葡萄早熟基因VvWRKY13在调控植物中乙烯生物合成中的应用的制作方法
本发明属于葡萄早熟有利基因挖掘和功能鉴定
技术领域:
,具体涉及葡萄早熟基因vvwrky13在调控植物中乙烯生物合成中的应用。
背景技术:
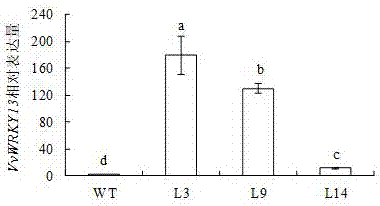
:葡萄作为一种世界广适性水果,以其独特的酿酒特性风靡全球。随着经济增长,中国葡萄酒消费呈上升趋势,葡萄种植面积随之增加。据“国际葡萄与葡萄酒组织”(internationalorganizationofvineandwine,oiv)发布的统计数据显示,2014年中国用于酿造葡萄酒的葡萄种植面积为79.9万公顷,占全球葡萄种植面积的11%,超过79.2万公顷的法国跃居世界第二。目前,葡萄产业已经成为我国农业产业化的一个重要产业。因此,探明葡萄果实生长发育的分子调控机制,对促进葡萄研究,提高我国葡萄的产量和品质,增加我国葡萄在国内和国际市场的竞争力具有极为重要的现实意义。葡萄果实的生长发育呈现出一种典型的双“s”型曲线,总体上分为3个生长阶段:第一生长周期(幼果膨大期)、停滞期(硬核期)和第二生长周期。葡萄果实的第一生长周期(幼果膨大期)从开花期持续将近60天。在这一生长周期结束时,浆果和种胚基本形成,浆果普遍呈椭圆形并生长到成熟果实体积的70%。第二个时期叫做停滞期(硬核期)。这一时期持续时间的长短与葡萄品种特性有关,无核葡萄这一时期相对短些,有核葡萄主要进行种胚的发育、种皮的硬化。此期还进行风味物质和芳香族化合物的合成、以及一些前体物质的形成。停滞期(硬核期)结束的标志是葡萄果皮的颜色开始发生变化,因此这个转折点也叫转色期。一般认为葡萄果实的停滞期在很大程度上决定了浆果成熟时的品质。之后,葡萄果实的发育进入最后一个时期——第二生长周期,浆果体积再次快速增大,含糖量急剧上升,含酸量迅速下降,单宁含量显著降低,花色苷和芳香族化合物大量合成,从而使果实呈现出特有的色、香、味,达到最佳的食用状态。目前,对果实生长发育的研究主要集中在第二生长周期即从果实转色到成熟的阶段,而对前两个时期的关注很有限。技术实现要素:本发明涉及葡萄早熟基因vvwrky13在调控植物中乙烯生物合成中的应用,同时不改变果实成熟时品质的功能和应用,可以为葡萄早熟有利基因挖掘做出贡献,从而解决缩短果实生长期的时候,难以保持果实品质的生产问题。为达到解决上述生产问题的目的,本发明采用以下技术方案予以实现:本发明提供了葡萄早熟基因vvwrky13在调控植物中乙烯生物合成中的应用。进一步的:所述植物为番茄。进一步的:将含有葡萄早熟基因vvwrky13的农杆菌菌株侵染番茄,进行共培养;将共培养后的外植体置于芽诱导培养基中进行继代培养;待外植体发芽后转入芽伸长培养基培养;之后将芽转入生根培养基中培养至获得转基因vvwrky13番茄植株。进一步的:在花后7天和转色期,乙烯在转基因vvwrky13番茄株系中的释放速率显著高于野生型番茄,vvwrky13影响番茄果实发育前期乙烯的生成量。进一步的:乙烯合成关键酶基因slacs1b、slaco1和slaco4在转基因vvwrky13番茄果实中的表达量显著升高,vvwrky13通过影响番茄果实发育过程中乙烯合成相关基因的表达来调控番茄果实中乙烯的生物合成。进一步的:转基因vvwrky13番茄植株种子具有典型的组成型三重反应表型。进一步的:转基因vvwrky13番茄植株中vvwrky13基因的相对表达量与野生型番茄相比显著增加,vvwkry13过表达促进了乙烯的过量生成。进一步的:基因vvwrky13的过表达缩短了果实发育过程中的膨大期。进一步的:基因vvwrky13过表达番茄株系果实从开花到转色比野生型早2-3天,vvwrky13缩短了果实幼果发育的时间。进一步的:基因vvwrky13过表达番茄株系果实中的番茄红素、可溶性糖和可滴定酸的含量与野生型相比均无变化。与现有技术相比,本发明的优点和有益技术效果是:1、本发明通过实验证明所述vvwrky13基因在果实的幼果膨大期大量表达,在停滞期和第二生长期表达量迅速降低;并且经转基因番茄表型性状验证只在幼果膨大期起作用,对停滞期和第二生长期几乎没有影响。2、本发明所述vvwrky13基因通过调控内源乙烯的生成量起作用,没有新物质的生成,安全可靠。3、本发明所述vvwrky13基因通过促进幼果膨大期内源乙烯的生成,缩短幼果膨大期,从而达到缩短果实发育期,促进果实早熟的效果。4、本发明所述vvwrky13基因在影响果实品质的重要阶段—停滞期和第二生长期--表达量很低,几乎不影响这两个时期果实的生长发育,从而最后对果实品质完全没有负面影响。5、完熟后果实的品质指标可溶性总糖、可滴定酸含量和番茄红素的含量均无明显变化。本发明选用浆果研究的模式作物—番茄为实验材料。通过对vvwrky13转基因番茄株系生长发育相关表型性状的研究,证明vvwrky13通过调控果实幼果膨大期乙烯的生成,缩短果实的幼果膨大期,从而使果实早熟但不改变果实的品质。附图说明图1是本发明中叶盘法转化观赏番茄品种‘micro-tom’的过程。从左至右的5幅图片分别是萌发的番茄幼苗、预培养的番茄子叶、发芽的愈伤组织、生根培养和转移至土壤中的转基因番茄植株。图2是本发明中观赏番茄品种‘micro-tom’的转基因株系中vvwrky13的相对表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图3是本发明中vvwkry13过表达番茄株系不同时期果实的乙烯生成量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图4是本发明中乙烯合成关键酶基因slacs1a在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图5是本发明中乙烯合成关键酶基因slacs1b在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图6是本发明中乙烯合成关键酶基因slacs2在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图7是本发明中乙烯合成关键酶基因slacs4在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图8是本发明中乙烯合成关键酶基因slacs6在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图9是本发明中乙烯合成关键酶基因slaco1在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图10是本发明中乙烯合成关键酶基因slaco3在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图11是本发明中乙烯合成关键酶基因slaco4在vvwkry13过表达番茄株系不同时期果实中的表达量。其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图12是本发明中vvwkry13过表达番茄株系的生长情况,其中a、b、c分别代表30天、45天和60天番茄植株的生长状况。wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图13是本发明中vvwkry13过表达番茄株系的果实,其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯合的转基因株系。图14是本发明中vvwkry13过表达番茄株系果实横切图,其中wt代表野生型番茄,l3、l9、l14分别是所筛选的3个独立的纯和的转基因株系。图15是本发明中栽培番茄品种‘ailsacraig’的vvwrky13过表达株系的pcr鉴定。s1-s3:转基因样品;w:野生型对照;o:阴性对照。图16是本发明中栽培番茄品种‘ailsacraig’的转基因株系中vvwrky13的相对表达量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图17是本发明中vvwrky13过表达番茄株系对乙烯的三重反应。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图18是本发明中acc对vvwrky13过表达番茄株系胚轴长度的影响。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图19是本发明中aoa对vvwrky13过表达番茄株系三重反应的影响。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图20是本发明中aoa对vvwrky13过表达番茄株系三重反应中胚轴长度的影响。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图21是本发明中转基因植株不同时期的果实中果胶酶基因slpg1的相对表达量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图22是本发明中转基因植株不同时期的果实中果胶酶基因slpg3的相对表达量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图23是本发明中vvwkry13过表达番茄株系完熟果实中番茄红素的含量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图24是本发明中vvwkry13过表达番茄株系完熟果实中可溶性总糖的含量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。图25是本发明中vvwkry13过表达番茄株系完熟果实中可滴定酸的含量。其中wt代表野生型番茄,l1、l2、l3分别是所筛选的3个独立的纯合的转基因株系。具体实施方式以下结合附图、附表和具体实施例对本发明的技术方案做进一步详细的说明。实施例1本发明所阐明的vvwrky13基因在乙烯生物合成中的功能和应用的证明,具体包括如下步骤:1、vvwrky13过表达番茄的转化和筛选:(1)取一定量的市售观赏番茄品种‘micro-tom’的种子,用75%的乙醇浸泡消毒30s,迅速倒掉。再用4%的次氯酸钠浸泡消毒15min,消毒后用无菌水洗涤7-8次,每次需充分洗涤,彻底去掉残留的消毒剂。种子洗涤干净后,将种子播在种子萌发培养基(1/2ms)。黑暗培养3-4天待种子发芽,发芽后移至25℃,光周期为16h/8h(光照/黑暗)的条件下培养。待长出子叶,子叶刚刚打开、未长出真叶之前的小苗用于转化。(2)将vvwrky13的cds序列(genbank登录号:jq692108)转入含35s启动子的super1300质粒。之后,将该质粒转入农杆菌菌株eha105中,并保存于-80℃的超低温冰箱中,备用。将保存于-80℃的含vvwrky13质粒的农杆菌菌株在yeb液体培养基中恢复培养和扩大培养。其中培养农杆菌所用的yeb液体培养基配方为:胰蛋白胨10g/l,酵母提取物1g/l,蔗糖5g/l,mgso4•7h2o0.5g/l,ph7.0。分装后121℃高压灭菌20min,待冷却至室温后加入50mg/l过滤灭菌的卡那霉素和利福平。(3)取步骤(1)未长出真叶的无菌子叶,剪去叶尖及叶柄部,将剩余子叶剪成3mm×3mm的大小的方块,置于无抗生素的ms固体培养基上,28℃预培养2天。(4)将步骤(2)扩摇后的菌液在5000r/min离心10min,沉淀用无菌水稀释至od600=0.5-0.6作为侵染液。将步骤(3)预培养2天后的外植体在侵染液中侵染5min。置于无抗生素的ms固体培养基上共培养,在28℃暗室共培养2天。共培养培养基成分是ms+2mg/lzt+0.2mg/liaa+100mg/l特美汀+10mg/l潮霉素。(5)将共培养2天后的外植体从暗室取出,全部置于芽诱导培养基,在光下培养7天后转入新的培养基中进行继代培养。第一次继代培养后,一般每隔2周进行下一次继代培养,直至外植体发芽完全。芽诱导培养基成分是ms+2mg/lzt+0.2mg/liaa。(6)经过芽诱导期后,待外植体发芽芽长约为2-3cm时转入芽伸长培养基中,培养3-4周。芽伸长培养基成分是ms+0.5mg/lzt+0.2mg/liaa+100mg/l特美汀+10mg/l潮霉素。(7)当芽伸长至4-5cm时,剪掉愈伤组织后将芽转入到生根培养中,培养3-4周。其中生根培养基成分是1/2ms+10mg/l潮霉素+2mg/liba。(8)将生根旺盛,生长到一定高度的小苗即时转入土盆里。叶盘法转化番茄的整个过程如图1所示。2、vvwrky13转基因番茄的筛选和纯合(1)提取土培阶段正常生长的番茄叶片dna,用基因的特异引物进行pcr鉴定。将检测得到的阳性植株进行正常培养,并标记为t1代。检测vvwrky13基因所用的pcr引物序列为:vvwrky13-fp1:5’-gctctagaatgtctactacttctcaagcc-3’;vvwrky13-rp1:5’-gctctagaatgtctactacttctcaagcc-3’;(2)为得到纯合的转基因株系,单株收获t1代种子并播种,得到t2代转基因番茄幼苗。(3)提取t2代转基因番茄叶片dna,去除产生基因分离植株,收获t2代种子。种植收获的种子,得到t3代的转基因番茄幼苗。(4)pcr检测t3代转基因番茄群体的分离情况。在t3代中不出现基因分离的株系即为纯合的转基因株系。检测vvwrky13基因所用的pcr引物序列与(1)中的一致。(5)采用qrt-pcr技术检测转基因株系中vvwrky13基因的相对表达量。检测结果如图2所示。检测vvwrky13基因表达量所用的qrt-pcr引物序列为:vvwrky13-fp:5’-ggttgccaacaatccct-3’;vvwrky13-rp:5’-gtcatctccaccgatacttc-3’;(6)收集纯合的转基因番茄株系的种子,用于后续实验。3、vvwrky13过表达番茄株系乙烯生成量、三重反应和乙烯合成基因表达量的检测(1)以野生型和vvwrky13异源过表达番茄植株果实为材料,分别选取大小均匀的果实3-4个,测定花后7天、转色期和完熟期果实的乙烯释放速率。结果如图3所示,在花后7天和转色期乙烯在vvwrky13异源过表达株系中的释放速率显著高于野生型,而在完熟期差别不明显,表明vvwrky13影响了番茄果实幼果的乙烯生成量。在乙烯合成途径中,acc合成酶与acc氧化酶为该途径的限速酶和关键酶。关键酶基因的表达决定着乙烯的生成量,以同时期的野生型和vvwrky13异源过表达番茄植株果实为材料,选取大小均匀的野生型和过表达植株的番茄果实3-4个,测定花后7d、转色期和完熟期果实的乙烯合成途径相关基因表达量。结果如图4-11所示,slacs1b、slaco1和slaco4在转基因果实中比野生型的相对表达量高;其余则无较大差异。综合分析表明,vvwrky13主要影响了番茄幼果发育过程中乙烯合成关键酶基因的表达,进而影响番茄乙烯的生成。检测slacs1b、slaco1和slaco4基因表达量所用的qrt-pcr引物序列为:slacs1b-fp:5’-tgttctgaacctggttggt-3’;slacs1b-rp:5’-taaaatcatccaatcttcga-3’;slaco1-fp:5’-gtacccgatcttgacga-3’;slaco1-rp:5’-gaagtatgatgcctcctg-3’;slaco4-fp:5’-ctgtcatatttccagcacc-3’;slaco4-rp:5’-aagttgacagtagtctccacag-3’。(2)植物对乙烯的三重反应是植物所特有的反应,可以用于乙烯的生物鉴定。被乙烯处理后的植物幼苗呈现出上下胚轴伸长生长的抑制,上胚轴的横向加粗,和顶点钩的扩大。将野生型和vvwkry13过表达番茄种子,无菌处理之后,点种在含有10μmacc和10μmol/laoa的ms培养基上,28℃正常培养12h后,进行黑暗照培养7d,观测其生长情况。结果显示:相对于野生型番茄,vvwrky13过表达番茄植株在没有外源乙烯处理下,上下胚轴生长受抑制,表现出对乙烯的组成型三重反应。在含有10μm乙烯合成抑制剂aoa的培养基上,vvwkry13过表达番茄株系和野生型番茄的三重反应差异变小。以上结果在表型上证明了vvwkry13过表达促进了乙烯的过量生成。4、vvwrky13过表达番茄株系果实各生长期长短的检测(1)观察同时播种的野生型番茄和vvwrky13过表达的番茄株系果实的发育情况。结果如图12、表1和表2所示,vvwrky13过表达株系番茄果实从开花到转色的时间大约提前2-3天,而转色到完熟时间均为10天。表明vvwrky13缩短了番茄果实幼果的发育时间。表1从开花至果实转色的时间统计株系转色时间(dpa)wt42.67a±1.15l136.33b±1.52l239.33ab±4.0l336.33b±1.52表2果实从转色至成熟的时间统计株系转色到成熟(d)wt10a±1.15l110a±1.52l210a±4.04l310a±1.52(2)多聚半乳糖醛酸酶属于果胶酶类,能降解细胞壁中的果胶使细胞壁分解,参与果实的软化,其合成酶基因为slpg。检测其合成基因slpg1和slpg3的表达量,发现其在野生型和vvwrky13过表达番茄株系中的相对表达量并无差异。vvwkry13过表达番茄株系的果实如图13和图14所示。检测slpg1和slpg3基因表达量所用的引物序列为:slpg1-fp:5’-tgaggaccaaatcggaatc-3’;slpg1-rp:5’-tgtcggactaagaaagaataacc-3’;slpg3-fp:5’-atacaacagttttcagcagttcaagt-3’;slpg3-rp:5’-ggttttccactttcccctactaa-3’。5、vvwrky13过表达番茄株系完熟果实品质的检测番茄红素属于类胡萝卜素,是一种天然的食用色素,主要集中在各种果菜和水果的红色果肉或组织当中,因其缺乏维生素原活性,对氧自由基具有粹灭作用,因此有预防癌症、养颜美容、预防心脑血管疾病,也是衡量番茄果实品质的关键指标之一。以完熟期野生型和vvwrky13过表达番茄株系果实为材料,测定完熟期果实中番茄红素的含量。结果显示番茄红素在野生型和vvwrky13过表达番茄植株果实中的含量没有发生明显变化。由此可知vvwrky13的过量表达并没有对番茄果实中的番茄红素含量产生影响。6、番茄甜度是番茄品质性状的重要组成部分,它主要取决于番茄果实中可溶性糖含量,因此可以用可溶性糖含量来评价番茄的品质。以完熟期野生型和vvwrky13过表达番茄株系的果实为材料,测定可溶性糖含量。结果显示vvwrky13转基因株系果实与野生型果实的糖含量没有发生明显改变,表明vvwrky13的过表达没有影响番茄果实中可溶性糖含量。7、果实中的酸主要是指总有效有机酸根的阴离子,在番茄中主要可滴定酸为柠檬酸,在番茄的风味中发挥着重要作用。我们以完熟期野生型和vvwrky13过表达番茄株系的果实为材料检测可滴定酸的含量。结果表明二者并无明显差异,说明vvwrky13不影响番茄中可滴定酸含量。综上表明葡萄vvwrky13通过促进幼果膨大期内源乙烯的生成,缩短幼果膨大期,从而达到缩短果实发育期,促进果实早熟的效果,但对果实品质没有负面影响。实施例2本发明所阐明的vvwrky13在乙烯生物合成中的功能和应用的证明,具体包括如下步骤:1、vvwrky13过表达番茄的转化和筛选:(1)取一定量的市售栽培番茄品种‘ailsacraig(ac)’的种子,用75%的乙醇浸泡消毒30s,迅速倒掉。再用4%的次氯酸钠浸泡消毒15min,消毒后用无菌水洗涤7-8次,每次需充分洗涤,彻底去掉残留的消毒剂。种子洗涤干净后,将种子播在种子萌发培养基(1/2ms)。黑暗培养3-4天待种子发芽,发芽后移至25℃,光周期为16h/8h(光照/黑暗)的条件下培养。待长出子叶,子叶刚刚打开、未长出真叶之前的小苗用于转化。(2)将保存于-80℃的含35s启动子质粒的eha105农杆菌菌株在yeb液体培养基中恢复培养和扩大培养。其中培养农杆菌所用的yeb液体培养基配方为:胰蛋白胨10g/l,酵母提取物1g/l,蔗糖5g/l,mgso4•7h2o0.5g/l,ph7.0。分装后121℃高压灭菌20min,待冷却至室温后加入50mg/l过滤灭菌的卡那霉素和利福平。(3)取步骤(1)未出真叶的无菌子叶,剪去叶尖及叶柄部,将剩余子叶剪成3mm×3mm的大小的方块,置于无抗生素的ms固体培养基上,28℃预培养2天。(4)将步骤(2)扩摇后的菌液在5000r/min离心10min,沉淀用无菌水稀释至od600=0.5-0.6作为侵染液。将预培养2天后的外植体在侵染液中侵染5min。置于无抗生素的ms固体培养基上共培养,在28℃暗室共培养2天。共培养培养基成分是ms+2mg/lzt+0.2mg/liaa+100mg/l特美汀+10mg/l潮霉素。(5)将共培养2天后的外植体从暗室取出,全部置于芽诱导培养基,在光下培养7天后转入新的培养基中进行继代培养。第一次继代培养后,一般每隔2周进行下一次继代培养,直至外植体发芽完全。芽诱导培养基成分是ms+2mg/lzt+0.2mg/liaa。(6)经过芽诱导期后,待外植体发芽芽长约为2-3cm时转入芽伸长培养基中,培养3-4周。芽伸长培养基成分是ms+0.5mg/lzt+0.2mg/liaa+100mg/l特美汀+10mg/l潮霉素。(7)当芽伸长至4-5cm时,剪掉愈伤组织后将芽转入到生根培养基中,培养3-4周。其中生根培养基成分是1/2ms+10mg/l潮霉素+2mg/liba。(8)将生根旺盛,生长到一定高度的小苗即时转入土盆里。2、vvwrky13转基因番茄的筛选和纯合(1)提取土培阶段正常生长的番茄叶片dna,用基因的特异引物进行pcr鉴定。结果如图15所示。将检测得到的阳性植株进行正常培养,并标记为t1代。检测vvwrky13基因所用的pcr引物序列为:vvwrky13-fp1:5’-gctctagaatgtctactacttctcaagcc-3’;vvwrky13-rp1:5’-gctctagaatgtctactacttctcaagcc-3’。(2)为得到纯合的转基因株系,单株收获t1代种子并播种,得到t2代转基因番茄幼苗。(3)提取t2代转基因番茄叶片dna,去除产生基因分离植株,收获t2代种子。种植收获的种子,得到t3代的转基因番茄幼苗。(4)pcr检测t3代转基因番茄群体的分离情况。在t3代中不出现基因分离的株系即为纯合的转基因株系。检测vvwrky13基因所用的pcr引物序列与(1)中的一致。(5)采用qrt-pcr技术检测转基因株系中vvwrky13基因的相对表达量。检测结果如图16所示。检测vvwrky13基因表达量所用的qrt-pcr引物序列为:vvwrky13-fp:5’-ggttgccaacaatccct-3’;vvwrky13-rp:5’-gtcatctccaccgatacttc-3’;(6)收集纯合的转基因番茄株系的种子,用于后续实验。3、vvwrky13过表达番茄株系乙烯生成量、三重反应和乙烯合成基因表达量的检测(1)以野生型和vvwrky13异源过表达番茄植株果实为材料,分别选取大小均匀的果实3-4个,测定花后7天、转色期和完熟期果实的乙烯释放速率。结果显示,在花后7天和转色期乙烯在vvwrky13异源过表达株系中的释放速率显著高于野生型,而在完熟期差别不明显,表明vvwrky13影响了番茄果实幼果的乙烯生成量。在乙烯合成途径中,acc合成酶与acc氧化酶为该途径的限速酶和关键酶。关键酶基因的表达决定着乙烯的生成量,以同时期的野生型和vvwrky13异源过表达番茄植株果实为材料,选取大小均匀的野生型和过表达植株的番茄果实3-4个,测定花后7d、转色期和完熟期果实的乙烯合成途径相关基因表达量。结果显示,slacs1b、slaco1和slaco4在转基因果实中比野生型的相对表达量高;其余则无较大差异。综合分析表明,vvwrky13主要影响了番茄幼果发育过程中乙烯合成关键酶基因的表达,进而影响番茄乙烯的生成。检测slacs1b、slaco1和slaco4基因表达量所用的qrt-pcr引物序列为:slacs1b-fp:5’-tgttctgaacctggttggt-3’;slacs1b-rp:5’-taaaatcatccaatcttcga-3’;slaco1-fp:5’-gtacccgatcttgacga-3’;slaco1-rp:5’-gaagtatgatgcctcctg-3’;slaco4-fp:5’-ctgtcatatttccagcacc-3’;slaco4-rp:5’-aagttgacagtagtctccacag-3’。(2)植物对乙烯的三重反应是植物所特有的反应,可以用于乙烯的生物鉴定。被乙烯处理后的植物幼苗呈现出上下胚轴伸长生长的抑制,上胚轴的横向加粗,和顶点钩的扩大。将野生型和vvwkry13过表达番茄种子,无菌处理之后,点种在含有10μmacc和10μmol/laoa的ms培养基上,28℃正常培养12h后,进行黑暗照培养7d,观测其生长情况。结果如图17-20所示:相对于野生型番茄,vvwrky13过表达番茄植株在没有外源乙烯处理下,上下胚轴生长受抑制,表现出对乙烯的组成型三重反应。在含有10μm乙烯合成抑制剂aoa的培养基上,vvwkry13过表达番茄株系和野生型番茄的三重反应差异变小。以上结果在表型上证明了vvwkry13过表达促进了乙烯的过量生成。4、vvwrky13过表达番茄株系果实各生长期长短的检测(1)观察同时播种的野生型番茄和vvwrky13过表达的番茄株系果实的发育情况。结果显示,vvwrky13过表达株系番茄果实从开花到转色的时间大约提前2-3天,而转色到完熟时间均为10天。表明vvwrky13缩短了番茄果实幼果的发育时间。(2)多聚半乳糖醛酸酶属于果胶酶类,能降解细胞壁中的果胶使细胞壁分解,参与果实的软化,其合成酶基因为slpg。检测其合成基因slpg1和slpg3的表达量。结果如图21和图22所示,其在野生型和vvwrky13过表达番茄株系中的相对表达量并无差异。检测slpg1和slpg3基因表达量所用的引物序列为:slpg1-fp:5’-tgaggaccaaatcggaatc-3’;slpg1-rp:5’-tgtcggactaagaaagaataacc-3’;slpg3-fp:5’-atacaacagttttcagcagttcaagt-3’;slpg3-rp:5’-ggttttccactttcccctactaa-3’。5、vvwrky13过表达番茄株系完熟果实品质的检测(1)番茄红素属于类胡萝卜素,是一种天然的食用色素,主要集中在各种果菜和水果的红色果肉或组织当中,因其缺乏维生素原活性,对氧自由基具有粹灭作用,因此有预防癌症、养颜美容、预防心脑血管疾病,也是衡量番茄果实品质的关键指标之一。以完熟期野生型和vvwrky13过表达番茄株系果实为材料,测定完熟期果实中番茄红素的含量。结果如图23所示,番茄红素在野生型和vvwrky13过表达番茄植株果实中的含量没有发生明显变化。由此可知vvwrky13的过量表达并没有对番茄果实中的番茄红素含量产生影响。(2)番茄甜度是番茄品质性状的重要组成部分,它主要取决于番茄果实中可溶性糖含量,因此可以用可溶性糖含量来评价番茄的品质。以完熟期野生型和vvwrky13过表达番茄株系的果实为材料,测定可溶性糖含量。结果如图24所示vvwrky13转基因株系果实与野生型果实的糖含量没有发生明显改变,表明vvwrky13的过表达没有影响番茄果实中可溶性糖含量。(3)果实中的酸主要是指总有效有机酸根的阴离子,在番茄中主要可滴定酸为柠檬酸,在番茄的风味中发挥着重要作用。我们以完熟期野生型和vvwrky13过表达番茄株系的果实为材料检测可滴定酸的含量。结果如图25所示,二者并无明显差异,说明vvwrky13不影响番茄中可滴定酸含量。综上表明葡萄vvwrky13通过促进幼果膨大期内源乙烯的生成,缩短幼果膨大期,从而达到缩短果实发育期,促进果实早熟的效果,但对果实品质没有负面影响。以上实施例仅用以说明本发明的技术方案,而非对其进行限制;尽管参照前述实施例对本发明进行了详细的说明,对于本领域的普通技术人员来说,依然可以对前述实施例所记载的技术方案进行修改,或者对其中部分技术特征进行等同替换;而这些修改或替换,并不使相应技术方案的本质脱离本发明所要求保护的技术方案的精神和范围。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 拟南芥SSCD1基因突变在调控植物体内茉莉酸合成中的应用的制作方法与工艺
- 一种基因调控网络的优化控制方法与流程
- 拟南芥WRKY家族转录因子AtWRKY44基因,蛋白编码序列及其应用的制作方法与工艺
- 针对免疫调控能力相关基因个体化配方食品及其制备方法与流程
- REVOLUTA基因在调控豆科植物小叶片数目和叶茎比中的应用的制作方法与工艺
- 玉米锌铁调控转运体ZmZIP3基因及其应用的制作方法与工艺
- 拟南芥基因SPOC1在调控植株开花期中的应用的制作方法与工艺
- 调控石榴花胚珠发育的PgAG基因及其用途的制作方法与工艺
- 光效调控基因HPE1及其应用的制作方法与工艺
- 菘蓝IiAP2/ERF049基因在调控木脂素类化合物合成中的应用的制作方法与工艺