T细胞受体的制作方法
发明领域
本发明涉及t细胞受体(tcr)。特别地,本发明涉及工程化的tcr,与相应的种系tcr序列相比,当作为外源tcr表达时,所述工程化的tcr具有高水平的细胞表面表达。本发明还涉及用于增加tcr的细胞表面表达的方法。
发明背景
t细胞受体(tcr)基因疗法是将抗原特异性tcr链转移到受体t细胞,从而将t淋巴细胞的特异性重新定向到感兴趣的靶抗原。
引入的tcr在细胞表面上表达的能力方面差异很大。引入的强表达的tcr与内源tcr共表达,或甚至可以胜过内源tcr的细胞表面表达。当与内源强tcr共表达时,引入的弱表达的tcr不存在于细胞表面(图1)。
在t细胞表面上抗原特异性tcr的高水平表达导致t细胞更高的亲合力,这有利于治疗性tcr疗法的功效。
因此,需要增加弱表达的tcr的细胞表面表达的方法和途径。
发明概述
本发明人已经确定了tcr框架区内许多关键氨基酸位置和残基,其增强tcr细胞表面表达。
因此,在第一方面,本发明提供了包含至以下氨基酸残基中的至少一个的工程化的tcr:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;
其中所述至少一个氨基酸残基不存在于相应的种系tcr氨基酸序列中。
所述至少一个氨基酸残基可以选自以下列表:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;和
β链的r43。
工程化的tcr可以包含α链的l96。
工程化的tcr可以包含β链的r9。
工程化的tcr可以包含β链的y10。
工程化的的tcr可以包含α链的t24。
工程化的tcr可以包含多个如本发明第一方面所定义的氨基酸残基。
工程化的tcr可以包含α链的l96;β链的r9和β链的y10。
工程化的tcr可以包含α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的每一个。
在第二方面,本发明提供了编码如本发明第一方面所定义的tcrα链和/或β链的核酸序列。
核酸序列可以包含通过内部自切割序列或内部核糖体进入位点连接的编码α链的核酸序列和编码β链的核酸序列。
在第三方面,本发明提供了包含根据本发明第二方面的核酸序列的载体。
载体可以是逆转录病毒载体、慢病毒载体或转座子。
在第四方面,本发明提供了包含根据本发明第二方面的核酸序列或根据本发明第三方面的载体的细胞。
细胞可以是t细胞、天然杀伤细胞或干细胞。
细胞可以衍生自从受试者分离的t细胞、天然杀伤细胞或干细胞。
在第五方面,本发明提供了包含表达tcr的细胞的药物组合物,所述tcr包含以下氨基酸残基中的至少一个:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
细胞可以表达包含多个如本发明第一方面所定义的氨基酸残基的tcr。
细胞可以表达由本发明第一方面所定义的工程化的tcr。
在第六方面,本发明涉及根据本发明第五方面的药物组合物,其用于治疗和/或预防疾病。
在第七方面,本发明提供根据本发明第三方面的载体或根据本发明第四方面的细胞,其用于制备用于治疗和/或预防疾病的药物。
在第八方面,本发明涉及用于治疗疾病的方法,其包括向受试者施用根据本发明第四方面的细胞的步骤。
在第九方面,本发明涉及用于产生根据本发明第四方面的细胞的方法,其包括用根据本发明第三方面的载体体外或离体转导细胞的步骤。
在第十方面,本发明提供了用于增加tcr的细胞表面表达的方法,其包括以下步骤:
(i)提供tcrα链和/或β链序列;
(ii)确定在选自以下的一个或多个位置的tcrα链和/或β链序列的氨基酸残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;并且
(iii)将步骤(ii)中列出的一个或多个位置处的氨基酸残基改变为选自以下的氨基酸残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
该方法可以包括改变tcr氨基酸序列使其包含多个如本发明第一方面所定义的氨基酸残基的步骤。
可以通过诱变编码tcr的核酸序列来改变tcr氨基酸序列。
在第十一方面,本发明涉及用于选择高表达tcr的方法,其包括以下步骤:
(i)提供一组tcr氨基酸序列;并且
(ii)选择包含选自以下的至少一个氨基酸残基的tcr氨基酸序列:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;
该方法可以包括选择包含多个如本发明第十一方面所定义的氨基酸残基的tcr氨基酸序列的步骤。
在第十二方面,本发明提供了用于确定tcr的强度的方法,其包括以下步骤:
(i)提供tcrα链和/或β链序列;并且
(ii)确定tcr序列是否包含选自以下的至少一个氨基酸残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;
其中将包含所述氨基酸残基中至少一个的序列确定为高表达tcr,并且将不包含所述氨基酸序列中至少一个的序列确定为低表达tcr。
在第十三方面,本发明提供计算机程序产品,其中以非瞬时方式存储计算机程序,当在处理装置上执行时其使得所述处理装置执行本发明的第十二方面的方法。
在另一方面,本发明提供了本发明的工程化的tcr、核酸序列或载体用于增加tcr的细胞表面表达的用途。
已经证明,在弱表达的tcr中用关键氨基酸残基取代导致表达极大提高。还已经证明,用本发明定义的‘强’氨基酸残基替换‘弱’残基的个别关键残基的单个氨基酸取代以及一系列氨基酸取代组合增加了tcr表达。
氨基酸的变化可通过诱变(例如pcr诱变)经济、快速和有效地实现。增加tcr细胞表面表达的这种方法不会干扰tcr对肽/mhc的识别,并且不影响tcr折叠或结构。因此,tcr表达增加,且对tcr结构的影响最小。
附图简述
图1.a)在也表达内源tcr的细胞中弱外源tcr和强外源tcr的描述。b)tcrα和tcrβ链的示意图-sp:信号肽;fr:框架区;cdr:互补决定区;ig:c区的免疫球蛋白样结构域;cp:连接肽;tm:跨膜结构域;ic:细胞内结构域
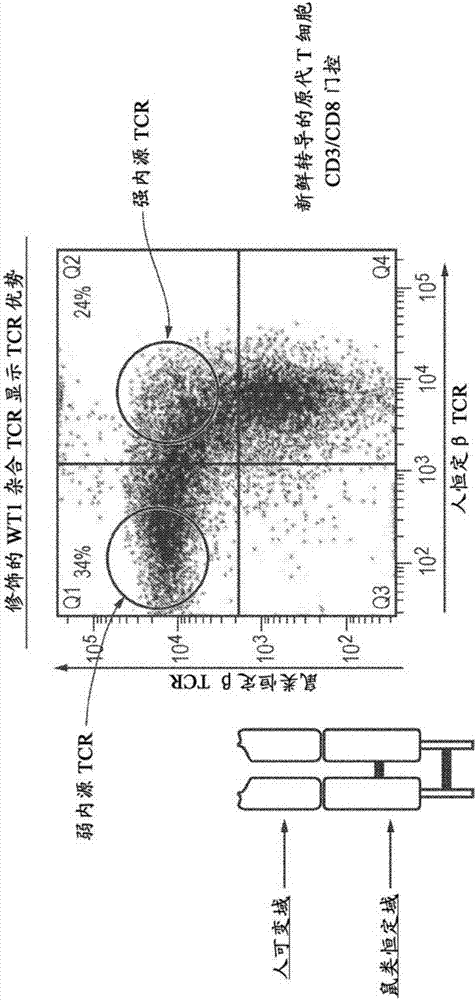
图2.用显示tcr优势的修饰的wt1tcr转导的原代t细胞
图3.强和弱内源tcr的tcr克隆分型
在表中显示的是(i)测序的强和弱α和β可变区段的总数和(ii)在强或弱内源tcr群体中显示优势的α可变区段和β可变区段,以及每个群体中每个可变区段的总数。已经使用imgt命名法对α和β区段进行了分类。
图4.用于评估通用强和弱表达tcr的特征的构建体的图示
图5.在非竞争性环境(没有tcr的jurkat细胞)中的强和弱tcr的表达
图6.在竞争性环境(表达wt1杂合tcr的jurkat细胞)中的强和弱tcr的表达
图7.表达修饰的wt1tcr的低、中和高表达的jurkat细胞中强和弱tcr的表达
图8.强表达和弱表达tcr的生物信息学分析。
a)来自克隆分型的测序信息用于比较已测序的所有强和弱tcr的每个位置上的所有残基。在tcr比对后,在使用trav38-1作为实例的图中将与弱tcr相比在强tcr中特定位置上具有高发生的残基指示为(*)。
b)强和弱tcr可变区段的预测3d结构的分析
图9.在非竞争性环境(不表达tcr的jurkat细胞)和竞争性环境(表达wt1杂合tcr的jurkat细胞)中,所有鉴定的关键残基突变以逆转强和弱表达tcr
图10.非竞争性环境(不表达tcr的jurkat细胞)中个别关键残基的改变影响外源tcr的表达水平
图11.竞争性环境(表达wt杂合tcr的jurkat细胞)中个别关键残基的改变影响外源tcr的表达水平
图12.用于提供或支持一些实施方案的通用计算设备
图13.没有tcr的jurkat细胞(a)和表达修饰的wt1tcr(b)的jurkat细胞,其中用编码所示tcr的pmp71逆转录病毒转导。转导后72h,用抗cd19、v5标签、myc标签和鼠类恒定βtcr抗体染色细胞。细胞对高cd19表达进行门控,并测定v5标签、myc标签和鼠类恒定βtcr表达。对于cd19+高表达细胞,还测定了v5标签、myc标签和鼠类恒定βtcr表达的mfi。(c)没有tcr的jurkat细胞的v5标签和myc标签的mfi。(d)表达修饰的wt1tcr的jurkat细胞的v5标签、myc标签和鼠类恒定βtcr的mfi。
图14.可变域残基变化为l96α、r9β和y10β提高了jurkat细胞中的cmv和wt1tcr表达
图15.可变域残基变化为l96α、r9β和y10β提高了抗原特异性细胞因子的产生
图16.可变域残基变化为l96α、r9β和y10β提高了抗原特异性细胞因子的产生
图17.用良好表达和差表达的tcr转导的细胞含有相似量的tcrmrna
图18.tcrα相对于tcrβ对tcr表达的贡献
发明详述
t细胞受体
在抗原加工过程中,抗原在细胞内被降解,然后通过主要组织相容性复合物(mhc)分子携带至细胞表面。t细胞能够识别抗原递呈细胞表面上的这种肽:mhc复合物。存在两类不同类别的mhc分子,mhci和mhcii,其将来自不同细胞区室的肽递送至细胞表面。
t细胞受体或tcr是存在于t细胞表面的分子,其负责识别与mhc分子结合的抗原。在95%的t细胞中tcr异二聚体由α和β链组成,而5%的t细胞具有由γ和δ链组成的tcr。
tcr与抗原和mhc的接合导致其t淋巴细胞通过一系列由相关酶、辅助受体和特化的辅助性分子介导的生化事件而活化。
tcr的每一条链都是免疫球蛋白超家族的成员,并且拥有一个n端免疫球蛋白(ig)-可变(v)结构域、一个ig恒定(c)结构域、跨膜/细胞膜跨越区以及在c端末端的短胞质尾(图1b)。
tcrα-链和β-链的可变域均具有三个高变或互补决定区(cdr)。cdr3是负责识别经加工的抗原的主要cdr,尽管α链的cdr1也显示出与抗原肽的n端部分相互作用,而β链的cdr1与肽的c端部分相互作用。cdr2被认为识别mhc分子。框架区(fr)定位在cdr之间。这些区域提供tcr可变区的结构。
tcr可变区的集合(repertoire)通过组合连接可变(v)、连接(j)和多样性(d)基因;和n区多样化(由酶脱氧核苷酸转移酶插入的核苷酸)而产生。
α和γ链由v和j区段之间的重组事件形成。β和δ链由涉及v、d和j区段的重组事件形成。
人tcrα基因座,其也包括tcrδ基因座,位于14号染色体上(14q11.2)。tcrβ基因座位于7号染色体上(7q34)。tcrα链的可变区通过46个不同vα(可变)区段之一和58个jα(连接)区段之一的重组而形成(koop等;1994;genomics;19:478-493)。tcrβ链的可变区由54个vβ、14个jβ和2个dβ(多样性)区段之间的重组形成(rowen等;1996;science;272:1755-1762)。
已经鉴定了每个tcr链基因座的v和j(和d,当适用时)基因区段,并且每个基因的种系序列是已知和注释的(例如参见scaviner&lefranc;2000;expclinimmunogenet;17:83-96和folch&lefranc;2000;expclinimmunogenet;17:42-54)。
α链的fr1、cdr1、fr2、cdr2、fr3和cdr3由vα基因编码。fr4由jα基因编码(图1b)。
β链的fr1、crd1、fr2、cdr2和fr3由vβ基因编码。cdr3由dβ基因编码,并且fr4由jβ基因编码(图1b)。
由于每个可变基因的种系序列是本领域已知的(参见如上所述的scaviner&lefranc;和如上所述的folch&lefranc),可以对特定tcr的vα和/或vβ进行测序,并可以鉴定在tcr中所用的种系v区段(参见例如hodges等;2003;jclinpathol;56:1-11,zhou等;2006;laboratoryinvestigation;86;314-321)
tcr结构域的恒定结构域由短的连接序列组成,其中半胱氨酸残基形成二硫键,在两条链之间产生连接。本发明的tcr可在α链和β链的每一条中具有额外的半胱氨酸氨基,使得该tcr在恒定结构域中包含两个二硫键(见下文)。
该结构允许tcr与其他分子结合,所述分子例如在哺乳动物中拥有三种不同链(γ、δ和ε)的cd3和ζ-链。这些辅助分子具有带负电荷的跨膜区,并且对于将信号从tcr传播至细胞内是非常重要的。cd3-和ζ-链与tcr一起形成称为t细胞受体复合物的复合物。
来自t细胞复合物的信号通过特异性辅助受体对mhc分子同时结合而增强。在辅助t细胞上,这一辅助受体是cd4(对ii类mhc是特异性的);而在细胞毒性t细胞上,这一辅助受体是cd8(对i类mhc是特异性的)。辅助受体不仅确保tcr对抗原的特异性,而且还允许延长抗原递呈细胞和t细胞之间的接合,以及募集参与活化的t淋巴细胞的信号传导的细胞内的关键分子(例如,lck)。
因此术语“t细胞受体”在常规意义上用于表示能够识别当由mhc分子递呈时的肽的分子。该分子可以是两条链α和β(或任选地γ和δ)的异二聚体,或其可以是单链tcr构建体。
本发明还提供了来自此类t细胞受体的α链或β链。
本发明的tcr可以是包含衍生自超过一种物种的序列的杂合tcr。例如,惊讶地发现了鼠类tcr被发现在人t细胞中比人tcr更有效地表达。因此,tcr可包含人可变区和鼠类恒定区。这一方法的缺陷是该鼠类恒定序列可能引发免疫应答,导致对所转移t细胞的排斥。然而,用于使得患者准备好过继性t细胞治疗的调理方案可以导致足够的免疫抑制,以允许表达鼠类序列的t细胞的植入。
根据本发明第一方面的工程化的tcr包含至少一个本文定义的氨基酸残基,其不由种系vα或vβ基因编码。换句话说,本发明的工程化的tcr包含α链和/或β链,其包含与由未改变的种系vα或vβ基因编码的相应的α链和/或β链相比较,在本文所述的一个或多个位置处的改变的氨基酸残基,其中改变的氨基酸残基氨基酸是本文所定义的残基。
本文鉴定的氨基酸残基根据国际immunogenetics信息系统’(imgt)编号。该系统是本领域众所周知的(lefrance等;2003;devcompimmunol;27:55-77),并且基于可变区结构的高度保守性。编号考虑并结合了fr和cdr的定义、x射线衍射研究的结构数据和高变环的表征。
在igmt编号系统中定义了fr和cdr区域的定界。fr1区域包含位置1-26(25-26个氨基酸,取决于v-gene组或亚组),具有23位的第1个cys。fr2区域包含位置39-55(16-17个氨基酸),具有41位的保守的trp。fr3区域包含位置66-104(36-39个氨基酸,取决于vgene组或亚组),具有89位的保守的疏水性氨基酸和104位的第二个cys。igmt编号系统的残基1是fr1中的第一个残基。igmt编号系统的残基104是fr3中的最后一个残基。
如此,本文使用的编号系统是指适用时氨基酸在整个α链或整个β链中的位置。
因此,igmt编号允许对tcr可变区内的氨基酸位置进行标准化描述,并与待进行种系编码的序列进行比较。
适用于产生根据本发明第一方面的工程化的tcr的方法是本领域已知的。
例如,可以进行诱变以改变编码tcr的核酸序列中的特定核苷酸。此类诱变将改变tcr的氨基酸序列,使得它包含至少一个根据本发明的氨基酸残基。
诱变方法的实例是quikchange方法(papworth等;1996;strategies;9(3);3-4)。该方法涉及使用一对互补诱变引物以在热循环反应中使用高保真非链置换dna聚合酶,例如pfu聚合酶扩增模板核酸序列。
本发明人已经确定tcr框架区的特定位置中特定氨基酸残基的存在改变了tcr的细胞表面表达。
如本文所用,细胞表面表达与表达强度同义。如此,‘强’或‘高’表达等同于tcr的高水平的细胞表面表达。‘弱’或‘低’表达等同于tcr的低水平的细胞表面表达。
增加tcr的细胞表面表达意味着相对于包含由种系序列编码的氨基酸序列的等同tcr,包含根据本发明的至少一个氨基酸残基的tcr具有更高水平的细胞表面表达。包含由种系序列编码的氨基酸序列的等同tcr是指未被改变为包含根据本发明的氨基酸残基的tcr,即未改变的tcr在特定位置具有野生型氨基酸残基。
在一个实施方案中,本发明的工程化的tcr可以具有比包含未修饰的种系tcr序列的相应tcr高至少1.5、2、2.5、3、4、5、6、7、8、9或10倍的细胞表面表达。
可以通过本领域已知的方法测定tcr的细胞表面表达。例如,可以使用本领域已知的常规流式细胞术方法测定tcr的细胞表面表达(参见例如shapiro;practicalflowcytometry;john2005;science)。
例如,tcr的细胞表面表达可以表示为细胞群体中tcr表达的平均荧光强度(mfi)(参见shapiro;如上所述)。在一个实施方案中,表达本发明的工程化的tcr的细胞群体的mfi可以比表达包含未修饰的种系tcr序列的相应tcr的相应细胞群体的mfi高至少1.5、2、2.5、3、4、5、6、7、8、9或10倍。
tcr的细胞表面表达可以表示为群体内在细胞表面上表达tcr的细胞百分比。可以使用本领域熟知的常规流式细胞术方法来测定此类百分比(参见例如shapiro;如上所述和henel等;2007;labmedicine;38;7;428-436)。
在一个实施方案中,本发明的工程化的tcr与包含未修饰的种系tcr序列的相应tcr相比可以由群体中至少2、至少5、至少10、至少15、至少20、至少25、至少30、至少40、至少50、至少60或至少70%的更多细胞表达。
在一个实施方案中,与包含未修饰的种系tcr序列的相应tcr相比,工程化的tcr的细胞表面表达增加,但是编码工程化的tcr或未修饰的种系tcr的mrna的相对水平基本相同。如本文所用,“基本上相同”可以意指mrna水平相差例如小于1.5倍。可以使用本领域已知的方法测定相对mrna水平;例如rt-qpcr,northern印迹和流式细胞术rna测定法(例如,如图17所示)。
因此,在第一方面,本发明提供了包含以下氨基酸残基中的至少一个的工程化的tcr:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;
其中所述至少一个氨基酸残基不存在于相应的种系tcr氨基酸序列中。
所述至少一个氨基酸残基可以选自以下列表:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;和
β链的r43。
在一个实施方案中,所述至少一个氨基酸残基可以选自以下列表:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;和
α链的s86。
工程化的tcr可以包含α链的l96。在一个实施方案中,工程化的tcr可以包含α链的l96和β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的l96;β链的r9和β链的y10。
工程化的tcr可以包含α链的v19;β链的r9和β链的y10。
工程化的tcr可以包含β链的r9。在一个实施方案中,工程化的tcr可以包含β链的r9和α链的l96;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含β链的y10。在一个实施方案中,工程化的tcr可以包含β链的y10和α链的l96;β链的r9;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的的tcr可以包含α链的t24。在一个实施方案中,工程化的tcr可以包含α链的t24和α链的l96;β链的r9;β链的y10;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的v19。在一个实施方案中,工程化的tcr可以包含α链的v19和α链的l96;β链的r9;β链的y10;α链的t24;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的t20。在一个实施方案中,工程化的tcr可以包含α链的t20和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的m50。在一个实施方案中,工程化的tcr可以包含α链的m50和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的t5。在一个实施方案中,工程化的tcr可以包含α链的t5和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的q8。在一个实施方案中,工程化的tcr可以包含α链的q8和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的s86。在一个实施方案中,工程化的tcr可以包含α链的s86和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的f39。在一个实施方案中,工程化的tcr可以包含α链的f39和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含α链的d55。在一个实施方案中,工程化的tcr可以包含α链的d55和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
工程化的tcr可以包含β链的r43。在一个实施方案中,工程化的tcr可以包含β链的r43和α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的至少一个。
本文所用的术语“至少一个”可以包括一个、二个、三个、四个、五个、六个、七个、八个、九个、十个、十一个、十二个、十三个、十四个、十五个、十六个、十七个、十八个、十九个、二十个或更多个如本文所述的氨基酸残基。
工程化的tcr可以包含多个上述氨基酸残基。换句话说,tcr可以包括至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个或至少20个上述氨基酸残基。
工程化的tcr可以在β链中包含r9和y10。
在一个实施方案中,工程化的tcr可以包含α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101中的每一个;
在一个实施方案中,工程化的tcr可以包含α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;和β链的r43中的每一个。
在一个实施方案中,工程化的tcr可以包含α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;和α链的s86中的每一个。
保守取代
本发明还包括工程化的tcr,其包含选自以下的氨基酸残基的保守取代:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
至少一个氨基酸残基可以选自:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;或
β链的r43。
至少一个氨基酸残基可以是本文所述的任何氨基酸残基或多个氨基酸残基。
可在残基的极性、电荷、溶解度、疏水性、亲水性和/或两亲性性质相似的基础上进行保守氨基酸取代,只要tcr的高表达得以保持。例如,带负电的氨基酸包括天冬氨酸和谷氨酸;带正电的氨基酸包括赖氨酸和精氨酸;以及具有不带电极性头基团(其具有类似亲水值)的氨基酸包括亮氨酸、异亮氨酸、缬氨酸、甘氨酸、丙氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、苯丙氨酸和酪氨酸。
例如可根据下表进行保守取代。第二列相同区块中以及优选在第三列相同行中的氨基酸可彼此取代:
本发明还包括同源取代(取代和替换在本文中均用于表示现存氨基酸残基与另一残基的互换),即类似-对-类似取代,诸如碱性对碱性、酸性对酸性、极性对极性等。
核酸序列
本发明还提供核苷酸序列,其编码本发明第一方面的工程化的tcr受体的核苷酸序列或其一部分,例如α链或β链的可变序列;α链和/或β链。
如本文所用,术语“多核苷酸”、“核苷酸”和“核酸”意在彼此同义。
本领域技术人员将理解,由于遗传密码的简并性,许多不同的多核苷酸和核酸可编码相同的多肽。此外,应当理解,技术人员可以使用常规技术进行核苷酸替换,其不影响本文所述的多核苷酸编码的多肽序列,以反映其中待表达多肽的任何特定宿主生物的密码子使用。
根据本发明的核酸可以包含dna或rna。它们可以是单链或双链的。它们也可以是这样的多核苷酸,在它们中包括合成或修饰的核苷酸。对寡核苷酸的许多不同类型的修饰是本领域已知的。这些包括甲基膦酸酯和硫代磷酸酯骨架,在分子的3'和/或5'端加入吖啶或多聚赖氨酸链。为了本文所述的用途的目的,应当理解,可以通过本领域可获得的任何方法修饰多核苷酸。可以进行此类修饰以便增强感兴趣多核苷酸的体内活性或存活期。
核苷酸序列可以是双链或单链的,并且可以是rna或dna。
核苷酸序列可以是经密码子优化的。不同的细胞在具体密码子的利用上是不同的。这种密码子偏好与细胞类型中特定trna的相对丰度的偏好相对应。通过改变序列中的密码子,从而将它们特制成与相应trna的相对丰富相匹配,可能增加表达。
包括hiv和其他慢病毒在内的许多病毒使用大量的罕见密码子,而通过将这些变成以对应于常用的哺乳动物密码子,可实现哺乳动物生产细胞中包装成分的表达增加。哺乳动物细胞以及多种其他生物的密码子使用表是本领域公知的。
密码子优化还可包括移除mrna不稳定基序和隐藏的剪接位点。
核酸序列可以包含使编码α链的核酸序列和β链的核酸序列能够从相同的mrna转录物表达的核酸序列。
例如,核酸序列可以包含编码α链和β链的核酸序列之间的内部核糖体进入位点(ires)。ires是允许在mrna序列中间翻译起始的核苷酸序列。
核酸序列可以包含通过内部自切割序列连接的编码α链的核酸序列和β链的核酸序列。
内部自切割序列可以是使包含α链的多肽和包含β链的多肽能够变得分离的任何序列。
切割位点可以是自切割的,使得当产生多肽时,其立即被切割成单个肽,而不需要任何外部切割活性。
术语“切割”在本文中用于方便,但是切割位点可以通过除经典切割之外的机制将肽分成单个实体。例如,对于口蹄疫病毒(fmdv)2a自切割肽(见下文),已经提出了各种模型来解释“切割”活性:宿主细胞蛋白酶的蛋白水解、自体蛋白水解或翻译效应(donnelly等(2001)j.gen.virol.82:1027-1041)。只要位于编码蛋白质的核酸序列之间时,切割位点导致蛋白质被表达为单独的实体,那么这种“切割”的确切机制对于本发明的目的来说就并不重要。
自切割肽可以是来自口疮病毒或心脏病毒的2a自切割肽。
载体
本发明还提供了包含本文所述的核苷酸序列的载体。
术语“载体”包括表达载体,即能够在体内或体外/离体表达的构建体。
病毒递送系统包括但不限于腺病毒载体、腺相关病毒(aav)载体、疱疹病毒载体、逆转录病毒载体、慢病毒载体、杆状病毒载体。
逆转录病毒是rna病毒,其具有与裂解性病毒(lyticvirus)不同的生命周期。在这点上,逆转录病毒是通过dna中间体复制的传染性实体。当逆转录病毒感染细胞时,其基因组通过逆转录酶转化为dna形式。该dna拷贝充当模板用于产生新的rna基因组、以及组装传染性病毒颗粒所需的病毒编码蛋白。
存在许多逆转录病毒,例如鼠类白血病病毒(mlv)、人免疫缺陷病毒(hiv)、马传染性贫血病毒(eiav)、小鼠乳腺癌病毒(mmtv)、劳氏肉瘤病毒(rsv)、fujinami肉瘤病毒(fusv)、莫洛尼氏鼠白血病病毒(mo-mlv)、fbr鼠骨肉瘤病毒(fbrmsv)、莫洛尼氏鼠肉瘤病毒(mo-msv)、abelson鼠白血病病毒(a-mlv)、禽髓细胞增生病毒29(mc29)、以及禽骨髓成红细胞增多症病毒(avianerythroblastosisvirus;aev),以及所有其他逆转录病毒科,包括慢病毒。
逆转录病毒的详细列表可在coffin等(“retroviruses”1997coldspringharbourlaboratorypress编辑:jmcoffin,smhughes,hevarmuspp758-763)中找到。
慢病毒也属于逆转录病毒科,但它们可感染分裂细胞和非分裂细胞(lewis等(1992)emboj.3053-3058)。
载体可以能够将根据本发明第二方面的核苷酸转移至细胞中,诸如t细胞中。理想情况下,该载体应当能够在t细胞中持续高水平表达,使得引入的tcr可成功地与内源tcr竞争有限的cd3分子池。
载体可以是逆转录病毒载体。该载体可基于或可衍生自mp71载体骨架。该载体可缺乏土拨鼠肝炎反应元件(woodchuckhepatitisresponseelement,wpre)的全长或截短版本。
为有效感染人细胞,病毒颗粒可以可用双嗜性包膜或长臂猿白血病病毒包膜进行包装。
在基因修改的细胞中,增加cd3分子供应可增加tcr表达。因此该载体还可包含cd3-γ、cd3-δ、cd3-ε和/或cd3-ζ的基因。该载体可仅包含cd3-ζ的基因。所述基因可通过自切割序列连接,诸如2a自切割序列。备选地,可提供一个或多个编码cd3基因的单独的载体用于与tcr-编码载体共转移。
细胞
本发明还涉及包含根据本发明的核苷酸序列的细胞。该细胞可表达本发明第一方面的t-细胞受体。
该细胞可以是t细胞。该t细胞可以是任何t细胞亚群,包括例如αβt细胞、γδt细胞或调节性t细胞。
该细胞可以是天然杀伤细胞。
该细胞可衍生自从受试者分离的细胞。该细胞可以是从受试者中分离的混合细胞群,诸如外周血淋巴细胞(pbl)群的一部分。
pbl群中的t细胞可通过本领域公知的方法进行活化,诸如应用抗-cd3和cd28抗体。
t细胞可以是cd4+辅助t细胞或cd8+细胞毒性t细胞。该细胞可在cd4+辅助t细胞/cd8+细胞毒性t细胞的混合群中。多克隆活化,例如使用抗cd3抗体任选地与抗cd28抗体组合,将引发cd4+和cd8+t细胞的增殖,但还可引发cd4+25+调节性t细胞的增殖。
在一个实施方案中,t细胞是cd8+细胞毒性t细胞。
在一个实施方案中,与表达包含未修饰的种系tcr序列的相应tcr的细胞相比,表达根据本发明的工程化的tcr的细胞可具有增加的功能活性。
增加的功能活性可以例如是指抗原与tcr结合后,细胞的增加的细胞因子产生。细胞因子可以选自ifnγ、il-2、gm-csf、tnfα、il-4、il-5、il-6、il-9、il-10和il-13。
该细胞因子可以是ifnγ和/或il-2。
如本文所用,“增加的细胞因子产生”是指-在抗原与tcr结合后-由表达根据本发明的工程化的tcr的细胞产生的细胞因子的量大于由表达包含未修饰的种系tcr序列的相应的tcr的等同细胞产生的量。
由表达根据本发明的工程化的tcr的细胞产生的细胞因子的量可以高于由表达包含未修饰的种系tcr序列的相应的tcr的等同细胞产生的量至少1.5倍、2倍、2.5倍、3倍、4倍、5倍、10倍、20倍或50倍。
可以使用本领域已知的方法,例如流式细胞术或elisa测定细胞产生的细胞因子的量。
本发明还提供了产生根据本发明的细胞的方法,其包括用根据本发明的载体体外或离体转染或转导细胞的步骤。
该细胞可从待过继性转移遗传修饰的细胞的受试者中分离。在这一方面,该细胞可通过从受试者中分离t细胞、任选地活化该t细胞、离体tcr基因转移,并随后通过过继性转移该tcr-转导的细胞而免疫治疗受试者而制得。
或者,该细胞可从不同的受试者分离,使得其为同种异体的。该细胞可分离自供体受试者。例如,如果该受试者在进行同种异体造血干细胞移植(allo-hsct),则该细胞可源自获得该hsc来源的供体。如果该受试者在进行或已进行实体器官移植,则该细胞可源自获得该实体器官来源的受试者。
或者,该细胞可以是、或衍生自干细胞,诸如造血干细胞(hsc)。将基因转移至hsc中不会导致在细胞表面表达tcr,因为干细胞不表达cd3分子。然而,当干细胞分化为迁移至胸腺的淋巴前体(lymphoidprecursor)时,cd3表达的启动将导致在胸腺细胞中引入的tcr的表面表达。这一方法的优点是成熟t细胞一旦产生,其仅表达引入的tcr,而很少表达或不表达内源tcr链,因为引入的tcr链的表达抑制了内源tcr基因片段重排形成功能性tcrα和β基因。
其他的益处是,基因修饰的干细胞是具有期望的抗原特异性的成熟t细胞的持续来源。因此该细胞可以是基因修饰的干细胞,其分化后产生表达本发明第一方面的tcr的t细胞。本发明还提供了通过诱导包含根据本发明的核苷酸序列的干细胞分化,从而产生表达本发明第一方面的tcr的t细胞的方法。
干细胞方法的缺陷是在胸腺中的t细胞发育期间,具有期望的特异性的tcr可能会被删除,或当在周围t细胞中表达时可能诱导耐受性。另一个可能的问题是干细胞中插入诱变的风险。
药物组合物
本发明还提供了包含根据本发明的载体或细胞的药物组合物。
药物组合物可另外包含药学上可接受的载体、稀释剂或赋形剂。药物组合物可任选地包含一种或多种另外的药学活性多肽和/或化合物。这样的制剂可以是例如适于静脉内输注的形式。
用途
本发明还涉及包含根据本发明的细胞的药物组合物,其用于治疗和/或预防疾病。
‘治疗’涉及本发明的细胞的治疗用途。本文中,细胞可以施用于具有现有疾病或病症的受试者,以便减轻、减少或改善与疾病相关的至少一种症状和/或减缓、减少或阻止疾病进展。
‘预防’涉及本发明的细胞预防性用途。本文中,此类细胞可以施用于尚未罹患疾病和/或没有显示任何疾病症状的受试者以预防或削弱疾病的起因或减少或预防与该疾病相关的至少一种症状的发展。该受试者可以具有疾病的倾向或被认为有形成疾病的风险。
本发明还提供了根据本发明的工程化的tcr;核酸序列或载体来增加tcr的细胞表面表达的用途。
本发明还提供了根据本发明的工程化的tcr;核酸序列或载体来提高细胞的功能活性的用途。功能活性可以是如本文所述的任何功能活性。
治疗方法
本发明提供了用于治疗和/或预防疾病的方法,其包括将本发明的细胞(例如在上述的药物组合物中)施用于受试者的步骤。
该方法可以包括以下步骤:
(i)分离含细胞的样品;
(ii)用本发明提供的核酸序列或载体转导或转染这些细胞;
(iii)向受试者施用来自(ii)的细胞。
本发明还提供了本发明的细胞,其用于治疗和/或预防疾病。
在本发明的优选实施方案中,本文所述的任何方法的受试者是哺乳动物,优选猫、狗、马、驴、绵羊、猪、山羊、牛、小鼠、大鼠、兔或豚鼠,但是最优选受试者是人。
对于本文所述的细胞的施用,可以使用本领域中常见或标准的任何细胞群体施用模式,例如,通过适当途径的注射或输注。在一个方面,人类每kg施用高达5x108个细胞。因此,例如,体重为100kg的人可以接受每次治疗高达5x1010个细胞的剂量。如果需要,可以在稍后时间重复剂量。
本发明还涉及根据本发明的载体或细胞在制备用于治疗和/或预防疾病的药物中的用途。
疾病
待通过本发明的方法和用途治疗和/或预防的疾病可以是诱导t细胞介导的免疫应答的任何疾病。
例如,待通过本发明的方法和用途治疗和/或预防的疾病可以是癌性疾病,例如膀胱癌、乳腺癌、结肠癌、子宫内膜癌、肾癌(肾细胞)、白血病、肺癌、黑素瘤、非霍奇金淋巴瘤、胰腺癌、前列腺癌和甲状腺癌。
待通过本发明的方法和用途治疗和/或预防的疾病可以是感染,例如细菌或病毒感染。
方法
本发明还提供了增加tcr的细胞表面表达的方法,其包括以下步骤:
(i)提供tcrα链和/或β链序列;
(ii)确定在选自以下的一个或多个位置处的tcrα链和/或β链的氨基酸残基:
α链的96;β链的9;β链的10;α链的24;α链的19;α链的20;α链的50;α链的5;α链的8;α链的86;α链的39;α链的55;β链的43;α链的66;β链的19;β链的21;β链的103;α链的3;α链的7;α链的9;α链的11;α链的16;α链的18;α链的21;α链的22;α链的26;α链的40;α链的47;α链的48;α链的49;α链的51;α链的52;α链的53;α链的67;α链的68;α链的74;α链的76;α链的79;α链的81;α链的83;α链的90;α链的92;α链的93;和α链的101;并且
(iii)将步骤(ii)中列出的一个或多个位置处的氨基酸残基改变为选自以下的氨基酸残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
如本文所述,每个可变基因的种系序列是本领域已知的(参见scaviner&lefranc;如上和folch&lefranc;如上),因此可以通过测序和与已知的种系序列比较来确定在tcr中使用的vα和/或vβ种系v区段(参见例如hodges等;如上,zhou等;2006;如上)。因此,在一个或多个位置确定tcrα链和/或β链的氨基酸残基的本发明步骤具体涉及测定存在于本文所述的一个或多个特定位置的氨基酸残基。
该方法可以包括确定存在于如本文所述的任何位置或多个位置处的氨基酸残基。
可以改变tcr的氨基酸序列,使得其包含本文所述的任何氨基酸残基或任何多个氨基酸残基。
在一个实施方案中,本发明提供了用于增加tcr的细胞表面表达的方法,其包括改变tcr氨基酸序列的步骤,使得其包含至少一个选自以下的残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
所述至少一个氨基酸残基可以选自:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;或
β链的r43。
可以改变tcr的氨基酸序列,使得其包含多个如上文定义的氨基酸残基。
可以使用本领域熟知的方法改变tcr的氨基酸序列。例如,可以通过编码tcr的核酸序列的诱变来改变tcr的氨基酸序列。
增加tcr的细胞表面表达意味着相对于包含由种系序列编码的氨基酸序列的等同tcr,包含至少一个根据本发明的氨基酸残基的tcr具有更高水平的细胞表面表达。包含由种系序列编码的氨基酸序列的等同tcr是指未被改变为包含根据本发明的氨基酸残基的tcr,即未改变的tcr在特定位置具有野生型氨基酸残基。
可以使用本文所述的任何方法来确定tcr的细胞表面表达。
本发明还涉及用于选择高表达tcr的方法,该方法包括以下步骤:
(i)提供一组tcr氨基酸序列;并且
(ii)选择包含至少一个选自以下的氨基酸残基的tcr氨基酸序列:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101。
所述至少一个氨基酸残基可以选自:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;或
β链的r43。
该方法可以包括选择包含如上定义的任何氨基酸残基或任何多个氨基酸残基的tcr的步骤。
在另一方面,本发明提供了用于确定tcr的强度的方法,其包括以下步骤:
(i)提供tcrα链和/或β链序列;并且
(ii)确定tcr序列是否包含至少一个选自以下的氨基酸残基:
α链的l96;β链的r9;β链的y10;α链的t24;α链的v19;α链的t20;α链的m50;α链的t5;α链的q8;α链的s86;α链的f39;α链的d55;β链的r43;α链的a66;β链的v19;β链的l21;β链的l103;α链的t3;α链的s7;α链的p9;α链的m11;α链的a16;α链的t18;α链的l21;α链的s22;α链的d26;α链的f40;α链的s47;α链的r48;α链的q49;α链的i51;α链的l52;α链的v53;α链的t67;α链的e68;α链的n74;α链的f76;α链的n79;α链的q81;α链的a83;α链的k90;α链的s92;α链的d93;和α链的m101;
其中将包含至少一个所述氨基酸残基的序列确定为高表达tcr,并且将不包含所述氨基酸序列中的至少一个的序列确定为低表达tcr。
所述至少一个氨基酸残基可以选自:
α链的l96;
β链的r9;
β链的y10;
α链的t24;
α链的v19;
α链的t20;
α链的m50;
α链的t5;
α链的q8;
α链的s86;
α链的f39;
α链的d55;或
β链的r43。
α链和/或β链序列可以是氨基酸序列或编码α链和/或β链的核酸序列。
该方法可以包括确定存在于如本文所述的任何位置或多个位置处的氨基酸残基。
当在细胞例如t细胞中作为外源tcr表达时,此类方法可用于预测tcr例如治疗性tcr的表面表达水平。
计算机程序产品
图12示意性地示出了可以用于实现上述技术的类型的通用计算装置100。在本发明的上下文中,这可以是例如构成较大系统的一部分的嵌入式处理器。通用计算装置100包括通过总线122连接在一起的中央处理单元102、随机存取存储器104和只读存储器106。它还包括网络接口卡108、硬盘驱动器110、显示驱动器112和监视器114以及具有通过公共总线122连接的键盘118和鼠标120的用户输入/输出电路116。在操作中,例如当形成较大系统的一部分时,中央处理单元102将执行可以例如存储在随机存取存储器104和/或只读存储器106中的计算机程序指令。程序指令可以另外地从硬盘驱动器110检索或经由网络接口卡108动态下载。所执行的处理结果可以经由连接的显示驱动器112和监视器114显示给用户或工程师。可以经由连接的用户输入输出电路116从键盘118或鼠标120接收用于控制通用计算装置100的操作的用户输入。应当理解,可以以各种不同的计算机语言写计算机程序。计算机程序可以本地存储在记录介质上或动态地下载到通用计算装置100。当在适当的计算机程序的控制下操作时,通用计算装置100可以执行上述技术并且可以被认为形成用于执行任何上述技术的设备。通用计算装置100的架构可以显著变化,而图13仅仅是一个示例。通用计算装置100还可以具有允许其提供指令执行环境(即,虚拟机)的配置。
现在将通过实施例进一步描述本发明,这些实施例旨在帮助本领域普通技术人员实施本发明,并不意图以任何方式限制本发明的范围。
实施例
实施例1:与强或弱tcr表达相关的tcr可变区段的测定
在用tcr转导t细胞后,引入的tcr相对于内源性tcr在细胞表面上的表达能力差异很大。强表达的外源性tcr与内源性tcr共表达,或甚至可以胜过内源性tcr的细胞表面表达。弱表达的外源tcr当与强tcr共表达时不存在于细胞表面,或表达很差(图1a)。
用编码wt1(wilms肿瘤1)tcr的pmp71载体转导激活的原代t细胞,所述载体含有鼠类化(murinised)的恒定域和恒定域中另外的半胱氨酸(整个tcr被密码子优化)。
该修饰的wt1tcr显示tcr显性(图2)。转导后72小时,用抗cd3和cd8抗体以及抗人tcr恒定域和抗鼠类tcr恒定域抗体染色t细胞。在cd3+和cd8+t细胞群体上门控t细胞。表达强的内源性tcr的cd8+t细胞被抗鼠类和抗人tcr恒定域抗体共染色。表达弱内源性tcr的cd8+t细胞仅被抗鼠类tcr恒定域抗体染色。
表达强和弱内源性tcr的cd8+t细胞群体通过fac分选和克隆分型以确定其可变的α和β链的使用。在图3所示的表中显示的是(i)被测序的强和弱的α和β可变区段的总数,以及(ii)在强或弱内源性tcr群体中显示显性的α可变区段和β可变区段,以及每个群体中每个可变区段的总数。使用imgt命名法对α和β区段进行了分类。
实施例2:在非竞争和竞争环境中表达通用强和弱tcr
克隆分型数据用于确定强和弱tcr群体中最普遍使用的α和β可变区段。使用该信息改造通用强和弱tcr。
强tcr是可变α38.2和可变β7.8。弱tcr是可变α13.2和可变β7.3。由于单克隆抗体对上述α和β可变区段不是商业上可用的,为了测定转导的t细胞中的tcr表达,v5标签存在于α链的开始处,并且myc标签存在于β链的开始处。此外,为了能够标准化转导效率,在构建体的末端存在截短的鼠类cd19(图4)。
两个构建体含有相同的α和β恒定域,并且整个构建体是非密码子优化的。此外,独特的限制性位点与α和β区段接界,以允许容易地交换α和β区段。将tcr克隆到pmp71逆转录病毒中用于转导实验。
在非竞争性环境中表达强和弱tcr
用编码通用强或弱tcr的pmp71载体转导不含内源性tcrα或β链的jurkat细胞。转导后72小时,用抗cd19、v5标签和myc标签抗体染色jurkat细胞(图5)。jurkat细胞对高cd19表达进行门控,并测定这些cd19+高表达细胞上的v5标签和myc标签表达。此外,还测定了cd19+高表达细胞的v5标签和myc标签的mfi。
在竞争性环境中表达强和弱tcr
用编码通用强或弱tcr的pmp71载体转导表达修饰的wt1tcr的jurkat细胞。转导后72小时,用抗cd19、v5标签和myc标签抗体染色jurkat细胞(图6)。jurkat细胞对高cd19表达进行门控,并测定了v5标签和myc标签表达。还测定了cd19+高表达细胞的v5标签和myc标签的mfi。
在表达低、中、高表达的修饰的wt1tcr的jurkat细胞中强和弱tcr的表达
用编码通用强或弱tcr的pmp71载体转导表达修饰的wt1tcr的jurkat细胞。转导后72小时,用抗cd19、鼠类tcr恒定域、v5标签和myc标签抗体染色jurkat细胞(图7)。jurkat细胞首先对高cd19表达进行门控,然后对鼠类tcr恒定域的高、中、低表达进行门控。对于所有三个单独的wt1tcr群体和wt1表达细胞作为整体,测定了v5标签和myc标签表达。还测定了v5标签和myc标签的mfi。
实施例3:生物信息学分析以鉴定与tcr表达强度相关的关键特征
使用来自克隆分型的测序信息来比较已测序的所有强和弱tcr的每个位置处的所有残基。在tcr比对后,与弱tcr相比,在强tcr中特定位置处具有高发生率的残基在图中以(*)表示,其使用trav38-1作为实例(图8a)。
在强tcr中显示出普遍性(prevalence)的残基被进一步分析和精制,以仅包括对稳定tcr链的结构和配对有影响的框架区中的残基。使用trav38-2和trbv7-8作为骨架,这些残基被高亮标记(*)(图8b)。上线是强tcr中的残基。底线是弱tcr中的等同残基。
实施例4:强表达和弱表达tcr的工程化
为了使用弱残基工程化强表达tcr,使用强tcr作为骨架,并且用相关位置处的弱tcr残基替换被预测对tcr强度重要的所有强残基。
为了使用强残基工程化弱tcr,使用弱tcr作为骨架,并且用相关位置处的弱tcr残基替换被预测对tcr强度重要的所有强残基。
用所述tcr转导没有tcr的jurkat细胞(顶行)和表达wt1鼠类化tcr的jurkat细胞(底行),并在72小时后用抗cd19、v5标签和myc标签抗体染色。jurkat细胞对高cd19表达进行门控,并测定了v5标签和myc标签表达。还测定了cd19+高表达细胞的v5标签和myc标签的mfi(图9)。
实施例5:突变个体残基对tcr表达的影响
强残基对tcr表达的个体影响
使用通用弱tcr作为骨架,并且使用快速变化pcr诱变技术制备一系列tcr,其仅具有预测的强残基之一替换相关位置处的弱tcr残基,同时保留其余的弱tcr骨架。
标记突变的tcr,使得字母表示氨基酸,数字是tcr框架中的位置,并且α或β表示残基在α链或β链中。最后一张图,底行,右侧有3个残基变化,即α链上的v19和β链上的r9、y10(图10和11)。
没有tcr的jurkat细胞(图10)和表达修饰的wt1tcr的jurkat细胞(图11),其中用编码所示tcr的pmp71逆转录病毒转导。转导后72小时,转导的细胞用抗cd19、v5标签和myc标签抗体染色。细胞对高cd19表达进行门控,并测定了v5标签和myc标签表达。
图13显示了没有tcr的jurkat细胞(a)和表达修饰的wt1tcr的jurkat细胞(b)的实验结果,其中用编码所示tcr的pmp71逆转录病毒转导。转导后72小时,细胞用抗cd19、v5标签、myc标签和鼠类恒定βtcr抗体染色。细胞对高cd19表达进行门控,并测定了v5标签、myc标签和鼠类恒定βtcr表达。还测定了cd19+高表达细胞的v5标签、myc标签和鼠类恒定βtcr表达的mfi。(c)没有tcr的jurkat细胞的v5标签和myc标签的mfi。(d)表达修饰的wt1tcr的jurkat细胞的v5标签、myc标签和小鼠恒定βtcr的mfi。
结果显示,α链的f39、α链的d55和β链的r43增加了tcr的表面表达。此外,上述三个残基的突变在jurkat细胞竞争实验中也显示降低在细胞表面上表达的wt1tcr的量。
实施例6:突变可变域残基对抗原特异性tcr中l96α、r9β和y10β的影响
jurkat细胞中的细胞表面表达
用编码未修饰的野生型cmv或wt1tcr或被突变以含有l96α、r9β和y10β的相同tcr的pmp71载体转导jurkat细胞(其不含有内源性tcrα或β链)。转导后72小时,用抗cd19abs和cmv四聚体或vβ2.1(以测定wt1可变β表达)abs染色jurkat细胞。
细胞首先对高cd19表达进行门控,然后测定cmv四聚体和或vβ2.1表达(图14)。
原代t细胞中的细胞表面表达
用编码未修饰的野生型cmv或wt1tcr或被突变以含有l96α、r9β和y10β的相同tcr的pmp71载体转导活化的原代t细胞。转导后72小时,t细胞用抗cd3、cd8和cd19abs和cmv四聚体或vβ2.1abs染色。细胞对cd3+和cd19高t细胞进行门控,然后使用四聚体或抗vβ2.1抗体分别检测cmv-和wt1-tcr以测定tcr表达(图15)。
用编码未修饰的野生型cmv或被突变以含有l96α、r9β和y10β的相同tcr的pmp71载体转导活化的原代t细胞。转导后5天,t细胞与表达pcmv肽或无关肽pwt235的apc共培养。共培养18小时后,收集上清液,并通过elisa测定il-2和ifn-γ的产生(图16)。
用良好表达和差表达的tcr转导的细胞含有相似量的tcrmrna
用编码通用强tcr或通用弱tcr或含有预测弱残基的通用强tcr或含有预测强残基的通用弱tcr的pmp71载体转导没有内源性tcr的jurkat细胞。affymetrixprimeflowrna测定用于测定表达恒定αmrna和恒定βmrna的cd19+高细胞的百分比。测定了四个群体的恒定α和βmrna的mfi。此外,还测定了表达v5标签的cd19+高jurkat细胞的百分比(图17)。
实施例7:tcrα对tcrβ对tcr细胞表面表达的贡献
用编码强tcr,或弱tcr,或被工程化成含有强α可变链和弱β可变链的tcr,或被工程化成含有弱α可变链和强β可变链的tcr的pmp71载体转导没有内源性tcr的jurkat细胞。转导后72h,用抗cd19、v5标签和myc标签抗体染色jurkat细胞。jurkat细胞对高cd19进行门控,并测定了v5标签和myc标签表达。还测定了cd19+高细胞中v5标签和myc标签的mfi(图18)。
在上面说明书中提及的所有公开物通过引用引入本文。本发明的所述方法和系统的各种修改和变形对本领域技术人员是显而易见的,而不会背离本发明的范围和精神。尽管本发明已结合特定优选的实施方案进行了描述,应当理解如权利要求所要求保护的本发明不应当被不正当地限定至此类具体实施方案。实际上,所描述的实施本发明的方式的对分子生物学、细胞免疫学或相关领域技术人员而言显而易见的各种修改也在以下权利要求的范围之内。
- 还没有人留言评论。精彩留言会获得点赞!