CD137特异性的蛋白的制作方法
背景技术:
cd137是一种肿瘤坏死因子受体(tnfr)超家族分子,其活性可能参与许多免疫介导的自身免疫性和炎性疾病。cd137也是癌症免疫治疗的靶标。
已证明cd137信号传导对抗原免疫响应的维持和扩张,以及对记忆t细胞的产生是基本的。鼠和人t细胞的许多研究表明,cd137促进增强的细胞增殖、存活和细胞因子产生(croft,2009,natrevimmunol9:271-285)。已有研究表明,一些cd137激动剂mab增加共刺激分子表达,并显著增强溶细胞性t淋巴细胞响应,从而导致在各种模型中的抗肿瘤功效。已经证明了cd137激动剂mab在预防和治疗方面的功效。此外,cd137单一疗法和联合疗法肿瘤模型已经建立了可持久的抗肿瘤保护性t细胞记忆响应(lynch,2008,immunolrev.22:277-286)。在多种本领域公认的自身免疫模型中,cd137激动剂也已显示出抑制自身免疫反应(vinay,2006,jmolmed84:726-736)。cd137的这种双重活性呈现了提供抗肿瘤活性同时抑制可能与破坏免疫耐受的免疫治疗方法相关的自身免疫性副作用的潜力。
因此,基于cd137在调节免疫响应中的作用,对结合人cd137、增加cd137介导的响应并由此为治疗或预防各种疾病和病症如癌症、感染性疾病和自身免疫性疾病提供潜在治疗的化合物存在长期未满足的需求。
因此,本发明的一个目的是提供这种化合物,其是源自脂质运载蛋白的突变蛋白。各种脂质运载蛋白的突变蛋白是一类快速扩张的治疗剂,并且通过高度复杂的人工工程可将各种脂质运载蛋白的突变蛋白构建为对与野生型脂质运载蛋白的天然配体不同的靶标显示出高亲和力和特异性(参见如,wo99/16873、wo00/75308、wo03/029463、wo03/029471和wo05/19256)。
定义
以下所列定义了在本说明书中使用的术语、短语和缩写。本文列出和定义的所有术语旨在涵盖所有语法形式。
如本文所使用,除另有规定外,“cd137”是指人cd137。cd137也称为“4-1bb”或“肿瘤坏死因子受体超家族成员9(tnfrsf9)”或“淋巴细胞活化诱导的(ila)”。人cd137是指由uniprotq07011定义的全长蛋白,其片段或其变体。
如本文所使用,“可检测的亲和力”是指以通常至少约10-5m或更低的亲和力常数结合选定靶标的能力。较低的亲和力通常不再能通过常规方法例如elisa测量,因此是次要的。
如本文所使用,可以通过本领域技术人员已知的多种方法测量本发明的蛋白(如,脂质运载蛋白的突变蛋白)或其融合多肽与选定靶标(在本发明中为cd137)的“结合亲和力”(并由此可以测定突变蛋白-配体复合物的kd值)。这种方法包括但不限于,荧光滴定、竞争elisa、如等温滴定量热法(itc)的量热法和表面等离子体共振(biacore)。这样的方法在本领域中是公知的,并且其实例也在下文详述。
还注意到,相应结合剂与其配体之间的复合物的形成受多个不同因素的影响,如各自结合配偶体的浓度、竞争者的存在、所采用的缓冲系统的ph和离子强度以及用于测定解离常数kd的实验方法(例如荧光滴定、竞争elisa或表面等离子体共振,仅举几个例子),或甚至用于评估实验数据的数学算法。
因此,本领域技术人员也清楚kd值(各自结合剂与其靶标/配体之间形成的复合物的解离常数)可以在某个实验范围内变化,这取决于用于测定特定脂质运载蛋白突变蛋白对给定配体的亲和力的方法和实验设置。这意味着,所测得的kd值可能会有轻微的偏差或公差范围,例如,取决于该kd值是通过表面等离子体共振(biacore)、竞争elisa、还是“直接elisa”测定的。
如本文所用,“突变蛋白”、“突变的”实体(蛋白或核酸)或“突变体”是指与天然存在(野生型)的核酸或蛋白的“参考”骨架相比,一个或多个核苷酸或氨基酸的交换、缺失或插入。所述术语还包含如本文所述的突变蛋白的片段和变体。优选地,本发明的脂质运载蛋白突变蛋白、其片段或变体保留了本文所述的结合cd137的功能。
本文所用的与本发明的突变蛋白有关的术语“片段”涉及n-末端和/或c-末端缩短的(即缺少n-末端和/或c-末端氨基酸中的至少一个)源自全长成熟人泪液脂质运载蛋白的蛋白质或肽。这种片段可包括成熟脂质运载蛋白一级序列的至少10个、更多如20或30个或者更多个连续氨基酸,并且通常在成熟脂质运载蛋白的免疫实验中可检测到。一般而言,如本文中关于本发明的脂质运载蛋白突变蛋白或根据本发明的组合物或本文所述的融合蛋白的相应蛋白配体cd137所使用的术语“片段”涉及n-末端和/或c-末端缩短的蛋白或肽配体,其保留了全长配体被根据本发明的突变蛋白识别和/或结合的能力。
本文所用的术语“诱变”是指,选择实验条件,以使得天然存在于成熟脂质运载蛋白给定序列位置处的氨基酸可被至少一个不存在于各天然多肽序列中的此特定位置处的氨基酸置换。术语“诱变”还包括通过缺失或插入一个或多个氨基酸而(另外的)改变序列区段的长度。因此,以下情况在本发明的范围内:例如,一个在选定序列位置处的氨基酸被三个随机突变的延伸段(stretch)替代,从而相比于野生型蛋白相应区段的长度,导致两个氨基酸残基的插入。这样的插入或缺失可以彼此独立地引入到在本发明中可以进行诱变的任何肽区段中。在本发明的一个示例性实施方式中,几个突变的插入可以引入到选定的脂质运载蛋白骨架的ab环中(参见以全文引用并入本文的国际专利申请wo2005/019256)。
术语“随机诱变”指无预定的单个氨基酸(突变)存在于某个序列位置处,而是至少两个氨基酸可以在诱变期间以某概率掺入在预限定的序列位置处。
“同一性”指的是衡量它们的相似性或关系的序列特性。本发明所用的术语“序列同一性”或“同一性”是指在本发明多肽序列与所讨论的序列(同源性)比对之后,相对于这两个序列中较长一个的残基数量的配对相同残基的百分比。通过将相同氨基酸残基的数量除以残基总数再使结果乘以100来量度序列同一性。
本文所用的术语“同源性”是其一般含义,并包括相同氨基酸以及认为是在本发明的多肽(如本发明的任意脂质运载蛋白突变蛋白)线性氨基酸序列中的等价位置处的保守置换的氨基酸(例如天冬氨酸残基交换谷氨酸残基)。
可以采用例如程序blastp,blastp2.2.5版(2002年11月16日;参见altschul,s.f.等人(1997)nucl.acidsres.25,3389-3402)测定序列同源性或序列同一性百分比。在本实施方案中,同源性百分比基于包括前肽序列的完整多肽序列比对(矩阵:blosum62;间隙权重:11.1;截止值设置为10-3),优选地在配对比较中使用野生型蛋白骨架作为参考。blastp程序输出的结果表示为以“阳性”(同源氨基酸)数除以程序选定的用于比对的氨基酸总数来计算的百分比。
具体而言,为了确定与野生型脂质运载蛋白不同的脂质运载蛋白(突变蛋白)氨基酸序列的氨基酸残基是否对应于野生型脂质运载蛋白氨基酸序列中的某一位置,技术人员可利用本领域熟知的手段和方法,如手动地或通过使用计算机程序例如使用blast2.0(其代表基本局部比对搜索工具)或clustalw或任意适用于产生序列比对的其它合适程序进行比对。因此,野生型脂质运载蛋白可作为“受试序列”或“参考序列”,而本文所述的与野生型脂质运载蛋白不同的脂质运载蛋白的氨基酸序列作为“查询序列”。本文中术语“参考序列”和“野生型序列”可互换使用。优选地,野生型脂质运载蛋白分别示于seqidno:1(tlc)或seqidno:2(ngal)。分别根据本发明的脂质运载蛋白突变蛋白是基于tlc还是ngal,可将相应的野生型脂质运载蛋白用作参考序列或野生型序列。
“间隙”是添加或缺失氨基酸所导致的比对中的空间。因此,两份完全相同的序列具有100%的同一性,但较不高度保守且具有缺失、添加或替换的序列可能具有较低程度的序列同一性。本领域的技术人员将会认识到,可以得到数个计算机程序,以用来使用标准参数测定序列同一性,例如blast(altschul等人(1997)nucleicacidsres.25,3389-3402)、blast2(altschul等人(1990)j.mol.biol.215,403-410)和smith-waterman(smith等人(1981)j.mol.biol.147,195-197)。
本发明中使用的术语“变体”涉及包含氨基酸序列修饰的蛋白质或肽的衍生物,所述氨基酸序列修饰例如通过置换、缺失、插入或化学修饰进行。在一些实施方式中,这种修饰确实不会降低蛋白质或肽的功能。这种变体包括蛋白质,其中一个或多个氨基酸被它们各自的d-立体异构体或除天然存在的20个氨基酸外的氨基酸(比如,例如,鸟氨酸、羟脯氨酸、瓜氨酸,高丝氨酸,羟赖氨酸、戊氨酸)替换。然而,这样的置换也可以是保守的,即氨基酸残基被替代为化学相似的氨基酸残基。保守置换的实例为以下组成员之间的替代:1)丙氨酸、丝氨酸和苏氨酸;2)天冬氨酸和谷氨酸;3)天冬酰胺和谷氨酰胺;4)精氨酸和赖氨酸;5)异亮氨酸、亮氨酸、甲硫氨酸和缬氨酸;和6)苯丙氨酸、酪氨酸和色氨酸。本文中关于本发明的脂质运载蛋白突变蛋白或根据本发明的组合物或本文所述的融合蛋白的相应蛋白配体cd137所使用的术语“变体”分别涉及分别与野生型cd137蛋白(例如本文所述的以swissprot保存的cd137参考蛋白)相比具有一个或多个(比如1、2、3、4、5、6、7、8、9、10、12、14、16、18、20、22、24、26、28、30、40、50、60、70、80或更多个)氨基酸置换、缺失和/或插入的cd137或其片段。cd137变体与野生型人cd137(例如本文所述的以swissprot保存的cd137参考蛋白)分别具有优选地至少50%、60%、70%、80%、85%、90%或95%的氨基酸同一性。
“天然序列”脂质运载蛋白是指与源自自然界的相应多肽具有相同氨基酸序列的脂质运载蛋白。因此,天然序列脂质运载蛋白可以具有来自任何生物体,特别是哺乳动物的相应天然存在的脂质运载蛋白的氨基酸序列。这种天然序列多肽可以从自然界中分离或者可以通过重组或合成手段产生。术语“天然序列”多肽具体涵盖脂质运载蛋白的天然存在的截短的或分泌的形式、天然存在的变体形式(例如替代地,脂质运载蛋白的剪接形式和天然存在的等位基因变体)。多肽“变体”是指与天然序列多肽具有至少约50%、60%、70%或至少约85%的氨基酸序列同一性的生物活性多肽。这种变体包括,例如,在多肽的n-或c-末端添加或缺失一个或多个氨基酸残基的多肽。通常,变体与天然序列多肽具有至少约70%,包括至少约80%,比如至少约85%的氨基酸序列同一性,包括至少约90%的氨基酸序列同一性或至少约95%的氨基酸序列同一性。作为示例性实例,可以在本发明的泪液脂质运载蛋白(tlc)突变蛋白中使前4个n-末端氨基酸残基(his-his-leu-leu)和最后2个c-末端氨基酸残基(ser-asp)缺失而不影响该蛋白的生物功能,如seqidnos:5-11。此外,作为另一个示例性实例,像对seqidno:16那样(lys-asp-pro,位置46-48),可以在本发明脂质运载蛋白2(ngal)突变蛋白中使某些氨基酸残基缺失而不影响该蛋白的生物功能。
当根据本发明使用时,术语“位置”是指本文描述的氨基酸序列中的氨基酸的位置或本文描述的核酸序列中的核苷酸的位置。为了理解本文中在一种或多种脂质运载蛋白突变蛋白氨基酸序列位置的上下文中使用的术语“对应”或“相应”,相应位置不仅由前述核苷酸/氨基酸的数量决定。因此,由于在(突变体或野生型)脂质运载蛋白的其它位置处的氨基酸缺失或添加,则可被置换的根据本发明的给定氨基酸的位置可能改变。类似地,由于在突变蛋白或野生型脂质运载蛋白5’-非翻译区(utr)(包括启动子和/或任何其它调节序列或基因(包括外显子和内含子))的其它位置处的核苷酸缺失或添加,则可被置换的根据本发明的给定核苷酸的位置可能改变。
因此,对于根据本发明的相应位置,优选地理解,核苷酸/氨基酸的位置在指定数字方面可以不同于类似的相邻核苷酸/氨基酸,但是所述一个或多个相应位置仍包含可被交换、缺失或添加的所述相邻核苷酸/氨基酸。
此外,对于基于根据本发明的参考骨架的脂质运载蛋白突变蛋白中的相应位置,优选地理解,核苷酸/氨基酸的位置在结构上对应于(突变体或野生型)脂质运载蛋白中的其它位置,尽管这些位置在指定数字方面可能不同,如技术人员鉴于脂质运载蛋白之间高度保守的整体折叠形式所理解地。
术语“白蛋白”包括所有的哺乳动物白蛋白,例如人血清白蛋白或牛血清白蛋白或大鼠血清白蛋白。
本文针对非天然靶标使用的术语“有机分子”或“小有机分子”表示包含至少两个碳原子的有机分子,但是优选地,它包含不超过7或12个可旋转碳键,具有100到2000道尔顿、优选地100到1000道尔顿的分子量范围,且任选地包含一个或两个金属原子。
本文使用的词“检测/可检测的”理解为在定量和定性水平上以及它们的组合。从而其包括目标分子的定量、半定量和定性测定。
“受试者”是指脊椎动物,优选是哺乳动物,更优选是人。本文所用术语“哺乳动物”是指归类为哺乳动物的任何动物,包括但不限于人、家畜和农用动物以及动物园、运动或宠物动物,例如绵羊、狗、马、猫、牛、大鼠、猪、如食蟹猕猴(cynomolgousmonkey)的猿等等,仅举几个示例性实例。优选地,本文所述哺乳动物是人。
“有效量”是足够实现有益或者所希望的结果的量。在一次或多次施用中可以施用有效量。
将“样品”定义为从任何受试者中采集的生物样品。生物样品包括但不限于血液、血清、尿液、粪便、精液或组织。
附图说明
图1:提供了通过表面等离子体共振(spr)对各种代表性脂质运载蛋白突变蛋白(图中所示的seqidno)与作为靶标的人cd137(fc-融合)的相互作用的结合-速率(on-rate)和解离-速率(off-rate)的典型测量。靶标通过抗-人igg-fc抗体固定,而抗-人igg-fc抗体使用标准胺偶联化学固定在传感器芯片上。使用脂质运载蛋白突变蛋白作为以不同浓度流经芯片表面的可溶性分析物。对于测试的所有突变蛋白,针对人靶标即人cd137-fc融合蛋白(hucd137-fc),存在清晰的spr结合信号,而阴性对照seqidno:3和seqidno:4未显示结合。表1提供了自对所有seqidno所示的数据拟合(1∶1结合模型)中得到的解离常数。
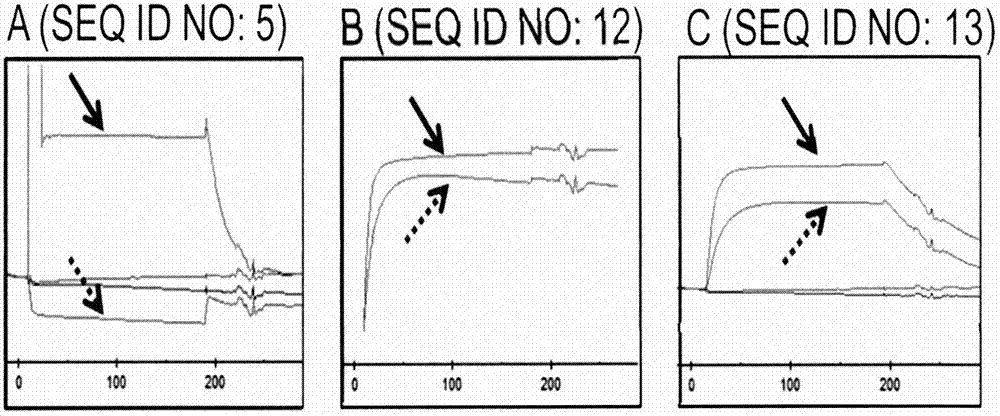
图2:提供了基于spr的实验的代表性实例,该实验设计为研究seqidno:5、seqidno:12和seqidno:13的突变蛋白是否干扰cd137配体(cd137l)与cd137的结合。通过在spr传感器芯片上产生cd137与cd137l的复合物,并检查测试的脂质运载蛋白突变蛋白是否能够结合该复合物来进行研究。作为参考,在不存在cd137l情况下,cd137与脂质运载蛋白突变蛋白一起温育。在附图中,仅提供了传感图的相关片段。采用实茎箭头(arrowwithasolidstem)标记了相应脂质运载蛋白突变蛋白与单独hucd137-fc结合的spr迹线。采用虚茎箭头(arrowwithabrokenstem)标记了相应脂质运载蛋白突变蛋白与已用cd137l饱和的hucd137-fc的结合的spr迹线。图2(a)显示,在存在cd137l下,seqidno:5不能结合hucd137-fc。图2(b)和图2(c)显示,在不存在和存在cd137l下,seqidno:12和seqidno:13均以非常相似的响应结合hucd137-fc,这表明在两种脂质运载蛋白突变蛋白和cd137l之间没有结合上的竞争。
图3:显示了进行荧光激活细胞分选(facs)研究的代表性实例,实施这些研究是为了评估代表性脂质运载蛋白突变蛋白(图中所示的seqidno)与哺乳动物细胞上表达的人cd137的特异性结合。将模拟-转染细胞作为阴性对照。
图4:描述了t细胞活化分析的结果,实施该t细胞活化分析是为了评估当涂布在塑料皿上时,通过诱导cd137信号传导的聚集(clustering)和伴随下游信号传导,一组代表性cd137-结合的脂质运载蛋白突变蛋白(seqidno:13、seqidno:14和seqidno:15)共刺激t细胞响应的能力。此外,检测了通过与可溶性脂质运载蛋白突变蛋白一起温育的t细胞的活化,从而研究相应的结合剂是否在不存在聚集的情况下展示出激动活性。在图4(a)中,将脂质运载蛋白突变蛋白和抗-人-cd3抗体一起涂布至塑料皿上,并随后在可溶性抗-人cd28存在下在涂布的表面上温育纯化的t细胞。在图4(b)中,将抗-人-cd3抗体涂布至塑料皿上,并随后在溶液中的脂质运载蛋白突变蛋白和可溶性抗-人cd28存在下在涂布的表面上温育纯化的t细胞。两种情况下,将上清液白细胞介素2(il-2)水平作为读出结果。采用seqidno:4作为阴性对照。在图4(a)的实验中,与阴性对照seqidno:4相比,由于seqidno:13、seqidno:14和seqidno:15的脂质运载蛋白突变蛋白的t细胞活化,上清液中il-2的浓度明显增加。对于图4(b)中利用溶液中的脂质运载蛋白突变蛋白的实验,与阴性对照seqidno:4相比,对于所测试的任何脂质运载蛋白突变蛋白,上清液中il-2浓度均未显著增加。总体而言,图4(a)和图4(b)显示所测试的脂质运载蛋白突变蛋白展现了所需的行为:通过塑料涂布的抗-cd137突变蛋白在t细胞表面上聚集cd137导致了所期望的t细胞共刺激,而相应的在溶液中的突变蛋白(如实施例4和图3所示,尽管结合cd137)未诱导任何t细胞共刺激。
图5:提供了采用seqidno:13的cd137-结合脂质运载蛋白突变蛋白作为测试分子的t细胞活化试验的结果。将seqidno:4用作阴性对照。采用另外的、次优的抗-cd3和抗-cd28刺激t细胞进行该实验,其中图5(a)中的读出结果为采用4hbrdu脉冲温育三天后t细胞的继续增殖,图5(c)为上清液中il-2浓度和图5(e)为上清液中ifn-g水平。可选地,仅采用次优的抗-cd3浓度,图5(b)中的读出结果为继续增殖,图5(d)为上清液中il-2浓度和图5(f)为上清液中ifn-g水平。这一实验证明了在采用抗-cd3/抗-cd28刺激和单独抗-cd3刺激下,增殖、il-2和ifn-γ水平的seqidno:13剂量依赖性增加。
具体实施方式
如本文所用,将“脂质运载蛋白”定义为重量为约18-20kda的单体蛋白,其具有圆柱形β折叠片超二级结构区,所述结构区包含通过在一端的多个(优选为4个)环逐对连接的多个(优选为8个)β链以由此限定结合口袋。在脂质运载蛋白家族成员中,正是所述环在本应刚性的脂质运载蛋白骨架中的这种多样性产生多种不同的结合模式,每一种模式能够容纳不同大小、形状和化学特征的靶标(例如上文的flower,d.r.(1996);上文的flower,d.r.等人(2000),或skerra,a.(2000)biochim.biophys.acta1482,337-350中的综述)。事实上,蛋白的脂质运载蛋白家族已经天然进化为结合范围广泛的配体,具有非常低水平的整体序列保守性(通常具有少于20%的序列同一性),但保留高保守性的整体折叠模式。不同脂质运载蛋白中位置间的对应关系为本领域技术人员所熟知。例如,见,美国专利no.7,250,297。
如上所述,脂质运载蛋白是由其超二级结构定义的多肽,超二级结构即包含通过在一端的4个环逐对连接的8个β链以由此限定结合口袋的圆柱形β折叠片超二级结构区。本发明不局限于本文具体公开的脂质运载蛋白突变蛋白。在这一方面,本发明涉及具有圆柱形β折叠片超二级结构区的脂质运载蛋白突变蛋白,该结构区包含通过在一端的4个环逐对连接的8个β链以由此限定结合口袋;其中所述4个环中的至少3个的每一个的至少一个氨基酸已经突变,并且其中所述脂质运载蛋白以可检测的亲和力有效结合cd137。
在一个特别的实施方案中,本发明公开的脂质运载蛋白突变蛋白是人泪液脂质运载蛋白(tlpc或tlc)(也称为脂质运载蛋白-1、泪液前-白蛋白或vonebner腺蛋白)的突变蛋白。本文所用术语“人泪液脂质运载蛋白”或“tlc”或“脂质运载蛋白-1”指具有swiss-prot/uniprot数据库登录号p31025(同种型1)的成熟的人泪液脂质运载蛋白。可将示于swiss-prot/uniprot数据库登录号p31025的氨基酸序列用作优选的“参考序列”,更优选地,将示于seqidno:1的氨基酸序列用作参考序列。
在另一特别的实施方案中,本文公开的脂质运载蛋白突变蛋白是人脂质运载蛋白2的突变蛋白。本文所用术语“人脂质运载蛋白2”或“人lcn2”或“人ngal”指具有swiss-prot/uniprot数据库登录号p80188的成熟的人嗜中性粒细胞明胶酶相关脂质运载蛋白(ngal)。本发明的人脂质运载蛋白2突变蛋白此处还可命名为“hngal突变蛋白”。可将示于swiss-prot/uniprot数据库登录号p80188的氨基酸序列用作优选的“参考序列”,更优选地,将示于seqidno:2的氨基酸序列用作参考序列。
在一些实施方案中,以可检测的亲和力结合cd137的脂质运载蛋白突变蛋白可包含至少一个用另一个氨基酸(例如丝氨酸残基)置换天然半胱氨酸残基的氨基酸置换。在一些其它实施方案中,以可检测的亲和力结合cd137的脂质运载蛋白突变蛋白可包含置换野生型脂质运载蛋白的一个或多个氨基酸的一个或多个非天然半胱氨酸残基。在进一步特别的实施方案中,根据本发明的脂质运载蛋白突变蛋白包含至少两个用半胱氨酸残基置换天然氨基酸的氨基酸置换,从而形成一个或多个半胱氨酸桥。在一些实施方案中,所述半胱氨酸桥可以连接至少两个环区。根据flower(flower,1996,上文;flower等人,2000,上文)和breustedt等人(2005,上文)在本文中使用这些区的定义。在相关的实施方案中,本发明教导了一种或多种脂质运载蛋白突变蛋白,其能够通过结合cd137来活化cd137的下游信号传导通路。
本发明的针对cd137或cd137特异性的蛋白包含任何数目的基于所定义的蛋白质骨架的特异性结合蛋白突变蛋白。优选地,交换、缺失或插入的核苷酸或氨基酸的数目分别是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20或更多(比如25、30、35、40、45或50),其中优选的是1、2、3、4、5、6、7、8、9、10或11和甚至更优选的是9、10或11。然而,优选的是本发明的脂质运载蛋白突变蛋白仍然能够结合cd137。
一方面,本发明包含以至少可检测的亲和力结合cd137的各种脂质运载蛋白突变蛋白。在此意义上,可认为cd137是参考野生型脂质运载蛋白的非天然配体,其中“非天然配体”是指在生理条件下不结合野生型脂质运载蛋白的化合物。通过在某些序列位置处用一个或多个突变工程化野生型脂质运载蛋白,本发明的发明人已经证明对非天然配体即cd137的高亲和力和高特异性是可能的。在一些实施方案中,在编码野生型脂质运载蛋白上某些序列位置处的1、2、3、4、5、6、7、8、9、10、11、12或甚至更多个核苷酸三联体上,可以通过用核苷酸三联体的亚集(subset)在这些位置处的置换来实施随机诱变。
此外,本发明的脂质运载蛋白突变蛋白可在对应于参考脂质运载蛋白线性多肽序列的某些序列位置的任意一个或多个(包括至少任意1个、2、3、4、5、6、7、8、9、10、11或12个)序列位置处具有突变的氨基酸残基。
在突变的氨基酸序列位置之外,本发明的蛋白可包含“亲本”蛋白骨架(例如脂质运载蛋白)的野生型(天然)氨基酸序列。在一些实施方案中,根据本发明的脂质运载蛋白突变蛋白还可在一个或多个序列位置处携带一个或多个氨基酸突变,只要这种突变不妨碍或干扰、至少基本上不妨碍或干扰突变蛋白的结合活性和折叠。可以采用已建立的标准方法(sambrook,j.等人(2001)molecularcloning:alaboratorymanual,3rded.,coldspringharborlaboratorypress,coldspringharbor,ny)在dna水平上非常简单地完成这种突变。氨基酸序列改变的示例性实例是插入或缺失以及氨基酸置换。这种置换可以是保守的,即氨基酸残基被化学性质(特别是关于极性以及尺寸)相似的氨基酸残基替换。保守置换的实例是以下组的成员之间的替代:1)丙氨酸、丝氨酸和苏氨酸;2)天冬氨酸和谷氨酸;3)天冬酰胺和谷氨酰胺;4)精氨酸和赖氨酸;5)异亮氨酸、亮氨酸、甲硫氨酸和缬氨酸;和6)苯丙氨酸、酪氨酸和色氨酸。另一方面,也可以将非-保守性改变引入氨基酸序列。此外,代替替换单一氨基酸残基,还可以使人泪液脂质运载蛋白的一级结构插入或缺失一个或多个连续氨基酸,只要这些缺失或插入导致稳定的折叠/功能的突变蛋白(例如,具有截短的n-和c-末端的tlc突变蛋白)。在这种突变蛋白中,例如,在多肽的n-或c-末端添加或缺失一个或多个氨基酸残基。一般而言,这种突变蛋白与成熟人泪液脂质运载蛋白的氨基酸序列具有约至少70%(包括至少约80%,比如至少约85%)的氨基酸序列同一性。作为示例性实例,本发明还涵盖如上定义的tlc突变蛋白,其中成熟人泪液脂质运载蛋白序列的前4个n-末端氨基酸残基(his-his-leu-leu;位置1-4)和/或成熟人泪液脂质运载蛋白线性多肽序列的最后2个c-末端氨基酸残基(ser-asp;位置157-158)缺失(seqidnos:5-11)。此外,作为另一个示例性实例,本发明还涵盖如上定义的ngal突变蛋白,其中成熟人泪液脂质运载蛋白2(hngal)线性多肽序列的氨基酸残基(lys-asp-pro;位置46-48)缺失(seqidno:16)。
当比较与其它脂质运载蛋白的序列同一性时,本文公开的脂质运载蛋白突变蛋白的氨基酸序列与参考脂质运载蛋白具有高序列同一性。在该一般情况下,本发明的脂质运载蛋白突变蛋白的氨基酸序列至少基本上类似于参考脂质运载蛋白的氨基酸序列,前提是在比对中可能存在由氨基酸的添加或缺失导致的间隙(如下所定义)。在一些实施方案中,本发明的基本上类似于参考脂质运载蛋白的序列的脂质运载蛋白突变蛋白的相应序列与参考脂质运载蛋白的序列具有至少70%的同一性或序列同源性、至少75%的同一性或序列同源性、至少80%的同一性或序列同源性、至少82%的同一性或序列同源性、至少85%的同一性或序列同源性、至少87%的同一性或序列同源性或者至少90%的同一性或序列同源性(包括至少95%的同一性或序列同源性),前提是保留改变的位置或序列并且一个或多个间隙是可能的。
如本文所使用,由于结合特异性不是绝对的,而是一种相对性质,因此,若本发明的脂质运载蛋白突变蛋白能够区分靶标和一个或多个参考靶标,则其“特异性结合”这种靶标(例如,cd137)。例如根据蛋白质印迹、elisa-、ria-、ecl-、irma-试验、facs、ihc和肽扫描来测定“特异性结合”。
在一个实施方案中,本发明的脂质运载蛋白突变蛋白在其n-末端和/或其c-末端与融合配偶体融合,该融合配偶体是延长所述突变蛋白血清半衰期的蛋白结构域。在进一步特别的实施方案中,该蛋白结构域是免疫球蛋白的fc部分、免疫球蛋白的ch3结构域、免疫球蛋白的ch4结构域、白蛋白结合肽或白蛋白结合蛋白。
在另一个实施方案中,本发明的脂质运载蛋白突变蛋白与延长所述突变蛋白血清半衰期的化合物缀合。更优选地,所述突变蛋白与选自以下的化合物缀合:聚亚烷基二醇分子、羟乙基淀粉、免疫球蛋白的fc部分、免疫球蛋白的ch3结构域、免疫球蛋白的ch4结构域、白蛋白结合肽和白蛋白结合蛋白。
在又一个实施方案中,本发明涉及包含编码本文公开的脂质运载蛋白突变蛋白的核苷酸序列的核酸分子。本发明涵盖包含所述核酸分子的宿主细胞。
a.cd137特异性脂质运载蛋白突变蛋白
一方面,本发明提供结合cd137的人脂质运载蛋白突变蛋白及其有益用途。本发明还提供制备本文所述的cd137结合蛋白以及包含这种蛋白的组合物的方法。本发明的cd137结合蛋白及其组合物可用于检测样品中cd137的方法中或用于结合受试者中cd137的方法中。之前未描述过这种具有伴随本发明提供用途的这些特征的人脂质运载蛋白突变蛋白。
1.示例性cd137特异性脂质运载蛋白突变蛋白
本发明的一个实施方案涉及一种脂质运载蛋白突变蛋白,例如,当通过基本上如实施例4所述的表面等离子体共振(spr)分析测量时,所述脂质运载蛋白突变蛋白能够以通过约300nm、100nm、75nm、50nm、25nm、10nm或甚至更低(例如2nm)的kd量度的亲和力结合cd137。
在另一个实施方案中,例如,当通过基本上如实施例6所述的facs分析测量时,所述脂质运载蛋白突变蛋白能够以约250nm或更低、约100nm或更低、约50nm或更低、约18nm或更低的ec50值结合cd137。
本发明的另一个实施方案提供一种脂质运载蛋白突变蛋白,其能够通过结合cd137来活化cd137的下游信号传导通路。
在一些实施方案中,例如,当在基本上如实施例7所述的功能t细胞活化分析中测量时,与阴性对照seqidno:4相比,本发明的脂质运载蛋白突变蛋白能够诱导更高的il-2浓度。
在一些其它实施方案中,例如,当在基本上如实施例8所述的功能t细胞活化分析中测量时,与阴性对照seqidno:4相比,本发明的脂质运载蛋白突变蛋白不导致更高的il-2浓度。
在一些实施方案中,例如,当在基本上如实施例9所述的功能t细胞活化分析中测量时,与阴性对照seqidno:4相比,本发明的脂质运载蛋白突变蛋白能够诱导更高的il-2和ifn-γ增殖。
一方面,本发明提供cd137-结合的人泪液脂质运载蛋白突变蛋白。
由此,本发明提供一种或多种tlc突变蛋白,其能够以通过约300nm或更低和甚至约100nm或更低的kd量度的亲和力结合cd137。
在一些实施方案中,这种tlc突变蛋白在对应于成熟人泪液脂质运载蛋白(seqidno:1)的线性多肽序列的位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153的一个或多个位置处包含突变的氨基酸残基。
在一些特别的实施方案中,这种tlc突变蛋白可在对应于成熟人泪液脂质运载蛋白的线性多肽序列的位置26-34、55-58、60-61、65、104-106和108的一个或多个位置处包含突变的氨基酸残基。
在进一步特别的实施方案中,这种tlc突变蛋白可进一步在对应于成熟人泪液脂质运载蛋白的线性多肽序列的位置101、111、114和153的一个或多个位置处包含突变的氨基酸残基。
在其它特别的实施方案中,所述tlc可在对应于成熟人泪液脂质运载蛋白的线性多肽序列的位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153的一个或多个位置处包含突变的氨基酸残基。
在一些进一步的实施方案中,所述tlc突变蛋白可在对应于成熟人泪液脂质运载蛋白的线性多肽序列的序列位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153的一个或多个序列位置处包含至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26或甚至更多个突变的氨基酸残基,并且其中所述多肽结合cd137,特别是结合人cd137。
在一些又进一步的实施方案中,本发明涉及一种多肽,其中所述多肽为tlc突变蛋白,与成熟人泪液脂质运载蛋白的线性多肽序列相比,所述tlc突变蛋白在序列位置526-34、55-58、60-61、65、104-106和108处包含至少1、2、3、4、5、6、7、8、9、10、11、12或甚至更多个突变的氨基酸残基,并且其中所述多肽结合cd137,特别是结合人cd137。
在一些实施方案中,根据本发明的脂质运载蛋白突变蛋白可包含至少一个用如丝氨酸残基置换天然半胱氨酸残基的氨基酸置换。在一些实施方案中,根据本发明的tlc突变蛋包含用另一个氨基酸如丝氨酸残基置换在位置61和/或153处的天然半胱氨酸残基的氨基酸置换。在本上下文中应注意已经发现(在相应的天然(naive)核酸文库的水平上)移除由半胱氨酸61和153形成的野生型泪液脂质运载蛋白的结构二硫键(参见breustedt等人,2005,上文)可以提供泪液脂质运载蛋白突变蛋白,其不仅稳定折叠,还能以高亲和力结合给定非-天然配体。在一些特别的实施方案中,根据本发明的tlc突变蛋白包含氨基酸置换cys61→ala、phe、lys、arg、thr、asn、gly、gln、asp、ash、leu、tyr、met、ser、pro或trp和cys153→ser或ala。已证明这种置换可用于阻止形成天然存在的连接cys61和cys153的二硫桥,并从而有助于处理所述突变蛋白。然而,结合cd137并具有在cys61和cys153之间形成的二硫桥的泪液脂质运载蛋白突变蛋白也是本发明的一部分。
在一些实施方案中,结构性二硫键的消除可提供进一步的优点,即允许将非天然人工二硫键(自发)生成或有意引入到本发明的突变蛋白中,由此增加该突变蛋白的稳定性。例如,在一些实施方案中,在位置61、101和153处的两个或全部三个半胱氨酸密码子被另一个氨基酸密码子替换。此外,在一些实施方案中,根据本发明的tlc突变蛋白包含位置101处的天然半胱氨酸残基被丝氨酸残基或组氨酸残基替换的氨基酸置换。
在一些实施方案中,根据本发明的突变蛋白包含天然氨基酸被关于成熟人泪液脂质运载蛋白的氨基酸序列的位置28或105处的半胱氨酸残基替换的氨基酸置换。
此外,在一些实施方案中,根据本发明的突变蛋白包含位置111处的天然精氨酸残基被脯氨酸残基替换的氨基酸置换。另外,在一些实施方案中,根据本发明的突变蛋白包含位置114处的天然赖氨酸残基被色氨酸残基或谷氨酸替换的氨基酸置换。
在一些实施方案中,根据本发明的cd137结合tlc突变蛋白在对应于成熟人泪液脂质运载蛋白(seqidno:1)的线性多肽序列的位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153中的一个或多个位置处包含以下突变的氨基酸残基中的一个或多个:ala5→val或thr;arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;thr42→ser;gly46→asp;lys52→glu;leu56→ala;ser58→asp;arg60→pro;cys61→ala;lys65→arg或asn;thr71→ala;val85→asp;lys94→arg或glu;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;lys121→glu;ala133→thr;arg148→ser;ser150→ile和cys153→ser。在一些实施方案中,根据本发明的tlc突变蛋白在成熟人泪液脂质运载蛋白的这些序列位置处包含两个或更多个,比如3、4、5、6、7、8、9、10、11、12,甚至更多个,比如13、14、15、16、17、18、19、20、21、22、23、24、25、26或全部突变的氨基酸残基。
在一些另外的实施方案中,与成熟人泪液脂质运载蛋白的线性多肽序列相比,结合cd137的tlc突变蛋白包含以下氨基酸置换组之一:
1.arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;leu56→ala;ser58→asp;arg60→pro;cys61→ala;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;cys153→ser;
2.ala5→thr;arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;leu56→ala;ser58→asp;arg60→pro;cys61→ala;lys65→arg;val85→asp;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;lys121→glu;ala133→thr;cys153→ser;157→pro;
3.arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;leu56→ala;ser58→asp;arg60→pro;cys61→ala;lys65→asn;lys94→arg;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;lys121→glu;ala133→thr;cys153→ser;
4.ala5→val;arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;leu56→ala;ser58→asp;arg60→pro;cys61→ala;lys65→arg;lys94→glu;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;lys121→glu;ala133→thr;cys153→ser;157→pro;
5.arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;thr42→ser;leu56→ala;ser58→asp;arg60→pro;cys61→ala;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;ser150→ile;cys153→ser;157→pro;
6.arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;lys52→glu;leu56→ala;ser58→asp;arg60→pro;cys61→ala;thr71→ala;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;ala133→thr;arg148→ser;ser150→ile;cys153→ser;157→pro;或
7.ala5→thr;arg26→glu;glu27→gly;phe28→cys;pro29→arg;glu30→pro;met31→trp;leu33→ile;glu34→phe;gly46→asp;leu56→ala;ser58→asp;arg60→pro;cys61→ala;thr71→ala;cys101→ser;glu104→val;leu105→cys;his106→asp;lys108→ser;arg111→pro;lys114→trp;ser150→ile;cys153→ser;157→pro。
在剩余区域,即与序列位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153不同的区域,在突变的氨基酸序列位置之外,本发明的tlc突变蛋白可包含野生型(天然)氨基酸序列。
在又进一步的实施方案中,根据本发明的tlc突变蛋白与成熟人泪液脂质运载蛋白(seqidno:1)的序列具有至少70%的序列同一性或至少70%的序列同源性。作为示例性实例,seqidno:7的突变蛋白与成熟人泪液脂质运载蛋白的氨基酸序列具有近81%的氨基酸序列同一性或序列同源性。
在进一步特别的实施方案中,本发明的tlc突变蛋白包含如seqidnos:5-11中任一个所示的氨基酸序列或其片段或变体。
在进一步特别的实施方案中,本发明的tlc突变蛋白与选自seqidnos:5-11的氨基酸序列具有至少75%、至少80%、至少85%或更高的序列同一性。
本发明还包含具有选自seqidnos:5-11的氨基酸序列的tlc突变蛋白的结构同源物,所述结构同源物与所述的tlc突变蛋白具有大于约60%,优选地大于65%、大于70%、大于75%、大于80%、大于85%、大于90%、大于92%和最优选地大于95%的序列同源性或序列同一性。
可以通过诱变天然存在形式的人泪液脂质运载蛋白获得本发明的tlc突变蛋白。在一些所述诱变的实施方案中,置换(或替换)是保守置换。然而,设想任何置换(包括非保守置换或来自下文的示例性置换中的一个或多个),只要所述脂质运载蛋白突变蛋白保留了其结合cd137的能力,和/或其与之后被置换的序列具有序列同一性,即所述脂质运载蛋白突变蛋白与成熟人泪液脂质运载蛋白(swiss-prot数据库登录号p31025)的氨基酸序列具有至少60%,比如至少65%、至少70%、至少75%、至少80%、至少85%或更高的序列同一性。
在一些另外的实施方案中,例如,当在基本上如实施例5所述的表面等离子体共振(spr)分析中测量时,本发明的hngal突变蛋白能够干扰cd137l与cd137的结合。
在一些特别的实施方案中,本发明提供以通过约200nm或更低的kd量度的亲和力结合cd137的脂质运载蛋白突变蛋白,其中所述脂质运载蛋白突变蛋白与seqidno:5的氨基酸序列具有至少90%或更高,比如95%的同一性。
另一方面,本发明涉及新的针对cd137或cd137特异性的特异性-结合的人脂质运载蛋白2(人lcn2或hngal)。
对此,本发明提供一种或多种hngal突变蛋白,其能够以通过约200nm或更低、约140nm或更低、约50nm或更低和甚至约10nm或更低的kd量度的亲和力结合cd137。更优选地,所述hngal突变蛋白可具有通过约5nm更低的kd量度的亲和力。
在一些实施方案中,本发明的hngal突变蛋白在对应于成熟hngal(seqidno:2)的线性多肽序列的位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、83、87、94、96、100、103、106、125、127、132和134的一个或多个位置处包含置换。
在特别的实施方案中,本发明的脂质运载蛋白突变蛋白在对应于成熟hngal(swiss-prot数据库登录号p80188;seqidno:2)的线性多肽序列的序列位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、83、87、94、96、100、103、106、125、127、132和134的序列位置处包含至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21或甚至更多个置换。优选地,设想本发明涉及一种脂质运载蛋白突变蛋白,其除了在对应于成熟人ngal的线性多肽序列的位置36、87和/或96的位置处包含一个或多个置换外,还在对应于成熟hngal的线性多肽序列的位置28、40-41、49、52、65、68、70、72-73、77、79、81、83、94、100、103、106、125、127、132和134的一个或多个位置处包含置换。
在一些又进一步的实施方案中,本发明涉及一种多肽,其中所述多肽是hngal突变蛋白,与成熟hngal(swiss-prot数据库登录号p80188;seqidno:2)的线性多肽序列相比,所述hngal突变蛋白在序列位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、87、96、100、103、106、125、127、132和134处包含至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21或甚至更多个突变的氨基酸残基,并且其中所述多肽结合cd137,特别是结合人cd137。
在一些实施方案中,本发明的cd137-结合hngal突变蛋白在成熟hngal(seqidno:2)的线性多肽序列的序列位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、83、87、94、96、100、103、106、125、127、132和134的任一个或多个处包含以下突变的氨基酸残基中的一个或多个:gln28→his;leu36→gln;ala40→ile;ile41→arg或lys;gln49→val、ile、his、ser或asn;tyr52→met;asn65→asp;ser68→met、ala或gly;leu70→ala、lys、ser或thr;arg72→asp;lys73→asp;asp77→met、arg、thr或asn;trp79→ala或asp;arg81→met、trp或ser;phe83→leu;cys87→ser;leu94→phe;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu和lys134→tyr。
在一些实施方案中,本发明的hngal突变蛋白在成熟hngal的这些序列位置处包含两个或更多个,比如3、4、5、6、7、8、9、10、11、12,甚至更多个,比如13、14、15、16、17、18、19、20、21、22、23、24或全部突变的氨基酸残基。
在一些另外的实施方案中,与成熟hngal的线性多肽序列相比,本发明的结合cd137的hngal突变蛋白包含下列氨基酸替换:
(a)gln28→his;leu36→gln;ala40→ile;ile41→lys;gln49→asn;tyr52→met;ser68→gly;leu70→thr;arg72→asp;lys73→asp;asp77→thr;trp79→ala;arg81→ser;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(b)gln28→his;leu36→gln;ala40→ile;ile41→arg;gln49→ile;tyr52→met;asn65→asp;ser68→met;leu70→lys;arg72→asp;lys73→asp;asp77→met;yrp79→asp;arg81→trp;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(c)gln28→his;leu36→gln;ala40→ile;ile41→arg;gln49→asn;tyr52→met;asn65→asp;ser68→ala;leu70→ala;arg72→asp;lys73→asp;asp77→thr;trp79→asp;arg81→trp;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(d)gln28→his;leu36→gln;ala40→ile;ile41→lys;gln49→asn;tyr52→met;asn65→asp;ser68→ala;leu70→ala;arg72→asp;lys73→asp;asp77→thr;trp79→asp;arg81→trp;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(e)gln28→his;leu36→gln;ala40→ile;ile41→lys;gln49→ser;tyr52→met;asn65→asp;ser68→gly;leu70→ser;arg72→asp;lys73→asp;asp77→thr;trp79→ala;arg81→met;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(f)gln28→his;leu36→gln;ala40→ile;ile41→lys;gln49→val;tyr52→met;asn65→asp;ser68→gly;leu70→thr;arg72→asp;lys73→asp;asp77→arg;trp79→asp;arg81→ser;cys87→ser;leu94→phe;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(g)gln28→his;leu36→gln;ala40→ile;ile41→arg;gln49→his;tyr52→met;asn65→asp;ser68→gly;leu70→thr;arg72→asp;lys73→asp;asp77→thr;trp79→ala;arg81→ser;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;
(h)gln28→his;leu36→gln;ala40→ile;ile41→lys;gln49→asn;tyr52→met;asn65→asp;ser68→gly;leu70→thr;arg72→asp;lys73→asp;asp77→thr;trp79→ala;arg81→ser;phe83→leu;cys87→ser;leu94→phe;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr;或
(i)g1n28→his;leu36→gln;ala40→ile;ile41→arg;gln49→ser;tyr52→met;asn65→asp;ser68→ala;leu70→thr;arg72→asp;lys73→asp;asp77→asn;trp79→ala;arg81→ser;cys87→ser;asn96→lys;tyr100→phe;leu103→his;tyr106→ser;lys125→phe;ser127→phe;tyr132→glu;lys134→tyr。
在剩余区域,即不同于序列位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、83、87、94、96、100、103、106、125、127、132和134的区域中,在突变的氨基酸序列位置之外,本发明的hngal突变蛋白可以包含野生型(天然的)氨基酸序列。
在另一个实施方案中,所述hngal突变蛋白与成熟人脂质运载蛋白2(swiss-prot数据库登录号p80188)的氨基酸序列具有至少70%或甚至更高的序列同一性。作为示例性实例,seqidno:17的突变蛋白与成熟hngal的氨基酸序列具有近86.5%的氨基酸序列同一性或序列同源性。
在进一步特别的实施方案中,根据本发明的脂质运载蛋白突变蛋白包含选自seqidnos:12-20的氨基酸序列或其片段或变体。
本发明的cd137-结合hngal突变蛋白的氨基酸序列可与选自seqidnos:12-20的序列具有高序列同一性,如至少70%、至少75%、至少80%、至少82%、至少85%、至少87%、至少90%的同一性,包括至少95%的同一性。
本发明还包含具有选自seqidnos:12-20的氨基酸序列的hngal突变蛋白的结构同源物,所述结构同源物与所述hngal突变蛋白具有大于约60%,优选地大于65%、大于70%、大于75%、大于80%、大于85%、大于90%、大于92%和最优选地大于95%的氨基酸序列同源性或序列同一性。
可以通过诱变天然存在形式的人脂质运载蛋白2获得本发明的hngal突变蛋白。在一些所述诱变的实施方案中,置换(或替换)是保守置换。然而,设想任何置换(包括非保守置换或来自下文的示例性置换中的一个或多个),只要所述脂质运载蛋白突变蛋白保留了其结合cd137的能力,和/或其与之后被置换的序列具有同一性,即所述脂质运载蛋白突变蛋白与成熟人脂质运载蛋白2(swiss-prot数据库登录号p80188)的氨基酸序列具有至少60%,比如至少65%、至少70%、至少75%、至少80%、至少85%或更高的同一性。
在一些另外的实施方案中,例如,当在基本上如实施例5所述的表面等离子体共振(spr)分析中测量时,本发明的tlc突变蛋白不干扰cd137l与cd137的结合。
在一些特别的实施方案中,本发明提供以通过约5nm或更低的kd量度的亲和力结合cd137的脂质运载蛋白突变蛋白,其中所述脂质运载蛋白突变蛋白与seqidno:13的氨基酸序列具有至少90%或更高,比如95%的同一性。
2.cd137特异性的脂质运载蛋白突变蛋白的应用
cd137是t细胞受体(tcr)活化上诱导的t细胞共刺激受体(nam等人,curr.cancerdrugtargets,5:357-363(2005);watts等人,anru.rev.immunol.,23:23-68(2005))。cd137除在活化的cd4+和cd8+t细胞上表达外,还在cd4+cd25+调节t细胞、天然杀伤(nk)和nk-t细胞、单核细胞、嗜中性粒细胞和树突状细胞上表达。已经在包括b细胞、单核细胞/巨噬细胞和树突状细胞的抗原呈递细胞上描述了cd137的天然配体cd137l(watts等人,annu.rev.immunol.,23:23-68(2005))。通过与其配体的相互作用,cd137导致了增加的tcr-诱导的t细胞增殖、细胞因子产生、功能成熟和延长的cd8+t细胞存活(nam等人,curr.cancerdrugtargets,5:357-363(2005);watts等人,annu.rev.immunol.,23:23-68(2005))。
cd137/cd137l相互作用涉及免疫响应的各个方面。这在抑制t细胞中活化诱导的细胞死亡中似乎是重要的(hurtado等人,j.immunol.158:2600,1997),但是消除了嗜中性粒细胞中其它细胞因子的抗凋亡作用(heinisch等人,eur.j.immunol.30:3441,2001)。因此,cd137可在免疫功能内环境稳态中起作用(ebata等人,eur.j.immunol.31:1210,2001),并可代表癌症或炎症反应的治疗中可被靶向的靶标共刺激系统(blazar等人,j.immunol.166:174,2001;takahashi等人,immunol.lett.76:183,2001;kim和broxmeyer,j.hematother.stemcellres.10:441,2001;kim等人,cancerres.61:2031,2001)。
因此,本发明的cd137-结合脂质运载蛋白突变蛋白的许多可能应用存在于医药中。另一方面,本发明涉及本文所公开的cd137-结合脂质运载蛋白突变蛋白在用于检测样品中cd137以及相应的诊断方法中的用途。
本发明还涉及如所述的一种或多种cd137-结合脂质运载蛋白突变蛋白在用于与cd137形成复合物中的用途。
因此,本发明的又一方面,所公开的脂质运载蛋白突变蛋白用于检测cd137。这种用途可包括以下步骤:使一种或多种所述突变蛋白与怀疑含有cd137的样品在合适条件下接触,由此允许在所述突变蛋白与cd137之间形成复合物,并通过合适信号检测所述复合物。
可检测信号可以通过如上文所述的标记产生,或通过由于结合(即复合物形成)本身而导致的物理特性的变化而产生。一个实例是表面等离子体共振,其数值在结合配偶体结合期间发生改变,所述配偶体中的一者固定在表面上,例如金箔上。
本文公开的cd137-结合脂质运载蛋白突变蛋白还可用于分离cd137。这种用途可包括以下步骤:使一种或多种所述突变蛋白与猜测含有cd137的样品在合适条件下接触,由此允许在所述突变蛋白与cd137之间形成复合物,并从所述样品中分离所述复合物。
在所公开的突变蛋白用于检测cd137以及分离cd137的用途中,可以将所述突变蛋白和/或cd137或者其结构域或片段固定在合适的固相上。
又一方面,本发明的特征在于包含根据本发明的cd137-结合脂质运载蛋白突变蛋白的诊断或分析试剂盒。
除了其在诊断中的用途,再一方面,本发明考虑包含本发明的突变蛋白和药学上可接受的赋形剂的药物组合物。
此外,本发明提供用作抗癌剂和/或免疫调节剂的结合cd137的人脂质运载蛋白突变蛋白。因此,设想将结合cd137的本发明的脂质运载蛋白突变蛋白用在治疗或预防诸如癌症、感染性疾病和自身免疫性疾病的人疾病的方法中。因此,还提供了治疗或预防有此需要的受试者中诸如癌症、感染性疾病和自身免疫性疾病的人疾病的方法,所述方法包括向所述受试者施用治疗有效量的结合cd137的本发明的脂质运载蛋白突变蛋白。
在t细胞中,cd137介导的信号传导导致了traf家族成员的募集和若干激酶包括ask-1、mkk、mapk3/mapk4、p38和jnk/sapk的活化。激酶活化之后是几种转录因子包括atf-2、jun和nf-κb的活化和核转运。除增加次优的tcr-诱导的增殖外,cd137介导的信号传导还保护t细胞,特别是保护cd8+t细胞免于活化诱导的细胞死亡(aicd)。
本发明涵盖本发明的cd137-结合脂质运载蛋白突变蛋白或包含这种脂质运载蛋白突变蛋白的组合物在结合cd137、共刺激t细胞和/或通过结合cd137来活化cd137的下游信号传导通路,包括增强il-2分泌和产生干扰素ifn-γ中的用途。
本发明的特征还在于一种结合cd137或共刺激t细胞的方法,所述方法包括应用一种或多种本发明的cd137-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
另外,本发明涉及一种活化cd137下游信号传导通路包括增强il-2分泌和产生干扰素ifn-γ的方法,所述方法包括应用一种或多种本发明的cd137-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
本发明还考虑一种诱导t淋巴细胞增殖的方法,所述方法包括应用一种或多种本发明的cd137-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
此外,cd137/cd137l相互作用的缺乏阻止了某些自身免疫性疾病的发展(seo等人,2003,2004)。除了良好表征的抗原呈递细胞-t细胞相互作用外,cd137/cd137l相互作用参与造血细胞和非造血细胞的网络。通过cd137l的信号传导在骨髓细胞及其细胞活性的分化中起关键作用,这表明cd137l信号触发并维持炎症。[immunenetwork2009;9(3):84-89]。
本发明涵盖能够与cd137l竞争地结合cd137的本发明的cd137-结合脂质运载蛋白突变蛋白或包含这种脂质运载蛋白突变蛋白的组合物在干扰cd137l与cd137的结合和/或天然cd137l信号传导中的用途。
本发明的特征还在于一种干扰cd137l与cd137的结合的方法,所述方法包括应用一种或多种本发明的cd137-竞争-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
此外,本发明涉及一种干扰cd137l天然信号传导的方法,所述方法包括应用一种或多种本发明的cd137-竞争-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
本发明还考虑一种降低促炎细胞因子和趋化因子产生的方法,所述方法包括应用一种或多种本发明的cd137-竞争-结合脂质运载蛋白突变蛋白或者一种或多种包含这种脂质运载蛋白突变蛋白的组合物。
b.本发明的脂质运载蛋白突变蛋白
脂质运载蛋白是天然进化从而结合配体的蛋白质性结合分子。脂质运载蛋白存在于许多生物体中,包括脊椎动物、昆虫、植物和细菌。脂质运载蛋白家族的成员(pervaiz,s.,&brew,k.(1987)fasebj.1,209-214)通常是小的分泌的蛋白并具有单个多肽链。它们的特征在于一系列不同的分子识别特性:它们结合多种分子(主要是疏水性分子,例如类视黄醇、脂肪酸、胆固醇、前列腺素、胆绿素、信息素、甜味剂(tastant)和着嗅剂(odorant))的能力;它们与特异细胞表面受体结合并形成大分子复合物。尽管过去将它们主要分类为转运蛋白,但现在明确了脂质运载蛋白完成多种生理功能。这些功能包括在视黄醇转运、嗅觉、信息素信号传导和前列腺素合成中的作用。脂质运载蛋白还涉及免疫响应的调节和细胞内环境稳态的介导(例如,flower,d.r.(1996)biochem.j.318,1-14和flower,d.r.等人(2000)biochim.biophys.acta1482,9-24中的综述)。
脂质运载蛋白之间的全序列保守性非常低,通常具有低于20%的序列同一性。形成强烈对比的是,其整体折叠模式高度保守。脂质运载蛋白结构的中心部分由单个八股反平行β折叠组成,所述β折叠自身闭合形成连续的氢键合β桶。该β桶形成中央腔体。桶的一端被穿过其底部的n末端肽段以及连接β折叠股的三个肽环立体封闭。β桶的另一端对溶剂开放并包含由四个柔性肽环形成的靶标结合位点。正是所述环在本应刚性的脂质运载蛋白骨架中的这种多样性产生多种不同的结合模式,每一种模式能够容纳不同大小、形状和化学特征的靶标(例如上文的flower,d.r.(1996);上文的flower,d.r.等人(2000),或skerra,a.(2000)biochim.biophys.acta1482,337-350中的综述)。
根据本发明的脂质运载蛋白突变蛋白可以是任何可选脂质运载蛋白的突变蛋白。可使用其突变蛋白的合适脂质运载蛋白(有时也命名为“蛋白质‘参考’骨架”或简称为“骨架”)的实例包括但不限于泪液脂质运载蛋白(脂质运载蛋白-1,vonebner腺蛋白)、视黄醇结合蛋白、嗜中性粒细胞、脂质运载蛋白型前列腺素d-合酶、β-乳球蛋白、后胆色素结合蛋白(bbp)、载脂蛋白d(apod)、嗜中性粒细胞明胶酶相关脂质运载蛋白(ngal)、泪液脂质运载蛋白(tlc)、α2-微球蛋白-相关蛋白(a2m)、24p3/uterocalin(24p3)、vonebners腺蛋白1(vegp1)、yonebners腺蛋白2(vegp2)和主要变应原canf1前体(all-1)。在相关的实施方案中,该脂质运载蛋白突变蛋白选自人嗜中性粒细胞明胶酶相关脂质运载蛋白(ngal)、人泪液脂质运载蛋白(tlc)、人载脂蛋白d(apod)和大菜粉蝶(pierisbrassicae)后胆色素结合蛋白。
当在本文的结合cd137的本发明的脂质运载蛋白突变蛋白的上下文中使用时,术语“特异的(specificfor)”包括该脂质运载蛋白突变蛋白分别针对cd137、结合cd137或者与cd137反应。因此,针对、结合或与其反应包括所述脂质运载蛋白突变蛋白分别特异性结合cd137。如本文所述,在上下文中术语“特异性地”是指,脂质运载蛋白突变蛋白与cd137蛋白反应,但至少基本上不与另一种蛋白反应。术语“另一种蛋白”分别包括任何非-cd137蛋白,包括本文公开的脂质运载蛋白指向的与cd137密切相关或同源的蛋白。然而,来自除人以外的物种(如在定义“受试者”的上下文中描述的那些)的cd137蛋白、片段和/或变体未被术语“另一种蛋白”排除。术语“基本上不结合”指本发明的脂质运载蛋白突变蛋白不结合另一种蛋白,即显示出小于30%、优选地20%、更优选地10%、特别优选地小于9%、8%、7%、6%或5%的交叉反应。如上定义的所述脂质运载蛋白是否特异性反应可以很容易地进行测试,特别是通过比较本发明的脂质运载蛋白突变蛋白与cd137的反应和所述脂质运载蛋白与其它(另一种)蛋白的反应。“特异性结合”也可以例如根据蛋白质印迹、elisa-、ria-、ecl-、irma测试、facs、ihc和肽扫描来测定。
当比较与另一种脂质运载蛋白(也见上文)的序列同一性时,根据本发明的脂质运载蛋白突变蛋白的氨基酸序列与相应脂质运载蛋白具有高序列同一性。在该一般情况下,根据本发明的组合的脂质运载蛋白突变蛋白的氨基酸序列与相应脂质运载蛋白(野生型或参考脂质运载蛋白)的氨基酸序列至少基本上相似。与相应脂质运载蛋白的序列至少基本上相似的根据本发明的组合的脂质运载蛋白突变蛋白的相应序列,在一定程度上与野生型(或参考)脂质运载蛋白类似,具有一个或多个氨基酸实施方案,与相应脂质运载蛋白的序列具有至少65%、至少70%、至少75%、至少80%、至少82%、至少85%、至少87%、至少90%的同一性,包括至少95%的同一性。在这一方面,本发明的脂质运载蛋白突变蛋白当然可包含本文所述的比较置换,其使该脂质运载蛋白突变蛋白能够分别结合cd137。脂质运载蛋白的典型突变蛋白相对于天然序列脂质运载蛋白在脂质运载蛋白配体结合位点开放端的四个环(参见上文)中包含一个或多个氨基酸突变。如上文所述,这些区域在决定脂质运载蛋白突变蛋白对所需靶标的结合特异性中至关重要。作为示例性实例,源自泪液脂质运载蛋白、ngal脂质运载蛋白或其同源物的多肽的突变蛋白在n-末端区域和/或β-桶结构端部处排列的三个肽环bc、de、和fg中的任何序列位置处可具有一个、两个、三个、四个或更多个突变的氨基酸残基,所述β-桶结构端部位于天然脂质运载蛋白结合口袋的对面。作为另一示例性实例,与泪液脂质运载蛋白的野生型序列相比,源自泪液脂质运载蛋白或其同源物的多肽的突变蛋白可在β-桶结构端部处排列的肽环de中没有突变的氨基酸残基。
与相应的天然脂质运载蛋白相比,根据本发明的脂质运载蛋白突变蛋白包含一个或多个(例如2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或甚至20个)置换,条件是这种脂质运载蛋白突变蛋白应该能够分别结合cd137。例如,脂质运载蛋白突变蛋白可在对应于具有例如泪液脂质运载蛋白、ngal脂质运载蛋白或本文公开的任何其它脂质运载蛋白的野生型序列的野生型脂质运载蛋白的不同位置的位置处(即在对应位置处)具有置换。在一些实施方案中,根据本发明的组合的脂质运载蛋白突变蛋白包含至少两个氨基酸置换,包括用精氨酸残基置换天然氨基酸的2、3、4、5或甚至更多个氨基酸置换。因此,以产生能够分别结合cd137的脂质运载蛋白突变蛋白为目标,对本文所述的蛋白质‘参考’骨架的核酸进行诱变。
并且,本发明的脂质运载蛋白突变蛋白可在其n-或c-末端,优选地c-末端包含异源氨基酸序列(例如seqidno:23,如strepii标签)而不影响该脂质运载蛋白突变蛋白的生物活性(分别结合其靶标,如cd137)。
同样,与相应的野生型脂质运载蛋白相比,本发明的脂质运载蛋白突变蛋白可在其n-末端缺失1、2、3、4或更多个氨基酸和/或在其c-末端缺失1、2或更多个氨基酸;例如seqidnos:5-11和16。
具体而言,为了确定与野生型脂质运载蛋白不同的脂质运载蛋白突变蛋白的氨基酸序列的氨基酸残基是否对应于野生型脂质运载蛋白的氨基酸序列中的某一位置,技术人员可利用本领域熟知的手段和方法,如手动地或通过使用计算机程序例如blast2.0(其代表基本局部比对搜索工具)或clustalw或任何适用于产生序列比对的其它合适程序进行比对。因此,野生型脂质运载蛋白可作为“受试序列”或“参考序列”,而本文所述的与野生型脂质运载蛋白不同的脂质运载蛋白的氨基酸序列作为“查询序列”。本文中术语“参考序列”和“野生型序列”可互换使用。
在一些实施方案中,置换(或替代)是保守置换。然而,可以设想任何置换(包括非-保守置换或者来自下文列出的示例性置换中的一个或多个),只要所述脂质运载蛋白突变蛋白保留了其分别结合cd137的能力,和/或其与之后被置换的序列具有同一性,即所述脂质运载蛋白突变蛋白与“原始”序列具有至少60%,比如至少65%、至少70%、至少75%、至少80%、至少85%或更高的同一性。
保守置换通常为以下置换,根据待突变的氨基酸列举,每一个待突变的氨基酸后面为一个或多个可作为保守性的替代:ala→gly、ser、val;arg→lys;asn→gln、his;asp→glu;cys→ser;gln→asn;glu→asp;gly→ala;his→arg、asn、gln;ile→leu、val;leu→ile、val;lys→arg、gln、glu;met→leu、tyr、ile;phe→met、leu、tyr;ser→thr;thr→ser;trp→tyr;tyr→trp、phe;val→ile、leu。其它置换也是可能的并可凭经验或根据其它已知的保守或非-保守置换确定。作为另一取向(orientation),以下八组的每一组包含氨基酸,该氨基酸通常可用于定义另一氨基酸的保守置换:
a.丙氨酸(ala)、甘氨酸(gly);
b.天冬氨酸(asp)、谷氨酸(glu);
c.天冬酰胺(asn)、谷氨酰胺(gln);
d.精氨酸(arg)、赖氨酸(lys);
e.异亮氨酸(ile)、亮氨酸(leu)、甲硫氨酸(met)、缬氨酸(val);
f.苯丙氨酸(phe)、酪氨酸(tyr)、色氨酸(trp);
g.丝氨酸(ser)、苏氨酸(thr);和
h.半胱氨酸(cys)、甲硫氨酸(met)。
如果这种置换导致了生物活性的改变,则可引入更多实质性改变(如以下或如下文关于氨基酸分类进一步描述)并就所需特征筛选产物。这种更实质性改变的实例是:ala→leu、ile;arg→gln;asn→asp、lys、arg、his;asp→asn;cys→ala;gln→glu;glu→gln;his→lys;ile→met、ala、phe;leu→ala、met、正亮氨酸;lys→asn;met→phe;phe→val、ile、ala;trp→phe;tyr→thr、ser;val→met、phe、ala。
脂质运载蛋白的生物性质中的实质性修饰通过选择它们在维持以下的效果上显著不同的置换完成:(a)置换区域中多肽骨架的结构,例如片(sheet)或螺旋构象;(b)靶点处分子的电荷或疏水性;或(c)侧链的体积(bulk)。基于一般侧链性质,将天然存在的残基分为以下的组:(1)疏水性:正亮氨酸、甲硫氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸;(2)中性亲水性:半胱氨酸、丝氨酸、苏氨酸;(3)酸性:天冬氨酸、谷氨酸;(4)碱性:天冬酰胺、谷氨酰胺、组氨酸、赖氨酸、精氨酸;(5)影响链取向的残基:甘氨酸、脯氨酸;和(6)芳香族:色氨酸、酪氨酸、苯丙氨酸。
非-保守置换需要将这些类别中一类的成员置换成另一类。通常还可用丝氨酸置换不参与维持相应脂质运载蛋白正确构象的任何半胱氨酸残基,以改善分子的氧化稳定性并防止异常交联。相反,可将半胱氨酸键添加至脂质运载蛋白以提高其稳定性。
采用已经建立的标准方法,可在核酸(如dna水平)上非常简单地完成任何突变(包括以上所讨论的插入)。氨基酸序列改变的示例性实例是插入或缺失以及氨基酸置换。这种置换可以是保守的,即氨基酸残基被替代为化学性质(特别是关于极性以及尺寸)相似的氨基酸残基。保守置换的实例为以下组的成员之间的替代:1)丙氨酸、丝氨酸和苏氨酸;2)天冬氨酸和谷氨酸;3)天冬酰胺和谷氨酰胺;4)精氨酸和赖氨酸;5)异亮氨酸、亮氨酸、甲硫氨酸和缬氨酸;和6)苯丙氨酸、酪氨酸和色氨酸。另一方面,也可以将非-保守性改变引入氨基酸序列。此外,代替替换单一氨基酸残基,还可以使泪液脂质运载蛋白的一级结构插入或缺失一个或多个连续氨基酸,只要这些缺失或插入导致稳定的折叠/功能的突变蛋白。
氨基酸序列的修饰包括单个氨基酸位置的定向诱变,以便通过引入某些限制性酶的切割位点来简化突变的脂质运载蛋白基因或其部分的亚克隆。此外,还可引入这些突变以进一步改善针对给定靶标(比如cd137)的脂质运载蛋白突变蛋白的亲和力。另外,如果需要,可以引入突变以便调节突变蛋白的某些特征,例如改善折叠稳定性、血清稳定性、蛋白质抗性或水溶解性或者降低聚集趋势。例如,可以将天然存在的半胱氨酸残基突变成其它氨基酸以防止二硫桥形成。还可以有意将其它氨基酸序列位置突变成半胱氨酸以便引入新的反应基团,例如用于与其它化合物如聚乙二醇(peg)、羟乙基淀粉(hes)、生物素、肽或蛋白质缀合,或用于形成非天然存在的二硫结合(linkage)。所产生的硫醇部分可以用于peg化或hes化突变蛋白,例如,以便增加相应脂质运载蛋白突变蛋白的血清半衰期。这种将半胱氨酸残基引入tlc突变蛋白的氨基酸序列的突变的示例性可能性包括置换thr40→cys、glu73→cys、arg90→cys、asp95→cys和glu131→cys。在任意氨基酸位置40、73、90、95和/或131侧面产生的硫醇部分可以用于peg化或hes化突变蛋白,例如,以便增加相应tlc突变蛋白的血清半衰期。
还可以将其它氨基酸序列位置突变成半胱氨酸以便引入新的反应基团,例如用于与其它化合物如聚乙二醇(peg)、羟乙基淀粉(hes)、生物素、肽或蛋白质缀合,或用于形成非天然存在的二硫结合(linkage)。
在一些实施方案中,若上述部分中的一个与本发明的脂质运载蛋白突变蛋白缀合,则与氨基酸侧链的缀合可能是有利的。合适的氨基酸侧链可以天然存在于人脂质运载蛋白的氨基酸序列中,或者可以通过诱变引入。若通过诱变引入了合适的结合位点,则一种可能性是用半胱氨酸残基替代在适当位置处的氨基酸。例如,这种突变包括人泪液脂质运载蛋白的野生型序列中thr40→cys、glu73→cys、arg90→cys、asp95→cys或glu131→cys置换中的至少一个。在这些位置中任意处新产生的半胱氨酸残基随后可用于将该突变蛋白与延长该突变蛋白血清半衰期的部分(例如peg或其活化衍生物)缀合。
对于人脂质运载蛋白2的突变蛋白,这种将半胱氨酸残基引入脂质运载蛋白(包括人脂质运载蛋白2突变蛋白)的氨基酸序列中的突变的示例性可能性包括在对应于人ngal野生型序列的序列位置14、21、60、84、88、116、141、145、143、146或158的序列位置中至少一个处引入半胱氨酸(cys)残基。在一些实施方案中,其中本发明的人脂质运载蛋白2突变蛋白具有这样一种序列,与swiss-prot/uniprot数据库登录号p80188的序列相比,在该序列中已经用另一氨基酸残基替代半胱氨酸,相应的半胱氨酸可重新引入该序列。作为示例性实例,在这种情况下,可以通过还原为在swiss-prot登录号p80188的序列中原始存在的半胱氨酸来在氨基酸位置87处引入半胱氨酸残基。在氨基酸位置14、21、60、84、88、116、141、145、143、146和/或158中任何的侧面处产生的硫醇部分可以用于peg化或hes化突变蛋白,例如,以便增加相应人脂质运载蛋白2突变蛋白的血清半衰期。
在另一个实施方案中,为了给将上述化合物中的一个缀合至根据本发明的脂质运载蛋白突变蛋白提供合适的氨基酸侧链,可以通过诱变引入人工氨基酸。一般而言,这些人工氨基酸设计为更具反应性,从而促进与所需化合物缀合。这种可通过人工trna引入的人工氨基酸的一个实例是对乙酰基-苯丙氨酸。
对于本文公开的突变蛋白的若干应用,以融合蛋白形式使用它们可能是有利的。在一些实施方案中,本发明的脂质运载蛋白突变蛋白在其n-末端或其c-末端融合至蛋白质、蛋白质结构域或肽,例如单个序列和/或亲和标签。
合适的融合配偶体的进一步实例是例如strep-标签
一般而言,可以用任何合适的化学物质或酶标记本发明的脂质运载蛋白突变蛋白,所述化学物质或酶直接或间接产生可检测化合物或化学、物理、光学或酶反应信号。物理反应和同时光学反应/标记物的实例是照射时的荧光发射或当使用放射活性标记时的x-射线发射。碱性磷酸酶、辣根过氧化物酶和β-半乳糖苷酶是催化显色反应产物形成的酶标记(同时是光学标记)的实例。一般而言,通常用于抗体(除了那些专门与免疫球蛋白的fc部分中的糖部分一起使用的)的所有标记也可用于与本发明的脂质运载蛋白突变蛋白缀合。本发明的脂质运载蛋白突变蛋白还可与任何合适的治疗活性剂缀合,例如用于将这种试剂靶向递送至给定细胞、组织或器官或用于选择性靶向如肿瘤细胞的细胞而不影响周围正常细胞。这种治疗活性剂的实例包括放射性核素、毒素、小有机分子和治疗性肽(例如作为细胞表面受体的激动剂/拮抗剂的肽或竞争给定细胞靶标上蛋白质结合位点的肽)。然而,本发明的脂质运载蛋白突变蛋白还可与治疗活性核酸(例如反义核酸分子、小干扰rna、微rna或核酶)缀合。可以采用本领域熟知的方法产生这种缀合物。
如上所述,在一些实施方案中,本发明的脂质运载蛋白突变蛋白可与延长该突变蛋白血清半衰期的部分缀合(这方面还参见pct公开文本wo2006/56464,其中参考具有ctla-4结合亲和力的人嗜中性粒细胞明胶酶相关脂质运载蛋白的突变蛋白描述了这种缀合策略)。所述延长血清半衰期的部分可以是聚亚烷基二醇分子、羟乙基淀粉、例如棕榈酸(vajo&duckworth2000,pharmacol.rev.52,1-9)的脂肪酸、免疫球蛋白的fc部分、免疫球蛋白的ch3结构域、免疫球蛋白的ch4结构域、白蛋白结合肽或白蛋白结合蛋白、转铁蛋白,仅举几个例子。白蛋白结合蛋白可以是细菌白蛋白结合蛋白、抗体、包括结构域抗体(例如,参见美国专利6,696,245)的抗体片段或具有白蛋白结合活性的脂质运载蛋白突变蛋白。因此,用于延长本发明的脂质运载蛋白突变蛋白的半衰期的合适缀合配偶体包括白蛋白结合蛋白,例如,细菌白蛋白结合结构域,如链球菌蛋白g
在其它实施方案中,白蛋白本身(osborn,b.l.等人,2002,j.pharmacol.exp.ther.303,540-548)或白蛋白的生物活性片段可用作本发明脂质运载蛋白突变蛋白的缀合配偶体。术语“白蛋白”包括所有的哺乳动物白蛋白,例如人血清白蛋白或牛血清白蛋白或大鼠白蛋白。可以按照us专利5,728,553或欧洲专利申请ep0330451和ep0361991中的描述重组产生所述白蛋白或其片段。可将重组的人白蛋白(recombumin
若白蛋白结合蛋白是抗体片段,则其可以是结构域抗体。将结构域抗体(dab)工程化以允许精确控制生物物理性质和体内半衰期,从而产生最佳安全性和有效性产品概况。结构域抗体例如可从domantisltd.(cambridge,ukandma,usa)商购获得。
将转铁蛋白用作延长本发明的脂质运载蛋白突变蛋白血清半衰期的部分,可将该突变蛋白遗传地融合至非-糖基化转铁蛋白的n或c末端或者两者。非-糖基化转铁蛋白具有14-17天的半衰期,且转铁蛋白-融合蛋白将类似地具有延长的半衰期。所述转铁蛋白载体还提供高生物利用度、生物分布和循环稳定性。这一技术可从biorexis(biorexispharmaceuticalcorporation,pa,usa)商购获得。用于用作蛋白质稳定剂/半衰期延长配偶体的重组人转铁蛋白(deltaferrintm)也可从novozymesdeltaltd.(nottingham,uk)商购获得。
若将免疫球蛋白的fc部分用于延长本发明的脂质运载蛋白突变蛋白的血清半衰期的目的,则可采用可从syntonixpharmaceuticals,inc(ma,usa)商购获得的synfusiontm技术。这种fc-融合技术的应用允许产生更长效的生物药物,并可以例如由两个拷贝的连接至抗体fc区的所述突变蛋白组成以改善药物动力学、溶解度和生产效率。
另一个延长本发明的脂质运载蛋白突变蛋白半衰期的选择是将长的、非结构化的、柔性富含甘氨酸的序列(例如具有约20至80个连续甘氨酸残基的聚甘氨酸)与突变蛋白的n-或c-末端融合。wo2007/038619中公开的这种方法,例如也称为“rpeg”(重组peg)。
若将聚亚烷基二醇用作缀合配偶体,则该聚亚烷基二醇可以是取代的、未取代的、线性的或支链的。其也可以是活化的聚亚烷基衍生物。合适的化合物的实例是如wo99/64016、美国专利6,177,074或美国专利6,403,564中关于干扰素所述的聚乙二醇(peg)分子,或如对于其它蛋白质所述的聚乙二醇(peg)分子,例如peg修饰的天冬酰胺酶、peg-腺苷脱氨酶(peg-ada)或peg-超氧化物歧化酶(参见,例如,fuertges等人(1990)theclinicalefficacyofpoly(ethyleneglycol)-modifiedproteinsj.control.release11,139-148)。这种聚合物(例如聚乙二醇)的分子量可以从大约300到大约70.000道尔顿,包括,例如,具有约10.000、约20.000、约30.000或约40.000道尔顿的分子量的聚乙二醇。此外,如例如美国专利6,500,930或6,620,413所述,为了延长血清半衰期的目的,可将例如淀粉或羟乙基淀粉(hes)的碳水化合物低聚-和聚合物与本发明的突变蛋白缀合。
此外,本文公开的脂质运载蛋白突变蛋白可与可赋予本发明的脂质运载蛋白突变蛋白新特征(例如酶活性或对其它分子的结合亲和力)的部分融合。合适融合配偶体的实例是碱性磷酸酶、辣根过氧化物酶、谷胱甘肽-s-转移酶、蛋白g的白蛋白结合结构域、蛋白a、抗体片段、低聚结构域或毒素。
特别地,可以将本文公开的脂质运载蛋白突变蛋白与单独的酶活性位点融合,使得所得融合蛋白的两个“组分”一起作用于给定治疗靶标。可将脂质运载蛋白突变蛋白的结合结构域附着至致病靶标,从而允许酶结构域消除该靶标的生物功能。
本发明还涉及核酸分子(dna和rna),该核酸分子包含编码本发明的脂质运载蛋白突变蛋白的核苷酸序列。由于遗传密码子的简并性允许指定相同氨基酸的其它密码子置换某些密码子,因此本发明不局限于编码本文所述脂质运载蛋白突变蛋白的特定核酸分子,而是涵盖包含编码功能性突变蛋白的核苷酸序列的全部核酸分子。在这一方面,本发明提供核苷酸序列,该核苷酸序列编码如seqidnos:24-39所示的本发明的一些脂质运载蛋白突变蛋白。
在本发明的一个实施方案中,所述方法包括使核酸分子在核苷酸三联体处经受诱变,该核苷酸三联体编码对应于人ngal(seqidno:2)的线性多肽序列的序列位置28、36、40-41、49、52、65、68、70、72-73、77、79、81、83、87、94、96、100、103、106、125、127、132和134的序列位置中至少一个,或甚至更多个。
在根据本发明的方法的另一个实施方案中,首先使编码人泪液脂质运载蛋白的核酸分子在人泪液脂质运载蛋白(seqidno:1)的线性多肽序列的氨基酸序列位置5、26-31、33-34、42、46、52、56、58、60-61、65、71、85、94、101、104-106、108、111、114、121、133、148、150和153中一个或多个处经受诱变。第二,使该编码人泪液脂质运载蛋白的核酸分子还在成熟人泪液脂质运载蛋白的线性多肽序列的氨基酸序列位置101、111、114和153中一个或多个处经受诱变。
本发明还包含编码本发明脂质运载蛋白突变蛋白的核酸分子,其在实验诱变的指定序列位置之外包含另外的突变。这种突变通常是耐受的或甚至可证明是有利的,例如若这些突变有利于所述突变蛋白的改善的折叠效率、血清稳定性、热稳定性或配体结合亲和力。
本申请公开的核酸分子可以“可操作地连接”至一个或多个调节序列以允许该核酸分子的表达。
核酸分子,如dna,如果它包括包含关于转录和/或翻译调节的信息的序列元件,并且这样的序列“可操作地连接”至编码多肽的核苷酸序列的话,其被称为“能够表达核酸分子”或能够“允许核苷酸序列的表达”。可操作的连接是这样的连接:在所述连接中,调节序列元件和待表达的序列以使基因能够表达的方式相连接。基因表达需要的调控区域的确切性质可能在种类间变化,但一般来说这些区域包括启动子,在原核生物中,所述启动子既包含启动子本身(即指导转录启动的dna元件),又包括当转录成rna时发出翻译启动信号的dna元件。这种启动子区域通常包括参与转录和翻译启动的5′非编码序列,如-35/-10盒以及原核生物中的shine-dalgarno元件或者真核生物中的tata盒、caat序列和5′-加帽元件。这些区域还可以包括增强子或阻遏元件以及用于将天然多肽靶向宿主细胞特定隔室的被翻译信号和前导序列。
此外,3′非编码序列可以包含参与转录终止、聚腺苷酸化等的调节元件。然而,如果这些终止序列在特定的宿主细胞内没有令人满意的功能,那么它们可能被该细胞中有功能的信号置换。
因此,本发明的核酸分子可以包括调节序列,如启动子序列。在一些实施方案中,本发明的核酸分子包括启动子序列和转录终止序列。合适的原核启动子是,例如,tet启动子,lacuv5启动子或t7启动子。可用于在真核细胞中表达的启动子的实例有sv40启动子或cmv启动子。
本发明的核酸分子也可以是载体的一部分或任何其它类型的克隆载体,如质粒、噬菌粒、噬菌体、杆状病毒、粘粒或人工染色体。
在一个实施方案中,核酸分子包含在质粒中。质粒载体指编码融合至目的cdna的温和噬菌体(如m13或fl)基因间区域或其功能部分的载体。用这样的噬菌粒载体和合适的辅助噬菌体(如m13k07、vcs-m13或r408)超感染细菌宿主细胞后,产生完整的噬菌体颗粒,由此使得可以将编码的异源cdna物理偶联至在噬菌体表面上展示的其相应多肽(参见例如lowman,h.b.(1997)annu.rev.biophys.biomol.struct.26,401-424,或rodi,d.j.和makowski,l.(1999)curr.opin.biotechnol.10,87-93)。
除上文描述的调节序列和编码本文所述脂质运载蛋白突变蛋白的核酸序列以外,这种克隆载体可以包括源自与用于表达的宿主细胞相容的物种的复制和控制序列,以及赋予转化或转染的细胞可选择表型的选择标记物。大量合适的克隆载体是本领域已知的,并可商购获得。
可将编码本文所述脂质运载蛋白突变蛋白的dna分子,特别是含有这种突变蛋白的编码序列的克隆载体,转化到能够表达该基因的宿主细胞中。可以使用标准技术进行转化。因此,本发明还涉及含有本文公开的核酸分子的宿主细胞。
在适于表达编码本发明融合蛋白的核苷酸序列的条件下培养转化的宿主细胞。合适的宿主细胞可以是原核细胞,如大肠杆菌(e.coli)或枯草芽孢杆菌(bacillussubtilis),或者是真核细胞,例如酿酒酵母(saccharomycescerevisiae)、巴斯德毕赤酵母(pichiapastoris)、sf9或high5昆虫细胞、永生化哺乳动物细胞系(例如hela细胞或cho细胞)或原代哺乳动物细胞。
本发明还涉及用于产生本文所述的脂质运载蛋白突变蛋白的方法,其中通过遗传工程方法,从编码该突变蛋白的核酸开始产生该突变蛋白、该突变蛋白的片段或该突变蛋白与另一多肽(如,另一脂质运载蛋白突变蛋白)的融合蛋白。该方法可以在体内进行,脂质运载蛋白突变蛋白可以例如在细菌或真核宿主生物体中产生,然后从该宿主生物体或其培养物中分离。还可以在体外产生蛋白质,例如使用体外翻译系统。
在体内产生脂质运载蛋白突变蛋白时,通过重组dna技术(上文已概述)将编码该突变蛋白的核酸引入合适的细菌或真核宿主生物体中。为此,首先使用已建立的标准方法,用克隆载体转化该宿主细胞,所述克隆载体包含编码本文所述脂质运载蛋白突变蛋白的核酸分子。然后在允许表达该异源dna并因而允许合成相应多肽的条件下培养该宿主细胞。之后,从细胞或培养基中回收该多肽。
在一些实施方案中,本申请公开的核酸分子,如dna,可以“可操作地连接”至本发明的另一核酸分子以允许本发明融合蛋白的表达。在这一方面,可操作的连接是这样的连接:在所述连接中,第一核酸分子的序列元件和第二核酸分子的序列元件以使融合蛋白能够表达为单一多肽的方式相连接。
此外,在一些实施方案中,可在本发明的ngal突变蛋白中移除cys76和cys175之间天然存在的二硫键。在本发明的tlc突变蛋白的一些实施方案中,可以移除cys61和cys153之间天然存在的二硫键。因此,这种突变蛋白可以在具有还原性氧化还原环境的细胞隔室中产生,例如,在革兰氏阴性细菌的细胞质中。
在本发明脂质运载蛋白突变蛋白包括分子内二硫键的情况下,可以优选的是,使用适合的信号序列将新生多肽引导至具有氧化性氧化还原环境的细胞隔室。这种氧化性环境可以由革兰氏阴性细菌(例如大肠杆菌)的外周胞质、在革兰氏阳性细菌的细胞外环境中或在真核细胞的内质网腔中提供,并且这种氧化性环境通常有利于结构性二硫键的形成。
然而,还可以在宿主细胞(优选大肠杆菌)的胞液中产生本发明的突变蛋白。在这种情况下,该多肽可以以可溶和折叠状态直接获得,也可以以包涵体的形式回收,然后体外复性。其它选择为使用具有氧化性细胞内环境的特定宿主菌株,其可以因而允许在胞液中形成二硫键(venturi等人(2002)j.mol.biol.315,1-8.)。
然而,本文所述的脂质运载蛋白突变蛋白可以不一定为仅通过利用基因工程生成或产生。而是,还可以通过化学合成或通过体外转录和翻译获得这种突变蛋白,所述化学合成例如merrifield固相多肽合成。例如可行的是,使用分子建模鉴定有希望的突变,然后体外合成希望的(设计的)多肽,然后研究对于cd137的结合活性。用于固相和/或液相合成蛋白的方法是本领域熟知的(参见例如bruckdorfer等人(2004)curr.pharm.biotechnol.5,29-43)。
在另一个实施方案中,本发明的脂质运载蛋白突变蛋白可以使用本领域技术人员已知的成熟方法通过体外转录/翻译产生。
本领域技术人员将会理解用于制备本发明预期的、但其蛋白或核酸序列在本文中没有明确地披露的脂质运载蛋白突变蛋白的方法。作为一个概述,氨基酸序列的这种修饰包括,例如,通过引入某些限制性酶的切割位点对单个氨基酸位置进行定点诱变,以便简化突变的脂质运载蛋白基因或其部分的亚克隆。此外,也可以引入这些突变以进一步提高脂质运载蛋白突变蛋白对其靶标(如分别是cd137)的亲和力。并且,如果必要,可以引入突变来调节突变蛋白的某些特征,如提高折叠稳定性、血清稳定性、蛋白抗性或水溶解性,或者减少聚集趋势。例如,天然存在的半胱氨酸残基可突变为其它氨基酸以防止形成二硫桥。
本文公开的脂质运载蛋白突变蛋白以及其衍生物可以类似于抗体或其片段用在许多领域。例如,该脂质运载蛋白突变蛋白可以用于以酶、抗体、放射活性物质或任何其它具有生物化学活性或确定的结合特征的基团进行标记。这样,它们各自的靶标或其缀合物或融合蛋白可以被检测或者与它们接触。此外,本发明的脂质运载蛋白突变蛋白可以用于通过已建立的分析方法(如elisa或蛋白质印迹)或通过显微术或免疫传感器(immunosensorics)来检测化学结构。在这个方面,检测信号可以通过使用合适的突变蛋白缀合物或融合蛋白直接产生,或者通过经由抗体对结合的突变蛋白进行免疫化学检测而间接产生。
对本领域技术人员而言,经审查以下的实施例及其附图(其并非意图用于限制),本发明的另外的目的、优点和特征将变得显而易见。因此,应当理解的是,虽然本发明通过示例性实施方案以及任选的特征进行特定的公开,但本领域技术人员可以采取本文所公开于其中的公开内容的修饰及变化,且认为这样的修饰及变化在本发明的范围内。
实施例
实施例1:特异性结合cd137的突变蛋白的选择和优化
本申请公开的代表性cd137特异性脂质运载蛋白突变蛋白选自基于人ngal和人tlc的天然突变体文库。采用不同的策略和靶标来获得cd137结合突变蛋白。所使用的重组靶标是来自人的cd137全细胞外结构域的可商购获得的fc融合物(hucd137-fc,r&dsystems838-4b)和人cd137的单个亚结构域,其全部以与人fc片段的融合物产生。作为可选的非-fc融合的靶标,我们采用了带his-标签的人cd137细胞外结构域(invitrogen,10041-h08h-250)。可选地,使用采用人cd137全cdna转染的cho细胞的基于细胞的淘选(cell-basedpanning)。使用标准程序进行基于蛋白质和基于细胞的淘选。对选择后获得的克隆进行实施例2所述的筛选过程。
实施例2:采用高通量elisa筛选识别特异性结合cd137的突变蛋白
将融合至c-末端strep-标签(seqidno:23,参见实施例3)的单个脂质运载蛋白突变蛋白用于接种2xyt/amp培养基并生长过夜(14-18h)至稳定期。随后,接种50μl来自稳定期培养物的2xyt/amp,并在37℃下温育3h,然后转移至22℃直至达到0.6-0.8的od595。通过加入10μl补充有1.2μg/ml脱水四环素的2xyt/amp诱导突变蛋白的产生。将培养物在22℃下温育直到第二天。加入40μl在pbs/t中的5%(w/v)bsa和在25℃下温育1h之后,培养物准备用于筛选分析。
通过在微量滴定板上涂布相关物种的hucd137-fc(5μg/ml,在pbs中)在4℃下过夜来测试分离的突变蛋白与人cd137的结合。用含有2%bsa的pbst封闭该板,将20μlbsa-封闭的培养物添加至微量滴定板并在25℃下温育1h。用与辣根过氧化物酶缀合的抗-strep标签抗体检测结合的突变蛋白(1h温育;iba,goettingen)。为了定量,加入20μlquantablu荧光过氧化物酶底物,并在330nm的激发波长和420nm的发射波长下测定所得荧光。
为了选择耐高温性的突变蛋白,将bsa封闭的培养物在60℃下温育1h,冷却至室温后将其加入至如前一段所述的cd137涂布的和bsa封闭的微量滴定板中。随后将所述突变蛋白按照前一段所述处理,并选择用于细菌表达、纯化和进一步表征。
实施例3:突变蛋白的表达
在2yt-amp培养基中的大肠杆菌中表达独特的突变蛋白,所述突变蛋白具有c-末端标签sawshpqfek(seqidno:21)或psawshpqfek(seqidno:22)(包括sa或psa连接子和strep-tag
实施例4:通过表面等离子体共振(spr)测定的突变蛋白结合人cd137-fc融合蛋白的亲和力
采用表面等离子体共振(spr)测量本文公开的代表性脂质运载蛋白突变蛋白的结合动力学和亲和力。
在37℃下,在采用hbs-ep+(1x;br-1006-69;gehealthcare)作为流动缓冲液的biacoret200仪器(gehealthcare)上实施代表性突变蛋白与人cd137-fc融合蛋白(hucd137-fc)的结合的spr分析。
在蛋白质测量之前,进行三个再生循环用于调节目的。通过施加3mmgcl2达60s然后施加ph1.7的10mm甘氨酸达180s来实现衍生化芯片表面的再生。在随后步骤中抗-人igg-fc抗体用于固定hucd137-fc并取自人抗体捕获试剂盒(gehealthcare,br-1008-39)。使用标准胺偶联化学和包含在试剂盒中的固定缓冲液(10mm醋酸钠ph5.0)将抗-人igg-fc抗体固定在cm5传感器芯片上,导致约13000个共振单位(ru)的配体密度。参考通道被相应处理。
以在hbs-ep+缓冲液中10μl/min的流速在此表面上捕获浓度为0.5μg/ml的hucd137-fc达180s。未对参考通道施加靶标蛋白。随后,以30μl/min的流速,以在hbs-ep+缓冲液中适当稀释系列施加脂质运载蛋白突变蛋白。如上所述实现衍生化芯片表面的再生。用biacoret200评估软件(v2.0)评估数据。使用双重参考并将1∶1的结合模型用于拟合原始数据。
图1示出了针对具有图中提供的对应seqidno的测试的脂质运载蛋白突变蛋白测定的spr迹线及拟合曲线。描绘了与hucd137-fc结合的数据。存在对人靶标清晰的spr结合信号,而阴性对照seqidno:3和seqidno:4未显示结合。下表1提供了自该数据的拟合得到的亲和力。
表1:
实施例5:表面等离子体共振(spr)分析测定人cd137l与突变蛋白在结合人cd137-fc融合蛋白中的竞争
对于本申请所述的结合cd137的脂质运载蛋白突变蛋白,通常可以有两种结合方式:第一种情况,突变蛋白(与cd137)的结合位点和人cd137配体(cd137l)与cd137的结合位点重叠。当这种脂质运载蛋白突变蛋白结合cd137时,这干扰了cd137l与cd137的结合,并伴随地导致了干扰天然cd137l信号传导(“竞争性结合”);第二种情况,突变蛋白的结合位点与cd137l的结合不重叠,并且这种脂质运载蛋白突变蛋白可结合cd137而不干扰cd137l的结合和天然cd137l信号传导(“非-竞争性结合”)。
通过其配体的cd137的聚集(clustering)活化了cd137的下游信号传导通路。在t细胞的情况下,cd137活化导致了t细胞活化响应的共刺激,如促炎细胞因子的增殖和产生。
诱导cd137聚集的另一种方式是采用固定的cd137结合剂。当涂布在板上(如在塑料培养皿上并在板中温育t细胞)时,竞争性和非-竞争性cd137结合剂均实现cd137聚集,由此活化下游信号传导。
因此,一方面,当按照本文所述应用时,竞争性和非-竞争性cd137结合剂均能活化cd137的下游信号传导。
另一方面,可将竞争性cd137结合剂用于抑制天然cd137/cd137l的相互作用,并由此抑制由cd137阳性细胞与cd137l表达细胞(例如,抗原呈递细胞)相遇而诱导的天然信号传导。当需要抑制不适当的强炎症或自身免疫反应时,这种作用方式是可取的。
为了证明本申请提供了竞争型突变蛋白和非-竞争型突变蛋白两者,我们采用表面等离子体共振(spr)实验。我们采用spr实验来研究cd137l与本文公开的三种代表性脂质运载蛋白突变蛋白在结合人cd137-fc融合蛋白(hucd137-fc)中的竞争。在这一试验中,研究了脂质运载蛋白突变蛋白是否能够结合预先形成的cd137和cd137l的复合物;如果不能,则证明cd137上该脂质运载蛋白突变蛋白的结合表位与cd137上cd137l的结合表位重叠。因此,相对于cd137/cd137l的相互作用,相应的突变蛋白竞争性地结合cd137。如果cd137l和脂质运载蛋白突变蛋白能够同时结合,则相对于cd137/cd137l的相互作用,该结合是非-竞争性的。
在37℃下,在采用hbs-ep+(1x;br-1006-69;gehealthcare)作为流动缓冲液的biacoret200仪器上实施竞争试验。采用生物素capture试剂盒(gehealthcare)将生物素化的hucd137-fc固定在芯片表面。采用标准的nhs化学将cd137-fc蛋白生物素化。在具有预先固定的互补ss-dna寡核苷酸的传感器芯片cap上捕获未稀释的生物素capture试剂(与ss-dna寡核苷酸(oligo)缀合的链霉亲和素)。此后,以5μl/min的流速施加2μg/ml的生物素化的cd137-fc蛋白质达300s。通过以10μl/min的流速施加250mmnaoh中的6m胍-hcl达120s实现芯片表面的再生。
在两个第一次测量周期中,确定了在实验条件下cd137l与cd137的成功结合,并且获得了在没有配体的情况下用于测试的脂质运载蛋白突变蛋白的单独结合的参考水平。在第三次周期中,按照如下详细叙述的,在加入脂质运载蛋白突变蛋白之前,用cd137l饱和cd137。
以500nm的浓度和30μl/min的流速将人cd137配体-fc(r&dsystems2295-4l-025/cf)配体施加至固定的cd137-fc蛋白达30s。再生后,以5μm的浓度和30μl/min的流速施加脂质运载蛋白突变蛋白达30s。最后,另一再生周期后,以500nm的浓度将人cd137配体-fc施加至固定的cd137-fc蛋白达30s,随后直接以5μm的浓度施加突变蛋白达30s,两者均以30μl/min的流速施加。通过以10μl/min的流速施加250mmnaoh中的6m胍-hcl达120s实现芯片表面的再生。目视分析所得传感图,确定结合的cd137配体-fc是否对突变蛋白与固定的cd137-fc蛋白的相互作用有影响。各周期的传感图是cd137配体-fc相互作用或作为对照单独施加的突变蛋白。
图2中提供了突变蛋白seqidno:5、seqidno:12和seqidno:13所得传感图的相关片段的代表性实例。采用实茎箭头(arrowwithasolidstem)标记了相应脂质运载蛋白突变蛋白与单独hucd137-fc结合的spr迹线。采用虚茎箭头(arrowwithabrokenstem)标记了脂质运载蛋白突变蛋白与已用cd137l饱和的hucd137-fc的结合的spr迹线。数据表明,例如,在存在cd137l的情况下,seqidno:5的突变蛋白不能结合hucd137-fc(图2(a))。相对地,在不存在和存在cd137l的情况下,seqidno:12的突变蛋白和seqidno:13的突变蛋白均以非常相似的响应结合hucd137-fc,这表明在脂质运载蛋白突变蛋白和cd137l之间没有结合上的竞争。数据汇总于表2。
表2:
实施例6:结合表达人cd137的细胞的脂质运载蛋白突变蛋白的facs分析
我们使用facs研究来评估脂质运载蛋白突变蛋白和阴性对照与用人cd137稳定转染的中国仓鼠卵巢(cho)细胞(cho-hucd137)的特异性结合。使用flp-in系统(invitrogen),根据制造商的说明产生该细胞系。将模拟转染的flp-incho细胞作为阴性对照。
在补充有10%胎牛血清(fcs,biochrom)和500μg/ml潮霉素b(roth)的ham′sf12培养基中保持转染的cho细胞。根据制造商的说明,在标准条件下(37℃,5%co2气氛)将细胞培养在细胞培养瓶中。为了分离贴壁细胞以用于传代培养或facs实验,根据制造商的说明使用accutase(paa)。
为了实施该实验,用脂质运载蛋白突变蛋白温育cd137-阳性和阴性flp-incho细胞,并采用可通过下述facs分析检测的抗-脂质运载蛋白一抗和荧光标记的二抗标记结合的突变蛋白。
将每孔1×105个细胞在含有5%胎牛血清的冰冷pbs(pbs-fcs)中预温育1h。随后,向细胞中加入通常为10μm至1nm的稀释系列的脂质运载蛋白突变蛋白和阴性对照,并在冰上继续温育1h。使用300g离心将细胞用冰冷的pbs洗涤两次,然后用兔抗-脂质运载蛋白一抗(pieris,多克隆兔抗-hngal和兔抗-htlc;pieris)在冰上温育30min。将细胞用冰冷的pbs洗涤两次,重悬浮于pbs-fcs中并在冰上用被荧光染料alexa488(lifetechnologies)标记的二抗抗-兔抗体温育30min。随后将细胞洗涤并用guavaeasycyteht流式细胞仪分析。通常,将门控设置为排除未成活细胞并记录5.000个事件。以荧光强度的几何平均值表示数值结果。
图3提供了所有测试的克隆的facs直方图(histogram)。在相应的小图(plot)中,描绘了相应脂质运载蛋白突变蛋白的seqidno。根据spr数据(图1,表1),所有突变蛋白均显示与细胞表达的cd137的明确结合。下表2中提供了自该数据的拟合得到的ec50。
表2:
实施例7:采用涂布的脂质运载蛋白突变蛋白的功能t细胞活化分析
我们采用t细胞活化分析来评估一组代表性cd137-结合脂质运载蛋白突变蛋白共刺激t细胞响应的能力。在实施例4中,所测试的突变蛋白(seqidno:13、seqidno:14和seqidno:15)跨越2nm至>23nm的spr亲和力(参见表1)。如实施例5所述,有几种诱导cd137聚集的方法,本实验中我们应用了固定的cd137结合剂。在本实验中,将脂质运载蛋白突变蛋白与抗-人cd3抗体(muronomab,janssen-cilag)一起涂布在塑料皿上,随后在可溶性抗-人cd28抗体(克隆28.2;ebioscience)存在下在涂布的表面上温育纯化的t细胞。将抗-cd3和抗-cd28抗体用于为可被cd137共刺激而共刺激的t细胞提供亚阈值刺激。我们测量了上清液中白细胞介素2(il-2)水平作为读出结果。增加的il-2产生是t细胞活化的标志之一,并且文献(fishert.s.等人,cancerimmunolimmunother(2012)61:1721-1733)中已经描述了通过与抗cd137抗体共刺激的il-2水平的增加。采用seqidno:4作为阴性对照。以下我们提供了本实验的详细描述。
根据biochrom的方案,通过经由聚蔗糖密度梯度(biocoll1.077g/ml,来自biochrom)的离心自血沉棕黄色层中分离健康志愿者供体的人外周血单核细胞(pbmc)。采用pant细胞纯化试剂盒(miltenyibiotecgmbh)和制造商的方案从所得的pbmc中分离t淋巴细胞,将纯化的t细胞重悬浮于由90%fcs和10%dmso组成的缓冲液中,立即用液氮冷冻并储存在液氮中直至进一步使用。对于分析,将t细胞解冻16h并在补充有10%fcs和1%青霉素-链霉素(lifetechnologies)的培养基(rpmi1640,lifetechnologies)中培养。
对于每个实验条件,采用一式三份实施以下程序。使用200μl0.5μg/ml抗-cd3抗体和25μg/ml兔抗-脂质运载蛋白抗体(多克隆兔抗-hngal,pieris)的混合物在4℃下涂布平底组织培养板过夜。后者用于允许通过亲和捕获固定脂质运载蛋白突变蛋白。第二天,用pbs洗涤孔两次,在37℃下在预涂布的板上捕获50μl浓度均为25μg/ml的seqidno:13、seqidno:14和seqidno:15的cd137-结合的脂质运载蛋白质突变蛋白达1h。同样使用seqidno:4并作为阴性对照。再次用pbs洗涤两次之后,向每个孔中加入100μl补充有2μg/mlhcd28抗体的培养基中的t细胞悬浮液(相当于5×104个t细胞)。板用透气密封件(4titude)覆盖,并在37℃下在潮湿的5%co2气氛中温育3天。随后,评估上清液中的il-2。
采用来自r&dsystems的il-2duoset试剂盒来定量合并的细胞培养物上清液中的人il-2水平。第一步,用pbs中稀释的1μg/ml“人il-2捕获抗体”(r&dsystem)在室温下涂布384孔板达2h。随后,使用biotekel405选择cw洗涤器(biotek),用80μlpbs-t(含有0.05%吐温20的pbs)洗涤孔5次。在另外含有1%酪蛋白(w/w)的pbs-t中阻断1h后,合并上清液和在培养基中稀释的浓度系列的il-2标准品在384孔板中在4℃下温育过夜。为了检测和定量捕获的il-2,在含有0.5%酪蛋白的pbs-t中加入100ng/ml生物素化山羊抗-hil-2-bio检测抗体(r&dsystem)与1μg/mlsulfotag标记的链霉亲和素(mesoscalediscovery)的混合物并在室温下温育1h。洗涤后,向每个孔中加入25μl读数缓冲液,并使用mesoscalediscovery读数器读取每个孔的电化学发光(ecl)信号。使用mesoscalediscovery软件进行分析和定量。
图4a中绘制了所得的数据。与阴性对照seqidno:4相比,由于seqidno:14和seqidno:15的脂质运载蛋白突变蛋白的t细胞活化使得上清液中存在明显增加的il-2浓度。本实验表明当涂布在塑料培养皿上时,全部测试的突变蛋白均能够共刺激t细胞响应。
实施例8:采用溶液中的脂质运载蛋白突变蛋白的功能t细胞活化分析
为了测试代表性脂质运载蛋白突变蛋白是否也通过简单的结合而非聚集来活化cd137,按照与实施例5类似的方式进行实施例7的分析,但使用溶解的脂质运载蛋白突变蛋白来代替捕获的脂质运载蛋白突变蛋白。在该分析中,如上所述涂布平底组织培养板,但是仅使用抗-cd3抗体。如上所述处理板直至t细胞添加步骤(包括2μg/mlhcd28),之后加入50μl的浓度为25μg/ml的溶液中的脂质运载蛋白突变蛋白。
图4b中绘制了所得的数据。与阴性对照seqidno:4相比,对于测试的任何脂质运载蛋白突变蛋白,没有由于t细胞活化导致的上清液中il-2浓度的显著增加。本实验表明在足以使全部cd137受体饱和的浓度下,溶液中的单体脂质运载蛋白突变蛋白不共刺激t细胞。
实施例9:采用涂布的脂质运载蛋白突变蛋白的功能t细胞活化分析
为了更详细地研究seqidno:13的突变蛋白共刺激t细胞响应的能力,我们采用如实施例7的t细胞活化分析。我们评估了采用4hbrdu脉冲温育三天后的t细胞继续增殖作为读出结果,并测量上清液il-2和干扰素γ(ifn-g水平)。除了增殖和il-2产生之外,t细胞活化的进一步标志是增加的ifn-g产生,并且在文献(jure-kunkel,m.等人,美国专利7288638)中已经描述了通过采用抗-cd137抗体共刺激的ifn-γ水平的增加。
作为阴性对照,采用野生型样脂质运载蛋白突变蛋白seqidno:4。就一些方面而言,该实验以与实施例8中所述的实验相同的方式进行。以下,我们提供了本实验的详细描述。
根据biochrom的方案,通过经由聚蔗糖密度梯度(biocoll1.077g/ml,来自biochrom)的离心自血沉棕黄色层中分离健康志愿者供体的人外周血单核细胞(pbmc)。采用pant细胞纯化试剂盒(miltenyibiotecgmbh)和制造商的方案从所得的pbmc中分离t淋巴细胞。将纯化的t细胞重悬浮于由90%fcs和10%dmso组成的缓冲液中,立即用液氮冷冻并储存在液氮中直至进一步使用。对于分析,将t细胞解冻16h并在补充有10%fcs和1%青霉素-链霉素(lifetechnologies)的培养基(rpmi1640,lifetechnologies)中培养。
对于每个实验条件,采用一式三份实施以下程序。使用200μl0.5μg/ml抗-cd3抗体和25μg/ml兔抗-脂质运载蛋白骨架抗体(多克隆兔抗-hngal,pieris)的混合物在4℃下涂布平底组织培养板过夜。后者用于允许通过亲和捕获固定seqidno:13。作为阴性对照,用5μg/ml的iggl同种型对照代替抗-cd3抗体与25μg/ml的兔抗-脂质运载蛋白骨架抗体一起涂布。第二天,用pbs洗涤孔两次,在37℃下在预涂布的板上将50μl从50μg/ml至0.8μg/ml的seqidno:13的稀释系列在7个步骤中捕获1h。作为阴性对照,以三个浓度(50μg/ml、25μg/ml、12.5μg/ml)捕获seqidno:4。作为进一步的阴性对照,将seqidno:13以50μg/ml捕获到已经用iggl同种型和抗hngal捕获抗体(见上文)涂布的孔。再次用pbs洗涤两次之后,向每个孔中加入100μl的培养基中的t细胞悬浮液(相当于5×104个t细胞)。这是在存在或不存在浓度为2μg/ml的hcd28抗体的情况下进行的。板用透气密封件(4titude)覆盖,并在37℃下在潮湿的5%co2气氛中温育3天。随后,评估上清液中的il-2和ifn-γ浓度以及增殖。
为了定量t细胞增殖,根据制造商的说明采用基于brdu掺入的化学发光细胞增殖elisa试剂盒(roche)。简言之,第三天,向每个孔中加入10μl的brdu标记溶液,并允许在潮湿的5%co2气氛中在37℃下进一步增殖4h。将板以300g离心10min且将一式三份上清液合并,立即储存在-20℃下以便用于之后的il-2和ifn-γ定量。随后将板在60℃下干燥1h。向每个孔中加入200μl的“fixdenat”溶液并将板在室温下温育30min。用过氧化物酶标记的抗brdu抗体通过在室温下2h温育标记引入的brdu。通过在pherastarfs读数器中定量化学发光过氧化物酶催化的反应来评估brdu水平。
采用来自r&dsystems的il-2duoset和ifn-γduoset试剂盒来定量合并的细胞培养物上清液中的人il-2和ifn-γ水平。对两种细胞因子类似地实施该程序,但仅在以下描述了针对il-2的。第一步,用pbs中稀释的1μg/ml“人il-2捕获抗体”(r&dsystem)在室温下涂布384孔板达2h。随后,使用biotekel405选择cw洗涤器(biotek),用80μlpbs-t(含有0.05%吐温20的pbs)洗涤孔5次。在另外含有1%酪蛋白(w/w)的pbs-t中阻断1h后,合并上清液和在培养基中稀释的浓度系列的il-2标准品在384孔板中在4℃下温育过夜。为了检测和定量捕获的il-2,在含有0.5%酪蛋白的pbs-t中加入100ng/ml生物素化山羊抗-hil-2-bio检测抗体(r&dsystem)与1μg/mlsulfotag标记的链霉亲和素(mesoscalediscovery)的混合物并在室温下温育1h。洗涤后,向每个孔中加入25μl读数缓冲液,并使用mesoscalediscovery读数器读取每个孔的电化学发光(ecl)信号。使用mesoscalediscovery软件进行分析和定量。
图5描绘了本实验的结果。图5a、5c和5e中分别提供了采用抗-cd3和抗-cd28抗体的实验的上清液中的增殖、il-2和ifn-γ的读出结果。图5b、5d和5f中提供了仅采用抗-cd3抗体实施的实验的相同读出结果。
在使用通过抗-cd3和抗-cd28抗体刺激的实验中,增殖率存在明显的剂量依赖性增加(图5a),其是相比阴性对照seqidno:4的高达14倍。在不存在抗-cd3mab(标记为“igg1”的柱)的情况下的增殖是可忽略的。关于il-2产生(图5c),也存在明显的剂量依赖性增加,在6.25μg/ml的seqidno:13的涂布浓度下成最大响应水平和在更高浓度下保持在相比阴性对照高达大约6倍的恒定水平。关于ifn-γ产生(图5e),该模式非常相似,与阴性对照相比,最大ifn-g水平达到高达2.5倍的值。
在采用仅通过抗-cd3mab刺激的实验中,我们再次发现了增殖率明显的剂量依赖性增加(图5a),其是相比阴性对照seqidno:4的高达4倍。在seqidno:13的6.25μg/ml涂布浓度下似乎具有最大响应,与阴性对照相比,其达到15倍的值。在更高和更低的浓度下,响应不太明显。关于ifn-γ产生,存在剂量依赖性增加,在6.25μg/ml的seqidno:13的涂布浓度下成最大响应水平和在更高浓度下保持在相比阴性对照高达大约2.5倍的恒定水平。
总之,本实施例9中所示的实验清楚地证明,在存在和不存在cd28刺激的情况下,关于增殖、il-2产生和ifn-γ产生,通过seqidno:13的突变蛋白的t细胞响应的显著共刺激。
本文示例性所描述的实施方式可以适当地在本文中未具体公开的任何一种或多种要素、一种或多种限制不存在的情况下实施。因此,例如,术语“包含”、“包括”、“含有”等应被宽泛地解读且无限制。另外,本文所采用的术语和表达被用作描述的术语而非限制的术语,并且无意图在使用这些术语和表达时,排除掉显示和描述的特征的任何等同物或其部分,而是认可各种修饰可以在本发明请求保护的范围内。因此,应当理解的是,虽然这些实施方式已通过优选的实施方式和任选的特征而被具体公开,本领域技术人员可以寻求其修饰和变化,并且这样的修饰和变化被认为是在本发明的范围内。本文所述的所有专利、专利申请、教科书和同行评审的出版物均通过引用全文并入本文。此外,当在通过引用并入本文的参考文献中的定义或使用的术语与本文所提供该术语的定义不一致或相反时,适用本文所提供的该术语的定义,而不适用该参考文献中该术语的定义。每个较窄种类和亚属分组落入类属的公开范围内,也形成本发明的一部分。这包括本发明的一般描述,且具有从该种类中取出任何主题的条件或负面限制,而不管该去除的材料是否在本文中具体陈述。此外,当特征是以马库什组的形式描述时,本领域技术人员将认识到,本发明还由此以该马库什组中任何单独成员或成员亚组的形式描述。进一步的实施方式将由以下的权利要求书中变得显而易见。
等同物:本领域的技术人员使用不超过常规实验就会认识到或者能够确定许多于此处描述的本发明的具体实施方式的等同物。以下的权利要求意欲涵盖这些等同物。在本说明书中提到的所有出版物、专利和专利申请在此完整引入本说明书中作为参考,就像每个单独的出版物、专利或专利申请具体地且分别指出引入本文作为参考一样。
- 还没有人留言评论。精彩留言会获得点赞!