针对HIV‑1V1V2ENV区域的广谱中和性单克隆抗体的制作方法
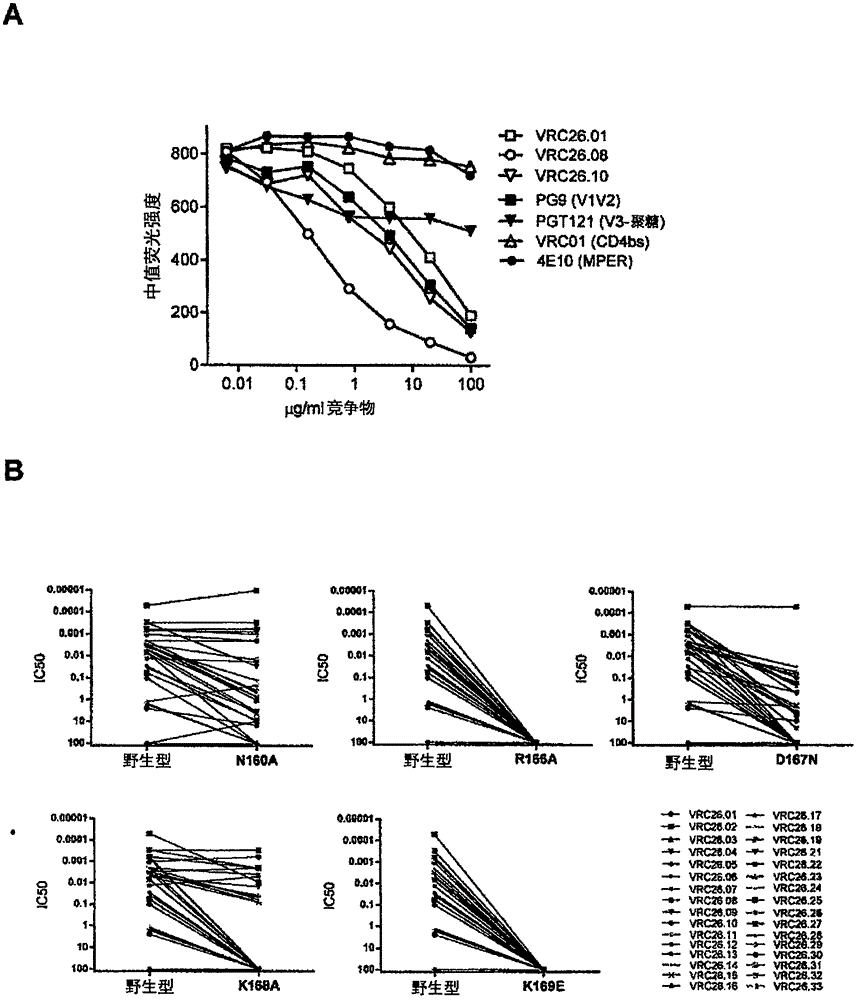
本发明涉及抗-HIV疗法和预防。具体地,本发明涉及针对HIV-1的广谱中和性抗体、编码这些抗体的核酸、包含所述核酸的载体、以及包含所述载体和/或抗体的细胞和药物组合物。本发明也涉及所述抗体和/或载体用于治疗和/或预防受试者中的HIV-1感染的用途。此外,本发明也涉及含有本发明的抗体的试剂盒。如果可以理解和再现导致其引起的分子事件,有效地中和HIV-1的抗体代表引导HIV-1疫苗接种策略的潜在模板(Haynes等人.2012;Kong等人.2012;Moir等人.2011;McCoy等人.2013;Overbaugh和Morris2012;Burton等人.2012)。基本上所有被感染的个体在感染的几个月内引起有效的抗体应答,但是这些抗体优先中和快速逃逸的自体病毒(Montefiori等人.1991;Richman等人.2003;Wei等人.2003;Bunnik等人.2008)。在感染的2-3年以后在约20%的供体中产生能够中和大多数HIV-1株的交叉反应性抗体(vanGils等人.2009;Gray等人.2011;Piantadosi等人.2009;Sather等人.2009;Doria-Rose等人.2010)。研究(和引起)广谱中和性抗体的挑战是,它们通常是亚优势应答,并且被无效应答或毒株特异性抗体应答在数量上远远胜过(Kong等人.2012;Overbaugh和Morris2012;Pantophlet等人.2006;Kwong等人.2012;Mascola和Haynes等人.2013)。重要的是,不清楚广谱中和性抗体是否从在数年的感染中成熟的早期株系特异性B细胞谱系形成,或者来自罕见随机事件的宽度结果是否刺激快速地变成与HIV-1交叉反应性的新B细胞谱系。描绘导致HIV-1-特异性抗体应答的形成的分子事件的一种方式包括,分离广谱中和性单克隆抗体和用下一代测序(NGS)检查B细胞遗传记录(Glanville等人.2009;Tian等人.2008;Briney等人.2012;Boyd等人.2010)。在中和抗体谱系形成以后在单个时间点的取样允许研究扩大的抗体家族和阐明具有较低体细胞突变水平的较早谱系序列(Wu等人.2011;Zhu等人.2012;Zhu等人.2013)。从感染时间以后早期用纵向取样可以获得甚至更大的洞察,如关于靶向HIV-1包膜糖蛋白(Env)的CD4-结合位点的CH103抗体谱系所证实的(Liao等人.2013)。在感染以后136周从抗原-特异性的B细胞分离几种成熟的CH103中和单克隆抗体。然后使用NGS鉴别CH103谱系的转录物,其在早至感染以后14周就被观察到,远远早于在该供体中检测到血浆交叉中和活性。值得注意的是,CH103的推断的未突变的共同祖先(UCA)结合自体传播的病毒/起始病毒,并响应于病毒多样化而进化以获得必要的体细胞突变以有效地中和HIV-1的异源株。由于CD4结合位点仅是HIV-1Env上的几个保守中和表位之一,需要另外的研究来理解靶向HIV-1的其它脆弱区域的有效中和抗体的遗传决定子和成熟途径。来自HIV-1感染的供体的血清中和抗体经常靶向HIV-1Env的V1V2区域(Gray等人.2011;Walker等人.2010;Lynch等人.2011;Tomaras等人.2011),并且在RV144HIV疫苗试验中证实针对该区域的结合抗体与保护有关(Haynes等人.2012)。另外,已经从几个供体分离出针对V1V2区域的有效中和单克隆抗体。这些包括:PG9和PG16,其中和70-80%的循环的HIV-1分离物(Walker等人.2009);CH01-04,其中和40-50%(Bonsignori等人.2011);和PGT141-145,其中和40-80%(Walker等人.2011)。所有这些抗体的特征在于长的第3-重链互补性决定区(CDRH3),其是突出的、阴离子的和酪氨酸硫酸化的(Pejchal等人.2010;McLellan等人.2011;Pancera等人.2013)。含有PG9和PG16的Env复合物的晶体和电子显微结构揭示,CDRH3会穿透HIV-1聚糖屏障,从而识别HIV-1刺突的膜-远端顶点处的季糖肽表位,其通过V1V2的结合从三个gp120原聚体形成(McLellan等人.2011;Pancera等人.2013;Julien等人.2013)。为了更精确地理解哪些因素在通向有效的V1V2-定向的抗体的途径中是关键性的,我们分析了供体CAP256,其在以前被证实会形成有效的V1V2-定向的血浆应答(Moore等人.2011;Moore等人.2013;Georgiev等人.2013)。通过纵向取样分析了该供体,发现在第15周是超感染的,并在一年时表现出适度中和宽度。到感染后三年,来自CAP256的血浆中和了所有HIV-1毒株中的77%,除了特别是来自亚型A和C的那些(Gray等人.2011)。我们在这里从该供体分离了有效的V1V2-定向的广谱中和性抗体,进行NGS以能够详细理解谱系进化,并确定晶体结构以限定其分子特征。还纵向分析了循环的血浆病毒以理解病毒Env进化和免疫应答之间的相互作用。结果允许精确地描述产生这类有效抗体的分子事件。技术实现要素:在本发明的第一方面,提供了一种分离的结合HIV的V1V2表位的抗-HIV抗体,其中所述抗体包含重链可变区和轻链可变区中的一个或两者,所述重链可变区包含SEQIDNO:121的共有氨基酸序列,所述轻链可变区包含SEQIDNO:122的共有氨基酸序列。在一个实施方案中,所述分离的抗-HIV抗体以小于0.5μg/ml的IC50浓度中和自体HIV病毒CAP256.SU或以小于0.5μg/ml的IC50浓度中和异源HIV病毒ZM53.12。在本发明的另一个方面,提供了一种分离的抗-HIV抗体,其选自CAP256-VRC26-UCA、CAP256-VRC26-I1、CAP256-VRC26-I2、CAP256-VRC26.01、CAP256-VRC26.02、CAP256-VRC26.03、CAP256-VRC26.04、CAP256-VRC26.05、CAP256-VRC26.06、CAP256-VRC26.07、CAP256-VRC26.08、CAP256-VRC26.09、CAP256-VRC26.10、CAP256-VRC26.11、CAP256-VRC26.12、CAP256-VRC26.25、CAP256-VRC26.26和CAP256-VRC26.27。在本发明的一个实施方案中,所述抗体是CAP256-VRC26.01,且所述抗体有效地中和至少19%的选自96ZM651.02、CAP210.E8、CAP244.D3、CAP45.G3、DU156.12、DU172.17、DU422.01、ZM109.4、ZM135.10a、ZM197.7、ZM214.15、ZM233.6、ZM249.1、ZM53.12、0260.v5.c36、BG505.w6m、KER2008.12、KER2018.11、MB201、MB539.2B7、Q168.a2、Q23.17、Q259.d2.17、Q461.e2、Q769.d22、Q842.d12、RW020.2、UG037.8、6535.3、AC10.29、CAAN.A2、PVO.04、QH0692.42、REJO.67、RHPA.7、SC422.8、THRO.18、TRJO.58、TRO.11、WITO.33、191821.E6.1、3016.V5.c36、6405.v4.c34、C1080.c3、CM244.ec1、CNE3和TH976.17的HIV-1毒株。在本发明的一个实施方案中,所述抗体是CAP256-VRC26.02,且所述抗体有效地中和至少17%的选自上述毒株的HIV-1毒株;或所述抗体是CAP256-VRC26.03,且所述抗体有效地中和至少36%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.04,且所述抗体有效地中和至少30%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.05,且所述抗体有效地中和至少21%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.06,且所述抗体有效地中和至少17%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.07,且所述抗体有效地中和至少13%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.08,且所述抗体有效地中和至少47%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.09,且所述抗体有效地中和至少47%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.10,且所述抗体有效地中和至少23%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.11,且所述抗体有效地中和至少23%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.12,且所述抗体有效地中和至少6%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.13,且所述抗体有效地中和至少6%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.14,且所述抗体有效地中和至少26%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.15,且所述抗体有效地中和至少32%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.16,且所述抗体有效地中和至少30%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.17,且所述抗体有效地中和至少30%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.18,且所述抗体有效地中和至少28%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.19,且所述抗体有效地中和至少47%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.20,且所述抗体有效地中和至少2%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.21,且所述抗体有效地中和至少13%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.22,且所述抗体有效地中和至少47%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.23,且所述抗体有效地中和至少6%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.24,且所述抗体有效地中和至少37%的选自上述毒株的HIV-1毒株;或所述抗体是CAP256-VRC26.25,且所述抗体有效地中和至少63%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.26,且所述抗体有效地中和至少59%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.27,且所述抗体有效地中和至少59%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.28,且所述抗体有效地中和至少41%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.29,且所述抗体有效地中和至少46%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.30,且所述抗体有效地中和至少28%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.31,且所述抗体有效地中和至少20%的选自上述毒株的HIV-1毒株;所述抗体是CAP256-VRC26.32,且所述抗体有效地中和至少20%的选自上述毒株的HIV-1毒株;或所述抗体是CAP256-VRC26.33,且所述抗体有效地中和至少22%的选自上述毒株的HIV-1毒株。另一个方面提供了一种分离的抗-HIV抗体,其包含选自SEQIDNO:1-60、SEQIDNO:170-173、SEQIDNO:178-181或SEQIDNO:186-189的氨基酸序列。在另一个实施方案中,所述抗-HIV抗体包含:i)包含SEQIDNO:3的氨基酸序列的重链序列和包含SEQIDNO:4的氨基酸序列的轻链序列;或者ii)包含SEQIDNO:7的氨基酸序列的重链序列和包含SEQIDNO:8的氨基酸序列的轻链序列;或者iii)包含SEQIDNO:11的氨基酸序列的重链序列和包含SEQIDNO:12的氨基酸序列的轻链序列;或者iv)包含SEQIDNO:15的氨基酸序列的重链序列和包含SEQIDNO:16的氨基酸序列的轻链序列;或者v)包含SEQIDNO:19的氨基酸序列的重链序列和包含SEQIDNO:20的氨基酸序列的轻链序列;或者vi)包含SEQIDNO:23的氨基酸序列的重链序列和包含SEQIDNO:24的氨基酸序列的轻链序列;或者vii)包含SEQIDNO:27的氨基酸序列的重链序列和包含SEQIDNO:28的氨基酸序列的轻链序列;或者viii)包含SEQIDNO:31的氨基酸序列的重链序列和包含SEQIDNO:32的氨基酸序列的轻链序列;或者ix)包含SEQIDNO:35的氨基酸序列的重链序列和包含SEQIDNO:36的氨基酸序列的轻链序列;或者x)包含SEQIDNO:39的氨基酸序列的重链序列和包含SEQIDNO:40的氨基酸序列的轻链序列;或者xi)包含SEQIDNO:43的氨基酸序列的重链序列和包含SEQIDNO:44的氨基酸序列的轻链序列;或者xii)包含SEQIDNO:47的氨基酸序列的重链序列和包含SEQIDNO:48的氨基酸序列的轻链序列;或者xiii)包含SEQIDNO:51的氨基酸序列的重链序列和包含SEQIDNO:52的氨基酸序列的轻链序列;或者xiv)包含SEQIDNO:55的氨基酸序列的重链序列和包含SEQIDNO:56的氨基酸序列的轻链序列;或者xv)包含SEQIDNO:59的氨基酸序列的重链序列和包含SEQIDNO:60的氨基酸序列的轻链序列;或者xvi)包含SEQIDNO:172的氨基酸序列的重链序列和包含SEQIDNO:173的氨基酸序列的轻链序列;或者xvi)包含SEQIDNO:180的氨基酸序列的重链序列和包含SEQIDNO:181的氨基酸序列的轻链序列;或者xvi)包含SEQIDNO:188的氨基酸序列的重链序列和包含SEQIDNO:189的氨基酸序列的轻链序列。在本发明的一个实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:3的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:3的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:3的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:4的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:4的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:4的位置90-101处的氨基酸的CDR3。备选地,在另一个实施方案中,所述分离的抗-HIV抗体包括重链和轻链,所述重链具有包含在SEQIDNO:7的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:7的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:7的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:8的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:8的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:8的位置90-101处的氨基酸的CDR3。在另一个实施方案中,所述分离的抗-HIV抗体可以包括重链和轻链,所述重链具有包含在SEQIDNO:11的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:11的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:11的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:12的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:12的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:12的位置90-101处的氨基酸的CDR3。备选地,在另一个实施方案中,所述分离的抗-HIV抗体可以包括重链和轻链,所述重链具有包含在SEQIDNO:15的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:15的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:15的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:16的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:16的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:16的位置90-101处的氨基酸的CDR3。在另一个实施方案中,所述分离的抗体包括重链和轻链,所述重链具有包含在SEQIDNO:19的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:19的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:19的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:20的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:20的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:20的位置90-98处的氨基酸的CDR3。在另一个实施方案中,所述分离的抗体包括重链和轻链,所述重链具有包含在SEQIDNO:23的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:23的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:23的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:24的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:24的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:24的位置90-101处的氨基酸的CDR3。在另一个实施方案中,所述分离的抗体可以包括重链和轻链,所述重链具有包含在SEQIDNO:27的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:27的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:27的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:28的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:28的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:28的位置90-101处的氨基酸的CDR3。在另一个实施方案中,所述分离的抗体包括重链和轻链,所述重链具有包含在SEQIDNO:31的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:31的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:31的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:32的位置23-30处的氨基酸的CDR1、包含在SEQIDNO:32的位置48-50处的氨基酸的CDR2和包含在SEQIDNO:32的位置90-98处的氨基酸的CDR3。另一个实施方案提供了一种分离的抗体,其包括重链和轻链,所述重链具有包含在SEQIDNO:35的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:35的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:35的位置97-134处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:36的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:36的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:36的位置90-98处的氨基酸的CDR3。根据另一个实施方案,所述分离的抗体包括重链和轻链,所述重链具有包含在SEQIDNO:39的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:39的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:39的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:40的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:40的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:40的位置90-101处的氨基酸的CDR3。在本发明的另一个实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:43的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:43的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:43的位置97-135处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:44的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:44的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:44的位置90-101处的氨基酸的CDR3。在本发明的另一个实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:47的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:47的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:47的位置97-135处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:48的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:48的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:48的位置90-101处的氨基酸的CDR3。在本发明的另一个实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:51的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:51的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:51的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:52的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:52的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:52的位置90-101处的氨基酸的CDR3。在本发明的一个备选实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:55的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:55的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:55的位置96-132处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:56的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:56的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:56的位置90-97处的氨基酸的CDR3。在本发明的另一个实施方案中,所述抗体包括重链和轻链,所述重链具有包含在SEQIDNO:59的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:59的位置51-58处的氨基酸的CDR2和包含在SEQIDNO:59的位置97-133处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:60的位置26-33处的氨基酸的CDR1、包含在SEQIDNO:60的位置51-53处的氨基酸的CDR2和包含在SEQIDNO:60的位置90-97处的氨基酸的CDR3。本发明的另一个实施方案提供了一种抗体,其包括重链和轻链,所述重链具有包含在SEQIDNO:172的位置31-35处的氨基酸的CDR1、包含在SEQIDNO:172的位置50-66处的氨基酸的CDR2和包含在SEQIDNO:172的位置99-134处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:173的位置23-35处的氨基酸的CDR1、包含在SEQIDNO:173的位置51-57处的氨基酸的CDR2和包含在SEQIDNO:173的位置90-101处的氨基酸的CDR3。本发明的另一个实施方案提供了一种抗体,其包括重链和轻链,所述重链具有包含在SEQIDNO:180的位置31-35处的氨基酸的CDR1、包含在SEQIDNO:180的位置50-66处的氨基酸的CDR2和包含在SEQIDNO:180的位置99-135处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:181的位置23-35处的氨基酸的CDR1、包含在SEQIDNO:181的位置51-57处的氨基酸的CDR2和包含在SEQIDNO:181的位置90-101处的氨基酸的CDR3。本发明的另一个实施方案提供了一种抗体,其包括重链和轻链,所述重链具有包含在SEQIDNO:188的位置31-35处的氨基酸的CDR1、包含在SEQIDNO:188的位置50-66处的氨基酸的CDR2和包含在SEQIDNO:188的位置99-135处的氨基酸的CDR3,所述轻链具有包含在SEQIDNO:189的位置23-35处的氨基酸的CDR1、包含在SEQIDNO:189的位置52-57处的氨基酸的CDR2和包含在SEQIDNO:189的位置90-101处的氨基酸的CDR3。本发明的第四方面提供了一种组合物,其包含本发明的分离的抗-HIV抗体或其片段。在一个实施方案中,本发明提供了一种核酸分子,其编码本发明的分离的抗-HIV抗体或其片段。其它实施方案提供了载体以及含有所述载体的细胞,所述载体包含编码本发明的分离的抗-HIV抗体或其片段的核酸分子。本发明的第五方面提供了一种组合物,其包含编码本发明的分离的抗-HIV抗体或其片段的核酸分子、包含所述核酸的载体或含有所述载体的细胞。本发明的另一个实施方案提供了一种药物组合物,其包含至少一种本发明的抗体或片段和药用载体。本发明的另一个实施方案提供了一种药物组合物,其包含至少一种编码本发明的分离的抗-HIV抗体或其片段的核酸分子、包含所述核酸的载体或含有所述载体的细胞和药用载体。在本发明的另一个方面,提供了一种针对HIV感染或HIV相关疾病进行免疫或治疗HIV感染或HIV相关疾病的方法,所述方法包括给需要免疫或治疗的受试者施用治疗有效量的至少一种本发明的HIV抗体。所述施用可以包括第二种治疗剂的施用。优选地,所述第二种治疗剂是抗病毒剂。在本发明的另一个方面,提供了一种针对HIV感染或HIV相关疾病进行免疫或治疗HIV感染或HIV相关疾病的方法,所述方法包括给需要免疫或治疗的受试者施用治疗有效量的至少一种编码本发明的分离的抗-HIV抗体或其片段的核酸分子、包含所述核酸的载体或含有所述载体的细胞。所述施用可以包括第二种治疗剂的施用。优选地,所述第二种治疗剂是抗病毒剂。本说明书还提供了一种疫苗,其包含特异性地结合本发明的抗体的表位。在本发明的另一个方面,提供了一种检测本发明的抗-HIV抗体或结合本发明的抗-HIV抗体的抗原或表位在患者中的存在的方法,所述方法包括:从所述患者分离生物样品,和针对所述抗-HIV抗体的存在、或结合本发明的抗-HIV抗体的抗原或表位或者含有编码所述抗体的DNA或mRNA中的至少一种的细胞的存在来测定所述生物样品。另一个方面提供了包含至少一种本发明的抗体或片段的试剂盒。附图说明现在将仅仅作为示例并参考以下附图来描述本发明的非限制性实施方案:图1:供体CAP256实现的广谱中和性的发展和中和抗体的分离。33种分离的抗体CAP256-VRC26.01-33的遗传特征和中和宽度和效能。相对于46种Env-假病毒的集合评估中和。图2:到HIV-1包膜糖蛋白的VIV2区域的CAP256-VRC26抗体的表位定位。(a)结合竞争指示CAP256-VRC26抗体在与V1V2-定向的抗体PG9相同的竞争组中。通过流式细胞计量术测量标记的CAP256-VRC26.08和未标记的竞争抗体对表达ZM53-Env的293T细胞的结合。(b)CAP256-VRC26抗体对ConC(野生型)和它的V2突变体的中和。每对点显示了一种病毒和各种突变体的IC50。图3:通过B细胞转录物的下一代测序(NGS)揭示的CAP256-VRC26谱系的成熟。重链和轻链的CAP256-VRC26克隆谱系的系统发生树。图4:形成中的CAP256-VRC26谱系的结构特征。在带状简图表示中显示了CAP256-VRC26.03的抗原结合片段(Fab)的晶体结构。图5:从UCA至CAP256-VRC26.01的发展。顶行显示了针对自体原代病毒(PI)、超级感染病毒(SU)和一组具有逃逸突变的SU病毒的抗体VRC26-UCA、VRC26-I1、VRC26-I2和VRC26.01的中和。底行显示了CAP256-VRC26.01谱系抗体对异源病毒的中和。针对显示的七种VRC26.01-敏感毒株的集合来评估中和。图6:CAP256-VRC26重链的序列。将33种B-细胞培养物衍生出的抗体推论的中间体的序列与种系V和J基因进行对比。图7:CAP256-VRC26λ轻链的序列。将33种B-细胞培养物衍生出的抗体推论的中间体的序列与种系V和J基因进行对比。图8:与其它众所周知的mAb相比的CAP256-VRC26.25对进化枝C的效能和宽度。图9:CAP256-VRC26.08和CAP256-VRC26.25对194种病毒的中和(IC50值)。对于<50的值,计算几何平均值。图10:CAP256血浆和mAb对异源病毒的中和。CAP256-VRC26mAb和来自在4个时间点取样的CAP256的血浆对31种病毒的中和。血浆数据来自Moore等人,2011。将抗体值显示为所有31种抗体的理论组合的50%抑制浓度(IC50)(计算为每种病毒的最低IC50)。血浆被显示为50%抑制稀释度(ID50),起始稀释度为1∶45。图11:针对Env克隆测试的31种CAP256-VRC26mAb中的每一种的中和(IC50值)。图12:V2表位多态性和逃逸突变对CAP256-VRC26中和的影响。SU和PI病毒以及突变成含有PI多态性162I、165V或169Q的SU病毒的CAP256-VRC26mAb中和。将V2表位序列(残基160-171)显示在左侧,并将中和IC50值显示在右侧。图13:CAP256-VRC26抗体的最小自身反应性。(a).在50和25μg/ml评估对Hep2细胞的染色。仅阳性对照mAb4E10显示阳性染色。(b)关于与心磷脂的结合的ELISA。4E10是强阳性的,CAP256-VRC26.03是弱阳性的,且其它CAP256-VRC26mAb和UCA与对照抗体VRC01一起是阴性的。序列表使用核苷酸碱基的标准字母缩写和氨基酸的标准三字母缩写显示了在附随的序列表中列出的核酸和氨基酸序列。本领域技术人员将理解,仅显示了每个核酸序列的一条链,但是通过对显示的链的任何提及来包括互补链。在附随的序列表中:SEQIDNO:1是CAP256-VRC26-UCA的抗体重链的氨基酸序列。SEQIDNO:2是CAP256-VRC26-UCA的抗体轻链的氨基酸序列。SEQIDNO:3是CAP256-VRC26-UCA的抗体重链可变结构域的氨基酸序列。SEQIDNO:4是CAP256-VRC26-UCA的抗体轻链可变结构域的氨基酸序列。SEQIDNO:5是CAP256-VRC26-I1的抗体重链的氨基酸序列。SEQIDNO:6是CAP256-VRC26-I1的抗体轻链的氨基酸序列。SEQIDNO:7是CAP256-VRC26-I1的抗体重链可变结构域的氨基酸序列。SEQIDNO:8是CAP256-VRC26-I1的抗体轻链可变结构域的氨基酸序列。SEQIDNO:9是CAP256-VRC26-I2的抗体重链的氨基酸序列。SEQIDNO:10是CAP256-VRC26-I2的抗体轻链的氨基酸序列。SEQIDNO:11是CAP256-VRC26-I2的抗体重链可变结构域的氨基酸序列。SEQIDNO:12是CAP256-VRC26-I2的抗体轻链可变结构域的氨基酸序列。SEQIDNO:13是CAP256-VRC26.01的抗体重链的氨基酸序列。SEQIDNO:14是CAP256-VRC26.01的抗体轻链的氨基酸序列。SEQIDNO:15是CAP256-VRC26.01的抗体重链可变结构域的氨基酸序列。SEQIDNO:16是CAP256-VRC26.01的抗体轻链可变结构域的氨基酸序列。SEQIDNO:17是CAP256-VRC26.02的抗体重链的氨基酸序列。SEQIDNO:18是CAP256-VRC26.02的抗体轻链的氨基酸序列。SEQIDNO:19是CAP256-VRC26.02的抗体重链可变结构域的氨基酸序列。SEQIDNO:20是CAP256-VRC26.02的抗体轻链可变结构域的氨基酸序列。SEQIDNO:21是CAP256-VRC26.03的抗体重链的氨基酸序列。SEQIDNO:22是CAP256-VRC26.03的抗体轻链的氨基酸序列。SEQIDNO:23是CAP256-VRC26.03的抗体重链可变结构域的氨基酸序列。SEQIDNO:24是CAP256-VRC26.03的抗体轻链可变结构域的氨基酸序列。SEQIDNO:25是CAP256-VRC26.04的抗体重链的氨基酸序列。SEQIDNO:26是CAP256-VRC26.04的抗体轻链的氨基酸序列。SEQIDNO:27是CAP256-VRC26.04的抗体重链可变结构域的氨基酸序列。SEQIDNO:28是CAP256-VRC26.04的抗体轻链可变结构域的氨基酸序列。SEQIDNO:29是CAP256-VRC26.05的抗体重链的氨基酸序列。SEQIDNO:30是CAP256-VRC26.05的抗体轻链的氨基酸序列。SEQIDNO:31是CAP256-VRC26.05的抗体重链可变结构域的氨基酸序列。SEQIDNO:32是CAP256-VRC26.05的抗体轻链可变结构域的氨基酸序列。SEQIDNO:33是CAP256-VRC26.06的抗体重链的氨基酸序列。SEQIDNO:34是CAP256-VRC26.06的抗体轻链的氨基酸序列。SEQIDNO:35是CAP256-VRC26.06的抗体重链可变结构域的氨基酸序列。SEQIDNO:36是CAP256-VRC26.06的抗体轻链可变结构域的氨基酸序列。SEQIDNO:37是CAP256-VRC26.07的抗体重链的氨基酸序列。SEQIDNO:38是CAP256-VRC26.07的抗体轻链的氨基酸序列。SEQIDNO:39是CAP256-VRC26.07的抗体重链可变结构域的氨基酸序列。SEQIDNO:40是CAP256-VRC26.07的抗体轻链可变结构域的氨基酸序列。SEQIDNO:41是CAP256-VRC26.08的抗体重链的氨基酸序列。SEQIDNO:42是CAP256-VRC26.08的抗体轻链的氨基酸序列。SEQIDNO:43是CAP256-VRC26.08的抗体重链可变结构域的氨基酸序列。SEQIDNO:44是CAP256-VRC26.08的抗体轻链可变结构域的氨基酸序列。SEQIDNO:45是CAP256-VRC26.09的抗体重链的氨基酸序列。SEQIDNO:46是CAP256-VRC26.09的抗体轻链的氨基酸序列。SEQIDNO:47是CAP256-VRC26.09的抗体重链可变结构域的氨基酸序列。SEQIDNO:48是CAP256-VRC26.09的抗体轻链可变结构域的氨基酸序列。SEQIDNO:49是CAP256-VRC26.10的抗体重链的氨基酸序列。SEQIDNO:50是CAP256-VRC26.10的抗体轻链的氨基酸序列。SEQIDNO:51是CAP256-VRC26.10的抗体重链可变结构域的氨基酸序列。SEQIDNO:52是CAP256-VRC26.10的抗体轻链可变结构域的氨基酸序列。SEQIDNO:53是CAP256-VRC26.11的抗体重链的氨基酸序列。SEQIDNO:54是CAP256-VRC26.11的抗体轻链的氨基酸序列。SEQIDNO:55是CAP256-VRC26.11的抗体重链可变结构域的氨基酸序列。SEQIDNO:56是CAP256-VRC26.11的抗体轻链可变结构域的氨基酸序列。SEQIDNO:57是CAP256-VRC26.12的抗体重链的氨基酸序列。SEQIDNO:58是CAP256-VRC26.12的抗体轻链的氨基酸序列。SEQIDNO:59是CAP256-VRC26.12的抗体重链可变结构域的氨基酸序列。SEQIDNO:60是CAP256-VRC26.12的抗体轻链可变结构域的氨基酸序列。SEQIDNO:61是CAP256-VRC26-UCA的抗体重链的一个示例性核酸序列。SEQIDNO:62是CAP256-VRC26-UCA的抗体轻链的一个示例性核酸序列。SEQIDNO:63是CAP256-VRC26-UCA的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:64是CAP256-VRC26-UCA的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:65是CAP256-VRC26-I1的抗体重链的一个示例性核酸序列。SEQIDNO:66是CAP256-VRC26-I1的抗体轻链的一个示例性核酸序列。SEQIDNO:67是CAP256-VRC26-I1的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:68是CAP256-VRC26-I1的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:69是CAP256-VRC26-I2的抗体重链的一个示例性核酸序列。SEQIDNO:70是CAP256-VRC26-I2的抗体轻链的一个示例性核酸序列。SEQIDNO:71是CAP256-VRC26-I2的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:72是CAP256-VRC26-I2的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:73是CAP256-VRC26.01的抗体重链的一个示例性核酸序列。SEQIDNO:74是CAP256-VRC26.01的抗体轻链的一个示例性核酸序列。SEQIDNO:75是CAP256-VRC26.01的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:76是CAP256-VRC26.01的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:77是CAP256-VRC26.02的抗体重链的一个示例性核酸序列。SEQIDNO:78是CAP256-VRC26.02的抗体轻链的一个示例性核酸序列。SEQIDNO:79是CAP256-VRC26.02的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:80是CAP256-VRC26.02的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:81是CAP256-VRC26.03的抗体重链的一个示例性核酸序列。SEQIDNO:82是CAP256-VRC26.03的抗体轻链的一个示例性核酸序列。SEQIDNO:83是CAP256-VRC26.03的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:84是CAP256-VRC26.03的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:85是CAP256-VRC26.04的抗体重链的一个示例性核酸序列。SEQIDNO:86是CAP256-VRC26.04的抗体轻链的一个示例性核酸序列。SEQIDNO:87是CAP256-VRC26.04的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:88是CAP256-VRC26.04的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:89是CAP256-VRC26.05的抗体重链的一个示例性核酸序列。SEQIDNO:90是CAP256-VRC26.05的抗体轻链的一个示例性核酸序列。SEQIDNO:91是CAP256-VRC26.05的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:92是CAP256-VRC26.05的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:93是CAP256-VRC26.06的抗体重链的一个示例性核酸序列。SEQIDNO:94是CAP256-VRC26.06的抗体轻链的一个示例性核酸序列。SEQIDNO:95是CAP256-VRC26.06的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:96是CAP256-VRC26.06的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:97是CAP256-VRC26.07的抗体重链的一个示例性核酸序列。SEQIDNO:98是CAP256-VRC26.07的抗体轻链的一个示例性核酸序列。SEQIDNO:99是CAP256-VRC26.07的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:100是CAP256-VRC26.07的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:101是CAP256-VRC26.08的抗体重链的一个示例性核酸序列。SEQIDNO:102是CAP256-VRC26.08的抗体轻链的一个示例性核酸序列。SEQIDNO:103是CAP256-VRC26.08的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:104是CAP256-VRC26.08的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:105是CAP256-VRC26.09的抗体重链的一个示例性核酸序列。SEQIDNO:106是CAP256-VRC26.09的抗体轻链的一个示例性核酸序列。SEQIDNO:107是CAP256-VRC26.09的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:108是CAP256-VRC26.09的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:109是CAP256-VRC26.10的抗体重链的一个示例性核酸序列。SEQIDNO:110是CAP256-VRC26.10的抗体轻链的一个示例性核酸序列。SEQIDNO:111是CAP256-VRC26.10的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:112是CAP256-VRC26.10的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:113是CAP256-VRC26.11的抗体重链的一个示例性核酸序列。SEQIDNO:114是CAP256-VRC26.11的抗体轻链的一个示例性核酸序列。SEQIDNO:115是CAP256-VRC26.11的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:116是CAP256-VRC26.11的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:117是CAP256-VRC26.12的抗体重链的一个示例性核酸序列。SEQIDNO:118是CAP256-VRC26.12的抗体轻链的一个示例性核酸序列。SEQIDNO:119是CAP256-VRC26.12的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:120是CAP256-VRC26.12的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:121是CAP256-VRC26抗体的抗体重链可变结构域的共有氨基酸序列。SEQIDNO:122是CAP256-VRC26抗体的抗体轻链可变结构域的共有氨基酸序列。SEQIDNO:123-167是用于制备用于454焦磷酸测序的扩增子的PCR引物。SEQIDNO:168是寡核苷酸引物EnvM的序列。SEQIDNO:169是寡核苷酸引物EnvAstop的序列。SEQIDNO:170是CAP256-VRC26.25的抗体重链的氨基酸序列。SEQIDNO:171是CAP256-VRC26.25的抗体轻链的氨基酸序列。SEQIDNO:172是CAP256-VRC26.25的抗体重链可变结构域的氨基酸序列。SEQIDNO:173是CAP256-VRC26.25的抗体轻链可变结构域的氨基酸序列。SEQIDNO:174是CAP256-VRC26.25的抗体重链的一个示例性核酸序列。SEQIDNO:175是CAP256-VRC26.25的抗体轻链的一个示例性核酸序列。SEQIDNO:176是CAP256-VRC26.25的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:177是CAP256-VRC26.25的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:178是CAP256-VRC26.26的抗体重链的氨基酸序列。SEQIDNO:179是CAP256-VRC26.26的抗体轻链的氨基酸序列。SEQIDNO:180是CAP256-VRC26.26的抗体重链可变结构域的氨基酸序列。SEQIDNO:181是CAP256-VRC26.26的抗体轻链可变结构域的氨基酸序列。SEQIDNO:182是CAP256-VRC26.26的抗体重链的一个示例性核酸序列。SEQIDNO:183是CAP256-VRC26.26的抗体轻链的一个示例性核酸序列。SEQIDNO:184是CAP256-VRC26.26的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:185是CAP256-VRC26.26的抗体轻链可变结构域的一个示例性核酸序列。SEQIDNO:186是CAP256-VRC26.27的抗体重链的氨基酸序列。SEQIDNO:187是CAP256-VRC26.27的抗体轻链的氨基酸序列。SEQIDNO:188是CAP256-VRC26.27的抗体重链可变结构域的氨基酸序列。SEQIDNO:189是CAP256-VRC26.27的抗体轻链可变结构域的氨基酸序列。SEQIDNO:190是CAP256-VRC26.27的抗体重链的一个示例性核酸序列。SEQIDNO:191是CAP256-VRC26.27的抗体轻链的一个示例性核酸序列。SEQIDNO:192是CAP256-VRC26.27的抗体重链可变结构域的一个示例性核酸序列。SEQIDNO:193是CAP256-VRC26.27的抗体轻链可变结构域的一个示例性核酸序列。具体实施方式现在将在下文中参考附图更完整地描述本发明,在所述附图中显示了本发明的一些、但是并非所有实施方案。描述的发明应当不限于公开的具体实施方案,并且在本发明范围内意图包括改进和其它实施方案。尽管本文采用了特定的术语,但是它们仅以一般性的和描述性的含义使用,而不用于限制的目的。在提供范围的值的情况下,应当理解,介于该范围的上限与下限之间的每个居间值(直至下限单位的十分之一,除非上下文另外清楚地指明),并且在该所述范围中的任意其它所述值或居间值被包括在本发明内。这些较小范围的上限和下限(其可以独立地包括在较下范围内)也被包括在本发明内,具有在所述范围中的任何具体地排除的限制。在所述范围包括一个或两个限值的情况下,排除那些被包括的端值中的任一个或两个端值的范围也被包括在本发明中。如贯穿本说明书和在下面的权利要求中使用的,单数形式“一个”、“一种”和“所述”包括复数形式,除非上下文另外清楚地指出。在本文中使用的术语和措辞用于描述的目的,且不应当视作限制。在本文中使用的术语“包含”、“含有”、“具有”和“包括”及其变体的应用意图包括在此后列出的项目和它们的等同物以及另外的项目。除非在本文中另外定义,否则本文使用的所有技术和科学术语具有本发明所属领域的普通技术人员所通常理解的相同含义。本发明提供了分离的抗-HIV抗体,其广谱中和HIV-1且其识别和结合HIV-1Env的V1V2表位。术语“抗体”包括单克隆抗体、多克隆抗体、多特异性抗体(例如,双特异性抗体和多反应性抗体)和抗体片段。因此,在本说明书中使用的术语“抗体”包括、但不限于任何特异性结合成员、免疫球蛋白种类和/或同种型(例如:IgG1、IgG2、IgG3、IgG4、IgM、IgA、IgD、IgE和IgM)或其抗体片段。在本领域中理解,抗体是包含通过二硫键互联的至少两个重(H)链和两个轻(L)链的糖蛋白或其抗原结合部分。重链包含重链可变区(VH)和重链恒定区(CH1、CH2和CH3)。轻链包含轻链可变区(VL)和轻链恒定区(CL)。重链和轻链的可变区包含框架区(FR)和互补性决定区(CDR)。四个FR是相对保守的,而CDR区域(CDR1、CDR2和CDR3)包含高变区。FR和CDR从NH2端至COOH端如下排列:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。重链和轻链的可变区含有与抗原相互作用的结合结构域。此外,所述恒定区可以介导免疫球蛋白与宿主组织或因子的结合。在“抗体”的定义中还包括嵌合抗体、人源化抗体、重组抗体、从转基因的非人动物产生的人抗体和使用本领域技术人员可得到的富集技术选自文库的抗体。本文中使用的术语“表位”是指抗体的互补位可以与其结合的、在抗原上的任何抗原决定簇。表位决定簇通常由分子(诸如氨基酸或糖侧链)的化学活性的表面组群(grouping)组成并且通常具有特定三维结构特征以及特定电荷特征。“抗体片段”包含完整抗体的一部分,诸如完整抗体的抗原结合区或可变区。抗体片段的例子包括,但不限于Fab、Fab′、F(ab′)2、Fv片段、scFV片段;双抗体;或线性抗体。抗体的木瓜蛋白酶消化会产生两个相同的“Fab”片段或抗原结合片段(每个具有单个抗原结合位点)和一个残余的“Fc”片段(其名称反映了它的容易结晶的能力)。抗体的胃蛋白酶处理会产生一个F(ab′)2片段,其具有两个抗原结合位点且其保留它的交联抗原的能力。术语“Fv”表示含有完全抗原识别和抗原结合位点的最小抗体片段。该片段含有紧密地非共价结合的一个重链可变区结构域和一个轻链可变区结构域的二聚体。这两个结构域的折叠导致六个高变环(三个环各自来自H和L链)的形成,它们促进用于抗原结合的氨基酸残基并给所述抗体赋予抗原结合特异性。但是,即使单个可变区(或仅包含三个对抗原特异性的CDR的Fv的一半)具有识别和结合抗原的能力,尽管以较低亲和力。“单链Fv”(“sFv”或“scFv”)是包含连接成单个多肽链的VH和VL抗体结构域的抗体片段。sFv多肽还可以包含在VH和VL结构域之间的多肽接头,其使sFv能够形成期望的结构用于抗原结合。“Fab”片段含有轻链的恒定结构域和重链的第一恒定结构域(CH1)。Fab′片段与Fab片段的差别在于几个残基在重链CH1结构域的羧基端处的添加,包括来自抗体铰链区的一个或多个半胱氨酸。Fab′-SH在本文中关于Fab′而命名,其中恒定结构域的半胱氨酸残基带有游离巯基。F(ab′)2抗体片段最初作为Fab′片段对而产生,所述Fab′片段对具有在它们之间的铰链半胱氨酸。抗体片段的其它化学偶联也是本领域已知的。变体抗体也都被包括在本发明的范围内。因而,在本申请中列举的序列的变体也都被包括在本发明的范围内。具有提高的亲和力的抗体序列的其它变体可以使用本领域已知的方法得到,并都被包括在本发明的范围内。本领域技术人员利用用于生产变体多肽的重组方法和/或合成化学技术可以修改多肽的氨基酸序列。例如,可以使用氨基酸置换得到具有进一步提高的亲和力的抗体。备选地,可以使用核苷酸序列的密码子优化来提高在用于生产抗体的表达系统中的翻译效率。这样的变体抗体序列与在本申请中列举的序列具有70%或更高的(即,75%、80%、85%、90%、95%、96%、97%、98%、99%或更大)序列同一性。相对于在本申请中列举的序列的全长,计算这样的序列同一性。本发明提供了单独的或与其它抗体组合的抗-HIV抗体,其在血清中具有宽中和活性且作为分离的抗体。术语“多肽”应当解读为包括“肽”和“蛋白”,反之亦然。本文中使用的“多肽”表示具有以下物质的氨基酸序列的重组或非重组多肽的氨基酸序列:i)天然多肽,ii)多肽的生物活性片段,或iii)多肽的生物活性变体。本文中使用的术语“分离的”是指已经从它的天然环境取出的核酸或抗体。已经“分离的”核酸、肽和蛋白因而包括通过标准纯化方法纯化的核酸和蛋白。该术语也包括通过在宿主细胞中的重组表达而制备的核酸、肽和蛋白以及化学合成的核酸和/或多肽。术语“受试者”和“患者”在本文中互换地用于指可能需要本文描述的药物组合物、治疗和疫苗的任何动物。受试者和患者因而包括、但不限于,灵长类动物(包括人类)、犬科动物、猫科动物、鼠和其它哺乳动物受试者。优选地,所述受试者是人类。从在其中使用所述术语的上下文显而易见,受试者和患者表示易于被人免疫缺陷病毒(HIV)感染的受试者或患者和/或被HIV感染的受试者或患者。在本发明的另一个实施方案中,所述交叉反应性的、广谱中和性的抗-HIV抗体包含重链可变区,所述重链可变区包含SEQIDNO:121的共有氨基酸序列。在另一个实施方案中,所述交叉反应性的、广谱中和性的抗-HIV抗体包含轻链可变区,所述轻链可变区包含SEQIDNO:122的共有氨基酸序列。本发明的另一个实施方案提供了一种分离的抗-HIV抗体,其包含SEQIDNO:1、5、9、13、17、21、25、29、33、37、41、45、49、53、57、170、178或186的重链序列和SEQIDNO:2、6、10、14、18、22、26、30、34、38、42、46、50、54、58、171、179或187的轻链序列;和一种分离的抗-HIV抗体,其包含SEQIDNO:3、7、11、15、19、23、27、31、35、39、43、47、51、55、59、172、180或188的重链可变区序列和SEQIDNO:4、8、12、16、20、24、28、32、36、40、44、48、52、56、60、173、181或189的轻链可变序列,或与其具有至少70%、或至少75%、或至少80%、或至少85%、或至少90%、或至少95%、或至少96%、或至少97%、或至少98%、或至少99%同一性的序列。在另一个实施方案中,本发明提供了一种分离的抗-HIV抗体,其包含SEQIDNO:3、7、11、15、19、23、27、31、35、39、43、47、51、55、59、172、180或188的重链序列和SEQIDNO:4、8、12、16、20、24、28、32、36、40、44、48、52、56、60、173、181或189的轻链序列中的一个或两者,且其中所述抗体以表3.1、3.2或3.3中所示的IC50浓度中和HIV病毒毒株的集合。在另一个实施方案中,本发明提供了一种分离的抗-HIV抗体,其选自CAP256-VRC261-UCA、CAP256-VRC26-I1、CAP256-VRC26-I2、CAP256-VRC26.01、CAP256-VRC26.02、CAP256-VRC26.03、CAP256-VRC26.04、CAP256-VRC26.05、CAP256-VRC26.06、CAP256-VRC26.07、CAP256-VRC26.08、CAP256-VRC26.09、CAP256-VRC26.10、CAP256-VRC26.11、CAP256-VRC26.12、CAP256-VRC26.13、CAP256-VRC26.14、CAP256-VRC26.15、CAP256-VRC26.16、CAP256-VRC26.17、CAP256-VRC26.18、CAP256-VRC26.19、CAP256-VRC26.20、CAP256-VRC26.21、CAP256-VRC26.22、CAP256-VRC26.23、CAP256-VRC26.24、CAP256-VRC26.25、CAP256-VRC26.26、CAP256-VRC26.27、CAP256-VRC26.28、CAP256-VRC26.29、CAP256-VRC26.30、CAP256-VRC26.31、CAP256-VRC26.32和CAP256-VRC26.33。在另一个实施方案中,本发明提供了一种分离的抗-HIV抗体,其包含含有选自CAP256-VRC261-UCA、CAP256-VRC26-I1、CAP256-VRC26-I2、CAP256-VRC26.01、CAP256-VRC26.02、CAP256-VRC26.03、CAP256-VRC26.04、CAP256-VRC26.05、CAP256-VRC26.06、CAP256-VRC26.07、CAP256-VRC26.08、CAP256-VRC26.09、CAP256-VRC26.10、CAP256-VRC26.11、CAP256-VRC26.12、CAP256-VRC26.25、CAP256-VRC26.26和CAP256-VRC26.27的HIV抗体的对应区域的氨基酸序列的重链CDR1、CDR2和CDR3区域和轻链CDR1、CDR2和CDR3区域。本发明也提供了制备分离的包含SEQIDNO:1、5、9、13、17、21、25、29、33、37、41、45、49、53、57、170、178或186的重链序列和SEQIDNO:2、6、10、14、18、22、26、30、34、38、42、46、50、54、58、171、179或187的轻链序列的抗-HIV抗体的方法,和生产分离的抗-HIV抗体的方法,所述分离的抗-HIV抗体包含SEQIDNO:3、7、11、15、19、23、27、31、35、39、43、47、51、55、59、172、180或188的重链可变区序列和SEQIDNO:4、8、12、16、20、24、28、32、36、40、44、48、52、56、60、173、181或189的轻链可变序列,或与它们具有至少70%、或至少75%、或至少80%、或至少85%、或至少90%、或至少95%、或至少96%、或至少97%、或至少98%、或至少99%同一性的序列。在另一个实施方案中,本发明提供了编码分离的抗-HIV抗体的分离的核酸、含有所述核酸的载体和宿主细胞、以及用于生产所述抗体的重组技术。本发明还提供了编码抗-HIV抗体CAP256-VRC261-UCA、CAP256-VRC26-I1、CAP256-VRC26-I2、CAP256-VRC26.01、CAP256-VRC26.02、CAP256-VRC26.03、CAP256-VRC26.04、CAP256-VRC26.05、CAP256-VRC26.06、CAP256-VRC26.07、CAP256-VRC26.08、CAP256-VRC26.09、CAP256-VRC26.10、CAP256-VRC26.11、CAP256-VRC26.12、CAP256-VRC26.25、CAP256-VRC26.26和CAP256-VRC26.27的重链和轻链的肽序列的多核苷酸变体。这些多核苷酸变体与本发明的多核苷酸序列相比可以具有至少70%、或至少75%、或至少80%、或至少85%、或至少90%、或至少95%、或至少96%、或至少97%、或至少98%、或至少99%或更大的序列同一性,如使用本文所述的方法所确定的。这样的连续序列可以编码CDR序列,或可以编码完整可变区。如本领域已知的,可变区序列可以与任何适当的恒定区序列融合。本领域技术人员会认识到,通过考虑密码子简并性、氨基酸相似性、读码框定位等,可以适当地调节这些值以确定由两个核苷酸序列编码的蛋白的对应同一性。术语“核酸”和“多核苷酸”在本文中互换地用于表示单链或双链RNA、DNA或混合的聚合物。对于抗体的重组生产,将编码它的核酸插入载体中用于进一步克隆(DNA的扩增)或用于表达。根据在实施例中阐述的方法,分离编码本发明的抗体的DNA。本领域技术人员会明白,许多载体可供用在抗体的重组生产中。载体组分通常包括、但不限于以下的一种或多种:信号序列、复制起点、一个或多个标记基因、增强子元件、启动子和转录终止序列。还可以重组地生产本发明的抗-HIV抗体,例如,作为具有异源或同源多肽的融合多肽,其包括在成熟蛋白或多肽的N-端处具有特定切割位点的信号序列或其它多肽、免疫球蛋白恒定区序列等。选择的异源信号序列优选地可以是被宿主细胞识别和加工(即,被信号肽酶切割)的那种。对于不会识别和加工天然抗体信号序列的原核宿主细胞,选择被原核信号序列置换的信号序列。当核酸放置成与另一个核酸序列在功能上关联时,所述核酸“可操作地连接”。例如,前序列或分泌型前导序列的DNA与多肽的DNA可操作地连接,条件是其表达为参与所述多肽的分泌的前蛋白;启动子或增强子与编码序列可操作地连接,条件是其影响所述序列的转录;或核糖体结合位点与编码序列可操作地连接,条件是其被定位为促进翻译。通常,“可操作地连接”是指,连接的DNA序列是连续的,并且,在分泌型前导序列的情况下,是连续的和在读码框中。然而,增强子不必须是连续的。连接通过在方便的限制性位点处的连接来实现。如果所述位点不存在,则合成的寡核苷酸适体或接头根据常规实践使用。本文中使用的表述“细胞”、“细胞系”和“细胞培养物”可互换使用,且所有这些名称都包括子代。因而,词语“转化体”和“转化的细胞”包括原代受试细胞和从其衍生出的培养物,不论转移的数目。还理解所有的后代的DNA含量可能不精确一致,这归因于有意或无意的突变。包括具有相同功能或生物活性的突变体后代,如在最初转化的细胞中所筛选的。在意指不同名称时,它从上下文是清楚的。用于克隆或表达DNA的合适宿主细胞是原核细胞、酵母细胞或高等真核细胞。将用于抗-HIV抗体生产的、用上述表达或克隆载体转化的宿主细胞在适当修饰的常规营养物培养基中培养,用于诱导启动子、选择转化体或扩增编码期望序列的基因。使用本领域普通技术人员已知的纯化技术,可以纯化从所述细胞制备的抗体组合物。优选地,将本发明的核苷酸序列和/或抗体施用至受试者体内,以便在受试者中产生免疫原性应答或提供预防作用。在某些实施方案中,可能期望在实验动物中表达本发明的核酸或施用本发明的抗体,诸如用于本发明的HIV-1免疫原性或预防性组合物的临床前试验。在其它实施方案中,将需要在人受试者中表达或施用本发明的抗体,诸如在临床试验中和用于本发明的免疫原性或预防性组合物的实际临床应用。在优选的实施方案中,所述受试者是人,例如被HIV-1感染或处于被HIV-1感染的风险中的人。根据另一个实施方案,本发明提供了一种治疗被病毒(例如,HIV)感染的哺乳动物的方法,所述方法包括给所述哺乳动物施用药物组合物,其包含本文中公开的抗-HIV抗体或表达本文中公开的抗-HIV抗体的核苷酸序列。根据一个实施方案,治疗被HIV感染的哺乳动物的方法包括,给所述哺乳动物施用包含本发明的抗体或其片段的药物组合物。本发明的组合物可以包括超过一种具有公开的特征的抗体(例如,多种抗体或抗体集合)。它还可以包括本领域已知的其它HIV中和抗体。根据另一个实施方案,治疗被HIV感染的哺乳动物的方法包括给所述哺乳动物施用药物组合物,其包含表达本发明的抗-HIV抗体或其片段的核苷酸序列、含有所述核酸的载体和含有所述载体的细胞。在体内应用中,本发明的抗体或核酸优选地作为药物组合物的组分来施用,所述药物组合物包含与药用载体混合的所述核苷酸序列、含有所述核酸的载体、含有所述载体的细胞和/或抗体。当用于体内疗法时,所述核苷酸序列、含有所述核酸的载体、含有所述载体的细胞和/或本发明的抗体以治疗有效量施用给患者。根据本领域已知的方法,例如,但不限于静脉内施用,将所述核苷酸序列、含有所述核酸的载体、含有所述载体和/或抗体的细胞施用给人患者。术语“药物组合物”在本文中用于限定具有适合用于施用给患者的形式、浓度和纯度水平的固体或液体组合物,在所述施用后,它可以引起生理学变化,所述变化治疗和/或改善HIV感染的影响。所述药物组合物还可以用作针对HIV-1的预防或治疗疫苗的一种或多种组分,和用于预防、改善或治疗AIDS。本发明的核酸和载体就提供基因疫苗(即这样的疫苗:其用于将编码本发明的抗体的核酸递送给受试者诸如人,使得所述抗体在受试者中被表达,以便引起免疫应答和/或提供针对HIV-1感染的直接保护)而言是特别有用的。备选地,可以将本发明的抗体施用给受试者,以便引起针对HIV-1的免疫应答和/或提供针对HIV-1感染的直接保护。通过任意合适的方式,包括胃肠外、皮下、腹膜内、肺内和鼻内施用,可以将本发明的抗体、核苷酸序列、含有所述核酸的载体和/或含有所述载体的细胞施用给受试者。所述抗体或其抗原结合部分可以单独施用或与另一种治疗剂(例如第二人抗体或其抗原结合部分)联合施用。在一个实施例中,所述第二种抗体或其抗原结合部分特异性地结合第二HIV表位,所述第二HIV表位不同于所述第一种抗体所结合的表位。在另一个实施例中,所述抗体与另一种药剂(例如,抗病毒剂)一起施用。抗病毒剂包括、但不限于,例如:阿巴卡韦、阿昔洛韦、阿德福韦、金刚烷胺、氨普那韦、Ampligen、阿拉韦罗、阿比朵尔、阿扎那韦、Atripla、波普瑞韦、西多福韦、双汰芝、达芦那韦、地拉韦啶、去羟肌苷、二十二醇、依度尿苷、依法韦仑、恩曲他滨、恩夫韦肽、恩替卡韦、进入抑制剂、泛昔洛韦、福米韦生、呋山那韦、膦甲酸、膦乙酸、融合抑制剂、更昔洛韦、伊巴他滨、Imunovir、碘苷、咪喹莫特、茚地那韦、肌苷、整合酶抑制剂、干扰素I、II或III、拉米夫定、洛匹那韦、洛韦胺、马拉韦罗、吗啉胍、美替沙腙、奈非那韦、奈韦拉平、Nexavir、核苷类似物、奥司他韦(达菲)、聚乙二醇干扰素α-2a、喷昔洛韦、培拉米韦、普来可那立、鬼臼毒素、蛋白酶抑制剂、拉替拉韦、逆转录酶抑制剂、利巴韦林、金刚乙胺、利托那韦、Pyramidine、沙奎那韦、司他夫定、替诺福韦、替诺福韦酯、替拉那韦、曲氟尿苷、三协唯、曲金刚胺、Truvada、伐昔洛韦(Valtrex)、伐昔洛韦、维立韦罗、阿糖腺苷、盐酸他立韦林、维立韦罗、扎西他滨、扎那米韦(Relenza)或齐多夫定或它们的任意组合。对于疾病的预防或治疗,抗体、核苷酸序列、含有所述核酸的载体和/或含有所述载体的细胞的适当剂量将取决于要治疗的疾病的类型,疾病的严重程度和进程,所述抗体、核苷酸序列、含有所述核酸的载体和/或含有所述载体的细胞是否为了预防目的而施用,以前的治疗,患者的临床史和对抗体的应答,以及主治医师的判断。将所述抗体、核苷酸序列、含有所述核酸的载体和/或含有所述载体的细胞一次或在一系列治疗中适当地施用给患者。术语“有效剂量”或“有效的剂量”被定义为足以实现或至少部分地实现期望的作用的量。术语“治疗有效剂量”被定义为足以治愈或至少部分地阻止疾病和它的征状和在遭受所述疾病的患者中的影响的量。使用的有效剂量取决于正在治疗的障碍的严重程度和受试者的免疫系统的一般状态。通过以低压冻干制剂或水溶液的形式将具有期望程度的纯度的抗体与任选的生理上可接受的载体、赋形剂或稳定剂混合,制备用于贮存的包含本发明的一种或多种抗体的药物组合物。以与良好医学实践一致的方式配制、定量施用和施用抗体组合物。在该背景下考虑的因素包括正在治疗的特定障碍、正在治疗的特定哺乳动物、个体患者的临床状况、障碍的起因、药剂的递送位点、施用方法、施用调度和医学从业人员已知的其它因素。要施用的抗体的“治疗有效量”由这样的考虑因素控制,且是降低被感染的个体中的病毒滴度所必需的最小量。通过以冻干制剂或水溶液的形式将具有期望程度的纯度的核酸、载体或细胞与任选的生理上可接受的载体、赋形剂或稳定剂混合,制备用于贮存的药物组合物,其包含一种或多种编码本发明的抗体的核酸、含有所述核酸的载体和/或含有所述载体的细胞。以与良好医学实践一致的方式配制、定量施用和施用所述组合物。在该背景下考虑的因素包括正在治疗的特定障碍、正在治疗的特定哺乳动物、个体患者的临床状况、障碍的起因、药剂的递送位点、施用方法、施用调度和医学从业人员已知的其它因素。要施用的核酸、载体或细胞的“治疗有效量”由这样的考虑因素控制,且是降低被感染的个体中的病毒滴度所必需的最小量。在本发明的另一个实施方案中,本发明的抗体可以用在诊断组合物中。诊断组合物是含有化合物或抗体(例如,标记的化合物或抗体)的组合物,所述化合物或抗体用于检测结合所述化合物的抗体或结合所述抗体的免疫原、抗原或表位在样品(诸如生物样品)中的存在;例如,抗-HIV抗体或HIV免疫原、抗原或表位。在本发明的另一个实施方案中,提供了制成品(诸如试剂盒),其含有可用于治疗上述障碍的物质。所述制成品包含容器和标签。合适的容器包括,例如,瓶子、管形瓶、注射器和试管。容器可以由多种材料制成,例如玻璃或塑料。所述容器容纳可有效地治疗所述病症的组合物,且可以具有无菌进入口(例如,容器可以是静脉内溶液袋或具有可被皮下注射针头刺穿的塞子的瓶子)。所述组合物中的活性剂是在如上所述的本发明的制剂中的一种或多种抗体。在容器上或伴随容器的标签指示,所述组合物用于治疗选择的病症。所述制成品还可以包含第二个容器,其包含药用缓冲液,诸如磷酸盐缓冲盐水、林格氏溶液和右旋糖溶液。它还可以包括从商业和用户观点看合乎需要的其它物质,包括其它缓冲液、稀释剂、过滤器、针头、注射器和具有使用说明书的包装说明书。研究受试者将CAPRISA参与者CAP256招募进2004年在南非KwaZulu-Natal建立的CAPRISA急性感染研究(vanLoggerenberg等人.2008)用于随访和随后的HIV血清转化鉴别。CAP256是在该组群中发展出中和宽度的7位妇女之一(Gray等人.2011)。CAPRISA002急性感染研究经过UniversityofKwaZulu-Natal(E013/04)、UniversityofCapeTown(025/2004)和UniversityoftheWitwatersrand(MM040202)的研究伦理委员会评审和批准。CAP256提供了关于研究参与的书面知情同意书。提供下述实施例作为举例说明且不作为限制。实施例1CAP256-VRC26家族基因的分离和表达将从第59、119和206周的CAP256血液抽吸物分离的PBMC染色,并在FACSAriaII上针对IgG+B细胞进行分选(Georgiev等人.2013)。将细胞以2个B细胞/孔铺板在384孔板中,并在有表达IL-2、IL-21和CD40L的辐照饲养细胞存在下培养14天(Huang等人(2012),(2013)。针对HIV-1ZM53.12和CAP45.G3Env-假病毒通过如在(Doria-Rose等人,2013.ProtocolExchange,印刷中)中所述的微量中和来筛选培养物上清液。筛选的孔的数目在表1中。将κ和λ轻链基因和IgG重链基因可变区从中和阳性孔扩增,亚克隆,表达,并纯化(Georgiev等人.2013)。将重链重构为IgG1。从第59周回收一种抗体,从第119周回收8种抗体,从第206周回收3种抗体。表1:自供体CAP256的B细胞克隆的总结PBMC的周59119206铺板的总B细胞(2/孔)150004500042000IgG阳性孔的百分比8.3%48%29%中和阳性孔的数目44934回收的抗体的数目183实施例2中和测定制备单轮的复制Env-假病毒,滴定,并用于感染TZM-b1靶细胞(Shu等人.2007,Montefiori等人.2009)。使用一组195种地理上和遗传上不同的Env-假病毒(其代表主要亚型和循环重组形式)确定CAP256-VRC26.08和.25的中和宽度(Georgiev等人.2013,Wu等人.2010)。将数据计算为相对于对照孔的发光单元的减小,并且对于单克隆抗体报告为以微克/微升计的50%抑制浓度(IC50),或对于血浆样品报告为倒数稀释度(ID50)。中和关联对一组29种HIV-1毒株(6535.3、AC10.29、CAAN.A2、CAP210.E8、CAP244.D3、CAP45.G3、DU156.12、DU172.17、DU422.01、PVO.04、Q168.a2、Q23.17、Q259.d2.17、Q461.e2、Q769.d22、Q842.d12、QH0692.42、REJO.67、RHPA.7、SC422.8、THRO.18、TRJO.58、TRO.11、WITO.33、ZM109.4、ZM135.10a、ZM197.7、ZM233.6、ZM53.12)计算CAP256-VRC26抗体的中和指纹和四个纵向血清时间点(在感染后59、106、159和220周)的中和模式之间的关联(Georgiev等人.2013)。对一组41种HIV-1毒株(6535.3、0260.v5.c36、6405.v4.c34、AC10.29、C1080.c3、CAAN.A2、CAP210.E8、CAP244.D3、CAP45.G3、CNE3、DU156.12、DU172.17、DU422.01、KER2008.12、KER2018.11、MB201.A1、MB539.2B7、PVO.04、Q168.a2、Q23.17、Q259.17、Q461.e2、Q769.d22、Q842.d12、QH0692.42、REJO.67、RHPA.7、RW020.2、SC422.8、TH976.17、THRO.18、TRJO.58、TRO.11、UG037.8、WITO.33、ZM109.4、ZM135.10a、ZM197.7、ZM214.15、ZM249.1、ZM53.12)计算CAP256-VRC26抗体的中和能力和靶向四个主要易损性(vulnerability)位点的抗体的参比集合(每种独特供体至多两种抗体)之间的关联。对一组28种HIV-1毒株(6535.3、AC10.29、CAAN.A2、CAP210.E8、CAP244.D3、CAP45.G3、DU156.12、DU172.17、DU422.01、PVO.04、Q168.a2、Q23.17、Q259.17、Q461.e2、Q769.d22、Q842.d12、QH0692.42、REJO.67、RHPA.7、SC422.8、THRO.18、TRJO.58、TRO.11、WITO.33、ZM109.4、ZM135.10a、ZM197.7、ZM53.12)计算四个纵向血清时间点的中和模式和参比抗体的中和指纹之间的关联。对于参比抗体,将来自多个中和实验的数据取平均值,并固定。所有关联是基于Spearman秩系数。病毒样颗粒ELISA如在Tong等人(2012)中所述执行VLPELISA。简而言之,用基于pCAGGS的表达Env的质粒和Env-缺陷型HIV-1基因组主链质粒pNL-LucR-E-,通过基于PEI的293T细胞共转染来生产VLP。以在转染上清液中的20倍浓度,将VLP包被在ELISA孔上。然后通过ELISA来评估MAb结合,省去PBS洗涤缓冲液中的去污剂,并用抗-人Fc碱性磷酸酶缀合物(Accurate,Westbury,NY)和SigmaFAST磷酸对硝基苯酯片(Sigma)探测。在405nm对平板进行读数。细胞表面Env结合用缺失细胞质尾巴的EnvZM53.12瞬时转染293T细胞(Pancera和Wyatt(2005))。2天后,将细胞用ViVid生存力染料(Invitrogen)染色,随后用与连续稀释的未标记的竞争抗体预混合的生物素化的CAP256-VRC26.01(10ug/ml)或CAP256-VRC26.08(0.8μg/ml)染色。温育和2次洗涤后,将细胞用1∶200稀释度的链霉抗生物素-PE(Invitrogen)染色。在BDLSRII(BectonDickinson)上分析细胞。将结合测量为每个样品的中值荧光强度(MFI)减去仅用链霉抗生物素-PE染色的细胞的MFI。实施例3抗体的多反应性分析如在Haynes等人(2005)中所述,确定抗体与心磷脂的结合。简而言之,按照生产商的方案使用QUANTALiteACAIgGIIIELISA试剂盒(ZeusScientific),将每种抗体在试剂盒样品稀释剂中稀释至100μg/ml,并在3倍连续稀释物中试验。显示的结果是至少2个独立ELISA的代表。在每个板上包括阳性和阴性对照,且在背景以上3倍的值认为是阳性的。按照生产商的方案用ANA/HEp-2CellCultureIFATestSystem(ZeusScientific)确定针对人上皮细胞系(HEp-2)的抗体反应性。将抗体在ZOBRA-NS稀释剂中稀释至50μg/ml和25μg/ml。在每个载玻片上包括阳性和阴性对照。在每个稀释度将抗体评分为阴性的、不确定的或阳性的(1+至4+)。结果是至少2个独立实验的代表。实施例4下一代测序根据Wu等人(2011)和Zhu等人(2012)的方法(有如所指示的轻微修改),制备用于下一代测序的扩增子。简而言之,使用Oligotex试剂盒(Qiagen)从1000-1500万个PBMC制备mRNA。使用SuperscriptII逆转录酶(Invitrogen)和oligo-dT(12-18)引物合成cDNA。用Phusion聚合酶执行各个PCR反应达30个循环。引物(表SNDRx)由对所有λ基因家族或VH3家族基因具有特异性的5-7个寡核苷酸的集合组成,且具有用于454下一代测序的衔接子。仅对于第176周,用针对所有VH家族的引物执行重链PCR,并将混合的λ和κ引物用于轻链(表2)。将PCR产物凝胶纯化(Qiagen)。在GSFLX测序仪(Roche-454LifeSciences,Bradford,CT,USA)上在每个反应的半芯片上(第176周为完整芯片)进行PCR产物的焦磷酸测序。平均而言,生产了约250,000个原始读数。表2:用于制备用于454焦磷酸测序的扩增子的PCR引物(A)用于除第176周以外所有时间点的引物,(B)仅用于第176周样品的引物。(A)(B)Antibodyomics流水线(Antibodyomicspipeline)使用在Python中执行的流水线,类似于我们以前报告的,处理原始454数据(Zhu等人.2013)。简而言之,针对长度来过滤读数,仅保持在300-600个核苷酸之间的那些。然后使用具有经验优化的参数的BLAST将种系V基因分配至各读数。抛弃没有在e-值=10-10的情况下发现其有V基因匹配的读数。对于分配至任何VH3-30或VL1-51等位基因(CAP256-VRC26种系基因)的读数,使用ClustalW2(Larkin等人.2007)计算与所述种系和各分离的抗体的序列同一性。使用在R的ggplot2将这些数据绘制为密度热图以产生同一性-趋异性图(未显示)。发现克隆相关序列将分配至与CAP256-VRC26、VH3-30和VL1-51相同的V基因的读数送至IMGTHigh-Vquest(Alamyar(2012))(http://www.imgt.org/IMGTindex/IMGTHighV-QUEST.html),并使用结果(包括自动化的序列校正)进一步筛选谱系相关序列。选择这样的读数用于进一步处理:其被分配至匹配CAP256-VRC26的J基因(JH3*02或JL1*01),且具有在V和J基因中的类似趋异性(±15%)、在V基因中的类似(±10%)核苷酸和氨基酸趋异性,且含有在整个可变区中的连续开放阅读框。接着,将来自所有时间点的读数合并,并使用CDHit(Li等人.2001)以97.25%序列同一性簇集(预期的454测序误差的标准差的2倍)(Zhu等人.2012)。对于每个簇,从最早可能的时间点选择代表性序列。来自它们所出现的最早时间点的簇代表的选择对于维持后续分析中的谱系发展的时间表的信息而言是关键性的。该操作产生8,485个独特重链序列和6,410个独特轻链序列。为了鉴别CAP256-VRC26谱系成员重链,我们使用12种分离的CAP256-VRC26抗体的重链序列对独特的454序列集合执行了供体内种系发生分析(Zhu等人.2012)。将707个序列鉴别为可能的谱系成员,其中27个在手工检查以后被抛弃,从而产生共计680个独特的CAP256-VRC26谱系重链序列。为了鉴别轻链谱系成员,要求在CDRL3中与分离的抗体之一具有至少92%序列同一性的筛选产生495个序列。使用Joinsolver(Souto-Carneiro(2004))详细地检查这些序列的V-J连接部,以确保关于分离的抗体已知的重组点匹配的那些。这会产生共计472个独特的CAP256-VRC26谱系轻链序列。系统发生树的计算使用MEGA5(Tamura等人.2011)选择GTR+G作为最佳数学模型用于从CAP256-VRC26谱系序列建造最大似然树。然后使用FASTML(Ashkenazy等人.2012)估计γ参数并建造重链和轻链序列(包括分离的抗体)的单独最大似然树,并将这些固定在种系V基因序列上。基于来自仅用12种分离的抗体构建的树的证据,手工移动轻链树的两个分支以匹配它们在重链树中的定位。使用DNAML和PHYLIP(PhylogenyInferencePackage)3.6版(Felsenstein,J.2005.PHYLIP(PhylogenyInferencePackage)3.6版.作者发布.DepartmentofGenomeSciences,UniversityofWashington,Seattle)的分析表明,这些重排没有显著地改变所述树的log-似然评分。为了建立重链系统发生树的集中形式,使用95%序列同一性阈值使CDRH3序列聚簇,并要求簇中的所有CDRH3具有相同的长度。分离的抗体和具有至少五个成员的单源簇由单个叶代表,而所有其它序列从树取下。在缺失内部结的情况下,将该结上面和下面的分支长度累加,使得所有剩余序列的树深度得到维持。UCA和推断的中间体使用DNAML最大似然软件包(http://cmgm.stanford.edu/phylip/dnaml.html)从所有重链和所有轻链谱系成员的系统发生树推断祖先序列(图3和图6和7),包括分离的抗体。计算的重链UCA与种系VH3-30*18等位基因相同。尽管VH3-30*03等位基因是仅一个核苷酸不同的,该供体的种系测序表明她携带*18等位基因且不携带*03等位基因(CatherineMitchell,个人通信)。为了试验CAP256-VRC26.01的发展中的中间体,从系统发生树选择两个内部结成为在进化距离方面大约等间距,并使用DNAML检索推断的序列。每个中间体的推断的重链和轻链的成功互补提示,所述谱系被454个数据充分取样,并且计算的系统发生树成功地捕获重链和轻链的偶联进化动力学。用Weblogo(Crooks等人.2004)产生了CDRH3的鉴别图(Logogram)。实施例5结晶通过在37℃用Lys-C将纯化的IgG消化2小时,制备VRC26.UCAFab。然后通过添加完全蛋白酶抑制剂淬灭反应。对于VRC26.01、VRC26.03、VRC26.04、VRC26.06、VRC26.07和VRC26.10Fab制备,在Lys235后插入HRV3C识别位点(GLEVLFQGP),并将纯化的IgG与HRV3C蛋白酶一起在4℃温育过夜。对于全部,使消化的抗体穿过蛋白A琼脂糖以除去Fc片段。将Fab在Superdex200凝胶过滤柱上进一步纯化,并将浓缩的等分试样在-80℃储存。使用CartesianHoneybee结晶机器人针对576种结晶条件筛选所有Fab。通过将0.2μl蛋白复合物与0.2μl蓄池溶液混合,在20℃在坐滴中通过蒸汽扩散方法使最初的晶体生长。通过将1.0μl蛋白复合物与1.0μl蓄池溶液混合,在悬滴中手工再现晶体。用27%PEG8000和0.1MHepes(pH7.5)的蓄池溶液使VRC26-UCA结晶,并用20%PEG400作为冷冻保护剂在液氮中快速冷冻。用32%PEG400、4%PEG3350和0.1M乙酸钠(pH5.5)的蓄池溶液使VRC26.01结晶,并用20%乙二醇作为冷冻保护剂在液氮中快速冷冻。用22%PEG8000、5%MPD和0.1M咪唑(pH6.5)的蓄池溶液使VRC26.03结晶,并用20%木糖醇作为冷冻保护剂在液氮中快速冷冻。用14%PEG3350、25%异丙醇和0.1MTris(pH8.5)的蓄池溶液使VRC26.04结晶,并用20%乙二醇作为冷冻保护剂在液氮中快速冷冻。用3M甲酸钠和0.1MTris(pH7.5)的蓄池溶液使VRC26.06结晶,并用20%木糖醇作为冷冻保护剂在液氮中快速冷冻。用4%PEG8000、0.1M乙酸锌和0.1MMES(pH6)的蓄池溶液使VRC26.07结晶,并用20%甘油作为冷冻保护剂在液氮中快速冷冻。用22%PEG4000、0.4M乙酸钠和0.1MTris(pH7.5)的蓄池溶液使VRC26.10结晶,并在没有冷冻保护剂的情况下在液氮中快速冷冻。在SER-CAT束线ID-22和BM-22(AdvancedPhotonSource,ArgonneNationalLaboratory)以的波长收集所有晶体的数据。用HKL2000套件(Otwinowski和Minor(1997))处理所有衍射数据,并分别在COOT(Emsley和Cowtan(2004))和PHENIX(Adams等人.2004)中执行模型构建和精化。对于VRC26.03Fab数据,使用具有来自PDBID3F12的检索模型的PHASER得到由每个不对称单元一个Fab分子组成的分子置换法。VRC26.03然后充当所有剩余VRC26Fab的检索模型。在精化过程中,使用由5%的数据组成的交叉确认(Rfree)测试组,并在精化模型中包括氢原子。在模型构建/精化过程中用MolProbity(Davis等人.2004)定期执行结构确认。用Pymol(DeLanoScientific,SanCarlos,California(2002))做出所有具有蛋白晶体结构的图解表示。实施例6结构建模(刺突和V1V2以及I1和I2)使用软件包UCSFChimera(Pettersen等人.2004)将V1V2、V3-聚糖和CD4-结合位点的确定位置直接映射到无配体的HIV-1BAL刺突的EM密度(EMD-5019)(Liu等人.2008)。如下限定CD4-结合位点:将VRC01-结合的BAL刺突的密度(EMD-5457)(Tran等人.2012)与无配体的定位图进行比对,并将VRC01-结合的gp120的晶体结构(PDBid3NGB)(Zhou等人.2010)拟合至所述密度。使得紧靠Fab结构的EM密度的颜色能够突出显示接触区域。使用相同的方法使用PGT128-结合的三聚体(EMD-1970)和晶体结构(PDBid3TYG)(Pejchal等人.2011)限定V3-聚糖区域,并使用PG9-结合的BG505SOSIP三聚体(EMD-2241)(Julien等人.2013)和V1V2-结合的PG9的晶体结构(PDBid3U4E)(McClellan等人.2011)限定V1V2区域。使用HIV-1刺突的3倍对称性,使用PG9-V1V2晶体结构与SOSIP三聚体的拟合建立V1V2的三聚体取向的模型。在沿着VRC26-UCA至VRC26.01途径的大约相等的成熟距离计算两个中间体。将与中间体有关的突变直接映射在VRC26.01的结构上。VRC26.01结构的35个残基中的14个是无序的,并且用Loopy(Soto等人.2008)模型化,并表示为灰色点。基于突出显示结构发展的时间线的纵向系统发生树,根据近似发生时间给中间体的突变赋予颜色。在GPS-TPS(Pan等人,http://tsp.biocuckoo.org)中进行酪氨酸硫酸化预测。实施例7单个基因组扩增、测序和克隆使用QiagenQIAampViralRNA试剂盒从血浆分离HIV-1RNA,并使用SuperScriptIII逆转录酶(Invitrogen,CA)逆转录成cDNA。从单个基因组模板(Salazar-Gonzalez(2008))扩增包膜基因,并将扩增子使用ABIPRISMBigDyeTerminatorCycleSequencingReadyReaction试剂盒(AppliedBiosystems,FosterCity,CA)直接测序,并在ABI3100自动化遗传分析仪上分析。使用Sequencherv.4.5软件(Genecodes,AnnArbor,MI)组装和编辑全长env序列。使用ClustalX(1.83版)执行多序列比对,并用BioEdit(7.0.9版)编辑,使用HighlighterforAminoAcidSequencesv1.1.0(beta)使序列比对可视化。对于选择压力的分析,并且为了解释SU和PI之间的重组,使用内部脚本基于推断的重组断点将序列分配进2个比对中(一个SU相关的比对,和一个PI相关的比对)。通过从一个参照向另一个参照的同一性的迁移来鉴别断点,并要求与对应的PI/SU相关病毒共有的至少两个连续多态性以便被考虑。然后使用具有GTR+CAT模型的FastTree(Price等人.2010)重构两次比对的种系发生,并固定在PI/SU上。使用在Hyphy(Pond等人.2005)中实现的、FastTree-产生的树用MEME(片断式多样化选择)(Murrell等人.2012)和DEPS(定向选择)(Kosakovsky等人.2008)检测选择压力的信号。从来自LosAlamosHIV数据库(http://www.hiv.lanl.gov/)的2012个过滤的网络比对(N=3990),计算具体氨基酸在一个位点处的频率和V2表位中的净电荷分布。用EnvM引物,5’-TAGCCCTTCCAGTCCCCCCTTTTCTTTTA-3’(SEQIDNO:168)(Gao等人.1996)和定向引物EnvAstop,5′-CACCGGCTTAGGCATCTCCTATGGCAGGAAGAA-3′(SEQIDNO:169)(Kraus等人.2010),使用PfuUltraII酶(Stratagene),通过SGA第一轮产物的再扩增将选择的包膜扩增子克隆进表达载体pcDNA3.1(定向)(Invitrogen)中。将克隆的env基因测序以证实它们精确地匹配测序的扩增子。如生产商所述,使用StratageneQuickChangeII试剂盒(Stratagene)在C-链内的关键残基处突变自体克隆。通过测序来证实突变,使用包膜克隆来产生单轮如上所述的复制Env-假病毒。实施例8自供体CAP256的有效中和抗体的分离CAPRISA急性感染组群包括每月针对HIV-1感染进行检查的高危妇女;62个发生血清转化,并招募进使用定期血液抽吸物的长期纵向研究中。在感染的3年以后,来自七位血清转化者的血浆可以中和超过40%的HIV-1毒株(Gray等人.2011)。这些抗体应答的映射指示大多数是聚糖依赖性的,靶向V1V2区域中的聚糖N160或第三可变区(V3)中的聚糖N332(Gray等人.2011)。在这些中,V1V2应答是最广阔和有效的,特别是在供体CAP256中,其针对一些病毒的血浆滴度超过1∶40,000(Gray等人.2011,Moore等人.2011和Moore等人.2013)。在感染后59、119和206周,通过高通量B细胞培养、微量中和实现的功能筛选和反转录-PCR,使用来自供体CAP256的血液记忆B细胞分离33种单克隆抗体以回收抗体可变区(Huang等人.2012、Huang等人.2013和Tiller等人.2008)(表1)。将这些抗体(命名为CAP256-VRC26.01-33)亚克隆并重构为IgG1抗体。所有33种是体相关的,并通过35-37个氨基酸(Kabat编号)(Kabat等人.1991)的特别长CDRH3来区分(图1)。重链和λ链显示相对于它们的种系编码的V-基因的4-15%的体细胞突变:Vλ1-51*02(对于轻链)和VH3-30*18(对于重链)。33种抗体显示不同程度的异源病毒中和,具有一定范围的宽度和效能(图1和表3.1、3.2和3.3)。它们对许多毒株(特别是亚型A和C病毒)是非常有效的,具有在纳摩尔范围内的50%抑制浓度(IC50)。最宽的CAP256-VRC26.25相对于种系VH基因突变了约12%,并中和总分离物的194个病毒集合的63%,几何平均IC50为0.03μg/ml(图9)。当血浆滴度较低时,在第59周分离的最早的抗体CAP256-VRC26.01表现出20%的跨进化枝中和(图1、表3.1、3.2和3.3),尽管相对于种系VH仅偏离约8%和相对于种系轻链偏离小于4%(图1)。实施例9CAP256-VRC26抗体识别V1V2链B和C为了理解CAP256血浆的中和与CAP256-VRC26谱系的抗体的中和之间的关联,我们确定了33种分离的抗体中的每一种的中和指纹,并将它们与来自1-4年CAP256的血浆的中和指纹进行对比。这些指纹是高度相关的;此外,所有33种抗体的组合说明了血浆的中和宽度(图10)。还将CAP256-VRC26抗体的中和指纹与以前表征的针对HIV-1易损性的主要位点的HIV-1-中和抗体的中和指纹进行了对比。用PG9和其它V1V2-定向的广谱中和性抗体观察到强关联,但是用靶向其它HIV-1ENV区域的抗体没有观察到。值得注意的是,CAP256-VRC26抗体没有表现出类似于其它V1V2定向的中和抗体的与gp120、gp140或带支架的V1V2)的23种构建体中的任一种的结合。但是,它们表现出与病毒样颗粒和细胞表面表达的gp160的强结合(图2),从而指示对Env的天然四级构象的高度特异性。为了确定抗体竞争组,我们因此使用细胞表面-Env结合测定。CAP256-VRC26.08(图2)和CAP256-VRC26.01与细胞表面gp160的结合被V1V2-定向的抗体PG9竞争,并被V3-聚糖抗体PGT121弱竞争,但是不被MPER-定向的抗体或CD4-结合位点定向的抗体竞争。我们然后聚焦于四链V1V2结构域的链B和C,因为这些链的残基以前被证实是供体CAP256血浆的中和所需要的(Moore等人.2011和Moore等人.2013),且因为V1V2-定向的中和抗体依赖于链B和C(McClellan等人.2011,Pancera等人.2013和Doria-Rosa等人.2012)。针对共有进化枝CEnv序列(ConC)和含有一系列V1V2链B和C点突变体的变体对病毒的中和,我们试验了CAP256-VRC26抗体(图2)。R166A和K169E突变消除了中和,其以前已经被证实会消除CAP256血浆的中和(Moore等人.2011)。其它残基具有在CAP256-VRC26家族成员之间变化的适度作用。例如,不同于V1V2-定向的抗体的PG9类别,CAP256-VRC26抗体对在N160以及N156处的聚糖损失部分地和变化地敏感(图2)。为了确证这些结果,我们使用了一对病毒,其为了获得对现有充分表征的广谱中和性的V1V2抗体的敏感性而设计(Doria-Rosa等人.2012)。如预期的,HIV-1毒株6405对CAP256-VRC26抗体是高度抗性的(图2b,左下)。6405的功能获得突变体(其含有在链B-C区域中的5个氨基酸变化(图2b,右下))被12种CAP256-VRC26抗体中的11种中和(图2b,下面)。这些突变向功能性病毒刺突37中的V1V2区域的模型的映射指示识别的表位靠近三聚体轴,从而为观察到的四级特异性提供了结构解释。总之,四级特异性、中和指纹、与PG9的竞争和长CDRH3指示,CAP256-VRC26抗体类似于来自其它供体的V1V2-定向的广谱中和性抗体,并识别四级表位,包括在HIV-1刺突的膜-远端顶点处的V1V2三聚体帽的链C。实施例10通过NGS的CAP256-VRC26谱系的时间表征为了表征CAP256-VRC26抗体谱系的发展,我们使用对VH3和Vλ家族特异性的引物,通过从每个反应500万个PBMC制备的cDNA的多路PCR,分析B细胞免疫球蛋白转录物。针对感染后15-206周之间的8个时间点,分析了PCR产物的454焦磷酸测序读数。尽管在15和30周没有检测到CAP256-VRC26谱系相关的转录物,在感染后38周发现了1,113种谱系相关的重链转录物和52种谱系相关的轻链转录物。在所有较后的时间点鉴别出编码重链和轻链的肉体变体的转录物。为了跟踪谱系内变化的纵向流行,我们利用了分配至与分离的抗体相同的VH3-30种系基因的所有重链读数的同一性-发散图。用第59周-分离的CAP256-VRC26.01作为同一性所指物,大重链序列群体作为第38周在同一性-趋异性图上的分离岛出现。在第48和59周之前,该岛已经繁殖和分裂,具有一些表现出与CAP256-VRC26.01的>95%同一性的序列。在第119周之前,该高同一性岛已经消失,并且较大的岛已经下降至~80%同一性的平均值,其在第206周之前进一步下降至~75%。类似地,用第119周-分离的CAP256-VRC26.08作为所指物,分离的序列的独特模式出现在第38周。流行和同一性都在第119周达到峰值,此时观察到两个分离岛:一个在~95%同一性,且另一个与CAP256-VRC26.08具有~85%同一性。关于其它CAP256-VRC26抗体观察到同一性的类似模式(未显示),流行的峰值对应于分离的周。为了提供CAP256-VRC26谱系的纵向发展的更精确理解,通过种系V基因序列构建和固定了所有独特的谱系转录物的最大似然系统发生树和(图3)。谱系在早期分叉,一个分支通向CAP256-VRC26.01,且第二个发展成CAP256-VRC26.02-12。重链和轻链系统发生树表现出12种B细胞培养物鉴别的抗体的匹配重链和轻链的类似位置,以及其中来自每个时间点的转录物出现在每个树内的类似模式。该一致性提示,NGS和产生的系统发生树反映了抗体重链和轻链的偶联进化行为,树体系结构亲密地靠近CAP256-VRC26抗体谱系的成熟。为了理解CAP256-VRC26谱系的起源,使用最大似然从系统发生树推断重链和轻链的未突变的共同祖先(UCA)(图3)。对于轻链,UCA具有12个氨基酸的CDRL3,与在CAP256-VRC26.01中发现的长度相同。对于重链,推断的UCA具有35个残基的CDRH3,显然是具有单个D-基因的VDJ重组(在每个连接部处具有34和31个核苷酸的N-核苷酸插入的IgHD3-3*01)的结果。该推断的UCA的确认来自几种转录物的鉴别,所述转录物含有35个氨基酸的CDR3、正确的V和J基因和少于3个来自种系的总核苷酸突变。因而,CAP256-VRC26谱系出现在第30和38周之间,具有通过重组整体上形成的显著35-残基CDRH3。该谱系快速地繁殖,在重链和轻链之间具有种系发生和时间一致性。我们还发现了HIV-1感染的时间与系统发生树上的深度之间的一般一致性;即来自较晚时间点的序列在树上更晚地出现,从而指示该B细胞谱系的连续进化。对于每个时间点,仅小部分分支给下一个时间点的分组序列提供根。该树结构似乎代表B细胞繁殖(在每个时间点的许多分组序列)和选择(来自非常少的当前序列的下一个时间点序列的固定)的“快照”。实施例11CAP256-VRC26谱系抗体的结构特征为了提供CAP256-VRC26谱系对与分子识别有关的特征的原子水平定义,我们确定了来自第59、119和206周的六种分离的抗体以及UCA的抗原结合片段(Fab)的晶体结构(表4)。所述结构揭示了延长的CDRH3,其突出到抗原结合表面以上(图4)且含有几个独特的特征,包括2-链β-折叠和CDRH3内二硫键,其具有位于每个链的C端的半胱氨酸。该二硫键不存在于UCA或CAP256-VRC26.01中,但是存在于所有更成熟的CAP256抗体中。预测在CDRH3的顶点附近的酪氨酸100H和100I被硫酸化,且电子密度证实了Tyr100H的O-硫酸化;但是,由于侧链杂乱,Tyr100I的硫酸化是不确定的。这些晶体结构还提供了对四年中谱系的发展的洞察。UCA和CAP256-VRC26.01的CDRH3突出到轻链上面,而更成熟的抗体的CDRH3指向重链。空间群和晶胞的多样性提示,在多种晶体包装条件下维持CDRH3突出的最初轨迹。这些结果提示UCA和以后的抗体之间CDRH3的取向的成熟。可能与此有关,CAP256-VRC26.01以后所有抗体中二硫键的出现与成熟的CDRH3取向的采用相关。通过实验证实了该二硫键的功能重要性:VRC26.03中的有关Cys残基的突变导致中和效能和宽度的缺失。另外,在缺乏二硫键的重链序列中在残基97处的保守甘氨酸在成熟的抗体中被突变成精氨酸,从而与两个半胱氨酸的出现相符合,这进一步限制了在CDRH3的基部的灵活性。CDRH3是一致地阴离子的,具有从-10至-4的范围内的有效电荷。因而,CAP256-VRC26谱系的抗体含有具有与以前确定的V1V2-定向的广谱中和性抗体类似的结构性能的CDRH3,所述结构性能包括酪氨酸-硫酸化、总阴离子特性、长-长度和突出到框架区上面。总之,结构分析指示,最初的B细胞重组会产生柔性的、酪氨酸-硫酸化的CDRH3环。成熟(包括CDRH3内二硫键的进化)似乎会选择与宽HIV-1识别更相容的构象。实施例12HIV-1和CAP256-VRC26谱系之间的发育相互影响超过约3年的感染的CAP256gp160序列分析表明由初级感染(PI)病毒和超级感染病毒(SU)之间的重组驱动的高多样性水平。SU病毒在感染后15周首次检测到(Moore等人.2013),且与PI病毒相差Env中的约17%。差异包括在V2残基165和169处的多态性,和在SU病毒中在位置160处的完整糖基化sequon,其更代表全部HIV-1毒株。在23和34周之间,除了源自PI病毒的重要V1V2区域以外,Envgp160重组体在较大程度上是SU-衍生的。在第30周再次检测到来自SU病毒的V1V2区段,且在感染后48周之前,PIV1V2在较大程度上被SUV1V2替代。位置169在SU病毒中处于强阳性多样化选择压力下(p=0.0001),这与在该C-链残基上的免疫压力一致。所有CAP256-VRC26.01-12抗体都中和SU病毒,但是没有中和PI病毒(除了CAP256-VRC26.06以外),从而提示SUV1V2引起这些抗体。在23-38周之间分离的Env克隆是较大抗性的,从而提示,得到PIV1V2表位的重组是主导性的早期逃逸机制。一个例外是高度敏感的34周克隆,其含有SU-样V1V2区域,具有N160聚糖(仅在1/26个第34周序列中见到)(图11)。此后,用含有N160糖基化sequon的SUV1V2跟踪来自第48和59周的克隆的灵敏度。最早抗体CAP256-VRC26.01(来自第59周)中和了SU病毒,但是令人惊讶地没有中和高度敏感的34-周克隆或来自第48和59周的更晚病毒(其含有SU-样V1V2)。该34周克隆具有K169I突变,其当引入SU病毒中时,会废除CAP256-VRC26.01中和,但是仅轻微影响以后的抗体(图12)。此外,针对K169I(以<亚型C病毒的2%存在)的定向选择(Bayes因子=77,095)的强证据提示,CAP256-VRC26.01或有关的抗体驱动早期逃逸突变,该谱系的成熟允许以后的抗体耐受169I。从94周以后的病毒通过R166S/K或K169E突变逃逸了所述谱系的所有12种分离的抗体(图12)。尽管R166S和K169E介导所有抗体的完全逃逸,4种最宽者(CAP256-VRC26.03、04、08、09)仍然表现出低水平的SUR166K突变体中和(图12)。除了在位置166处的最常见的精氨酸以外,耐受赖氨酸(以所有HIV-1病毒的约13%存在)的能力可能占据这4种抗体的更大宽度(图1)。中和逃逸也与V2表位中从+3(SU)至176周的罕见0的净电荷变化有关(仅在约4%的HIV-1病毒中见到)。这与抗体CDRH3形成对比,所述抗体CDRH3随着时间变成带更少的负电荷(-10至-4,图4),从而反映了病毒表位和抗体互补位的继续共进化。尽管病毒逃逸,中和宽度继续增加,从而提示,敏感的病毒在低水平持续,且继续刺激该谱系。总之,这些结果表明了病毒和抗体之间的复杂相互作用,低流行病毒(含有SU-样V2表位)逃逸CAP256-VRC26谱系的原初B细胞。循环重组体的V2表位中的急剧变化先于增加的中和宽度,更晚的CAP256-VRC26抗体将48和59-周克隆中和,从而提示这些病毒促进抗体成熟。实施例13CAP256-VRC26.01的快速发展为了理解有效的V1V2-定向的抗体的发展所需的关键要素,我们聚焦于CAP256-VRC26.01,即最早分离的抗体。该抗体会中和15%的不同HIV-1毒株,且具有有效的V1V2-定向的抗体的中和所需的所有分子特征(图4)。因此,我们推断了UCA和CAP256-VRC26.01之间的途径上的两种发展中间体(VRC26-I1和VRC26-I2)的重链和轻链(图6和7)。为了评估CAP256-VRC26.01前体和自体病毒之间的相互作用,我们重构了UCA、I1和I2作为IgG并测量了它们的中和活性。UCA表现出弱的、但是可再现的SU中和,且没有表现出PI的中和。VRC26-I1、VRC26-I2和CAP256-VRC26.01抗体表现出逐渐提高的针对SU的效能。在每种情况下,中和被V1V2链C上的特定突变减少或消除(图5)。中和的宽度也逐渐增加,VRC26-I2中和了7种CAP256-VRC26.01-敏感的异源病毒中的6种(图5)。与UCA和中间体有关的B-细胞转录物的频率支持导致CAP256-VRC26.01的该连续发生途径。我们产生了VRC26-I1和I2的结构模型,并将它们与实验确定的UCA和CAP256-VRC26.01的结构进行了对比。I1含有在重链(共145个残基)中的12个突变和在轻链(共110个残基)中的3个突变。突变主要积累在结合表面上的暴露于溶剂的位置,从而提示在结合中的功能作用。对于重链,总氨基酸突变的大约半数发生在CDRH3区域中(图6和7和表5)。总之,与UCA相比,CAP256-VRC26.01含有共255个氨基酸中的39个氨基酸变化。该发展途径允许从单个病毒的边界中和(SU)至异源HIV-1毒株的交叉中和的中和增加。在该发展过程中,CDRH3维持它的长度、酪氨酸硫酸化和一般的阴离子特性。表5:从VRC26-UCA至中间体VRC26-I1和VRC26-I2至VRC26.01的重链核苷酸和氨基酸变化.实施例14CAP256供体中的长抗原结合环的自身反应性和频率CAP256-VRC26谱系中的一个潜在限速发展步骤是产生UCA的35-残基CDRH3的重组事件。据估计,具有编码长(≥24残基,IMGT定义)或非常长(≥28残基)CDRH3的重组抗体基因的人B细胞分别占原初B细胞的~3.5%和0.4%(Briney等人.2012)。这些长B细胞受体已经与自身反应性相关联,并且发生中央和周围排除,从而导致甚至更小的IgG+记忆B细胞群体(Briney等人.2012,Wardemann等人.2003和Ditzel等人.1996)。因此,我们关于对自身抗原的反应性试验了CAP256-VRC26UCA和15种CAP256-VRC26克隆的抗体(Haynes等人.2005)。对于UCA或对于任何克隆抗体,没有注意到与Hep2细胞的反应性。同样地,对于UCA和对于15种选择的抗体中的14种,没有注意到心磷脂反应性;仅对于CAP256-VRC26.03观察到边界反应性(图13)。这提示,耐受性和克隆排除不是CAP256-VRC26谱系的发展中的因素,这不同于关于一些其它HIV-1广谱中和性抗体(Haynes等人.2005,Verkoczy等人.2010)和关于具有长CDRH3环的其它抗体(Briney等人.2012,Wardemann等人.2003和Ditzel等人.1996)已经发现的结果。我们还探究了在该供体中是否可能以更大的频率发现长CDRH3抗体。总B细胞的NGS指示,<0.4%的序列具有大于28个氨基酸长度的CDRH3,从而提示CAP256供体不具有非常高频率的具有长CDRH3区域的克隆谱系。实施例15疫苗和治疗暗示HIV-1V1V2区域是血清中和抗体的一种常见靶标,并且是HIV-1Env上的四种已知的保守中和表位之一(Overbaugh和Morris,2012和Kwong和Mascola,2012)。因而,V1V2中和抗体的引起是HIV-1疫苗设计的一个主要目标。值得注意的是,在RV144Thai疫苗试验中,将增加水平的针对V1V2区域的结合抗体与降低的感染风险相关联(Haynes等人.2012),并且病毒筛选分析表明在该相同区域中的免疫压力(Rolland等人.2012)。尽管在RV144试验中使用的疫苗没有引起与这里和别处描述的那些类似的广谱中和性的V1V2-定向的抗体(Walker等人.2009,Bonsignori等人.2011和Walker等人.2011),所述疫苗的适度保护效力在V1V2区域(作为疫苗引起的抗体的靶标)中具有突出的强兴趣。理论上,这样的抗体应答将是有效地中和的,并与类似于这里所述的那些的不同HIV-1毒株反应。以前确定地表征的V1V2中和抗体和CAP256-VRC26谱系都具有非常长的CDRH3区域,所述区域似乎是穿透聚糖屏障和连接V1V2表位所必需的。一个重要的未得到答案的问题是这些长抗原结合环如何起源。直到现在,不清楚CDRH3区域是通过亲和力成熟过程中的插入而延长,还是通过V(D)J重组完全形成并因此以原初B细胞受体的水平存在。从纵向样品执行抗体基因转录物的NGS的能力允许我们推断CAP256-VRC26谱系的UCA,并确定它含有在遇到HIV-1抗原之前预形成的必需长抗原结合环。我们还推断了导致CAP256-VRC26谱系的刺激和进化的病毒学事件。类似于供体CH505和产生广谱中和性的抗-CD4结合位点抗体的CH103谱系,CAP256中的自体病毒在宽度发展之前表现出广泛变异(Liao等人.2013)。令人感兴趣的是,UCA没有识别供体CAP256中的原发感染病毒,但是较弱地中和在15周以后出现的超级感染病毒,并且含有最常见的进化枝CV1V2序列。随后的抗体-病毒相互作用(包括病毒中和逃逸)似乎会驱动体细胞突变和更交叉反应性的中和的发展。最后,由CAP256-VRC26谱系揭示的V1V2-定向的广谱中和性抗体的独特个体发生提示,在没有惊人水平的体细胞突变的情况下可以实现中和效能和宽度。例如,具有~8%V基因体细胞突变的CAP256-VRC26.03会中和30%的HIV-1毒株(图1)。因而,引起V1V2-定向的抗体的主要要求似乎是由具有突出的阴离子CDRH3的相对罕见的原初B细胞受体识别的适当抗原性V1V2表位的可用性。尽管有些中和抗体似乎需要数年的成熟(Haynes等人.2012,Burton等人.2012,Kwong等人.2012和Mascola等人.2013),我们证实,非常意外的是,V1V2定向的B细胞谱系可以相对快速地获得HIV-1中和宽度。V1V2表位的四级特异性可以帮助该谱系避免诱饵策略,因为gp120的病毒碎片和单体形式不可能结合这类抗体。因而,鉴别能够连接原初B细胞与长CDRH3的抗原的关键特征是靶向该易损性位点的疫苗的设计中的关键步骤。待确定的是,仅仅通过提供适当的三聚体V1V2构建体是否可以引起中和V1V2抗体,或者是否需要反映病毒进化的多个/连续免疫原。总之,这里报道的CAP256-VRC26谱系的发生途径的精确描述会提供尝试引起有效的V1V2-定向的HIV-1-中和抗体的基础。参考文献Adams,P.D.等人(2004)JSynchrotronRadiat11,53-55.Alamyar,E.,等人(2012)Immunomeresearch8,26.Ashkenazy,H.等人(2012)Nucleicacidsresearch40,580-584.Bonsignori,M.,等人(2011)Journalofvirology85,9998-10009.Boyd,S.D.等人(2010)JImmunol184,6986-6992.Briney,B.S.等人(2012)Genesandimmunity13,469-473.Briney,B.S.等人(2012)PloSone7,e36750.Bunnik,E.M.等人(2008)Journalofvirology82,7932-7941.Burton,D.R.等人(2012).Cellhostµbe12,396-407.Crooks,G.E.,等人(2004)Genomeresearch14,1188-1190.Davis,I.W.,等人(2004)Nucleicacidsresearch32,615-619.Ditzel,H.J.等人(1996)JImmunol157,739-749..Doria-Rose,N.A.,等人(2010)Journalofvirology84,1631-1636.Doria-Rose,N.A.,等人(2012)Journalofvirology86,8319-8323.Emsley,P.&Cowtan,K.(2004)ActaCrystallogrDBiolCrystallogr60,2126-2132.Gao,F.等人(1996)Journalofvirology70.Georgiev,I.S.,等人(2013)Science340,751-756.Glanville,J.等人(2009)ProcNatlAcadSci.106,20216-20221.Gray,E.S.等人(2011)Journalofvirology85,4828-4840.Haynes,B.F.等人(2005)Science308,1906-1908.Haynes,B.F.等人(2012)Naturebiotechnology30,423-433.Huang,J.等人(2012)Nature491,406-412.Huang,J.等人(2013)NatureProtocols.Julien,J.P.等人(2013)ProcNatlAcadSci110,4351-4356.Kabat,E.A.等人(1991)SequencesofProteinsofImmunologicalInterest,(U.S.DepartmentofHealthandHumanService,NationalInstitutesofHealth,BethesdaMD).Kong,L.&Sattentau,Q.J.(2012)JAIDSClinicResS8,003.Kosakovsky等人(2008)Molecularbiologyandevolution25,1809-1824.Kraus,M.H.,等人(2010)Virology397,346-357.Kwong,P.D.&Mascola,J.R.(2012)Immunity37,412-425.Larkin,M.A.等人(2007)Bioinformatics23,2947-2948.Li,W.,等人(2001)Bioinformatics17,282-283.Liao,H.X.,等人(2013)Nature496,469-476.Liu,J.,等人(2008)Nature455,109-113.Lynch,R.M.等人(2011)Journalofvirology85,905-915.Mascola,J.R.&Haynes,B.F.(2013)Immunologicalreviews254,225-244.McCoy,L.E.&Weiss,R.A.(2013)TheJournalofexperimentalmedicine210,209-223.McLellan,J.S.等人(2011)Nature480,336-343.Moir,S.等人(2011)Naturestructural&molecularbiology18,1317-1321.Montefiori,D.C.等人(1991)Virology182,635-643.(1991).Montefiori,D.C.(2009)MethodsMolBiol485,395-405.Moore,P.L.,等人(2011)Journalofvirology85,3128-3141.Moore,P.L.等人(2013)Journalofvirology87,4882-4894.Murrell,B.等人(2012)PLoSgenetics8.Otwinowski,Z.&Minor,W.(1997)Methodsinenzymology276,307-326.Overbaugh,J.&Morris,L.(2012)TheAntibodyResponseagainstHIV-1.ColdSpringHarborperspectivesinmedicine2.Pancera,M.&Wyatt,R.(2005)Virology332,145-156.Pancera,M.,等人(2013)Naturestructural&molecularbiology20,804-813.Pantophlet,R.&Burton,D.R.(2006)Annualreviewofimmunology24,739-769.Pejchal,R.等人(2010)ProcNatlAcadSci107,11483-11488.Pejchal,R.等人(2011)Science334,1097-1103.Pettersen,E.F.等人(2004)JComputChem25,1605-1612.Piantadosi,A.,等人(2009)Journalofvirology83,10269-10274.Pond,S.L.,等人(2005)Bioinformatics21,676-679.Price,M.N.,等人(2010)PloSone5.Richman,D.D.等人(2003)ProcNatlAcadSci.100,4144-4149.Rolland,M.,等人(2012)Nature490,417-420.Salazar-Gonzalez,J.F.等人(2008)Journalofvirology82,3952-3970.Sather,D.N.,等人(2009)Journalofvirology83,757-769.Shu,Y.等人(2007)Vaccine25,1398-1408.Soto,C.S.,等人(2008)Proteins70,834-843.Souto-Carneiro,M.M.,等人(2004)JournalofImmunology172,6790-6802.Tamura,K.等人(2011)Molecularbiologyandevolution28,2731-2739.Tian,C.等人(2008)JImmunol180,3279-3288.Tiller,T.,等人(2008)JImmunolMethods329,112-124.Tomaras,G.D.等人(2011)Journalofvirology85,11502-11519.Tong,T.,等人(2012)Journalofvirology86,3574-3587.Tran,E.E.,等人(2012)PLoSpathogens8.vanGils,M.J.等人(2009)AIDS23,2405-2414.vanLoggerenberg,F.等人(2008)PloSone3,e1954,doi:10.1371/journal.pone.0001954.Verkoczy,L.,等人(2010)ProcNatlAcadSci107,181-186.Walker,L.M.,等人(2009)Science326,285-289.Walker,L.M.等人(2010)PLoSpathogens6.Walker,L.M.,等人(2011)Nature477,466-470.Wardemann,H.等人(2003)Science301,1374-1377.Wei,X.,等人(2003)Nature422,307-312.Wu,X.等人(2010)Science329,856-861.Wu,X.等人(2011)Science333,1593-1602.Zhou,T.等人(2010)Science329,811-817.Zhu,J.等人(2012)Frontiersinmicrobiology3,315.Zhu,J.等人(2013)ProcNatlAcadSci.当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1