用于扩增细胞群的方法、试剂盒及装置与流程
本发明要求于2014年4月16日向美国专利商标局提交的美国临时专利申请61/980,506“用于扩增细胞群的方法、试剂盒及装置(Methods,Kits And Apparatus For Expanding A Population Of Cells)”的优先权权益,其内容特此出于所有目的通过引用整体并入本文。
发明领域
本发明涉及细胞群诸如淋巴细胞群的扩增(增殖)。本发明大体上提供了一种用于细胞群扩增(增殖)的新方法及试剂,其需要受体结合分子(诸如文本所述的第一试剂)与细胞表面的受体分子结合,从而向细胞提供初级激活信号。本发明采用一种多聚化剂,其具有固定(结合)结合于其上的、向细胞提供初级激活信号的第一试剂。这种初级激活信号可足以激活细胞以扩增/增殖。此种第一试剂能可逆或不可逆地与多聚化剂结合。该多聚化剂还可以具有固定(结合)于其上的、刺激细胞表面辅助分子的第二试剂。该第二试剂当与细胞表面的辅助分子结合时,可由此刺激已激活的细胞扩增。此外,第二试剂能可逆或不可逆地与多聚化剂结合。多聚化剂可以被固定在固体载体上或是可溶的。一方面,本文所公开的方法是细胞群的连续扩增,其中一个完整的淋巴细胞群被刺激/扩增,然后在适合的固定相中通过层析除去扩增所必需的试剂,被扩增/刺激的细胞任选地用如T细胞受体或嵌合抗原受体(chimeric antigen receptor,CAR)进行转染,并用与所引入的T细胞受体或嵌合抗原受体结合的不同刺激分子进行二次刺激扩增。本发明还涉及一种用于选定细胞群扩增的装置。
背景技术:
体外增殖T细胞群技术的发展对T细胞识别抗原和T细胞激活理解方面的许多进展是至关重要的。用于人抗原特异性T细胞克隆繁殖的培养方法的开发已经用于界定被T细胞识别的由病原体和肿瘤表达的抗原,从而建立免疫疗法以治疗多种人类疾病。抗原特异性T细胞可在体外扩增用于过继性细胞免疫疗法或癌症疗法,其中输注这样的T细胞已在荷瘤宿主中显示出抗肿瘤活性。此外,过继性免疫疗法也被用于治疗免疫妥协个体的病毒感染。
近几年已建立的在缺乏外源性生长因子和佐细胞的情况下体外扩增人T细胞的方法在美国专利US 6,352,694 B1和欧洲专利EP 0 700 430 B1中有所描述。这些专利中公开了一种用于诱导T细胞群增殖的体外方法。该方法包括使T细胞群与固体表面接触,所述固体表面具有直接固定于其上的:(a)第一试剂,其向T细胞提供初级激活信号,从而激活T细胞;和(b)第二试剂,其刺激T细胞表面的辅助分子,从而刺激已激活的T细胞。第一试剂和第二试剂与T细胞的结合诱导T细胞增殖/扩增。在美国专利US 6,352,694 B1和欧洲专利EP 0 700 430 B1中所述的优选第一试剂是单克隆抗CD3抗体,其在T细胞中与TCR/CD3(TCR=T细胞受体(T cell receptor))复合物结合,从而刺激TCR/CD3复合物相关的信号。根据这两篇专利所述的优选第二试剂是与T细胞呈递的辅助分子CD28结合的单克隆抗CD28抗体。该第二试剂与CD28辅助分子的结合提供了对激活的T细胞扩增/增殖所必需的必要共刺激。同时,市售的CD3/CD28(Invitrogen)可用于T细胞扩增。CD3/CD28CTSTM是均质的、4.5μm超顺磁性无菌无热原的、涂覆有抗人T细胞的CD3和CD28细胞表面分子的经亲和纯化的单克隆抗体混合物的聚苯乙烯珠。
然而,这种磁珠,例如,在临床试验或治疗目的所要求的条件下,难以整合到扩增细胞的方法中,因为必须保证这些磁珠在向患者施用扩增的T细胞于之前被完全去除。因此,本发明的目的在于为研究、诊断特别是治疗的目的提供一种用于扩增细胞群诸如调节性T细胞或中枢记忆T细胞的替代方法。理想地,这种新方法也应当适于整合到可用于快速简便扩增为治疗应用所需的细胞群的自动化过程中。
这一目的是通过独立权利要求的主题,尤其是独立权利要求中所述的方法、试剂盒、排布和装置实现的。
发明概述
本发明提供了用于体外扩增所需细胞群的方法、试剂盒、排布和装置,所述细胞表面具有受体分子,其一旦与合适的试剂结合便会向细胞群提供初级激活信号,并从而激活细胞群扩增(增殖)。因此,本发明的方法也可用于诱导细胞群增殖。
第一方面,本发明提供了一种扩增细胞群的体外方法,其包括使包含细胞群的样品与多聚化剂接触,
其中所述多聚化剂具有可逆固定(结合)于其上的、向细胞提供初级激活信号的第一试剂;
其中所述多聚化剂包含至少一个结合位点Z1用于第一试剂的可逆结合,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够可逆地与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,以及
其中所述第一试剂与细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活细胞。
第二方面,本发明提供了一种扩增细胞群的体外方法,其包括使包含细胞群的样品与多聚化剂接触,
其中所述多聚化剂是可溶形式的,并且具有固定(结合)于其上的、向细胞提供初级激活信号的第一试剂;
其中所述多聚化剂包含至少一个结合位点Z1用于第一试剂的结合,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的键与多聚化剂相结合,以及
其中所述第一试剂与细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活细胞。
第三方面,本发明提供了用于扩增细胞群的试剂盒,所述试剂盒包括:
(i)多聚化剂,
其中,所述多聚化剂包含至少一个结合位点Z用于第一试剂的可逆结合,
(ii)第一试剂,其与细胞表面受体分子结合,由此向细胞提供初级激活信号从而激活细胞,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够可逆地与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,以及
(iii)第二试剂,其刺激细胞表面的辅助分子,
其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够可逆地与多聚化剂的结合位点Z2结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合,
其中所述第二试剂与细胞表面的辅助分子结合,从而刺激已激活的细胞。
第四方面,本发明提供了一种用于扩增细胞群的试剂盒,所述试剂盒包括:
(i)多聚化剂,
其中所述多聚化剂是可溶形式的,并且包含至少一个结合位点Z用于第一试剂的可逆结合,
(ii)第一试剂,其与细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活细胞,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合。
第五方面,本发明提供了一种连续扩增淋巴细胞群的体外方法,其中所述淋巴细胞群包含T细胞,所述方法包括:
使含有包含T细胞的淋巴细胞群样品与多聚化剂接触,
其中所述多聚化剂是可溶形式,并且具有可逆固定于其上的(i)向T细胞提供初级激活信号的第一试剂和(ii)刺激T细胞表面辅助分子的第二试剂,
其中所述多聚化剂包含至少一个结合位点Z1用于第一试剂的可逆结合,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够可逆地与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,
其中所述多聚化剂包含至少一个结合位点Z2用于第二试剂的可逆结合,
其中所述第二试剂包含至少一个结合配偶体C2,其中所述结合配偶体C2能够可逆地与多聚化剂的结合位点Z2结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的可逆键与多聚化剂相结合,
其中所述第一试剂与T细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活T细胞,
其中所述第二试剂与T细胞表面的辅助分子结合,由此刺激已激活的细胞,从而所述第一试剂和所述第二试剂一同诱导T细胞扩增。
第六方面,本发明提供了一种包含生物反应器和层析固定相的排布,
其中所述生物反应器适于细胞的扩增,
其中所述固定相适于细胞分离和试剂去除,所述固定相为凝胶过滤基质和/或亲和层析基质,其中所述凝胶过滤和/或亲和层析基质包括亲和试剂,其中所述亲和试剂包含与第一试剂中所含结合配偶体C1特异性结合的结合位点Z1,和/或所述亲和试剂包含与第二试剂中所含结合配偶体C2特异性结合的结合位点Z2,由此适于将第一试剂和/或第二试剂、第一结合配偶体C1和/或游离的第二结合配偶体C2固定于固定相上,
其中,所述生物反应器和所述固定相是流体连接的。
第七方面,本发明提供了一种用于纯化和扩增细胞群的装置,所述装置包含至少一个根据第六方面所述的包含生物反应器和层析固定相的排布。
第八方面,本发明提供了一种能够扩增细胞群的多聚化剂,
其中所述多聚化剂是可溶形式的,并且包含至少一个结合位点Z1用于向细胞提供初级激活信号的第一试剂的可逆结合,
其中所述多聚化剂具有可逆固定(结合)于其上的、向细胞提供初级激活信号所述第一试剂,
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够可逆地与多聚化剂的至少一个结合位点Z1结合,
其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合。
第九方面,本发明提供了一种能够扩增细胞群的组合物,所述组合物包含:
(i)第一多聚化剂,
其中所述第一多聚化剂是可溶形式的,并且包含至少一个结合位点Z1用于向细胞提供初级激活信号的第一试剂的可逆结合,
其中所述第一多聚化剂具有可逆固定(结合)于其上的、向细胞提供初级激活信号所述第一试剂;
其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够可逆地与多聚化剂的至少一个结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,以及
(ii)第二多聚化剂,
其中所述第二多聚化剂是可溶形式的,并且包含至少一个结合位点Z2用于对细胞表面辅助分子进行刺激的第二试剂的可逆结合,
其中所述多聚化剂具有可逆固定(结合)于其上的、刺激细胞表面辅助分子的所述第二试剂,
其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的至少一个结合位点Z2结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合。
附图说明
参考以下详细描述并结合考虑非限制性实施例和附图,将能更好得理解本发明。附图示出了本发明方法的具体实施方式。不希望受到理论的束缚,附图包括了关于下述扩增机制的结论。所给出的结论仅用于说明的目的,并仅仅用于使扩增方法在分子水平上可实现可视化。
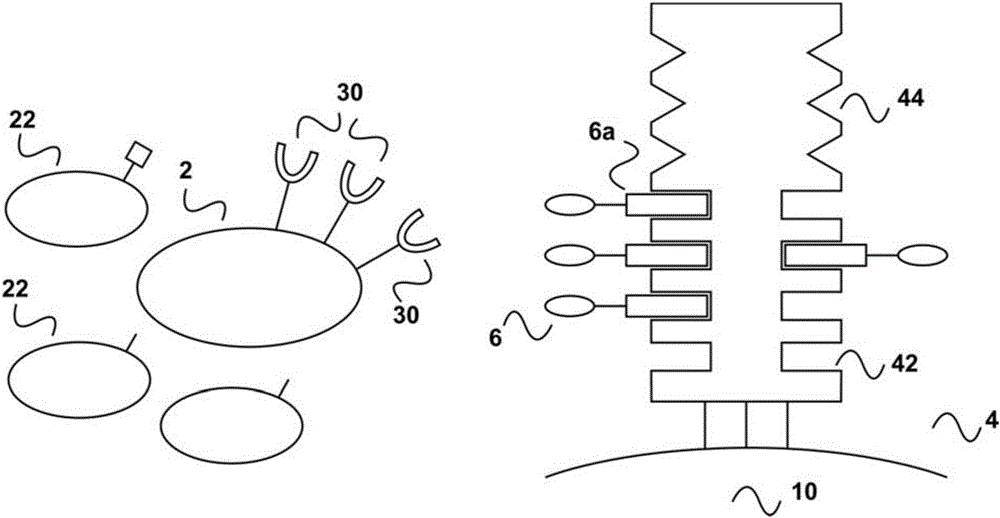
图1示出了一种扩增细胞群的体外方法的实施方式,所述细胞群具有细胞表面受体,其与第一试剂的结合可提供细胞扩增的激活信号。
如图1a所示,包含携带表面受体分子(30)的细胞群(2)的样品与多聚化剂(4)接触。细胞群(2)是与其它缺乏表面受体分子(30)的细胞群(22)混合在一起的。多聚化剂(4)具有可逆固定(结合)于其上的、向细胞提供初级激活信号的第一试剂(6)。多聚化剂(4)包含至少一个结合位点Z1(42)用于第一试剂(6)的可逆结合,而第一试剂(6)包含至少一个结合配偶体C1(6a),其中,结合配偶体C1(6a)能够可逆地与多聚化剂的结合位点Z1(44)结合。因此,为了固定,第一试剂(6)通过结合配偶体C1(6a)和结合位点Z1(42)之间形成的可逆键与多聚化剂(4)相结合。在图1所示的实施例中,多聚化剂(4)具有该实施例中未使用的第二结合位点Z2(44)。多聚化剂(4)本身被固定在固体载体(10)上,例如磁珠、聚合珠、细胞培养板或反应器的表面。细胞群(2)例如诸如B细胞群的淋巴细胞群可以通过CD40受体而被激活(参见例如Carpenter等人,Journal of Translational Medicine 2009,7:93“Activation of human B cells by the agonist CD40antibody CP-870,893and augmentation with simultaneous toll-like receptor 9stimulation)。在这个示例中,细胞表面分子(30)是CD40,第一试剂(6)可以是任何提供所需激活信号的CD40结合分子,例如,单克隆抗体CP-870,893或其抗体结合片段,诸如单价Fab片段。第一试剂(6)的结合配偶体C1可以是例如与例如抗体分子两条多肽链(重链或轻链)其中一条的C端融合或缀合的任何亲和肽。结合配偶体C1(6a)可以是例如,诸如肽Trp-Ser-His-Pro-Gln-Phe-Glu-Lys(SEQ ID NO:01)的链霉亲和素结合肽,也称为美国专利US 5,506,121中所描述的“Strep标记”(Strep-),或者例如,具有包含两个或多个单独结合模块的连续排布的链霉亲和素结合肽,如国际专利公开WO 02/077018或美国专利US 7,981,632中所描述的。当使用链霉亲和素结合肽作为结合配偶体C1时,多聚化剂(4)是与链霉亲和素肽(=第一结合配偶体C1(6a))经由其(生物素)如图示于图1的结合位点Z1(42)可逆结合的任意链霉亲和素突变蛋白。这种多聚化剂可以是在野生型链霉亲和素序列位点第44至47位上包含氨基酸序列Va144-Thr45-Ala46-Arg47(SEQ ID NO:02)的链霉亲和素突变蛋白(类似物),或者是在野生型链霉亲和素序列位点第44至47位上包含氨基酸序列lle44-Gly45-Ala46-Arg47(SEQ ID NO:03)的链霉亲和素突变蛋白(类似物),这两者均在例如美国专利US 6,103,493中有所描述,并且可以商标Strep-购得。在图1的实施例中,多聚化剂(4)可以进一步包括多聚钙调蛋白或谷胱甘肽-S-转移酶,两者均与钙调蛋白结合肽或谷胱甘肽形成可逆键。因此,结合位点Z2(44)可由钙调蛋白或谷胱甘肽-S-转移酶构成。这种例如钙调蛋白与链霉亲和素突变蛋白形成的蛋白缀合物可通过标准蛋白质化学制备,例如,通过使用双功能接头。
如图1b中所示,在使细胞群(2)与多聚化剂(4)接触并通常是用多聚化剂(4)孵育细胞群后,细胞群(2)经由第一试剂(6)与多聚化剂形成复合物/相结合。第一试剂与细胞表面受体分子诸如该实施例中的CD40特异性结合,并提供例如B细胞扩增的细胞扩增激活信号。包含在初始样品中的、缺乏特异性细胞表面分子(30)的其它细胞群(22)无法与多聚化剂结合。在这方面,应当注意的是,细胞群(2)通常在其表面具有多个拷贝的细胞表面分子(30),并且这些多个重拷贝的结合通常是激活所需要的。因此,多聚化剂(4)通常提供不止一个的结合位点Z1,这样使得多个第一试剂(6)可以可逆地结合以实现第一试剂的“多聚化”,即意味着将足够密度的第一试剂呈现给细胞群(2)(未在图1的图式中示出)。在这方面,应当注意的是,本文使用的多聚化试剂本身可具有多个结合位点Z1,例如,链霉亲和素突变蛋白(为同源四聚体)其天然态具有四个这样的结合位点Z1。然而也有可能是,多聚化剂是基于一种本身仅有一个结合位点Z1用于结合配偶体C1可逆结合的化合物。这样的实例是多聚钙调蛋白。钙调蛋白本身仅具有一个钙调蛋白结合肽的结合位点。然而,钙调蛋白可以被生物素化,然后与链霉亲和素寡聚物发生反应(也参见下文),从而提供其中多个钙调蛋白分子在“支架(scaffold)”上以高浓度呈现的多聚化剂,由此提供多聚钙调蛋白。
如图1c所示,孵育(通常进行一段适宜的时间以实现所需细胞群的扩增)后,第一试剂(6)的结合配偶体C1(6a)和多聚化剂(4)的结合位点Z1之间的结合通过破坏各自的可逆键而被破坏。可以通过向含有与多聚化剂结合的细胞群(2)的孵育/反应混合物中加入竞争物来实现这种破坏。为了竞争性破坏(可以理解为竞争性洗脱)第一试剂的结合配偶体C1(6a)和多聚化剂的结合位点Z1(22)之间的可逆键,孵育混合物/细胞群可以与游离的第一结合配偶体C1(20)或所述第一结合配偶体C的类似物接触,所述第一结合配偶体C的类似物能够破坏第一结合配偶体C1(6a)和结合位点Z1(22)之间的键。在结合配偶体C1为与链霉亲和素的生物素结合位点相结合的链霉亲和素结合肽的实施例中,第一游离配偶体C1(20)可以是相应的游离链霉亲和素结合肽或者是竞争性结合的类似物。这样的类似物可以是例如生物素或诸如脱硫生物素的生物素衍生物。
如图1d所示,第一游离配偶体(20)或其类似物的加入导致结合配偶体C1(6a)从多聚化剂(4)上被置换下来,由于结合配偶体C1被包含在第一试剂(6)中,从而导致第一试剂(6)从多聚化剂(4)上被置换下来。第一试剂(6)的这种置换转而导致第一试剂(6)从细胞表面受体(30)上解离,特别是,如果第一试剂和细胞表面受体(30)之间的键的结合亲和力解离常数(Kd)在10-2M至10-13M的范围内并因此也是可逆的。由于这种解离,细胞群(2)的刺激也被终止。因此,本发明提供了这样的优势:细胞群的刺激或扩增的时间段可被精确控制,因此细胞群的功能状态也可被严密控制。关于这点,应当注意的是,抗体分子对其抗原(包括例如细胞表面受体分子,诸如该实施例中的CD40)的结合亲和力通常处于Kd为10-7M至10-13M的亲和力值范围内。因此,常规的单克隆抗体可用作本发明中的第一试剂(当然也可用作如下文所述的第二试剂)。为了避免导致更强结合的任何不期望的亲和作用,单克隆抗体也可以以其单价抗体片段例如Fab片段或单链Fv片段的形式使用。
此外,由于第一试剂从细胞表面分子(30)解离,本发明还具有附加的优势:被刺激的细胞群在刺激期结束时不含刺激试剂和本方法中使用的所有其它试剂,也就是说第一试剂(6)以及结合配偶体C1或其类似物的游离第一配偶体(20)可通过国际专利申请WO 2013/124474中所述的“去除筒(removal cartridge)”从被刺激的细胞群(2)上被轻易去除,而被固定于固体载体诸如生物反应器表面或磁珠上的多聚化剂(4)被保留。因此,回到游离试剂(6)和游离第一配偶体(20)的去除,依照WO 2013/124474中对“去除筒”的描述(参见例如其中的图4),本文图1d中获得的洗脱样品可上样到WO 2013/124474中所述的第二层析柱上。该层析柱具有合适的固定相,其既是亲和层析基质,同时也能作为凝胶渗透基质。该亲和层析基质具有固定于其上的亲和试剂。在当前实施例中,该亲和试剂可以是例如链霉亲和素、链霉亲和素突变蛋白、亲和素、亲和素突变蛋白或其混合物。第一试剂(6)、结合配偶体C1的游离第一配偶体(20)(在此也被称为“竞争试剂”)与亲和试剂结合,由此被固定在层析基质上。结果,含有已分离的扩增细胞群(2)的洗脱样品正耗尽第一试剂(6)和竞争试剂(20)。不含任何试剂的扩增的细胞群(2)此时处于留待进一步使用的条件下,例如,用于诊断应用(例如,进一步的FACSTM分选)或者用于任何基于细胞的治疗应用。
图2示出了本发明扩增方法的另一个实施方式。如图2a所示,样品包含携带两个特异性细胞表面分子(30)和(32)的细胞群(2)。细胞表面分子(30)参与向细胞群提供初级激活信号,而细胞表面分子(32)是参与向细胞提供刺激的细胞表面的辅助分子。所述细胞群可以是,例如,细胞表面分子(30)为TCR/CD3复合物且细胞表面分子(32)为辅助分子CD28的T细胞群。作为初级激活信号的TCR/CD3复合物的结合和作为共刺激物的CD28的结合对于T细胞的扩增/增殖是必要的。T细胞群(2)是与其它缺乏表面受体分子(30)和(32)的细胞群(22)混合在一起的。另外,在本实施方式中,细胞群(2)与多聚化剂(4)相接触。多聚化剂(4)具有可逆固定(结合)于其上的、向细胞提供初级激活信号的第一试剂(6)。此外,多聚化剂具有可逆固定(结合)于其上的、刺激作为细胞表面辅助分子的CD28的第二试剂(8)。
多聚化剂(4)包含至少一个结合位点Z1(42)用于第一试剂(6)的可逆结合,并且第一试剂(6)包含至少一个结合配偶体C1(6a),其中结合配偶体C1(6a)能够可逆地与多聚化剂的结合位点Z1(44)结合。因此,为了固定,第一试剂(6)通过结合配偶体C1(6a)和结合位点Z1(42)之间形成的可逆键与多聚化剂(4)相结合。此外,在图2所述的实施例中,第二试剂(8)包含结合配偶体C2(8a),其中所述结合配偶体C2能够被可逆地与多聚化剂(4)的结合位点Z2(44)结合。第二试剂(8)通过结合配偶体C2(8a)和结合位点Z2(44)之间形成的可逆键与多聚化剂(4)相结合。在本实施例中,第一试剂(6)可能是单克隆抗CD3抗体或其抗原结合片段如Fab片段。第二试剂(8)可能是单克隆抗CD28抗体或其抗原结合片段如Fab片段。第一结合配偶体(6a)可能是与抗CD3抗体或抗CD3抗体片段融合或缀合的链霉亲和素结合肽(6a)。第二结合配偶体(8a)可能是也与CD28抗体或CD28结合抗体片段缀合或融合的钙调蛋白结合肽。关于这点,应当注意的是,抗例如CD3或CD28的单克隆抗体是众所周知的(参见例如,如上所述的美国专利US 6,352,694 B或欧洲专利EP 0 700 430 B1)并且可购自许多供应商例如Santa Cruz Biotechnology(Santa Cruz,CA,USA)、Life Technologies(Carlsbad,CA,USA)、BD Biosciences(San Jose,CA,USA)、Biolegend(San Diego,CA,USA)或者Miltenyi Biotec(Bergisch Gladbach,Germany),仅仅举几个例子。因此,这种单克隆抗体可用作第一试剂和第二试剂,并且可以与例如结合配偶体C1或C2化学偶联(缀合)。或者,也可以从杂交瘤细胞系克隆可变结构域的基因或者使用氨基酸序列已知的抗体,重组产生各自的抗体片段例如Fab片段或者Fv。当在关于产生单克隆抗CD3抗体的杂交瘤细胞系OKT3(CRL-8001TM,如美国专利US 4,361,549所描述的)和抗CD28抗体28.3(如Vanhove等人BLOOD,15July 2003,Vol.102,No.2,pages 564-570和基因库(GenBank)登录号AF451974.1中所描述)两者的实施例部分中使用本文所述的这种方法时,通过用于重组生产的各自表达载体很方便地提供结合配偶体C1和C2,这样抗体片段在轻链或重链的C端携带作为融合肽的结合配偶体C1或C2(关于这点,如Arakawa等人J.Biochem.120,657-662(1996)中所描述的抗体OKT3的重链可变结构域和轻链可变结构域的氨基酸序列出于说明的目的以SEQ ID NO 17和18及所附序列表示出,而如上述Vanhove等人中所描述的抗CD28抗体28.3的可变结构域的氨基酸序列在所附序列表中以SEQ ID NO 19(VH)和20(VL)示出)。克隆抗体分子的可变结构域并重组生产各自抗体片段的这种方法也是本领域技术人员而言所熟知的,参见例如,Skerra,A.(1994)A general vector,pASK84,for cloning,bacterial production,and single-step purification of antibody Fab fragments.Gene 141,79-84,或者Skerra,A.(1993)Bacterial expression of immunoglobulin fragments.Curr Opin Immunol.5,256-562。最后,如图2实施例中所述,还可以通过下列众所周知的进化方法来产生具有抗给定靶物诸如CD3或CD28的抗体样特性人工结合分子的抗体分子,诸如噬菌体展示(例如,Kay,B.K.等人(1996)Phage Display of Peptides and Proteins–A Laboratory Manual,1st Ed.,Academic Press,New York NY;Lowman,H.B.(1997)Annu.Rev.Biophys.Biomol.Struct.26,401–424,或者Rodi,D.J.和Makowski,L.(1999)Curr.Opin.Biotechnol.10,87–93中所综述的)、核糖体展示(Amstutz,P.等人(2001)Curr.Opin.Biotechnol.12,400-405中所综述的)或者如Wilson,D.S.等人(2001)Proc.Natl.Acad.Sci.USA 98,3750-3755中所报道的mRNA展示。
在图2所示的实施例中,多聚化剂(4)具有两个不同的结合位点Z1(42)和Z2(44)。当结合配偶体C1(6a)是链霉亲和素结合肽时,多聚化剂(4)的结合位点Z1(42)由合适的与链霉亲和素肽(6a)可逆结合的链霉亲和素突变蛋白提供。由于结合配偶体C2是钙调蛋白结合肽,多聚化剂(4)的结合位点Z2(44)由多聚钙调蛋白提供。多聚化剂(4)可以是单个分子,例如多聚钙调蛋白与链霉亲和素的缀合物(该替代选择可能通常是在可溶性多聚化的情况下采用),或者也可以由两个独立分子组成。当多聚化剂(44)被固定于如图2所示的固体载体时,后一种选择是优选的。在这种情况下,链霉亲和素突变蛋白和钙调蛋白的混合物被涂覆(固定)在固体载体上,例如,按结合位点Z1和Z2为1:1的摩尔比。关于这点,应当注意的是,由于钙调蛋白固定在固体载体上,无需再制备如上所述的多聚钙调蛋白,但是钙调蛋白固定在表面上足以呈现足够高密度的钙调蛋白(如上所述,钙调蛋白仅有一个钙调蛋白结合肽的结合位点),以确保细胞群(2)的结合。例如,在这种情况下,具有两个抗CD28结合位点的二价抗体片段或者自身具有两个相同结合位点的完整抗体可以用作第二试剂(8)。
如图2b所示,在使T细胞群(2)与多聚化剂(4)接触并通常用多聚化剂(4)孵育细胞群之后,T细胞群(2)经由第一试剂(6)和第二试剂(8)与多聚化剂形成复合物/相结合。第一试剂(6)和第二试剂(8)与TCR/CD3复合物和辅助分子CD28特异性结合,由此诱导T细胞增殖/扩增。
如图2c所示,在孵育(通常进行一段适宜的时间以实现所需细胞群的扩增)之后,第一试剂(6)的结合配偶体C1(6a)和多聚化剂(4)的结合位点Z1之间的结合通过破坏各自的可逆键而被破坏。同样地,第二试剂(8)的结合配偶体C2(8a)和多聚化剂(4)的结合位点Z2之间的结合通过破坏各自的可逆键而被破坏。第一试剂(6)的结合配偶体C1(6a)和结合位点Z1之间的可逆键可被生物素(其充当游离的第一配偶体的类似物(20))破坏,而由于钙调蛋白与钙调蛋白结合肽的结合是钙离子(Ca2+)依赖型,因此第二试剂(8)的结合配偶体C2(8a)和结合位点Z2之间的可逆键可通过加入金属螯合剂(钙离子螯合剂)诸如EDTA或EGTA(其充当游离的第二配偶体的类似物(20))而被破坏。当然,这意味着与细胞群(2)的接触是在含Ca2+的缓冲液中进行。
如图2d所示,第一游离配偶体和第二游离配偶体的类似物(20)的加入,分别导致结合配偶体C1(6a)和C2(8a)从多聚化剂(4)上被置换下来,因而导致第一试剂(6)和第二试剂(8)从多聚化剂(4)上被置换下来。第一试剂(6)和第二试剂(8)的这种置换转而导致第一试剂(6)和第二试剂(8)从TCR/CD3复合物和辅助分子CD28上解离,由此终止细胞群(2)的刺激/扩增。因此,如上所述,本发明提供了可精确控制T细胞群刺激或扩增的时间段从而严密控制T细胞群功能状态的优势。在如图1d所示的洗脱细胞后,第一试剂(6)、第二试剂(8)以及结合配偶体C1的游离第一配偶体和结合配偶体C2的第二游离配偶体的类似物(20)可经由如国际专利申请WO 2013/124474中所述的”去除筒”从被刺激过的细胞群(2)上被轻易地除去。此外,重要的是,假使初始样品是例如由聚蔗糖(Ficoll)梯度得到的PMBC形式的淋巴细胞群,那么T细胞群(2)此时便可用于如本文所定义的连续扩增。由于扩增的细胞群(例如,通过CD3/CD28的初始刺激)在扩增期间可用例如T细胞受体(TCR)或嵌合抗原受体(CAR,也被称为人造T细胞受体)转染,然后经基因修饰的细胞可从初始刺激中释放出来,并随后被第二类刺激例如重新引入的受体所刺激。这些第二刺激可以包括以肽/MHC分子形式存在的抗原刺激物、遗传引入受体的同源(交联)配体(例如,CAR的天然配体)或在新受体的框架内直接结合(例如,通过在受体内识别恒定区)的任意配体(例如抗体)。因此,从这种连续扩增中获得的T细胞群可用于过继性细胞转移。
图3示出了本发明扩增方法的又一个实施方式。此外,该实施例所用的样品包含携带两个特异性细胞表面分子(30)和(32)的T细胞群(2),其中细胞表面分子(30)是TCR/CD3复合物且细胞表面分子(32)是辅助分子CD28。在图3a中,示出的是与多聚化剂(4)接触后的T细胞群(2)。此外,在该实施例中,多聚化剂(4)具有可逆固定(结合)于其上作为第一试剂(6)的、向T细胞提供初级激活信号的抗CD3抗体或其抗原结合片段,以及作为第二试剂(8)的、刺激辅助分子CD28的抗CD28抗体或其抗原结合片段。
图3实施例中所示的多聚化剂(4)仅包括一类结合位点Z1(42)用于第一试剂(6)和第二试剂(8)两者的可逆结合。第一试剂(6)和第二试剂(8)两者都包含至少一个结合配偶体C1(6a、8a),其中结合配偶体C1(6a)和结合配偶体(8a)两者都能够可逆地与多聚化剂的结合位点Z1(44)结合。因此,为了固定,第一试剂(6)和第二试剂(8)通过结合配偶体C1(6a)和结合配偶体C2与结合位点Z1(42)之间形成的可逆键分别与多聚化剂(4)相结合。结合配偶体C1和C2可以是不同的或相同的。例如,结合配偶体C1可以是序列为Trp-Ser-His-Pro-Gln-Phe-Glu-Lys((SEQ ID NO:01),“Strep标记(Strep-)”)的链霉亲和素结合肽,而结合配偶体C2可以是序列为Trp-Ser-His-Pro-Gln-Phe-Glu-Lys-(GlyGlyGlySer)3-Trp-Ser-His-Pro-Gln-Phe-Glu-Lys((SEQ ID NO:04),也被称为“di-tag3”)或为Trp-Ser-His-Pro-Gln-Phe-Glu-Lys-(GlyGlyGlySer)2-Trp-Ser-His-Pro-Gln-Phe-Glu-Lys((SEQ ID NO:05),也被称为“the di-tag2”)的链霉亲和素结合肽,如Junttila等人,Proteomics 5(2005),1199-1203或美国专利7,981,632中所描述的。所有这些链霉亲和素结合肽都与相同的结合位点相结合,即链霉亲和素的生物素结合位点。如果一个或多个这种链霉亲和素结合肽用作结合配偶体C1和C2,那么多聚化剂(4)是一种链霉亲和素突变蛋白。如图3所示,可溶性多聚化剂(4)被使用。在链霉亲和素突变蛋白的情况下,这种可溶性多聚化剂可以是例如链霉亲和素或亲和素的寡聚物或聚合物,或者链霉亲和素或亲和素的任意突变蛋白(类似物)的寡聚物或聚合物。所述寡聚物可以包含三个或更多个链霉亲和素、亲和素或其突变蛋白的单体。所述寡聚物或聚合物可通过多糖交联。这种链霉亲和素或亲和素的寡聚物或聚合物、或链霉亲和素或亲和素突变蛋白的寡聚物或聚合物可通过在第一步中将羧基残基引入多糖例如葡聚糖来制备,基本上如“Noguchi,A.,Takahashi,T.,Yamaguchi,T.,Kitamura,K.,Takakura,Y.,Hashida,M.&Sezaki,H,(1992).Preparation and properties of the immunoconjugate composed of anti-human colon cancer monoclonal antibody and mitomycin C dextran conjugate.Bioconjugate Chemistry 3,132-137”中所述。在第二步中,采用常规的碳二亚胺化学方法,链霉亲和素或亲和素或其突变蛋白经由内部赖氨酸残基和/或游离N端的伯氨基基团与葡聚糖骨架上的羧基基团偶联。可替代地,链霉亲和素或亲和素或任意链霉亲和素或亲和素的突变蛋白的交联寡聚物或聚合物也可以经由双功能接头诸如戊二醛通过交联或通过文献中所述的其它方法获得。
作为结合配偶体C1和C2使用,与多聚化剂的相同结合位点结合(42)相结合的部分具有的优势在于,如图3b所示,(第一结合配偶体C1以及第二结合配偶体C2的)相同游离配偶体或其类似物可用于终止T细胞群(2)的扩增,并将T细胞群(2)从多聚化剂上释放出来。在图3的实施例中,第一和第二配偶体C1和C2的类似物诸如生物素或生物素的衍生物(亚氨基生物素或脱硫生物素)可方便地用于T细胞群(2)的扩增终止和洗脱。
如图3c所示,在如图1d所示洗脱细胞后,第一试剂(6)、第二试剂(8)以及生物素作为结合配偶体C1的游离第一配偶体和结合配偶体C2的游离第二配偶体的类似物,可以通过如国际专利申请WO 2013/124474中所述的”去除筒”从被刺激的细胞群(2)上轻易地去除。此外,使用可溶性多聚化剂(4)的实施方式具有另一个优点,即能够避开任何固体载体诸如磁珠。这意味着将不存在已激活的T细胞被这种磁珠污染的风险。这也意味着符合GMP标准的过程与已知的方法相比更容易建立,诸如使用则必须采用额外的措施以确保最终的扩增T细胞群不含磁珠。此外,可溶性多聚化剂的使用使得其极易从已激活的细胞群(T细胞、B细胞或天然杀伤细胞)中去除,因为细胞可通过离心简单沉淀,而含有可溶性多聚化剂的上清液则可弃除。可替代地,可溶性多聚化剂可在国际专利申请WO 2013/124474的去除筒的凝胶渗透基质中从已扩增的细胞群中被去除。由于不存在固相(例如磁珠),本发明还提供了一种用于细胞扩增的自动封闭系统,其可整合到已知的细胞扩增系统中,诸如可购于GE Healthcare(Little Chalfont,Buckinghamshire,United Kingdom)的W25型Xuri细胞扩增系统(Xuri Cell Expansion System W25)和WAVE生物反应器2/10系统(WAVE Bioreactor2/10System),或者可购于TerumoBCT Inc.(Lakewood,CO,USA)的细胞扩增系统(Cell Expansion System)。
图4示出了CD3+ T应答细胞在体外用可逆固定于涂覆有链霉亲和素突变蛋白Strep-的磁珠上的αCD3和αCD28 Fab片段刺激后增殖的实验结果。图4A是示出了被刺激细胞大小分布(前向散射)的直方图,图4B示出了代表据每次细胞分裂时的细胞数所示的增殖程度的直方图,细胞分裂次数如图4B顶部所标示(0代表未分裂的细胞;5代表经历了至少5次分裂的细胞),图4C示出了刺激4天后的培养皿照片。
图5示出了Jurkat细胞内不同的细胞内钙动员的结果,所述Jurkat细胞是用αCD3抗体OKT3或用Strep-多聚化的OKT3的Fab片段(本文中也称为Fab多聚体)标记的。图5A:Jurkat细胞与钙敏染料Indo-1-AM一起上样,通过注入αCD3mAb(黑色正方形)触发钙释放,或者在进行或不进行预先D-生物素破坏(分别为深灰色三角形和浅灰色圆形)下通过注入αCD3OKT3 Fab多聚体(衍生自亲本细胞系OKT3)触发钙释放,与注入PBS(白色倒三角形)相比较。离子霉素(ionomycine)的应用为阳性对照。基于FL6/FL7比率的变化,通过流式细胞术来监控细胞内Ca2+浓度的时间分辨变化。图5B:经Indo-1-AM标记的Jurkat细胞被如图4a所述的不同的αCD3刺激所激活(OKT3:上方曲线图,αCD3 Fab多聚体:中间曲线图),随后(t=140s)进行D-生物素介导的αCD3 Fab多聚体信号的破坏。注射PBS(下方曲线图)或离子霉素作为阴性或阳性对照。数据代表三个不同的实验。
图6示出了用抗CD3OKT3 Fab多聚体对细胞进行可逆染色的结果。新鲜分离的PBMC用单克隆抗体(左侧散点图,Fab多聚体的亲本克隆)或同源的PE标记的Fab多聚体染色,并在用D-生物素处理之前(左二列)或之后(中间列)进行分析。在后续的清洗步骤后,使用新鲜的PE标记的Strep-(右二列)检测剩余的Fab单体。进行二次Fab多聚体染色的可逆染色细胞作为对照(右列)。只有活(PI阴性)细胞才能显示出来。散点图中的数字表示栅极内的细胞百分比。
图7示出了通过抗CD28 Fab片段的可逆结合所分离的细胞,所述抗CD28 Fab片段与Strep-多聚化,并用荧光标记藻红蛋白进行标记。CD28+细胞通过如国际专利申请WO2013/011011中所述的Fab多聚体磁性细胞选择法从新鲜分离的PMBC中被选择/分离出来。在选择之前,用同源的荧光αCD28多聚体(左侧散点图)或者用直接抗免疫球蛋白κ轻链的抗体(左二散点图,α-IgκmAb)对细胞进行对照染色。在选择之后,用D-生物素处理细胞,随后清洗去除磁珠和Fab单体。随后用CD28 Fab多聚体(右二散点图)或用α-IgκmAb(右散点图)对被释放的CD28+细胞进行(重新)染色,以检测可能残余的Fab单体。只有活的(PI阴性)CD3+细胞才能显示出来。散点图中的数字表示栅极内的细胞百分比。
图8示出了CD3+ T应答细胞在体外用可逆固定于充当可溶性多聚化剂的可溶性寡聚Strep-上的可逆性αCD3/αCD28 Fab片段刺激后增殖的实验结果。对于图8所示的实验结果,300.000个CD3+应答T细胞(Tresp)用2μM羧基荧光素琥珀酰亚胺酯(Carboxyfluorescein succinimidyl ester,CFSE)进行标记,并用不同量的固定有αCD3 Fab片段和αCD28 Fab组合的可溶性Streptactin寡聚物制剂进行刺激,所述αCD3 Fab片段和αCD28 Fab两者均在重链上携带充当链霉亲和素结合肽的Strep标记的。(“1x”对应于用0.5μgαCD3 Fab和0.5μgαCD28 Fab功能化的3μg多聚Strep-tactin;数字表示“1x”的倍数)。剩余的未经刺激的或用空白Strep-tactin多聚体(无Fab)进行刺激的Tresp细胞作为阴性对照。将Tresp细胞一式两份连同300.000个CD3阴性自饲养细胞(经30Gy辐照)一起接种到48孔板上1ml添加有20U/ml白细胞介素2(IL-2)的细胞培养基中。细胞在37℃下进行孵育,不更换培养基,5天后根据CFSE稀释法通过FACS分析对增殖情况进行分析(图8B)。图8A示出了培养5天后细胞的大小分布。直方图示出了活的CD3+细胞,而图8C示出培养后用1mM D-生物素处理并清洗后由刺激试剂释放的细胞。通过使用荧光标记藻红蛋白标记的Strep-(ST-PE)进行重新染色来分析单体Fab片段的解离和去除,并示出了代表性直方图。图8D示出了使用Neubauer计数室计算出的5天后活(台盼蓝阴性)细胞的绝对数,并按各自的刺激条件绘图。中位细胞数在图8D中示出;误差线表示标准偏差(SD)。图8E示出了刺激5天后培养板的照片。
图9示出了本发明的连续扩增方法的图解(图9a),而图9b简单介绍了连续扩增的一些特征和优势。
图10示出了可与本发明的扩增方法一同使用的本发明的一种排布。这种排布(100)包括生物反应器(50)、第一“去除筒”(70)和第二“去除筒”(90)。生物反应器(50)与第一去除筒(70)流体式连接,而第一去除筒与第二去除筒(90)为流体连接。这种排布(100)可以是本文所述的自动化细胞扩增纯化设备的一部分。
本文所述的扩增方法在生物反应器(50)中进行,例如,图3中所述的使用可溶性多聚化剂的扩增方法。在这种情况下,在通过加入竞争物(20)(结合配偶体C1的游离配偶体或其类似物)终止细胞群(2)的激活/扩增之后,从生物反应器中释放的反应混合物包含扩增的细胞群(2)、第一试剂(6)、第二试剂(8)以及可溶性多聚化剂(4)。在该实施例中,第一试剂(6)是包括作为结合配偶体C1的链霉亲和素结合肽在内的CD3结合抗体片段,第二试剂(8)是包括作为结合配偶体C1的链霉亲和素结合肽在内的CD28结合抗体片段,以及竞争物(20)(结合配偶体C1的游离类似物)是生物素。这种反应混合物被施加到去除筒(70)上。这种第一去除筒(70)是如国际专利申请WO 2013/124474所述的去除筒,其包括具有合适固定相的层析柱。所述固定相可用作亲和层析基质,并且同时可用作凝胶渗透基质。这种亲和层析基质具有固定于其上的亲和试剂。在本实施例中,所述亲和试剂可以是例如链霉亲和素、链霉亲和素突变蛋白、亲和素、亲和素突变蛋白或其混合物。因此,第一试剂(6)和第二试剂(8)通过其链霉亲和素结合肽与亲和试剂结合。此外,生物素作为竞争物(20)与亲和试剂结合。因此,这三种试剂全部被固定在第一去除筒的层析基质上,而扩增的细胞群(2)和可溶性多聚化剂(4)通过固定相。然后这种“流经物”被施加到第二去除筒(90)上。此外,这种第二去除筒(90)包括固定相。这种固定相其上包含能够与多聚化剂(4)的结合位点Z1(42)结合的第二亲和试剂。这种亲和试剂可能是例如与固定相共价结合的生物素。这种固定相可能是例如获自Affiland S.A.(Ans-Liege,Belgium)的D-生物素SepharoseTM琼脂糖凝胶。因此,可溶性多聚化剂(4)将被结合(保留)在第二去除筒(90)的固定相上,而扩增的细胞群(2)通过固定相且不含任何试剂。细胞群(2)此时处于留待进一步使用的状态下,例如,用于诊断应用(例如,进一步FACSTM分选)或者用于任何基于细胞的治疗应用。这里应当注意的是,改变排布(100)中第一“去除筒”(70)和第二“去除筒”(90)的顺序当然也是可以的,这样,生物反应器(50)与第二去除筒(90)(直接)流体连接在,而将第一去除筒(70)安排在其后并与第二去除筒(90)流体式连接。在此排布中,多聚化剂(4)将首先从细胞群(2)中去除,随后是第一试剂(6)、第二试剂(8)和例如竞争物(20)被去除。这样的排布也被包含在本发明中并且也可以是本文所述的自动化细胞扩增纯化设备的一部分。
图11示出了可与本发明的扩增方法一同使用的本发明排布的另一个实施方式。这种排布(110)包括生物反应器(50)、第一“去除筒”(70)和第二“去除筒”(90)。生物反应器(50)与第一去除筒(70)流体式连接,且第一去除筒与第二去除筒(90)流体式连接。此外,第二去除筒(90)与生物反应器(50)流体式连接。这种排布(110)也可以是本文所述的自动化细胞扩增纯化设备的一部分。当例如与采用可溶性多聚化剂(4)的扩增方法一同使用时,经纯化的扩增细胞群(2)以第二去除筒(90)的洗脱液获得。由于去除筒(90)与生物反应器(50)流体式连接,因此细胞群(2)可转回至生物反应器(50),例如进行本文所述的连续克隆扩增,通过例如用T细胞受体基因对细胞群进行转染,随后采用本发明的扩增方法进一步(二次)扩增。
图12示出了可与本发明的扩增方法一同使用的本发明的排布的又一个实施方式。这种排布(120)包括生物反应器(50)、第一“去除筒”(70)和第二“去除筒”(90)。生物反应器(50)与第一去除筒(70)流体式连接,并且第一去除筒与第二去除筒(90)流体式连接。类似于图11中所示的实施方式,第二去除筒(90)与生物反应器(50)流体式连接。然而,如国际专利申请WO 2013/124474中所述的“选择筒”(92)被排布在第二去除筒(90)和生物反应器(50)之间。因此,细胞群(2)中包含的细胞亚群(2a)可通过这种如WO 2013/124474中所述的“选择筒”(92)被选择/富集。这种细胞亚群(2a)可或被转移至生物反应器(50),例如以进行本文所述的连续扩增。作为替代地(未示出),这种细胞亚群(2a)可用于基于细胞的治疗。这里再次需要注意的是,使用本文所述的可溶性多聚化剂得以设计出功能性封闭的、且由此不易受污染的自动化细胞纯化扩增设备。此外,由于可溶性多聚化剂避免了对固相材料诸如磁珠的需要,因此这种细胞纯化设备可设计为连续流动设备。
图13示出了纯化的CD4+和CD8+ T应答细胞(Tresp)在体外被可逆固定于作为可溶性多聚化剂的两种可溶性寡聚Strep-突变蛋白上的αCD3/αCD28 Fab片段或αCD3/αCD28/αCD8刺激而增殖的扩增动力学。第一种寡聚Strep-是实施例5中获得的寡聚链霉亲和素突变蛋白(n≥3)的一部分(也被称为“常规骨架”,在图13中以尖端朝下的三角形符号表示),第二种用做可溶性多聚化剂的寡聚链霉亲和素突变蛋白是一种通过使可溶性寡聚链霉亲和素突变蛋白与生物素化的人血清白蛋白(human serum albumin,HSA)发生反应而得到寡聚物。这种基于HSA的可溶性多聚化剂在本文中也被称为“大骨架”。在图13的实验中,扩增是在不更换培养基的情况下进行的。CD4+ T应答细胞的结果在图13A中示出,CD8+ T应答细胞的结果在图13B中示出。关于这点,应当注意的是,实验上使用的可溶性多聚化剂通过可逆地与第一试剂、任选地与第二和第三试剂结合而被功能化,在图中被称为“多聚体”。
图14示出了纯化的CD4+和CD8+ T应答细胞(Tresp)在体外被可逆固定于作为可溶性多聚化剂的两种可溶性寡聚Strep-上的αCD3/αCD28Fab片段刺激而增殖的扩增动力学。第一种寡聚Strep-是实施例5中获得的寡聚链霉亲和素突变蛋白(n≥3)的一部分(也被称为“常规骨架”,在图14中以尖端朝上的三角形符号表示),第二种用做可溶性多聚化剂的寡聚链霉亲和素突变蛋白是基于HSA的可溶性多聚化剂(上述的“大骨架”。在图14的实验中,扩增是在更换培养基的情况下进行的。CD4+ T应答细胞的结果在图14A中示出,CD8+ T应答细胞的结果在图14B中示出。
图15示出了图13和14中纯化的CD4+和CD8+ T应答细胞增殖的扩增动力学所得结果的综合数据,其中,图15A示出了CD4+ T细胞的结果,图15B示出了CD8+ T细胞的结果。在第3天更换培养基的情况下的培养用直线表示,而虚线示出了第3天不更换培养基的情况下的扩增程度的值。图15中所示的数据用输入的细胞数归一化。只有用寡聚链霉亲和素突变蛋白(n≥3)刺激的Tresp、用市售Dynabeads(阳性对照)刺激的Tresp以及未经刺激的T细胞(阴性对照)的数据被示出,而具有“大骨架”的多聚化剂却没有数据。
图16示出了纯化的CD4+和CD8+ T应答细胞在体外被可逆固定于如实施例5中所述的可溶性寡聚链霉亲和素突变蛋白(n≥3)上的αCD3/αCD28Fab片段刺激而激活后T细胞的早期群簇形成。图16A示出了CD4+ T细胞的结果,而图16B示出了CD8+ T细胞的结果。用可溶性多聚化剂(寡聚链霉亲和素突变蛋白)刺激的Tresp、用市售Dynabeads刺激的Tresp(阳性对照)以及未经刺激的T细胞(阴性对照)的数据被示出。
图17示出了在体外被与实施例5中所述的可溶性寡聚链霉亲和素突变蛋白(n≥3)可逆结合的αCD3/αCD28 Fab片段所多克隆刺激的CD3+中枢记忆T细胞(Tcm)(CD3+CD62L+CD45RA-Tcm)的扩增动力学和表型。图17中示出的曲线图代表了据每个时间点上收获的细胞数所示的增殖程度,其中,图17A示出了只添加IL-2的培养基中的增殖,图17B示出了添加了IL-2和IL-15的培养基中的增殖。图17C示出了在这些可变细胞因子环境中培养14后CD62L和CD127的表面表达水平的流式细胞分析。
图18示出了从大量纯化的CD3+CD62L+CD45RA-Tcm应答细胞中进行选择性抗原特异性(Ag特异性)扩增的动力学,其中,所述CD3+CD62L+CD45RA-Tcm应答细胞在体外被肽:MHC分子复合物(充当向细胞提供初级激活信号的第一试剂)和αCD28 Fab片段(充当与细胞表面辅助分子结合的第二试剂)二者刺激,未刺激的T细胞(阴性对照)也被示出。抗原特异性肽与MHC分子的复合物和αCD28 Fab片段两者可逆固定于同一种如实施例5中所述的可溶性寡聚链霉亲和素突变蛋白(n≥3)。用于图18A中抗原特异性扩增的肽为肽CRVLCCYVL(SEQ ID NO:06)和被HLA-C702MHC分子(如Ameres等人,PLOS Pathogens,May 2013,vol.9,issue 5,e1003383所述)限制的即刻早期1蛋白的第309-317位氨基酸,代表对巨细胞病毒(cytomegalovirus,CMV)具有特异性的HLA-C7/IE-1抗原表位。呈递肽的MHC I分子在其重链C端携带链霉亲和素结合肽(SAWSHPQFEK(GGGS)2GGSAWSHPQFEK(SEQ ID NO:07),也就是从IBA GmbH(Germany)购得的“双Strep标记(Twin-Strep-)”。图18A示出了对用肽:MHC-I复合物来增殖的部分Ag特异性细胞的示例性流式细胞分析,其中所述肽:MHC-I复合物对HLA-C7/IE-1抗原表位具有特异性,并作为第一试剂可逆固定于可溶性寡聚链霉亲和素突变蛋白,并向细胞提供初级激活信号。图18B至图18E中的曲线图示说明了据每个时间点上收获的特异性肽:MHCI多聚体-阳性细胞数所示的其他Ag特异性的扩增动力学,这与图18A类似,即使用抗原特异性肽与MHC I分子的不同复合物作为可逆固定于可溶性寡聚链霉亲和素突变蛋白上、向细胞提供初级激活信号的第一试剂。更详细地,图18B示出了使用对CMV的pp65抗原表位有特异性的肽:MHC-I复合物(被HLA-A2402限制的第341-350位氨基酸(QYDPVAALF,(SEQ ID NO:08)))进行扩增的Ag特异性细胞的扩增结果,图18C示出了使用另一种对CMV的pp65抗原表位具有特异性的肽:MHC-I复合物(被HLA-B702限制的第265-274位氨基酸RPHERNGFTV,(SEQ ID NO:09))进行增殖的Ag特异性细胞的扩增结果,图18D示出了使用对腺病毒的Hexon 5抗原表位具有特异性的肽:MHC-I复合物(被HLA-B702限制的第114-124位氨基酸(CPYSGTAYNSL,(SEQ ID NO:10)))进行增殖的Ag特异性细胞的扩增结果,图18E示出了使用对CMV的HLA-B7/IE-1309-317抗原表位具有特异性的肽:MHC-I复合物进行增殖的Ag特异性细胞的扩增结果(示例性的FACS数据可参见上图18A)。所有承载双标记的肽:MHC分子均可从IBA GmbH购得。关于这点,携带“双Strep标记(Twin-Strep-)”作为其C端的HLA-A*2402、HLA-B*0702和HLA-C*0702分子的氨基酸序列在所附序列表中的SEQ ID NO:21、22和23中示出,而β2微球蛋白的氨基酸序列(其与α链一起形成,这意味着HLA编码相应的MHC I分子(HLA encoded molecules the respective MHC I molecule))在所附序列表中的SEQ ID NO:24中示出。此外,图18F示出了对来自图18D的、经HLA-B7/Hexon5114-124刺激/扩增的细胞进行培养14天后,CD62L和CD127表面表达水平的示例性流式细胞分析。
图19示出了从纯化的CD3+CD62L+CD45RA-Tcm应答细胞中进行选择性Ag特异性扩增的动力学,其中,所述CD3+CD62L+CD45RA-Tcm应答细胞在体外被可逆固定于可溶性寡聚链霉亲和素突变蛋白上作为第一和第二试剂的a)抗原特异性肽MHC I复合物和b)αCD28 Fab片段刺激。为了这个目的,使用3μl的被0.5μg配有链霉亲和素结合肽的肽:MHC I类复合物(特异性肽表示被HLA-B0702限制的腺病毒的Hexon 5蛋白的第114-124位氨基酸(CPYSGTAYNSL,SEQ ID NO:10),见上文)和0.5μgαCD28 Fab功能化的Streptactin多聚化剂制剂对500.000个CD3+CD62L+CD45RA-应答Tcm细胞(Tresp)进行Ag特异性刺激。作为替代,4.5μl Streptactin多聚化剂制剂负载有0.5μg肽:MHC I类复合物、0.5μgαCD8 Fab和0.5μgαCD28 Fab。为了比较,使用3μl负载有0.5μgαCD3 Fab和0.5μgαCD28 Fab组合的Streptactin多聚化剂制剂(1mg/ml)进行多克隆刺激。作为上述刺激条件的另一种替代方案,使用了4.5μl负载有0.5μgαCD3 Fab、0.5μgαCD8 Fab和0.5μgαCD28 Fab的Streptactin多聚体制剂。未经处理(未经刺激的)的Tresp细胞作为阴性对照,而经Dynabeads多克隆刺激的Tresp细胞作为阳性对照。Tresp细胞被接种到48孔板上添加有30U/ml IL-2和5ng/ml IL-15的1ml细胞培养基中。细胞在37℃下孵育,每3天更换一次培养基,并于7天和14天后分析细胞计数。图19中所示的照片代表在Ag特异性刺激第5天时群簇形成的程度,以腺病毒HLA-B7/Hexon 5抗原表位为例证。
图20示出了纯化CD8+ T应答细胞的扩增产量和表型,其中所述CD8+ T应答细胞在体外用可逆固定于两种起可溶性多聚化剂作用的可溶性寡聚Strep-上的αCD3/αCD28 Fab片段刺激。第一种寡聚Strep-是实施例5中获得的寡聚链霉亲和素突变蛋白(常规骨架)的一部分,第二种用作可溶性多聚化剂的寡聚链霉亲和素突变蛋白是上文所述的可溶性寡聚物,在此被称为“大”骨架。在这些实验中,寡聚常规链霉亲和素突变蛋白(n≥3)的一部分也被用作多聚化剂,其可被单一的Fab片段(图20A和图20B中的第三根柱)或者αCD3和αCD28 Fab片段组合所功能化。除了用αCD3/αCD28 Fab片段组合刺激之外,还有额外的αCD8 Fab片段(从IBA GmbH购得,Germany)被固定,以测试是否可能优先刺激特定T细胞亚群。图20A示出了代表据第6天收获的细胞数所示的增殖程度的柱状图,其中,所述细胞数与阴性对照(未经刺激的纯化CD8+ T应答细胞)相比较,并以阳性对照(用市售Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)刺激的纯化CD8+ T应答细胞)进行归一化。图20B示出了对细胞培养后CD8和T细胞表面分子CD45RO(这是T细胞增殖和激活的指征)的表面表达水平的流式细胞分析。采用单因素方差分析法对各种刺激条件进行比较,并未检测到显著性差异(n.s.)。
图21示出了纯化的CD8+ T应答细胞扩增的产量和表型,其中,所述CD8+ T应答细胞在体外被可逆固定于起可溶性多聚化剂作用的可溶性寡聚Strep-上的αCD3/αCD28 Fab片段刺激,所述可溶性寡聚Strep-被单一的Fab片段或Fab片段组合(上文已有描述)所功能化。在这些实验中,CD8+ T应答细胞被可溶性多聚化剂(实施例5中的可溶性寡聚Strep-(1mg/ml))刺激,所述可溶性多聚化剂被不同量的αCD3和αCD28 Fab片段(任选地连同上述αCD8 Fab片段)所功能化。术语“1x”对应于被0.5μgαCD3 Fab片段单独功能化的1.5μg多聚化Streptactin和被0.5μgαCD28 Fab单独功能化的1.5μg多聚化Streptactin,或负载有0.5μgαCD3 Fab片段和0.5μgαCD28 Fab的3μl寡聚Streptactin制剂,或负载0.5μg strep标记的αCD3、0.5μg strep标记的αCD8和0.5μg strep标记的αCD28 Fab的4.5μl Streptactin多聚体制剂。相应地,术语“2x”对应于被1μgαCD3 Fab片段单独功能化的3.0μg多聚化Streptactin和被1μgαCD28 Fab单独功能化的3.0μg多聚化Streptactin,这意味着使用了两倍于固定化αCD3 Fab片段化的量。未经处理的Tresp细胞作为阴性对照,而经市售Dynabeads(其上不可逆地固定有αCD3和αCD28单克隆抗体的磁珠)刺激的纯化CD8+ T应答细胞作为阳性对照。图21A示出了代表据第5天收获的细胞数所示的增殖程度的柱状图,其中所述细胞数与阴性对照相比较,并以阳性对照进行归一化。图21B示出了细胞培养后CD8和CD45RO表面表达水平的FACS分析。
图22示出了转导的Jurkat细胞胞内信号级联的激活,其中所述转导的Jurkat细胞被修饰以表达αCD19嵌合抗原受体(CAR),并被实施例5中用作可溶性多聚化剂的寡聚Strep-刺激。CAR的特异性通常来源于由单克隆抗体(mAb)抗原结合区组装的scFv部位,所述单克隆抗体(mAb)与靶物/肿瘤相关抗原诸如CD19特异性结合,并使其与T细胞特异性信号相连(如Hudecek等人,Clin Cancer Res.2013June 15;19(12):3153–3164所述)。在这个实验中,含有αCD19CAR天然配体的CD19胞外域(extracellular domain,ECD)以及识别αCD19-CAR中IgG4间隔区的多克隆αIgG F(ab)2片段(从Jackson Immuno Research商购获得的驴抗人F(ab)2)也被用作向Jurkat细胞提供初级激活信号的第一试剂。与可溶性寡聚链霉亲和素突变蛋白的可逆固定由与CD19的ECD的C端融合的链霉亲和素肽SAWSHPQFEK(GGGS)2GGSAWSHPQFEK(SEQ ID NO:07)提供,或由生物素化的αIgG的(Fab)2片段提供(由于链霉亲和素突变蛋白“m2”以降低的亲和力与生物素结合,这种结合是可逆的并且可例如被加入的过量的游离生物素所置换)。在图22A的对照实验中,300.000个CD3+Jurkat应答细胞(Jresp)用不同量的、被αCD3 Fab和αCD28 Fab功能化(“x1”对应于被0.5μg aCD3-和0.5μg aCD28 Fab功能化的3μg多聚化Streptactin——多克隆Streptamer多聚体)的寡聚Streptactin(1mg/ml)制剂的混合物进行刺激。在图22B的实验中,3μl寡聚Streptactin制剂被0.5μg(x1)或1μg(x2)的CD19胞外域(ECD)功能化,或3μl寡聚Streptactin制剂负载有0.5μg(x1)或1μg(x2)识别IgG4间隔区的αIgG(二者均为CAR特异性多聚体)。经Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)或PMA和离子霉素刺激的Jresp作为阳性对照。Jresp细胞被接种在1.5ml Eppendorf管的200μl添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,并在刺激0min至20min后,置于冰上裂解。
图23示出了在体外被αCD3/αCD28 Fab片段刺激的纯化CD3+ T应答细胞的扩增,所述αCD3/αCD28 Fab片段可逆地固定于实施例5中被用作可溶性多聚化剂的可溶性寡聚Strep-上。在一项实验中,除αCD3/αCD28 Fab片段以外,从IBA GmbH(Germany)商购获得的αCD8 Fab片段(目录编号 6-8000-203)也被固定于链霉亲和素突变蛋白的可溶性寡聚物,以测试是否可能用其上还可逆固定有αCD8 Fab片段的本发明多聚化剂在体外优先刺激大量CD3+培养物中的CD8+ T细胞亚群。更详细地,使用负载有0.5μgαCD3和0.5μgαCD28 Fab组合的3μl寡聚链霉亲和素制剂(1mg/ml)对500.000个纯化的CD3+应答T细胞(Tresp)进行刺激。作为一种替代方法,4.5μl Streptactin寡聚物负载有如上所述的0.5μgαCD3、0.5μgαCD8 Fab和0.5μgαCD28 Fab。未经刺激的Tresp细胞作为阴性对照,经Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)刺激的Tresp细胞作为阳性对照。
图24示出了产生可被用作本发明的可溶性多聚化剂的寡聚链霉亲和素突变蛋白的示例性策略。图24A示出了,在第一步中,使用在野生型链霉亲和素序列位点第44至47位上包含氨基酸序列lle44-Gly45-Ala46-Arg47(SEQ ID NO:03)的链霉亲和素突变蛋白“m2”(SAm2)用于产生具有“常规骨架”的寡聚链霉亲和素突变蛋白。在第二步中,具有“大骨架”的寡聚可溶性链霉亲和素突变蛋白可通过链霉亲和素突变蛋白与生物素化的载体蛋白诸如人血清白蛋白(HSA)的偶联或通过链霉亲和素突变蛋白与合成载体诸如PEG的偶联而产生。图24B:人血清白蛋白(HSA)的生物素化。
发明详述
本发明提供了用于扩增细胞群或用于诱导T细胞增殖的方法、试剂盒和装置。
本文使用的术语“细胞群”包括所有可通过细胞表面受体与向细胞提供初级激活信号的第一试剂结合而扩增的细胞。还可能的是,为了扩增细胞群,可能需要第二试剂与第二细胞表面受体(辅助因子)结合以产生扩增细胞所需的共刺激信号。在一些实施方式中,细胞群可以是淋巴细胞群,包括但不限于B细胞群、T细胞群或天然杀伤细胞群。细胞群的示例性实例是携带CD40或CD137的B细胞(这两种细胞群仅结合提供激活信号的第一试剂配体就可以进行增殖,例如4-1BB,或者αCD40抗体分子或者αCD137抗体分子(参见例如Zhang等人,2010,J Immunol,184:787-795))。可用于B细胞扩增的试剂(第一或第二试剂)的其他示例性实例是与IgG、CD19、CD28或CD14,例如αCD19、αIgG、αCD28或αCD14抗体分子结合的试剂。还可设想,用于B细胞扩增的第一或第二试剂可以包括toll样受体或白细胞介素诸如IL-21的配体(参见例如Dienz O,等人 2009.J.Exp.Med.206:69)。应当注意的是,B细胞的脂多糖依赖性激活也包括在本发明中,因为脂多糖也可用作第一试剂,并且可用如文本所述的结合配偶体C1装配。合适细胞群的其它示例性实例包括T细胞,其通过第一试剂与TCR/CD3的结合以及第二试剂与T细胞上的辅助分子诸如CD28的结合而被激活后扩增。在这种情况下,第一试剂刺激T细胞中的TCR/CD3复合物相关信号,而第二试剂通过与作为辅助因子的CD28结合而提供次级刺激。可用于T细胞扩增的试剂可包括白细胞介素,诸如IL-2、IL-7、IL-15或IL-21(参见例如Cornish等人 2006,Blood.108(2):600-8、Bazdar和Sieg,2007,Journal of Virology,2007,81(22):12670-12674、Battalia等人,2013,Immunology,139(1):109-120)。可用于T细胞扩增的试剂的其它示例性实例为与CD8、CD45或CD90,诸如αCD8、αCD45或αCD90抗体结合的试剂。T细胞群的示例性实例包括抗原特异性T细胞、辅助T细胞、细胞毒性T细胞、记忆T细胞(记忆T细胞的一个示例性实例是CD62L+CD8+特异性中枢记忆T细胞)或调节性T细胞(Treg的一个示例性实例是CD4+CD25+CD45RA+ Treg细胞)。本文使用的术语“T细胞(群)”也可以包括含有嵌合抗原受体(CAR)的T细胞,嵌合抗原受体(CAR)又称为人造T细胞受体或嵌合T细胞受体。因此,包含嵌合抗原受体的T细胞群也可以使用本发明的方法、试剂及设备进行扩增。这一点也可参见实施例15,其中,表达嵌合CD19特异性抗原受体(CAR)的JurkaT细胞通过使用本发明的可溶性多聚化剂而被刺激。合适细胞群的另一个示例性实例包括天然杀伤细胞(NK细胞),其例如可使用与CD16或CD56,诸如αCD16或αCD56抗体相结合的试剂进行扩增。这种αCD16抗体的示例性实例是具有SEQ ID NO:25中所列的VH序列和SEQ ID NO:26中所列的VL序列的抗体3G8(参见例如Hoshino等人,Blood.1991Dec 15;78(12):3232-40)。用于NK细胞扩增的另一种试剂可以是IL-15(参见例如Vitale等人 2002.The Anatomical Record.266:87-92)。合适细胞群的又一个示例性实例包括单核细胞,其可以例如使用与CD14诸如αCD14抗体分子相结合的试剂进行扩增。细胞群可以是任意哺乳动物来源,包括但不限于人、兔、豚鼠、松鼠、仓鼠、猫、狗、狐猴、山羊、猪、马、恒河猴,猕猴或黑猩猩。
因此,根据上文所述,本发明涉及在缺乏外源性生长因子诸如淋巴因子和佐细胞的情况下选择性诱导细胞群诸如B细胞、T细胞或天然杀伤细胞的离体扩增方法。此外,这些细胞诸如B细胞或T细胞的增殖可在不需要抗原的情况下进行诱导,由此提供了具有多克隆反应性的扩增细胞群诸如T细胞群。本文公开的方法可以在延长的时间段内提供选定T细胞群例如CD4+或CD8+ T细胞的持续扩增,以产生这些细胞相对于初始T细胞群的数量的成倍增长。一般而言,在如本文所述的淋巴细胞群的(克隆)扩增的情况下,所有子代可以与被选定扩增的细胞群共享相同的抗原特异性。
同样,根据上文所述,本发明提供了用于抗原特异性T细胞群扩增的方法。为了产生抗原特异性T细胞群,T细胞与适当形式的抗原相接触以触发T细胞中的初级激活信号,即抗原被呈递到T细胞中以致于信号在T细胞中经TCR/CD3复合物被触发。例如,抗原可通过抗原呈递细胞与MHC分子一起被呈递到T细胞。抗原呈递细胞,诸如B细胞、巨噬细胞、单核细胞、树突细胞、朗格汉斯细胞或其它可以向T细胞呈递抗原的细胞,可与T细胞在抗原(例如可溶性抗原)存在下进行孵育,抗原这样抗原呈递细胞将呈递至T细胞。可替代地,表达感兴趣抗原的相关的细胞可与T细胞一起孵育。例如,表达肿瘤相关抗原的肿瘤细胞可与T细胞一起孵育以诱导肿瘤特异性应答。类似地,被病原体例如病毒感染的、呈递病原体抗原的细胞可与T细胞一起孵育。在T细胞群的抗原特异性激活之后,可依照本发明的方法扩增细胞。例如,在抗原特异性建立以后,T细胞可根据本文所述的方法通过与抗CD3抗体(用作第一试剂)和抗CD28抗体(用作第二试剂)一起培养而进行扩增。在另一个实施方式中,第一试剂可以是MHC I:肽复合物,其可与抗原特异性T细胞群结合。在这样一个实施方式中,可以使用任何已知的且可与各自MHC I分子复合的抗原特异性肽。这一点可参见实施例11和12,其中例证了针对四种不同的抗原特异性细胞,从大量CD3+中枢记忆T细胞中选择性抗原特异性扩增Tcm应答细胞。或者,触发细胞扩增的受体的天然配体也可以用作第一试剂。这一点可参见实施例15,其中,CD19胞外域导致经改造得以表达嵌合CD19结合抗原受体(CAR)的转导Jurkat细胞胞内信号级联激活。
细胞群样品可以来自任何合适的来源,通常可以是身体组织或者体液诸如血液的全部样品。在后一种情况中,样品可以是例如外周血单核细胞(peripheral blood mononucleated cell,PBMC)群,其可通过标准分离方法如血细胞的聚蔗糖梯度获得。然而,待扩增的细胞群也可以是纯化的形式,并且可以使用如美国专利US 7,776,562、US 8,298,782、国际专利申请WO02/054065或国际专利申请WO2013/011011中所述的可逆细胞染色/分离技术进行分离。或者,细胞群也可以通过细胞分选经由负磁性免疫细胞粘连获得,正如美国专利US 6,352,649 B1或欧洲专利EP 0 700 430 B1中所述的。如果本文所述的分离方法用于基础研究,那么样品可以是体外细胞培养实验的细胞。样品通常被制备成流体的形式,例如溶液或分散液。
根据上文所述,在一个实施方式中,本发明提供了一种扩增细胞群的体外方法,其包括使含有细胞群的样品与多聚化剂接触。多聚化剂具有可逆固定(结合)于其上的、向细胞提供初级激活信号的第一试剂,其中所述多聚化剂包含至少一个结合位点Z1用于第一试剂的可逆结合。第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1可逆结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合。第一试剂与细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活细胞。
在另一个实施方式中,本发明提供了一种方法,其中所述多聚化剂具有可逆固定(结合)于其上的、刺激细胞表面辅助分子的第二试剂。第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的结合位点Z2可逆结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的可逆键与多聚化剂相结合。第二试剂与细胞表面辅助分子结合,由此刺激已激活的细胞。在这个实施方式中,第一试剂可刺激T细胞中的TCR/CD3复合物相关信号,并且可以是特异性结合CD3的结合剂。在这个实施方式中,T细胞上的辅助分子可以是CD28,而与辅助分子结合的第二试剂是特异性结合CD28的结合剂。在这种情况下,特异性结合CD3的第一试剂可以选自抗CD3抗体、抗CD3抗体的二价抗体片段、抗CD3抗体的单价抗体片段和具有抗体样结合特性的蛋白质CD3结合分子。此外,特异性结合CD28的第二试剂可以选自抗CD28抗体、抗CD28抗体的二价抗体片段、抗CD28抗体的单价抗体片段和具有抗体样结合特性的蛋白质CD28结合分子。所述二价抗体片段可以是(Fab)2’片段,或二价单链Fv片段,而单价抗体片段可以选自Fab片段、Fv片段和单链Fv片段(scFv)。具有抗体样结合特性的蛋白质CD3或CD28结合分子可以是适体、基于脂质运载蛋白(lipocalin)家族多肽的突变蛋白、glubody、基于锚蛋白支架的蛋白、基于晶体支架的蛋白、adnectin和亲和多聚体(avimer)。
一般而言,本发明使用的第一和第二试剂可以是例如抗体、抗体片段和具有抗体样功能的蛋白质结合分子。(重组)抗体片段的实例是Fab片段、Fv片段、单链Fv片段(scFv)、二价抗体片段诸如(Fab)2'片段、双特异抗体(diabody)、三链抗体(triabody)(Iliades,P.,等人,FEBS Lett(1997)409,437-441)、十链抗体(decabody)(Stone,E.,等人,Journal of Immunological Methods(2007)318,88-94)以及其它结构域抗体(Holt,L.J.,等人,.Trends Biotechnol.(2003),21,11,484-490)。在一些实施方式中,第一或第二试剂的一个或多个结合位点可以是二价蛋白质人造结合分子,诸如也称为“双运载蛋白(duocalin)”的二聚脂质运载蛋白突变蛋白。在一些实施方式中,受体结合剂可以具有单个第二结合位点,即它可以是单价的。单价第一或第二试剂的实例包括但不限于单价抗体片段、具有抗体样结合特性的蛋白质结合分子或MHC分子。单价抗体片段的实例包括但不限于Fab片段、Fv和单链Fv片段(scFv)片段,包括二价单链Fv片段。
如上所述,具有抗体样功能的蛋白质结合分子的实例是基于脂质运载蛋白家族多肽的突变蛋白(参见例如,WO 03/029462、Beste等人,Proc.Natl.Acad.Sci.U.S.A.(1999)96,1898-1903)。脂质运载蛋白,诸如胆汁三烯结合蛋白、人中性粒细胞明胶酶相关脂质运载蛋白、人载脂蛋白D或人泪液脂质运载蛋白具有的天然配体结合位点,其可被修饰以使它们与给定靶物结合。可用作特异性结合受体分子的受体结合剂的、具有抗体样结合特性的蛋白质结合分子的其他实例是包括但不限于所谓的glubody(参见例如,国际专利申请WO 96/23879)、基于锚蛋白支架的蛋白(Mosavi,L.K.,等人,Protein Science(2004)13,6,1435-1448)或基于晶体支架的蛋白(例如,国际专利申请WO 01/04144)、如Skerra,J.Mol.Recognit.(2000)13,167-187中所述的蛋白、AdNectin、tetranectin和亲和多聚体(avimer)。亲和多聚体,包括由人类受体结构域家族经外显子改组进化而来的多价亲和多聚体蛋白,包含作为若干细胞表面受体多域串出现的所谓A结构域(Silverman,J.,等人,Nature Biotechnology(2005)23,1556-1561)。Adnectin,衍生自人纤连蛋白结构域,包含三个可被工程化成用于免疫球蛋白样结合至靶物的三个环状结构(loop)(Gill,D.S.&Damle,N.K.,Current Opinion in Biotechnology(2006)17,653-658)。Tetranectin,衍生自相应的人同源三聚体蛋白,同样在C型凝集素结构域中包含可被工程化成用于所需结合的环状结构区域(出处同上)。可作为蛋白质配体的类肽,是不同于多肽的寡聚(N-烷基)甘氨酸,其侧链与酰胺氮连接而不是α碳原子。类肽通常对蛋白酶及其它修饰酶耐受,并且比多肽具有高得多的细胞渗透性(参见例如,Kwon,Y.-U.和Kodadek,T.,J.Am.Chem.Soc.(2007)129,1508-1509)。合适的蛋白质结合分子的另外一些实例是EGF样结构域、三环结构域(Kringle-domain)、I型纤连蛋白结构域、II型纤连蛋白结构域、III型纤连蛋白结构域、PAN结构域、G1a结构域、SRCR结构域、Kunitz/牛胰胰蛋白酶抑制剂结构域、淀粉酶抑肽(tendamistat)、Kazal型丝氨酸蛋白酶抑制剂结构域、三叶型(P型)结构域、C型血管假性血友病因子(von Willebrand factor)结构域、过敏毒素样结构域、CUB结构域、I型甲状腺球蛋白重复、A型LDL受体结构域、Sushi结构域、Link结构域、I型血小板反应蛋白结构域、免疫球蛋白结构域或免疫球蛋白样结构域(例如,结构域抗体或骆驼重链抗体)、C型凝集素结构域、MAM结构域、A型血管假性血友病因子结构域、生长调节素B结构域、WAP型四二硫核心结构域、C型F5/8结构域、血液结合素结构域、SH2结构域、SH3结构域、层粘连蛋白型EGF样结构域、C2结构域、“κ抗体(Kappabody)”(参见Ill.等人,Protein Eng(1997)10,949-57)、所谓的“微抗体(minibody)”(Martin等人,EMBO J(1994)13,5303-5309)、双特异抗体(参见Holliger等人,PNAS USA(1993)90,6444-6448)、所谓的“Janusis”(参见Traunecker等人,EMBO J(1991)10,3655-3659,或者Traunecker等人,Int J Cancer(1992)Suppl 7,51-52)、纳米抗体(nanobody)、微体(microbody)、affilin、亲和体(affibody)、打结素(knottin)、泛素、锌指蛋白、自体荧光蛋白或富亮氨酸重复蛋白。具有抗体样功能的核酸分子的实例是适体。适体折叠成确定的三维基序,并显示出对给定靶结构的高亲和性。
现在转向多聚化剂,多聚化剂的结合位点Z1和Z2可以是相同的(也可参见图3的实施例3)。在这种情况下,可以使用单一的多聚化剂。
在使用可逆结合的第一试剂和任选第二试剂的实施方式中,多聚化剂可以固定在固体表面上。任何固体表面(载体)可用于固定多聚化剂。可固定多聚化剂的固体表面的示例性实例包括磁珠、聚合珠、细胞培养板、微量滴定板、膜或中空纤维。中空纤维例如用作购自TerumoBCT Inc.(Lakewood,CO,USA)的细胞扩增系统中的生物反应器。多聚化剂通常与固体载体共价连接,但是如果需要,非共价相互作用也可用于固定化,例如固定于塑料基质上。如下文更详细的说明,多聚化剂可以是例如与链霉亲和素结合肽可逆结合的链霉亲和素或亲和素突变蛋白。这种链霉亲和素突变蛋白可与任何表面共价连接,例如用于层析纯化树脂(珠),并且可以从IBA GmbH,商业购得这种形式,例如,Strep-琼脂糖凝胶、Strep-大容量Strep-或Strep-易商业购得的多聚化剂的其它示例性实例是固定化金属离子亲和层析(immobilized metal affinity chromatography,IMAC)树脂,诸如树脂(Westburg,Leusden,The Netherlands),其一般可用于可逆固定寡聚组氨酸标记的(his标记的)蛋白,在这里意指,用于携带第一结合配偶体C1或第二结合配偶体C2的第一或第二试剂与低聚组氨酸标记物诸如五组氨酸或六组氨酸标记物的可逆结合。多聚化剂的其它实例是可购于GE Life Sciences钙调蛋白琼脂糖凝胶,其可与含有钙调蛋白结合肽作为结合配偶体C1或C2或琼脂糖凝胶的第一或第二试剂一起使用,并与谷胱甘肽偶联。在这种情况下,结合配偶体C1或C2是谷胱甘肽-S-转移酶。
在本发明方法的其它实施方式中,多聚化剂可以是可溶形式。原则上,可以使用与固定于固体载体上的多聚化剂相同的多聚化剂。可溶形式的多聚化剂可以例如是链霉亲和素突变蛋白寡聚物、钙调蛋白寡聚物、提供至少两个螯合基团K的化合物(寡聚物),其中所述至少两个螯合基团能够与过渡金属离子结合,由此使A部分能够与寡聚组氨酸亲和标记物、多聚谷胱甘肽-S-转移酶或生物素化的载体蛋白结合。
如上所述,所述第一和第二试剂除具有能够与各自细胞表面受体分子结合的结合位点以外,还具有结合配偶体C1或C2(为便于参考,以下将其称为“结合配偶体C”)。此结合配偶体C能够与多聚化剂的结合位点Z结合(Z表示多聚化剂的结合位点Z1或结合位点Z2)。包括在第一或第二试剂中的结合配偶体C和多聚化剂的结合位点Z之间形成的非共价键可以具有任何期望的强度和亲和力,只要其在进行本发明方法的条件下是可被破坏的或可逆的。包括在受体结合剂中的结合配偶体C和多聚化剂的结合位点Z之间的结合力的解离常数(KD)可以是在约10-2M至约10-13M的范围内的值。因此,这种可逆键KD值可以例如为约10-2M至约10-13M、或约10-3M至约10-12M、或约10-4M至约10-11M、或约10-5M至约10-10M。这种键的KD值以及受体结合剂的结合位点B和受体分子之间形成的键的KD、koff和kon率可通过任何合适的手段确定,例如,通过荧光滴定、平衡透析或表面等离子体共振。所述受体分子结合剂可以包括至少一个,包括两个、三个或更多个第二结合配偶体C,所述亲和试剂可以包括至少两个,诸如三个、四个、五个、六个、七个、八个或更多个针对受体分子结合剂中包括的结合配偶体的结合位点。如美国专利US 7,776,562、US 8,298,782或国际专利申请WO 2002/054065中所述,可以选择结合配偶体C和具有一个或更多相应结合位点Z的亲和试剂的任意组合,只要结合配偶体C和亲和试剂的结合位点Z能够可逆地结合或多聚化成(多价)复合物,这通常伴随着亲和力效应。
包括在第一或第二试剂中的结合配偶体可以是寡肽、多肽、蛋白质、核酸、脂质、糖、寡糖或多糖。这样的结合配偶体对多聚化剂结合位点的亲和力比其它物质高。各自结合配偶体的实例包括但不限于免疫球蛋白分子及其片段和具有抗体样功能的蛋白质结合分子。
在一些实施方式中,包括在第一或第二试剂中的结合配偶体C包括生物素,而亲和试剂包括与生物素可逆结合的链霉亲和素类似物或亲和素类似物。
在一些实施方式中,包括在第一或第二试剂中的结合配偶体C包括与链霉亲和素或亲和素可逆结合的生物素类似物,而亲和试剂包括包括与各自生物素类似物可逆结合的链霉亲和素、亲和素、链霉亲和素类似物或亲和素类似物。
在一些进一步的实施方式中,包括在第一或第二试剂中的结合配偶体C包括链霉亲和素或亲和素结合肽,而亲和试剂包括包括与各自链霉亲和素或亲和素结合肽可逆结合的链霉亲和素、亲和素、链霉亲和素类似物或亲和素类似物。
在一些实施方式中,包括在第一或第二试剂中的结合配偶体可以包括链霉亲和素结合肽Trp-Ser-His-Pro-Gln-Phe-Glu-Lys(SEQ ID NO:01),而亲和试剂可以包括在野生型链霉亲和素序列位点第44至47位上含有氨基酸序列Va144-Thr45-Ala46-Arg47(SEQ ID NO:02)的链霉亲和素突变蛋白(类似物),或者在野生型链霉亲和素序列位点第44至47位上含有氨基酸序列lle44-Gly45-Ala46-Arg47(SEQ ID NO:03)的链霉亲和素突变蛋白(类似物),两者均如例如美国专利US 6,103,493中所述,并且以商标名Strep-可商业购得。链霉亲和素结合肽可以例如是单一的肽,诸如如美国专利US 5,506,121中所述的“Strep标记(Strep-)”,或者是具有两个或多个独立结合分子连续排布的链霉亲和素结合肽,正如国际专利申请WO 02/077018或美国专利US 7,981,632中所述的。
在一些实施方式中,第一或第二试剂的结合配偶体C包括技术人员已知为亲和标记物的部分。在这样的实施方式中,亲和试剂包括已知能与亲和标记物结合的相应结合配偶体,例如,抗体或抗体片段。作为已知亲和标记物的一些示例性实例,包括在第一或第二试剂中的结合配偶体可以包括寡聚组氨酸、免疫球蛋白结构域、麦芽糖结合蛋白、谷胱甘肽-S-转移酶(GST)、甲壳质结合蛋白(chitin binding protein,CBP)或硫氧还蛋白、钙调蛋白结合肽(CBP)、FLAG’肽、HA标记物(序列:Tyr-Pro-Tyr-Asp-Val-Pro-Asp-Tyr-Ala,(SEQ ID NO:11))、VSV-G标记物(序列:Tyr-Thr-Asp-Ile-Glu-Met-Asn-Arg-Leu-Gly-Lys,(SEQ ID NO:12))、HSV标记物(序列:Gln-Pro-Glu-Leu-Ala-Pro-Glu-Asp-Pro-Glu-Asp,(SEQ ID NO:13))、T7抗原表位(Ala-Ser-Met-Thr-Gly-Gly-Gln-Gln-Met-Gly,(SEQ ID NO:14))、麦芽糖结合蛋白(MBP)、序列为Gln-Pro-Glu-Leu-Ala-Pro-Glu-Asp-Pro-Glu-Asp(SEQ ID NO:13)的单纯疱疹病毒糖蛋白D的HSV抗原表位、序列为Glu-Gln-Lys-Leu-Ile-Ser-Glu-Glu-Asp-Leu(SEQ ID NO:15)的转录因子c-myc的“myc”抗原表位、V5标记物(序列:Gly-Lys-Pro-Ile-Pro-Asn-Pro-Leu-Leu-Gly-Leu-Asp-Ser-Thr,SEQ ID NO:16),或谷胱甘肽-S-转移酶(GST)。在这样的实施方式中,多聚化剂的一个或多个结合位点(在这种情况下,是抗体或抗体片段)与抗原形成的复合物可通过添加游离的抗原即游离肽(抗原表位标记物)或游离的蛋白质(诸如MBP或CBP)而被竞争性破坏。亲和标记物也可以是寡核苷酸标记物。这种寡核苷酸标记物可例如用于与连接至或包括在亲和试剂中的、具有互补序列的寡核苷酸杂交。
在一些实施方式中,包括在第一或第二试剂的结合配偶体C和多聚化剂的一个或多个结合位点之间的结合在二价、三价或四价阳离子存在时发生。在这一点上,在一些实施方式中,多聚化剂包含通常借助于合适的螯合剂容纳(例如,与之复合)的二价、三价或四价阳离子。在这样的实施方式中,包括在受体结合剂中的结合配偶体可以包括含有(例如,复合有)二价、三价或四价阳离子。各自金属螯合剂的实例包括但不限于乙二胺、乙二胺四乙酸(EDTA)、乙二醇四乙酸(EGTA)、二乙烯三胺五乙酸(DTPA)、N,N-双(羧甲基)甘氨酸(也称为次氮基三乙酸,NTA)、或1,2-双(2-氨基苯氧基)乙烷-N,N,N',N'-四乙酸(BAPTA)。作为一个实例,EDTA与大多数一价、二价、三价和四价的金属离子形成复合物,诸如,钙(Ca2+)、锰(Mn2+)、铜(Cu2+)、铁(Fe2+)、钴(Co3+)和锆(Zr4+),而BAPTA对Ca2+有特异性。作为一个示例性实例,本领域使用的标准方法是使寡聚组氨酸标记物与铜(Cu2+)、镍(Ni2+)、钴(Co2+)或锌(Zn2+)之间形成复合物,这通过螯合剂次氮基三乙酸(NTA)实现。
在一些实施方式中,包括在第一或第二试剂中的结合配偶体C包括钙调蛋白结合肽,而亲和试剂包括如美国专利US 5,985,658或者如本文参照例如图2所述的多聚钙调蛋白。在一些实施方式中,包括在第一或第二试剂中的结合配偶体C包括FLAG肽,而亲和试剂包括与FLAG肽结合的抗体,例如,与如美国专利US 4,851,341中所述的单克隆抗体4E11结合的FLAG肽。在一个实施方式中,包括在第一或第二试剂中结合配偶体C包括寡聚组氨酸标记物,而亲和试剂包括与寡聚组氨酸标记物结合的抗体或过渡金属离子。所有这些结合复合物的破坏可以通过金属离子螯合作用完成,例如钙螯合,例如通过添加EDTA或EGTA(同上所述)完成。钙调蛋白、抗体诸如4E11或螯合金属离子或游离螯合剂可通过常规方法进行多聚化,例如,第一步,通过与链霉亲和素或亲和素或其聚合物的生物素化和络合作用或者通过将羧基残基引入多糖例如葡聚糖,基本上如Noguchi,A,等人Bioconjugate Chemistry(1992)3,132-137中所述;第二步,使用常规碳二亚胺化学法,将钙调蛋白或抗体或螯合金属离子或游离螯合剂经由伯氨基连接到多糖例如葡聚糖主链的羧基上。在这样的实施方式中,包括在第一或第二试剂中的结合配偶体C与多聚化剂的一个或多个结合位点Z之间的结合可通过金属离子螯合作用而破坏。金属螯合作用可例如通过添加EGTA或EDTA而完成。
在一些实施方式中,尤其是,如果多聚化剂是可溶形式的并且是基于链霉亲和素或亲和素的,那么它可以是链霉亲和素或亲和素的寡聚物或聚合物或者链霉亲和素或亲和素的任意突变蛋白(类似物)的寡聚物或聚合物。结合位点Z是亲和素或链霉亲和素的天然生物素结合。各自的寡聚物或聚合物可以通过多糖交联。在一个实施方式中,在第一步中,通过将羧基残基引入多糖例如葡聚糖中制备链霉亲和素的寡聚物或聚合物、或亲和素的寡聚物或聚合物、或链霉亲和素类似物的寡聚物或聚合物、或亲和素类似物的寡聚物或聚合物,基本上如Noguchi,A,等人,Bioconjugate Chemistry(1992)3,132-137中所述。然后在第二步中,使用常规碳二亚胺化学法,可将链霉亲和素或亲和素或其类似物可以经由其内部的赖氨酸残基和/或游离N端的伯氨基连接到葡聚糖主链的羧基上。此外,链霉亲和素或亲和素的交联寡聚物或聚合物、或者链霉亲和素或亲和素的任意突变蛋白(类似物)的交联寡聚物或聚合物也可以经由诸如戊二醛的双官能分子作为接头,通过交联单独的链霉亲和素或亲和素分子(本文中链霉亲和素或亲和素的四聚同源二聚物被称为“单独分子”或各自寡聚物或聚合物的最小结构单元),或者通过本领域的其它方法获得。例如,通过第一步将硫醇基引入链霉亲和素突变蛋白(这可例如通过链霉亲和素突变蛋白和2-亚氨基噻吩(iminothiolane)(Trauts试剂)发生反应而实现),并且在单独的反应中激活链霉亲和素突变蛋白中可获得的氨基,可以产生链霉亲和素突变蛋白的寡聚物。氨基的激活可通过链霉亲和素突变蛋白与市售异源双功能交联剂诸如磺基琥珀酰亚胺基-4-(N-马来酰亚胺甲基)环己烷-1-羧酸酯(磺基SMCC)或琥珀酰亚胺基-6-[(β-马来酰亚胺丙酰氨基)己酸酯(SMPH)发生反应而实现。在第二步中,将如此获得两个反应产物混合在一起,导致包含在一批经修饰的链霉亲和素突变蛋白中的硫醇基与另一批经修饰的链霉亲和素突变蛋白中已激活的(通过马来酰亚胺的功能)氨基发生反应。通过这个反应,链霉亲和素突变蛋白的多聚物/寡聚物形成。这些寡聚物可以具有大于3的任何合适数目的“单独分子”或“链霉亲和素结构单元”,并且寡聚反应程度可根据反应条件变化(参见图24)。这两批经修饰的链霉亲和素突变蛋白发生反应后,寡聚可溶性多聚化剂通常通过体积排阻色谱法进行分离,并且任何所期望的部分都可用作多聚化剂。通常,寡聚物不具有(且无需具有)单一的分子量,但它们通常遵循统计重量分布诸如高斯分布。任何具有多于三个链霉亲和素同源四聚体(结构单元;(n≥3))的寡聚物可用作可溶性多聚化剂。寡聚物可以具有例如3至25个链霉亲和素突变蛋白同源四聚体。具有分子量为约50kDa的链霉亲和素突变蛋白,诸如下文更详细描述的突变蛋白“m1”或“m2”,这些可溶性寡聚物的分子量为约150kDa至约1250kDa。由于每个链霉亲和素分子/突变蛋白具有四个生物素结合位点,因此这样的多聚化剂提供12至100个如本文所述的结合位点Z1(和Z2)。
根据以上公开内容,除了这种只包含交联的链霉亲和素同源四聚体的寡聚多聚化剂以外,可以使四聚链霉亲和素突变蛋白与载体发生反应而获得本发明所用的多聚化剂。除了上述与多糖的反应以外,也可以使用生理学上或药学上可接受的蛋白质诸如血清白蛋白(例如人血清白蛋白(HSA)或牛血清白蛋白(BSA))作为载体蛋白。在这种情况下,链霉亲和素突变蛋白(无论是单个同源四聚体,还是n≥3的寡聚物形式)可与载体蛋白经由非共价相互作用偶联。为了这个目的,生物素化的BSA(其可购自多个供应商例如ThermoFisher Scientific、Sigma Aldrich或Vectorlabs,这里仅举几个例子)可与链霉亲和素突变蛋白发生反应。当这样做时,一些链霉亲和素寡聚物将会经由几个或多个生物素结合位点(Z1、Z2)与生物素化的载体蛋白非共价结合,留下大部分寡聚物结合位点(Z1、Z2)可用于结合诸如第一试剂和任选第二试剂以及任何如本文所述的另外的试剂。因此,通过这种方法,可方便地制备具有多个结合位点Z1的可溶性多聚化剂(参见图24)。或者,链霉亲和素突变蛋白(无论是单个同源四聚体,还是n≥3的寡聚物形式)可与合成载体诸如聚乙二醇(PEG)分子共价偶联。任何适合的PEG分子均可用于此目的,只要这些PEG分子和各自的多聚化剂是可溶的。通常,分子量高达1000Da的PEG分子均可溶于水或本发明可用的培养基。这种基于PEG的多聚化剂通过使用市售已激活的PEG分子(例如,可购自NOF North America Corporation,Irvine,California,USA的PEG-NHS衍生物,或可购自Creative PEGWorks,Chapel Hills,North Carolina,USA的已激活的PEG衍生物)与链霉亲和素突变蛋白中的氨基可以很容易制备。
在链霉亲和素或野生型链霉亲和素(wt-链霉亲和素)中,Argarana等人,Nucleic Acids Res.14(1986)1871-1882中所公开的氨基酸序列被提及。链霉亲和素突变蛋白是通过一个或多个氨基酸的替换、缺失或添加而有别于野生型链霉亲和素序列的多肽,但其保留了野生型链霉亲和素的结合特性。链霉亲和素样多肽和链霉亲和素突变蛋白是与野生型链霉亲和素基本上免疫学等同的多肽,并且特别能够以与野生型链霉亲和素相同或不同的亲和力结合生物素、生物素衍生物或生物素类似物。链霉亲和素样多肽或链霉亲和素突变蛋白可以含有不是野生型链霉亲和素一部分的氨基酸,或者其可仅包括野生型链霉亲和素的一部分。链霉亲和素样多肽也可以是与野生型链霉亲和素的多肽不完全相同的多肽,因为宿主不具有将宿主产生的多肽转化成野生型链霉亲和素的结构所需的酶。术语“链霉亲和素”也包括链霉亲和素四聚体和链霉亲和素二聚体,特别是链霉亲和素同源四聚体、链霉亲和素同源二聚体、链霉亲和素异源四聚体和链霉亲和素异源二聚体。每个亚基通常具有生物素或生物素类似物的结合位点,或者链霉亲和素结合肽的结合位点。链霉亲和素或链霉亲和素突变蛋白的实例在例如WO 86/02077、DE 19641876Al、US 6,022,951、WO 98/40396或WO 96/24606中有所提及。
在一个优选的实施方式中,用作多聚化剂的链霉亲和素突变蛋白是那些如美国专利US 6,103,493和德国专利DE 196 41 876.3中所述的链霉亲和素突变蛋白。基于野生型链霉亲和素的氨基酸序列,这些链霉亲和素突变蛋白在氨基酸位点第44至53位的区域内具有至少一个突变。最小链霉亲和素的突变蛋白是优选的,其N端起始于野生型链霉亲和素第10至16位氨基酸区域,C端终止于野生型链霉亲和素第133至142位氨基酸区域。这样的优选链霉亲和素突变蛋白的实例具有第44位的疏水性脂肪族氨基酸而非Glu、第45位的任意氨基酸、第46位的疏水性脂肪族氨基酸或/和第47位的碱性氨基酸而非Val。链霉亲和素突变蛋白可以是在野生型链霉亲和素序列位点第44至47位上包含氨基酸序列Va144-Thr45-Ala46-Arg47(SEQ ID NO:02)的突变蛋白,或者是在野生型链霉亲和素序列位点第44至47位上包含氨基酸序列lle44-Gly45-Ala46-Arg47(SEQ ID NO:03)的链霉亲和素突变蛋白(类似物)。这种突变蛋白在例如美国专利US 6,103,493中有所描述,并且可以从IBA GmbH以突变蛋白“m1”和突变蛋白“m2”的形式商业购得,其商标为Strep-
在一些实施例中,可以使用根据本发明的方法来排除先前用于细胞扩增的试剂样品。第一或第二试剂以及各自的游离配偶体(竞争试剂)可以例如存在于上述扩增方法的洗脱液中。使用本发明的方法,这样的试剂可被至少基本上(包括完全地)从样品(例如从细胞群)中去除。作为一个示例性实例,如上所定义的第一或第二试剂可从样品中排除至低于例如FACS或Western Blot印迹法检出限的水平。竞争试剂(游离的第一或第二结合配偶体或其类似物)可用于终止并控制扩增,将细胞群从多聚化剂中释放出来。这种竞争试剂可以具有能够在WO 2013/124474中所述的“去除筒”中与亲和试剂的结合位点Z特异性结合的结合位点。在这个实施方式中,本发明的各方法也可用于排除第一和第二试剂以及竞争试剂,包括去除它们。
根据本发明所述的方法可在细胞群生存力至少基本不受影响的任何温度下进行。当在本文中提及至少基本上无害或无毒或至少基本上不影响生存力的条件时,是指这样的条件,即在其中进行扩增的、具有完全生存力的细胞群百分比不低于至少70%,包括至少75%、至少80%、至少85%、至少90%、至少92%、至少95%、至少97%、至少98%、至少99%或至少99.5%。在一些实施方式中,根据本发明所述的方法在约20℃或更高的温度下进行。根据待扩增的细胞群,合适的温度范围可以例如是从约20℃至约45℃,包括从约25℃至约40℃,或从约32℃至37℃。在一些实施方式中,根据本发明所述的方法在恒定的温度下进行,或在选定的温度值上±约5℃、±约4℃、±约3℃、±约2℃、±约1℃或±约0.5℃。本领域的技术人员能够根据经验并考虑细胞特性和扩增条件来确定合适的温度。通常人类细胞是在例如37℃温度下进行扩增。
在进一步的实施方式中,本发明提供了一种扩增细胞群的体外方法,其包括使含有细胞群的样品与多聚化剂接触,其中所述多聚化剂是可溶形式的,并且具有固定(结合)于其上的、向细胞提供初级激活信号的第一试剂。多聚化剂包含至少一个结合位点Z1用于结合第一试剂,其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1结合。第一试剂通过结合配偶体C1和结合位点Z1之间形成的键与多聚化剂相结合,并且第一试剂与细胞表面受体分子结合,由此向细胞提供初级激活信号从而激活细胞。这里应明确指出的是,当使用可溶性多聚化剂时,结合配偶体C1和结合位点Z1之间的键无需是可逆的。
在第二种方法的一个实施方式中,多聚化剂具有固定(结合)于其上的、刺激细胞表面辅助分子的第二试剂,其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的结合位点Z2结合。第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合,其中所述第二试剂与细胞表面辅助分子结合,由此刺激已激活的细胞。
在第二种方法的一个实施方式中,结合配偶体C1和结合位点Z1之间形成的键可以是不可逆的和/或结合配偶体C2和结合位点Z2之间形成的键也可以是不可逆的。
在第二种方法的一个不同的实施方式中,结合配偶体C1和结合位点Z1之间形成的键可以是可逆的。结合配偶体C2和结合位点Z2之间形成的键也可以是可逆的。在这种情况下,所述结合位点Z1和所述结合配偶体C1之间的可逆结合和/或所述结合位点Z2和所述结合配偶体C2之间的可逆结合的解离常数(Kd)在10-2M至10-13M的范围内。
在基于可溶性多聚化剂的第二种方法中,第一和第二试剂、多聚化剂和所有其它试剂以及细胞群也可以以与上述利用第一或第二试剂和多聚化剂间可逆结合的方法相同的方式使用。
本发明进一步提供了用于扩增细胞群的试剂盒,所述试剂盒包括:
(i)多聚化剂,其中所述多聚化剂包含至少一个结合位点Z用于第一试剂的可逆结合,
(ii)第一试剂,其与细胞表面受体分子结合,由此向细胞提供初级激活信号从而激活细胞,其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1可逆结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,和
(iii)第二试剂,其刺激细胞表面辅助分子,其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的结合位点Z2可逆结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合,其中所述第二试剂与细胞表面辅助分子结合从而刺激已激活的细胞。
本发明还提供了用于扩增细胞群的试剂盒,所述试剂盒包括:
(i)多聚化剂,其中所述多聚化剂是可溶形式的并且包含至少一个结合位点Z用于第一试剂的可逆结合,
(ii)第一试剂,其与细胞表面受体分子结合,由此向细胞提供初级激活信号从而激活细胞,其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合。
这第二种试剂盒可以进一步包括:(iii)第二试剂,其刺激细胞表面辅助分子,其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的结合位点Z2结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合。
本文所公开的试剂盒特别地用于当细胞群是淋巴细胞群时。
根据上文公开的内容,本发明还提供了新型多聚化剂和包含能够扩增细胞群的多聚化剂的新型组合物。这种能够扩增细胞群的多聚化剂是可溶形式的,并且包含至少一个结合位点Z1用于可逆结合向细胞提供初级激活信号的第一试剂,其中所述多聚化剂具有可逆固定(结合)于其上的所述向细胞提供初级激活信号的第一试剂;其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的至少一个结合位点Z1可逆结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合。这里应当注意的是,这种多聚化剂可以具有固定于其上的本文所述的任何第一试剂。
本发明的多聚化剂可以进一步包含至少一个结合位点Z2用于可逆结合刺激细胞表面辅助分子的第二试剂,其中,所述多聚化剂具有可逆固定(结合)于其上的、刺激细胞表面辅助分子的第二试剂,其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的至少一个结合位点Z2结合。在这个实施方式中,第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合。
同样根据上方公开的内容,这种多聚化剂能够扩增淋巴细胞群或包含在淋巴细胞群中的亚群。待扩增的淋巴细胞群可以是任何合适的细胞群,例如,B细胞群、T细胞群或天然杀伤细胞群。T细胞群可以是抗原特异性T细胞群、辅助T细胞群、细胞毒性T细胞、记忆T细胞、调节性T细胞群或天然杀伤T细胞群。因此,在这种多聚化剂的实施方式中,第一试剂能够刺激T细胞中的TCR/CD3复合物相关信号。因此,多聚化剂中存在的第一试剂可以是与CD3特异性结合的结合剂,而与辅助分子结合的第二试剂可以是与CD28或CD137特异性结合的结合剂。
在多聚化剂的实施方式中,与CD3特异性结合的第一试剂可以是抗CD3抗体、抗CD3抗体的二价抗体片段、抗CD3抗体的单价抗体片段、和/或具有抗体样结合特性的蛋白质CD3结合分子。在这些实施方式中,与CD28或CD137特异性结合的第二试剂可以是抗CD28抗体、抗CD28抗体的二价抗体片段、抗CD28抗体的单价抗体片段、具有抗体样结合特性的蛋白质CD28结合分子、抗CD137抗体、抗CD137抗体的二价抗体片段、抗CD137抗体的单价抗体片段、具有抗体样结合特性的蛋白质CD137结合分子、4-1BB配体及其任意混合物。因此,本发明的多聚化剂一般可具有固定于其上的一种第一试剂和一种第二试剂混合物,例如,抗CD3抗体作为第一试剂,以及例如,抗CD28抗体和4-1BB配体作为(联合)第二试剂。
如果多聚化剂被用于扩增B细胞,那么固定在多聚化剂上的第一试剂可以是与CD40或CD137特异性结合的结合剂。根据本文公开的内容,在这样的实施方式中,与CD40或CD137特异性结合的第一结合剂可选自抗CD40抗体、抗CD40抗体的二价抗体片段、抗CD40抗体的单价抗体片段和具有抗体样结合特性的蛋白质CD40结合分子,或者抗CD137抗体、抗CD137抗体二价抗体片段、抗CD137抗体的单价抗体片段、具有抗体样结合特性的蛋白质CD137结合分子和CD40配体(CD154)。
此外,根据本发明的大致公开内容,在如本文所述的多聚化剂中,多聚化剂的结合位点Z1和Z2可以是完全相同的。如上所述,这样的多聚化剂可以包括链霉亲和素的寡聚物或聚合物、亲和素的寡聚物或聚合物、可逆结合生物素的链霉亲和素类似物的寡聚物或聚合物、可逆结合生物素的亲和素类似物的寡聚物或聚合物、含有至少两个螯合基团K的试剂,其中所述至少两个螯合基团能够与过渡金属离子结合,由此使所述试剂能够与寡聚组氨酸亲和标记物、多聚谷胱甘肽-S-转移酶、多聚钙调蛋白和生物素化的载体蛋白结合。
本文提供了一种能够扩增细胞群的新型组合物,其可包括:
(i)第一多聚化剂,其中所述第一多聚化剂是可溶形式的,并且包含至少一个结合位点Z1用于向细胞提供初级激活信号的第一试剂的可逆结合,其中所述第一多聚化剂具有可逆固定(结合)于其上的所述向细胞提供初级激活信号的第一试剂,其中所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的至少一个结合位点Z1可逆结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,和
(ii)第二多聚化剂,其中所述第二多聚化剂是可溶形式的,并且包含至少一个结合位点Z2用于对细胞表面辅助分子进行刺激的第二试剂的可逆结合,其中所述多聚化剂具有可逆固定(结合)于其上的所述刺激细胞表面辅助分子的第二试剂,其中所述第二试剂包含结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的至少一个结合位点Z2结合,其中所述第二试剂通过结合配偶体C2和结合位点Z2之间形成的键与多聚化剂相结合。
这样的新型组合物是例如实施例13中使用的反应混合物,其中两种单独的多聚化剂单独用αCD3 Fab片段或单独用αCD28 Fab片段功能化。关于这点应注意的是,这样的组合物在实施例13中显示出具有与其上共同固定有第一试剂和第二试剂两者的单一多聚化剂相同的扩增效率。因此,联合使用两种或多种仅被一种试剂(例如,一种第一试剂或一种第二试剂)单独功能化的多聚化剂与使用一种具有固定于其上的第一试剂和第二试剂两者的联合多聚化剂在用于扩增上功能等同。关于这点,应当指出的是,本发明的多聚化剂可被多种意在用于扩增选定细胞群的试剂(例如,一种、两种、三种、四种或甚至更多种的试剂)所功能化。第三或第四试剂可以例如提供用于扩增所期望细胞亚群的刺激。关于这点可参见例如实施例13,其中可溶性多聚化剂被三种试剂可逆地功能化,即αCD3 Fab片段作为第一试剂、αCD28 Fab片段作为第二试剂和αCD8 Fab片段作为第三试剂以富集在CD3+ T细胞群(淋巴细胞)样品中的CD8+ T细胞亚群。通过使用这种全都可逆固定于同一个多聚化剂上的试剂组合,本发明允许存在从样品例如含有多种不同细胞亚群的样品中优选扩增或选择性富集任何所期望的细胞(亚)群的可能性。然而关于这点应当注意的是,出于此目的也可以使用三种不同的多聚化剂,例如,仅被αCD3 Fab片段功能化的第一多聚化剂、被αCD28 Fab片段功能化的第二多聚化剂和被αCD8 Fab。同样地,可以仅用两种不同的多聚化剂,即仅被αCD3 Fab片段多功能化的第一多聚化剂和被αCD28 Fab片段和αCD8 Fab片段两者功能化的第二多聚化剂。因此,本发明允许以模块化的方式设计任何种类的想要的扩增试剂。
本发明还提供了一种连续扩增淋巴细胞群的体外方法,其中所述淋巴细胞群包括T细胞。这种方法包括:
使含有包含T细胞的淋巴细胞群样品与多聚化剂接触,
其中,所述多聚化剂是可溶形式的,并且具有可逆固定于其上的(i)向T细胞提供初级激活信号的第一试剂和(ii)刺激T细胞表面辅助分子的第二试剂,
其中,所述多聚化剂包含至少一个结合位点Z1用于第一试剂的可逆结合,
其中,所述第一试剂包含至少一个结合配偶体C1,其中所述结合配偶体C1能够与多聚化剂的结合位点Z1可逆结合,其中所述第一试剂通过结合配偶体C1和结合位点Z1之间形成的可逆键与多聚化剂相结合,
其中,所述多聚化剂包括至少一个结合位点Z2用于第二试剂的可逆结合,
其中,所述第二试剂包含至少一个结合配偶体C2,其中所述结合配偶体C2能够与多聚化剂的结合位点Z2可逆结合,其中所述第一试剂通过结合配偶体C2和结合位点Z2之间形成的可逆键与多聚化剂相结合,
其中,所述第一试剂与T细胞表面的受体分子结合,由此向细胞提供初级激活信号从而激活细胞T细胞,
其中,所述第二试剂与T细胞表面的辅助分子结合,从而刺激已激活的细胞,由此所述第一试剂和所述第二试剂一同诱导T细胞扩增。
在这个方法中,使含有淋巴细胞群进而包含T细胞的样品与具有固定于其上的第一和第二试剂的可溶性多聚化剂接触,导致T细胞与多聚化剂特异性结合。
含有包含T细胞的淋巴细胞群样品与多聚化剂的接触可在生物反应器中进行,诸如中空纤维生物反应器(例如,细胞扩增系统的中空纤维生物反应器)或塑料袋生物反应器(例如,来自GE Healthcare的Xuri细胞扩增系统W25中使用的)。
这个方法进一步包括使淋巴细胞群(含有经由第一试剂和第二试剂与多聚化剂结合的T细胞的反应混合物)与以下项接触:(i)能够破坏第一结合配偶体C1和结合位点Z1间键的游离第一结合配偶体C1或其类似物和(ii)能够破坏第二结合配偶体C2和结合位点Z2间键的游离第二结合配偶体C2或其类似物。通过这样做,所述第一试剂结合配偶体C1和所述结合位点Z1之间的可逆键以及所述第二试剂结合配偶体C2和所述多聚化剂结合位点Z2之间的可逆键被破坏,从而在洗脱液中释放经由第一试剂和第二试剂与多聚化剂结合的T细胞并停止T细胞的扩增。
在这个方法中,含有扩增T细胞群的洗脱液(反应混合物,其中扩增反应已通过加入游离第一配偶体或其类似物被终止)可在合适的(第一)固定相上进行层析。所述(第一)固定相可以是如国际专利申请WO 2013/124474中所述的凝胶过滤基质和/或亲和层析基质。这种凝胶过滤和/或亲和层析基质包含亲和试剂,其中所述亲和试剂包含与第一试剂或第二试剂中所包含的结合配偶体C1和/或C2特异性结合的结合位点Z1和/或Z2。通过这样做,第一试剂、第二试剂、第一结合配偶体C1和/或游离第二结合配偶体C2被固定在固定相上。在这种方法中,所述第一固定相与生物反应器流体式连接。
在这种连续扩增的一个实施方式中,多聚化剂的结合位点Z1和Z2是相同的。此外,可以使用单一的多聚化剂。当使用可溶性多聚化剂时,T细胞群(或一般的扩增T细胞群)从可溶性多聚化剂中分离。分离/除去可以利用第二固定相进行。为此,在施加到上述第一固定相之前或之后,含有T细胞和可溶性多聚化剂的混合物在合适的固定相上进行层析。这种第二固定相可以是凝胶过滤基质和/或亲和层析基质,其中所述凝胶过滤和/或亲和层析基质包含亲和试剂。包含在层析树脂上的亲和试剂包括与多聚化剂的结合位点Z1和/或结合位点Z2(如果存在的话)(特异性地)结合的结合配偶体D,从而将多聚化剂固定在固定相上。如果使用基于链霉亲和素的多聚化剂并且第一和第二试剂两者都具有链霉亲和素结合肽作为结合配偶体C1或C2,那么这种第二固定相上的亲和试剂中所包含的结合配偶体D可以是生物素。用作多聚化剂的链霉亲和素的可溶性寡聚物或链霉亲和素突变蛋白的可溶性寡聚物然后和通常与层析基质共价偶联的生物素结合,诸如市售的生物素琼脂糖凝胶(biotin-sepharoseTM)。
在这种连续扩增的方法中,第一试剂可以刺激T细胞中的TCR/CD3复合物相关信号,并且第一试剂可以因此是与CD3特异性结合的结合剂。此外,T细胞上的辅助分子可以是CD28。在这种情况下,与辅助分子结合的第二试剂是与CD28特异性结合的结合剂。
在这种连续扩增的方法中,T细胞可以在扩增期间或者之后用例如T细胞受体(TCR)或嵌合抗原受体(CAR,也称为人造T细胞受体)进行转染。这种用于引入所需受体基因的转染可以用例如任何合适的逆转录病毒载体进行。经遗传修饰的细胞群然后从初始刺激物(例如,CD3/CD28刺激物)中释放出来,随后被第二类的刺激物刺激,例如通过重新引入的受体。这种第二类刺激物可以包括肽/MHC分子形式的抗原刺激物、经遗传引入的受体的同源(交联)配体(例如,CAR的天然配体)、或在新受体的框架内直接结合(例如,通过识别受体内的恒定区)的任意配体(诸如抗体)。这方面参阅Cheadle等人的“Chimeric antigen receptors for T-cell based therapy”Methods Mol Biol.2012;907:645-66或者Barrett等人的Chimeric Antigen Receptor Therapy for Cancer Annual Review of Medicine Vol.65:333-347(2014).
在这种方法中,包含T细胞的淋巴细胞群可以是外周血单核细胞(PBMC)群或经富集或经纯化的T细胞群。淋巴细胞群可以例如源自全血、或源自非流动的血液分离产品或冷冻组织制品。
在这种基于可溶性多聚化剂的连续扩增方法中,第一和第二试剂、多聚化剂以及所有其它试剂和细胞群也可以以与上述利用第一或第二试剂和多聚化剂间可逆结合的方法相同的方式使用。
本发明进一步指向一种包含生物反应器和用于层析的第一固定相的排布。生物反应器适用于细胞扩增,所述固定相适用于细胞分离和去除试剂。第一固定相是凝胶过滤基质和/或亲和层析基质,其中所述凝胶过滤和/或亲和层析基质包含亲和试剂,其中所述亲和试剂包含与第一试剂中所含的结合配偶体C1特异性结合的结合位点Z1,和/或所述亲和试剂包含与第二试剂中所含的结合配偶体C2特异性结合的结合位点Z2。所述第一固定相由此适合于将第一试剂和/或第二试剂、第一结合配偶体C1和/或游离第二结合配偶体C2固定于其上。此外,生物反应器和固定相是流体式连接的。这种排布可用于上述连续扩增,并且可以整合到已知细胞扩增系统中,诸如细胞扩增系统或Xuri细胞扩增系统W25。
在这种排布中,第一固定相被包括在层析柱中或是平面固定相。这种排布可以进一步包括与第一固定相流体式连接的第二固定相。第二固定相可以是凝胶过滤基质和/或亲和层析基质,其中所述凝胶过滤和/或亲和层析基质包含亲和试剂。这种亲和试剂可以包含与多聚化剂的结合位点Z1(特异性地)结合的结合配偶体D,由此适于将多聚化剂固定在固定相上。
本发明进一步指向一种用于纯化和扩增细胞群的装置,所述装置包括至少一个如上所定义的包含生物反应器和用于层析的第一固定相或第二固定相的排布。
所述装置还可以进一步包括流体式连续连接的多个包含生物反应器的装置和固定相的排布。
所述装置可以包括样品入口,其与包含生物反应器和层析固定相的排布中的生物反应器流体式连接。所述装置还可以包括已纯化扩增的靶细胞的样品出口,所述样品出口与至少一个包含生物反应器和层析固定相的排布中最后一个排布的固定相流体式连接。
最后,所述装置可被设计成功能性封闭的系统。
本领域普通技术人员从本发明公开的内容将容易理解,同样可能根据本发明利用目前存在或之后开发的、与本文所述的相应的示例性实施方式发挥基本相同功能或取得基本相同结果的物质、手段、用途、方法或步骤的其他组合。
实验实施例
实施例1:用可逆固定于涂覆有链霉亲和素突变蛋白Strep-磁珠上的αCD3/αCD28 Fab片段进行的CD3+ T应答细胞的刺激/扩增。
用3μM CFSE对300.000个CD3+CD62L应答T细胞(Tresp,通过连续磁富集从非流动供体血液分离产品中分离得到)进行标记,并用 15μl磁珠制剂(10mg磁性微粒/ml,负载有35μg磁珠)中的5μl对细胞进行刺激,其中所述磁珠制剂负载有单独的0.5μgαCD3 Fab片段、单独的0.5μgαCD28 Fab片段、或0.5μgαCD3 Fab片段和0.5μgαCD28 Fab的混合物。
所用的αCD3 Fab片段来源于由杂交瘤细胞系OKT3产生的CD3结合单克隆抗体。杂交瘤细胞系OKT3和OKT3抗体在美国专利US 4,361,549中有所描述,细胞系以登录号CRL-8001TM保藏。所用的CD28 Fab来源于单克隆抗人CD28抗体CD28.3(Vanhove等人,BLOOD,15July 2003,Vol,102,No.2,pages 564-570)。这种抗体CD28.3可变结构域的核苷酸序列以合成单链Fv构建抗人CD28抗体scFv28.3的形式保藏于基因库中,登录号AF451974.1。
如国际专利申请WO2013/011011和WO 2013/124474中所述,两种Fab片段在E.coli中重组生产,其携带IgG1保守序列作为恒定结构域(CH1和Cκ)。两种Fab片段的重链在羧基端与两个链霉亲和素结合模块(SAWSHPQFEK(GGGS)2GGSAWSHPQFEK(SEQ ID NO:07),可以“双Strep标记(Twin-Strep-)”购于IBA GmbH,Germany)的顺序排列相融合。所述αCD3 Fab片段用作第一试剂,其具有用作结合配偶体C1的链霉亲和素结合肽,而所述αCD28 Fab片段用作第二试剂,其具有用作结合配偶体C2的链霉亲和素结合肽。所述(四聚)链霉亲和素突变蛋白“Strep-”作为其上可逆固定有两种Fab片段多聚化剂。
在扩增实验中,以用空白磁珠(无Fab)刺激的Tresp细胞作为阴性对照。将Tresp细胞连同300.000个CD3自饲养细胞(经30Gy辐照)一起一式三份接种到48孔板上3ml添加有10U/ml白细胞介素2(IL-2)的完全细胞培养基(RPMI(Gibco),添加有10%(v/v)胎牛血清、L-谷氨酰胺、b-巯基乙醇、HEPES、青霉素、链霉素和庆大霉素)中。细胞在37°下孵育,不更换培养基,并在4天后通过FACS分析法进行分析。用100μM D-生物素孵育10分钟后进行FACS染色和分析。每种情况的代表曲线在图4中示出。曲线示出了活的CD3+细胞,其中CD3+细胞用碘化丙啶(PI)染色以区分活/死。图4A是示出了被刺激细胞大小分布(前向散射)的直方图。图4A示出了,当在其上固定有0.5μgαCD3 Fab片段和0.5μgαCD28 Fab混合物的磁珠存在下进行孵育,并在体外被可逆固定于涂覆有链霉亲和素突变蛋白Strep-的磁珠上的αCD3/αCD28 Fab片段刺激后,Tresp细胞的特定细胞群被刺激和扩增(与未经刺激的“仅磁珠”对照相比较,其大小/数量增加)。图4B示出了代表据每次细胞分裂的细胞数(如于图4B顶部所标识,0代表未分裂的细胞;5代表经历了至少5次分裂的细胞)所示增殖程度的增殖染料CFSE稀释度直方图。从图4B可以看出,被其上固定有0.5μgαCD3 Fab片段和0.5μgαCD28 Fab混合物的磁珠所刺激的T细胞群大多经历了三次细胞分裂,并表现出比单独的单一刺激更为均匀的增殖模式(未分裂的峰“0”内细胞数较少)。增加的增殖绝对数(更多的细胞在用αCD3和αCD28功能化的磁珠刺激4天均匀增殖)也可以通过更多的培养基消耗示出,如图4C所示,指示剂颜色变为黄色。
实施例2:对Jurkat细胞中不同的细胞内钙动员的分析
在此检测了用αCD3抗体克隆OKT3或与Strep-多聚化的OKT3的Fab片段标记的Jurkat细胞内诱发的不同的细胞内钙动员的实时流式细胞分析(low-cytometric analysis)。
为此,Jurkat细胞负载有钙敏染料Indo-1-AM,通过注射αCD3单克隆抗体OKT3(由杂交瘤细胞系OKT3产生,见上文,黑色正方形)或αCD3Fab片段(源自亲本细胞系OKT3)触发钙释放,其中,αCD3 Fab片段通过其链霉亲和素结合肽与荧光缀合有藻红蛋白的可溶性Strep-Tactin的可逆结合而多聚化。在完整的多聚OKT3 Fab-Strep-Tactin复合物的情况下,钙释放触发持续一段与亲本抗体克隆(深灰色三角形)相同的时间。细胞的激活可以通过注射经的D-生物素处理的、预解离的Fab-Strep-Tactin复合物(浅灰色圆形)而完全避免,等同于注射PBS阴性对照(白色倒三角形)。应用离子霉素作为钙内流的阳性对照。基于FL6/FL7比率的变化,通过流式细胞术来对细胞内Ca2+浓度的时间解析的变化进行监控。从图5A中可以看出,亲本抗体OKT3以及OKT3的多聚单价Fab片段两者均影响钙释放,这意味着OKT3的多聚单价Fab片段基本上具有与亲本抗体相同的功能。值得注意的是,如果在加入Streptactin-OKT3 Fab片段之前,将生物素添加到固定有OKT3 Fab片段的Strep-tactin中,那么多聚OKT3 Fab片段无法触发钙释放。在这种情况下,生物素破坏了作为多聚化剂的Strep-tactin和OKT3 Fab片段之间形成的可逆键。因此单价Fab片段从多聚化剂上被置换出来,并且解离后其不能再通过与Jurkat细胞的CD3结合而触发钙释放。
在图5B所示的实验中,indo-1-AM标记的Jurkat细胞被源自OKT3的αCD3 Fab-Strep-Tactin复合物所激活,如图5A中所示。注射完整的(上图)或预解离的复合物(下图)分别作为阳性或阴性对照。此外,用完整的Fab-Strep-Tactin复合物刺激细胞后接着注射D-生物素(在t=140s近激活峰处),导致αCD3 Fab多聚体信号传导的突然中断(中图)。将离子霉素注射入预解离的Fab复合物群作为阳性对照。数据代表三不同的实验。重要的是,图5B示出了加入样品中的D-生物素迅速将Fab片段从Strep-tactin多聚化剂中置换出来,从而有效地终止钙释放,即便是在正在进行的钙刺激下,并且证明了解离的OKT3 Fab片段不再具有生物学活性。同样地,在将Streptactin-OKT3 Fab样品加入Jurkat细胞之前,将生物素添加到Strep-tactin-OKT3 Fab片段多聚体中,多聚OKT3 Fab片段也无法触发钙释放。
实施例3:经CD3 Fab多聚体进行的细胞可逆染色
本实施例检测了经CD3 Fab多聚体对细胞进行的可逆染色。新鲜分离的PBMC用αCD3单克隆抗体克隆OKT3(左侧散点图,Fab多聚体的亲本克隆)或同源藻红蛋白(PE)标记的OKT3 Fab多聚体进行染色,并在D-生物素处理之前(左二列)或之后(中间列)进行分析。在后续的清洗步骤后,使用新鲜PE标记的Strep-(右二列)检测剩余的Fab单体。对经可逆染色的细胞进行二次Fab多聚体染色作为对照(右列)。只有在用于区分活/死的碘化丙啶(propidium iodide PI)染色中为阴性的活CD3细胞在图6中示出。散点图中的数字表示栅极内的细胞百分比。这个实验表明,用与多聚化剂Streptactin多聚化的抗CD3 Fab片段对CD3+ PBMC进行的染色通过加入D-生物素完全可逆,并且单独的单价Fab片段无法与存在于PBMC上的CD3分子结合。
实施例4:通过CD28 Fab多聚体进行的细胞可逆分离
本实施例示出了通过与Strep-磁性颗粒(磁性微粒购自IBA GmbHGermany)多聚化的抗CD28 Fab片段的可逆结合进行细胞分离。为此目的,使用了来源于上文实施例1所述的抗体CD28.3的Fab片段。通过Fab多聚体磁性细胞选择法将CD28+细胞从新鲜分离的PMBC中选择/分离出来,基本上正如国际专利申请WO2013/011011中所述。在选择之前,用同源荧光αCD28多聚体(左侧散点图)或用直接抗免疫球蛋白κ轻链的抗体(左二散点图,α-IgκmAb)对细胞进行对照染色作为染色对照。在选择之后,细胞用D-生物素处理,随后清洗去除磁珠和Fab单体。被释放CD28+细胞随后用CD28 Fab多聚体(右二散点图)或用α-IgκmAb(右侧散点图)(重新)进行染色,以检测可能残余的Fab单体。只有活的(PI阴性)CD3+细胞被显示出来。散点图中的数字表示栅极内的细胞百分比。图7表明使用这种多聚抗CD28 Fab片段可以将CD28+细胞能够从PMBC中分离出来,并且所有分离试剂包括抗CD28 Fab单体可以在选择后被去除。
实施例5:用可逆固定于可溶性Strep-tactin上的αCD3/αCD28 Fab片段刺激/扩增CD3+ T应答细胞
在本实施例中,CD3+ T应答细胞(通过磁性选择法从由聚蔗糖梯度得到的新鲜PBMC样品中分离的)在体外被可逆固定于充当可溶性多聚化剂的可溶性寡聚Strep-上的αCD3/αCD28 Fab片段刺激后扩增。寡聚Strep-是根据制造商(Thermo Scientific)的方法流程,通过使Strep-与磺基SMCC(磺基琥珀酰亚胺基4-(N-马来酰亚胺甲基)环己烷-1-羧酸酯,产品#22122,Thermo Scientific)与亚氨基噻吩(iminothiolane)(产品#26101,Thermo Scientific)聚合而获得的。寡聚链霉亲和素可通过体积排阻色谱法与单体(未反应的)和二聚链霉亲和素突变蛋白分离,并将如此得到的寡聚链霉亲和素突变蛋白(n≥3)部分用作可溶性多聚化剂。
为了体外扩增,300.000个CD3+应答T细胞(Tresp)用2μM羟基荧光素琥珀酰亚胺酯(Carboxyfluorescein succinimidyl ester,CFSE)标记,并用不同量的可溶性Strep-寡聚物制剂进行刺激,所述可溶性Strep-寡聚物制剂上固定有上述αCD3OKT3 Fab片段和抗体28.3的αCD28 Fab片段(两者在重链上都携带上述作为链霉亲和素结合肽的双Strep标记(Twin-Strep-))的组合(“1x”对应于被0.5μgαCD3-和0.5μgαCD28单体Fab片段功能化的3μg多聚化Streptactin,数字“0.5x”、“2x”和“5x”表示分别为“1x”的n倍量)。剩余的未被刺激的Tresp细胞或用空白Strep-tactin多聚体(无Fab)刺激的Tresp细胞作为阴性对照。将Tresp细胞连同300.000个CD3阴性自饲养细胞(经30Gy辐照)一式两份接种到48孔板上1ml添加有20U/ml IL-2的细胞培养基中。细胞在37℃下孵育,不更换培养基,并在5天后通过FACS分析法按照CFSE稀释度来对增殖情况进行分析。图8A示出了在5天后与阴性对照相比培养物中增殖细胞的大小分布增加。图8B示出了当与其上固定有αCD3 Fab和αCD28 Fab片段混合物的可溶性寡聚Strep-(与图4中的固体Streptactin磁性颗粒相比)一起孵育时,CD3+Tresp细胞被适当地刺激并剧增殖。图8A和图8B的结果表明,在这些体外条件下,大多数CD3+ T应答细胞在表面CD28和TCR/CD3复合物与可逆固定于可溶性Strep-寡聚物上的αCD3和αCD28 Fab片段结合之后进行分裂(2至5次细胞分裂)。在体外扩增后,可溶性Fab-Strep-Tactin刺激试剂在D-生物素处理后被解离并去除。通过用藻红蛋白标记的Strep-(ST-PE)对细胞重新染色而对单体Fab片段的解离和去除进行流式细胞分析。与适当的仅用ST-PE标记的阴性对照(浅灰色直方图)相比,代表性的直方图(深灰色直方图)被示出。从图8C中可以看出,两种Fab片段都已经完全地解离并已从扩增的细胞中完全去除。图8D示出了5天后活(台盼蓝阴性)细胞的绝对数。该绝对数通过使用Neubauer计数室计数得到,并按各自的刺激条件绘图。中位细胞数在图8D中示出;误差线表示标准偏差(SD)。图8D表明,所有固定于可溶性Strep-tactin多聚化剂上的αCD3 Fab片段和αCD28 Fab片段的混合物在扩增CD3+细胞上同样有效,并导致绝对细胞数的约4倍增长。
实施例6:在体外被可逆的aCD3/aCD28 Fab-Streptamer多聚体刺激的纯化CD4+和CD8+ T应答细胞增殖(不更换培养基)的动力学
本实施例中检测了在体外被可逆固定于可溶性寡聚链霉亲和素突变蛋白上的αCD3/αCD28 Fab片段刺激的纯化CD4+和CD8+ T应答细胞(Tresp)增殖的扩增动力学。为此,两种不同大小的可溶性寡聚Strep-突变蛋白被用作可溶性多聚化剂。第一种寡聚Strep-是在实施例5中获得的寡聚链霉亲和素突变蛋白的一部分(n≥3)(也被称为“常规骨架”,如图13中尖端朝下的三角形符号所示)。用作可溶性多聚化剂第二种寡聚链霉亲和素突变蛋白是与生物素化的人血清白蛋白反应的寡聚链霉亲和素突变蛋白(n≥3)(也被称为“大骨架”)。
在本实施例中,分别用如上所述的这两种不同的Streptamer多聚体,即实施例5所述的Streptactin骨架(使用浓度为1mg寡聚链霉亲和素突变蛋白/ml的溶液)或大Streptactin骨架(0.1mg/ml)刺激500.000个纯化的CD4+或CD8+应答T细胞(Tresp)。3μl两种不同的骨架都负载有前述实施例中使用的0.5μgαCD3和0.5μgαCD28 Fab的组合,其在Fab片段重链的C端携带链霉亲和素结合肽SAWSHPQFEK(GGGS)2GGSAWSHPQFEK(SEQ ID NO:07)。此外,4.5μl常规Streptactin骨架负载有0.5μgαCD3 Fab片段、0.5μgαCD8 Fab片段(IBA GmbH在其Fab片段的C端也携带有链霉亲和素结合肽SAWSHPQFEK(GGGS)2GGSAWSHPQFEK(SEQ ID NO:07))和0.5μgαCD28 Fab片段。未经处理的(未刺激的)Tresp细胞作为阴性对照,经市售Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)刺激的Tresp细胞作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基(RPMI 1640(Gibco),添加有10%(v/v)胎牛血清、0.025%(w/v)L-谷氨酰胺、0.025%(w/v)L-精氨酸、0.1%(w/v)HEPES、0.001%(w/v)庆大霉素、0.002%(w/v)链霉素、0.002%(w/v)青霉素)中。细胞在37℃下孵育,不更换培养基,并在1、3和6天后进行细胞计数分析。在图13的实验中,扩增在不更换培养基的情况下进行。CD4+ T应答细胞的结果在图13A中示出,CD8+ T应答细胞的结果在图13B中示出,其中曲线图代表据每个时间点收集的CD4+ Tresp(图13A)和CD8+ Tresp(图13B)的细胞数所示的增殖程度。
从图13A中可以看出,所述其上可逆固定有αCD3和αCD28 Fab片段的“较小”可溶性多聚化剂提供了与Dynabeads(其是目前为止用于扩增T细胞的标准试剂)相同量的CD4+ T细胞的扩增,而所述“较大”寡聚可溶性Streptactin提供了与Dynabeads相比甚至更好的扩增。这种改善可能是由以下导致的:可溶性“较大寡聚多聚化剂”能够同时结合比“较小”可溶性寡聚物更多的T细胞,由此能够比“较小”寡聚物刺激更多的CD4+ T细胞。
从图13B中可以明显看出,使用本发明的可溶性多聚化剂,CD8+ T细胞可以在最初的3天内至少与Dynabeads一样有效地扩增。值得注意的是,在这个时间段中,使用了除αCD3和αCD28 Fab片段(作为第一和第二试剂)以外还携带可逆固定于其上的αCD8 Fab片段的可溶性多聚化剂的扩增实验在这些培养条件下显示出了最佳的扩增程度。这表明有可能通过使用对特定亚细胞群具有特异性的刺激物(这里是αCD8 Fab片段)来提高或调整扩增的选择性,从而能获得大量所需的(亚)细胞群。
因此,综上所述,实施例6表明本发明所用的可溶性多聚化剂的功能就触发T细胞扩增而言,与用于此目的的、使用Dynabeads的现有标准方法相当。然而,由于可通过加入竞争剂控制(并且终止,如果需要)刺激,诸如在第一和第二试剂与多聚化剂之间基于链霉亲和素的可逆相互作用的情况下加入生物素,因此本发明提供了显著优于Dynabeads技术的优势,因为扩增条件可进行优化(例如可以在3天后停止图13B实验中的刺激)。此外,由于可溶性多聚化剂可以很容易地从反应中去除(例如,在扩增反应后,通过将试剂固定在生物素化的柱子上),本发明的扩增方法可在封闭系统中实施并自动化,而不必处理磁珠诸如Dynabeads的去除问题,该封闭系统例如是用于治疗目的的细胞的GMP生产所需的。
实施例7:在体外被可逆的aCD3/aCD28 Fab-Streptamer多聚体刺激的纯化CD4+和CD8+ T应答细胞增殖(更换培养基)的动力学
本实施例中也检测了在体外被可逆固定于可溶性寡聚链霉亲和素突变蛋白上的αCD3/αCD28 Fab片段刺激的纯化CD4+和CD8+ T应答细胞(Tresp)增殖的扩增动力学。为此,两种不同大小的可溶性寡聚Strep-突变蛋白被用作可溶性多聚化剂。第一种寡聚Strep-是在实施例5中获得的寡聚链霉亲和素突变蛋白的一部分(n≥3)(也被称为“常规骨架”,如图13中以尖端朝下的三角形符号所示)。被用作可溶性多聚化剂的第二种寡聚链霉亲和素突变蛋白通过使实施例5中获得的寡聚Strep-tactin(n≥3)与生物素化的人血清白蛋白发生反应而获得。这种可溶性寡聚多聚化剂也被称为“大骨架”。
在本实施例中,分别用如上所述的这两种不同的Streptamer多聚体,即用实施例5所述的Streptactin骨架(1.0mg/ml)或用大Streptactin骨架(0.1mg/ml)刺激400.000个纯化的CD4+或CD8+应答T细胞(Tresp)。3μl两种不同的骨架都负载有如上所述的0.5μgαCD3和0.5μgαCD28 Fab片段的组合。此外,4.5μl实施例5所述的Streptactin骨架负载有0.5μgαCD3 Fab片段、0.5μgαCD8 Fab片段和0.5μgαCD28 Fab片段,如上文所述。未经处理的(未刺激的)Tresp细胞作为阴性对照,用Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体)刺激的Tresp细胞作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,在第3天更换培养基,并在1、3和6天后进行细胞计数分析。CD4+ T应答细胞的结果在图14A中示出,CD8+ T应答细胞的结果在图14B中示出,其中曲线图代表了据每个时间点收集的CD4+ Tresp(图14A)和CD8+ Tresp(图14B)的细胞数所示的增殖程度。
从图14A中可以看出,本发明的其上可逆固定有αCD3和αCD28 Fab片段的可溶性多聚化剂提供了比Dynabeads更好的CD4+ T细胞的扩增。
从图14B中可以明显看出,使用本发明的可溶性多聚化剂,CD8+ T细胞可以在最初6天内至少与Dynabeads一样有效地扩增。值得注意的是,在这个时间段中,使用携带有αCD3和αCD28 Fab片段(作为第一和第二试剂)的较大可溶性多聚化剂的扩增实验在这些培养条件下显现出了最佳的扩增程度。这可能也是由以下导致的:可溶性“较大寡聚多聚化剂”能够同时结合比“较小”可溶性寡聚物更多的T细胞,由此能够比“较小”寡聚物刺激更多的CD4+ T细胞。
实施例8:更换或不更换培养基情况下纯化CD4+和CD8+ T细胞培养物的扩增动力学
在本实施例中,将实施例6和实施例7中关于“较小”可溶性多聚化剂以及阳性和阴性对照的的综合数据用输入的细胞数归一化。并未获取关于“较大”多聚化剂的归一化数据。如实施例6和实施例7中所释,400.000至500.000个CD4+或CD8+应答T细胞(Tresp)被3μl的Streptactin多聚体制剂(1mg/ml;其上固定有0.5μgαCD3 Fab片段和0.5μgαCD28 Fab片段)刺激。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads刺激的Tresp细胞作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。Tresp细胞被一式两份接种在48孔板中1ml补充以30U/ml IL-2的细胞培养基上。细胞在37℃下孵育,在第3天更换培养基(图15中的直线)或不更换培养基(图15中的虚线),并在1、3和6天后进行细胞计数分析。从图15A的归一化数据可以看出,可逆固定有αCD3和αCD28 Fab片段的“较小”可溶性多聚化剂产生了约2.5倍的CD4+T细胞扩增,而使用Dynabeads的扩增产生了约1.8倍的扩增率。因此,使用本发明的可溶性多聚化剂甚至还提供了在CD4+ T细胞扩增方面超过Dynabeads的改善。类似地,图15B确认了通过使用本发明的可溶性多聚化剂,CD8+ T细胞可以在最初3天内至少与使用Dynabeads一样有效地扩增。
实施例9:在体外被可逆的aCD3/aCD28 Fab Streptamer多聚体刺激的纯化CD4+和CD8+ T应答细胞激活后的早期群簇形成
在本实施例中,用3μl负载有0.5μgαCD3-和0.5μgαCD28 Fab组合的寡聚Streptactin多聚化剂制剂(1mg/ml)刺激400.000个CD4+或CD8+应答T细胞(Tresp)。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads刺激的Tresp细胞作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育并在1天和2天后通过显微镜进行分析。CD4+ Tresp(图16A)和CD8+ Tresp(图16B)针对Dynabeads(中间一行)和Streptamer多聚体(下边一行)的刺激被分别示出。照片表示群簇形成的程度:为了更好的可视性,在图16A和图16B中用圆圈标出了经可溶性链霉亲和素突变蛋白寡聚物刺激的示例性群簇。Dynabeads刺激中的群簇经深色刺激颗粒的积聚变得明显易见。显然,对于CD4+和CD8+ T细胞两者而言,当使用本发明的采用可溶性寡聚多聚化剂的扩增方法时,早期群簇形成。
实施例10:多克隆刺激/扩增的大量CD3+中枢记忆T细胞(Tcm)的扩增动力学和表型
在本实施例中,用3μl负载有0.5μgαCD3和0.5μgαCD28 Fab组合的如实施例5所述的可溶性寡聚Streptactin的制剂(1mg/ml)刺激500.000个CD3+CD62L+CD45RA应答Tcm细胞(Tresp)。此外,使用4.5μl负载有0.5μgαCD3、0.5μgαCD8 Fab和0.5μgαCD28 Fab的Streptactin多聚体制剂作为额外的刺激条件。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体)刺激的Tresp细胞作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml仅添加有30U/ml IL-2或添加有30U/ml IL-2和5ng/ml IL-15的细胞培养基上。细胞在37℃下孵育,每3天更换一次培养基,并在7天和14天后进行细胞计数分析。曲线图代表了据每个时间点收获的细胞数所示的扩增程度,图17A中为仅添加有IL-2的培养基,图17B中为添加有IL-2和IL-15的培养基。在图17A和图17B两图中可以看出,具有可逆结合于其上的CD3 Fab片段和αCD28 Fab片段的可溶性多聚化剂产生了比Dynabeads更好的细胞扩增。如图17C中对在可变细胞因子环境中培养14天后的CD62L和CD127表面表达水平进行的流式细胞分析进一步所示,使用本发明的可溶性多聚化剂的实验方法,在此选择的两种条件下,保持着比使用Dynabeads的扩增更高含量的CD127表达型长期存活的记忆T细胞。这阐明了本发明方法的又一个优势。
实施例11:从大量CD3+中枢记忆T细胞中对Tcm应答细胞进行的选择性抗原特异性扩增(动力学和表型)
本实施例中检测了从纯化的CD3+CD62L+CD45RA-Tcm应答细胞中进行的选择性抗原特异性(Ag特异性)扩增的动力学和表型。
更详细地,在体外用肽:MHC分子复合物(作为向细胞提供初级激活信号的第一试剂)和αCD28 Fab片段(作为刺激细胞表面辅助分子的第二试剂)两者刺激CD3+CD62L+CD45RA-Tcm应答细胞。抗原特异性肽与MHC分子的复合物和αCD28 Fab片段两者均被可逆地固定在实施例5所述的可溶性寡聚链霉亲和素突变蛋白(n≥3)上。用于抗原特异性扩增的肽为肽CRVLCCYVL(SEQ ID NO:06),即刻早期蛋白1的第309-317位氨基酸(如Ameres等人,PLOS Pathogens,May 2013,vol.9,issue 5,e1003383所述),其呈现对巨细胞病毒(CMV)具有特异性的HLA-C7/IE-1抗原表位。呈递所述肽的MHC I分子在α链(重链)的C端携带链霉亲和素结合肽(SAWSHPQFEK(GGGS)2GGSAWSHPQFEK,(SEQ ID NO:07),可以“双Strep标记(Twin-Strep-)”购于IBA GmbH,Germany)。
为此,使用3μl被0.5μg配有链霉亲和素结合肽的肽:MHC I类复合物和0.5μg上述αCD28 Fab所功能化的可溶性寡聚Streptactin多聚化剂制剂对500.000个CD3+CD62L+CD45RA应答Tcm细胞(Tresp)进行Ag特异性刺激。作为替代,4.5μl Streptactin多聚化剂制剂负载有0.5μg这些肽:MHC I类复合物、0.5μg CD8αFab和0.5μgαCD28 Fab。为了比较,使用3μl负载有0.5μgαCD3 Fab和0.5μgαCD28 Fab组合的Streptactin多聚化剂制剂(1mg/ml)进行多克隆刺激。同样作为上述刺激条件的替代,使用4.5μl可逆负载有0.5μgαCD3 Fab、0.5μgαCD8 Fab和0.5μgαCD28 Fab的Streptactin多聚化剂制剂。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)多克隆刺激的Tresp细胞作为阳性对照。将Tresp细胞接种到48孔板上1ml添加有30U/ml IL-2和5ng/ml IL-15的细胞培养基中。细胞在37℃下孵育,每3天更换一次培养基,并在7天和14天后进行细胞计数分析。对经可溶性strept-tactin寡聚物刺激/扩增的Ag特异性细胞的一部分进行示例性流式细胞分析(图18A),其中所述可溶性strept-tactin寡聚物上固定有含HLA-C7/IE-1抗原表位(针对CMV)的肽:MHC-I复合物,表明这些抗原特异性T细胞被特异性扩增。图18B至图18E的曲线图(其代表据每个时间点收获的肽:MHCI多聚体阳性细胞的细胞数所示的不同Ag特异性的扩增程度,类似于图18A中所示的扩增实验)表明,使用相应Ag特异性肽和MHC I分子复合物的多聚化剂提供了最多数目的扩增细胞(范围从对于识别被HLA-A2402限制的CMV的pp65抗原表位(第341-350位氨基酸(QYDPVAALF,(SEQ ID NO:08))的Ag特异性细胞的细胞数的20倍增长(参见图18B)到识别CMV的HLA-B7/IE-1309-317抗原表位(CRVLCCYVL(SEQ ID NO:06))的Ag特异性细胞数的98倍增长(参见图18E)),由此表明本发明的扩增方法完全适用于Ag特异性细胞的扩增。最后,图18F中示出了HLA-B7/Hexon5抗原表位(针对腺病毒)培养14天后对CD62L和CD127表面表达水平进行的示例性流式细胞分析,其进一步确认了使用本发明可溶性多聚化剂的实验方法,在多克隆且Ag特异性刺激的条件下,保持了更高含量的CD127表达型长期存活的记忆T细胞。
实施例12:大量中枢记忆T细胞的选择性Ag特异性扩增动力学和表型
本实施例检测了从使用可逆固定于可溶性寡聚链霉亲和素突变蛋白上的、作为第一和第二试剂的a)抗原特异性肽MHC I复合物和b)αCD28 Fab片段体外刺激的纯化CD3+CD62L+CD45RA-Tcm应答细胞中的选择性Ag抗原特异性扩增的动力学。
为此,使用3μl被0.5μg配有链霉亲和素结合肽(该特异性肽代表被HLA-B07限制的腺病毒Hexon 5蛋白的第 114-124位氨基酸(CPYSGTAYNSL,SEQ ID NO:10))的肽:MHC I类复合物和0.5μgαCD28 Fab所功能化的Streptactin多聚化剂制剂对500.000个CD3+CD62L+CD45RA应答Tcm细胞(Tresp)进行Ag特异性刺激。作为替代,4.5μl Streptactin多聚化剂制剂负载有0.5μg这种肽:MHC I类复合物、0.5μgαCD8 Fab和0.5μgαCD28 Fab。为了比较,使用3μl负载有0.5μgαCD3 Fab和0.5μgαCD28 Fab组合的Streptactin多聚化剂制剂(1mg/ml)进行多克隆刺激。同样作为上述刺激条件的替代,使用4.5μl负载有0.5μgαCD3 Fab、0.5μgαCD8 Fab和0.5μgαCD28 Fab的Streptactin多聚化剂制剂。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads多克隆刺激的Tresp细胞作为阳性对照。将Tresp细胞接种到48孔板上1ml添加有30U/ml IL-2和5ng/ml IL-15的细胞培养基中。细胞在37℃下孵育,每3天更换一次培养基经历,并在7天和14天后进行细胞计数分析。图19中示出的照片代表了在第5天时群簇形成的程度,示出了示例性的针对腺病毒的HLA-B7/Hexon 5抗原表位的Ag特异性刺激。从图19中可以看出,这样的腺病毒抗原特异性细胞可从最初的CD3+CD62L+CD45RA-Tcm应答细胞群中被特异性地扩增出来。
实施例13:扩增的CD8+ T细胞的产量和表型——可溶性多聚化剂的大小变化和用于刺激的αCD8-Fab的加入
本实施例检测了在体外被可逆固定于可溶性寡聚链霉亲和素突变蛋白上αCD3/αCD28 Fab片段刺激的纯化CD8+ T应答细胞的扩增。此外,本实施例还检测了多聚化剂中添加αCD8-Fab对提高CD8+ T细胞扩增特异性的影响。
为此,分别使用两种不同的基于Streptactin的多聚化剂,即实施例5所述的小寡聚Streptactin多聚化剂(1mg/ml)或上述较大Streptactin寡聚物(0.1mg/ml)刺激300.000个纯化的CD8+应答T细胞(Tresp)。3μl的两种不同的多聚化剂(骨架)均负载有上述0.5μgαCD3和0.5μgαCD28 Fab片段组合。此外,4.5μl较小Streptactin多聚化剂(骨架)负载有上述的0.5μgαCD3、0.5μgαCD8 Fab和0.5μgαCD28 Fab片段。此外,使用3μl仅被0.5μgαCD3 Fab片段单独功能化或0.5μgαCD28 Fab片段单独功能化的“较小”Streptactin多聚化剂(骨架)。未经刺激的Tresp细胞作为阴性对照,经Dynabeads刺激的Tresp作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,每3天更换一次培养基,并在6天后进行分析。图20A示出了据第6天收获的细胞数所示的增殖程度,其中所述细胞数与阴性对照相比较,并以阳性对照进行归一化。图20A示出了使用本发明的可溶性多聚化剂的CD8+ T细胞扩增比使用Dynabeads扩增产生了更高产量的CD8+ T细胞。在细胞培养后CD8表面表达水平(图20B)和CD45RO表面表达水平(图20C)的FACS分析表明,本发明的多聚化剂或Dynabeads扩增了相同表型的CD8+T细胞(使用单因素ANOVA分析对不同的刺激条件进行比较,并未检测到显著性差异(n.s.))。与使用Dynabeads相比,使用本发明扩增方法的CD8+细胞产量的提高可能是由于可溶性多聚化剂比固定在Dynabeads上的抗体可以更好地接近其位于细胞表面的靶受体。当从初始样品中扩增稀少的细胞群时,这种产量提高会变得十分有利。
此外,将使用其上联合固定有0.5μgαCD3和0.5μgαCD28 Fab片段两者的多聚化剂所获得的扩增产量(图20B中的左二列)与使用两种仅被αCD3 Fab片段单独功能化或αCD28 Fab片段单独功能化的多聚化剂所获得的扩增产量(图20B中左三列)进行比较,可以看出两个实验具有相同的扩增效率。因此,这些实验表明使用一种其上联合固定有第一试剂和第二试剂两者的多聚化剂在用于扩增的功能上等同于使用两种分别仅负载有第一试剂或第二试剂的单独多聚化剂。
实施例14:扩增的CD8+ T细胞的产量和表型——其上固定有不同比例的αCD3-和αCD28 Fab片段的单独可溶性多聚化剂的滴定
本实施例中检测了在体外被片段以不同量可逆固定于可溶性寡聚链霉亲和素突变蛋白上的αCD3/αCD28 Fab片段刺激的扩增CD8+ T应答细胞(Tresp)的产量和表型。
为此,使用不同量的被αCD3 Fab单独功能化或αCD28 Fab单独功能化的“小”寡聚Streptactin多聚化剂制剂(1mg/ml)的混合物(“1x”对应于被0.5μgαCD3单独功能化的1.5μg Streptactin多聚化剂和被0.5μgαCD28 Fab片段单独功能化的1.5μg多聚化Streptactin)、或3μl负载有0.5μgαCD3和0.5μgαCD28 Fab的Streptactin多聚化剂制剂、或4.5μl负载有0.5μgαCD3、0.5μg strep-标记的αCD8和0.5μgαCD28 Fab的Streptactin多聚化剂制剂对300.000个CD8+应答T细胞(Tresp)进行刺激。未经处理的Tresp细胞作为阴性对照,经Dynabeads刺激的Tresp作为阳性对照。将Tresp细胞接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,不更换培养基,并在5天后进行分析。图21A示出了据第6天收获的细胞数所示的增殖程度,其中,所述细胞数与与阴性对照相比较,并以阳性对照进行归一化。图21A表明使用本发明的不同可溶性多聚化剂的CD8+T细胞扩增比使用Dynabeads的扩增产生了更高产量的CD8+T细胞(特别是5x条件中的累计试剂总量导致细胞的最佳扩增,特别是随着时间的推移,或者随着由开始细胞分裂引起的细胞总数的增加)。在细胞培养后CD8表面表达水平(图21B)和CD45RO表面表达水平(图21C)的FACS分析表明,本发明的不同多聚化剂或市售Dynabeads扩增了相同表型的CD8+ T细胞。
实施例15:Streptamer多聚体刺激aCD19-CAR转导的Jurkat细胞后胞内信号级联的激活
本实施例检测了转导的Jurkat细胞胞内信号级联的激活,所述Jurkat细胞被修饰以表达肿瘤特异性嵌合抗原受体(CAR)即这里的CD19,并使用作为可溶性多聚化剂的如实施例5所述的寡聚Strep-进行刺激。
为此,用以下试剂对300.000个JurkaT应答细胞(Jresp)进行刺激:(A)不同量的被本文所述的αCD3 Fab和αCD28 Fab片段功能化的Streptactin多聚化剂制剂(1mg/ml)的混合物(“x1”对应于被0.5μgαCD3-和0.5μgαCD28 Fab功能化的3μg Streptactin多聚化剂——这提供了“基于Streptactin的多克隆多聚化剂”),或(B)3μl被0.5μg(x1)或1μg(x2)CD19胞外域(ECD)功能化的Streptactin多聚化剂制剂(CD19胞外域是αCD19-CAR的天然配体——这提供了“基于Streptactin的CAR特异性多聚化剂”),或3μl负载有0.5μg(x1)或1μg(x2)αIgG的Streptactin多聚化剂制剂(αIgG识别αCD19-CAR中的IgG4间隔区——这也提供了“基于链霉亲和素突变蛋白的CAR特异性多聚化剂)。装配有六组氨酸标记物的CD19的ECD从Sino Biological/Life technologies(SEQ ID NO:27)获得,并通过将CD19的ECD与衔接分子His-STREPPER(IBA GmbH,Germany,订单编号2-0920-005)按分子比率1:1混合并于室温下孵育15分钟而被功能化成可与基于链霉亲和素的多聚化剂结合。该His-STREPPER衔接分子含有与六组氨酸标记物结合的螯合部分和链霉亲和素结合肽,从而临时性提供靶分子,在此CD19的ECD具有可与基于链霉亲和素突变蛋白的多聚化剂可逆结合的链霉亲和素结合肽。用Dynabeads(具有不可逆固定于其上的αCD3-和αCD28-单克隆抗体的磁珠)或用PMA和离子霉素刺激的Jresp作为阳性对照。将Jresp细胞接种到1.5ml Eppendorf离心管的200μl添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,并在刺激0至20分钟后置于冰上进行裂解。检测到磷酸化的ERK表明MAPK信号传导活跃,对管家β-肌动蛋白的染色表明在每种条件下和在每个时间点时上样的总蛋白量相同。图22A示出了Jurkat细胞经“多克隆Streptactin多聚化剂”的激活情况,图22B示出了Jurkat细胞经两种“基于Streptactin的CAR特异性多聚化剂”的激活情况,从图22A和图22B的比较中可以看出,Jurkat细胞可以通过CD19胞外域与CD19特异性嵌合抗原受体结合而被激活/扩增。由于对T细胞的基因下游加工几乎仅仅在预先选定的细胞群中进行,因此,所引入的CAR经IgG4间隔区结构域(在各种具有不同特异性的CAR内是保守的)发生交联引起的一般性激活拓宽了可逆细胞刺激/扩增在这些体外细胞处理情况下的适用性。
因此,这个实验表明,原则上任何通过向细胞群提供初级激活信号的试剂(配体)的结合而被激活的细胞群都可以使用如本文所述的可逆固定在多聚化剂上的第一试剂进行扩增。
实施例16:加入αCD8-Fab刺激扩增的CD3+ T细胞的产量和亚群组成
本实验示出了纯化的CD3+ T应答细胞在体外经可逆固定于实施例5所述作为可溶性多聚化剂的可溶性寡聚Strep-上的αCD3/αCD28 Fab片段刺激的扩增情况。在一项实验中,除了αCD3/αCD28 Fab片段以外,从IBA GmbH,Germany购得的αCD8 Fab片段(目录编号6-8000-203)可被固定在链霉亲和素突变蛋白的可溶性寡聚物上,以检测是否可能在体外用可逆的αCD3/αCD28 Fab-Streptamer多聚体优先刺激特定的T细胞亚群。更详细地,用3μl负载有0.5μgαCD3和0.5μgαCD28 Fab组合的寡聚链霉亲和素制剂(1mg/ml)对500.000个纯化的CD3+应答T细胞(Tresp)进行刺激。作为一种替代的方法,4.5μl Streptactin寡聚物负载有0.5μgαCD3、0.5μg strep-标记的αCD8 Fab和0.5μg strep-标记的αCD28 Fab。未经刺激的Tresp细胞作为阴性对照,经Dynabeads(其上不可逆固定有αCD3和αCD28单克隆抗体的磁珠)刺激的Tresp细胞作为阳性对照。从图23A中可以看出,可逆负载有αCD3 Fab片段、αCD28 Fab片段以及还有αCD8 Fab片段的多聚化剂提供了最高数量的扩增CD3+ T细胞。扩增细胞数为1.1x106的产量比使用市售Dynabeads对这些T细胞进行扩增高约30%。此外更重要的是,如图23B所示,与使用Dynabeads或本发明的如本文所述仅携带αCD3 Fab片段和αCD28 Fab作为第一和第二试剂的可溶性多聚化剂进行的两种扩增相比,使用携带αCD3 Fab片段、αCD28 Fab片段和αCD8 Fab片段的多聚化剂的CD8+ T细胞量是最高的。因此,本实验还表明了本发明的优势在于除了向所期望的细胞群提供初级激活信号的第一试剂以及可选地提供共刺激信号的第二试剂以外,另一种用于特异性激活所期望细胞群的试剂也可以固定在多聚化剂上。因此,通过这样做,本发明提供了从例如包含各种不同亚群的样品中优先扩增或选择性富集任何所期望的(亚)细胞群的可能性。
实施例17:从单个细胞池中对Tcm应答细胞进行的平行抗原特异性扩增
本实施例检测了从在体外经多种可逆的肽:MHC/αCH28 Fab Streptamer多聚体刺激的T应答细胞单个细胞中进行的平行抗原特异性(Ag特异性)扩增的动力学。
使用3μl被相应的携带有链霉亲和素结合肽的肽:MHC I类复合物以及0.5μg也携带有链霉亲和素结合肽的αCD28 Fab所功能化的Streptactin多聚体,对500.000个CD3+CD62L+CD45RA应答Tcm细胞(Tresp)同时进行用于每种特异性的多重特异性的刺激。作为一种替代方法,对每种特异性使用4.5μl被如本文所述的0.5μg携带有链霉亲和素结合肽的肽:MHC I、0.5μgαCD8 Fab和0.5μgαCD28 Fab所功能化的基于Streptactin的多聚化剂。为了比较,使用3μl可逆负载有0.5μgαCD3 Fab和0.5μgαCD28 Fab组合的基于Streptactin的多聚化剂制剂(1mg/ml)进行多克隆刺激。同样,作为上述刺激条件的替代,可以使用4.5μl可逆负载有0.5μgαCD3 Fab、0.5μgαCD8 Fab和0.5μgαCD28 Fab(其中每以个都携带有链霉亲和素结合肽)的基于Streptactin的多聚化剂制剂。未经处理的(未刺激的)Tresp细胞作为阴性对照,经Dynabeads(涂覆有αCD3-和αCD28-mAb的磁珠)多克隆刺激的Tresp细胞作为阳性对照。将Tresp细胞接种到48孔板上1ml添加有30U/ml IL-2和5ng/ml IL-15的细胞培养基中。细胞在37℃下孵育,每3天更换一次培养基,并在7天和14天后进行细胞计数分析。
实施例18:在体外用被αCD3/αCD8/αCD28 Fab片段可逆功能化的基于链霉亲和素的多聚化剂所刺激的CD3+T应答细胞中优选扩增CD8+T细胞
使用以下试剂对300.000个CD3+应答T细胞(Tresp)进行刺激:3μl负载有0.5μgαCD3和0.5μgαCD28 Fab组合的多聚化Streptactin制剂(1mg/ml)或使用大Streptactin骨架的多聚化剂制剂,或者4.5μl负载有0.5μgαCD3、0.5μgαCD8 Fab和0.5μgαCD28 Fab的基于Streptactin的多聚化剂制剂,或者3μl仅具有0.5μgαCD3 Fab或仅具有0.5μgαCD28 Fab的基于Streptactin的多聚化剂制剂的混合物(各Fab片段也携带链霉亲和素结合肽)。未经处理的Tresp细胞作为阴性对照,经Dynabeads(涂覆有αCD3-和αCD28-mAb的磁珠)刺激的Tresp作为阳性对照。将Tresp细胞一式两份接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,3天后更换培养基,6天后进行分析。
实施例19:在体外用被αCD3和αCD28 Fab片段可逆功能化的基于链霉亲和素的多聚化剂所刺激的CD3+T应答细胞中优选扩增CD8+T细胞
使用以下试剂对300.000个CD3+应答T细胞(Tresp)进行刺激:不同量的被αCD3 Fab片段单独功能化和被αCD28 Fab片段单独功能化的基于Streptactin的多聚化剂制剂(1mg/ml)的混合物(1.5μg基于Streptactin的多聚化剂被0.5μgαCD3 Fab单独功能化,1.5μg基于Streptactin的多聚化剂被0.5μgαCD28 Fab片段单独功能化),或者不同量的被αCD3 Fab片段和αCD28 Fab片段具有或不具有αCD8 Fab片段(各Fab片段也携带有链霉亲和素结合肽)功能化的基于Streptactin的多聚化剂制剂的混合物(3μg基于Streptactin的多聚化剂被0.5μgαCD3-和0.5μgαCD28 Fab片段功能化——无αCD8 Fab片段),或者4.5μl负载有0.5μgαCD3 Fab片段、0.5μgαCD8 Fab片段和0.5μgαCD28 Fab片段(其中Fab片段也携带有链霉亲和素结合肽)的Streptactin多聚化剂制剂。未经处理的Tresp细胞作为阴性对照,经Dynabeads(涂覆有αCD3-和αCD28-mAb的磁珠)刺激的Tresp作为阳性对照。将Tresp细胞接种到48孔板上1ml添加有30U/ml IL-2的细胞培养基中。细胞在37℃下孵育,3天后更换培养基,并在6天后进行分析。
在本说明书中列举或讨论在先出版的文献不应当地被视为承认该文献是现有技术的一部分或是公知常识。
本文示例性描述的本发明以适当地在缺乏本文未具体公开的任何一个或多个元件、一个或多个限制的情况下实施。因此,例如,术语“包括”、“包含”、“含有”等应该在广义上理解且没有限制。此外,本文使用的术语和表述被用作描述而非限制,并且在使用这些术语和表述时,没有意图排除所示和所述特征或其部分的任何等同物,而应认识到各种修改均可能在发明要求保护的范围内。因此,应当理解,尽管已经通过示例性实施方案和可选特征具体公开了本发明,但是本领域技术人员可以寻求对本文所公开和体现的本发明做出修改和变化,这样的修改和变化被认为在本发明的范围内。
本文已经全面且一般性地描述了本发明。属于一般公开内容中的每个更窄的种类和亚属组也构成本发明的一部分。这包括本发明的一般性描述,其具有从该属中去除任何主题的附带条件或否定限制,无论所排除的材料是否在本文有具体描述。
其他实施方权利要求范围内。此外,在本发明的特征或方面以马库什组的方式进行描述时,本领域技术人员将认识到本发明也由此以马库什组的任何个体成员或成员亚组的形式进行描述。
序列表
<110> 朱诺治疗有限公司
<120> 用于扩增细胞群的方法、试剂盒及装置
<130> LC16310021P
<150> US 61/980,506
<151> 2014-04-16
<160> 27
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> PRT
<213> 人工
<220>
<223> Strep-标记
<400> 1
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 2
<211> 4
<212> PRT
<213> 人工
<220>
<223> 链霉亲和素突变蛋白 (类似物) 1
<400> 2
Val Thr Ala Arg
1
<210> 3
<211> 4
<212> PRT
<213> 人工
<220>
<223> 链霉亲和素突变蛋白 (类似物) 2
<400> 3
Ile Gly Ala Arg
1
<210> 4
<211> 28
<212> PRT
<213> 人工
<220>
<223> di-标记3
<400> 4
Trp Ser His Pro Gln Phe Glu Lys Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser Trp Ser His Pro Gln Phe Glu Lys
20 25
<210> 5
<211> 24
<212> PRT
<213> 人工
<220>
<223> di-标记2
<400> 5
Trp Ser His Pro Gln Phe Glu Lys Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Trp Ser His Pro Gln Phe Glu Lys
20
<210> 6
<211> 9
<212> PRT
<213> 人工
<220>
<223> 即刻早期1蛋白 (第309-317位氨基酸)
<400> 6
Cys Arg Val Leu Cys Cys Tyr Val Leu
1 5
<210> 7
<211> 30
<212> PRT
<213> 人工
<220>
<223> 双Strep-标记
<400> 7
Ser Ala Trp Ser His Pro Gln Phe Glu Lys Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Ser Gly Gly Ser Ala Trp Ser His Pro Gln Phe Glu Lys
20 25 30
<210> 8
<211> 9
<212> PRT
<213> 人工
<220>
<223> CMV的pp65抗原表位 (第341-350位氨基酸)
<400> 8
Gln Tyr Asp Pro Val Ala Ala Leu Phe
1 5
<210> 9
<211> 11
<212> PRT
<213> 人工
<220>
<223> CMV的pp65抗原表位 (第265-274位氨基酸)
<400> 9
Arg Pro His Glu Arg Asn Gly Phe Thr Val Leu
1 5 10
<210> 10
<211> 11
<212> PRT
<213> 人工
<220>
<223> 腺病毒的hexon5抗原表位 (第114-124位氨基酸)
<400> 10
Lys Pro Tyr Ser Gly Thr Ala Tyr Asn Ala Leu
1 5 10
<210> 11
<211> 9
<212> PRT
<213> 人工
<220>
<223> HA-标记
<400> 11
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
1 5
<210> 12
<211> 11
<212> PRT
<213> 人工
<220>
<223> VSV-G-标记
<400> 12
Tyr Thr Asp Ile Glu Met Asn Arg Leu Gly Lys
1 5 10
<210> 13
<211> 11
<212> PRT
<213> 人工
<220>
<223> HSV-标记
<400> 13
Gln Pro Glu Leu Ala Pro Glu Asp Pro Glu Asp
1 5 10
<210> 14
<211> 10
<212> PRT
<213> 人工
<220>
<223> T7 抗原表位
<400> 14
Ala Ser Met Thr Gly Gly Gln Gln Met Gly
1 5 10
<210> 15
<211> 10
<212> PRT
<213> 人工
<220>
<223> myc 抗原表位
<400> 15
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
1 5 10
<210> 16
<211> 14
<212> PRT
<213> 人工
<220>
<223> V5-标记
<400> 16
Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp Ser Thr
1 5 10
<210> 17
<211> 119
<212> PRT
<213> 小鼠
<400> 17
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 18
<211> 106
<212> PRT
<213> 小鼠
<400> 18
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn
100 105
<210> 19
<211> 116
<212> PRT
<213> 小鼠
<400> 19
Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala Ser Val Arg
1 5 10 15
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr Ile Ile His
20 25 30
Trp Ile Lys Leu Arg Ser Gly Gln Gly Leu Glu Trp Ile Gly Trp Phe
35 40 45
Tyr Pro Gly Ser Asn Asp Ile Gln Tyr Asn Ala Lys Phe Lys Gly Lys
50 55 60
Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Val Tyr Met Glu Leu
65 70 75 80
Thr Gly Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys Ala Arg Arg
85 90 95
Asp Asp Phe Ser Gly Tyr Asp Ala Leu Pro Tyr Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val
115
<210> 20
<211> 108
<212> PRT
<213> 小鼠
<400> 20
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Val Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Thr Asn Glu Asn Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Ala Ala Thr His Leu Val Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Thr Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Gly Asn Tyr Tyr Cys Gln His Phe Trp Gly Thr Pro Cys
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 21
<211> 314
<212> PRT
<213> 人工
<220>
<223> HLA-A*2402
<400> 21
Met Gly Ser His Ser Met Arg Tyr Phe Ser Thr Ser Val Ser Arg Pro
1 5 10 15
Gly Arg Gly Glu Pro Arg Phe Ile Ala Val Gly Tyr Val Asp Asp Thr
20 25 30
Gln Phe Val Arg Phe Asp Ser Asp Ala Ala Ser Gln Arg Met Glu Pro
35 40 45
Arg Ala Pro Trp Ile Glu Gln Glu Gly Pro Glu Tyr Trp Asp Glu Glu
50 55 60
Thr Gly Lys Val Lys Ala His Ser Gln Thr Asp Arg Glu Asn Leu Arg
65 70 75 80
Ile Ala Leu Arg Tyr Tyr Asn Gln Ser Glu Ala Gly Ser His Thr Leu
85 90 95
Gln Met Met Phe Gly Cys Asp Val Gly Ser Asp Gly Arg Phe Leu Arg
100 105 110
Gly Tyr His Gln Tyr Ala Tyr Asp Gly Lys Asp Tyr Ile Ala Leu Lys
115 120 125
Glu Asp Leu Arg Ser Trp Thr Ala Ala Asp Met Ala Ala Gln Ile Thr
130 135 140
Lys Arg Lys Trp Glu Ala Ala His Val Ala Glu Gln Gln Arg Ala Tyr
145 150 155 160
Leu Glu Gly Thr Cys Val Asp Gly Leu Arg Arg Tyr Leu Glu Asn Gly
165 170 175
Lys Glu Thr Leu Gln Arg Thr Asp Pro Pro Lys Thr His Met Thr His
180 185 190
His Pro Ile Ser Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly
195 200 205
Phe Tyr Pro Ala Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp
210 215 220
Gln Thr Gln Asp Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly
225 230 235 240
Thr Phe Gln Lys Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln
245 250 255
Arg Tyr Thr Cys His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr
260 265 270
Leu Arg Trp Glu Pro Pro Pro Ser Gly Ser Ser Ala Trp Ser His Pro
275 280 285
Gln Phe Glu Lys Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
290 295 300
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
305 310
<210> 22
<211> 314
<212> PRT
<213> 人工
<220>
<223> HLA-B*0702
<400> 22
Met Gly Ser His Ser Met Arg Tyr Phe Tyr Thr Ser Val Ser Arg Pro
1 5 10 15
Gly Arg Gly Glu Pro Arg Phe Ile Ser Val Gly Tyr Val Asp Asp Thr
20 25 30
Gln Phe Val Arg Phe Asp Ser Asp Ala Ala Ser Pro Arg Glu Glu Pro
35 40 45
Arg Ala Pro Trp Ile Glu Gln Glu Gly Pro Glu Tyr Trp Asp Arg Asn
50 55 60
Thr Gln Ile Tyr Lys Ala Gln Ala Gln Thr Asp Arg Glu Ser Leu Arg
65 70 75 80
Asn Leu Arg Gly Tyr Tyr Asn Gln Ser Glu Ala Gly Ser His Thr Leu
85 90 95
Gln Ser Met Tyr Gly Cys Asp Val Gly Pro Asp Gly Arg Leu Leu Arg
100 105 110
Gly His Asp Gln Tyr Ala Tyr Asp Gly Lys Asp Tyr Ile Ala Leu Asn
115 120 125
Glu Asp Leu Arg Ser Trp Thr Ala Ala Asp Thr Ala Ala Gln Ile Thr
130 135 140
Gln Arg Lys Trp Glu Ala Ala Arg Glu Ala Glu Gln Arg Arg Ala Tyr
145 150 155 160
Leu Glu Gly Glu Cys Val Glu Trp Leu Arg Arg Tyr Leu Glu Asn Gly
165 170 175
Lys Asp Lys Leu Glu Arg Ala Asp Pro Pro Lys Thr His Val Thr His
180 185 190
His Pro Ile Ser Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Gly
195 200 205
Phe Tyr Pro Ala Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp
210 215 220
Gln Thr Gln Asp Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Arg
225 230 235 240
Thr Phe Gln Lys Trp Ala Ala Val Val Val Pro Ser Gly Glu Glu Gln
245 250 255
Arg Tyr Thr Cys His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr
260 265 270
Leu Arg Trp Glu Pro Pro Pro Ser Gly Ser Ser Ala Trp Ser His Pro
275 280 285
Gln Phe Glu Lys Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
290 295 300
Ser Ala Trp Ser His Pro Gln Phe Glu Lys
305 310
<210> 23
<211> 321
<212> PRT
<213> 人工
<220>
<223> HLA-C*0702
<400> 23
Met Ala Leu Thr Glu Thr Trp Ala Cys Ser His Ser Met Arg Tyr Phe
1 5 10 15
Asp Thr Ala Val Ser Arg Pro Gly Arg Gly Glu Pro Arg Phe Ile Ser
20 25 30
Val Gly Tyr Val Asp Asp Thr Gln Phe Val Arg Phe Asp Ser Asp Ala
35 40 45
Ala Ser Pro Arg Gly Glu Pro Arg Ala Pro Trp Val Glu Gln Glu Gly
50 55 60
Pro Glu Tyr Trp Asp Arg Glu Thr Gln Lys Tyr Lys Arg Gln Ala Gln
65 70 75 80
Ala Asp Arg Val Ser Leu Arg Asn Leu Arg Gly Tyr Tyr Asn Gln Ser
85 90 95
Glu Asp Gly Ser His Thr Leu Gln Arg Met Ser Gly Cys Asp Leu Gly
100 105 110
Pro Asp Gly Arg Leu Leu Arg Gly Tyr Asp Gln Ser Ala Tyr Asp Gly
115 120 125
Lys Asp Tyr Ile Ala Leu Asn Glu Asp Leu Arg Ser Trp Thr Ala Ala
130 135 140
Asp Thr Ala Ala Gln Ile Thr Gln Arg Lys Leu Glu Ala Ala Arg Ala
145 150 155 160
Ala Glu Gln Leu Arg Ala Tyr Leu Glu Gly Thr Cys Val Glu Trp Leu
165 170 175
Arg Arg Tyr Leu Glu Asn Gly Lys Glu Thr Leu Gln Arg Ala Glu Pro
180 185 190
Pro Lys Thr His Val Thr His His Pro Leu Ser Asp His Glu Ala Thr
195 200 205
Leu Arg Cys Trp Ala Leu Gly Phe Tyr Pro Ala Glu Ile Thr Leu Thr
210 215 220
Trp Gln Arg Asp Gly Glu Asp Gln Thr Gln Asp Thr Glu Leu Val Glu
225 230 235 240
Thr Arg Pro Ala Gly Asp Gly Thr Phe Gln Lys Trp Ala Ala Val Val
245 250 255
Val Pro Ser Gly Gln Glu Gln Arg Tyr Thr Cys His Met Gln His Glu
260 265 270
Gly Leu Gln Glu Pro Leu Thr Leu Ser Trp Glu Pro Ser Ser Gln Pro
275 280 285
Thr Ile Gly Ser Ala Trp Ser His Pro Gln Phe Glu Lys Gly Gly Gly
290 295 300
Ser Gly Gly Gly Ser Gly Gly Ser Ala Trp Ser His Pro Gln Phe Glu
305 310 315 320
Lys
<210> 24
<211> 100
<212> PRT
<213> 人
<400> 24
Met Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala
1 5 10 15
Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His
20 25 30
Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu
35 40 45
Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr
50 55 60
Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala
65 70 75 80
Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp
85 90 95
Asp Arg Asp Met
100
<210> 25
<211> 118
<212> PRT
<213> 小鼠
<400> 25
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Arg Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Ser Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Ala Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Gln Ile Asn Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 26
<211> 111
<212> PRT
<213> 小鼠
<400> 26
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Phe Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Thr Thr Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Ala Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 27
<211> 283
<212> PRT
<213> 人工
<220>
<223> 具有His-标记的人CD19胞外域
<400> 27
Pro Glu Glu Pro Leu Val Val Lys Val Glu Glu Gly Asp Asn Ala Val
1 5 10 15
Leu Gln Cys Leu Lys Gly Thr Ser Asp Gly Pro Thr Gln Gln Leu Thr
20 25 30
Trp Ser Arg Glu Ser Pro Leu Lys Pro Phe Leu Lys Leu Ser Leu Gly
35 40 45
Leu Pro Gly Leu Gly Ile His Met Arg Pro Leu Ala Ile Trp Leu Phe
50 55 60
Ile Phe Asn Val Ser Gln Gln Met Gly Gly Phe Tyr Leu Cys Gln Pro
65 70 75 80
Gly Pro Pro Ser Glu Lys Ala Trp Gln Pro Gly Trp Thr Val Asn Val
85 90 95
Glu Gly Ser Gly Glu Leu Phe Arg Trp Asn Val Ser Asp Leu Gly Gly
100 105 110
Leu Gly Cys Gly Leu Lys Asn Arg Ser Ser Glu Gly Pro Ser Ser Pro
115 120 125
Ser Gly Lys Leu Met Ser Pro Lys Leu Tyr Val Trp Ala Lys Asp Arg
130 135 140
Pro Glu Ile Trp Glu Gly Glu Pro Pro Cys Leu Pro Pro Arg Asp Ser
145 150 155 160
Leu Asn Gln Ser Leu Ser Gln Asp Leu Thr Met Ala Pro Gly Ser Thr
165 170 175
Leu Trp Leu Ser Cys Gly Val Pro Pro Asp Ser Val Ser Arg Gly Pro
180 185 190
Leu Ser Trp Thr His Val His Pro Lys Gly Pro Lys Ser Leu Leu Ser
195 200 205
Leu Glu Leu Lys Asp Asp Arg Pro Ala Arg Asp Met Trp Val Met Glu
210 215 220
Thr Gly Leu Leu Leu Pro Arg Ala Thr Ala Gln Asp Ala Gly Lys Tyr
225 230 235 240
Tyr Cys His Arg Gly Asn Leu Thr Met Ser Phe His Leu Glu Ile Thr
245 250 255
Ala Arg Pro Val Leu Trp His Trp Leu Leu Arg Thr Gly Gly Trp Lys
260 265 270
Ala His His His His His His His His His His
275 280
- 还没有人留言评论。精彩留言会获得点赞!
- 琼脂糖在pcr扩增中的应用、pcr扩增试剂盒及pcr扩增方法
- 一种基于重组酶聚合酶扩增技术原理的组织胞浆菌感染分子诊断试剂盒及其应用
- 试剂盒及其在生殖道病原体扩增检测中的用图
- 一种基于核糖体dna its1基因检测华支睾吸虫囊蚴的pcr扩增试剂盒及扩增引物的制作方法
- 一种检测简单异尖线虫和华支睾吸虫囊蚴的双重pcr扩增试剂盒及其扩增引物的制作方法
- 一种基于线粒体coi基因检测华支睾吸虫囊蚴的pcr扩增试剂盒及扩增引物的制作方法
- 一种用于血液直接荧光pcr扩增的反应液及其试剂盒的制作方法
- 同时扩增人常染色体和y染色体str基因座的荧光标记复合扩增试剂盒及其应用
- 一种人基因组dna34个基因座的复合扩增的试剂盒的制作方法
- 一种具有增强鉴别能力的常染色体str基因座荧光标记复合扩增试剂盒及其应用