异银杏双黄酮在制备促进肿瘤细胞自噬的抗肿瘤药物中的应用的制作方法
本发明涉及癌症药物技术领域,尤其涉及异银杏双黄酮在制备促进肿瘤细胞自噬的抗肿瘤药物中的应用。
背景技术:
肿瘤(tumour)是指机体在各种致瘤因子作用下,局部组织细胞增生所形成的新生物,因为这种新生物多呈占位性块状突起,也称赘生物(neoplasm)。癌症是危害人类生命健康的重大疾病之一,据最新who统计,仅2012年世界范围内约有820万人死于癌症,其中中国新增307万癌症患者并造成约220万人死亡,分别占全球总量的21.9%和26.8%。近年来,肿瘤的术前或放疗前化学治疗的引入越来越受国内外学者的重视,随着肿瘤治疗学的日趋完善以及化疗新药不断产生,人们逐渐认识到化学治疗在癌症治疗中的重要地位,辅助性化疗及同步放化疗已成癌症治疗不可或缺的重要治疗手段。化学治疗虽能降低癌症死亡率,但现有的抗肿瘤药具有杀伤肿瘤细胞的同时对正常细胞也有较大的杀伤性,毒副反应较大,肿瘤仍然是严重威胁人们生命和生活质量的疾病之一。同时,面对如此严峻的现实问题,长期以来,各国的政府部门、研究机构、制药企业一直对恶性肿瘤的治疗方法和抗肿瘤药物的研究开发高度重视,并不惜投入巨额资金。因此,开发出新的疗效好,且安全的肿瘤治疗药物对于救治肿瘤病人意义重大。
双黄酮类化合物是银杏中的一种黄酮类化合物。银杏叶中存在多种双黄酮类化合物,包括银杏黄酮、银杏叶素、银杏素、异银杏黄酮、蛇床子素、四氢黄酮、红杉黄酮和等色氨酸。其中异银杏双黄酮分子式为c32h22o10,分子量为566.518,cas号为548-19-6。被报道在免疫和骨髓瘤、乳腺癌、宫颈癌方面具有一定作用,但是其他癌症的治疗并见过报道。
细胞自噬过程通常起源于各种应激条件,如饥饿、缺氧、氧化应激、蛋白质聚集、内质网应激等。这些信号通路的共同靶点是ulk复合物(由ulk1/2、atg13、fip200和atg101组成),并导致ulk复合物易位到内质网结构域。进入内质网后,ulk复合物通过pi3k复合物i(由beclin-1、pik3c3/vps34、pik3r4/p150、ambra1和atg14/barkor组成)启动吞噬细胞的成核。目前,参与吞噬细胞形成的膜的来源尚不清楚,可能是内质网、线粒体、高尔基体和质膜。吞噬体形成后,它会扩张并分离受损的细胞器(包括内质网、核糖体和线粒体等)和错误折叠的蛋白质。当吞噬细胞扩张时,细胞膜弯曲,最终形成双层膜结构自噬体。atg12-atg5-atg16l1复合物和lc3磷脂酰乙醇胺(pe)结合物在吞噬细胞的伸长和闭合过程中起重要作用。最后,自噬体与溶酶体融合形成自噬体,自噬体中的物质被溶酶体中的酸性水解酶降解。此外,降解过程中产生的营养物质被释放回细胞质,并被细胞重新利用。尽管自噬使细胞能够在癌症不同阶段所面临的应激条件下存活,但过度的自噬和自噬性细胞死亡可能杀死癌细胞并限制其进展和转移。
很多肿瘤细胞的生存依赖于糖酵解产生的atp,尤其是在低氧环境下。这种依赖性为基于糖酵解设计靶向药物提供了潜在的策略。肝细胞癌,同其他癌症类型一样,临床上发现很多糖酵解相关的关键酶和蛋白质在肿瘤组织中出现上调,包括葡萄糖转运蛋白家族,特别是葡萄糖转运蛋白1(glut1)。这些酶和蛋白质构成了潜在的治疗靶点。事实上,对于其中一些靶点,已经报道了几种抑制剂,如2-脱氧葡萄糖、伊马替尼或类黄酮。糖酵解抑制剂作为一类新型的抗癌药物,其开发前景仍有待进一步探讨和挖掘。
技术实现要素:
本发明的目的是针对现有技术中的不足,提供异银杏双黄酮在制备促进肿瘤细胞自噬的抗肿瘤药物中的应用。
为实现上述目的,本发明采取的技术方案是:
提供异银杏双黄酮在制备促进肿瘤细胞自噬的抗肿瘤药物中的应用,上述异银杏双黄酮通过抑制葡萄糖转运蛋白表达,促进肿瘤细胞自噬发挥抗肿瘤作用。
进一步地,所述肿瘤为肝癌、肺癌、食道癌、胃癌、结直肠癌、胰腺癌、白血病、脑瘤或皮肤癌。
进一步地,所述抗肿瘤药物还包括药物学上可接受载体。
进一步地,所述抗肿瘤药物的剂型为注射剂、胶囊剂、片剂、粉剂、颗粒剂、茶剂或口服液。
本发明采用以上技术方案,与现有技术相比,具有如下技术效果:
本发明发现异银杏双黄酮可以显著性的抑制癌细胞增殖,迁移以及克隆形成能力,同时裸鼠成瘤实验表明异银杏双黄酮能够有效抑制裸鼠皮下肿瘤细胞的体积和重量增长。进一步的机制探究发现,异银杏双黄酮可以抑制葡萄糖转运蛋白家族基因的表达,进而导致细胞处于饥饿状态,从而激活ampk通路促进肿瘤细胞发生杀伤性自噬;本发明为肿瘤治疗提供了潜在的药物。
附图说明
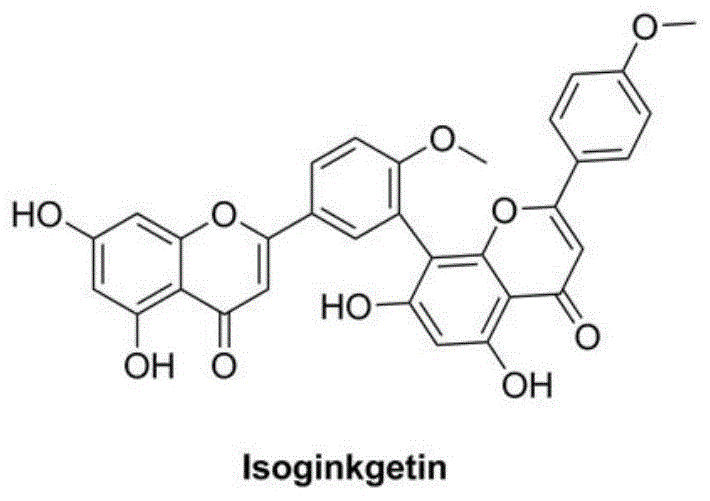
图1为异银杏双黄酮的分子式;
图2中(a)为异银杏双黄酮可以特异性杀伤各类肝癌细胞的统计结果图,(b)为异银杏双黄酮可以显著抑制肝癌细胞hepg2的克隆形成能力结果图;
图3中(a、b)为异银杏双黄酮抑制肝癌细胞hepg2迁移的结果和统计图,(c、d)为异银杏双黄酮抑制肝癌细胞huh7侵袭的结果和统计图;
图4为实施例2中裸鼠成瘤实验的结果图;
图5中(a)为异银杏双黄酮促进lc3ii/lc3i的比例结果图;(b)为异银杏双黄酮促进自噬相关基因p62和lc3的mrna水平统计图;(c)为异银杏双黄酮促进自噬溶酶体的形成;(d)为抑制自噬能减缓异银杏双黄酮对肝癌细胞的杀伤作用统计图;
图6中rna-seq结果图,其中,(a)展示异银杏双黄酮处理细胞后有1253个上调的基因和1488个下调的基因;(b)热图展示差异基因表达,红色表示高表达基因,蓝色表示低表达基因;(c)差异基因的信号通路分析;(d)差异基因的信号通路分析;
图7中(a)异银杏双黄酮处理后葡萄糖转运蛋白家族基因的表达受到抑制;(b)荧光d-葡萄糖同系物展示异银杏双黄酮处理后细胞的葡萄糖摄取受到抑制;(c)异银杏双黄酮处理后ampk下游蛋白ulk1的磷酸化升高。
具体实施方式
下面结合附图和具体实施例对本发明作进一步说明,但不作为本发明的限定。需要说明的是,在不冲突的情况下,本发明中的实施例及实施例中的特征可以相互组合。
实施例1异银杏双黄酮对肝癌细胞生长增殖、克隆形成及侵袭迁移的影响
1.1.cck8实验
传5000个/孔肝癌细胞到96孔板,24小时候后加入1、5、10、20、30、40μm的异银杏双黄酮,每组样品传五个平行。培养24h后加入化合物处理48h,然后每孔加入10μl的cck8溶液,培养箱培养4h后。最后在酶标仪上选取450nm波长读取吸光值。
1.2.细胞克隆形成实验
传600个/孔到24孔板,然后连续培养两周,随后经常观察,当培养皿中出现肉眼可见的克隆时,终止培养。弃去上清液,用pbs小心浸洗2次。加4%多聚甲醛固定15分钟。然后吸走固定液,加适量giemsa应用染色液染10~30分钟,然后用pbs清洗三次,空气干燥。拍照整理数据。
1.3.transewll实验
6×105hepg2细胞在无血清或生长因子的培养基重悬后培养在matrigel室(24孔;孔径8μm;bdbiosciences)的上室中,并且在下室中使用补充有10%胎牛血清的培养基作为引诱剂。孵育36小时后,取出小室,吸走上室培养基,并用干净的棉签擦拭上室,在24孔板另一个干净的孔中加入1ml无钙pbs,将小室放入pbs中,清洗两次。再加入600μl甲醇室温固定30min。固定结束后将小室倒扣在桌面风干5min。配制0.1%结晶紫溶液(pbs溶解,质量体积比),取600μl结晶紫溶液到24孔板内,将风干后的小室正面朝放进结晶紫溶液中,室温染20min。用pbs清洗三次,并用棉签擦拭上室。将小室取出倒扣在桌面上风干5min。
实验结果:首先将各类肝癌细胞系h22,hep1-6,hepg2,huh7和永生化的肝正常细胞系lo-2铺到96孔板中,24小时候后加入1、5、10、20、30、40μm的异银杏双黄酮,然后cck8检测各类细胞的增殖情况,结果如图2a所示,在20μm浓度时,异银杏双黄酮可以特异性杀伤各类肝癌细胞系,而对lo-2没有影响。同时,异银杏双黄酮可以显著抑制肝癌细胞hepg2的克隆形成能力(结果如图2b所示)。并且,异银杏双黄酮能够抑制肝癌细胞hepg2的迁移(结果如图3a、b)及抑制肝癌细胞huh7的侵袭(结果如图3c、d)。
实施例2异银杏双黄酮体内抗肝癌药效评价结果
2.1裸鼠成瘤实验
购买裸鼠,裸鼠类型balb/c-nu,四周龄,雄性。根据实验目的确定老鼠数量,保证每种类型的细胞至少可以接种十个点。将裸鼠放置在spf级动物房内饲养。提前做预实验确定合适的细胞量,以hepg2细胞为例,每个点注射8×106个细胞,半个月左右肿瘤已肉眼可见。确定好细胞量后,胰酶消化收集细胞,pbs清洗一遍,再用pbs重悬细胞,细胞计数,保证细胞密度为8×106个/ml,将装有细胞的ep管置于冰上,经过严格的消毒后拿进动物房。用1ml注射器(拿掉针头)轻轻吹打细胞系,然后吸上细胞后套上针头,轻轻弹注射器并排出气泡,向老鼠的四肢腋下以z字形方式注射100μl的pbs细胞悬液。每隔两三天到动物房内观察老鼠状态以及肿瘤大小,当肿瘤肉眼可见时,将长肿瘤的老鼠随机分为四组:对照组;低异银杏双黄酮组(25mg/kg);高异银杏双黄酮组(50mg/kg)和lenvatinib组(5mg/kg)。然后连续两周腹腔注射药物,每次观察时都记录下肿瘤的长和宽,待肿瘤大小长得很大的时候,处死老鼠,解剖取出瘤子,称量体重并拍照。
通过裸鼠成瘤实验发现异银杏双黄酮能显著抑制肝癌细胞的成瘤大小和体积(图4a、b所示),同时阳性对照lenvatinib组在抑制肿瘤大小的同时对小鼠脾指数有抑制作用,而异银杏双黄酮则没有(图4c所示)。并且,通过he染色发现异银杏双黄酮抑制肿瘤的血管生成(图4d所示)。
实施例3异银杏双黄酮具有促进肝癌细胞自噬的抗肝癌活性,且其抗肝癌作用与细胞自噬密切相关
3.1免疫荧光共聚焦实验
细胞固定:吸走培养基,预冷pbs洗一遍,加入500μl3.7%的甲醛,室温放置15-20min。随后dapi染色:1mlpbs中加入1μldapi染料(1:1000),充分混匀。将盖玻片正面朝上放回12孔板,加入500μl含dapi的pbs,轻轻晃动培养皿,室温静置1-2min,然后用pbs洗三次,超纯水洗一次,每次5min。最后封片:在载玻片上滴适量封片剂,然后将盖玻片以倾斜的形式缓缓倒扣在载玻片上,室温避光过夜。可以冻存在-80℃或用共聚焦免疫荧光显微镜观察。
3.2稳定过表达细胞系的构建
以肝癌细胞hepg2为例,传2×105个细胞到六孔板内,至少传六个孔。细胞培养36h后分别加入puromycin,终浓度分别为0、0.2、0.5、1.0、2.0、4.0μg/ml,培养48h后观察细胞状态,最终确定puromycin终浓度为4μg/ml。病毒包装是在293t细胞内完成的,第一天晚上传代293t,第二天上午做转染。传代步骤及转染方法同磷酸钙转染法。转染12h后更换新的培养基(因为ca2+会阻碍病毒包装)。第三天晚上传代hepg2细胞,传2×105个细胞到六孔板内。第四天上午,吸干hepg2细胞的培养基,将293t包装后的病毒上清培养基用注射器吸上来,并用0.45μm的滤器过滤后加入hepg2细胞中。最后再加入1000×的聚凝胺(polybrene,10mg/ml),摇匀后放入培养箱培养24h,更换培养基再培养12h(病毒感染时间过程有可能导致细胞状态不好)。未感染病毒的细胞作为对照。注意:所有与病毒接触过的实验材料都必须泡84消毒液。加入puromycin使其终浓度为4μg/ml,48h后观察细胞状态,若对照细胞全死而感染细胞还有活的,就将活的细胞进行传代培养;如对照细胞没死,可以进行第二次加puromycin筛选。蛋白免疫印记或rt-pcr法检测细胞稳定表达情况。
实验结果:在hepg2细胞中加入异银杏双黄酮组处理后,westernblot发现异银杏双黄酮能显著提高lc3ii/lc3i的比例(图5a所示)、且促进自噬相关基因p62和lc3的mrna水平的表达(图5b所示)。
进一步,通过慢病毒包装系统建立了rfp-gfp-lc3的稳定表达的hepg2细胞系。在稳定表达系中通过免疫荧光共聚焦实验证明异银杏双黄酮能促进自噬溶酶体的形成(图5c所示)。
通过加入自噬抑制剂氯喹(cq)和异银杏双黄酮联用,cck8实验表明抑制自噬后能显著减少异银杏双黄酮对肝癌细胞的杀伤作用,提示异银杏双黄酮是通过自噬杀伤肿瘤细胞的(图5d所示)。
实施例4异银杏双黄酮能够抑制葡萄糖转运蛋白家族基因的表达促进肝癌细胞自噬,从而抑制肝癌细胞的生长增殖和侵袭迁移
4.1葡萄糖摄入检测实验
以肝癌细胞hepg2为例,传2×105个细胞到12孔板内,培养24小时后,pbs清洗三次,加入无血清培养基培养三个小时,然后加入0.5mm的荧光d-葡萄糖同系物2-nbdg,培养箱孵育20min后荧光显微镜下观察细胞。
为了进一步探究异银杏双黄酮促进自噬的分子机制,通过二代测序(rna-seq)发现(图6a-d所示),异银杏双黄酮能显著抑制葡萄糖转运蛋白家族基因的表达(图7a所示),荧光d-葡萄糖同系物实验表明异银杏双黄酮能抑制细胞内葡萄糖的摄入(图7b所示),westernblotting实验表明异银杏双黄酮能激活ampk下游的ulk1的磷酸化水平(图7c所示),提示异银杏双黄酮是通过抑制葡萄糖转运蛋白家族基因的表达,进而导致细胞处于饥饿状态,从而激活ampk通路促进肝癌细胞发生杀伤性自噬。
上所述仅为本发明较佳的实施例,并非因此限制本发明的实施方式及保护范围,对于本领域技术人员而言,应当能够意识到凡运用本发明说明书内容及图示所作出的等同替换和显而易见的变化所得到的方案,均应当包含在本发明的保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!