OsARD1基因在提高水稻耐水淹方面的应用的制作方法
技术领域
本发明属于水稻整体生长发育情况改进的技术领域,涉及OsARD1基因在水稻耐水淹方面的应用,特别是能够增强水稻耐水淹的能力,减轻水稻遭遇洪涝灾害的损失,对保障世界人民粮食安全及农业可持续发展具有重要意义。
背景技术:
水稻是我国第一大粮食作物,也是单产水平最高的粮食作物,其种植面积不足粮食作物的30%,产量却占粮食总产量的40%以上;我国有60%以上人口以稻米为主食。在水稻的生产过程中往往遭受到诸多生物的和非生物的危害。水稻是一种半水生植物,喜欢在温暖湿润的地带生长,水稻生长的地区常常多雨,所以洪涝灾害是制约水稻生产与发展的非生物逆境胁迫因素之一。弄清水稻耐淹的分子机理,提高水稻的耐淹能力,培育耐水淹的水稻品种,减轻水稻遭遇洪涝灾害的损失,对保障世界人民粮食安全及农业可持续发展具有重要意义。
当植物受到淹水胁迫时,植物会对胁迫产生应答,为了适应氧气、二氧化碳及光都缺乏的水淹环境,植物通过两种策略[1]来适应淹水的环境,一种是低氧逃逸策略(low O2 escape strategy, LOES),即在水淹时植物的茎和叶片快速伸长,使植物更快的接触空气,恢复正常的有氧代谢,这种策略的不足是植物的快速生长会消耗大量的有机物,如果对于长期或者较深的淹水时,由于植物体内大量有机物的消耗可能在还没有接触空气时植物已经死亡。另一种完全相反的策略是静止策略,即在较深的淹水情况下,植物的生长减慢,保存体内的有机物,待到水位退去以后再进行生长。植物激素乙烯在植物淹水时发挥了重要的作用,低氧的环境可以促进乙烯的合成,研究表明乙烯能够在水淹的时候促进植物的伸长[2],乙烯在淹水时促进植物细胞的伸长是通过与其他的激素相互作用来实现的,乙烯的增加会抑制脱落酸(ABA)的活性,从而促进赤霉素(GA)的活性,GA促进细胞的分裂和伸长[3],所以乙烯在植物淹水情况下起到十分重要的作用。
顺式还原酮加双氧酶(Acireductone Dioxygenase,ARD)是乙烯合成途径中的一个重要的酶,首先,顺式还原酮加双氧酶催化顺式还原酮生成KMTB (2-Keto-4-methylthiobutyrate),KMTB进而被催化形成甲硫氨酸(Met)。Met是乙烯合成途中的重要物质,植物中乙烯的合成是首先通过Met催化形成S-腺苷甲硫氨酸,S-腺苷甲硫氨酸由ACC合成酶催化形成乙烯前体1-aminocyclopropane-1-carboxylic acid (ACC),ACC被氧化后形成乙烯[4],所以ARD位于乙烯合成的上游。在细菌里,Dai等[5]发现在肺炎克雷伯菌(Klebsiela pneumoniae)中有两种ARD酶,这两种酶的蛋白质构成完全相同,由同一个基因编码,但由于结合的金属离子不同,却行使了完全相反的功能。很多研究表明真核生物中参与甲硫氨酸循环的两个ARD蛋白同样行使着完全不同的功能,Fe-ARD催化顺式还原酮形成KMTB进而促进甲硫氨酸(Met)的合成,而Ni-ARD参与催化顺式还原酮生成甲基丙酸乙酯,顺式还原酮的消耗使甲硫氨酸的合成减少[6]。目前OsARD1具体的生物学功能仍然未知,前人研究结果表明水淹可以诱导OsARD1的表达[7]。我们进一步构建了OsARD1的过表达载体获得了其过表达植株,发现过量表达OsARD1能够增强水稻耐水淹的能力。
技术实现要素:
本发明公开了一种水稻耐水淹基因OsARD1核苷酸序列,其特征在于OsARD1基因的核苷酸序列为SEQ ID No.1, OsARD1所编码的氨基酸序列为SEQ ID No.2。
本发明进一步公开了水稻耐水淹基因OsARD1核苷酸序列在提高水稻的耐水淹能力方面的应用。
本发明公开的水稻耐水淹基因OsARD1核苷酸序列如下:
ATGGAGAACGAATTCCAGGATGGTAAGACGGAGGTGATAGAAGCATGGTACATGGATGATAGCGAAGAGGACCAGAGGCTTCCTCATCACCGCGAACCCAAAGAATTCATTCATGTTGATAAGCTTACAGAACTAGGAGTAATCAGCTGGCGCCTAAATCCTGATAACTGGGAGAATTGCGAGAACCTGAAGAGAATCCGCGAAGCCAGAGGTTACTCTTATGTGGACATTTGTGATGTGTGCCCAGAGAAGCTGCCAAATTATGAAACTAAGATCAAGAGTTTCTTTGAAGAACACCTGCATACCGATGAAGAAATACGCTATTGTCTTGAAGGGAGTGGATACTTTGATGTGAGAGACCAAAATGATCAGTGGATTCGTATAGCACTGAAGAAAGGAGGCATGATTGTTCTGCCTGCAGGGATGTACCACCGCTTTACGTTGGACACCGACAACTATATCAAGGCAATGCGACTGTTTGTTGGCGATCCTGTTTGGACACCCTACAACCGTCCCCATGACCATCTTCCTGCAAGAAAGGAGTTTTTGGCTAAACTTCTCAAGTCAGAAGGTGAAAATCAAGCAGTTGAAGGCTTCTGA
OsARD1所编码的氨基酸序列为:
MENEFQDGKTEVIEAWYMDDSEEDQRLPHHREPKEFIHVDKLTELGVISWRLNPDNWENCENLKRIREARGYSYVDICDVCPEKLPNYETKIKSFFEEHLHTDEEIRYCLEGSGYFDVRDQNDQWIRIALKKGGMIVLPAGMYHRFTLDTDNYIKAMRLFVGDPVWTPYNRPHDHLPARKEFLAKLLKSEGENQAVEGF。
附图说明
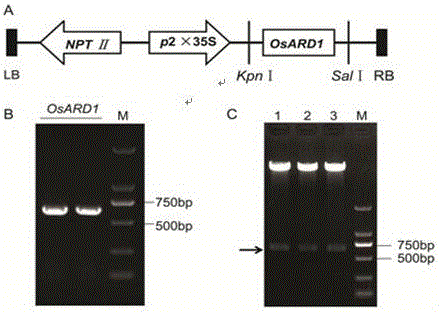
图1. 为OsARD1过表达载体的构建;其中A.过表达载体的结构;LB:T-DNA的左边界;NPTⅡ:新霉素磷酸转移酶抗性标记基因;2×35S:两个串联排列的花椰菜病毒(CaMV)35S启动子;OsARD1:OsARD1的全长ORF片段;RB:T-DNA的右边界。B.目的基因OsARD1全长ORF片段的PCR扩增。C.过表达重组载体酶切鉴定,1-3为3个不同的单克隆。M为Marker标记;
图2为OsARD1过表达转基因植株的表型及表达分析;其中A. T0代过表达转基因植株中NPTⅡ基因的PCR检测;1-22为获得的40株中的22个转基因单株,P: pCAMBIA2300空载体质粒; WT: 转入pCAMBIA2300空载体对照;CK: 正常未转化的野生型中花11;所用引物为载体中NPTⅡ基因特异引物;B. 部分T0代过表达转基因植株的表达分析;WT为空载体对照转基因植株;1-18为1-6号株系的18株过表达转基因植株; C. T1代转基因植株表达分析。WT为空载对照体转基因植株,#2~#6为4个独立的过表达转基因植株. D. 过表达转基因植株表型;WT为空载对照体转基因植株;OsARD1-OV为过表达转基因植株;
图3为盐处理情况下表型分析;其中A. 水淹处理9天后,野生型和过表达植株处理表型;(WT为转入空载体的转基因植株,OsARD1-OV为过表达转基因植株) B. 水淹处理9天后,野生型和过表达植株地上部分表型比较;C. 水淹处理9天后,地上部分长度比较; D. 水淹条件下,OsARD1表达分析比较;E. 水淹条件下,OsAdh1表达分析比较;F. 水淹条件下,OsSub1C表达分析比较; (WT为转入空载体的转基因植株,OsARD1-OV为过表达转基因植株)。
具体实施方式
下面通过具体的实施方案叙述本发明。除非特别说明,本发明中所用的技术手段均为本领域技术人员所公知的方法。另外,实施方案应理解为说明性的,而非限制本发明的范围,本发明的实质和范围仅由权利要求书所限定。对于本领域技术人员而言,在不背离本发明实质和范围的前提下,对这些实施方案中的物料成分和用量进行的各种改变或改动也属于本发明的保护范围。本发明所用原料及试剂均有市售。其中的粳稻品种中花11有市售。
实施例1
1 材料和方法
1.1 材料
粳稻品种中花11(Oryza sativa L. ssp. Japonica cv. Zhonghua 11)及其转基因植株用于本研究,田间材料均种植于天津市天津师范大学试验田,田间保持日常管理。用于处理的材料均种植于人工气候箱中(Climacell MMM,德国)。
1.2 OsARD1过表达载体的构建
OsARD1目的基因用RT-PCR方法获得,总RNA从60天的成熟水稻叶片中提取,用M-MLV反转录酶(Vazyme,南京)反转录合成cDNA,以其为模板,用OsARD1特异性引物
OsARD1-F: 5’-TTAggtaccTTCCACCCCGCAATCCACAT-3’
和OsARD1-R: 5’-GTTgtcgacGTGCAGGAGCCCAACAAAAC-3’(小写字母部分为酶切位点序列)进行PCR扩增获得OsARD1完整的ORF(Open Reading Frame)。
PCR扩增程序为:95℃ 1 min;94℃ 30s,62℃ 30s,72℃ 30s,运行35个循环,所用DNA聚合酶为高保真特性的酶(Vazyme,南京)。
目的片段凝胶检测正确后进行纯化回收,然后与pCAMBIA2300空载体进行连接,获得重组载体。对重组载体进行酶切及测序验证正确后转入农杆菌(EHA105)中,通过农杆菌的遗传转化法获得OsARD1过表达转基因植株,同时转入pCAMBIA2300空载体作为对照,转化方法参照Hiei等的转化方法(Hiei et al., 1994)。
1.3 转基因植株的分子检测
待T0代过表达转基因植株在田间长到约60天后,我们对其进行了转基因分子检测。用CTAB法(张凤娟等, 2004)提取叶片基因组DNA,然后用pCAMBIA2300载体中独有的新霉素磷酸转移酶基因(NPTⅡ)对获得的过表达转基因植株进行检测。
所用引物为NPTⅡ-F: 5’-TTCTCACTGAAGCGGGAAGGG-3’
和NPTⅡ-R: 5’-GCGATACCGTAAAGCACCAGG-3’;
PCR扩增程序为:94℃ 4 min;94℃ 30s,57℃ 30s,72℃ 30s,运行30个循环。
1.4 水淹胁迫处理及表达分析
将种在土里刚刚发芽一天的野生型和OsARD1-OV过表达转基因幼苗同时放入35cm深的蒸馏水中,观察野生型植株与过表达转基因植株处理的表型变化,每天观察表型并拍照记录。
对于淹水相关基因的表达分析,将种在土里正常生长培养的10天大小的野生型和过表达转基因幼苗,放入35cm深的蒸馏水中经行处理,水淹7天,退水后正常恢复生长3天。分别提取淹水0、1、3、5、7天和退水后恢复1天和3天的野生型和OsARD1-OV转基因幼苗的总RNA进行表达分析,每次取样时间为上午10点。
2 结果与分析
2.1 OsARD1过表达载体的构建
从水稻成熟叶片中提取总RNA,反转录获得cDNA后利用RT-PCR的方法获得OsARD1的全长ORF,经琼脂糖凝胶检测后发现目的片段大小与预期相符(图2-B)。将凝胶纯化回收后的目的片段与含有2×35S启动子的空载体pCAMBIA2300经限制性内切酶KpnⅠ和SalⅠ进行双酶切,利用T4 DNA连接酶16 ℃过夜连接获得重组载体。重组载体经酶切检测,酶切出预期的目的条带(图2-C),表明OsARD1的全长ORF片段已成功的连入过表达空载体中,获得最终的过表达载体(图2-A)。测序验证后,转入农杆菌中以备后续转基因实验。
2.2 过表达转基因植株的获得及表达分析
利用农杆菌介导的遗传转化方法,将重组载体转入野生型中花11中获得OsARD1 过表达转基因植株。通过转化获得了6个独立株系共40株T0代转基因植株,经转基因分子检测后发现有37株为转基因阳性植株(图3-A),田间观察过表达转基因植株与空载体对照表型,二者没有明显差异。我们进一步提取了过表达转基因植株的总RNA,用RT-PCR方法分析了6个独立株系的18个转基因植株中目的基因OsARD1的表达,结果表明,OsARD1的表达在不同株系过表达转基因植株中的表达明显提高(图3-B),说明过表达载体在转基因植株中起到了过量表达的作用,正常工作。收获T0代转基因植株种子后,来年我们选取了4个过表达株系在大田里进行种植获得T1代转基因株系,并检测了T1代植株中目的基因的表达情况,发现OsARD1在4个过表达株系中表达量均明显提高(图3-C),进一步表明构建的过表达载体能够正常工作,而且可以稳定的遗传到后代。进一步观察T1代转基因植株的表型,与对照相比,仍未见明显差异,图为#5株系与野生型的表型(图3-D)。由于前人研究OsARD1受水淹胁迫的诱导,我们推测OsARD1可能与淹水的耐受性相关,因此我们接下来用水淹胁迫处理观察转基因植株的表型。
2.3 过表达OsARD1提高了水稻对淹水的耐受能力
将种在土里刚刚发芽一天的过表达和野生型幼苗放入35 cm深的蒸馏水中进行淹水处理,结果显示,OsARD1过表达转基因幼苗的胚芽鞘和叶片的长度均明显长于野生型对照植株,过表达转基因植株的叶片在第六天的时候就露出了水面,接触了空气,而野生型幼苗叶片生长到第8天才露出水面,表明OsARD1过表达转基因植株在淹水条件下的生长速度快于野生型对照。为了进一步直观的观察过表达转基因植株和野生型的生长情况,在水淹9天后,将幼苗从土中拔出后,可以明显的观察到过表达转基因植株茎和叶片的长度长于野生型幼苗,通过进一步测量可知,OsARD1过表达转基因幼苗比野生型幼苗的长度平均长7 cm。我们进一步探究了水淹相关基因在野生型和过表达转基因植株中的表达情况。在水淹的情况下,OsARD1在野生型植株表达明显升高了,在退水恢复正常生长后OsARD1的表达下降,表明OsARD1受水淹诱导。而在过表达转基因植株中,水淹处理后OsARD1的表达量升高的更加显著,并且在退水恢复期OsARD1的表达也没有降低。水淹情况下,乙醇发酵途径中关键基因OsAdh1的突变体胚芽鞘的伸长会受到抑制[7]。在我们的水淹表达分析实验中,OsAdh1基因的表达水平在过表达转基因植株中均高于野生型对照,这与过表达转基因植株胚芽鞘长度长于野生型的表型相符。而对于水稻耐水淹基因Sub1C,水淹处理前三天过表达转基因植株中的表达量高于野生型,水淹后3天要低于野生型植株,表明过表达OsARD1能更快诱导水稻耐淹基因Sub1C的表达,增强了水稻的耐淹性。
经过水淹实验我们发现,过表达转基因植株胚芽鞘和叶片的长度均长于野生型对照植株,所以水淹情况下,OsARD1过表达转基因植株可以通过让胚芽鞘和叶片的更快速的伸长,使植株更快地接触空气和光,从而恢复植株正常的有氧代谢和光合作用,保证有足够的养分提供水淹部分的生长。表明过量表达OsARD1能在一定程度上增强水稻的耐水淹能力。
参考文献:
[1] Bailey-Serres J, Voesenek LA. Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol. 2008, 59: 313-339.
[2] Jackson MB. Ethylene-promoted elongation: an adaptation to submergence stress. Ann Bot. 2008, 101(2): 229-248.
[3] Fukao Takeshi, Bailey-Serres Julia. Ethylene—A key regulator of submergence responses in rice. Plant Sci. 2008, 175:43–51
[4] Rzewuski Guillaume, Sauter Margret. Ethylene biosynthesis and signaling in rice. Plant Sci. 2008,175(1): 32-42.
[5] Dai Y, Wensink P C, Abeles R H.One Protein, Two Enzymes. J Biol Chem, 1998, 274(3): 1193-1195.
[6] Friedman EJ, Wang HX, Jiang K, Perovic I, Deshpande A, Pochapsky TC, Temple BR, Hicks SN, HardenTK, JonesAM.Acireductone dioxygenase 1 (ARD1)isan effector ofthe heterotrimeric G protein beta subunit in Arabidopsis. J Biol Chem. 2011, 286(34): 30107-30118.
[7] Hiroaki Saika, Hideo Matsumura, Tesuo Takano, Nobuhiro Tsumi, Mikio Nakazono. A Point Mutation of Adh1 Gene is Involved in the Repression of Coleoptile Elongation under Submergence in Rice. Breediong Science,2006, 56(1):69-74。
SEQUENCE LISTING
<110> 天津师范大学
<120> OsARD1基因在提高水稻耐水淹方面的应用
<160> 2
<170> PatentIn version 3.5
<210> 1
<211> 600
<212> DNA
<213> 人工序列
<400> 1
atggagaacg aattccagga tggtaagacg gaggtgatag aagcatggta catggatgat 60
agcgaagagg accagaggct tcctcatcac cgcgaaccca aagaattcat tcatgttgat 120
aagcttacag aactaggagt aatcagctgg cgcctaaatc ctgataactg ggagaattgc 180
gagaacctga agagaatccg cgaagccaga ggttactctt atgtggacat ttgtgatgtg 240
tgcccagaga agctgccaaa ttatgaaact aagatcaaga gtttctttga agaacacctg 300
cataccgatg aagaaatacg ctattgtctt gaagggagtg gatactttga tgtgagagac 360
caaaatgatc agtggattcg tatagcactg aagaaaggag gcatgattgt tctgcctgca 420
gggatgtacc accgctttac gttggacacc gacaactata tcaaggcaat gcgactgttt 480
gttggcgatc ctgtttggac accctacaac cgtccccatg accatcttcc tgcaagaaag 540
gagtttttgg ctaaacttct caagtcagaa ggtgaaaatc aagcagttga aggcttctga 600
<210> 2
<211> 199
<212> PRT
<213> 顺式还原酮加双氧酶
<400> 2
Met Glu Asn Glu Phe Gln Asp Gly Lys Thr Glu Val Ile Glu Ala Trp
1 5 10 15
Tyr Met Asp Asp Ser Glu Glu Asp Gln Arg Leu Pro His His Arg Glu
20 25 30
Pro Lys Glu Phe Ile His Val Asp Lys Leu Thr Glu Leu Gly Val Ile
35 40 45
Ser Trp Arg Leu Asn Pro Asp Asn Trp Glu Asn Cys Glu Asn Leu Lys
50 55 60
Arg Ile Arg Glu Ala Arg Gly Tyr Ser Tyr Val Asp Ile Cys Asp Val
65 70 75 80
Cys Pro Glu Lys Leu Pro Asn Tyr Glu Thr Lys Ile Lys Ser Phe Phe
85 90 95
Glu Glu His Leu His Thr Asp Glu Glu Ile Arg Tyr Cys Leu Glu Gly
100 105 110
Ser Gly Tyr Phe Asp Val Arg Asp Gln Asn Asp Gln Trp Ile Arg Ile
115 120 125
Ala Leu Lys Lys Gly Gly Met Ile Val Leu Pro Ala Gly Met Tyr His
130 135 140
Arg Phe Thr Leu Asp Thr Asp Asn Tyr Ile Lys Ala Met Arg Leu Phe
145 150 155 160
Val Gly Asp Pro Val Trp Thr Pro Tyr Asn Arg Pro His Asp His Leu
165 170 175
Pro Ala Arg Lys Glu Phe Leu Ala Lys Leu Leu Lys Ser Glu Gly Glu
180 185 190
Asn Gln Ala Val Glu Gly Phe
195
- 还没有人留言评论。精彩留言会获得点赞!