一种高催化效率且耐高温木聚糖酶突变体及其制备方法和应用与流程
本发明涉及基因工程领域,具体地涉及一种高催化效率且耐高温木聚糖酶突变体及其制备方法和应用。
背景技术:
纤维素、半纤维素和木质素是构成植物细胞壁的主要成分,约占了世界上50%生物量。半纤维素主要存在于细胞壁的表面,纤维素包裹在半纤维素中,而半纤维素又与木质素共价连接形成网状结构。木聚糖是半纤维素的主要成分,是自然界中含量仅次于纤维素的可再生资源,几乎占地球有机碳含量的三分之一。木聚糖的结构复杂,其主链是由吡喃木糖以β-d-1,4-木糖苷键连接起来,并带有多种取代基。
由于木聚糖的多样性和结构的复杂性,它的完全水解需要多种酶的协同作用才能完成。包括木聚糖酶、木糖苷酶以及水解侧链取代基团的α-d-葡萄糖醛酸苷酶、乙酰木聚糖酯酶、阿魏酸酯酶、香豆酸酯酶等。通过这些酶的协同作用,木聚糖能够有效地被水解。其中,木聚糖酶能够降解木聚糖主链的β-1,4-糖苷键,是木聚糖降解过程中发挥最重要作用的酶。木聚糖酶在饲料工业、食品工业、造纸工业以及能源、纺织、医药等行业均具有广泛的应用价值。
木聚糖酶的分子改良方面主要集中在热稳定性机理研究及改良。ding等(2013)通过分子动力学模拟的方法研究来源于streptomyceslividans的木聚糖酶sl-xlna的热不稳定性区段,确定影响sl-xlna热稳定性的因素。gallardo等通过强制进化的方法使来源于paenibaciiusbarcinonensis的木聚糖酶xyn10b的热稳定性提高20倍。kamondi等通过familyshuffling的方法,将两个10家族的木聚糖酶进行随机片段的替换,发现有4个杂合体的最适温度得到了提高。但在第10家族木聚糖酶其它性质的机制及改良方面的研究还很少,尤其在第10家族木聚糖酶高效催化机制及改良方面的研究还未见报道。
技术实现要素:
本发明的一个目的是对材料xyl10c截掉n端序列后获得的高催化效率木聚糖酶以及材料xyl10c中e175位点突变后获得的高催化效率木聚糖酶突变体的氨基酸序列进行保护,并详细研究n端序列以及e175位点在xyl10c的高催化效率中所扮演的角色。
本发明的再一目的是提供包含e175饱和突变体基因的重组载体。
本发明的另一目的是提供包含e175饱和突变体基因的重组菌株。
突变体的野生型氨基酸序列全长如seqidno.1所示。
msfhsllisgllasvavavpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqselptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
突变体的野生型成熟氨基酸序列如seqidno.2所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqselptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求1突变体的野生型氨基酸去掉n端序列如seqidno.3所示。
wglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqselptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175h氨基酸序列如seqidno.4所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqshlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175n氨基酸序列如seqidno.5所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsnlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175d氨基酸序列如seqidno.6所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsdlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175v氨基酸序列如seqidno.7所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsvlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175l氨基酸序列如seqidno.8所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsllptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175q氨基酸序列如seqidno.9所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsqlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
权利要求2突变体e175m氨基酸序列如seqidno.10所示。
vpkeawgitvtetktvsttiiatvtelgtcsstitsptsdatttttssatntnptttllatpqpsnwglnnaaradgklwfgtaadipgleqddryymkeynnthdfggttpanimkfmftepeqnvfnftgaqefldiafashklvrchnliwqsmlptwvtnpttnwtnetlskvlqnhvytlvshfgdqcyswdvvnealsddpagsyqnniwfdtigpeyvamafeyaekavkdhklnvklyyndynieypgpkstaaqnivkelkarniqidgvgleshfiagetpsqatqitnmadftsldidvavteldvrlylppnatseaqqvadyyatvaacaatercigitvwdfddtyswvpstfagqgyadlffqpdgpntplvkkaaydgclqalqhkaesp
本发明还是提供一种制备所述木聚糖酶突变体的方法,其技术方案如下:
1、设计突变引物如表1所示,获得突变体的完整基因片段;
2、将获得突变基因片段克隆到表达载体ppic9上,获得突变体重组质粒;
3、将突变体重组质粒转化毕赤酵母gs115,诱导表达,获得高催化效率
聚糖酶。
本发明还提供了一种制备木聚糖酶的方法,包括以下步骤:
1、用上述的重组载体转化酵母宿主细胞,得重组菌株;
2、培养重组菌株,诱导重组木聚糖酶表达;
3、回收并纯化所表达的木聚糖酶。
本发明通过对成熟的木聚糖酶xyl10c截掉n端以及e175位点实施饱和突变构建了包括去掉e175位点在内的21个突变体,将突变体重组载体转化毕赤酵母gs115,小管诱导表达发现,除了xyl10c-cute,xyl10c-e175w三个突变体的小管上清没有检测到活性之外,其余19个突变体都有活性。
对19个有活性的突变体和野生酶xyl10c进行大瓶诱导,蛋白浓缩纯化后进行了基本酶学性质的测定,如表2所示,以榉木木聚糖测定发现,e175位点饱和突变中的18个突变体以及突变体xyl10c-cutn最适温度在85度左右,,最适ph在4.0-4.5左右,彼此之间的ph作用范围相类似。
突变体xyl10c-cutn、e175h、e175n、e175d、e175v、e175l、e175q、e175m的比活较野生酶xyl10c分别提高了2.1倍、2.4倍、1.7倍、1.9倍、2.0倍、1.4倍2.5倍和4.3倍。突变体xyl10c-cutn、e175h、e175n、e175d、e175v、e175l、e175q、e175m的催化效率较野生酶xyl10c分别提高了1.7倍、2.3倍、2.1倍、1.4倍、1.9倍、1.7倍、2.3倍和4.9倍。
本发明证明确认了材料xyl10c中的关键催化相关的位点e175位点,并证明了该位点对于该酶高催化效率高比活的重要性,即e175位点在该酶的高比活高催化效率中扮演着重要的角色。对于研究材料xyl10c的高效催化机制提供了重要的线索,同时对于提高该家族的其他木聚糖酶的催化效率提供了可靠的参考位点,证明了该位点在本家族木聚糖酶催化效率中的决定性作用。
附图说明
图1:gh10家族木聚糖酶xyl10c-cutn的晶体结构以及e175位点在三维结构中所处的位置。
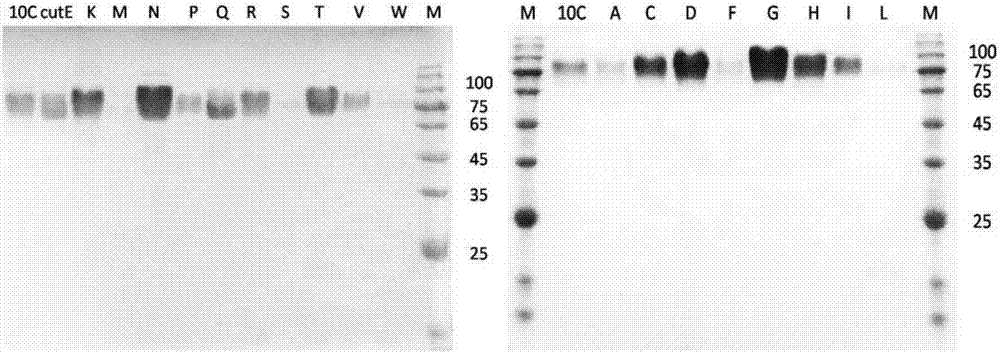
图2:野生酶xyl10c,突变体xyl10c-cutn以及18个有活性的突变体的蛋白纯化电泳图片。
图3:(图3-1和图3-2)野生酶xyl10c,突变体xyl10c-cutn以及18个有活性的突变体的最适ph。
图4:(图4-1和图4-2)野生酶xyl10c,突变体xyl10c-cutn以及18个有活性的突变体的最适温度。
具体实施方式
以下结合附图对本发明作进一步的描述。
表1.以xyl10c为母本的xyl10c-cutn以及e175饱和突变体构建引物
试验材料和试剂
1、菌株及载体:表达宿主pichiapastorisgs115,表达质粒载体ppic9r为本实验室保存。
2、酶类及其它生化试剂:内切酶购自fermentas公司,重组酶购自全式金公司,榉木木聚糖购自sigma公司。其它都为国产分析纯试剂(均可从普通生化试剂公司购买得到)。
3、培养基:
(1)lb培养基:0.5%酵母提取物,1%蛋白胨,1%nacl,ph7.0。
(2)ypd培养基:1%酵母提取物,2%蛋白胨,2%葡萄糖。
(3)md固体培养基:2%葡萄糖,1.5%琼脂糖,1.34%ynb,0.00004%biotin。
(4)bmgy培养基:1%酵母提取物,2%蛋白胨,1%甘油(v/v),1.34%ynb,0.00004%biotin。
(5)bmmy培养基:1%酵母提取物,2%蛋白胨,1.34%ynb,0.00004%biotin,0.5%甲醇
(v/v)。
实施例1突变体xyl10c–cutn重组载体的制备
以bisporasp.mey-1来源的gh10家族木聚糖酶xyl10c-ppic9r为模板,在氨基酸序列wglnn所对应的核酸序列处设计引物xyl10c-cutn—f以及整个氨基酸序列结尾处所对应的核酸序列设计引物xyl10c-cutn—r,扩增产物经过ecorⅰ和notⅰ双酶切后直接与切开的ppic9r质粒进行连接,转化transi-t1感受态。
实施例2以xyl10c为母本的e175饱和突变体重组载体的构建
在基因xyl10c的e175位点处设计饱和突变引物e175x—f和e175x—r,以xyl10c--ppic9r为模板以xyl10c-f和e175x—r以及e175x—f和xyl10c-r为引物pcr,扩增出上下游片段后,再以xyl10c-f和xyl10c-r为引物进行overlap,产物经过ecorⅰ和notⅰ双酶切后直接与切开的ppic9r质粒进行连接,转化transi-t1感受态。挑取单克隆测序发现,只有突变体e175fe175me175te175w以及cute没有出现,因此针对剩余的几个突变体重新和成引物,方法同上。
实施例3木聚糖酶突变体重组菌株的制备
将与表达载体ppic9r连接测序正确的重组载体用内切酶bglii线性化并转化毕赤酵母gs115,获得重组酵母菌株gs115/xyl10c-cutn,gs115/xyl10c-e175x。
挑取取含有重组质粒的gs115菌株,接种于2mlbmgy培养基的10ml小管中,置于30℃,220rpm摇床培养48h;后将培养液3000g离心5min,弃上清,沉淀用1.5ml含有0.5%甲醇的bmmy培养基重悬,并再次置于30℃,220rpm条件下诱导培养48h。最后取上清用于酶活性检测。
挑取上清酶活最高的gs115菌株,接种于30mlypd培养基中培养48h进行种子放大培养,后按1%接种体积接种于300mlbmgy培养基的1l三角瓶中,置于30℃,220rpm摇床培养48h;后将培养液3000g离心5min,弃上清,沉淀用200ml含有0.5%甲醇的bmmy培养基重悬,并再次置于30℃,220rpm条件下诱导培养。每隔12h补加1ml甲醇,使菌液中的甲醇浓度保持在0.5%,同时取上清用于酶活性检测。
实施例4重组木聚糖酶突变体和野生型的活性分析
dns法:具体方法如下:在给定的ph、温度条件下,1ml的反应体系包括100μl适当的稀释酶液,900μl底物,反应10min,加入1.5mldns终止反应,沸水煮5min。冷却后540nm测定od值。1个酶活单位(u)定义为在给定的条件下,每分钟木聚糖生成1μmol还原端所需的酶量。
实施例5重组木聚糖酶突变体和野生型的性质测定
1、重组木聚糖酶突变体和野生型的最适ph测定方法如下:
将实施例3纯化的重组木聚糖酶突变体和野生型在不同的ph下进行酶促反应以测定其最适ph。底物榉木木聚糖用不同ph的0.2mol/l柠檬酸-磷酸氢二钠缓冲液中80℃下进行木聚糖酶活力测定。结果(图3)表明,重组木聚糖酶突变体和野生型的最适反应ph大致在4.0-4.5左右,且在ph3.0-6.0范围内有相近的相对酶活力。
2、重组木聚糖酶突变体和野生型的最适温度测定方法如下:
重组木聚糖酶突变体和野生型的最适温度的测定为在0.2mol/l柠檬酸-磷酸氢二钠缓冲液各自最适ph缓冲液体系及不同温度下进行酶促反应。酶反应最适温度测定结果(图4)表明,重组木聚糖酶突变体和野生型的最适温度在80-95℃左右。
3、重组木聚糖酶突变体和野生型的动力学参数测定方法如下:
参照本实验室王坤的方法(王坤,2013),测定反应的一级反应时间。确定测定km及vmax的反应时间为5min。用不同浓度的木聚糖(10,8,5,2.5,2,1,0.75,0.5mg/ml)为底物,在同一条件(80℃、ph4.0)下测定酶活性,计算出相应的反应速度,利用grafit7软件计算km值及vmax。
计算结果如表2所示。
表2,xyl10c与所有突变体基本酶学性质汇总
- 还没有人留言评论。精彩留言会获得点赞!