一种提高spCEMA对植物病原菌抗性的方法及转基因烟草材料与流程
本发明涉及一种提高spcema对植物病原菌抗性的方法,还涉及spcema::erbd的植物表达载体。属于植物基因工程技术领域。
背景技术:
在农业生产中,植物病害尤其是真菌病害一直是限制农作物产量提高的重要因素。长期以来,广泛采用的防治手段主要为化学防治,但由于病原菌变异速度快,会对各种化学农药产生耐药性或抗药性,同时农药所带来的环境污染和生态平衡问题也不容忽视。因此,为保障食品安全,减少化学防治方法对环境的破坏,急需开发高效并对环境安全的植物病害防治手段。
近年来,基因工程在植物抗病的研究上已经取得了初步的进展。研究表明,在植物中表达不同来源的抗菌肽基因能显著增强植物对病原菌的抗病性(gaoetal.,2000;jaynesetal,1993;jhaetal.,2009)。但利用抗菌肽获得的转基因植物材料抗病能力较差,还不能满足生产的的需要。为获得高效的植物抗病基因,除继续在不同物种中筛选、克隆抗菌肽基因外,利用分子进化手段对抗菌肽进行定向改良,在现有基础上提高其抗菌效果是一条较为有效的途径。现有研究发现,不同的抗菌肽具有不同的抑菌机制,包括结合并破坏真菌细胞膜,穿过细胞膜进入细胞中抑制不同的靶标等。因此,与真菌细胞膜的互着是抗菌肽发挥抑菌活性的重要步骤。我们设想,如能增加抗菌肽在真菌细胞膜上的富集能力,将提高抗菌肽的抑菌效果。在真菌中,麦角甾醇是细胞膜上独特的脂类成分,能够调控真菌细胞膜的流动性和渗透性,是细胞生存的关键因子(sanglardetal.,2003)。cema是由杀菌肽ceropina的n-端8个氨基酸和经改造的蜂毒素c-末端组成的阳离子抗菌肽,我们实验室进一步在其n端添加信号肽获得了spcema(牛国清等,2002)。将spcema导入植物中,可提高植物抗病性,但提高幅度仍有限。在本发明中,以真菌细胞膜上的麦角甾醇为靶标,利用分子改良手段在现有抗菌肽spcema上添加麦角甾醇结合域erbd(luccaetal.,1998),提高对植物病原真菌的靶向性和富集能力,从而达到理想的抗菌效果。
技术实现要素:
针对现有技术存在的上述不足,本发明的目的在于提供一种提高spcema对植物病原菌抗性的方法,解决目前植物抗病基因工程中抗菌肽的抑菌效果不理想的问题,利用分子改良手段在spcema的c末端融合麦角甾醇结合域erbd,提高抗菌肽在植物病原菌细胞膜上的靶向性和富集能力,从而进一步增强转基因植物对病原菌的抗性。
本发明还提供该融合抗菌肽基因的植物表达载体及转基因烟草材料。
为实现上述目的,本发明采用如下技术方案:一种提高spcema对植物病原菌抗性的方法,以植物病原真菌细胞膜上的麦角甾醇为靶点,利用分子改良手段在spcema的c末端融合麦角甾醇结合域erbd,并获得相应的融合抗菌肽基因;如seqidno.1所示的核苷酸序列和seqidno.2所示的氨基酸序列,构建植物表达载体。
进一步,具体步骤为:由于麦角甾醇结合域erbd和spcema的分子量小,采用人工合成的方式(常州基宇生物科技有限公司)获得两端分别添加bamhi与ecori酶切位点、中间用linker序列连接的融合抗菌肽,并连接于大肠杆菌克隆载体puc57上,命名为puc57-spcema::erbd。随后,bamhi与ecori酶切puc57-spcema::erbd,回收spcema::erbd片段,并与植物表达载体进行连接,获得正确的spcema::erbd植物表达载体。
一种融合抗菌肽基因的转基因烟草材料,利用构建的上述植物表达载体,经遗传转化后导入野生型烟草中组成性表达。
相比现有技术,本发明具有如下有益效果:
1、本发明利用分子改良手段在spcema的c末端融合了麦角甾醇结合域erbd,并获得一个新的融合抗菌肽基因。与野生型抗菌肽相比,改良后的抗菌肽能更加有效的抑制植物病原真菌的萌发及生长。
2、本发明将融合抗菌肽基因导入野生型烟草中,获得相应的转基因烟草。结果显示,相比野生型spcema,获得的spcema::erbd转基因烟草对植物病原菌的抗性显著提高。表明经分子改良后的spcema::erbd在植物抗病基因工程中具有较好的应用前景。
3、随着转基因技术的发展,越来越多的抗菌肽已被应用到植物抗病基因工程中。然而,天然抗菌肽仅有限提高转基因植株对病原菌的抗性,还不能满足生产需要。因此,植物抗病基因工程的关键是获得高效、特异的抗病基因。本发明中利用植物病原菌细胞与植物细胞在结构及组成成份上存在差异,以植物病原真菌细胞膜上特有的麦角甾醇为靶点,通过定向改良手段在spcema的c末端融合麦角甾醇结合域erbd,获得相应的融合抗菌肽,提高其对植物病原真菌的靶向性和富集能力,在相同浓度下,增强抗菌肽对病原真菌的抑制效果,同时减少对非靶标细胞的危害。
附图说明
图1spcema::erbd植物表达载体的酶切验证;
图中m2000表示marker2000(bioer公司)m15表示marker15(fermentas公司);泳道1-5表示不同spcema::erbd植物表达载体转化子经限制性内切酶bamhi和ecori酶切的结果。
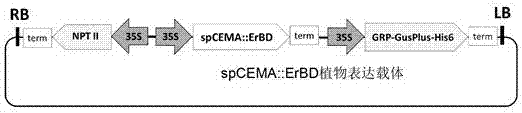
图2spcema::erbd植物表达载体图谱;
其中grp-gusplus-his6代表gusplus报告基因,该基因在n端融合了grp信号肽,在c端融合了his6序列标签;nptii代表新霉素磷酸转移酶基因,具有卡那霉素抗性;camv35s:来源于花椰菜花叶病毒的植物组成性启动子;lb:t-dna左边界;rb:t-dna右边界。
图3转基因烟草的gus染色;
transgenic代表的是转基因烟草的gus染色结果;non-transgenic代表的是非转基因烟草的gus染色结果。
图4转基因烟草表达量筛选;
横坐标表示不同转基因烟草的转化子,其中spcema::erbd-4和spcema::erbd-42代表的是spcema::erbd转基因株系,spcema-44和spcema-46代表的是spcema转基因株系,wt表示野生型烟草植株;纵坐标表示不同植株中外源基因的相对表达水平。
图5转基因烟草叶片离体抗病性比较;
a:转基因烟草叶片对大丽轮枝菌的抗性比较;b:转基因烟草叶片对烟草赤星病的抗性比较。wt表示野生型烟草叶片;spcema表示spcema转基因烟草叶片;spcema::erbd表示spcema::erbd转基因烟草叶片;water表示经水处理的野生型叶片。
图6转基因烟草叶片总蛋白提取液对油菜黑斑病原菌的抗性比较;
wt表示野生型烟草叶片总蛋白提取液;spcema表示spcema转基因烟草叶片总蛋白提取液;spcema::erbd表示spcema::erbd转基因烟草叶片总蛋白提取液;kpb表示磷酸盐缓冲液。
具体实施方式
下面结合具体实施例和附图对本发明作进一步详细说明。
本发明提供一种提高spcema对植物病原菌抗性的方法,利用分子改良手段将麦角甾醇结合域erbd,融合到spcema的c末端,其具有如seqidno.1所示的核苷酸序列和如seqidno.2所示的氨基酸序列。
【实施例1】spcema::erbd基因的获取
由于麦角甾醇结合域erbd和spcema的分子量小,采用人工合成的方式(常州基宇生物科技有限公司)获得两端分别添加bamhi与ecori酶切位点、中间用linker序列连接的融合抗菌肽,如序列1所示1-165bp为spcema序列;166-189bp为linker序列;190-240bp为erbd序列;241-243bp为终止密码子序列,如序列2所示1-55aa为spcema编码;56-63aa为linker编码;64-80为erbd编码。并连接于大肠杆菌克隆载体puc57上,命名为puc57-spcema::erbd。
【实施例2】spcema::erbd植物表达载体的构建
为验证通过分子改良后的spcema::erbd能否进一步提高植株的抗病能力,于是构建了融合抗菌肽的植物表达载体,步骤如下:
植物表达载体是由西南大学生物技术中心利用pcambia2300和pbi121自主改造而得的一个双元表达的基础载体。该载体的hptii基因用xhoi单酶切后替换为nptii基因(设计引物从pbi121载体中扩增获得,引物设计两端带xhoi位点),限制性酶切和测序结果验证了nptii基因的方向。在pbi121载体中扩增获得camv35s启动子和nos终止子,同时这两个元件两端也引入了相应的酶切位点,这两个元件最后分别导入pcambia的多克隆位点,形成camv35s::mcs::nos单元。该植物表达载体含有1套2×camv35s启动子控制nptii基因的植物表达元件、1套camv35s启动子控制报告基因grp-gusplus-his6的植物表达元件和一套camv35s启动子控制目标基因的植物表达元件,可实现kan和gus活性的双标记筛选。在多克隆位点(multiplecloningsite,mcs)插入外源基因spcema::erbd,可以实现spcema::erbd的超量表达(图2)。
烟草遗传转化用培养基
共培养培养基(ph5.4):ms无机+b5有机+30g/l蔗糖+2.0mg/l6-ba(6-苄氨基嘌呤)+0.1mg/lnaa(吲哚乙酸)+100µmas(乙酰丁香酮)+2g/lgelrite;
筛选培养基(ph5.8):ms无机+b5有机+30g/l蔗糖+2.0mg/l6-ba(6-苄氨基嘌呤)+0.1mg/lnaa(吲哚乙酸)+400mg/lcef(头孢噻肟钠)+100mg/lkm(卡那霉素)+2g/lgelrite;
生根培养基(ph5.8):ms无机+b5有机+30g/l蔗糖+200mg/lcef(头孢噻肟钠)+2g/lgelrite。
【实施例3】烟草的遗传转化
⑴转化用农杆菌浸染液的制备
分别挑取含融合抗菌肽基因的根癌农杆菌单菌落,接种于20mlyeb(5g/l蔗糖,1g/l细菌用酵母抽提物,10g/l细菌用胰化蛋白胨,0.5g/lmgso4·7h2o,ph7.0;50mg/lkm(卡那霉素),125mg/lsm(链霉素)),28℃、200rpm培养过夜,然后按5%的比例将菌液接种入20ml不含抗生素的液体yeb,28℃、200rpm培养至od600约为0.8-1.2。收集菌液,离心后弃上清,用同等体积的msb液体重悬菌体,然后将菌液全部收集到一个无菌的三角瓶,放在摇床上震荡培养1h后,即制备成农杆菌浸染液。
⑵烟草遗传转化
参考horsch等(science,asimpleandgeneralmethodfortransferringgenesintoplants,1985,227:1229-1231)的方法,以无菌苗的幼嫩叶片为外植体,利用根癌农杆菌介导法进行遗传转化。
具体操作为:烟草种子1%次氯酸钠溶液消毒10min,无菌自来水冲洗5-6次,25℃、16h光照/8h黑暗的光周期于固体萌发培养培养,待无菌苗长至5片叶时,取幼嫩叶片作为农杆菌介导遗传转化的外植体。农杆菌浸染液浸染外植体30min后倾去菌液,无菌吸水纸吸去外植体表面多余菌液,然后接种入铺有一层无菌滤纸的共培养培养基,25℃暗共培养2d。共培养完成后,将外植体接种入筛选培养基中进行分化培养,25℃、16h光照/8h黑暗的光周期培养,每2周继代一次。产生km抗性幼芽后,将幼芽切下接种入生根培养基,获得km抗性再生植株。待根长至3-5cm时,对再生植株进行gus染色确认,阳性植株移栽入温室生长成苗。
gus染色液配方
500mg/lx-gluc(x-gluc溶于dmso,浓度为12.5mg/ml),na2edta(0.01mol/l),k3fe(cn)6(0.1mol/l),k4fe(cn)6(0.1mol/l),tritonx-1001%(v/v),pbs缓冲液(0.14mol/lph7)。
【实施例4】转基因烟草的gus染色
用打孔器在再生烟草植株的幼嫩叶片上取样,于37℃恒温箱中进行gus组织化学染色。3小时后用75%酒精对染色后的样品进行脱色,并于体视镜下拍照。结果表明,已成功将目的基因转入野生型烟草中(图3)。
【实施例5】转基因烟草的表达量检测
为检测外源基因在转基因烟草中的表达情况,对温室中各转化子植株提取rna,并反转录成cdna,利用rt-pcr进行表达量检测,并基于bio-radcfxmanager和excel进行相应数据的分析。所用引物为(p1:5’-tgcttccttttcttggttc-3’;p2:5’-caacttcaaagctggcaat-3’)。rt-pcr反应体系:5.0μluniversalsybrgreensupermix(bio-rad),4.0μlcdna,0.5μlp1,0.5μlp2,总体积10μl。rt-pcr反应参数:95℃(3min);40个循环:94℃(10sec),56℃(30sec),72℃(30sec)。结果如图4所示,经rt-pcr的表达量检测分析,筛选得到了表达量较高的spcema::erbd和spcema转基因烟草株系各2个。
为了在同一水平上比较spcema:erbd与野生型抗菌肽spcema的转基因烟草抗病性差异,选取了表达量水平尽可能一致的spcema::erbd-4和spcema-44植株进行抗病鉴定分析。
【实施例6】转基因烟草叶片离体抗病性比较
首先,检测了转基因烟草叶片对大丽轮枝菌的抗性。用不带针头的1ml注射器在烟草离体叶片上创建轻微的伤口,随后移液枪吸取10µl浓度为107个孢子/ml的落叶型大丽轮枝菌的孢子悬浮液注入到伤口处,并在叶柄处包裹上湿润的吸水纸。将处理后的叶片置于塑料筐中,并覆盖上一层透明薄膜以保持湿度,于28℃恒温箱内培养15天后观察发病情况。结果如图5a所示,接种病原菌后的野生型烟草叶片发病严重,接种处的叶片呈现明显的褐变现象,并出现大量的真菌菌丝,而接种水的野生型烟草叶片未出现任何病症,排除了由机械损伤引起叶片病变的可能;野生型spcema转基因烟草有轻微的病变症状,接种处的叶片开始变褐,同时可以观察到少量的大丽轮枝菌菌丝;而相比野生型spcema,spcema::erbd转基因烟草叶片在接种处未发现明显的病变及菌丝。表明融合抗菌肽对大丽轮枝菌具有更强的抗性。
同时,也进行了对烟草赤星病菌的抗性鉴定。取温室健康生长烟草的第三片叶,利用叶片喷施法接种浓度为107个孢子/ml的烟草赤星病孢子悬浮液,并在叶柄处包裹上湿润的吸水纸。将处理后的叶片置于塑料筐中,并覆盖上一层透明薄膜以保持湿度,于28℃恒温箱内培养15天后观察发病情况。结果如图5b所示,接种后的野生型烟草叶片发病严重,已呈现明显的枯萎症状;野生型spcema转基因烟草轻微发病,其叶片的边缘开始变色、枯萎;而相比野生型spcema,spcema::erbd转基因烟草叶片上未发现明显的病斑。表明融合抗菌肽对烟草赤星病菌具有更强的抗性。
【实施例7】转基因烟草植株总蛋白提取液对病原菌的抗性比较
为了更加直观的比较spcema::erbd与野生型spcema转基因烟草的抗性差异,利用0.03mpbs缓冲液(ph5.8)提取植株总蛋白进行体外抑菌实验。bradford法测定各样品中总蛋白的含量,并利用蛋白提取液将各植物总蛋白的浓度统一到1mg/ml,随后比较它们对油菜黑斑病原菌孢子的抑菌效果。
在灭菌处理的90mm培养皿内铺上一层大小合适的滤纸,加灭菌水湿润,然后在滤纸上放上两根牙签,放上灭菌处理的载玻片。将6µl的烟草蛋白提取液(1mg/ml)、1µl油菜黑斑孢子重悬液(107个/ml)以及3µlpdb滴加在载玻片的中间,于26℃共培养10h后,进行电镜观察。结果发现(图6):油菜黑斑孢子在pbs缓冲液和野生型烟草总蛋白提取液中能够正常萌发生长;在野生型spcema转基因烟草总蛋白提取液中,绝大部分的孢子能够正常萌发,但其菌丝的生长受到了抑制;而融合抗菌肽转基因烟草总蛋白提取液能够明显抑菌孢子的萌发。上述结果表明,麦角甾醇结合域的添加能够进一步提高spcema对油菜黑斑病原菌孢子的抑菌效果。
sequencelisting
<110>西南大学
<120>一种提高spcema对植物病原菌抗性的方法及转基因烟草材料
<160>2
<170>patentinversion3.5
<210>1
<211>243
<212>dna
<213>人工序列
<220>
<223>seqidno.1所示的核苷酸序列
<400>1
atggagaagaagtctcttgctggcttgtgcttccttttcttggttctttttgttgctcaa60
gaaattgtggtgactgaagctaagtggaagttgttcaagaagatcggtatcggtgctgtt120
ttgaaggttttgactactggattgccagctttgaagttgaccaagggtggttctggtggt180
ggttctggttttaagttgagagctaagattaaggttagattgagagctaagattaagttg240
tag243
<210>2
<211>80
<212>prt
<213>人工序列
<220>
<223>seqidno.2所示的氨基酸序列
<400>2
mekkslaglcflflvlfvaqeivvteakwklfkkigigavlkvlttglpalkltkggsgg60
gsgfklrakikvrlrakikl80
- 还没有人留言评论。精彩留言会获得点赞!