一种TCR敲除的靶向NY-ESO-1的T细胞受体基因修饰T细胞及其制备方法和应用与流程
本发明涉及医学生物领域,特别涉及一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞及其制备方法和应用。
背景技术:
在众多肿瘤抗原家族中,癌-睾丸抗原(cancer-testisantigen,cta)特异性强,广泛表达于肿瘤组织。cta在肿瘤不同时期和不同分化水平的表达水平不同,可能与肿瘤的恶化、进展有关。其中,ny-eso-1(newyorkesophagealsquamouscellcarcimonma1)属于cta家族,被认为是目前免疫原性最强的肿瘤抗原之一,在多种肿瘤组织,如恶性黑素瘤、食道癌、卵巢癌等表达,在其他正常的细胞中几乎不表达而备受关注。
近些年来,免疫细胞治疗已经成为继手术、放疗、化疗之后在恶性肿瘤治疗中具有巨大前景的一种治疗方法。t细胞受体(tcellreceptor,tcr)基因修饰t细胞技术(tcr-t)为当前过继性细胞回输治疗技术最新的免疫细胞技术之一,因其能够在体内能激活自身免疫系统,持续性地靶向肿瘤细胞进行杀伤,最终达到清除恶性肿瘤细胞的目的而受到广泛的关注和研究。然而有研究表明,通过tcr-t技术转导的tcr与t细胞内源的tcr的亚基(例如α链与β链)之间存在错配,造成产生的错配的tcr的靶向识别能力丧失或识别自身抗原或者主要组织相容性复合体(majorhistocompatibilitycomplex,mhc)而引起自身免疫反应。同时,获取到对肿瘤抗原高特异的、高亲和性的tcr基因,以及tcr的亚基间的正确配对仍面临着巨大挑战。目前还未有靶向ny-eso-1的t细胞受体同时避免tcr-t技术转导的tcr与t细胞内源的tcr的亚基之间错配的和研究。
技术实现要素:
有鉴于此,本发明提供了一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞,所述tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞敲除了tcr基因,有利于防止t细胞中的内源性和外源性的tcr的α链和β链间的错配,保持高效且特异性的杀伤癌细胞的性能,并且对正常细胞不会造成损伤,避免了自身免疫反应。
第一方面,本发明提供了一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的制备方法,包括:
(1)提供cd3阳性t淋巴细胞,敲除所述cd3阳性t淋巴细胞的tcr基因,得到tcr敲除的cd3阳性t淋巴细胞;
(2)提供靶向ny-eso-1的t细胞受体tcr-ny-eso-1的编码基因,包括从5’端到3’端顺次连接的α链的编码基因、2a肽的编码基因和β链的编码基因,其中,所述α链的编码基因包括编码如seqidno:1所示的氨基酸序列的基因序列,所述β链的编码基因包括编码如seqidno:2所示的氨基酸序列的核苷酸序列,所述2a肽的编码基因包括编码如seqidno:3所示的氨基酸序列的核苷酸序列;
(3)将所述tcr-ny-eso-1的编码基因插入到plenti载体中,得到plenti-tcr-ny-eso-1重组质粒;
(4)包装所述plenti-tcr-ny-eso-1重组质粒,得到重组慢病毒;
(5)将所述重组慢病毒转染所述tcr敲除的cd3阳性t淋巴细胞,得到tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞。
可选的,步骤(1)中,所述cd3阳性t淋巴细胞是从人源外周血单个核细胞中分离获得。
可选的,所述人源外周血单个核细胞来源于自体静脉血、自体骨髓、脐带血和胎盘血等。
进一步可选的,来源于癌症患者手术一个月后、放化疗一个月后采集的新鲜外周血或骨髓。
具体的,所述cd3阳性t淋巴细胞的获得过程如下:向外周血单个核细胞中按一定比例加入cd3/cd28免疫磁珠,孵育一段时间后,放入磁铁进行筛选,得到免疫磁珠包被的cd3阳性t淋巴细胞,去除磁珠后,获得cd3阳性t淋巴细胞。
可选的,所述α链的编码基因序列包括如seqidno:4所示的核苷酸序列。
可选的,所述α链的编码基因应该考虑简并碱基,即如seqidno:1所示的氨基酸序列的编码基因包括如seqidno:4所示的核苷酸序列,保护范围还应该保护与seqidno:4具有碱基简并性质的核苷酸序列,这些核苷酸序列对应的氨基酸序列仍然为seqidno:1。
可选的,所述β链的编码基因序列包括如seqidno:5所示的核苷酸序列。
可选的,所述β链的编码基因应该考虑简并碱基,即如seqidno:2所示的氨基酸序列的编码基因包括如seqidno:5所示的核苷酸序列,保护范围还应该保护与seqidno:5具有碱基简并性质的核苷酸序列,这些核苷酸序列对应的氨基酸序列仍然为seqidno:2。
可选的,所述2a肽的编码基因序列包括如seqidno:6所示的核苷酸序列。
可选的,所述2a肽的编码基因序列应该考虑简并碱基,即如seqidno:3所示的氨基酸序列的编码基因包括如seqidno:6所示的核苷酸序列,保护范围还应该保护与seqidno:6具有碱基简并性质的核苷酸序列,这些核苷酸序列对应的氨基酸序列仍然为seqidno:3。
其中,上述“从5’端到3’端顺次连接”具体为:所述α链的编码基因序列的3’端与2a肽的编码基因序列的5’端相连,所述2a肽的编码基因序列的3’端与所述β链的5’端相连。
其中,所述2a肽为可“自我剪切”的短小肽链,所述2a肽可在蛋白翻译中会被剪切。所述2a肽具有以下优点:(1)2a肽序列短,能够有效地实现连接基因之间的共表达;(2)位于2a下游的基因同样可以获得很高的表达水平。由于传统的ires序列后基因的表达效率显著低于ires序列前基因的表达,所以与ires相比,本发明所述的2a肽更具有优势。
其中,所述tcr-ny-eso-1的编码基因插入到plenti载体中mlui和ecorⅰ酶切位点之间,且位于plenti载体的延伸因子1α(ef1α)之后,以ef1α为启动子。所述tcr-ny-eso-1的编码基因插入到plenti载体时,所述tcr-ny-eso-1的编码基因的5’端可加入起始密码子(如atg)与plenti载体中mlui酶切位点相连,3’端可加入终止密码子(如taa)与plenti载体中ecorⅰ酶切位点相连。
可选的,所述包装所述plenti-tcr-ny-eso-1重组质粒,得到重组慢病毒包括:
将所述plenti-tcr-ny-eso-1重组质粒与包膜质粒和包装质粒共同转染宿主细胞,得到所述重组慢病毒。
可选的,所述包膜质粒为pmd2g,所述包装质粒为pspax2。
所述包膜质粒pmd2g编码水疱性口炎病毒糖蛋白衣壳,所述水疱性口炎病毒糖蛋白衣壳协助重组慢病毒向细胞膜粘附,并保持重组慢病毒的感染性。
可选地,所述宿主细胞可以包括hek293t细胞、293细胞、293t细胞、293ft细胞、sw480细胞、u87mg细胞、hos细胞、cos1细胞或cos7细胞。
进一步可选的,所述宿主细胞为hek293t细胞。
本发明所述重组慢病毒可以进一步含有来自其它病毒的被膜蛋白。例如,作为这种蛋白质,最好是来自感染人类细胞的病毒被膜蛋白。对这种蛋白质没有特别的限定,可例举出逆转录病毒的兼嗜性病毒手皮膜蛋白等,例如可以使用来自小鼠白血病病毒(mumlv)4070a株的被膜蛋白。另外,也可以使用来自mumlv10al的被膜蛋白。另外,作为疱疹病毒科的蛋白,可以举出例如,单纯性疱疹病毒的gb、gd、gh、gp85蛋白,eb病毒的gp350、gp220蛋白等。作为嗜肝病毒科的蛋白,可以例举出b型肝炎病毒的s蛋白等。所述被膜蛋白还可为麻疹病毒糖蛋白与其他单链抗体融合后形成。
重组慢病毒的包装通常采用瞬时转染或采用细胞系包装。瞬时转染时可以用作包装细胞使用的人类细胞株,例如包括293细胞、293t细胞、293ft细胞、293ltv细胞、293ebna细胞及其他的从293细胞分离的克隆;sw480细胞、u87mg细胞、hos细胞、c8166细胞、mt-4细胞、molt-4细胞、hela细胞、ht1080细胞、te671细胞等。也可以采用来源于猴子的细胞株,例如,cos1细胞、cos7细胞、cv-1细胞、bmt10细胞等。而且,通常采用的磷酸钙和pei转染试剂,还有一些转染试剂如lipofectamine2000、fugene和s93fectin也被经常使用。
重组慢病毒的包装也采用一些慢病毒包装细胞系,如使用最普遍的env糖蛋白、vsvg蛋白或hiv-1gag-pol蛋白所产生的稳定细胞系。
为了安全起见,大规模使用的慢病毒载体系统都是采用分割基因组的方法,即将起不同辅助功能的基因定位于不同的质粒。目前有四质粒系统(编码gag-pol基因、rev基因、vsvg基因、sin转移基因分别位于四个不同的质粒)、三质粒系统(去掉了编码rev基因的质粒,在gag-pol质粒中gag-pol基因采用了在人细胞中偏爱性的密码子)和二质粒系统(慢病毒载体包装所必需的辅助基因位于同一个质粒上,这些辅助基因是单一的基因序列;另一个则是转基因质粒)。也有超过四质粒系统的慢病毒包装系统在使用。
可选的,所述敲除cd3阳性t淋巴细胞的tcr基因的tcr基因采用电转染和crispr/cas9技术敲除所述cd3阳性t淋巴细胞的tcr基因的tcr基因。
进一步地,所述敲除cd3阳性t淋巴细胞的tcr基因的tcr基因,包括以下步骤:
将所述cas9的编码基因插入至pcdna3.1载体中,得到pcdna3.1-cas9重组质粒,以所述pcdna3.1-cas9重组质粒为模板进行体外转录得到cas9mrna;
提供靶向tcr基因的sgrna对应的基因序列,将所述靶向tcr基因的sgrna对应的基因序列插入至pcdna3.1载体中,得到pcdna3.1-tcr-sgrna重组质粒,以所述pcdna3.1-tcr-sgrna重组质粒为模板进行体外转录得到sgrna;
将所述cas9mrna和所述靶向tcr基因的sgrna与所述cd3阳性t淋巴细胞进行混合,并置于电转仪中进行电转,完成所述cd3阳性t淋巴细胞的tcr基因的敲除。
可选的,所述靶向tcr基因的sgrna对应的基因序列包括如seqidno:7所示的核苷酸序列。
可选的,所述cas9mrna和所述tcr基因的sgrna的质量比为1:1-1:5。
进一步可选的,所述cas9mrna和所述tcr基因的sgrna的质量比为1:3。
本发明第一方面提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的制备方法,通过敲除t淋巴细胞上的tcr基因,并制备靶向ny-eso-1的t细胞受体,得到tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞。本发明所述制备方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞可以有效防止t细胞中的内源性和外源性的tcr的α链和β链间的错配,避免了自身免疫反应,保持高效且特异性的杀伤ny-eso-1阳性肿瘤细胞的性能。
第二方面,本发明提供了采用如第一方面所述的制备方法制备得到的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞,所述tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞不含tcr基因,所述tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞包括靶向ny-eso-1的t细胞受体tcr-ny-eso-1,所述靶向ny-eso-1的t细胞受体tcr-ny-eso-1包括α链和β链,所述α链包括如seqidno:1所示的氨基酸序列,所述β链包括如seqidno:2所示的氨基酸序列。
其中,所述tcr-ny-eso-1包括的α链和β链可以形成稳定的异源二聚体结构,对ny-eso-1蛋白具有很强的特异性,不会靶向正常细胞或组织,增加了其安全性并减小了脱靶效应的风险。
其中,本发明所述tcr-ny-eso-1可以特异性识别并结合ny-eso-1蛋白,且具有最佳的靶点亲和力。所述靶点亲和力是指基因改造的tcr对于靶点(例如本发明中,tcr-ny-eso-1对ny-eso-1蛋白)的结合强度。靶点亲和力对tcr的影响巨大,如果靶点亲和力的结合强度过低,则无法诱导t细胞对癌细胞的特异性杀伤;反之,如果结合强度过高,则丧失特异性,t细胞会杀伤除了肿瘤细胞外的其他低表达tcr靶点的正常细胞。
本发明所述tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞可以高效识别并杀伤包括恶性黑素瘤、食道癌、卵巢癌等表达有ny-eso-1的癌细胞。
第三方面,本发明提供了一种如第一方面所述的制备方法制备得到的或如第二方面所述的一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞在制备预防、诊断和治疗恶性肿瘤的药物中的应用。
可选的,应用于表达ny-eso-1的实体瘤的预防、诊断和治疗的药物中,例如恶性黑素瘤、食道癌、卵巢癌等。
所述应用具体为:提供了一种试剂盒,所述试剂盒包括如第一方面所述的制备方法制备得到的或如第二方面所述的一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞。
本发明的有益效果:
本发明提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞可以防止t细胞中的内源性和外源性的tcr的α链和β链间的错配,避免了自身免疫反应,保持高效且特异性的杀伤肿瘤细胞的性能,并且对正常细胞不会造成损伤。所述tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞还可以专一性的靶向ny-eso-1,促进t细胞在患者体内的扩增,能够高效且特异性的杀伤ny-eso-1阳性癌细胞,对正常的细胞不会造成损伤。
附图说明
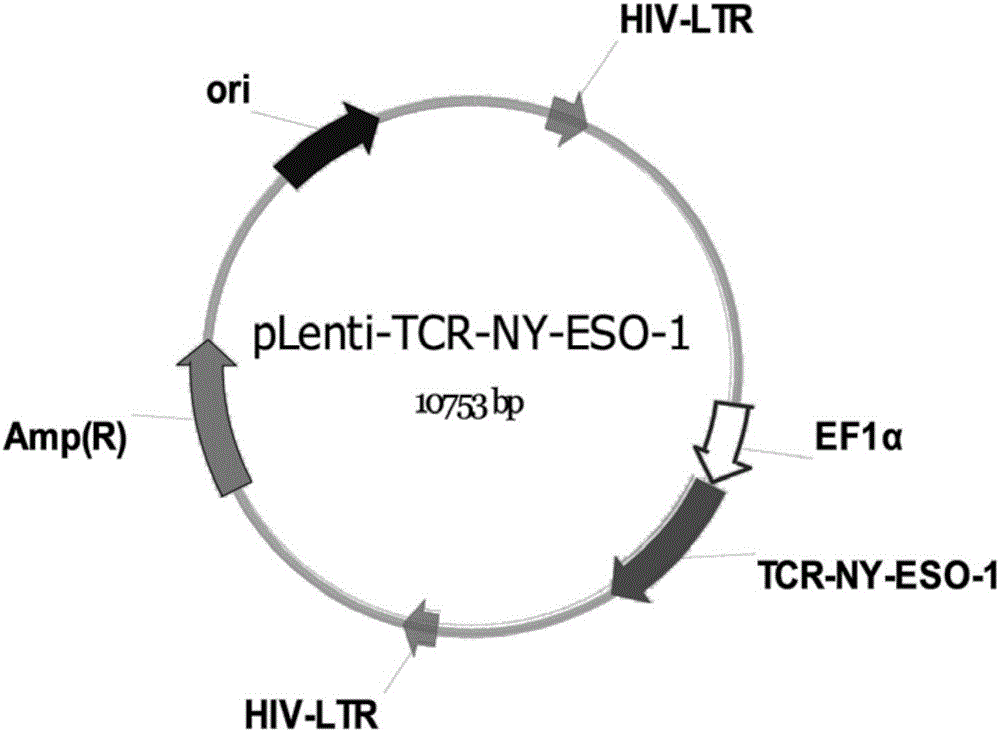
图1为本发明实施例提供的plenti-tcr-ny-eso-1重组质粒的质粒图谱。
图2为本发明实施例提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的阳性率;图2中(a)为阴性对照组,图2中(b)为本发明实施例提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的实验组。
图3为本发明实施例提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的体外肿瘤细胞杀伤效果图。
图4为本发明实施例提供的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞治疗肿瘤小鼠的效果图。
具体实施方式
以下所述是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也视为本发明的保护范围。
实施例一
一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的制备方法,包括以下步骤:
(1)tcr敲除的cd3阳性t淋巴细胞的制备
a)pbmc(外周血单个核细胞)的分离
pbmc来源于自体静脉血、自体骨髓、脐带血和胎盘血等。最好是来源于癌症患者手术一个月后、放化疗一个月后采集的新鲜外周血或骨髓。
抽取病人血液,送样至血液分离室;采集外周血单个核细胞,ficoll离心分离后取中间层细胞;经pbs洗涤后,得到pbmc。
b)免疫磁珠法分离抗原特异性t淋巴细胞
取上述pbmc,加入不含血清的基础培养基,配成细胞悬液;按磁珠与细胞的比例为3:1,加入cd3/cd28免疫磁珠,室温孵1-2h;采用磁铁对孵育好磁珠的细胞进行筛选;pbs洗涤,去除免疫磁珠后,得到cd3阳性t淋巴细胞。
c)敲除tcr基因
将所述cas9的编码基因插入至pcdna3.1载体中,得到pcdna3.1-cas9重组质粒,以所述pcdna3.1-cas9重组质粒为模板,利用mmessage
提供靶向tcr基因的sgrna对应的基因序列,其序列如seqidno:7所示;将所述靶向tcr基因的sgrna对应的基因序列插入至pcdna3.1载体中,得到pcdna3.1-tcr-sgrna重组质粒,以所述pcdna3.1-tcr-sgrna重组质粒为模板进行体外转录得到sgrna;
将所述cas9mrna和所述靶向tcr基因的sgrna序列与上述得到的cd3阳性t淋巴细胞进行混合,并置于电转仪中进行电转,敲除所述cd3阳性t淋巴细胞的tcr基因;将电转后的t细胞进行培养,得到tcr敲除的cd3阳性t淋巴细胞。
利用流式细胞仪测定上述tcr敲除的cd3阳性t淋巴细胞的制备的tcr表达量,计算敲除率,结果发现tcr敲除的cd3阳性t淋巴细胞的制备中tcr基因的敲除率达68%。
(2)制备靶向ny-eso-1的t细胞受体tcr-ny-eso-1的基因序列
分别制备α链、2a肽和β链的编码基因序列,所述α链的编码基因序列包括如seqidno:4所示的核苷酸序列,所述β链的编码基因序列包括如seqidno:5所示的核苷酸序列,所述2a肽的编码基因序列包括如seqidno:6所示的核苷酸序列。
通过pcr的方法将上述α链、2a肽和β链的编码基因序列依次从5’端到3’端连接到一起,得到靶向ny-eso-1的t细胞受体tcr-ny-eso-1的编码基因。
(3)构建plenti-tcr-ny-eso-1重组质粒
将靶向ny-eso-1的t细胞受体tcr-ny-eso-1的编码基因插入到plenti载体的mlui和ecorⅰ酶切位点之间,并在plenti载体ef1α之后,以ef1α为启动子。所述tcr-ny-eso-1的编码基因序列插入到plenti载体时,所述tcr-ny-eso-1的编码基因序列的5’端可加入起始密码子(如atg)与plenti载体中mlui酶切位点相连,3’端可加入终止密码子(如taa)与plenti载体中ecorⅰ酶切位点相连。然后转入大肠杆菌感受态细胞dh5α,进行阳性克隆pcr鉴定和测序鉴定。经过pcr产物凝胶电泳检测和测序鉴定符合目的片段大小和序列,构建如图1所示的plenti-tcr-ny-eso-1重组质粒。
(4)重组慢病毒构建
将plenti-tcr-ny-eso-1重组质粒、包装质粒pspax2、包膜质粒pmd2g三者共转染入培养好的hek293t细胞。第48h收获含病毒的上清,经0.45μm滤膜过滤,-80℃超低温冰箱中保存;第72h二次收获含病毒的上清,0.45μm滤膜过滤,与第48h收获的病毒上清合并后一起加入超速离心管中,逐一放入至beckman超速离心机内,设置离心参数为25000rpm,离心时间为2h,离心温度控制在4℃;离心结束后,弃去上清,尽量去除残留在管壁上的液体,加入病毒保存液,轻轻反复吹打重悬;经充分溶解后,高速离心10000rpm,离心5min后,取上清荧光法测定滴度,病毒按照100μl,2×108个/ml分装,保存于-80℃超低温冰箱,得到重组慢病毒。
(5)tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的制备
取上述tcr敲除的cd3阳性t淋巴细胞,加入与tcr敲除的cd3阳性t淋巴细胞相应的病毒滴度的上述重组慢病毒进行培养。培养的第3天,进行细胞计数和换液,调整细胞浓度为1×106个/ml,接种,培养;培养的第5天,观察细胞状态,如果细胞密度增大,则稀释细胞浓度为1×106个/ml,检测细胞活性,继续培养。扩增培养到第9-11天,收集细胞,得到tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞。
为了评估本发明所描述的上述方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞效果,进行如下效果实施例。
效果实施例一:评估本发明所制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的阳性率
将经过本发明方法制备tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞(实验组)与未经制备的t淋巴细胞(阴性对照组),使用流式细胞仪检测其阳性率,结果如图2所示,其中图2中(a)为阴性对照组,即未经制备的t细胞,图2中(b)为实验组,即为本发明制得的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞。由图2中(a)与(b)比较可得到,本发明所制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的阳性率为49.9%。
效果实施例二:评估tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的体外肿瘤细胞杀伤情况
将经过本发明方法制得的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞(实验组)与未经制备的t淋巴细胞(阴性对照组)的体外肿瘤杀伤效果进行比较,具体的:在体外将效应细胞(tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞或未经制备的t淋巴细胞)与靶细胞(hct116细胞)按数量比为1:10、1:3、1:1、3:1和10:1比例,在37℃,5%co2下进行共培养,在培养后的第15-18小时,收集细胞,进行流式染色,检测细胞杀伤情况,结果如图3所示。从图3中可看出,经过本发明所述的方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的肿瘤杀伤力均在15%以上,甚至可达60%,远远高于阴性对照组,这说明经本发明方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞具有强的肿瘤杀伤能力。
效果实施例三:评估tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞的小鼠体内肿瘤细胞杀伤情况
将经过本发明方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞(实验组)、未经制备的t淋巴细胞(阴性对照组)以及生理盐水(空白对照组),在小鼠肿瘤模型中,给每只小鼠尾静脉注射1×106个hct116细胞(n=9),绘制小鼠的生存曲线,结果如图4所示。从图4可以看出,经过本方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞使得小鼠在培养85天以后存活率还高于40%,远远超过阴性对照组和空白对照组。图4的结果表明,经过本方法制备的tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞能够更好的保护小鼠免于因肿瘤导致的死亡。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
序列表
<110>深圳宾德生物技术有限公司
<120>一种tcr敲除的靶向ny-eso-1的t细胞受体基因修饰t细胞及其制备方法和应用
<160>7
<170>siposequencelisting1.0
<210>1
<211>269
<212>prt
<213>人工序列(artificialsequence)
<400>1
gluthrleuleuglyvalserleuvalileleutrpleuglnleuala
151015
argvalasnserglnglnglyglugluaspproglnalaleuserile
202530
glngluglygluasnalathrmetasncyssertyrlysthrserile
354045
asnasnleuglntrptyrargglnasnserglyargglyleuvalhis
505560
leuileleuileargserasngluargglulyshisserglyargleu
65707580
argvalthrleuaspthrserlyslysserserserleuleuilethr
859095
alaserargalaalaaspthralasertyrphecysalathraspgly
100105110
alaglylysserthrpheglyaspglythrthrleuthrvallyspro
115120125
asnileglnlysproaspproalavaltyrglnleuargaspserlys
130135140
serserasplysservalcysleuphethrasppheaspserglnthr
145150155160
asnvalserglnserlysaspseraspvaltyrilethrasplysthr
165170175
valleuaspmetargsermetaspphelysserasnseralavalala
180185190
trpserasnlysseraspphealacysalaasnalapheasnasnser
195200205
ileileproalaaspthrphepheproserproglusersercysasp
210215220
vallysleuvalglulysserphegluthraspthrasnleuasnphe
225230235240
glnasnleuservalileglypheargileleuleuleulysvalala
245250255
glypheasnleuleumetthrleuargleutrpserser
260265
<210>2
<211>308
<212>prt
<213>人工序列(artificialsequence)
<400>2
metaspsertrpthrleucyscysvalserleucysileleuvalala
151015
lyshisthraspalaglyvalileglnserproarghisgluvalthr
202530
glumetglyglngluvalthrleuargcyslysproileserglyhis
354045
asptyrleuphetrptyrargglnthrmetmetargglyleugluleu
505560
leuiletyrpheasnasnasnvalproileaspaspserglymetpro
65707580
gluaspargpheseralalysmetproasnalaserpheserthrleu
859095
lysileglnprosergluproargaspseralavaltyrphecysala
100105110
serthrileglyalaglnproglnhispheglyaspglythrargleu
115120125
serileleugluaspleuasnlysvalpheproprogluvalalaval
130135140
phegluproserglualagluileserhisthrglnlysalathrleu
145150155160
valcysleualathrglyphepheproasphisvalgluleusertrp
165170175
trpvalasnglylysgluvalhisserglyvalserthraspprogln
180185190
proleulysgluglnproalaleuasnaspserargtyrcysleuser
195200205
serargleuargvalseralathrphetrpglnasnproargasnhis
210215220
pheargcysglnvalglnphetyrglyleusergluasnaspglutrp
225230235240
thrglnaspargalalysprovalthrglnilevalseralagluala
245250255
trpglyargalaaspcysglyphethrservalsertyrglnglngly
260265270
valleuseralathrileleutyrgluileleuleuglylysalathr
275280285
leutyralavalleuvalseralaleuvalleumetalametvallys
290295300
arglysaspphe
305
<210>3
<211>21
<212>prt
<213>人工序列(artificialsequence)
<400>3
glyserglygluglyargglyserleuleuthrcysglyaspvalglu
151015
gluasnproglypro
20
<210>4
<211>807
<212>dna
<213>人工序列(artificialsequence)
<400>4
gaaactctcctgggagtgtctttggtgattctatggcttcaactggctagggtgaacagt60
caacagggagaagaggatcctcaggccttgagcatccaggagggtgaaaatgccaccatg120
aactgcagttacaaaactagtataaacaatttacagtggtatagacaaaattcaggtaga180
ggccttgtccacctaattttaatacgttcaaatgaaagagagaaacacagtggaagatta240
agagtcacgcttgacacttccaagaaaagcagttccttgttgatcacggcttcccgggca300
gcagacactgcttcttacttctgtgctacggacggggcaggcaaatcaacctttggggat360
gggactacgctcactgtgaagccaaatatccagaagcctgaccctgccgtgtaccagctg420
agagactctaaatccagtgacaagtctgtctgcctattcaccgattttgattctcaaaca480
aatgtgtcacaaagtaaggattctgatgtgtatatcacagacaaaactgtgctagacatg540
aggtctatggacttcaagagcaacagtgctgtggcctggagcaacaaatctgactttgca600
tgtgcaaacgccttcaacaacagcattattccagcagacaccttcttccccagcccagaa660
agttcctgtgatgtcaagctggtcgagaaaagctttgaaacagatacgaacctaaacttt720
caaaacctgtcagtgattgggttccgaatcctcctcctgaaagtggccgggtttaatctg780
ctcatgacgctgcggctgtggtccagc807
<210>5
<211>924
<212>dna
<213>人工序列(artificialsequence)
<400>5
atggactcctggaccctctgctgtgtgtccctttgcatcctggtagcaaagcacacagat60
gctggagttatccagtcaccccggcacgaggtgacagagatgggacaagaagtgactctg120
agatgtaaaccaatttcaggacacgactaccttttctggtacagacagaccatgatgcgg180
ggactggagttgctcatttactttaacaacaacgttccgatagatgattcagggatgccc240
gaggatcgattctcagctaagatgcctaatgcatcattctccactctgaagatccagccc300
tcagaacccagggactcagctgtgtacttctgtgccagcactatcggggctcagccccag360
cattttggtgatgggactcgactctccatcctagaggacctgaacaaggtgttcccaccc420
gaggtcgctgtgtttgagccatcagaagcagagatctcccacacccaaaaggccacactg480
gtgtgcctggccacaggcttcttccctgaccacgtggagctgagctggtgggtgaatggg540
aaggaggtgcacagtggggtcagcacggacccgcagcccctcaaggagcagcccgccctc600
aatgactccagatactgcctgagcagccgcctgagggtctcggccaccttctggcagaac660
ccccgcaaccacttccgctgtcaagtccagttctacgggctctcggagaatgacgagtgg720
acccaggatagggccaaacccgtcacccagatcgtcagcgccgaggcctggggtagagca780
gactgtggctttacctcggtgtcctaccagcaaggggtcctgtctgccaccatcctctat840
gagatcctgctagggaaggccaccctgtatgctgtgctggtcagcgcccttgtgttgatg900
gccatggtcaagagaaaggatttc924
<210>6
<211>63
<212>dna
<213>人工序列(artificialsequence)
<400>6
ggcagtggagagggcagaggaagtctgctaacatgcggtgacgtcgaggagaatcctggc60
cca63
<210>7
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>7
tcaacctttggggatgggactac23
- 还没有人留言评论。精彩留言会获得点赞!
- 阻断UBE2M介导Neddylation修饰在肾细胞癌治疗中的应用的制造方法与工艺
- 无载体共组装肿瘤靶向抗癌纳米药物及其制备方法与应用与流程
- 一种用于判断靶向单克隆抗体治疗肿瘤疗效的预测分子的制造方法与工艺
- 一种靶向CD47的免疫检查点抑制剂药物组合物及其制备方法与流程
- 具有淋巴靶向功能的生物自组装纳米晶注射剂及制备方法与流程
- 用于检测肝癌的基因标志物及其用途的制造方法与工艺
- 特异性shRNA筛选及其靶向Ang‑2基因抑制肺癌细胞的验证方法与流程
- 一种多功能自动化生物细胞提取分离控制系统及采用该系统实现的细胞提取分离方法与流程
- 一种融合蛋白及光敏剂复合物及其制备方法和应用与流程
- 具有可视化和显示光斑边界功能的光动力治疗系统的制造方法与工艺
- 一种用于判断靶向单克隆抗体治疗肿瘤疗效的预测分子的制造方法与工艺
- 一种检测肺癌相关基因热点突变的试剂盒及其应用的制造方法与工艺
- 一种为肺癌提供离体个体化药物测试的方法及培养基与流程
- 一种谷胱甘肽/超声双重刺激响应纳米聚合物囊泡、其制备方法及应用与流程
- 一种多重刺激响应交联型聚合物纳米水凝胶、其制备方法及应用与流程
- 一种靶向CD47的免疫检查点抑制剂药物组合物及其制备方法与流程
- 具有淋巴靶向功能的生物自组装纳米晶注射剂及制备方法与流程
- 用于检测肝癌的基因标志物及其用途的制造方法与工艺
- 特异性shRNA筛选及其靶向Ang‑2基因抑制肺癌细胞的验证方法与流程
- 一种具有靶向功能仿病毒结构高分子囊泡及其制备与应用的制造方法与工艺