莲纤维素合酶基因NnuCESA4的应用的制作方法
本发明属于植物基因工程
技术领域:
,涉及一种纤维素合酶基因的应用,特别涉及一种莲纤维素合酶基因NnuCESA4的应用。
背景技术:
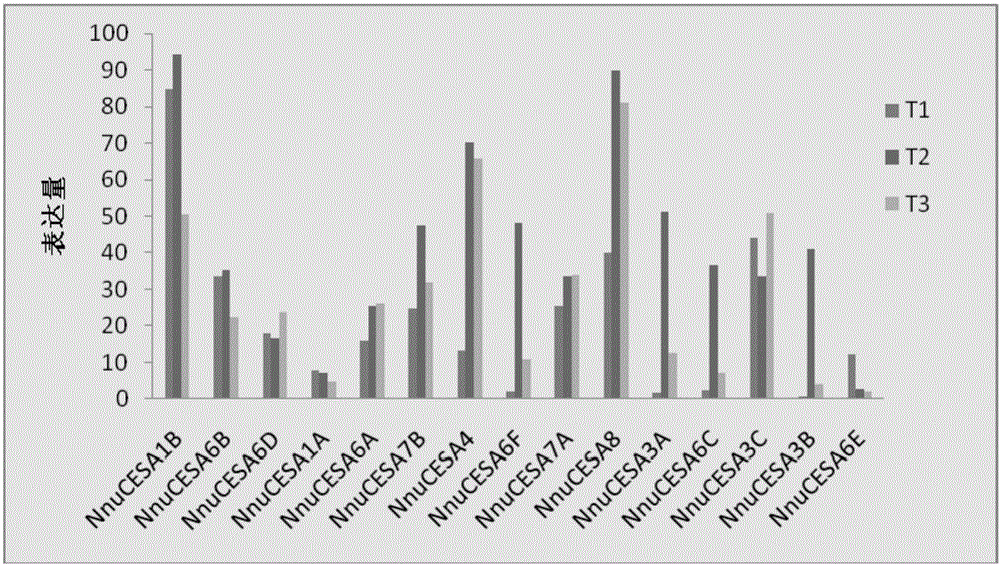
:纤维素是细胞壁的主要组成成分之一,也是评价造纸原料性能优劣的一个重要指标。植物细胞壁主要纤维素、半纤维素、木质素三大成分组成,这三大组分是地球上最丰富的可再生资源,而纤维素是细胞壁的第一大组分,由均一的吡喃式D-葡萄糖以β-1,4糖苷键连接而成,其葡萄糖残基约2000~25000个(PaulyM,KeegstraK(2010)Plantcellwallpolymersasprecursorsforbiofuels.CurrentOpinioninPlantBiology,95:305-312;PaulyM,KeegstraK(2008)Cell-wallcarbohydratesandtheirmodificationasaresourceforbiofuels.ThePlantJournal,54(4):559-568)。而这种β-1,4糖苷链由纤维素合酶催化合成。Richmond和Somerville在拟南芥中已发现了10个纤维素合酶的编码基因(现命名为AtCESA)(RichmondTA,SomervilleCR.(2000)Thecellulosesynthasesuperfamily1.PlantPhysiology,124:496-498)。AtCESA1,AtCESA2,AtCESA3,AtCESA5,AtCESA6和AtCESA9与植物初生壁的生物合成有关(ArioliT,PengL,BetznerA,etal.(1998)MolecularanalysisofcellulosebiosynthesisinArabidopsis.Science,279:717-720;BurnJE,etal.FunctionalanalysisofthecellulosesynthasegenesCESA1,CESA2,andCESA3inarabidopsis.PlantPhysiology,129(2):797-807;PagantS,etal.(2002)KOBITO1encodesanovelplasmamembraneproteinnecessaryfornormalsynthesisofcelluloseduringcellexpansioninarabidopsis.ThePlantCellOnline,14(9):2001-2013),次生壁纤维素合酶基因复合体由AtCESA4,AtCESA7和AtCESA8构成,这些基因的突变体都表现为坍塌的木质部(irregularxylem,irx)(TurnerSR,SomervilleCR.(1997)CollapsedxylemphenotypeofArabidopsisidentifiesmutantsdeficientincellulosedepositioninthesecondarycellwall.PlantCell,9:689-701;SzyjanowiczPMJ,MckinnonI,TaylorNG,etal.(2004)Theirregularxylem2mutantisanallele ofkorriganthataffectsthesecondarycellwallofArabidopsisthaliana.ThePlantJournal,37(5):730-740)。拟南芥cesa4突变体和cesa8突变体次生细胞壁中的纤维素沉积出现严重缺陷导致植株出现坍塌的木质部细胞(TurnerSR,SomervilleCR(1997)CollapsedxylemphenotypeofArabidopsisidentifiesmutantsdeficientincellulosedepositioninthesecondarycellwall.PlantCell,9:689-701;TaylorNG,HowellsRM,HuttlyAK,etal.(2002)InteractionsamongthreedistinctCesAproteinsessentialforcellulosesynthesis.100(3):1450-1455)。目前,CESA的功能推测主要来自于基因表达、UDP-G结合实验(PearJR,KawagoeY,SchreckengostWE,etal.(1996)HigherplantscontainhomologsofthebacterialcelAgenesencodingthecatalyticsubunitofcellulosesynthase.ProcNatlAcadSciUSA,93(22):12637-12642)、突变分析(FagardM,DesnosT,DesprezT,etal.(2000)PROCUSTE1encodesacellulosesynthaserequiredfornormalcellelongationspecificallyinrootsanddark-grownhypocotylsofArabidopsis.ThePlantCell,12(12):2409-2424)及基因沉默实验(Burtonetal,2000)。莲(Nelumbonucifera)属睡莲科(Nymphaeaceae)莲属(Nelumbo)多年生水生宿根草本植物。莲叶柄中含有大量的纤维素,从中间折断可拉出大量细长而有韧性的莲纤维。莲纤维的性能非常特殊,Pan等报道,两段掰开的叶柄之间的纤维丝能被拉长至少十厘米而不断开,每根纤维丝由二十根左右的细纤维丝组成,平行并以螺旋形式的分布在莲叶柄的导管分子中(PanY,HanG,MaoZ(2011)TheanatomyoflotusfibersfoundinpetiolesofNelumbonucifera.AquaticBotany,95:167-171)。莲纤维丝的成分中纤维素、半纤维素、木质素的含量分别是41.4±0.29%,25.87±0.64%,19.56±0.32%(PanY,HanG,MaoZ(2011)StructuralcharacteristicsandphysicalpropertiesoflotusfibersobtainedfromNelumbonuciferapetioles.CarbohydPolym,85:188-195)。克隆出莲的纤维素合酶基因,并利用转基因技术研究其功能表达,将有利于深入研究莲纤维丝及纤维素的合成机理,进而有目的地利用莲纤维素合酶基因去改善一些林木树种用于增加其纤维素含量,提高可利用性。莲以及莲纤维丝的药学应用、化学性能和物理结构均被研究并报道过,也有莲基因水平的研究,莲的基因组于2013年公布(RayM,RobertV,YanlingL,etal.(2013)Genomeofthelong-livingsacredlotus(NelumbonuciferaGaertn.).GenomeBiology,14(5):R41),这为莲相关基因的克隆与功能研究提供了便利。技术实现要素:为了克服现有技术的缺点与不足,本发明的目的在于提供一种莲纤维素合酶基因NnuCESA4的应用。鉴于莲的叶柄中含有大量纤维丝且叶柄内部、外部纤维素含量不一致的现象,为从基因表达程度方面入手进行深入研究其纤维素合成,将莲叶柄内、外和叶片三个部位做转录组测序,进而研究其纤维素合酶基因的表达。本发明的主要目的是将转录组中发现的一段在叶柄部位表达量比较高的纤维素合酶基因的表达情况。根据转录组测序得到的序列设计引物以莲叶柄总cDNA为模板将此基因克隆出来并进行测序,对测序结果行BLAST,发现其与拟南芥的AtCESA4基因序列相似度最高,因此我们将之命名为NnuCESA4。然后从ABRC(ArabidopsisBiologicalResourceCenter)订购到拟南芥cesa4突变体(Salk_029940c),并通过转基因技术将莲中的NnuCESA4基因转入到相应的拟南芥突变体中检测其功能互补情况,推断该基因的功能表达情况,以便后续可有目的性的将该基因转入到一些林木树种用于增加其纤维素含量,提高可利用性。本发明通过构建过表达载体及通过转基因技术来研究其功能表达,确定该基因具有纤维素合酶功能并促进植物次生细胞壁的纤维素合成。本发明的目的通过下述技术方案实现:本发明提供一种莲纤维素合酶基因NnuCESA4在促进纤维素合成中的应用。本发明提供一种莲纤维素合酶基因NnuCESA4在促进次生细胞壁中纤维素合成中的应用。所述的莲纤维素合酶基因NnuCESA4的氨基酸序列如SEQIDNO:1所示。所述的莲纤维素合酶基因NnuCESA4的核苷酸序列如SEQIDNO:2所示。上述应用的具体过程如下:1.基因的获得根据转录组数据,得到了15个莲的CESA基因序列,对这15个CESA基因进行了RT-qPCR,发现其中一条序列表达量特别高并且在叶柄内外部位表达量远远高于叶片,根据转录组测序结果设计引物以莲叶柄总cDNA为模板将此基因克隆出来并进行测序,对测序结果行BLAST,发现本序列与拟南芥的AtCESA4序列相似度最高,因此我们猜测其与次生细胞壁纤维素的合成密切相关,并将该基因命名为NnuCESA4,进而搜索已公布的莲基因组序列,发现本基因序列与NNU_10513序列相同。所述的引物序列如下:NnuCESA4-F:5′-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATGAC CGGCCTAATCGCCG-3′;NnuCESA4-R:5′-GGGGACCACTTTGTACAAGAAAGCTGGGTCTTAGCACTCCACTCCACATTGC-3′。2.载体的构建将扩增的NnuCESA4的cDNA构建至过表达载体pEarlyGate100中组成双元表达载体,得到过表达重组质粒;该载体携带花椰菜花叶病毒(CaMV)的35S启动子,并具有除草剂抗性,可供转基因植物的筛选。3.转化农杆菌将上述得到的过表达重组质粒采用冻融法转化至根癌农杆菌GV3101中,选取阳性菌并保存,准备转染拟南芥。4.转化拟南芥cesa4:由于拟南芥cesa4纯合突变体植株矮小,果荚畸形,几乎不结种子,故选用无表型的cesa4杂合突变体进行转基因并收取种子以备下一代阳性植株筛选,将上述得到的含有目的基因的阳性农杆菌以浸染法转化45天盛花期的cesa4杂合突变体植株,果荚成熟后收集种子,用除草剂抗性筛选转基因阳性植株,并通过基因鉴定选取基因背景为cesa4纯合突变体的阳性植株,即为转基因植株。以拟南芥cesa4纯合突变体作为阴性对照。5.转基因植株表型观察拟南芥cesa4纯合突变体植株矮小,莲座叶窄短呈深绿色,果荚畸形短小,几乎无种子,纤维素含量较低,并具有严重坍塌的木质部结构。通过转莲基因NnuCESA4入拟南芥cesa4纯合突变体,对相同条件下种植的生长6到7周的转基因植株、野生型植株及cesa4纯合突变体植株的主茎高度、莲座叶大小及果荚状态进行比较观察并记录。转基因植株长出了少量比野生型短小但能正常结种子的果荚,莲座叶长度比cesa4纯合突变体增长了92%,并且茎秆高度也增长了47%。为了观察木质部变化,对转基因植株的主茎横切片进行甲苯胺蓝染色,在光学显微镜下观察,cesa4纯合突变体原本坍塌比较严重的木质部细胞在莲NnuCESA4基因的转入后得到了部分恢复。用硝酸乙醇法测定拟南芥主茎的纤维素含量,测定结果证明转基因植株的纤维素含量比cesa4纯合突变体增长了1.16倍,但尚未恢复至野生型程度。结果表明:通过进化树分析和转基因技术及互补植株表型分析确定莲基因NnuCESA4具有纤维素合酶功能,能参与次生细胞壁中纤维素的合成。本发明相对于现有技术,具有如下的优点及效果:(1)本发明扩增得到莲基因NnuCESA4,并得到了含有莲基因NnuCESA4的过表达载体。(2)本发明得到了莲基因NnuCESA4转拟南芥cesa4纯合突变体的阳性转基因植株。(3)本发明通过转基因植株、纯合突变体植株与野生型植株对比,发现该基因能部分互补拟南芥cesa4纯合突变体的表型,具有纤维素合酶的功能,促进纤维素的合成,并确定该基因参与植物次生细胞壁中纤维素的合成。附图说明图1是转录组测序中15个NnuCESA基因在不同部位的表达量;其中,T1,T2,T3分别是叶片、叶柄外部和叶柄内部。图2是15个NnuCESA基因的在不同部位的RT-qPCR结果;其中,T1,T2,T3分别是叶片、叶柄外部和叶柄内部。图3是表达载体构建示意图。图4是cesa4纯合突变体、转基因植株和野生型植株表型对比图;其中,图A中从左至右分别为cesa4纯合突变体,转基因植株和野生型植株;图B中从下至上三排分别为cesa4纯合突变体,转基因植株和野生型植株的莲座叶片;图C中从下至上三排分别为cesa4纯合突变体,转基因植株和野生型植株的果荚。图5是cesa4纯合突变体、转基因植株和野生型植株茎部横切切片对比图。其中,A,B,C分别为cesa4纯合突变体,转基因植株和野生型植株的茎部横切面甲苯胺蓝染色图,箭头所指的部位为木质部细胞。具体实施方式下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。下列实施例中未注明具体实验条件的实验方法,通常按照常规实验条件或按照制造厂商所建议的实验条件。实施例1:莲NnuCESA4基因序列的获得先将莲花叶柄内、外和叶片三个部位做转录组测序得到各基因片段在不同 部位的表达量,找出15个CESA基因片段的表达量(结果见图1),并做RT-qPCR验证(结果见图2),各基因在不同部位的相对表达量结果与转录组测序的结果趋势基本一致,其中一段CESA基因序列表达量特别高并且在茎秆内部和外部的表达量均高于叶片部位,根据转录组测序结果设计引物以叶柄总cDNA为模板将此基因克隆出来并进行测序,对测序结果行BLAST,发现该基因与拟南芥中参与次生细胞壁纤维素合成的基因AtCESA4相似度极高,因此我们推测该基因应该参与次生细胞壁中纤维素的合成,并将其命名为NnuCESA4,通过序列比对发现,本基因序列与莲基因组网站(http://lotus-db.wbgcas.cn/)中的基因NNU_10513序列相同。实施例2:莲叶柄的总RNA的提取用RNA提取试剂盒(OMEGA)分别提取莲叶柄的总RNA。详细步骤参考试剂盒说明书。实施例3:莲总cDNA的获得以莲叶柄总RNA为模板采用OMEGA反转录试剂盒得莲总cDNA,详细步骤按照试剂盒使用说明书操作。实施例4:目的片段的扩增以莲基因NnuCESA4的mRNA为模板设计两端引物,并在引物两端加上用于构建载体的Gateway接头引物。上下游引物分别为:NnuCESA4-F:5′-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATGACCGGCCTAATCGCCG-3′;NnuCESA4-R:5′-GGGGACCACTTTGTACAAGAAAGCTGGGTCTTAGCACTCCACTCCACATTGC-3′;以莲叶柄总cDNA为模板,在退火温度为52℃及延伸温度为72℃的程序下进行PCR扩增,扩增产物经电泳鉴定后纯化并保存。实施例5:载体构建连接采用Gateway双元载体构建方法,将实施例4中获得的扩增产物与载体质粒pDONR207(购买于ABRC,http://abrc.osu.edu/)进行BP重组反应,重组产物转化DH5α感受态细胞(市售)并得到阳性菌,扩大培养并提取质粒后进行 质粒测序鉴定。将测序正确的重组质粒与pEarlyGate100(购买于ABRC,http://abrc.osu.edu/)(该载体携带花椰菜花叶病毒(CaMV)的35S启动子,并具有除草剂抗性,可供转基因植物的筛选)进行LR重组反应,重组产物转化大肠杆菌DH5α并得到阳性菌,扩大培养后提取质粒并测序,将测序正确的重组质粒保存备用。表达载体构建示意图如图3所示。测序结果如SEQIDNO:2所示,与基因NnuCESA4(基因ID为NNU_10513)中序列相同。实施例6:转化根癌农杆菌GV31011.将根癌农杆菌(Agrobacteriumtumefaciens)GV3101感受态细胞(市售)冰上融化,加入1至5μL(<300μg)实施例5中的重组质粒,轻弹混匀,冰浴15min。2.将上述混合物转移至液氮中急冻5min,再37℃热激5min,迅速转移至冰上5min。3.加入800μL无抗性的LB液体培养基,28℃,200rpm摇4h。4.5000rpm,离心1min,弃700μL上清,其余重悬,取200μL涂布于LB固体培养基(Rif+:50μg/mL,Gen+:15μg/mL),28℃倒置培养2天。5.选取阳性菌落并小摇保菌。实施例7:农杆菌转染拟南芥1.选用抽苔十天后的拟南芥(Arabidopsisthaliana)cesa4杂合突变体植株,保持较高湿度,准备转化。2.在LB培养基中分别复苏上述带有重组质粒的根癌农杆菌,28℃,220rpm小遥24h。3.5000rpm离心15min收集菌体。同时配制1L重悬液(5%(w/v)Sucrose,0.02~0.05%Silwet-77)。4.弃上清,残留菌液用5%(w/v)蔗糖重悬清洗,8000rpm,离心10min收集菌体,此步骤可再复一次。5.用重悬液重悬菌体至OD600位0.8,重悬液转移至15cm培养皿中。6.将待转化拟南芥的花序侵入重悬液中1min。7.将已转化植株倒置平放,22℃,相对湿度60~70%暗培养一天后,转为正常培养,一个月后收取转化种子。实施例8:筛选转基因阳性植株将实施例7中得到的种子播种,萌芽10天左右喷施除草剂,得到存活植株后通过DNA鉴定确定目的基因是否转入及植株基因型背景,做好标记,转基因后的cesa4纯合突变体可结少量种子,待果荚成熟后单株收种并保存。实施例9:拟南芥观察表型选取生长6~7周的野生型、cesa4纯合突变体及转基因植株来观察对比植株表型变化,每种类型的植株选取十株以上并用标尺来测量其茎秆高度、莲座叶长度,观察并拍照记录果荚大小。结果如图4和表1所示,结果表明,cesa4纯合突变体的畸形果荚较多,几乎无种子;转基因植株长出了少量比野生型短小但能正常结种子的果荚,莲座叶长度比cesa4纯合突变体增长了92%,并且茎秆高度也增长了47%。表1野生型植株、纯合突变体和转基因植株纤维素含量比较结果野生型cesa4纯合突变体转基因植株茎秆纤维素含量(μg/mg)300±8.97A78.625±2.33C169.8±5.16B莲座叶长度(cm)4.41±0.13A1.13±0.13C2.17±0.09B茎秆高度(cm)33.4±2.37A10.01±1.48C14.73±1.24B注:数值代表平均值+标准差(n=4),不同的字母代表显著性差异(P<0.05;T检验)。实施例10:拟南芥主茎纤维素含量比较纤维素提取采用硝酸乙醇法。具体步骤为:先将收集的拟南芥主茎粉碎,烘干后经苯醇抽提,称取2g经苯醇抽提的试样,把试样置于250mL洁净干燥的锥形瓶中,加入25mL硝酸乙醇混合液,装上回流装置,放在沸水浴上加热1h。在加热过程中,应随时摇荡锥形瓶,以防止试样跳动。煮沸1h后,移去冷凝管,将锥形瓶自水浴上取下,静置片刻。待残渣沉积瓶底后,用倾泻法滤经已恒重的1G2玻璃滤器,尽量不使试样流出。用真空泵将滤器中的滤液吸干,再用玻璃棒将流入滤器的残渣移入锥形瓶中重复施行上述步骤数次,直至纤维变白为止。最后将锥形瓶内容物全部移入滤器,用10mL硝酸乙醇混合液洗涤残渣,再用热水洗涤,至洗涤液遇甲基橙不呈酸性反应为止。最后用乙醇洗涤两次,吸干洗液,将滤器移入烘箱,于105±2℃烘干至恒重。纤维素含量计算:纤维素含量=(m1-m2)/m0(1-w)×100%,式中m1为绝干纤维素与玻璃滤器的质量,m2为绝干空玻璃滤器质量,m0为风干试样的质量,w为试样的含水量。纤维素含量的结果见表1。从表1中可知,测 定结果证明转基因植株的纤维素含量比cesa4纯合突变体增长了1.16倍,但尚未恢复至野生型程度。实施例11:拟南芥茎段横切片的制作与观察分别选取生长6~7周的野生型、cesa4纯合突变体及转基因植株主茎,在土壤表层以上3cm取1cm茎段,用3%(w/v)的琼脂糖包埋茎段,在LeicaVT1000S震动切片机上切片,厚度40μm,甲苯氨蓝染色1~2min,置于载玻片上于光学显微镜下观察比较并拍照。结果如图5所示,A,B,C分别为cesa4纯合突变体,转基因植株和野生型植株的茎部横切面甲苯胺蓝染色图,箭头所指的部位为木质部细胞,图A中的木质部细胞坍塌变形比较严重,图B中的木质部细胞已得到部分恢复,但尚有部分畸形塌陷的细胞存在,未完全恢复至野生型。上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1