrthrhisproglnphevalilealathrargargleugluasp130135140ileleuglyproaspglyasnthrproglnphegluasnileserval145150155160tyrasntyrphevaltrpthrhistyrtyrservallyslysthrphe165170175leuglythrglyglngluserpheglyaspvalasppheserhisglu180185190glyproalapheleuthrtrphisargtyrhisleuleuglnleuglu195200205argaspmetglnglumetleuglngluproserpheserleuprotyr210215220trpasnphealathrglylysasnvalcysaspvalcysthraspasp225230235240leumetglyserargserasnpheaspserthrleuileserproasn245250255servalpheserglntrpargvalvalcysgluserleugluglutyr260265270aspthrleuglythrleucysasnserthrgluglyglyproilearg275280285argasnproalaglyasnvalglyargproalavalglnargleupro290295300gluproglnaspvalthrglncysleugluvalargvalpheaspthr305310315320proprophetyrserasnserthraspserpheargasnthrvalglu325330335glytyrseralaprothrglylystyraspproalavalargserleu340345350hisasnleualahisleupheleuasnglythrglyglyglnthrhis355360365leuserproasnaspproilephevalleuleuhisthrphethrasp370375380alavalpheaspglutrpleuargargtyrasnalaaspileserthr385390395400pheproleugluasnalaproileglyhisasnargglntyrasnmet405410415valprophetrpproprovalthrasnthrglumetphevalthrala420425430proaspasnleuglytyralatyrgluvalglntrpproglyglnglu435440445phethrvalsergluglyserasplysthrhisthrcysproprocys450455460proalaproglualaalaglyglyproservalpheleuphepropro465470475480lysprolysaspthrleumetileserargthrprogluvalthrcys485490495valvalvalaspvalserhisgluaspprogluvallyspheasntrp500505510tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu515520525gluglntyrasnserthrtyrargvalvalservalleuthrvalleu530535540hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn545550555560lysalaleuglyalaproileglulysthrileserlysalalysgly565570575glnproarggluproglnvaltyrthrleuproprocysargaspglu580585590leuthrlysasnglnvalserleutrpcysleuvallysglyphetyr595600605proseraspilealavalglutrpgluserasnglyglnprogluasn610615620asntyrlysthrthrproprovalleuaspseraspglyserphephe625630635640leutyrserlysleuthrvalasplysserargtrpglnglnglyasn645650655valphesercysservalmethisglualaleuhisasnhistyrthr660665670glnlysserleuserleuserproglylysserglyglyleuasnasp675680685ilepheglualaglnlysileglutrphisglu690695<210>35<211>225<212>prt<213>人工序列<220><223>fc(臼)<400>35asplysthrhisthrcysproprocysproalaproglualaalagly151015glyproservalpheleupheproprolysprolysaspthrleumet202530ileserargthrprogluvalthrcysvalvalvalaspvalserhis354045gluaspprogluvallyspheasntrptyrvalaspglyvalgluval505560hisasnalalysthrlysproargglugluglntyrasnserthrtyr65707580argvalvalservalleuthrvalleuhisglnasptrpleuasngly859095lysglutyrlyscyslysvalserasnlysalaleuglyalaproile100105110glulysthrileserlysalalysglyglnproarggluproglnval115120125cysthrleuproproserargaspgluleuthrlysasnglnvalser130135140leusercysalavallysglyphetyrproseraspilealavalglu145150155160trpgluserasnglyglnprogluasnasntyrlysthrthrpropro165170175valleuaspseraspglyserphepheleuvalserlysleuthrval180185190asplysserargtrpglnglnglyasnvalphesercysservalmet195200205hisglualaleuhisasnargphethrglnlysserleuserleuser210215220pro225<210>36<211>393<212>prt<213>人工序列<220><223>egfrviiiecd-avi-his<400>36leugluglulyslysglyasntyrvalvalthrasphisglysercys151015valargalacysglyalaaspsertyrglumetglugluaspglyval202530arglyscyslyslyscysgluglyprocysarglysvalcysasngly354045ileglyileglygluphelysaspserleuserileasnalathrasn505560ilelyshisphelysasncysthrserileserglyaspleuhisile65707580leuprovalalapheargglyaspserphethrhisthrproproleu859095aspproglngluleuaspileleulysthrvallysgluilethrgly100105110pheleuleuileglnalatrpprogluasnargthraspleuhisala115120125phegluasnleugluileileargglyargthrlysglnhisglygln130135140pheserleualavalvalserleuasnilethrserleuglyleuarg145150155160serleulysgluileseraspglyaspvalileileserglyasnlys165170175asnleucystyralaasnthrileasntrplyslysleupheglythr180185190serglyglnlysthrlysileileserasnargglygluasnsercys195200205lysalathrglyglnvalcyshisalaleucysserprogluglycys210215220trpglyprogluproargaspcysvalsercysargasnvalserarg225230235240glyargglucysvalasplyscysasnleuleugluglygluproarg245250255gluphevalgluasnserglucysileglncyshisproglucysleu260265270proglnalametasnilethrcysthrglyargglyproaspasncys275280285ileglncysalahistyrileaspglyprohiscysvallysthrcys290295300proalaglyvalmetglygluasnasnthrleuvaltrplystyrala305310315320aspalaglyhisvalcyshisleucyshisproasncysthrtyrgly325330335cysthrglyproglyleugluglycysprothrasnglyprolysile340345350proservalaspglyglyserprothrproprothrproglyglygly355360365serglyleuasnaspilepheglualaglnlysileglutrphisglu370375380alaargalahishishishishishis385390<210>37<211>5<212>prt<213>人工序列<220><223>egfrviiip056.021hcdr1<400>37sertyrtrpileala15<210>38<211>17<212>prt<213>人工序列<220><223>egfrviiip056.021hcdr2<400>38valilehisprotyraspseraspthrargtyrserproserphegln151015gly<210>39<211>10<212>prt<213>人工序列<220><223>egfrviiip056.021hcdr3<400>39valserargsersertyralapheasptyr1510<210>40<211>119<212>prt<213>人工序列<220><223>egfrviiip056.021vh<400>40gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>41<211>17<212>prt<213>人工序列<220><223>egfrviiip056.021lcdr1<400>41lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>42<211>7<212>prt<213>人工序列<220><223>egfrviiip056.021lcdr2<400>42trpalaserthrarggluser15<210>43<211>10<212>prt<213>人工序列<220><223>egfrviiip056.021lcdr3<400>43glnglnvalhisserglyproprovalthr1510<210>44<211>114<212>prt<213>人工序列<220><223>egfrviiip056.021vl<400>44aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095valhisserglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>45<211>5<212>prt<213>人工序列<220><223>egfrviiip056.052hcdr1<400>45asntyrtrpilegly15<210>46<211>17<212>prt<213>人工序列<220><223>egfrviiip056.052hcdr2<400>46thriletyrproglyaspseraspargargtyrserproserphegln151015gly<210>47<211>10<212>prt<213>人工序列<220><223>egfrviiip056.052hcdr3<400>47valserargsersertyralapheasptyr1510<210>48<211>119<212>prt<213>人工序列<220><223>egfrviiip056.052vh<400>48gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrthrphemetasntyr202530trpileglytrpvalargglnmetproglylysglyleuglutrpmet354045glythriletyrproglyaspseraspargargtyrserproserphe505560glnglyglnvalthrleuseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>49<211>17<212>prt<213>人工序列<220><223>egfrviiip056.052lcdr1<400>49lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>50<211>7<212>prt<213>人工序列<220><223>egfrviiip056.052lcdr2<400>50trpalaserthrarggluser15<210>51<211>10<212>prt<213>人工序列<220><223>egfrviiip056.052lcdr3<400>51glnglnvalhisserglyproprovalthr1510<210>52<211>114<212>prt<213>人工序列<220><223>egfrviiip056.052vl<400>52aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095valhisserglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>53<211>5<212>prt<213>人工序列<220><223>egfrviiip047.019hcdr1<400>53seriletrpilehis15<210>54<211>17<212>prt<213>人工序列<220><223>egfrviiip047.019hcdr2<400>54thriletyrproglyaspseraspthrargtyrserproserphegln151015gly<210>55<211>9<212>prt<213>人工序列<220><223>egfrviiip047.019hcdr3<400>55thrglyproglyleualapheasptyr15<210>56<211>118<212>prt<213>人工序列<220><223>egfrviiip047.019vh<400>56gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheproserile202530trpilehistrpvalargglnmetproglylysglyleuglutrpmet354045glythriletyrproglyaspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargthrglyproglyleualapheasptyrtrpglyglnglythr100105110leuvalthrvalserser115<210>57<211>17<212>prt<213>人工序列<220><223>egfrviiip047.019lcdr1<400>57lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>58<211>7<212>prt<213>人工序列<220><223>egfrviiip047.019lcdr2<400>58trpalaserthrarggluser15<210>59<211>9<212>prt<213>人工序列<220><223>egfrviiip047.019lcdr3<400>59glnglnsertyrserthrproilethr15<210>60<211>113<212>prt<213>人工序列<220><223>egfrviiip047.019vl<400>60aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095sertyrserthrproilethrpheglyglnglythrlysvalgluile100105110lys<210>61<211>5<212>prt<213>人工序列<220><223>egfrviiip057.012hcdr1<400>61asntyrtrpileala15<210>62<211>17<212>prt<213>人工序列<220><223>egfrviiip057.012hcdr2<400>62ileiletyrproaspaspseraspthrargtyrserproserphegln151015gly<210>63<211>12<212>prt<213>人工序列<220><223>egfrviiip057.012hcdr3<400>63alathrasnilealaserglyglytyrpheasptyr1510<210>64<211>121<212>prt<213>人工序列<220><223>egfrviiip057.012vh<400>64gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserphealaasntyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyileiletyrproaspaspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargalathrasnilealaserglyglytyrpheasptyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>65<211>17<212>prt<213>人工序列<220><223>egfrviiip057.012lcdr1<400>65lysserserglnservalleutrpasnserasnasnlysasntyrleu151015ala<210>66<211>7<212>prt<213>人工序列<220><223>egfrviiip057.012lcdr2<400>66trpalaserlysarggluser15<210>67<211>9<212>prt<213>人工序列<220><223>egfrviiip057.012lcdr3<400>67glnglnsertyrseralaproilethr15<210>68<211>113<212>prt<213>人工序列<220><223>egfrviiip057.012vl<400>68aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutrpasn202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserlysarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095sertyrseralaproilethrpheglyglnglythrlysvalgluile100105110lys<210>69<211>5<212>prt<213>人工序列<220><223>egfrviiip057.011hcdr1<400>69argargtrpileala15<210>70<211>17<212>prt<213>人工序列<220><223>egfrviiip057.011hcdr2<400>70ileiletyrproglyaspseraspthrargtyrserproserphegln151015gly<210>71<211>12<212>prt<213>人工序列<220><223>egfrviiip057.011hcdr3<400>71alathrasnilealaserglyglytyrpheasptyr1510<210>72<211>121<212>prt<213>人工序列<220><223>egfrviiip057.011vh<400>72gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrasnpheglyargarg202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyileiletyrproglyaspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargalathrasnilealaserglyglytyrpheasptyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>73<211>17<212>prt<213>人工序列<220><223>egfrviiip057.011lcdr1<400>73lysserserglnservalleutrpasnserasnasnlysasntyrleu151015ala<210>74<211>7<212>prt<213>人工序列<220><223>egfrviiip057.011lcdr2<400>74trpalaserlysarggluser15<210>75<211>9<212>prt<213>人工序列<220><223>egfrviiip057.011lcdr3<400>75glnglnsertyrseralaproilethr15<210>76<211>113<212>prt<213>人工序列<220><223>egfrviiip057.011vl<400>76aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutrpasn202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserlysarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095sertyrseralaproilethrpheglyglnglythrlysvalgluile100105110lys<210>77<211>5<212>prt<213>人工序列<220><223>egfrviiip056.027hcdr1<400>77asnasntrpileala15<210>78<211>17<212>prt<213>人工序列<220><223>egfrviiip056.027hcdr2<400>78valiletyrproglyaspserasplysargtyrserproserphegln151015gly<210>79<211>10<212>prt<213>人工序列<220><223>egfrviiip056.027hcdr3<400>79valserargsersertyralapheasptyr1510<210>80<211>119<212>prt<213>人工序列<220><223>egfrviiip056.027vh<400>80gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrthrpheglyasnasn202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvaliletyrproglyaspserasplysargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>81<211>17<212>prt<213>人工序列<220><223>egfrviiip056.027lcdr1<400>81lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>82<211>7<212>prt<213>人工序列<220><223>egfrviiip056.027lcdr2<400>82trpalaserthrarggluser15<210>83<211>10<212>prt<213>人工序列<220><223>egfrviiip056.027lcdr3<400>83glnglnvalhisserglyproprovalthr1510<210>84<211>114<212>prt<213>人工序列<220><223>egfrviiip056.027vl<400>84aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095valhisserglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>85<211>5<212>prt<213>人工序列<220><223>egfrviiip063.056hcdr1<400>85sertyrtrpileala15<210>86<211>17<212>prt<213>人工序列<220><223>egfrviiip063.056hcdr2<400>86valilehisprotyraspseraspthrargtyrserproserphegln151015gly<210>87<211>10<212>prt<213>人工序列<220><223>egfrviiip063.056hcdr3<400>87valserargsersertyralapheasptyr1510<210>88<211>119<212>prt<213>人工序列<220><223>egfrviiip063.056vh<400>88gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>89<211>17<212>prt<213>人工序列<220><223>egfrviiip063.056lcdr1<400>89lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>90<211>7<212>prt<213>人工序列<220><223>egfrviiip063.056lcdr2<400>90trpalaserthrarggluser15<210>91<211>10<212>prt<213>人工序列<220><223>egfrviiip063.056lcdr3<400>91glnglnglnargaspglyproprovalthr1510<210>92<211>114<212>prt<213>人工序列<220><223>egfrviiip063.056vl<400>92aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095glnargaspglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>93<211>5<212>prt<213>人工序列<220><223>egfrviiip064.078hcdr1<400>93sertyrtrpileala15<210>94<211>17<212>prt<213>人工序列<220><223>egfrviiip064.078hcdr2<400>94valilehisprotyraspseraspthrargtyrserproserphegln151015gly<210>95<211>10<212>prt<213>人工序列<220><223>egfrviiip064.078hcdr3<400>95valserargleusertyralaleuasptyr1510<210>96<211>119<212>prt<213>人工序列<220><223>egfrviiip064.078vh<400>96gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargleusertyralaleuasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>97<211>17<212>prt<213>人工序列<220><223>egfrviiip064.078lcdr1<400>97lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>98<211>7<212>prt<213>人工序列<220><223>egfrviiip064.078lcdr2<400>98trpalaserthrarggluser15<210>99<211>10<212>prt<213>人工序列<220><223>egfrviiip064.078lcdr3<400>99glnglnvalhisserglyproprovalthr1510<210>100<211>114<212>prt<213>人工序列<220><223>egfrviiip064.078vl<400>100aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095valhisserglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>101<211>5<212>prt<213>人工序列<220><223>egfrviiip065.036hcdr1<400>101sertyrtrpileala15<210>102<211>17<212>prt<213>人工序列<220><223>egfrviiip065.036hcdr2<400>102valilehisprotyraspseraspthrargtyrserproserphegln151015gly<210>103<211>10<212>prt<213>人工序列<220><223>egfrviiip065.036hcdr3<400>103valserargsersertyralaleuasptyr1510<210>104<211>119<212>prt<213>人工序列<220><223>egfrviiip065.036vh<400>104gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralaleuasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>105<211>17<212>prt<213>人工序列<220><223>egfrviiip065.036lcdr1<400>105lysserserglnservalleutyrserserasnasnlysasntyrleu151015ala<210>106<211>7<212>prt<213>人工序列<220><223>egfrviiip065.036lcdr2<400>106trpalaserthrarggluser15<210>107<211>10<212>prt<213>人工序列<220><223>egfrviiip065.036lcdr3<400>107glnglnvaltyrserglyproprovalthr1510<210>108<211>114<212>prt<213>人工序列<220><223>egfrviiip065.036vl<400>108aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095valtyrserglyproprovalthrpheglyglnglythrlysvalglu100105110ilelys<210>109<211>672<212>prt<213>人工序列<220><223>egfrviiivh-ch1(ee)-cd3orig/cd3optvl-ch1-fc(杵,pglala)<400>109gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserseralaserthrlysglyproservalphe115120125proleualaproserserlysserthrserglyglythralaalaleu130135140glycysleuvalgluasptyrpheprogluprovalthrvalsertrp145150155160asnserglyalaleuthrserglyvalhisthrpheproalavalleu165170175glnserserglyleutyrserleuserservalvalthrvalproser180185190serserleuglythrglnthrtyrilecysasnvalasnhislyspro195200205serasnthrlysvalaspglulysvalgluprolyssercysaspgly210215220glyglyglyserglyglyglyglyserglnalavalvalthrglnglu225230235240proserleuthrvalserproglyglythrvalthrleuthrcysgly245250255serserthrglyalavalthrthrserasntyralaasntrpvalgln260265270glulysproglyglnalapheargglyleuileglyglythrasnlys275280285argalaproglythrproalaargpheserglyserleuleuglygly290295300lysalaalaleuthrleuserglyalaglnprogluaspglualaglu305310315320tyrtyrcysalaleutrptyrserasnleutrpvalpheglyglygly325330335thrlysleuthrvalleuserseralaserthrlysglyproserval340345350pheproleualaproserserlysserthrserglyglythralaala355360365leuglycysleuvallysasptyrpheprogluprovalthrvalser370375380trpasnserglyalaleuthrserglyvalhisthrpheproalaval385390395400leuglnserserglyleutyrserleuserservalvalthrvalpro405410415serserserleuglythrglnthrtyrilecysasnvalasnhislys420425430proserasnthrlysvalasplyslysvalgluprolyssercysasp435440445lysthrhisthrcysproprocysproalaproglualaalaglygly450455460proservalpheleupheproprolysprolysaspthrleumetile465470475480serargthrprogluvalthrcysvalvalvalaspvalserhisglu485490495aspprogluvallyspheasntrptyrvalaspglyvalgluvalhis500505510asnalalysthrlysproargglugluglntyrasnserthrtyrarg515520525valvalservalleuthrvalleuhisglnasptrpleuasnglylys530535540glutyrlyscyslysvalserasnlysalaleuglyalaproileglu545550555560lysthrileserlysalalysglyglnproarggluproglnvaltyr565570575thrleuproprocysargaspgluleuthrlysasnglnvalserleu580585590trpcysleuvallysglyphetyrproseraspilealavalglutrp595600605gluserasnglyglnprogluasnasntyrlysthrthrproproval610615620leuaspseraspglyserphepheleutyrserlysleuthrvalasp625630635640lysserargtrpglnglnglyasnvalphesercysservalmethis645650655glualaleuhisasnhistyrthrglnlysserleuserleuserpro660665670<210>110<211>447<212>prt<213>人工序列<220><223>egfrviiivh-ch1(ee)-fc(臼,pglala)<400>110gluvalglnleuvalglnserglyalagluvallyslysproglyglu151015serleulysilesercyslysglyserglytyrserpheaspsertyr202530trpilealatrpvalargglnmetproglylysglyleuglutrpmet354045glyvalilehisprotyraspseraspthrargtyrserproserphe505560glnglyglnvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alaargvalserargsersertyralapheasptyrtrpglyglngly100105110thrleuvalthrvalserseralaserthrlysglyproservalphe115120125proleualaproserserlysserthrserglyglythralaalaleu130135140glycysleuvalgluasptyrpheprogluprovalthrvalsertrp145150155160asnserglyalaleuthrserglyvalhisthrpheproalavalleu165170175glnserserglyleutyrserleuserservalvalthrvalproser180185190serserleuglythrglnthrtyrilecysasnvalasnhislyspro195200205serasnthrlysvalaspglulysvalgluprolyssercysasplys210215220thrhisthrcysproprocysproalaproglualaalaglyglypro225230235240servalpheleupheproprolysprolysaspthrleumetileser245250255argthrprogluvalthrcysvalvalvalaspvalserhisgluasp260265270progluvallyspheasntrptyrvalaspglyvalgluvalhisasn275280285alalysthrlysproargglugluglntyrasnserthrtyrargval290295300valservalleuthrvalleuhisglnasptrpleuasnglylysglu305310315320tyrlyscyslysvalserasnlysalaleuglyalaproileglulys325330335thrileserlysalalysglyglnproarggluproglnvalcysthr340345350leuproproserargaspgluleuthrlysasnglnvalserleuser355360365cysalavallysglyphetyrproseraspilealavalglutrpglu370375380serasnglyglnprogluasnasntyrlysthrthrproprovalleu385390395400aspseraspglyserphepheleuvalserlysleuthrvalasplys405410415serargtrpglnglnglyasnvalphesercysservalmethisglu420425430alaleuhisasnhistyrthrglnlysserleuserleuserpro435440445<210>111<211>221<212>prt<213>人工序列<220><223>egfrviiivl-cl(rk)<400>111aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnservalleutyrser202530serasnasnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserthrarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095glnargaspglyproprovalthrpheglyglnglythrlysvalglu100105110ilelysargthrvalalaalaproservalpheilepheproproser115120125asparglysleulysserglythralaservalvalcysleuleuasn130135140asnphetyrproargglualalysvalglntrplysvalaspasnala145150155160leuglnserglyasnserglngluservalthrgluglnaspserlys165170175aspserthrtyrserleuserserthrleuthrleuserlysalaasp180185190tyrglulyshislysvaltyralacysgluvalthrhisglnglyleu195200205serserprovalthrlysserpheasnargglyglucys210215220<210>112<211>186<212>prt<213>智人<400>112glnaspglyasngluglumetglyglyilethrglnthrprotyrlys151015valserileserglythrthrvalileleuthrcysproglntyrpro202530glysergluileleutrpglnhisasnasplysasnileglyglyasp354045gluaspasplysasnileglyseraspgluasphisleuserleulys505560gluphesergluleugluglnserglytyrtyrvalcystyrproarg65707580glyserlysprogluaspalaasnphetyrleutyrleuargalaarg859095valcysgluasncysmetglumetaspvalmetservalalathrile100105110valilevalaspilecysilethrglyglyleuleuleuleuvaltyr115120125tyrtrpserlysasnarglysalalysalalysprovalthrarggly130135140alaglyalaglyglyargglnargglyglnasnlysgluargpropro145150155160provalproasnproasptyrgluproilearglysglyglnargasp165170175leutyrserglyleuasnglnargargile180185<210>113<211>177<212>prt<213>食蟹猴(macacafascicularis)<400>113glnaspglyasngluglumetglyserilethrglnthrprotyrgln151015valserileserglythrthrvalileleuthrcysserglnhisleu202530glyserglualaglntrpglnhisasnglylysasnlysgluaspser354045glyaspargleupheleuproglupheserglumetgluglnsergly505560tyrtyrvalcystyrproargglyserasnprogluaspalaserhis65707580hisleutyrleulysalaargvalcysgluasncysmetglumetasp859095valmetalavalalathrilevalilevalaspilecysilethrleu100105110glyleuleuleuleuvaltyrtyrtrpserlysasnarglysalalys115120125alalysprovalthrargglyalaglyalaglyglyargglnarggly130135140glnasnlysgluargproproprovalproasnproasptyrglupro145150155160ilearglysglyglnglnaspleutyrserglyleuasnglnargarg165170175ile<210>114<211>513<212>prt<213>智人<400>114glnpheproargglncysalathrvalglualaleuargserglymet151015cyscysproaspleuserprovalserglyproglythraspargcys202530glyserserserglyargglyargcysglualavalthralaaspser354045argprohisserproglntyrprohisaspglyargaspaspargglu505560valtrpproleuargphepheasnargthrcyshiscysasnglyasn65707580pheserglyhisasncysglythrcysargproglytrpargglyala859095alacysaspglnargvalleuilevalargargasnleuleuaspleu100105110serlysgluglulysasnhisphevalargalaleuaspmetalalys115120125argthrthrhisproleuphevalilealathrargargsergluglu130135140ileleuglyproaspglyasnthrproglnphegluasnileserile145150155160tyrasntyrphevaltrpthrhistyrtyrservallyslysthrphe165170175leuglyvalglyglngluserpheglygluvalasppheserhisglu180185190glyproalapheleuthrtrphisargtyrhisleuleuargleuglu195200205lysaspmetglnglumetleuglngluproserpheserleuprotyr210215220trpasnphealathrglylysasnvalcysaspilecysthraspasp225230235240leumetglyserargserasnpheaspserthrleuileserproasn245250255servalpheserglntrpargvalvalcysaspserleugluasptyr260265270aspthrleuglythrleucysasnserthrgluaspglyproilearg275280285argasnproalaglyasnvalalaargprometvalglnargleupro290295300gluproglnaspvalalaglncysleugluvalglyleupheaspthr305310315320proprophetyrserasnserthrasnserpheargasnthrvalglu325330335glytyrseraspprothrglylystyraspproalavalargserleu340345350hisasnleualahisleupheleuasnglythrglyglyglnthrhis355360365leuserproasnaspproilephevalleuleuhisthrphethrasp370375380alavalpheaspglutrpleuargargtyrasnalaaspileserthr385390395400pheproleugluasnalaproileglyhisasnargglntyrasnmet405410415valprophetrpproprovalthrasnthrglumetphevalthrala420425430proaspasnleuglytyrthrtyrgluileglntrpproserargglu435440445pheservalprogluileilealailealavalvalglyalaleuleu450455460leuvalalaleuilepheglythralasertyrleuileargalaarg465470475480argsermetaspglualaasnglnproleuleuthraspglntyrgln485490495cystyralagluglutyrglulysleuglnasnproasnglnserval500505510val<210>115<211>919<212>prt<213>智人<400>115leugluglulyslysglyasntyrvalvalthrasphisglysercys151015valargalacysglyalaaspsertyrglumetglugluaspglyval202530arglyscyslyslyscysgluglyprocysarglysvalcysasngly354045ileglyileglygluphelysaspserleuserileasnalathrasn505560ilelyshisphelysasncysthrserileserglyaspleuhisile65707580leuprovalalapheargglyaspserphethrhisthrproproleu859095aspproglngluleuaspileleulysthrvallysgluilethrgly100105110pheleuleuileglnalatrpprogluasnargthraspleuhisala115120125phegluasnleugluileileargglyargthrlysglnhisglygln130135140pheserleualavalvalserleuasnilethrserleuglyleuarg145150155160serleulysgluileseraspglyaspvalileileserglyasnlys165170175asnleucystyralaasnthrileasntrplyslysleupheglythr180185190serglyglnlysthrlysileileserasnargglygluasnsercys195200205lysalathrglyglnvalcyshisalaleucysserprogluglycys210215220trpglyprogluproargaspcysvalsercysargasnvalserarg225230235240glyargglucysvalasplyscysasnleuleugluglygluproarg245250255gluphevalgluasnserglucysileglncyshisproglucysleu260265270proglnalametasnilethrcysthrglyargglyproaspasncys275280285ileglncysalahistyrileaspglyprohiscysvallysthrcys290295300proalaglyvalmetglygluasnasnthrleuvaltrplystyrala305310315320aspalaglyhisvalcyshisleucyshisproasncysthrtyrgly325330335cysthrglyproglyleugluglycysprothrasnglyprolysile340345350proserilealathrglymetvalglyalaleuleuleuleuleuval355360365valalaleuglyileglyleuphemetargargarghisilevalarg370375380lysargthrleuargargleuleuglngluarggluleuvalglupro385390395400leuthrproserglyglualaproasnglnalaleuleuargileleu405410415lysgluthrgluphelyslysilelysvalleuglyserglyalaphe420425430glythrvaltyrlysglyleutrpileprogluglyglulysvallys435440445ileprovalalailelysgluleuargglualathrserprolysala450455460asnlysgluileleuaspglualatyrvalmetalaservalaspasn465470475480prohisvalcysargleuleuglyilecysleuthrserthrvalgln485490495leuilethrglnleumetpropheglycysleuleuasptyrvalarg500505510gluhislysaspasnileglyserglntyrleuleuasntrpcysval515520525glnilealalysglymetasntyrleugluaspargargleuvalhis530535540argaspleualaalaargasnvalleuvallysthrproglnhisval545550555560lysilethrasppheglyleualalysleuleuglyalagluglulys565570575glutyrhisalagluglyglylysvalproilelystrpmetalaleu580585590gluserileleuhisargiletyrthrhisglnseraspvaltrpser595600605tyrglyvalthrvaltrpgluleumetthrpheglyserlysprotyr610615620aspglyileproalasergluileserserileleuglulysglyglu625630635640argleuproglnproproilecysthrileaspvaltyrmetilemet645650655vallyscystrpmetileaspalaaspserargprolyspheargglu660665670leuileileglupheserlysmetalaargaspproglnargtyrleu675680685valileglnglyaspgluargmethisleuproserprothraspser690695700asnphetyrargalaleumetaspglugluaspmetaspaspvalval705710715720aspalaaspglutyrleuileproglnglnglyphepheserserpro725730735serthrserargthrproleuleuserserleuseralathrserasn740745750asnserthrvalalacysileaspargasnglyleuglnsercyspro755760765ilelysgluaspserpheleuglnargtyrserseraspprothrgly770775780alaleuthrgluaspserileaspaspthrpheleuprovalproglu785790795800tyrileasnglnservalprolysargproalaglyservalglnasn805810815provaltyrhisasnglnproleuasnproalaproserargasppro820825830histyrglnaspprohisserthralavalglyasnproglutyrleu835840845asnthrvalglnprothrcysvalasnserthrpheaspserproala850855860histrpalaglnlysglyserhisglnileserleuaspasnproasp865870875880tyrglnglnaspphepheprolysglualalysproasnglyilephe885890895lysglyserthralagluasnalaglutyrleuargvalalaprogln900905910serserglupheileglyala915<210>116<211>1186<212>prt<213>智人<400>116leugluglulyslysvalcysglnglythrserasnlysleuthrgln151015leuglythrphegluasphispheleuserleuglnargmetpheasn202530asncysgluvalvalleuglyasnleugluilethrtyrvalglnarg354045asntyraspleuserpheleulysthrileglngluvalalaglytyr505560valleuilealaleuasnthrvalgluargileproleugluasnleu65707580glnileileargglyasnmettyrtyrgluasnsertyralaleuala859095valleuserasntyraspalaasnlysthrglyleulysgluleupro100105110metargasnleuglngluileleuhisglyalavalargpheserasn115120125asnproalaleucysasnvalgluserileglntrpargaspileval130135140serserasppheleuserasnmetsermetasppheglnasnhisleu145150155160glysercysglnlyscysaspprosercysproasnglysercystrp165170175glyalaglyglugluasncysglnlysleuthrlysileilecysala180185190glnglncysserglyargcysargglylysserproseraspcyscys195200205hisasnglncysalaalaglycysthrglyproarggluseraspcys210215220leuvalcysarglyspheargaspglualathrcyslysaspthrcys225230235240proproleumetleutyrasnprothrthrtyrglnmetaspvalasn245250255progluglylystyrserpheglyalathrcysvallyslyscyspro260265270argasntyrvalvalthrasphisglysercysvalargalacysgly275280285alaaspsertyrglumetglugluaspglyvalarglyscyslyslys290295300cysgluglyprocysarglysvalcysasnglyileglyileglyglu305310315320phelysaspserleuserileasnalathrasnilelyshisphelys325330335asncysthrserileserglyaspleuhisileleuprovalalaphe340345350argglyaspserphethrhisthrproproleuaspproglngluleu355360365aspileleulysthrvallysgluilethrglypheleuleuilegln370375380alatrpprogluasnargthraspleuhisalaphegluasnleuglu385390395400ileileargglyargthrlysglnhisglyglnpheserleualaval405410415valserleuasnilethrserleuglyleuargserleulysgluile420425430seraspglyaspvalileileserglyasnlysasnleucystyrala435440445asnthrileasntrplyslysleupheglythrserglyglnlysthr450455460lysileileserasnargglygluasnsercyslysalathrglygln465470475480valcyshisalaleucysserprogluglycystrpglyproglupro485490495argaspcysvalsercysargasnvalserargglyargglucysval500505510asplyscysasnleuleugluglygluproarggluphevalgluasn515520525serglucysileglncyshisproglucysleuproglnalametasn530535540ilethrcysthrglyargglyproaspasncysileglncysalahis545550555560tyrileaspglyprohiscysvallysthrcysproalaglyvalmet565570575glygluasnasnthrleuvaltrplystyralaaspalaglyhisval580585590cyshisleucyshisproasncysthrtyrglycysthrglyprogly595600605leugluglycysprothrasnglyprolysileproserilealathr610615620glymetvalglyalaleuleuleuleuleuvalvalalaleuglyile625630635640glyleuphemetargargarghisilevalarglysargthrleuarg645650655argleuleuglngluarggluleuvalgluproleuthrprosergly660665670glualaproasnglnalaleuleuargileleulysgluthrgluphe675680685lyslysilelysvalleuglyserglyalapheglythrvaltyrlys690695700glyleutrpileprogluglyglulysvallysileprovalalaile705710715720lysgluleuargglualathrserprolysalaasnlysgluileleu725730735aspglualatyrvalmetalaservalaspasnprohisvalcysarg740745750leuleuglyilecysleuthrserthrvalglnleuilethrglnleu755760765metpropheglycysleuleuasptyrvalarggluhislysaspasn770775780ileglyserglntyrleuleuasntrpcysvalglnilealalysgly785790795800metasntyrleugluaspargargleuvalhisargaspleualaala805810815argasnvalleuvallysthrproglnhisvallysilethraspphe820825830glyleualalysleuleuglyalagluglulysglutyrhisalaglu835840845glyglylysvalproilelystrpmetalaleugluserileleuhis850855860argiletyrthrhisglnseraspvaltrpsertyrglyvalthrval865870875880trpgluleumetthrpheglyserlysprotyraspglyileproala885890895sergluileserserileleuglulysglygluargleuproglnpro900905910proilecysthrileaspvaltyrmetilemetvallyscystrpmet915920925ileaspalaaspserargprolysphearggluleuileilegluphe930935940serlysmetalaargaspproglnargtyrleuvalileglnglyasp945950955960gluargmethisleuproserprothraspserasnphetyrargala965970975leumetaspglugluaspmetaspaspvalvalaspalaaspglutyr980985990leuileproglnglnglyphepheserserproserthrserargthr99510001005proleuleuserserleuseralathrserasnasnserthrval101010151020alacysileaspargasnglyleuglnsercysproilelysglu102510301035aspserpheleuglnargtyrserseraspprothrglyalaleu104010451050thrgluaspserileaspaspthrpheleuprovalproglutyr105510601065ileasnglnservalprolysargproalaglyservalglnasn107010751080provaltyrhisasnglnproleuasnproalaproserargasp108510901095prohistyrglnaspprohisserthralavalglyasnproglu110011051110tyrleuasnthrvalglnprothrcysvalasnserthrpheasp111511201125serproalahistrpalaglnlysglyserhisglnileserleu113011351140aspasnproasptyrglnglnaspphepheprolysglualalys114511501155proasnglyilephelysglyserthralagluasnalaglutyr116011651170leuargvalalaproglnserserglupheileglyala117511801185<210>117<211>225<212>prt<213>智人<400>117asplysthrhisthrcysproprocysproalaprogluleuleugly151015glyproservalpheleupheproprolysprolysaspthrleumet202530ileserargthrprogluvalthrcysvalvalvalaspvalserhis354045gluaspprogluvallyspheasntrptyrvalaspglyvalgluval505560hisasnalalysthrlysproargglugluglntyrasnserthrtyr65707580argvalvalservalleuthrvalleuhisglnasptrpleuasngly859095lysglutyrlyscyslysvalserasnlysalaleuproalaproile100105110glulysthrileserlysalalysglyglnproarggluproglnval115120125tyrthrleuproproserargaspgluleuthrlysasnglnvalser130135140leuthrcysleuvallysglyphetyrproseraspilealavalglu145150155160trpgluserasnglyglnprogluasnasntyrlysthrthrpropro165170175valleuaspseraspglyserphepheleutyrserlysleuthrval180185190asplysserargtrpglnglnglyasnvalphesercysservalmet195200205hisglualaleuhisasnhistyrthrglnlysserleuserleuser210215220pro225<210>118<211>10<212>prt<213>人工序列<220><223>连接基<400>118glyglyglyglyserglyglyglyglyser1510<210>119<211>11<212>prt<213>人工序列<220><223>连接基<400>119aspglyglyglyglyserglyglyglyglyser1510<210>120<211>107<212>prt<213>智人<400>120argthrvalalaalaproservalpheilepheproproseraspglu151015glnleulysserglythralaservalvalcysleuleuasnasnphe202530tyrproargglualalysvalglntrplysvalaspasnalaleugln354045serglyasnserglngluservalthrgluglnaspserlysaspser505560thrtyrserleuserserthrleuthrleuserlysalaasptyrglu65707580lyshislysvaltyralacysgluvalthrhisglnglyleuserser859095provalthrlysserpheasnargglyglucys100105<210>121<211>105<212>prt<213>智人<400>121glnprolysalaalaproservalthrleupheproproserserglu151015gluleuglnalaasnlysalathrleuvalcysleuileseraspphe202530tyrproglyalavalthrvalalatrplysalaaspserserproval354045lysalaglyvalgluthrthrthrproserlysglnserasnasnlys505560tyralaalasersertyrleuserleuthrprogluglntrplysser65707580hisargsertyrsercysglnvalthrhisgluglyserthrvalglu859095lysthrvalalaprothrglucysser100105<210>122<211>328<212>prt<213>智人<400>122alaserthrlysglyproservalpheproleualaproserserlys151015serthrserglyglythralaalaleuglycysleuvallysasptyr202530pheprogluprovalthrvalsertrpasnserglyalaleuthrser354045glyvalhisthrpheproalavalleuglnserserglyleutyrser505560leuserservalvalthrvalproserserserleuglythrglnthr65707580tyrilecysasnvalasnhislysproserasnthrlysvalasplys859095lysvalgluprolyssercysasplysthrhisthrcysproprocys100105110proalaprogluleuleuglyglyproservalpheleuphepropro115120125lysprolysaspthrleumetileserargthrprogluvalthrcys130135140valvalvalaspvalserhisgluaspprogluvallyspheasntrp145150155160tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu165170175gluglntyrasnserthrtyrargvalvalservalleuthrvalleu180185190hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn195200205lysalaleuproalaproileglulysthrileserlysalalysgly210215220glnproarggluproglnvaltyrthrleuproproserargaspglu225230235240leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr245250255proseraspilealavalglutrpgluserasnglyglnprogluasn260265270asntyrlysthrthrproprovalleuaspseraspglyserphephe275280285leutyrserlysleuthrvalasplysserargtrpglnglnglyasn290295300valphesercysservalmethisglualaleuhisasnhistyrthr305310315320glnlysserleuserleuserpro325当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1