小麦抗白粉病相关蛋白TaEDS1‑B1及其编码基因与应用的制作方法
本发明属于生物
技术领域:
,具体涉及一种小麦抗白粉病相关蛋白taeds1-b1及其编码基因与应用。
背景技术:
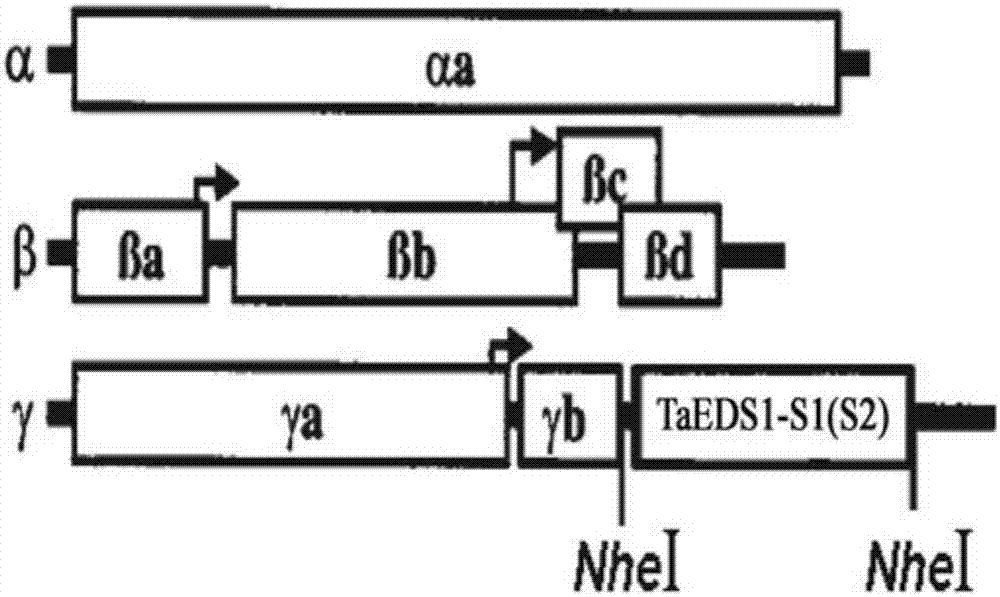
:小麦白粉病是由blumeriagraminisf.sp.tritici(bgt)引起的一种重要的真菌病害。小麦白粉病病菌可侵染小麦植株地上各个器官,但以危害叶片为主,严重时可侵染叶鞘、茎秆及穗部。白粉病主要影响叶片光合作用及植株的新陈代谢。发病较早、较重时导致植株不抽穗或抽出的穗短小,减少亩穗数、穗粒数和千粒重,一般可减产5%-10%,在严重流行年份,减产达30%以上(johnson,j.w.,baenziger,p.s.,yamazaki,w.t.,smith,r.t.effectsofpowderymildewonyieldandqualityofisogeniclinesof‘chancellor’wheat,cropscience,1979,19(3):349-352.)。白粉病是小麦最严重的病害之一,在中国及世界各小麦生产国广泛发生,这给白粉病的防治带来非常大的困难和经济成本。目前,对于小麦白粉病的防治仍以喷施化学农药为主。虽然通过喷施农药可在一定程度上减轻白粉病的危害,但是会带来生产成本增加和环境污染等问题。小麦白粉菌具有小种多、变异快、侵染时期长、气流传播、适应范围广等特点。新小种的出现常导致大批生产品种抗性的丧失。因此不断培育和推广抗病品种是减少病害损失最经济、有效和环保的方法(庄巧生,中国小麦品种改良及系谱分析。北京。中国农业出版社,2003年第一版)。抗病基因的克隆与功能鉴定对于植物抗病分子机制的研究和进行分子抗病育种具有十分重要的意义。eds1(enhanceddiseasesusceptibility1)基因最初是parker等在筛选由prr(patternrecognitionreceptor)类基因介导的霜霉病敏感突变体过程中发现的(parker,j.e.,holub,e.b.,frost,l.n.,falk,a.,gunn,n.d.,anddaniels,m.j.characterizationofeds1,amutationinarabidopsissuppressingresistancetoperonosporaparasiticaspecifiedbyseveraldifferentrppgenes.plantcell,1996,8:2033-2046.)。后来,falk等通过转座子标签法克隆了该基因并证明该基因编码一种酯酶(falk,a.,feys,b.j.,frost,l.n.,jones,j.d.,daniels,m.j.,andparker,j.e.eds1,anessentialcomponentofrgene-mediateddiseaseresistanceinarabidopsishashomologytoeukaryoticlipases.proc.natl.acad.sci.usa,1999,96:3292-3297.),并陆续证明eds1蛋白是抵御活体和半活体寄生病原菌抗性的一种正调控因子(wiermer,m.,feys,b.j.,andparker,j.e.plantimmunity:theeds1regμlatorynode.currentopinioninplantbiology,2005,8:383-389.)。除了拟南芥以外,在其它双子叶植物中也发现了eds1的同源基因,如烟草、番茄、葡萄、海岛棉等。在单子叶植物玉米、水稻和大麦等植物中也发现了eds1类基因的存在。在小麦中该基因还未被克隆,因此在小麦中克隆eds1基因,并验证其抗白粉病功能具有重要意义,可为培育小麦抗白粉病品种提供新的基因资源和抗源材料。技术实现要素:本发明的第一个目的是提供一种蛋白质。本发明提供的蛋白质,将其命名为taeds1-b1,是如下a)或b)或c)或d)的蛋白质:a)氨基酸序列是序列2所示的蛋白质;b)在序列2所示的蛋白质的n端和/或c端连接标签得到的融合蛋白质;c)将序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的蛋白质;d)与序列2所示的氨基酸序列具有75%或75%以上的同源性且具有相同功能的蛋白质。其中,序列2由620个氨基酸残基组成。为了使a)中的蛋白质便于纯化,可在序列表中序列2所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1、标签的序列标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tagii8wshpqfekc-myc10eqkliseedl上述c)中的蛋白质,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。上述c)中的蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。上述c)中的蛋白质的编码基因可通过将序列1所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。上述d)中,“同源性”包括与本发明的序列2所示的氨基酸序列具有75%或更高,或80%或更高,或85%或更高,或90%或更高,或95%或更高同源性的氨基酸序列。本发明的第二个目的是提供与taeds1-b1蛋白质相关的生物材料。本发明提供的与taeds1-b1蛋白质相关的生物材料为下述a1)至a12)中的任一种:a1)编码taeds1-b1蛋白质的核酸分子;a2)含有a1)所述核酸分子的表达盒;a3)含有a1)所述核酸分子的重组载体;a4)含有a2)所述表达盒的重组载体;a5)含有a1)所述核酸分子的重组微生物;a6)含有a2)所述表达盒的重组微生物;a7)含有a3)所述重组载体的重组微生物;a8)含有a4)所述重组载体的重组微生物;a9)含有a1)所述核酸分子的转基因植物细胞系;a10)含有a2)所述表达盒的转基因植物细胞系;a11)含有a3)所述重组载体的转基因植物细胞系;a12)含有a4)所述重组载体的转基因植物细胞系。上述生物材料中,a1)所述核酸分子为如下1)或2)或3)所示的基因:1)其编码序列是序列1所示的cdna分子或序列3所示的dna分子;2)与1)限定的核苷酸序列具有75%或75%以上同一性,且编码taeds1-b1蛋白质的cdna分子或基因组dna分子;3)在严格条件下与1)或2)限定的核苷酸序列杂交,且编码taeds1-b1蛋白质的cdna分子或基因组dna分子。其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。其中,序列1由1863个核苷酸组成,编码序列2所示的氨基酸序列。本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码taeds1-b1的核苷酸序列进行突变。那些经过人工修饰的,具有编码taeds1-b1的核苷酸序列75%或者更高同一性的核苷酸,只要编码taeds1-b1且具有相同功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列2所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。上述生物材料中,所述载体可为质粒、黏粒、噬菌体或病毒载体。上述生物材料中,所述微生物可为酵母、细菌、藻或真菌,如农杆菌。上述生物材料中,所述转基因植物细胞系、转基因植物组织和转基因植物器官均不包括繁殖材料。本发明的第三个目的是提供taeds1-b1蛋白质或上述相关生物材料的新用途。本发明提供了taeds1-b1蛋白质或上述相关生物材料在调控植物白粉病抗性中的应用。本发明还提供了taeds1-b1蛋白质或上述相关生物材料在培育白粉病抗性降低的转基因植物中的应用。本发明还提供了taeds1-b1蛋白质或上述相关生物材料在培育白粉病抗性提高的转基因植物中的应用。本发明的第四个目的是提供一种培育白粉病抗性降低的转基因植物的方法。本发明提供的培育白粉病抗性降低的转基因植物的方法包括将抑制taeds1-b1蛋白质表达的物质导入受体植物中,得到转基因植物的步骤;所述转基因植物的白粉病抗性低于所述受体植物。上述方法中,所述抑制权利要求1所述的蛋白质表达的物质为如下a)或b):a)bsmv病毒载体α、bsmv病毒载体β和含有序列1自5'端第1133-1424位所示的dna分子的bsmv病毒载体γ;b)bsmv病毒载体α、bsmv病毒载体β和含有序列1自5'端第1530-1783位所示的dna分子的bsmv病毒载体γ。上述方法中,所述转基因植物的白粉病抗性低于所述受体植物具体体现在转基因植物的白粉菌孢子堆比受体植物多。本发明的第五个目的是提供抑制taeds1-b1蛋白质表达的物质。本发明提供的抑制taeds1-b1蛋白质表达的物质为如下a)或b):a)bsmv病毒载体α、bsmv病毒载体β和含有序列1自5'端第1133-1424位所示的dna分子的bsmv病毒载体γ;b)bsmv病毒载体α、bsmv病毒载体β和含有序列1自5'端第1530-1783位所示的dna分子的bsmv病毒载体γ。本发明的第六个目的是提供一种培育白粉病抗性提高的转基因植物的方法。本发明提供的培育白粉病抗性提高的转基因植物的方法包括提高受体植物中taeds1-b1蛋白质的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的白粉病抗性高于所述受体植物。上述方法中,所述转基因植物的白粉病抗性高于所述受体植物体现在如下a1)或a2):a1)转基因植物的白粉病菌的吸器指数低于受体植物;a2)转基因植物的白粉病菌的分生孢子数低于受体植物。上述方法中,所述提高受体植物中taeds1-b1蛋白质的表达量和/或活性的方法为在受体植物中过表达taeds1-b1蛋白质;所述过表达的方法为将所述taeds1-b1蛋白质的编码基因导入受体植物;所述taeds1-b1蛋白质的编码基因的核苷酸序列是序列1所示的dna分子。在本发明的具体实施例中,所述taeds1-b1蛋白质的编码基因是通过pegad-taeds1-b1导入受体植物中,所述pegad-taeds1-b1为将序列1所示的taeds1-b1基因编码区全长cdna序列插入载体pegad的agei和xmai的酶切识别位点之间,且保持pegad载体其它序列不变的重组载体。上述方法中,所述受体植物为单子叶植物或双子叶植物。所述单子叶植物可为小麦,具体为小麦品种山疙瘩;所述双子叶植物可为拟南芥,具体为拟南芥突变体eds1。上述方法中,所述转基因植物理解为不仅包含将所述taeds1-b1基因转化目的植物得到的第一代转基因植物,也包括其子代。对于转基因植物,可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述转基因植物包括种子、愈伤组织、完整植株和细胞。本发明从小麦农家品种山疙瘩中克隆到eds1基因,将其命名为taeds1-b1基因,并构建了用于沉默小麦中taeds1-b1基因的病毒诱导性载体和过表达taeds1-b1基因的pegad-taeds1-b1载体,分别得到沉默taeds1-b1小麦和转taeds1-b1拟南芥。通过试验证明:沉默taeds1-b1小麦接种白粉病菌后,小麦叶片上出现明显的白粉病孢子堆,反应型为3-4级;转taeds1-b1小麦中过表达的taeds1-b1基因可以互补部分拟南芥eds1基因的功能,使拟南芥突变体eds1植株由感白粉病转为抗白粉病。由此证明小麦中的taeds1-b1基因具有抗白粉病的功能。附图说明图1为taeds1基因与大麦条纹花叶病毒(bsmv)重组载体的结构示意图。分别将taeds1基因的沉默片段taeds1-s1和taeds1-s2反向插入γ链的限制性内切酶nhei酶切位点,分别构建得到taeds1-b1基因的两个bsmv-vigs载体系统bsmv:taeds1-s1和bsmv:taeds1-s2,α和β链与重组载体bsmv:gf则共同构成gfp载体系统bsmv:gfp。图2为bsmv-vigs的抗白粉病普通小麦地方品种山疙瘩植株中taeds1基因的相对表达量及接种白粉病菌系e18后的叶片表型。a.taeds1基因的相对表达量,说明沉默片段s1和s2均可极显著下调taeds1基因的表达(“**”代表p≤0.01);b.接种白粉病菌系e18后的叶片表型,说明下调taeds1基因表达的山疙瘩植株失去对白粉病菌系e18的抗性,表现为感病(反应型为3~4级)。其中,mock:模拟接种的空白对照植株;bsmv:gfp:转gfp基因的负对照植株;bsmv:taeds1-s1和bsmv:taeds1-s2分别代表利用沉默片段s1和s2下调taeds1基因表达的植株。图3为载体pjit166-taeds1-b1的结构示意图。taeds1-b1基因通过限制性内切酶ncoi和xmai(与smai是同尾酶)切割与载体pjit166连接。图4为瞬时表达taeds1-b1基因的感白粉病普通小麦品种chancellor叶片表皮细胞接种白粉菌菌系e18后的吸器形成及吸器指数变化。a.共表达taeds1-b1基因和gus基因的抗病细胞,箭头指示附着在细胞表面未萌发的分生孢子;b.只表达gus基因的感病细胞,箭头指示白粉菌分生孢子萌发形成的吸器;c.瞬时表达taeds1-b1基因对吸器指数的影响。说明共表达taeds1-b1基因和gus基因细胞(gus+taeds1-b1)的吸器指数较只表达gus基因(gus)细胞的吸器指数显著降低。图5为载体pegad-taeds1-b1的结构示意图。taeds1-b1基因通过限制性内切酶agei和xmai(与smai是同尾酶)切割与载体pegad连接。图6为taeds1-b1基因在转基因拟南芥突变体eds1中的表达及其对白粉病抗性的影响。a.taeds1-b1基因在转基因拟南芥突变体eds1中的表达;b.taeds1-b1基因在拟南芥突变体eds1中表达对白粉病(大量接菌)抗性的影响,说明阳性转taeds1-b1基因拟南芥植株叶片(eds1-taeds1-b1)对白粉病的抗性较受体植株(eds1)明显增强,接近于野生型(ws);c.大量接菌条件下,taeds1-b1基因在拟南芥突变体eds1中表达对白粉病菌生长的影响,说明阳性转taeds1-b1基因拟南芥植株(eds1-taeds1-b1)叶片组织中的白粉病菌丝和分生孢子较受体(eds1)明显减少;c.拟南芥突变体eds1和阳性转taeds1-b1基因拟南芥植株(eds1-taeds1-b1)少量接种白粉菌3天后分生孢子与受体植株(eds1)及野生型(ws)的比较。说明阳性转taeds1-b1基因拟南芥植株(eds1-taeds1-b1)每菌落分生孢子数较拟南芥突变体eds1极显著降低(p≤0.01)。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径获得。下述实施例中的定量实验,均设置3次重复,结果取平均值。下述实施例中的pgem-teasyvector载体为promega公司的产品,货号为a1360。下述实施例中的bsmv-vigs病毒载体(包括α、β和γ质粒)和gfp(bsmv:gfp)质粒在文献“holzbergs.,brosiop.,grossc.,pogueg.barleystripemosaicvirus-inducedgenesilencinginamonocotplant.plantj.,2002,30:315-327.”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的白粉菌golovinomycescichoracearum(ucsc1)在文献“张海泉,马淑琴。34份粗山羊草抗白粉病性鉴定及遗传分析。植物保护,2008,34(2):37-40。”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的小麦品种chancellor(triticumaestivumvar.chancellor)在文献“翟雯雯,段霞瑜,周益林,马慧乾。我国小麦地方品种蚂蚱麦、小白冬麦、游白兰、红卷芒抗白粉病性遗传分析。植物保护,2008,1:37-40。”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的载体pjit166和pjit166-gus在文献“frederica.,carolel.t.,carstenl.,niuy.,wangd.w.,andrewj.m.virusinductionofheatshockprotein70reflectsageneralresponsetoproteinaccumulationintheplantcytosol.plantphysiology,2005,138:529-536.”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的载体pegad载体在马毅的硕士学位论文“拟南芥akin11基因的克隆、定位以及功能分析。湖南大学,2007。”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的小麦品种山疙瘩(triticumaestivumvar.shangeda)在文献“薛飞,段霞瑜,周益林,吉万全。部分小麦农家品种抗白粉病基因推导与遗传多样性分析。麦类作物学报,2009,29(2):228-235。”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的拟南芥野生型(ws型)和拟南芥突变体eds1均在文献“parkerj.e.,holube.b.,frostl.n.,falka.,gunnn.d.,anddanielsm.j.characterizationofeds1,amutationinarabidopsissuppressingresistancetoperonosporaparasiticaspecifiedbyseveraldifferentrppgenes.plantcell,1996,8:2033-2046.”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的gkpbuffer含有50mm甘氨酸(glycine)、30mmk2hpo4(ph9.2)、1%(质量百分比)膨润土(bentonite)和1%(质量百分比)硅藻土(celite)。下述实施例中的白粉病菌系e18在文献“huashi,qiujingshen,yipingqi,haojieyan,haozhennie,yongfangchen,tingzhao,fumiakikatagiri,anddingzhongtang.br-signalingkinase1physicallyassociateswithflagellinsensing2andregulatesplantinnateimmunityinarabidopsis.theplantcell,2013,25:1143–1157.”中公开过。公众可从中国科学院遗传与发育生物学研究所获得。下述实施例中的重组载体bsmv:gfp(holzbergs.,brosiop.,grossc.,pogueg.barleystripemosaicvirus-inducedgenesilencinginamonocotplant.plantj.,2002,30:315-327.),公众可从中国科学院遗传与发育生物学研究所获得。实施例一、小麦taeds1-b1基因的克隆与序列结构分析一、小麦taeds1-b1基因编码区cdna和基因组dna序列的获得1、小麦基因组dna和总rna的提取(1)小麦基因组dna的提取按照常规的ctab法提取小麦基因组dna。具体步骤如下:取约0.2g新鲜的7天龄山疙瘩幼苗叶片,在液氮中研磨成精细的粉末,转移到灭菌的2ml离心管中。加入1.0mlctab提取缓冲液(1.3%ctab,133mmtris/hclph8.0,13mmedta,0.93mnacl,0.66%pvp3600,0.18mβ-巯基乙醇),混匀。65℃温浴1.0h,其间不时混合样品,以充分提取。待样品冷却至室温,加入1ml氯仿/异戊醇(v/v=24:1),轻轻颠倒混匀1h。4℃,12000rpm离心15min,将上清液转移至一新的离心管中,加入等体积氯仿/异戊醇(v/v=24:1)重新抽提一次。吸取上清液至一新离心管,加入3μlrnasea(10mg/ml),室温下在摇床上消化45min。缓慢加入420μl异丙醇,轻轻上下摇动,直到出现白色dna沉淀。用枪头挑出dna,用1mlwashⅰ(76%乙醇、200mmnaac)溶液浸泡、洗涤15min。瞬时离心,弃上清,用washⅱ(76%乙醇,10mmnh4ac)溶液浸泡、洗涤5min。瞬时离心,弃上清,吸干残余液体,将dna沉淀室温吹干至透明。加入100μl灭菌水,待dna充分溶解后,测定dna浓度和质量。(2)小麦总rna的提取及cdna的获得采用trizol试剂按常规方法提取普通小麦农家品种山疙瘩(triticumaestivumvar.shangeda,高抗白粉病)的总rna,并利用常规反转录方法对获得的山疙瘩总rna进行反转录,获得cdna。trizol试剂购自invitrogen公司(货号:15596-018),提取材料为山疙瘩7天龄幼苗的叶片,具体的提取操作程序和条件按产品说明书进行。反转录使用takara公司反转录试剂盒(货号:rr047a),具体的操作程序和条件按试剂盒的说明书进行。2、taeds1-b1基因编码区全长cdna和基因组dna序列的获得(1)全长cdna的获得以上述步骤1中获得的cdna为模板,利用引物对taeds1-f和taeds1-r进行pcr扩增,得到pcr扩增产物。引物对taeds1-f和taeds1-r的核苷酸序列如下:taeds1-f:5'-atgccgatggacaccccgccgcc-3';taeds1-r:5'-ttacgaaggcacaagtctcgcgatg-3'。pcr反应体系:10×反转录缓冲液5.0μl,dntp4.0μl,taeds1-f(10μm)2.0μl,taeds1-r(10μm)2.0μl,cdna1.0μl,taq酶1.0μl(1.0u),最后用水补足至50.0μl。pcr反应条件:94℃1min;然后94℃30s,62℃30s,72℃2min,35个循环;最后72℃7min。将上述pcr扩增产物分离纯化后连接到pgem-tvector(promega)载体上,得到重组质粒pgem-t-taeds1-b1,并对其进行测序。测序结果表明,pcr扩增得到大小为1863bp的dna序列即为普通小麦的一个eds1基因编码区的全长cdna序列,将该基因命名为taeds1-b1。taeds1-b1的cdna序列即为序列表中的序列1。相应地将taeds1-b1基因编码的蛋白命名为taeds1-b1蛋白。taeds1-b1蛋白由620个氨基酸残基组成,其氨基酸序列即为序列表中的序列2。(2)基因组dna序列的获得以步骤1获得的基因组dna为模板,同样利用引物对taeds1-f和taeds1-r进行pcr扩增。反应体系中除模板由cdna换成基因组dna外,其它成分和量均与cdna扩增相同,反应条件亦相同。将pcr扩增产物进行回收、克隆和测序。测序结果表明,pcr扩增得到大小为3122bp的dna序列,即为普通小麦的taeds1-b1基因编码区的基因组序列,taeds1-b1基因编码区的基因组dna序列为序列表中的序列3。二、小麦taeds1-b1基因的序列结构分析比较taeds1-b1基因编码区的基因组dna序列与cdna序列,显示taeds1-b1基因由3个外显子和2个内含子组成。三、taeds1-b1基因的染色体定位在ensemblplants网站(http://plants.ensembl.org/hmmer/index.html)上比对证明taeds1-b1基因定位于小麦5al染色体臂上。实施例二、利用vigs技术验证taeds1-b1基因的抗白粉病功能一、taeds1-b1基因沉默植株的获得1、诱导taeds1-b1基因bsmv-vigs载体系统的构建(1)沉默序列的获得1)沉默序列taeds1-s1的获得以实施例1中制备的重组质粒pgem-t-taeds1-b1为模板,利用引物对taeds1-s1f和taeds1-s1r进行pcr扩增,得到大小为292bp的pcr扩增产物(对应于序列1自5’端起第1133-1424位核苷酸序列),将其命名为沉默序列taeds1-s1。上述引物对taeds1-s1-f和taeds1-s1-r的核苷酸序列如下(下划线所示序列为限制性内切酶nhei的酶切识别位点):taeds1-s1-f:5'-tacgctagcgtggccttgaactgag-3';taeds1-s1-r:5'-tacgctagctctggaagctcacgcc-3'。2)沉默序列taeds1-s2的获得以实施例一中制备的重组质粒pgem-t-taeds1-b1为模板,利用引物对taeds1-s2-f和taeds1-s2-r进行pcr扩增,得到大小为254bp(序列1自5’末端起第1530-1783位核苷酸的序列)的pcr扩增产物,将其命名为沉默序列taeds1-s2。上述引物对taeds1-s2-f和taeds1-s2-r的核苷酸序列如下(下划线所示序列为限制性内切酶nhei的酶切识别位点):taeds1-s2-f:5'-tacgctagccaccggctcctacct-3';taeds1-s2-r:5'-tacgctagcagctgctcagaaacac-3'。(2)沉默载体的构建按照常规分子生物学方法,参见“颜子颖,王海林译《精编分子生物学实验指南》,2001,科学出版社”中介绍的方法构建沉默载体。具体操作步骤如下:1)γ-taeds1-s1沉默载体的构建将上述步骤(1)获得的沉默序列taeds1-s1反向插入bsmv-vigs病毒载体γ的nhei酶切位点间,且保持bsmv-vigs病毒载体γ的其它序列不变,得到重组载体γ-taeds1-s1。以引物对taeds1-s1-f和γ-strain-p对重组载体γ-taeds1-s1进行pcr扩增和测序鉴定,阳性克隆即为将taeds1-b1基因编码区cdna序列(序列表中的序列1)自5’末端起第1133-1424位核苷酸的序列按基因表达的相反方向插入bsmv-vigs病毒载体γ链的nhei酶切位点间,且保持bsmv-vigs病毒载体γ的其它序列不变,得到的载体。γ-strain-p引物序列如下:5'-caactgccaatcgtgagtagg-3'。2)γ-taeds1-s2沉默载体的构建将上述步骤(1)获得的沉默序列taeds1-s2反向插入bsmv-vigs病毒载体γ的nhei酶切位点间,且保持bsmv-vigs病毒载体γ的其它序列不变,得到重组载体γ-taeds1-s2。以引物对taeds1-s2-f和γ-strain-p对重组载体γ-taeds1-s2进行pcr扩增和测序鉴定,阳性克隆为将序列表中的序列1自5’末端起第1530-1783位核苷酸的序列按基因表达的相反方向插入bsmv-vigs病毒载体γ的nhei酶切位点间,且保持bsmv-vigs病毒载体γ的其它序列不变得到的载体。(3)bsmv-vigs载体系统bsmv-vigs病毒载体α、β和γ-gfp载体共同构成病毒载体系统bsmv:gfp。bsmv-vigs病毒载体α、β和重组载体γ-taeds1-s1共同构成可沉默taeds1-b1基因的病毒沉默载体系统bsmv:taeds1-s1。bsmv-vigs系统载体α、β和重组载体γ-taeds1-s2共同构成可沉默taeds1-b1基因的病毒沉默载体系统bsmv:taeds1-s2。上述构建的大麦条纹花叶病毒诱导基因沉默(bsmv-vigs)的载体系统结构如图1所示,分别为载体系统中α,β和γ链的结构。2、bsmv体外转录(1)载体的线性化用mlui酶切bsmv病毒载体α链、γ-gfp载体、重组载体γ-taeds1-s1和重组载体γ-taeds1-s2,用spei酶切bsmv病毒载体β链,分别得到线性化质粒。(2)以上述步骤(1)获得的线性化质粒为模板进行体外转录,分别得到体外转录的bsmv病毒载体α、β、γ-gfp、γ-taeds1-s1和γ-taeds1-s2。体外转录反应按照ribomaxtmlargescalernaproductionsystem-t7(promega公司产品,货号:p1300)说明书进行操作。转录反应体系和条件分别为:反应总体积20.0μl,包括线性化质粒6.5μl,5×transcriptionbuffer4.0μl,cap1.5μl(promega公司产品,货号:p1718),rntppremix6.0μl,enzymemix2.0μl,37℃反应4h,转录产物置-70℃保存备用。3、bsmv接种将小麦山疙瘩播种于营养土中,待生长至二叶期,分别取8-10μlbsmv:taeds1-s1和bsmv:taeds1-s2重组病毒载体溶液涂抹接种于小麦山疙瘩平展的第二叶上,10min后用灭菌超纯水喷施叶面,覆保鲜膜保湿24h。之后转为22℃正常条件培养,分别得到转bsmv:taeds1-s1植株和bsmv:taeds1-s2植株。同时,部分植株接种bsmv:gfp重组病毒载体溶液,得到转bsmv:gfp植株,部分植株涂抹1×gkpbuffer,得到模拟接种植株。上述bsmv:taeds1-s1重组病毒载体溶液是将体外转录的bsmv-vigs载体α、β和γ-taeds1-s1经depc水稀释3倍后等量混合,再加入等体积的2×gkpbuffer得到的溶液。上述bsmv:taeds1-s2重组病毒载体溶液是将体外转录的bsmv-vigs载体α、β和γ-taeds1-s2用depc水稀释3倍后等体积混合,再加入等体积的2×gkpbuffer得到的溶液。上述bsmv:gfp重组病毒载体溶液是将体外转录的bsmv-vigs载体α、β和γ-gfp用depc水稀释3倍后等量混合,再加入等体积的2×gkpbuffer得到的溶液。上述转bsmv:taeds1-s1和bsmv:taeds1-s2的植株即为沉默taeds1-b1基因的小麦植株,转bsmv:gfp的植株即为阴性对照植株,涂抹gkpbuffer的模拟接种植株即为空白对照植株。4、沉默taeds1-b1基因小麦的rt-pcr验证bsmv-vigs系统沉默taeds1-b1基因效果的具体检测方法如下:将上述步骤3获得的转bsmv:taeds1-s1和转bsmv:taeds1-s2植株、转bsmv:gfp植株及模拟接种植株(mock)在正常条件下培养10天后,取第三叶提取总rna,反转录后通过定量pcr检测taeds1基因的相对表达量。设置actin为内参基因,相对表达量通过δδct法计算得到,由abi7500型荧光定量pcr仪自带计算公式计算。检测taeds1相对表达量的定量pcr引物对的核苷酸序列如下:rtaeds1-f:5'-ctggggacctcgtaccgc-3';rtaeds1-r:5'-gaaccatccaagtgcatcg-3';检测内参基因taactin的引物对的核苷酸序列如下:actin-f:5'-caacgagctccgtgtcgca-3';actin-r:5'-gaggaagcgtgtatccctcatag-3'。taeds1基因的相对表达量的检测结果如图2a所示。从图2a中可以看出,转bsmv:taeds1-s1和转bsmv:taeds1-s2植株中taeds1-b1基因的相对表达量较阴性对照植株(bsmv:gfp)和空白对照植株(mock)都极显著地降低了,说明本实验所选择的两个沉默序列taeds1-s1和taeds1-s2都是有效的。二、taeds1-b1基因沉默植株的白粉病抗性分析将有效沉默了taeds1-b1基因表达的植株(bsmv:taeds1-s1和bsmv:taeds1-s2)、阴性对照植株(bsmv:gfp),以及空白对照植株(mock)在第四叶展开后(约16天龄)接种白粉病菌菌系e18,接种14天后观察第四叶发病情况。反应型鉴定按zhang(zhangh,luoh,etal.geneticstudyonstriperustresistanceofawheat-thinopyronintermediumsubstitutionline.[j]triticeaecrops2003,23(1):31-33)的六级分级标准:0级:免疫,叶片无病斑;0;级:近免疫,叶片出现过敏性坏死斑,无孢子堆;1级:高抗,叶片上有少量病斑(一般直径小于1mm),菌丝层稀薄可见绿色叶面,偶见较大病斑,但仍透绿,产孢量极少;2级:中抗,叶片病斑较少(一般直径小于1mm),但菌丝层较厚,不透绿,能产生一定量的孢子;3级:中感,叶片病斑较多(一般直径大于1mm),菌丝层厚,产孢量大,但病斑不连片;4级:高感,叶片病斑多,菌丝层厚,病斑连片,产孢量大。结果如图2b所示。模拟接种的空白对照植株(mock)的叶片没有任何症状,反应型为0级(免疫)。转bsmv:gfp的阴性对照植株叶片可见明显的病毒过敏反应(hr)斑,但没有孢子产生,反应型为0;级。而taeds1基因沉默效果明显的转bsmv:taeds1-s1和转bsmv:taeds1-s2植株没有明显的hr反应,出现大量的白粉病菌孢子堆,反应型为3-4级,呈感病表型。以上结果表明taeds1-b1基因表达量的降低使得小麦山疙瘩对白粉病菌菌系e18的抗性丧失,沦为感病。由此证明taeds1-b1是一个参与小麦抗白粉病反应过程的重要基因。实施例三、利用小麦叶片表皮细胞瞬时表达技术验证taeds1-b1的功能小麦抗白粉病功能的发挥是单细胞的自主行为,并且白粉病菌只侵染寄主叶片的表皮细胞。因此,表皮细胞瞬时表达是验证小麦抗白粉病基因功能的一项有效技术。将目标基因通过基因枪随机导入到离体小麦叶片表皮细胞中,并同时共转化一种报告基因来标记阳性转化细胞,转化后高密度接种白粉菌孢子,在单细胞水平上分析阳性细胞内抗病相关基因表达对白粉菌侵染的影响。具体实施步骤如下:1、小麦叶片表皮细胞瞬时表达载体构建1)以实施例1中制备的pgem-t-taeds1-b1质粒为模板,利用taeds1-b166-ncoi-f3和taeds1-b166-xmai-r2引物对进行pcr扩增,得到大小为1889bp的扩增产物,该序列含有taeds1-b1基因编码区cdna全长序列(1863bp,如序列表中的序列1所示)。taeds1-b166-ncoi-f3和taeds1-b166-xmai-r2引物对的核苷酸序列如下(下划线所示序列分别为ncoi和xmai的酶切识别位点):taeds1-166-ncoi-f3:5'-agtccatggctatgccgatggac-3';taeds1-166-xmai-r2:5'-tcacccgggttacgaaggcacaagtc-3'。2)用ncoi和xmai对步骤1)获得的pcr扩增产物进行双酶切,得到大小为1863bp的dna片段即为taeds1-b1基因编码区cdna全长序列;用ncoi和xmai对载体pjit166进行双酶切,得到载体大片段;将1863bp的cdna序列与载体大片段连接,得到重组载体pjit166-taeds1-b1,并对其进行测序验证。正确的pjit166-taeds1-b1载体即为将完整的taeds1-b1基因编码区cdna序列(序列表中序列1)插入pjit166载体的ncoi和xmai酶切位点之间,且保持pjit166载体的其它序列不变得到的载体,pjit166-taeds1-b1载体的结构图如图3所示。2、小麦叶片的基因枪转化1)常规种植和培养感病小麦品种chancellor至7天龄,取幼苗第一片叶尖部,平铺于苯板(1%琼脂,灭菌,加入经过无菌过滤的苯并咪唑,至终浓度为85μm)中央,每板放5-7片叶子。2)称取8.5毫克金粉(1.0μm,bio-rad公司)于1.5ml的离心管中,65℃烘干4h。加入70%的乙醇水溶液,震荡5min,静置15min,离心2sec,弃上清;加入1.0ml无菌去离子水,震荡2min,静置1min,离心2sec,弃上清。重复3次;加入1.0ml50%(v/v)的甘油水溶液,震荡充分混匀。置-20℃保存备用。3)取等摩尔的pjit166-gus质粒和pjit166-taeds1-b1质粒,共≤1.0μg,补无菌去离子水至5.0μl;取50.0μl制备好的金粉(使用前震荡5min)加入混合好的质粒,震荡3min,并边震荡边逐滴加入50.0μl2.5mcacl2,再迅速加入20.0μl0.1m亚精胺,继续震荡3min,静置1min,离心2sec,弃上清;加入140.0μl70%乙醇,震荡混匀,离心2sec,弃上清;加入140.0μl100%乙醇,震荡混匀,离心2sec,弃上清;加入15.0μl100%乙醇,震荡混匀,置冰上备用。4)利用基因枪法(pds-100/hedeliverysystembio-red)转化小麦叶片。将载体盘置于载体盘holder(购自bio-rad公司)中,再次震荡混合上述金粉质粒混合物,轻轻涂于载体盘(购自bio-rad公司)上;将900psi可裂盘(购自bio-rad公司)放入螺旋头上,旋紧,滴加异丙醇密封;将终止屏放入终止屏holder(购自bio-rad公司)上,倒置放入载体盘,旋紧固定旋钮,置于基因枪内最上一层;将载有叶片的苯板放于托盘上,将托盘放在载体盘的下两层;抽真空到27inch,轰击叶片;将轰击过的叶片排列整齐,用3m透气胶带封口,放入培养箱(白天22℃,16h,晚上18℃,8h)中培养4h,得到转taeds1-b1基因的chancellor叶片。按照上述步骤1)-4)操作,分别将pjit166-gus载体和空pjit166载体共转化离体小麦叶片表皮细胞中,分别获得转pjit166-gus载体的chancellor叶片和转空pjit166载体的chancellor叶片作为对照。5)染色:培养48h后,将叶片放入15ml离心管中,倒入x-gluc染液,抽真空3次,每次5min,37℃放置24h。6)脱色:倒去染色液,尽量控干染液。加入脱色液,室温放置两天。7)染菌:将叶片放入大量水中,浸泡清洗1h;用考马斯亮蓝染色几秒钟,清水冲洗两次;将叶片置于载玻片上,用50%甘油固定,盖上盖玻片,镜检。8)统计:只统计被x-gulc染蓝的细胞,对于有孢子附着,但是没有吸器形成的细胞,记为抗病细胞;有明显吸器形成的细胞记为感病细胞。按下列公式计算吸器指数:吸器指数=感病细胞/(感病细胞+抗病细胞)×100%。3、白粉病菌接种及吸器指数统计分别对转taeds1-b1基因的chancellor叶片、转pjit166-gus载体的chancellor叶片和转空pjit166载体的chancellor叶片接种白粉病菌系e18。具体步骤如下:将上述转基因叶片在苯板上培养4h,高密度接种白粉病菌系e18的分生孢子,继续放入培养箱培养44h。将培养后的叶片放入15ml离心管中,加入x-gluc染液(0.1mna2hpo4/nah2po4,10mmna-edta,5mmk4fe[cn6],5mmk3fe[cn6],0.1%tritonx-100(v/v),20%甲醇(v/v),1g/lx-gluc,调至ph=7.0),抽真空3次,每次5min,37℃放置24h。弃干净染色液。加入脱色液(3倍储液:50%甘油,25%乳酸,25%的ddh2o,使用前加入两倍体积的100%乙醇),室温放置48h。将叶片放入水中浸泡清洗1h;用考马斯亮蓝[0.6%(w/v)考马斯亮蓝(servabluer)溶于甲醇或乙醇中]染色几秒钟,清水冲洗两次;将叶片置于载玻片上,用50%的甘油水溶液固定,加盖玻片,镜检。统计被x-gluc染成蓝色的细胞:有孢子附着,但没有吸器形成的细胞记为抗病细胞,而有明显吸器形成的细胞记为感病细胞,并按下列公式计算吸器指数:吸器指数=感病细胞/(感病细胞+抗病细胞)×100%。结果如图4所示。图4a为小麦叶片细胞瞬时表达实验获得的抗病细胞,箭头为附着在细胞表面未萌发的白粉病菌系e18的分生孢子。图4b为感病细胞,箭头为白粉病菌系e18的分生孢子萌发形成的吸器。图4c为吸器指数统计结果。由图4c可见,瞬时表达taeds1-b1基因的感病小麦品种chancellor叶片表皮细胞(gus+taeds1-b1)的吸器指数(34.26%)较阴性对照细胞(gus)的吸器指数(63.92%)显著降低。由此可见,taeds1-b1基因对白粉病菌的吸器形成具有明显的抑制作用,说明taeds1-b1基因具有提高小麦抗白粉病的功能。实施例四、利用转基因拟南芥验证taeds1-b1基因的抗白粉病功能一、转taeds1-b1基因拟南芥的获得1、表达载体的构建(1)以实施例1中的pgem-t-taeds1-b1质粒为模板,利用eds1-agei-f和eds1-xmali-r引物对进行pcr扩增,得到pcr扩增产物。引物eds1-agei-f和eds1-xmali-r的核苷酸序列如下(下划线序列分别为限制性内切酶agei和xmai的酶切识别位点):eds1-agei-f:5'-accggtatgccgatggacaccc-3';eds1-xmali-r:5'-tcacccgggttacgaaggcacaagtc-3'。上述pcr产物大小为1878bp的片段,该片段含有taeds1-b1基因编码区全长cdna序列(1863bp,如序列表中的序列1所示)。(2)用限制性内切酶agei和xmai双酶切上述pcr扩增片段(1878bp),得到酶切dna片段,即taeds1-b1基因编码区全长cdna序列(1863bp,序列表中序列1)。同时用限制性内切酶agei和xmai双酶切pegad载体(由美国加州大学林辰涛实验室赠送),得到载体大片段。将上述酶切dna片段与载体大片段连接,得到重组质粒,命名为pegad-taeds1-b1(结构图谱如图5所示)。经测序表明,pegad-taeds1-b1即为将序列1所示的taeds1-b1基因编码区全长cdna序列插入载体pegad的agei和xmai的酶切识别位点之间,且保持pegad载体其它序列不变的重组载体。2、农杆菌介导法转化拟南芥(1)农杆菌转化载体的构建采用液氮冻融法转化农杆菌gv3101株系(由biovectorco.,ltd,质粒载体菌株细胞基因保藏中心购买,货号为biovector008)。具体操作如下:取冻存的3管(eppendorf)感受态细胞系(每管200μl),融化后分别加入1μg重组质粒pegad-taeds1-b1,混匀后放置冰上20-30min,放入液氮中1min,转入37℃(2-5min)融化后,加入1.0mllb液体培养基。28℃低速振荡培养(150rpm)4-5h,10000rpm离心30sec,去上清。加入100μllb液体培养基,悬浮菌体后涂板(含50mg/ml卡那霉素,50mg/ml利福平),置28℃培养至转化子长出。挑取单菌落进行pcr检测,挑取阳性的单菌落接种于液体lb(含50mg/ml卡那霉素,50mg/ml利福平)中,28℃振荡培养过夜。提取质粒,酶切,电泳检测。取阳性克隆菌液700μl,加入等体积30%的甘油,置-70℃长期保存。(2)农杆菌介导法转化拟南芥转化受体材料为即将开花(约30天龄)的拟南芥突变体eds1。转化前,将已授粉花以及荚果去掉,并把最上面的顶生花序去掉,只留下刚刚露白的小花。取步骤(1)获得的阳性克隆于含抗生素的lb培养基中28℃活化1-2天。按1:100比例接入500mllb培养基中(kan+rip),振荡培养至od600值0.8-1.5之间。室温4000rpm离心8min,弃上清。将沉淀悬浮于相应体积的渗透培养基中,使od600值在0.8左右,即为转化液。将种植有拟南芥的方盘倒置,使花序浸泡在转化液中,3min后取出,用保鲜膜覆盖,移到光照较弱处培养,第二天揭膜。培养3-4周后,收获种子并在干燥环境中存放1-2周。(3)转化子的筛选筛选前先进行消毒,即用70%乙醇浸泡1分钟,用无菌水洗3次。消毒后的种子均匀涂布在固体筛选培养基(ms+25%basta)表面。4℃低温处理72h,移入22℃恒温培养室,在16h光照/8h黑暗周期、光强2500lux条件下培养。由于只有含外源基因的转基因种子才能在含有basta选择培养基上萌发和生长,所以一般根据种子萌发和植株生长状态即可初步判断是否为转化子。萌发后7-8天阳性苗为正常的绿色,并有较长的根系,而阴性苗则为较小的黄化苗,且基本不生根。阳性株移栽至花盆中继续生长,成熟后分单株收获种子(t1代)。收取的t1代种子分单株同样在含basta选择培养基上培养,观察分离比例,挑取统计分离比符合3:1的转基因系继续繁殖,并分单株收获t2代种子。t2代种子继续播种于含basta选择培养基上,从t3代的分离情况来推测t2代是否是纯合株,即t3代不发生分离的t2代植株即为纯合体。选取单拷贝插入的转taeds1-b1基因拟南芥纯合体家系进行后续研究。(4)转基因植株的分子鉴定以转基因植株基因组dna为模板,利用引物对taeds1-522f和taeds1-1275r进行pcr鉴定。引物对taeds1-522f和taeds1-1275r的核苷酸序列如下:taeds1-522f:5’-acagccattttgcgtgac-3’;taeds1-1275r:5’-ctcgttcagggacttgagggctt-3’。pcr反应条件为:94℃变性30sec、58℃退火30sec、72℃延伸60sec,扩增35个循环。由于小麦的taeds1-b1基因与拟南芥eds1基因同源性较低,所以只有在含有外源基因,即转taeds1-b1基因拟南芥的阳性植株中才能够扩增出相应的产物,而对照植株无任何扩增产物。二、转taeds1-b1基因拟南芥的抗白粉病鉴定1、半定量rt-pcr检测转基因拟南芥中taeds1-b1基因的表达分别以拟南芥野生型(ws)、突变体eds1和阳性转taeds1-b1基因拟南芥(eds1-taeds1-b1)的cdna为模板,利用taeds1-522f和taeds1-1275r引物对进行半定量rt-pcr扩增,检测taeds1-b1的表达量。同时以拟南芥基因actin2作为内参,内参基因的扩增引物序列为actin2-f:5’-tcggtggttccattcttgct-3’和actin2-r:5’-gctttttaagcctttgatcttgagag-3’。对扩增产物进行1%琼脂糖电泳分离和紫外分析,根据条带亮度调节pcr扩增的上样量,达到所有样品亮度均一,并与内参基因actin2一致。设20、25、30和35梯度循环数来检测pcr平台期。选择达到平台期前的合适循环数扩增基因,上样量与内参基因actin2一致,亮度的强弱即代表基因表达量的多少。rt-pcr检测结果如图6a所示,taeds1-b1基因在阳性转taeds1-b1基因拟南芥(eds1-taeds1-b1)中具有较高的表达量,而在野生型ws和突变体eds1中没有表达。2、转taeds1-b1基因拟南芥白粉病抗性的大量接菌鉴定对生长4周左右(温度为22℃,9h/15h光周期)的拟南芥野生型(ws)、突变体eds1和阳性转taeds1-b1基因拟南芥(eds1-taeds1-b1)进行白粉菌g.cichoracearum(ucsc1)的大量接种,8天后观察叶片表型。同时取部分叶片进行台盼蓝染色,观察菌丝生长情况。台盼蓝染色步骤如下:所取叶片在酒精/乳粉/台盼蓝溶液(10ml水、20ml乙醇、10ml苯酚、10ml83%乳酸、10mg台盼蓝)中煮沸2min。水洗后加入水合三氯乙醛(2.5g/ml水)室温放置4-5h。水洗,50%甘油浸泡,显微镜下观察菌丝生长情况。大量接菌鉴定结果如图6b所示。接种8天后,拟南芥野生型(ws)叶片上几乎没有白粉菌孢子出现,突变体eds1叶片上则有大量的白粉菌孢子堆出现,而转taeds1-b1基因拟南芥株系(eds1-taeds1-b1)的叶片上与野生型拟南芥相似,仅有少量的白粉菌孢子堆。同时对叶片进行台盼蓝染色后,显微镜下观察可见拟南芥野生型(ws)叶片上没有白粉菌的分生孢子产生,甚至几乎看不到菌丝生长。拟南芥突变体eds1叶片上则有大量的菌丝和分生孢子(无性繁殖的结构),而转taeds1-b1基因拟南芥株系(eds1-taeds1-b1)的叶片上仅有较少的菌丝和少量的分生孢子(图6c),与阴性对照植株(eds1)差异明显。3、转taeds1-b1拟南芥抗白粉病的微量接菌鉴定为定量描述转taeds1-b1基因拟南芥对白粉病的抗性,在大量接菌实验的基础上进一步进行了微量接菌和单菌落分生孢子数统计。选取抗性好的转taeds1-b1基因拟南芥株系与野生型(ws)和突变体eds1一同播种于恒温培养箱中(温度为22℃,9h/15h光周期),生长到28天时进行白粉菌g.cichoracearum(ucsc1)的微量接菌。5天后取叶片(每个样品取30个叶片)进行台盼蓝染色(染色方法同大量接菌鉴定),显微镜下观察并统计每个菌落的分生孢子数。统计结果如图6d所示,野生型(ws)叶片上每个菌落的平均分生孢子数为14个,突变体eds1为103个,而转taeds1-b1基因拟南芥的平均分生孢子数为28个。转taeds1-b1基因拟南芥株系每菌落的平均分生孢子数较阴性对照突变体eds1呈极显著减少。以上结果表明小麦taeds1-b1基因可以部分互补拟南芥eds1基因的功能,使拟南芥突变体eds1植株由感白粉病转为抗白粉病。由此证明小麦的taeds1-b1基因具有抗白粉病的功能,在小麦抗病育种中有利用价值。序列表<110>中国科学院遗传与发育生物学研究所<120>小麦抗白粉病相关蛋白taeds1-b1及其编码基因与应用<160>3<210>1<211>1863bp<212>dna<213>人工序列<220><223><400>1atgccgatggacaccccgccgccgcgcggggcggcgctgtccgacgacgaccgcctcctc60gtcgcccactgcgccgcgctttccttcccctccgcctcccgctccccgacctccgctgcc120tccgcctccttccaggtgcaccacgcgtcccacccctacccctgcgccgccttcgtcttc180cagcccacctggtccgccgccggctggatgccctcctcccccgacgccggcggccgcgcg240ccgtttggggacgctgaggtcgaccccgcgctcttcccgtcgctgcgcagcgtcgggagc300ggcgtccccgcgcgcgccagcgccgccttcgtcgccgccttcaggggcctgctcgacggc360tcgccgctccaaaccgaggtatctaaagcggtggcagaggagaaacgcgtcgtattcaca420ggccattcatctggaggttcaatagccacccttgctgccatatggtttcttgagaactgc480acgagacggggaagtgtcaatcaagcacagccattttgcgtgacctttggggcacctctt540gttggcgataatgttttcaaccatgctgttagaagggagggctggtcgcagtgtattttg600catttcatcatgccactggacatcatcccacgaataccgctggccccccttgaatcctct660agagaacaaattcaatctgttctggattggttgtctcctcacagtccaaacttctcacct720gttgggaattcgcttgttattcctgagttttacgaaactttgttgagaagcaccctatct780attgccagctacgaggcctgttctttcatgggatgcactagctcaatcctagggacgctg840acttcctttattgagctatccccttacagaccctgtgggacttatcttttcttgacaagc900accgaacaactgattattctcacaaactcagatgctgtattacagttgctattttactgc960cttcaattggatccccaacaacaattgcttgatgctgccgccagaagtttaagtgctcac1020tgggaatatgaatcgattaagcaaaatgtgatgcaagagatagtttgtgtggattaccta1080agagcaatttcatcatcccttcctggtagacagatgaatgggacagcaattggtggcctt1140gaactgagtaaggaagctatgctgagccttgctgctgctgcacaatgggagaagcaaaga1200gagataaaccaagcaaagatagatgcaaactgcagtaaaattcaggaagccctcaagtcc1260ttgaatgagtacaaaagaacatgcgagctgcatgaagtgagctactatgattccttcaag1320cttcagcgggaagtgcatgacttcaattcaaatgtacggaggctggaactagccggcctt1380tgggacgagataattgagatgttgcgaaggcgtgagcttccagacgcgttcgaagggcga1440gaagagtgggtgaatctgggaacctcgtaccgtcggctggttgagcccctggacattgca1500aactactacaggcattccaagaacgaggacaccggctcctacctctccaagggcaggccg1560agacgttacaagtatactcagaaatggcgcgagcagttgcaccgcatccccttgggttcc1620agcctcgagtcgtgcttctgggcaatgtccgaggagctgcaggctgagatgatcaacggc1680aaatcattcgaggatttgaaagacagggtgggtaaactggagagtgatgcacttggatgg1740ttcacctcgggaaatcttggcagggatgtgtttctgagcagctcatcctttgtgatatgg1800tggaagacactcccggagcagcacaggtccgcgtcgtgcatcgcgagacttgtgccttcg1860taa1863<210>2<211>620<212>prt<213>人工序列<220><223><400>2metprometaspthrproproproargglyalaalaleuseraspasp151015aspargleuleuvalalahiscysalaalaleuserpheproserala202530serargserprothrseralaalaseralaserpheglnvalhishis354045alaserhisprotyrprocysalaalaphevalpheglnprothrtrp505560seralaalaglytrpmetproserserproaspalaglyglyargala65707580propheglyaspalagluvalaspproalaleupheproserleuarg859095servalglyserglyvalproalaargalaseralaalaphevalala100105110alapheargglyleuleuaspglyserproleuglnthrgluvalser115120125lysalavalalagluglulysargvalvalphethrglyhisserser130135140glyglyserilealathrleualaalailetrppheleugluasncys145150155160thrargargglyservalasnglnalaglnprophecysvalthrphe165170175glyalaproleuvalglyaspasnvalpheasnhisalavalargarg180185190gluglytrpserglncysileleuhispheilemetproleuaspile195200205ileproargileproleualaproleugluserserarggluglnile210215220glnservalleuasptrpleuserprohisserproasnpheserpro225230235240valglyasnserleuvalileprogluphetyrgluthrleuleuarg245250255serthrleuserilealasertyrglualacysserphemetglycys260265270thrserserileleuglythrleuthrserpheilegluleuserpro275280285tyrargprocysglythrtyrleupheleuthrserthrgluglnleu290295300ileileleuthrasnseraspalavalleuglnleuleuphetyrcys305310315320leuglnleuaspproglnglnglnleuleuaspalaalaalaargser325330335leuseralahistrpglutyrgluserilelysglnasnvalmetgln340345350gluilevalcysvalasptyrleuargalaileserserserleupro355360365glyargglnmetasnglythralaileglyglyleugluleuserlys370375380glualametleuserleualaalaalaalaglntrpglulysglnarg385390395400gluileasnglnalalysileaspalaasncysserlysileglnglu405410415alaleulysserleuasnglutyrlysargthrcysgluleuhisglu420425430valsertyrtyraspserphelysleuglnarggluvalhisaspphe435440445asnserasnvalargargleugluleualaglyleutrpaspgluile450455460ileglumetleuargargarggluleuproaspalaphegluglyarg465470475480gluglutrpvalasnleuglythrsertyrargargleuvalglupro485490495leuaspilealaasntyrtyrarghisserlysasngluaspthrgly500505510sertyrleuserlysglyargproargargtyrlystyrthrglnlys515520525trparggluglnleuhisargileproleuglyserserleugluser530535540cysphetrpalametserglugluleuglnalaglumetileasngly545550555560lysserphegluaspleulysaspargvalglylysleugluserasp565570575alaleuglytrpphethrserglyasnleuglyargaspvalpheleu580585590serserserserphevaliletrptrplysthrleuprogluglnhis595600605argseralasercysilealaargleuvalproser 610615620<210>3<211>3122bp<212>dna<213>人工序列<220><223><400>3atgccgatggacaccccgccgccgcgcggggcggcgctgtccgacgacgaccgcctcctc60gtcgcccactgcgccgcgctttccttcccctccgcctcccgctccccgacctccgctgcc120tccgcctccttccaggtgcaccacgcgtcccacccctacccctgcgccgccttcgtcttc180cagcccacctggtccgccgccggctggatgccctcctcccccgacgccggcggccgcgcg240ccgtttggggacgctgaggtcgaccccgcgctcttcccgtcgctgcgcagcgtcgggagc300ggcgtccccgcgcgcgccagcgccgccttcgtcgccgccttcaggggcctgctcgacggc360tcgccgctccaaaccgaggtgagggccttctgccattcataaacacggcttctcggcaca420gcgttttatttttttattttttatttatttttaaataaaccgctgtgccgagaagccgtg480tttgccgcacagaagtaaaatgcaacctccgagaccttttatgctttgctatttccttcc540gcggtaggaaataggagcgtattgtttcaggaaaccttcttcctctcgtgtctagaacaa600cagcttcattccgtctgtgaaagaaatatttcaattttcaaatgctccctttgtaccaag660aaaaccttgttgctccgaattctctacaggctacaattatagtaaggacttgttgtgaat720tgttgcttcagaaattgtgtcacaccgagcaatgggatgcagtcctgtggattggagtgt780ttggagatcagtgtagtggttgtgtctagaaagggactacctctttttcttctcccatat840tgttataatacttttgaagatttatttctttggccggttttgttttcatagtgcttctac900ggttcagttagtttgatttgagcttatgaaacagaatgagggagatgatgagatccatgg960ccactatagtttcagactgatagtagtagctaaaaggctacgaggtagtttggcttggtg1020cggactaatgcccggtgaaaatatgctttattaatttgatgacaggaaaatgaactcgcc1080tccgtgagctcagcaattgtaccaagttcctctccacttacttcttcctcttgcggtggt1140tatagacctttcatagtcttacatcgtgttatgttatcttcgtatcaacagagtgatagt1200cttgtcatggtgaatttcagtgtaactgcattatcacctgaaggaaaagtggtgtttttt1260tttatcgctttgtattacaggtatctaaagcggtggcagaggagaaacgcgtcgtattca1320caggccattcatctggaggttcaatagccacccttgctgccatatggtttcttgagaact1380gcacgagacggggaagtgtcaatcaagcacagccattttgcgtgacctttggggcacctc1440ttgttggcgataatgttttcaaccatgctgttagaagggagggctggtcgcagtgtattt1500tgcatttcatcatgccactggacatcatcccacgaataccgctggccccccttgaatcct1560ctagagaacaaattcaatctgttctggattggttgtctcctcacagtccgaacttctcac1620ctgttgggaattcgcttgttattcctgagttttacgaaactttgttgagaagcaccctat1680ctattgccagctacgaggcctgttctttcatgggatgcactagctcaatcctagggacgc1740tgacttcctttattgagctatccccttacagaccctgtgggacttatcttttcttgacaa1800gcaccgaacaactgattattctcacaaactcagatgctgtattacagttgctattttact1860gccttcaattggatccccaacaacaattgcttgatgctgccgccagaagtttaagtgctc1920actgggaatatgaatcgattaagcaaaatgtgatgcaagagatagtttgtgtggattacc1980taagagcaatttcatcatcccttcctggtagacagatgaatgggacagcaattggtggcc2040ttgaactggtaagcaatggctcctcttcttccataaacaactgtggattgccactgtaaa2100tatctgttattttggtgcactgtatgtgtgaaggcagtcttagtctggaaactctcggtg2160ttgccttttagcatctaatcatttatttagggtgtagccaaatgcaacagatatgtttgt2220accttaggataaatggcaatgtacacatcaaatttgtatcagtaactgtttccgataaca2280caattctccagttactagttccatttccatctccataaaaagtgttttccctaagtttaa2340accactagtaaatttaataaccagtccaatcacataatattacagttgtcactttcatat2400tacagagtaaagaagctatgctgagccttgctgctgctgcacaatgggagaagcaaagag2460agataaaccaagcaaagatagatgcagactgcagtaaaattcaggaagccctcaagtcct2520tgaatgagtacaaaagaacatgcgagctgcatgaagtgagctactatgattccttcaagc2580ttcagcgggaagtgcatgacttcaattcaaatgtacggaggctggaactagccggccttt2640gggacgagataattgagatgttgcgaaggcgtgagcttccagacgcgttcgaagggcgag2700aagagtgggtgaatctggggacctcgtaccgtcggctggttgagcccctggacattgcaa2760actactacaggcattccaagaacgaggacaccggctcctacctctccaagggcaggccga2820gacgttacaagtatactcagaaatggcgcgagcagttgcaccgcatccccttgggttcca2880gcctcgagtcgtgcttctgggcaatgtccgaggagctgcaggctgagatgatcaacggca2940aatcattcgaggatttgaaagacagggtgggtaaactggagagtgatgcacttggatggt3000tcacctcgggaaatcttggcagggatgtgtttctgagcagctcatcctttgtgatatggt3060ggaagacactcccggagcagcacaggtccgcgtcgtgcatcgcgagacttgtgccttcgt3120aa3122当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1