玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用的制作方法
本发明涉及植物基因工程领域,具体涉及玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用。
背景技术:
在拟南芥中MGT家族功能差异显著,拟南芥中MGT1定位于质膜,MGT2定位于液泡膜,MGT10定位于叶绿体,均为高亲和性;MGT3定位于液泡膜,MGT7定位于内质网,均为低亲和性;而MGT5定位于线粒体,为双亲和性,MGT7在根中表达,目前还未见报道来自玉米的MGT10基因。
技术实现要素:
本发明的目的是提供玉米低镁条件下特异性转运镁离子基因ZmMGT10。
本发明的再一目的是提供玉米低镁条件下特异性转运镁离子基因ZmMGT10的应用。
根据本发明的玉米低镁条件下特异性转运镁离子基因ZmMGT10,其核苷酸序列如SEQ ID NO:1所示。
SEQ ID NO:1
TATGGGGAGGTTATCGGGAAGCGGGTCCAGGAAGCCGCCTCCCTTCCTCTCGCCCTCCTCATCATTCTCCTCCTCGTCGTACTCCAAGCGCTCCCGCACTGTTCGGCGCCTCCCGTCGCTGCCCAGGCCGCCTCTCGCGCCGCCGGCGCCGCACCCCGCTGGGAGGACGAGGAGGAGGAAGGCACCCACCCGGCTGTGGATGAGGATGGATCGGTGGGGCAGGTGCGAGGTGTTCATGACTAACGGGGCCTTCGTCGCGGAACGCTCCGGGGTGCACGCGCGCGACCTGCGCATCGTTGGGCCCCTTCTCTCTCGCTGCCCCGGCATCCTCGCTCGAGAGAAGGCCATGGTAATCAATCTAGAGTTCATAAGAGCTATTGTAACTGCGGACGAAGTCCTGCTGCTGGAACCTCTTGCTCAGGAGGTTATCCCTTTCATCGATAAATTGAGGCGGCATTTTCCATTGAAGAGCCTGGAGGTTGATGTTGGGGCCACACAAGTGGGTAATGTGAATGGCAAGCATGCTAAAACTGGTGCAGAGTGTGAGCTCCCCCTCCCCTTTGAGTTTCAGGTGTTGGAGCTCGCCCTAGAAGCTGTATGCTTGTCATTTCATTCAAGCCTATCTGATCTGAATAGGCACACCATCTTTGTGATGGATGAGTTGACTAAGAATGTGAGCACCAGGAATCTTGAACGTGTTCGAAGTCTGAAAAGAAATCTAACCTCCCTGCTCGCTGGTGTGCAAAAGGTCAGAGATGAAGTAGAGCATCTTTTGGATCATAATGAGAATATGGCACAACTGCACCTATCCAGGAAGCAAATAAAATGTCCGCAGGATGAAATTCTACTGGCTTCTGCTGCTTTAAATAGCAATCTTCCTTCAAAAACAAAGCTGGGTACACCAAATTCTGTTGTTAACCAAGCCATGGGCATTGCTATGACTGCGCCACTGGCCGACAATGTAGGAGACCTAGAGATATTACTTGAATCTTATTTCATGCAGCTGGATGGAATTCGCAACAGAATTATGATGGTCCGAGGATATATTGTTGACACAGAAGACTACATCAACATACAACTTGACAACCAACGCAATGAACTCATTCAGTTCCATCTTGTACTGATCATTGTATCATTTGGCATAGCTATGAACACATTGATAGCTGGAGCGTTTGCCATGAACATGCCTCACAATGGGGAGATGAAGAAGTTTGTAGGCCCGTTTTGGCCATTTGTTGGGGCCACATCATCTTTCTGCTTGTTGGTCAGCGTTGTTTTACTGGGATATGCAAGGGGGAACAGGCTACTTGGCAGTTGA
本发明提供了ZmMGT10在低镁浓度下介导植物根对Mg2+的吸收来增强植物对低镁胁迫的抗性。
根据本发明的具体实施方式,提供了包含上述玉米低镁条件下特异性转运镁离子基因ZmMGT10的重组表达载体。
根据本发明的具体实施方式,提供了包含上述玉米低镁条件下特异性转运镁离子基因ZmMGT10的重组细胞系。
根据本发明的具体实施方式,提供了上述玉米低镁条件下特异性转运镁离子基因ZmMGT10的应用,尤其用于增强植物抵抗低镁胁迫的应用。
根据本发明的玉米低镁条件下特异性转运镁离子基因ZmMGT10,在低镁浓度下特异性表达,而低镁影响了植物的正常生长,因此ZmMGT10基因具有增强植物抵抗低镁胁迫的能力。
附图说明
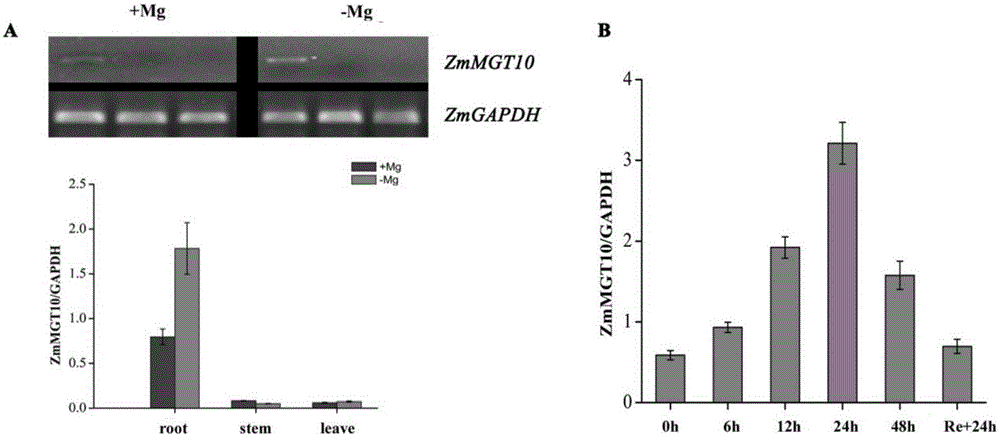
图1显示ZmMGT10基因的表达模式,其中,ZmMGT10在镁供应(+Mg)和镁缺乏(-Mg)在根茎叶中的表达模式;B.ZmMGT10表达对外界镁缺乏的响应。
图2显示显示了ZmMGT10功能互补MM281细菌突变株的生长状况。
图3显示野生型材料和转基因材料低镁胁迫表型,其中,A.野生型材料和转基因材料在正常和低镁条件下的生长表型;B.野生型材料和转基因材料在正常和低镁条件下的主根长度;C.野生型材料和转基因材料在正常和低镁条件下的生物量;D.野生型材料和转基因材料在正常和低镁条件下的叶绿素浓度。
图4显示不同外源Mg2+处理下野生型材料和转基因材料在根和梢中的Mg2+含量,其中,A.根中Mg2+含量B.梢中Mg2+含量。
图5显示野生型和转基因植株的Mg2+吸收,其中,A.时间依赖的野生型和转基因植株的Mg2+吸收;B.浓度依赖的野生型和转因植株的Mg2+吸收。
具体实施方式
实施例1
1、玉米ZmMGT10基因的的克隆
挑选饱满的玉米种子消毒并清洗干净,然后发芽并培养幼苗。釆用Invitrogen公司生产的Trizol试剂提取RNA,利用Takara公司的第一链反转录试剂盒(PrimeScriptⅡ1st Strand cDNA Synthesis Kit)进行反转录合成第一链cDNA。以合成的第一链cDNA为模板,PCR扩增基因的CDS序列(引物序列:forward:5’-ATGGGGAGGTTATCGGGAAG-3’;reverse:5’-TCAACTGCCAAGTAGCCTGTTC-3’)。克隆目的基因,并测序验证。
2、玉米ZmMGT10基因的表达模式
图1显示了ZmMGT10基因的表达模式,其中,“A”为ZmMGT10在镁供应即+Mg条件下的表达模式,“B”为镁缺乏即-Mg调节下在根茎叶中的表达模式。如图1的A所示,ZmMGT10基因主要在根中表达,在茎和叶中不表达或超低表达,ZmMGT10基因在根中的表达受外界Mg2+缺乏强烈诱导。如图1的B所示Mg2+缺乏上调了ZmMGT10基因的表达,且ZmMGT10的表达量在24h达到峰值,表达量几乎是对照0h的6倍,随后ZmMGT10的表达量开始减少。此外,当Mg2+缺乏处理24h小时的植株重新恢复Mg2+供应24h后,ZmMGT10在根中的表达量几乎恢复到了对照0h时的水平。图1所示结果表明,ZmMGT10是一个根特异性表达基因,且其表达响应了早期外界Mg2+缺乏。
3、ZmMGT10具有镁离子转运能力
为了确定ZmMGT10蛋白是否具有Mg2+转运能力,构建了原核表达载体pTrc99A::ZmMGT10,并将其与pTrc99A空载体、pTrc99A::AtMGT1重组质粒载体分别转入鼠伤寒沙门氏菌MM281突变菌株中在含不同浓度Mg2+的1×N-minimal固体培养基上进行功能互补分析,其中CorA、MgtA/MgtB、MgtE基因失活,不能在低于10mM Mg2+条件下生长。图2显示了ZmMGT10功能互补MM281细菌突变株的生长状况。如图2中的A所示,作为阴性对照的MM281和转pTrc99A空载体的MM281菌株在低于10mM Mg2+的培养基上不能生长,而转pTrc99A::AtMGT1作为阳性对照的MM281菌株在Mg2+浓度低至0.1mM培养基上的生长情况与其他高浓度Mg2+条件下的生长情况无明显差异;转pTrc99A::ZmMGT10菌株能在低于0.1mM Mg2+条件下恢复MM281菌株的生长缺陷,但其恢复能力明显要弱于AtMGT1。为进一步验证平板实验结果,监测了MM281、转pTrc99A空载体、pTrc99A::ZmMGT10和pTrc99A::AtMGT1的MM281在含不同浓度Mg2+的1×N-minimal液体培养基中的生长情况。结果如图2中的B所示,不同菌株在液体培养基中的生长情况与平板培养结果一致。这些结果表明,ZmMGT10是一个镁离子转运蛋白,具有转运镁离子的能力。
4、过表达ZmMGT10增强了转基因拟南芥对镁缺乏的容忍
筛选到转35S:ZmMGT10基因的拟南芥阳性后,对野生型和T1代转基因拟南芥进行了低镁条件下的表型观察实验。图3显示了野生型材料和转基因材料低镁胁迫表型。将在MS培养基和含50mg/L卡那霉素MS培养基上生长两周的野生型和转基因拟南芥幼苗转移到全营养液,培养3天,其中,全营养液含2mM MgSO4.7H2O,随后两种材料各分为两组,分别在含2mM MgSO4.7H2O(+Mg)和含0.01mM MgSO4.7H2O(-Mg)的营养液中培养3周后观察表型。结果如图3中的A所示,在正常培养条件下野生型拟南芥植株和转基因拟南芥植株间没有明显的表型差异,而在低镁条件下,野生型植株的生长略弱于转基因植物,且野生型植株出现了心叶变黄的典型的植物镁素缺乏症状。此外,在正常生长条件下的野生型和转基因材料的植株大小略大于低镁条件下植株的大小,且叶子的颜色也略绿于低镁条件植株叶子的颜色。对两种材料的主根长度进一步测量,结果如图3中的B所示,在低镁生长条件下,野生型植株的主根长明显短于转基因植株的主根长;而在正常生长条件下,野生型植株的主根长和转基因植株的主根长无明显差异。此外,在正常生长条件下的两种材料的主根长均明显长于低镁生长条件下两种材料的主根长。生物量分析显示,在低镁生长条件下,野生型植株的鲜重明显低于转基因植株的鲜重;而在正常生长条件下,野生型植株的鲜重和转基因植株的鲜重无明显差异。如图3中的C所示,同样,在正常生长条件下的两种材料的鲜重均明显高于低镁生长条件下两种材料的鲜重。叶片叶绿素浓度分析显示,在低镁生长条件下,野生型植株叶片的叶绿素浓度明显低于转基因植株叶片的叶绿素浓度;而在正常生长条件下,野生型植株叶片的叶绿素浓度与转基因植株叶片的叶绿素浓度间无明显差异。此外,如图3中的D所示,在正常生长条件下的两种材料的叶片叶绿素浓度均高于低镁生长条件下两种材料叶片叶绿素浓度。这些结果表明,低镁影响了植物的正常生长,ZmMGT10基因具有增强植物抵抗低镁胁迫的能力。
5、过表达ZmMGT10增加转基因拟南芥低镁条件下镁离子积累
为了进一步探索在低镁条件下ZmMGT10转基因拟南芥植株具有更强的生长能力是否与它具有更高的Mg2+含量有关,检测了正常生长条件和低镁生长条件下两种材料全株、地上部分、根的Mg含量。将在MS培养基和含50mg/L卡那霉素MS培养基上生长两周的野生型和转基因拟南芥幼苗转移到全营养液培养3天,全营养液含2mM MgSO4.7H2O,随后两种材料各分为两组,分别在含2mM MgSO4.7H2O(+Mg)和含0.01mM MgSO4.7H2O(-Mg)的营养液中培养3周后收获根和梢用于Mg2+含量检测。图4显示不同外源Mg2+处理下野生型材料和转基因材料在根和梢中的Mg2+含量。如图4所示,在2mM Mg2+生长条件下,野生型和转基因植株的根和梢中的Mg2+含量无明显差异。然而,当在0.01mM生长条件下,所有植株在根和梢中的Mg2+含量均明显低于正常生长条件下根和梢中的Mg2+含量,但转基因植株根和梢中的Mg2+含量均明显高于野生型植株。这些结果表明,在低镁生长条件下,ZmMGT10转基因拟南芥植株具有更强的生长能力与它比野生型植株积累更多的Mg2+有关,ZmMGT10介导了低镁条件下根对外界Mg2+的吸收。
6、过表达ZmMGT10增加转基因拟南芥低镁下根对镁离子的吸收
由于植物中的Mg2+主要是通过镁离子转运蛋白从外界吸收后转运进植物体内的,而镁含量测定结果表明,在低镁生长条件下转基因材料镁含量明显高于野生型材料,说明ZmMGT10在低镁条件下介导了根对外界Mg2+的吸收,转基因植株比野生型植株有更强的Mg2+吸收能力。测试野生型植株和转基因株系,用于Mg2+摄取分析。对于时间依赖的Mg2+摄取分析,经Mg2+饥饿处理的野生型和转基因植株在包含0.01mM和2mM MgSO4.7H2O营养液中培养10、30、60、120、180、240min,收获根用于Mg2+含量测定。对于浓度依赖的Mg2+摄取分析,经Mg2+饥饿处理的野生型和转基因植株在包含0、0.01、0.05、0.1、0.5、1、2、3mM MgSO4.7H2O营养液中培养2h,收获根用于Mg2+含量测定。图5显示了野生型和转基因植株的Mg2+吸收。时间依赖的Mg2+摄取分析结果如图5所示,在2mM Mg2+生长条件下,野生型植株和转基因植株具有相似的Mg2+积累模式,而在0.01mM Mg2+条件下,野生型和转基因植株Mg2+积累量均明显低于2mM条件下植株的Mg2+积累量,但转基因植株的Mg2+积累量明显高于野生型植株,表明过表达ZmMGT10增强了低镁条件下拟南芥根对Mg2+的吸收。浓度依赖的Mg2+摄取分析结果如图5所示,在低镁浓度条件下,转基因植株的Mg2+摄取率明显高于野生型植株,但在高镁浓度条件下,两种材料的Mg2+摄取率无明显差异,表明ZmMGT10是在低镁浓度下介导植物根对Mg2+的吸收。所有这些结果表明,ZmMGT10通过在低镁浓度下介导植物根对Mg2+的吸收来增强植物对低镁胁迫的抗性。
<110> 四川农业大学
<120> 玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用
<160> 1
<210> 1
<211> 1318
<212> DNA
<213> 链霉菌
<400> 1
tatggggagg ttatcgggaa gcgggtccag gaagccgcct cccttcctct cgccctcctc 60
atcattctcc tcctcgtcgt actccaagcg ctcccgcact gttcggcgcc tcccgtcgct 120
gcccaggccg cctctcgcgc cgccggcgcc gcaccccgct gggaggacga ggaggaggaa 180
ggcacccacc cggctgtgga tgaggatgga tcggtggggc aggtgcgagg tgttcatgac 240
taacggggcc ttcgtcgcgg aacgctccgg ggtgcacgcg cgcgacctgc gcatcgttgg 300
gccccttctc tctcgctgcc ccggcatcct cgctcgagag aaggccatgg taatcaatct 360
agagttcata agagctattg taactgcgga cgaagtcctg ctgctggaac ctcttgctca 420
ggaggttatc cctttcatcg ataaattgag gcggcatttt ccattgaaga gcctggaggt 480
tgatgttggg gccacacaag tgggtaatgt gaatggcaag catgctaaaa ctggtgcaga 540
gtgtgagctc cccctcccct ttgagtttca ggtgttggag ctcgccctag aagctgtatg 600
cttgtcattt cattcaagcc tatctgatct gaataggcac accatctttg tgatggatga 660
gttgactaag aatgtgagca ccaggaatct tgaacgtgtt cgaagtctga aaagaaatct 720
aacctccctg ctcgctggtg tgcaaaaggt cagagatgaa gtagagcatc ttttggatca 780
taatgagaat atggcacaac tgcacctatc caggaagcaa ataaaatgtc cgcaggatga 840
aattctactg gcttctgctg ctttaaatag caatcttcct tcaaaaacaa agctgggtac 900
accaaattct gttgttaacc aagccatggg cattgctatg actgcgccac tggccgacaa 960
tgtaggagac ctagagatat tacttgaatc ttatttcatg cagctggatg gaattcgcaa 1020
cagaattatg atggtccgag gatatattgt tgacacagaa gactacatca acatacaact 1080
tgacaaccaa cgcaatgaac tcattcagtt ccatcttgta ctgatcattg tatcatttgg 1140
catagctatg aacacattga tagctggagc gtttgccatg aacatgcctc acaatgggga 1200
gatgaagaag tttgtaggcc cgttttggcc atttgttggg gccacatcat ctttctgctt 1260
gttggtcagc gttgttttac tgggatatgc aagggggaac aggctacttg gcagttga 1318
- 还没有人留言评论。精彩留言会获得点赞!
- 玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用的制造方法与工艺
- CRISPR‑Cas9特异性敲除猪SLA‑3基因的方法及用于特异性靶向SLA‑3基因的sgRNA与制造工艺
- 毛酸浆的分子特异性标记引物及鉴别方法与制造工艺
- 一种检测CYP2C9基因多态性的ARMS引物的制造方法与工艺
- 构建在海马体区域特异性敲除IKKα基因的小鼠模型的方法及打靶载体和试剂盒与制造工艺
- 使用重叠非等位基因特异性引物和等位基因特异性阻断剂寡核苷酸的组合物进行等位基因特异性扩增的制造方法与工艺
- 一种利用线粒体D-loop区特异区域鉴定鼋的标记引物及方法
- 一种用于检测大豆拟茎点种腐病菌的特异性lamp引物组合物及其应用
- 一种针对膨胀污泥中Eikelboom Type 0092型丝状菌群特异性引物和方法
- 脯氨酸特异性蛋白酶基因及其应用
- 一种泰国库蠓的特异基因及其分子鉴定方法
- 一种美丽库蠓特异基因及其分子鉴定方法
- 一种特异于禽类原始生殖细胞的基因转移系统及方法
- CRISPR-Cas9特异性敲除猪SALL1基因的方法及用于特异性靶向SALL1基因的sgRNA的制作方法
- CRISPR-Cas9特异性敲除猪CMAH基因的方法及用于特异性靶向CMAH基因的sgRNA的制作方法
- CRISPR-Cas9特异性敲除猪SLA-2基因的方法及用于特异性靶向SLA-2基因的sgRNA的制作方法
- 一种安宁铗蠓的特异基因及其分子鉴定方法
- CRISPR-Cas9特异性敲除猪GGTA1基因的方法及用于特异性靶向GGTA1基因的sgRNA的制作方法
- 一种蓝腹库蠓的特异基因及其分子鉴定方法
- 扬子鳄抗病毒潜力检测的ⅰ类mhc基因的特异性引物及分型方法
- 一种检测水稻粒宽等位基因的特异性PCR分子标记和方法与流程
- 丝状真菌中多个基因拷贝的同时位点特异性整合的制造方法与工艺
- 一种特异性敲减人BRCC3基因慢病毒的构建及在肺癌中的应用的制造方法与工艺
- 烟粉虱MED隐种高温耐受相关基因特异性探针及其应用的制造方法与工艺
- CRISPR/SaCas9特异性敲除人CXCR4基因的方法与流程
- 玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用的制造方法与工艺
- CRISPR‑Cas9特异性敲除猪SLA‑3基因的方法及用于特异性靶向SLA‑3基因的sgRNA与制造工艺
- 构建在海马体区域特异性敲除IKKα基因的小鼠模型的方法及打靶载体和试剂盒与制造工艺
- 使用重叠非等位基因特异性引物和等位基因特异性阻断剂寡核苷酸的组合物进行等位基因特异性扩增的制造方法与工艺
- 用于检测烟草对tmv抗性的n基因特异性引物对、检测方法及试剂盒的制作方法