蛋白质TaTAR2;1在调控植物生物量和根系发育中的应用的制作方法
本发明涉及生物
技术领域:
,具体涉及蛋白质tatar2;1在调控植物生物量和根系发育中的应用。
背景技术:
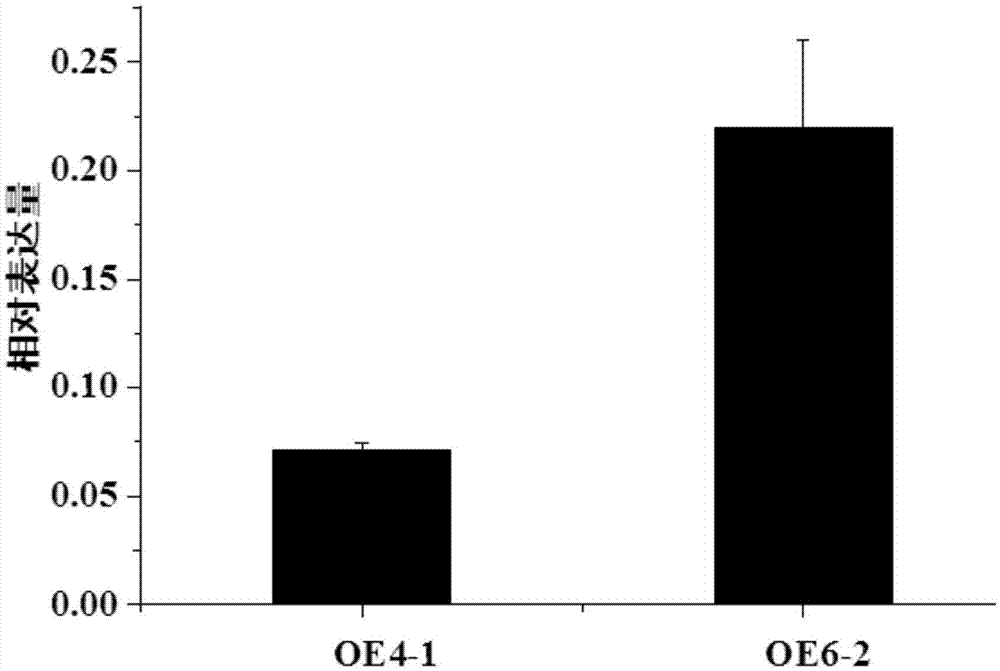
:氮素是植物必需的最重要的大量元素之一,是植物体内核酸、蛋白质、磷脂、叶绿素等重要化合物的主要组成成分,在物质代谢和能量转化中起重要作用。植物对氮素的缺乏可导致植物生长缓慢,产量下降,使粮食作物籽粒蛋白质含量下降,从而降低品质。氮肥的使用,是几十年来作物产量大幅提高的重要原因,但是自90年代以来,我国氮肥使用率继续大幅提高的同时,粮食总产和单产增长却放缓甚至停滞不前。与欧洲等国家相比,中国、印度、埃及等国家的氮肥利用效率呈逐年递减的趋势。近年对高产作物品种的研究也发现,尽管一些新的作物品种具有很好的产量,氮肥利用率却不高,高产不高效成为高产育种的一个重要问题,同时大幅度提高我国粮食单产和养分利用效率是我国农业可持续发展的关键。同时多达45–50%的氮肥未被农作物吸收利用而损失进入环境,导致资源浪费和环境污染。发达的根系是植物高效吸收水分以及营养元素的重要生理基础之一,例如在小麦中,氮高效小麦品种科农9204和小偃54在表层和底层土壤中的根长比小麦品种京411的发达,吸氮效率也高;小麦-黑麦1bl/1rs易位系在小麦育种中得到广泛利用,这与黑麦1rs可显著增加小麦表层和深层根系生物量、促进氮等养分吸收有关。鉴于根系在高效吸收水肥方面的重要性,科学家认为遗传改良农作物根系形态构型是实现第二次绿色革命、进一步提高产量的关键。技术实现要素:本发明要解决的技术问题是如何调控植物生物量和/或植物根系发育。为解决上述问题,本发明首先提供了蛋白质tatar2;1在调控植物生物量和/或调控植物根系发育中的应用;所述蛋白质tatar2;1为a1)或a2)或a3):a1)氨基酸序列是序列表中序列2所示的蛋白质;a2)在序列表中序列2所示的蛋白质的n端或/和c端连接标签得到的融合蛋白质;a3)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与调控植物生物量和/或植物根系发育相关的蛋白质。其中,序列表中序列2可由431个氨基酸残基组成。为了使a1)中的蛋白质便于纯化,可在序列表中序列2所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1、标签的序列标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tagii8wshpqfekc-myc10eqkliseedl上述a3)中的蛋白质tatar2;1,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。上述a3)中的蛋白质tatar2;1可人工合成,也可先合成其编码基因,再进行生物表达得到。上述a3)中的蛋白质tatar2;1的编码基因可通过将序列表中序列1所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。编码所述蛋白质tatar2;1的核酸分子在调控植物生物量和/或调控植物根系发育中的应用也属于本发明的保护范围。其中,所述编码所述蛋白质tatar2;1的核酸分子可以是dna,如cdna、基因组dna或重组dna;所述编码所述蛋白质tatar2;1的核酸分子也可以是rna,如mrna或hnrna等。所述编码所述蛋白质tatar2;1的核酸分子可为如下b1)或b2)或b3):b1)核苷酸序列是序列表中序列1所示的dna分子;b2)与b1)限定的核苷酸序列具有75%或75%以上同一性,且编码所述蛋白质tatar2;1的dna分子;b3)在严格条件下与(b1)或(b2)限定的核苷酸序列杂交,且编码所述蛋白质tatar2;1的dna分子。其中,序列表中序列1由1296个核苷酸组成,编码序列表中序列2所示的氨基酸序列。本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的蛋白质tatar2;1的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的蛋白质tatar2;1的核苷酸序列具有75%或者更高同一性的核苷酸,只要编码蛋白质tatar2;1且具有蛋白质tatar2;1的功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列表中序列2所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。以上任一所述应用中,所述调控植物生物量可为增加植物生物量。以上任一所述应用中,所述调控植物根系发育可为促进植物根系发育。以上任一所述应用中,所述增加植物生物量可为地上部生物量增加和/或根系生物量增加。以上任一所述应用中,所述促进植物根系发育可为可见侧根数增多和/或总侧根长增加和/或主根长增加。以上任一所述地上部生物量可为每株植株的地上部分鲜重。以上任一所述根系生物量可为每株植物的根系鲜重。以上任一所述应用中,所述植物可为如下c1)至c4)中的任一种:c1)双子叶植物;c2)单子叶植物;c3)十字花科植物;c4)拟南芥。为解决上述问题,本发明还提供了一种培育转基因植物的方法。本发明所提供的培育转基因植物的方法,包括将编码所述蛋白质tatar2;1的核酸分子导入受体植物中,得到转基因植物的步骤;所述转基因植物与所述受体植物相比地上部生物量增加和/或根系生物量增加和/或可见侧根数增多和/或总侧根长增加和/或主根长增加。其中,所述编码所述蛋白质tatar2;1的核酸分子可以是dna,如cdna、基因组dna或重组dna;所述编码所述蛋白质tatar2;1的核酸分子也可以是rna,如mrna或hnrna等。上述方法中,所述编码所述蛋白质tatar2;1的核酸分子可为如下b1)或b2)或b3):b1)核苷酸序列是序列表中序列1所示的dna分子;b2)与b1)限定的核苷酸序列具有75%或75%以上同一性,且编码所述蛋白质tatar2;1的dna分子;b3)在严格条件下与(b1)或(b2)限定的核苷酸序列杂交,且编码所述蛋白质tatar2;1的dna分子。上述方法中,所述地上部生物量可为单株地上部分鲜重。所述根系生物量可为单株根系鲜重。上述方法中,所述植物可为如下c1)至c4)中的任一种:c1)双子叶植物;c2)单子叶植物;c3)十字花科植物;c4)拟南芥。所述拟南芥具体可为columbia-0生态型拟南芥。上述方法中,所述“将编码所述蛋白质tatar2;1的核酸分子导入受体植物中”可通过向受体植物中导入重组载体实现;所述重组载体可为将所述蛋白质tatar2;1的编码基因插入出发质粒得到的重组质粒。所述重组载体具体可为重组质粒pcambia2300-tatar2;1。所述重组质粒pcambia2300-tatar2;1具体可为在载体pcambia2300的多克隆位点插入序列表中序列1所示的dna分子得到的重组质粒。所述重组质粒pcambia2300-tatar2;1具体可为将载体pcambia2300的xbaⅰ和salⅰ识别序列间的片段(载体pcambia2300被限制性核酸内切酶xbaⅰ和salⅰ切成一个大片段和一个小片段,该dna为该小片段)替换为序列表中序列1所示的dna分子得到的重组质粒。实验证明,本发明所提供的蛋白质tatar2;1能增加植物生物量和/或促进植物根系发育:与野生型植物相比,转tatar2;1基因植物的可见侧根数、总侧根长、主根长、地上部生物量和根系生物量均显著增加。因此,可以利用蛋白质tatar2;1调控植物生物量和/或调控植物根系发育,本发明对植物生物量和/或植物根系发育的新材料的选育具有重要应用价值。附图说明图1为pcambia2300/tatar2;1的部分元件示意图图2为oe4-1和oe6-2中tatar2;1基因相对表达量。图3为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的生长情况。图4为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的可见侧根数统计结果。图5为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的总侧根长统计结果。图6为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的主根长统计结果。图7为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的地上部鲜重统计结果。图8为野生型拟南芥(col-0)、oe4-1和oe6-2在高氮(hn)和低氮(ln)条件下的根系鲜重统计结果。具体实施方式下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中的高氮培养基的各组分见表2,低氮培养基的各组分见表3。表2.高氮培养基的组分表3.低氮培养基的组分根癌农杆菌gv3101为普如汀生物技术(北京)有限公司产品,产品目录号为biovector-375。野生型拟南芥(arabidopsisthaliana)(columbia-0生态型)购自arabidopsisbiologicalresourcecenter(abrc,网址为:http://www.abrc.com/),材料编号为cs6673。公众可从中国科学院遗传与发育生物学研究所获得,以重复本申请实验。拟南芥(arabidopsisthaliana)(columbia-0生态型)在下文中简称野生型拟南芥或col-0。小麦“科农199”(简称科农199)记载于如下文献中:李俊明,张相岐,张爱民.高产广适小麦新品种——科农199.《麦类作物学报》,2007.公众可从中国科学院遗传与发育生物学研究所获得,以重复本申请实验。载体pcambia2300为北京鼎国昌盛生物技术有限责任公司产品,货号为mcv036-n;载体peasytm-blunt为北京全式金生物技术有限公司产品,产品目录号为cb501-02。实施例1、转tatar2;1基因拟南芥的获得及表型鉴定一、重组载体和重组农杆菌的构建1、tatar2;1基因的获得(1)采用trizol法提取科农199的幼嫩叶片的总rna,将该总rna用逆转录酶反转录出第一链cdna。2、人工合成引物f1:5’-tctagaatgccactatggcgagtcttc-3’(下划线为限制性内切酶xbaⅰ识别序列)和r1:5’-gtcgacccatttgccattcgaaactgct-3’(下划线为限制性内切酶salⅰ识别序列)。3、以步骤1获得的cdna为模板,以步骤2合成的f1和r1为引物进行pcr扩增,得到约1500bp的双链dna分子。反应程序:98℃2min,(98℃30s,58℃30s,68℃45s)×35循环,72℃10min。4、将步骤3中得到的双链dna分子连接至载体peasytm-blunt,得到重组质粒peasytm-blunt-tatar2;1。5、完成步骤4后,用限制性内切酶xbaⅰ和salⅰ双酶切重组质粒peasytm-blunt-tatar2;1,回收约1500bp的片段。6、用限制性内切酶xbaⅰ和salⅰ双酶切载体pcambia2300,回收约8700bp的载体骨架。7、将步骤5得到的片段与步骤6得到的载体骨架连接,得到重组质粒pcambia2300-tatar2;1(部分元件的示意图见图1)。根据测序结果,对重组质粒pcambia2300-tatar2;1进行结构描述如下:将载体pcambia2300的xbaⅰ和salⅰ识别序列间的片段(载体pcambia2300被限制性核酸内切酶xbaⅰ和salⅰ切成一个大片段和一个小片段,该dna为该小片段)替换为序列表中序列1所示的dna分子。重组质粒pcambia2300-tatar2;1表达序列表中序列2所示的tatar2;1蛋白。重组质粒pcambia2300-tatar2;1中还含有一个由35s启动子启动nptii基因表达的表达盒(见图1),因此可利用硫酸卡那霉素(kanamycinsulfate)筛选转化植株。将重组质粒pcambia2300-tatar2;1导入根癌农杆菌gv3101,得到重组农杆菌,命名为gv3101/pcambia2300-tatar2;1。将载体pcambia2300导入根癌农杆菌gv3101,得到重组农杆菌,命名为gv3101/pcambia2300。二、拟南芥转tatar2;1基因植株的再生和筛选1、采用拟南芥花序浸花转化法(记载于如下文献中clough,s.j.,andbent,a.f..floraldip:asimplifiedmethodforagrobacterium-mediatedtransformationofarabidopsisthaliana.plantj.(1998)16,735-743.),将步骤一制备的gv3101/pcambia2300-tatar2;1转至野生型拟南芥中,获得t1代转tatar2;1基因拟南芥种子。2、将步骤1获得的t1代转tatar2;1基因拟南芥种子播种于含有40mg/l硫酸卡那霉素的1/2ms培养基上,能够正常生长的拟南芥(抗性苗)即为t1代转tatar2;1基因阳性苗,t1代转tatar2;1基因阳性苗收到的种子即为t2代转tatar2;1基因拟南芥种子。3、将步骤2筛选出的不同株系的t2代转tatar2;1基因拟南芥种子播种于含有40mg/l硫酸卡那霉素的1/2ms培养基上进行筛选,如果某株系中能够正常生长的拟南芥(抗性苗)的数目与不能够正常生长的拟南芥(非抗性苗)的数目比例为3:1,则该株系为tatar2;1基因插入一个拷贝的株系,该株系中的抗性苗收到的种子即为t3代转tatar2;1基因拟南芥种子。4、将步骤3筛选出的t3代转tatar2;1基因拟南芥种子再次播种于含有40mg/l硫酸卡那霉素的1/2ms培养基上进行筛选,均为抗性苗的即为t3代纯合转tatar2;1基因拟南芥。将其中2个t3代纯合转tatar2;1基因拟南芥株系分别命名为oe4-1和oe6-2,并进行后续实验。按照上述方法,将gv3101/pcambia2300-tatar2;1替换为gv3101/pcambia2300,其他步骤均相同,得到t3代纯合转空载体拟南芥的植株,简称转空载体拟南芥。三、real-timepcr检测tatar2;1基因的相对表达量实验重复三次,每次重复的步骤如下:1、样本的获得取oe4-1的t3代植株的幼嫩叶片,放入液氮保存,得到样本1。取oe6-2的t3代植株的幼嫩叶片,放入液氮保存,得到样本2。取转空载体拟南芥的t3代植株的幼嫩叶片,放入液氮保存,得到样本3。取野生型拟南芥植株的幼嫩叶片,放入液氮保存,得到样本4。2、real-timepcr检测tatar2;1基因的相对表达量利用rneasyplantminikit提取试剂盒(qiagen公司产品)提取样本1的总rna,将该总rna用逆转录酶反转录出第一链cdna,然后将该cdna稀释10倍作为模板,根据premixextaqtmii(perfectrealtime)试剂盒(takara公司产品)说明书的步骤,real-timepcr检测样本1中tatar2;1基因的相对表达量(以actin基因作为内参基因)。real-timepcr使用的仪器为eprealplex(eppendorf公司产品,德国)。将样本1中actin基因的表达量作为1,样本1中tatar2;1基因的相对表达量见图2。鉴定tatar2;1基因的引物为5’-tgtgtctgtcgcattgaatgtc-3’和5’-atgttgctgatcaggagagatgac-3’。内参引物为5’-cctcgtctcgaccttgctggg-3’和5’-gagaacaagcaggaggacggc-3’。按照上述步骤,将样本1分别替换为样本2、样本3和样本4,其它步骤均相同,得到样本2、样本3和样本4中tatar2;1基因的相对表达量(图2)。实验结果表明,oe4-1的t3代植株中tatar2;1基因的相对表达量为1.718,oe6-2的t3代植株中tatar2;1基因的相对表达量为2.466,野生型拟南芥和转空载体拟南芥的t3代植株中tatar2;1基因的相对表达量均为0。可见,oe4-1的t3代植株和oe6-2的t3代植株均为转tatar2;1基因拟南芥植株。四、转tatar2;1基因拟南芥的表型鉴定以野生型拟南芥种子、转空载体拟南芥的t3代种子、oe4-1的t3代种子和oe6-2的t3代种子为实验材料,鉴定拟南芥的表型。实验重复三次取平均值,每个株系每次重复20株。每次重复步骤如下:1、将野生型拟南芥种子、转空载体拟南芥的t3代种子、oe4-1的t3代种子或oe6-2的t3代种子先用1%次氯酸钠室温灭菌10min,无菌蒸馏水冲洗5遍,然后4℃避光放置4天(目的使种子发芽一致)。2、将完成步骤1的种子播种于1/2ms培养基,22℃竖直培养4天,挑选生长状态一致的拟南芥幼苗转移至低氮培养基(ln)或高氮培养基(hn)上,竖直培养6天,观察植株的生长情况并统计可见侧根数、总侧根长、主根长、地上部鲜重和根系鲜重。野生型拟南芥、oe4-1的t3代植株和oe6-2的t3代植株在高氮培养基或低氮培养基下的生长情况见图3。各植株的可见侧根数的统计结果见图4。结果表明,在低氮培养基中,oe4-1的t3代植株的可见侧根数为9.0条,oe6-2的t3代植株的可见侧根数为9.6条,野生型拟南芥的可见侧根数为5.8条;在高氮培养基中,oe4-1的t3代植株的可见侧根数为7.1条,oe6-2的t3代植株的可见侧根数为7.0条,野生型拟南芥的可见侧根数为4.4条。各植株的总侧根长的统计结果见图5。结果表明,在低氮培养基中,oe4-1的t3代植株的总侧根长为6.2cm,oe6-2的t3代植株的总侧根长为6.9cm,野生型拟南芥的总侧根长为3.0cm;在高氮培养基中,oe4-1的t3代植株的总侧根长为4.0cm,oe6-2的t3代植株的总侧根长为3.9cm,野生型拟南芥的总侧根长为1.6cm。各植株的主根长的统计结果见图6。结果表明,在低氮培养基中,oe4-1的t3代植株的主根长为6.1cm,oe6-2的t3代植株的主根长为6.1cm,野生型拟南芥的主根长为5.3cm;在高氮培养基中,oe4-1的t3代植株的主根长为6.0cm,oe6-2的t3代植株的主根长为6.1cm,野生型拟南芥的主根长为5.2cm。各植株的地上部每株鲜重的统计结果见图7。结果表明,在低氮培养基中,oe4-1的t3代植株的单株拟南芥地上部的平均鲜重为1.79mg,oe6-2的t3代植株的单株拟南芥地上部的平均鲜重为1.98mg,单株野生型拟南芥地上部的平均鲜重为1.48mg;在高氮培养基中,oe4-1的t3代植株的单株拟南芥地上部的平均鲜重为4.90mg,oe6-2的t3代植株的单株拟南芥地上部的平均鲜重为5.14mg,单株野生型拟南芥地上部的平均鲜重为3.39mg。各植株的根系每株鲜重的统计结果见图8。结果表明,在低氮培养基中,oe4-1的t3代植株的单株拟南芥根系的平均鲜重为2.02mg,oe6-2的t3代植株的单株拟南芥根系的平均鲜重为2.05mg,单株野生型拟南芥根系的平均鲜重为1.46mg;在高氮培养基中,oe4-1的t3代植株的单株拟南芥根系的平均鲜重为1.50mg,oe6-2的t3代植株的单株拟南芥根系的平均鲜重为1.53mg,单株野生型拟南芥根系的平均鲜重为1.03mg。野生型拟南芥和转空载体拟南芥的t3代植株在高氮培养基或低氮培养基下的可见侧根数、总侧根长、主根长、单株拟南芥地上部每株鲜重和根系每株鲜重无显著差异。结果表明,与野生型拟南芥相比,在高氮培养基或低氮培养基中培养的oe4-1的t3代植株或oe6-2的t3代植株的可见侧根数、总侧根长、主根长、地上部生物量(单株拟南芥地上部的平均鲜重)和根系生物量(单株拟南芥根系的平均鲜重)均显著增加。可见,将tatar2;1基因导入拟南芥中,在高氮和低氮条件下均会增加生物量和促进根系发育。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1