用于靶向基因转移的方法和组合物与流程
本申请根据35u.s.c.§119(e)要求2016年8月16日提交的美国临时申请no.62/375,666的权益,其全部内容通过引用并入本文。
政府支持声明
本发明是在美国国立卫生研究院授予的资金号5103757的政府支持下完成的。政府拥有本发明的某些权利。
发明领域
本发明涉及来自腺相关病毒(aav)的修饰衣壳蛋白以及包含其的病毒衣壳和病毒载体。特别地,本发明涉及修饰的aav衣壳蛋白和包含其的衣壳,其可引入病毒载体中以对目标靶组织赋予所需的转导概况。
发明的背景
从动物组织和腺病毒原液中分离的新的腺相关病毒(aav)毒株已扩展了可用于治疗性基因转移应用的aav载体组。目前正全面努力绘制这些aav分离株在动物模型中的组织趋向性。将aav载体的归巢导向选择性器官的能力对于基因治疗和其他治疗应用有用。
腺相关病毒(aav)已成为病毒基因转移的所选载体,并已在临床试验中显示出巨大的希望。重要的是通过视网膜下递送成功治疗视网膜。通过玻璃体内递送满足微创注射途径的开发,但通过该途径递送aav导致不良的转导结果。内界膜(ilm)产生分隔玻璃体和视网膜的屏障。
本发明解决了本领域对具有所需靶向特征的核酸递送载体的需求。
发明概述
本发明提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav4衣壳蛋白的腺相关病毒(aav)血清型4(aav4)载体,其中aav4衣壳蛋白包含在氨基酸残基k530处的置换和/或进一步包含在氨基酸残基s584、n585、s586和n587中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:4的氨基酸序列(aav4衣壳蛋白的氨基酸序列)。
本发明还提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav5衣壳蛋白的腺相关病毒(aav)血清型5(aav5)载体,其中aav5衣壳蛋白包含在氨基酸残基k517处的置换和/或进一步包含在氨基酸残基s575、s576、t577和t578中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:5的氨基酸序列(aav5衣壳蛋白的氨基酸序列)。
本文另外提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav7衣壳蛋白的腺相关病毒(aav)血清型7(aav7)载体,其中aav7衣壳蛋白包含在氨基酸残基k533处的置换和/或进一步包含在氨基酸残基a587、a588、n589和r590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:7的氨基酸序列(aav7衣壳蛋白的氨基酸序列)。
本发明进一步提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav8衣壳蛋白的腺相关病毒(aav)血清型8(aav8)载体,其中aav8衣壳蛋白包含在氨基酸残基k533处的置换和/或进一步包含在氨基酸残基q587、q588、n589和t590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:8的氨基酸序列(aav8衣壳蛋白的氨基酸序列)。
本文还提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav9衣壳蛋白的腺相关病毒(aav)血清型9(aav9)载体,其中aav9衣壳蛋白包含在氨基酸残基k531处的置换和/或进一步包含在氨基酸残基q587、a588、n589和t590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:9的氨基酸序列(aav9衣壳蛋白的氨基酸序列)。
此外,本发明提供了治疗受试者眼睛病症或缺陷的方法,其包括向受试者玻璃体内施用本发明的病毒载体,其中病毒载体包含编码有效治疗受试者眼睛疾病或缺陷的治疗性蛋白的核酸分子或治疗性dna。
本发明的这些和其他方面在下面所述的发明描述中更详细地解决。
附图简述
图1:注射后12周玻璃体内递送raav2载体及其hs结合缺陷型变体后的绿色荧光蛋白(gfp)荧光。眼底图像的定量显示raav2(hs结合)和raav2i8(消融的hs结合)之间的表达减少300倍。注射raav2的视网膜的免疫组织化学(ihc)主要在rgc中显示荧光,其中inl中具有较少的gfp阳性体细胞。显示图表,其中误差棒表示标准误差平均值(sem),并且通过非参数t检验检测显著性(**p<0.01)。
图2:视网膜的示意图描绘了玻璃体内递送后的raav输送。
图3:与离体人视网膜的病毒结合的qpcr分析。结果被定量为每个细胞基因组的载体基因组。raav2(hs结合)载体显示最大存在于视网膜处,其他地方发现的转基因很少。由raav2i8(消融的hs结合)递送的转基因在所有收集的组织中存在均低,但是与脉络膜和巩膜中的raav2递送转基因相比均显示出显著的增加。误差棒表示标准偏差。
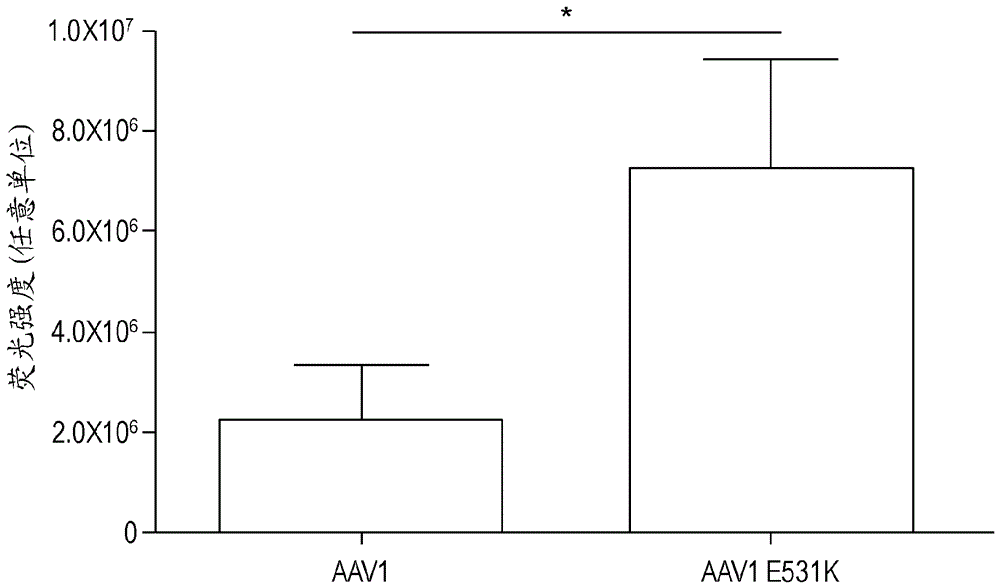
图4:注射后8周玻璃体内递送raav1的hs结合变体后的gfp荧光。眼底图像的定量显示,与raav1(非hs结合)衣壳相比,用raav1-e531k(hs结合)衣壳增加3倍。显示图表,其中误差棒表示sem和基于非参数t检验的显著性(*p<0.05)。
图5:嵌合衣壳表明趋向性受hs结合以外的其他基序的影响。使用嵌合raav2.5衣壳将raav1的元件应用于raav2,并针对玻璃体内递送成像。定量眼底荧光用于衣壳的收集。误差棒表示sem。
图6:使用可溶性肝素阻断hek293细胞的raav转导的体外竞争测定。将病毒与递增剂量的可溶性肝素一起温育,并以每个细胞10,000vg的感染复数施用于细胞培养物。raav2显示出转导的剂量依赖性降低,这是raav1或raav1-e531k所未观察到的。在所有条件下,raav1-e531k的转导量均低于raav1。误差棒显示为sem。
发明详述
现在将参考附图描述本发明,附图中显示了本发明的代表性实施方案。然而,本发明可以以不同的形式实施,并且不应被解释为限于本文阐述的实施方案。相反,提供这些实施方案,使得本公开彻底和完整,并且向本领域技术人员充分传达本发明的范围。
除非另外定义,否则本文使用的所有技术和科学术语具有与本发明所属领域的普通技术人员通常所理解的含义相同的含义。本文中对本发明的描述中使用的术语仅用于描述特定实施方案的目的,而不是意欲限制本发明。
本发明基于意外的发现,即视网膜细胞和/或视网膜色素上皮细胞的玻璃体内转导可通过在aav衣壳上添加硫酸乙酰肝素结合基序而增强。因此,在一个实施方案中,本发明提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav4衣壳蛋白的腺相关病毒(aav)血清型4(aav4)载体,其中aav4衣壳蛋白包含在氨基酸残基k530处的置换和/或进一步包含在氨基酸残基s584、n585、s586和n587中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:4的氨基酸序列(aav4衣壳蛋白的氨基酸序列)。
本发明还提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav5衣壳蛋白的腺相关病毒(aav)血清型5(aav5)载体,其中aav5衣壳蛋白包含在氨基酸残基k517处的置换和/或进一步包含在氨基酸残基s575、s576、t577和t578中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:5的氨基酸序列(aav5衣壳蛋白的氨基酸序列)。
本文另外提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav7衣壳蛋白的腺相关病毒(aav)血清型7(aav7)载体,其中aav7衣壳蛋白包含在氨基酸残基k533处的置换和/或进一步包含在氨基酸残基a587、a588、n589和r590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:7的氨基酸序列(aav7衣壳蛋白的氨基酸序列)。
本发明进一步提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav8衣壳蛋白的腺相关病毒(aav)血清型8(aav8)载体,其中aav8衣壳蛋白包含在氨基酸残基k533处的置换和/或进一步包含在氨基酸残基q587、q588、n589和t590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:8的氨基酸序列(aav8衣壳蛋白的氨基酸序列)。
本文还提供了将核酸分子引入受试者的视网膜细胞和/或视网膜色素上皮细胞中的方法,其包括玻璃体内施用包含aav9衣壳蛋白的腺相关病毒(aav)血清型9(aav9)载体,其中aav9衣壳蛋白包含在氨基酸残基k531处的置换和/或进一步包含在氨基酸残基q587、a588、n589和t590中的一个或多个处的任意组合的置换,其中残基的编号基于seqidno:9的氨基酸序列(aav9衣壳蛋白的氨基酸序列)。
此外,本发明还提供了治疗受试者眼睛病症或缺陷的方法,其包括向受试者玻璃体内施用本发明的病毒载体,其中病毒载体包含编码有效治疗受试者眼睛疾病或缺陷的治疗性蛋白的核酸分子或治疗性dna。
在进一步的实施方案中,本发明的方法可以用包含衣壳蛋白的aav载体进行,所述衣壳蛋白已经如下所述进行了修饰。具体而言,在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、490k、491t、527k、528d、529d、530e、d531e、532k、s585r、s586g、s587n、t588r)可以被绘制到aav1衣壳蛋白上和/或aav1衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:g469n、470m、s471a、472v、p474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或缺失突变(aav2编号:t265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、490k、491t、527k、528d、529d、530e、531e、532k、585r、586g、587n、588r)可以被绘制到aav2衣壳蛋白上和/或aav2衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:d469n、i470m、r471a、d472v、s474g、y500f)、具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、l488v、489s、490k、491t、527k、528d、529d、530e、531e、532k、s585r、s586g、n587n、t588r)可以被绘制到aav3b衣壳蛋白上和/或aav3b衣壳蛋白可以任何组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:s469n、470m、s471a、n472v、a474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:k484r、485q、486q、g487r、f488v、489s、490k、491t、g527k、p528d、a529d、d530e、s531e、532k、s585r、n586g、s587n、n588r)可以被绘制到aav4衣壳蛋白上和/或aav4衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:469n、f470m、s471a、n472v、k474g、s500f)、具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、t485q、g487r、w488v、n489s、l490k、g491t、l527k、q528d、g529d、s530e、n531e、t532k、s585r、s586g、t587n、t588r)可以被绘制到aav5衣壳蛋白上和/或aav5衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:r469n、y470m、471a、n472v、y474g、s500f)、具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、490k、491t、527k、528d、529d、530e、531e、r532k、a585r、a586g、587n、t588r)可以被绘制到aav7衣壳蛋白上和/或aav7衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:t469n、470m、471a、e472v、a474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、t490k、491t、527k、528d、529d、530e、531e、r532k、q585r、q586g、n587n、t588r)可以被绘制到aav8衣壳蛋白上和/或aav8衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:t469n、470m、471a、n472v、a474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或置换突变(aav2编号:s265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、t490k、491t、527k、e528d、g529d、530e、d531e、r532k、s585r、a586g、q587n、a588r)可以被绘制到aav9衣壳蛋白上和/或aav9衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:469n、470m、471a、472v、474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或置换突变(aav2编号:s265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:484r、485q、486q、487r、488v、489s、t490k、491t、527k、528d、529d、530e、531e、r532k、q585r、a586g、587n、t588r)可以被绘制到aav10衣壳蛋白上和/或aav10衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:469n、470m、s471a、a472v、a474g、500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或置换突变(aav2编号:s265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:k484r、485q、486q、487r、f488v、489s、490k、491t、g527k、p528d、s529d、d530e、g531e、d532k、n585r、a586g、t587n、t588r)可以任意组合被绘制到aav11衣壳蛋白上和/或aav11衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:d469n、f470m、471a、f472v、r474g、a500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在一些实施方案中,硫酸乙酰肝素结合基序(aav2编号:k484r、485q、486q、k487r、f488v、489s、490k、n491t、g527k、a528d、g529d、d530e、s531e、d532k、n585r、a586g、t587n、t588r)可以被绘制到aav12衣壳蛋白上和/或aav12衣壳蛋白可以任意组合包含tyr突变(aav2编号:y252f、y272f、y444f、y500f、y700f、y704f、y730f)、半乳糖基序(aav2编号:d469n、f470m、471a、f472v、r474g、a500f)具有氨基酸缩写lalgettrpa的肽的插入(aav2编号:587)、或插入突变(aav2编号:265)。氨基酸残基编号基于aav2的氨基酸序列(seqidno:2)。
在本发明的方法中,病毒载体可包含编码治疗性蛋白的核酸分子和/或治疗性dna。
本发明进一步提供治疗受试者眼睛病症或缺陷的方法,其包括向接受有效治疗受试者眼睛病症或缺陷的治疗性蛋白或治疗性dna的受试者玻璃体内施用本发明的病毒载体。
可根据本发明的方法治疗的眼睛病症或缺陷的非限制性实例包括年龄相关性黄斑变性、莱伯氏先天性黑蒙症(amarousis)1型、莱伯氏先天性黑蒙症2型、色素性视网膜炎、视网膜劈裂、失色症(achromatopsia)、色盲、先天性静止性夜盲症或其任意组合。
定义:
在本文的描述和所附权利要求中使用以下术语:
除非上下文另有明确说明,否则单数形式“一个/种(a)”、“一个/种(an)”和“该”同样意欲包括复数形式。
此外,如本文所用,术语“约”在提及诸如多核苷酸或多肽序列的长度、剂量、时间、温度等的量的可测量值时,意指涵盖特定量的±20%、±10%、±5%、±1%、±0.5%或甚至±0.1%的变化。
同样,如本文所用,“和/或”是指并且涵盖一个或多个相关所列项目的任意和所有可能的组合、以及当在替代方案(“或”)中解释时缺乏组合。
如本文所用,过渡短语“基本上由......组成”(和语法变体)意指权利要求的范围应被解释为涵盖权利要求中所述的特定材料或步骤、“以及那些不会对所要求保护的发明的基础和新颖特征产生实质性影响的材料或步骤”。因此,当在本发明的权利要求中使用时,术语“基本上由......组成”并不意欲被解释为等同于“包含”。
除非上下文另有说明,否则特别意欲本文中描述的发明的各种特征可以任意组合使用。
此外,本发明还预期在本发明的一些实施方案中,可排除或省略本文所述的任何特征或特征组合。
为了进一步说明,例如,如果说明书表明特定氨基酸可以选自a、g、i、l和/或v,则该语言还表明氨基酸可以选自这些氨基酸的任何子集,例如a、g、i或l;a、g、i或v;a或g;仅l;等,如同每个这样的子组合已在本文中明确阐述。此外,这样的语言还表明可以放弃一种或多种特定的氨基酸。例如,在特定的实施方案中,氨基酸不是a、g或i;不是a;不是g或v;等,如同每个这样的可能的放弃已在本文中明确阐述。
如本文所用,术语“减少”、“减小”、“降低”和类似术语意指减少至少约25%、35%、50%、75%、80%、85%、90%、95%、97%或更多。
如本文所用,术语“增强”、“增大”、“增长”和类似术语表示增加至少约5%、10%、20%、25%、50%、75%、100%、150%、200%、300%、400%、500%或更多。这些术语也可用于提及倍数增加,例如,一倍、两倍、三倍、四倍、五倍、六倍、七倍、八倍、九倍、十倍等。
如本文所用,术语“细小病毒”涵盖细小病毒科,包括自主复制的细小病毒和依赖病毒。自主细小病毒包括细小病毒属、红病毒属、浓核病毒属、重复病毒属和康特拉病毒属(contravirus)的成员。示例性自主细小病毒包括但不限于小鼠的微小病毒、牛细小病毒、犬细小病毒、鸡细小病毒、猫泛白细胞减少症病毒、猫细小病毒、鹅细小病毒、h1细小病毒、番鸭细小病毒、b19病毒以及现在已知或以后发现的任何其他自主细小病毒。其他自主细小病毒是本领域技术人员已知的。参见例如,bernardn.fields等人,virology,第2卷,第69章(第4版,lippincott-ravenpublishers)。
如本文所用,术语“腺相关病毒”(aav)包括但不限于aav1型、aav2型、aav3型(包括3a和3b型)、aav4型、aav5型、aav6型、aav7型、aav8型、aav9型、aav10型、aav11型、aav12型、禽aav、牛aav、犬aav、马aav、绵羊aav以及现在已知或以后发现的任何其他aav。参见例如,bernardn.fields等人,virology,第2卷,第69章(第4版,lippincott-ravenpublishers)。已经鉴定了许多相对新的aav血清型和进化枝(参见例如,gao等人(2004)j.virology78:6381-6388;moris等人(2004)virology33-:375-383;和表1)。
各种血清型的aav和自主细小病毒的基因组序列、以及天然末端重复序列(tr)、rep蛋白和衣壳亚基的序列是本领域已知的。这样的序列可以在文献或诸如genbank®database的公共数据库中找到。参见例如,genbank登录号nc_044927,nc_002077,nc_001401,nc_001729,nc_001863,nc_001829,nc_001862,nc_000883,nc_001701,nc_001510,nc_006152,nc_006261,af063497,u89790,af043303,af028705,af028704,j02275,j01901,j02275,x01457,af288061,ah009962,ay028226,ay028223,nc_001358,nc_001540,af513851,af513852,ay530579;其公开内容通过引用并入本文用于教导细小病毒和aav核酸以及氨基酸序列。还参见例如,srivistava等人,(1983)j.virology45:555;chiorini等人,(1998)j.virology71:6823;chiorini等人,(1999)j.virology73:1309;bantel-schaal等人,(1999)j.virology73:939;xiao等人,(1999)j.virology73:3994;muramatsu等人,(1996)virology221:208;shade等人,(1986)j.virol.58:921;gao等人,(2002)proc.nat.acad.sci.usa99:11854;moris等人,(2004)virology33-:375-383;国际专利公开wo00/28061,wo99/61601,wo98/11244;和美国专利no.6,156,303;其公开内容通过引用并入本文用于教导细小病毒和aav核酸以及氨基酸序列。也参见表1。
bernardn.fields等人,virology,第2卷,第69和70章(第4版,lippincott-ravenpublishers)更详细地描述了自主细小病毒和aav的衣壳结构。还参见,aav2(xie等人(2002)proc.nat.acad.sci.99:10405-10),aav4(padron等人(2005)j.virol.79:5047-58),aav5(walters等人(2004)j.virol.78:3361-71)和cpv(xie等人(1996)j.mol.biol.6:497-520和tsao等人(1991)science251:1456-64)的晶体结构的描述。
如本文所用,术语“趋向性”是指病毒优先进入某些细胞或组织,任选随后表达(例如,转录和任选翻译)由细胞中病毒基因组携带的序列,例如,对于重组病毒,表达目标异源核酸。本领域技术人员将理解,在不存在反式作用因子的情况下,来自病毒基因组的异源核酸序列的转录可能不被起始,例如对于诱导型启动子或以其他方式受调控核酸序列。在raav基因组的情况下,来自病毒基因组的基因表达可以来自稳定整合的原病毒、来自非整合的附加体、以及病毒可以在细胞内采取的任何其他形式。
除非另有说明,“有效转导”或“有效趋向性”或类似术语可通过参照合适的对照来确定(例如,分别为对照的至少约50%,60%,70%,80%,85%,90%,95%,98%,99%,100%或更多的转导或趋向性)。合适的对照将取决于多种因素,包括所需的趋向性概况(tropismprofile)。
除非另有说明,本文所用的术语“多肽”涵盖肽和蛋白两者。
“多核苷酸”是核苷酸碱基的序列,并且可以是rna、dna或dna-rna杂合序列(包括天然存在和非天然存在的核苷酸两者),但在代表性的实施方案中为单链或双链dna序列。
如本文所用,“分离的”多核苷酸(例如,“分离的dna”或“分离的rna”)意指至少部分地与天然存在的生物体或病毒的至少一些其他组分(例如,通常发现的与所述多核苷酸结合的细胞或病毒结构组分或其他多肽或核酸)分离的多核苷酸。在代表性实施方案中,与起始材料相比,“分离的”核苷酸被富集至少约10倍、100倍、1000倍、10,000倍或更多。
同样,“分离的”多肽意指至少部分地与天然存在的生物体或病毒的至少一些其他组分(例如,通常发现的与所述多肽结合的细胞或病毒结构组分或其他多肽或核酸)分离的多肽。在代表性实施方案中,与起始材料相比,“分离的”多肽被富集至少约10倍、100倍、1000倍、10,000倍或更多。
如本文所用,“分离”或“纯化”(或语法等同物)病毒载体,意指病毒载体至少部分地与起始材料中的至少一些其他组分分离。在代表性实施方案中,与起始材料相比,“分离的”或“纯化的”病毒载体被富集至少约10倍、100倍、1000倍、10,000倍或更多。
“治疗性蛋白”是可以减轻、减少、预防、延迟和/或稳定由细胞或受试者中蛋白的不存在或缺陷引起的症状的蛋白和/或是以其他方式向受试者赋予益处的蛋白。
如本文所用,“治疗性rna分子”或“功能性rna分子”可以是反义核酸,核酶(例如,如美国专利no.5,877,022中所述),影响剪接体介导的反式剪接的rna(参见,puttaraju等人(1999)naturebiotech.17:246;美国专利no.6,013,487;美国专利no.6,083,702),包括sirna、shrna或mirna的干扰rna(rnai),其介导基因沉默(参见,sharp等人,(2000)science287:2431),和任何其他非翻译rna,例如本领域已知的“指导”rna(gorman等人(1998)proc.nat.acad.sci.usa95:4929;yuan等人的美国专利no.5,869,248)等。
术语“治疗(treat)”、“治疗(treating)”或“治疗(treatmentof)”(及其语法变体)意指受试者状况的严重程度降低,至少部分改善或稳定和/或实现至少一种临床症状的某些减轻、缓解、减少或稳定,和/或疾病或病症进展中存在延迟。
术语“预防(prevent)”,“预防(preventing)”和“预防(prevention)”(及其语法变体)意指预防和/或延迟受试者中疾病、病症和/或临床症状的发作和/或相对于不存在本发明方法的情况下将发生的情况的疾病、病症和/或临床症状的发病严重程度的降低。预防可以是完全的,例如完全没有疾病、病症和/或临床症状。预防也可以是部分的,使得受试者中疾病、病症和/或临床症状的发生和/或发病的严重程度小于不存在本发明的情况下将发生的情况。
如本文所用,“治疗有效”量是足以对受试者提供某些改善或益处的量。或者说,“治疗有效”量是提供受试者的至少一种临床症状的某些减轻、缓解、减少或稳定的量。本领域技术人员将理解,治疗效果不需要是完全的或治愈的,只要对受试者提供某些益处即可。
如本文所用,“预防有效”量是足以预防和/或延迟受试者中疾病、病症和/或临床症状的发作和/或相对于在不存在本发明方法的情况下将发生的情况而减少和/或延迟受试者中疾病、病症和/或临床症状的发病的严重程度的量。本领域技术人员将理解,预防水平不需要是完全的,只要对受试者提供某些益处即可。
术语“异源核苷酸序列”和“异源核酸分子”在本文中可互换使用,并且是指非天然存在于病毒中的核酸分子和/或核苷酸序列。通常,异源核酸包含开放阅读框,其编码目标蛋白、蛋白片段、肽或非翻译rna(例如,用于递送至细胞或受试者)。
如本文所用,术语“病毒载体”、“载体”或“基因递送载体”是指这样的病毒(例如,aav)颗粒,其作为核酸递送载体起作用,并且其包含包装在病毒粒子内的载体基因组(例如,病毒dna[vdna])。或者,在某些情况下,术语“载体”可用于指单独的载体基因组/vdna。
“raav载体基因组”或“raav基因组”是包含一种或多种异源核酸序列的aav基因组(即,vdna)。raav载体通常仅需要顺式的末端重复序列(tr)来产生病毒。所有其他病毒序列均是非必需的,并且可以反式提供(muzyczka(1992)curr.topicsmicrobiol.immunol.158:97)。通常,raav载体基因组将仅保留一个或多个tr序列以最大化可被载体有效包装的转基因的大小。结构和非结构蛋白编码序列可以反式提供(例如由载体例如质粒,或通过将序列稳定整合到包装细胞中)。在本发明的实施方案中,raav载体基因组包含至少一种末端重复序列(tr)序列(例如,aavtr序列)、任选两个tr(例如,2个aavtr),其通常将在载体基因组的5'和3'末端处并且侧接异源核酸序列,但无需与之邻接。tr可以彼此相同或不同。
术语“末端重复序列”或“tr”包括形成发夹结构并起到反向末端重复序列的作用(即,介导所需功能例如复制、病毒包装、整合和/或原病毒拯救等)的任何病毒末端重复序列或合成序列。tr可以是aavtr或非aavtr。例如,非aavtr序列如其他细小病毒(例如犬细小病毒(cpv)、小鼠细小病毒(mvm)、人细小病毒b-19)的那些或任何其他合适的病毒序列(例如用作sv40复制起点的sv40发夹)可被用作tr,其可以通过截短、置换、缺失、插入和/或添加而进一步修饰。此外,tr可以是部分或完全合成的,例如,如samulski等人的美国专利no.5,478,745中所述的“双重d序列”。
“aav末端重复序列”或“aavtr”可以来自任何aav,包括但不限于血清型1、2、3、4、5、6、7、8、9、10或11或现在已知或以后发现的任何其他aav(参见例如表1)。aav末端重复序列不需要具有天然末端重复序列(例如天然aavtr序列可以通过插入、缺失、截短和/或错义突变来改变),只要该末端重复序列介导所需功能例如复制、病毒包装、整合和/或原病毒拯救等即可。
本发明的病毒载体还可以是“靶向”病毒载体(例如,具有定向趋向性)和/或“杂合”细小病毒(即,其中病毒tr和病毒衣壳来自不同的细小病毒),如国际专利公开wo00/28004和chao等人(2000)moleculartherapy2:619中所述。
本发明的病毒载体还可以是双链体细小病毒颗粒,如国际专利公开wo01/92551(其公开内容通过引用全部并入本文)中所述。因此,在一些实施方案中,双链(双链体)基因组可被包装到本发明的病毒衣壳中。
此外,病毒衣壳或基因组元件可含有其他修饰,包括插入、缺失和/或置换。
如本文所用,术语“氨基酸”涵盖任何天然存在的氨基酸、其修饰形式和合成氨基酸。
天然存在的、左旋(l-)氨基酸显示于表2。
或者,氨基酸可以是经修饰的氨基酸残基(非限制性实例显示于表3)和/或可以是通过翻译后修饰(例如乙酰化、酰胺化、甲酰化、羟基化、甲基化、磷酸化或硫酸化)来修饰的氨基酸。
此外,非天然存在的氨基酸可以是如wang等人annurevbiophysbiomolstruct.35:225-49(2006))所描述的“非天然”氨基酸。这些非天然氨基酸可有利地用于将目标分子化学连接至aav衣壳蛋白。
修饰的aav衣壳蛋白以及包含其的病毒衣壳和病毒载体
本发明提供包含氨基酸序列中的突变(即修饰)的aav衣壳蛋白以及包含修饰的aav衣壳蛋白的病毒衣壳和病毒载体。本发明人已经发现,本文所述的氨基酸位置处的修饰例如置换可赋予包含修饰的aav衣壳蛋白的病毒载体一种或多种所需特性,包括但不限于表面上具有硫酸肝素的细胞的选择性转导和视网膜细胞和/或视网膜色素上皮细胞的增强的转导。
在具体的实施方案中,本发明的修饰的aav衣壳蛋白包含天然aav4衣壳蛋白的氨基酸序列或来自另一aav(包括但不限于aav5、aav7、aav8和aav9)的衣壳蛋白的相应区域中的一个或多个突变(例如,置换)。
如本文所用,氨基酸序列中的“突变”或“修饰”包括置换、插入和/或缺失,每个置换、插入和/或缺失可涉及一个、两个、三个、四个、五个、六个、七个、八个、九个、十个或更多氨基酸。在具体的实施方案中,该修饰是置换。例如,在具体的实施方案中,aav4衣壳蛋白序列在氨基酸位置530、584、585、586和/或587处以任意组合进行修饰。
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为np_044927(seqidno:4)的aav4衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为yp_680426(seqidno:2)的aav2衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为aas99264(seqidno:9)的aav9衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为yp_068409(seqidno:5)的aav5衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸编号基于genbank登录号为np_049542(seqidno:1)的aav1衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为yp_077178(seqidno:7)的aav7衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为yp_077180(seqidno:8)的aav8衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为aat46337(seqidno:10)的aav10衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为aat46339(seqidno:11)的aav11衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为nc_001863(seqidno:3)的aav3b衣壳蛋白的氨基酸序列:
在如本文所述的某些实施方案中,氨基酸残基编号基于genbank登录号为abi16639(seqidno:12)的aav12衣壳蛋白的氨基酸序列:
本发明的修饰的病毒衣壳蛋白可以是但不限于aav衣壳蛋白,其中来自一种aav衣壳蛋白的氨基酸被置换成另一种aav衣壳蛋白,并且置换和/或插入的氨基酸可以来自任何来源,并且进一步可以是天然存在的或者是部分或完全合成的。此外,本发明的aav衣壳蛋白可具有天然氨基酸序列或合成氨基酸序列。
如本文所述,来自许多aav的衣壳蛋白的核酸和氨基酸序列是本领域已知的。因此,例如,对于包括例如aav5、aav7、aav8和aav9的任何其他aav衣壳蛋白而言,“对应于”参考aav4衣壳蛋白的氨基酸位置530、584、585、586和587的氨基酸可被容易地确定(例如,通过使用本领域熟知的序列比对)。“对应于”天然aav4衣壳蛋白中的这些位置的其他aav血清型或修饰的aav衣壳中的氨基酸位置对于本领域技术人员而言是显而易见的,并且可使用序列比对技术(参见例如wo2006/066066的图7)和/或晶体结构分析(padron等人(2005)j.virol.79:5047-58)容易地确定。可以在其他aav血清型的这些各自的位置处置换天然氨基酸的氨基酸残基的实例列举在表2和3中。
本发明预期本发明的修饰的衣壳蛋白可以通过修饰现在已知或以后发现的任何aav的衣壳蛋白来产生。此外,待修饰的aav衣壳蛋白可以是天然存在的aav衣壳蛋白(例如aav4、aav5、aav7、aav8或aav9衣壳蛋白或表1中所示的任何aav),但不限于此。本领域技术人员将理解,对aav衣壳蛋白的多种操作是本领域已知的,并且本发明不限于天然存在的aav衣壳蛋白的修饰。例如,与天然存在的aav相比,待修饰的衣壳蛋白可能已经具有改变(例如,衍生自天然存在的aav衣壳蛋白,例如aav1、aav2、aav3a、aav3b、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11和/或aav12或现在已知或以后发现的任何其他aav血清型)。这样的aav衣壳蛋白也在本发明的范围内。
因此,在具体实施方案中,待修饰的aav衣壳蛋白可衍生自天然存在的aav,但进一步包含插入和/或置换到衣壳蛋白中的一种或多种外源序列(例如,对天然病毒是外源的),和/或已通过缺失一个或多个氨基酸而进行了改变。
因此,当在本文中提及特定aav衣壳蛋白(例如,aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11或aav12衣壳蛋白或来自表1所示任意aav的衣壳蛋白等)时,意欲包括天然衣壳蛋白以及具有除本发明的修饰之外的改变的衣壳蛋白。这样的改变包括置换、插入和/或缺失。在具体的实施方案中,与天然aav衣壳蛋白序列相比,衣壳蛋白包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个,小于20个,小于30个,小于40个,小于50个,小于60个,或小于70个插入其中的氨基酸(非本发明的插入)。在本发明的实施方案中,与天然aav衣壳蛋白序列相比,衣壳蛋白包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个,小于20个,小于30个,小于40个,小于50个,小于60个,或小于70个氨基酸置换(非本发明所述的氨基酸置换)。在本发明的实施方案中,与天然aav衣壳蛋白序列相比,衣壳蛋白包含1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个,小于20个,小于30个,小于40个,小于50个,小于60个,或小于70个氨基酸的缺失(非本发明的氨基酸缺失)。
因此,例如,术语“aav4衣壳蛋白”包括具有天然aav4衣壳蛋白序列(参见genbank登录号nc_044927)的aav衣壳蛋白、以及在天然aav4衣壳蛋白序列中包含置换、插入和/或缺失(如本文所述)的那些。
在具体实施方案中,aav衣壳蛋白具有天然aav衣壳蛋白序列或具有与天然aav衣壳蛋白序列至少约90%、95%、97%、98%或99%相似或同一的氨基酸序列。例如,在具体实施方案中,“aav4”衣壳蛋白涵盖天然aav4衣壳蛋白序列以及与天然aav4衣壳蛋白序列至少约90%、95%、97%、98%或99%相似或同一的序列。
确定两个或更多个氨基酸序列之间的序列相似性或同一性的方法是本领域已知的。序列相似性或同一性可以使用本领域已知的标准技术确定,包括但不限于smith&waterman,adv.appl.math.2,482(1981)的局部序列同一性算法,通过needleman&wunsch,j.mol.biol.48,443(1970)的序列同一性比对算法,通过pearson&lipman,proc.natl.acad.sci.usa85,2444(1988)的相似性搜索方法,通过这些算法的计算机化执行(gap、bestfit、fasta和tfasta,在wisconsingeneticssoftwarepackage中,geneticscomputergroup,575sciencedrive,madison,wi),devereux,nucl.acidres.12,387-395(1984)等人的最佳拟合序列程序,或通过检查。
另一种合适的算法是blast算法,其描述于altschul等人j.mol.biol.215,403-410,(1990)和karlin等人proc.natl.acad.sci.usa90,5873-5787(1993)中。特别有用的blast程序是wu-blast-2程序,其获自altschul等人methodsinenzymology,266,460-480(1996);blast.wustl/edu/blast/readme.html。wu-blast-2使用几个搜索参数,所述参数任选设置为默认值。参数是动态值并且由程序本身根据具体序列的组成和正被搜索的目标序列的具体数据库的组成来建立;然而,可调整数值以增加灵敏度。
此外,额外有用的算法是如altschul等人(1997)nucleicacidsres.25,3389-3402所报道的带缺口的blast。
如例如美国专利no.5,863,541中所述,修饰的病毒衣壳可用作“衣壳载体”。可以通过修饰的病毒衣壳包装并转移到细胞中的分子包括异源dna、rna、多肽、小有机分子、金属或其组合。
异源分子定义为在aav感染中不天然存在的那些,例如,并非由野生型aav基因组编码的那些。此外,治疗上有用的分子可以与嵌合病毒衣壳的外部结合,用于将分子转移到宿主靶细胞中。这样的结合分子可包括dna、rna、小有机分子、金属、碳水化合物、脂质和/或多肽。在本发明的一个实施方案中,治疗上有用的分子与衣壳蛋白共价连接(即缀合或化学偶联)。共价连接分子的方法是本领域技术人员已知的。
在其他实施方案中,可在施用递送编码目标的多肽或功能rna的核酸的病毒载体之前和/或同时(例如,在彼此的几分钟或几小时内)施用病毒衣壳以阻断某些细胞位点。例如,可以递送本发明的衣壳以阻断具体细胞上的细胞受体,并且可以随后或同时施用递送载体,这可以减少阻断细胞的转导,并且增强其他靶标(例如cns祖细胞和/或神经母细胞)的转导。
根据代表性实施方案,修饰的病毒衣壳可以在根据本发明所述的修饰的病毒载体之前和/或同时施用于受试者。此外,本发明提供了包含本发明的修饰的病毒衣壳的组合物和药物制剂;任选地,该组合物还包含本发明的修饰的病毒载体。
本文进一步提供了包含本文所述的aav衣壳蛋白的aav衣壳以及包含该aav衣壳的病毒载体。本文还提供了在药学上可接受的载体中包含本发明的病毒载体的组合物。
本发明还提供了编码本发明的修饰的病毒衣壳和衣壳蛋白的核酸分子(任选地,分离的核酸分子)。进一步提供了包含核酸分子的载体和包含本发明的核酸分子和/或载体的细胞(体内或培养的)。合适的载体包括但不限于病毒载体(例如腺病毒、aav、疱疹病毒、牛痘、痘病毒、杆状病毒等)、质粒、噬菌体、yac、bac等。这样的核酸分子、载体和细胞可用作例如用于产生如本文所述的修饰的病毒衣壳或病毒载体的试剂(例如,辅助包装构建体或包装细胞)。
根据本发明所述的病毒衣壳可以使用本领域已知的任何方法产生,例如通过从杆状病毒的表达产生(brown等人(1994)virology198:477-488)。
根据本发明所述的对aav衣壳蛋白的修饰是“选择性”修饰。该方法与先前使用aav血清型之间的整个亚基或大结构域交换的工作形成对比(参见例如国际专利公开wo00/28004和hauck等人(2003)j.virology77:2768-2774)。在具体实施方案中,“选择性”修饰导致小于约20、18、15、12、10、9、8、7、6、5、4、3、2或1个连续氨基酸的插入和/或置换和/或缺失。
本发明的修饰的衣壳蛋白和衣壳可进一步包含现在已知或以后鉴定的任何其他修饰。
例如,本发明的aav衣壳蛋白和病毒衣壳可以是嵌合的,因为它们可以包含来自另一种病毒(任选地另一种细小病毒或aav)的衣壳亚基的全部或一部分,例如,如国际专利公开wo00/28004中所述。
病毒衣壳可以是靶向病毒衣壳,其包含靶向序列(例如,置换或插入病毒衣壳中),其引导病毒衣壳与存在于期望靶组织上的细胞表面分子相互作用(参见例如,国际专利公开wo00/28004和hauck等人(2003)j.virology77:2768-2774);shi等人humangenetherapy17:353-361(2006)[描述在aav衣壳亚基的位置520和/或584处的整合素受体结合基序rgd的插入];和美国专利no.7,314,912[描述在aav2衣壳亚基的氨基酸位置447、534、573和587之后的含有rgd基序的p1肽的插入])。耐受插入的aav衣壳亚基内的其他位置是本领域已知的(例如,grifman等人moleculartherapy3:964-975(2001)描述的位置449和588)。
在代表性实施方案中,靶向序列可以是将感染引导至特定细胞类型的病毒衣壳序列(例如,自主细小病毒衣壳序列、aav衣壳序列或任意其他病毒衣壳序列)。
作为另一个非限制性实例,可以将肝素结合结构域(例如,呼吸道合胞病毒肝素结合结构域)插入或置换到通常不结合hs受体的衣壳亚基(例如aav4、aav5)中,以使肝素与所得突变体结合。
在代表性实施方案中,外源靶向序列可以是编码肽的任何氨基酸序列,所述肽改变包含修饰的aav衣壳蛋白的病毒衣壳或病毒载体的趋向性。在具体的实施方案中,靶向肽或蛋白可以是天然存在的,或者可替代地是完全或部分合成的。示例性靶向序列包括与细胞表面受体和糖蛋白结合的配体和其他肽,如rgd肽序列、缓激肽、激素、肽生长因子(例如表皮生长因子、神经生长因子、成纤维细胞生长因子、血小板衍生生长因子、胰岛素样生长因子i和ii等)、细胞因子、黑素细胞刺激激素(例如α、β或γ)、神经肽和内啡肽等、以及其保留将细胞靶向至其同源受体的能力的片段。其他说明性肽和蛋白包括p物质、角质形成细胞生长因子、神经肽y、胃泌素释放肽、白细胞介素2、鸡蛋清溶菌酶、促红细胞生成素、促性腺释放激素、皮质抑素、β-内啡肽、亮氨酸脑啡肽、强腓肽b(rimorphin)、α-新脑啡肽、血管紧张素、pneumadin、血管活性肠肽、神经降压素、胃动素以及它们的如上所述的片段。作为另一个替代方案,来自毒素的结合结构域(例如破伤风毒素或蛇毒素,例如α-银环蛇毒素等)可以作为靶向序列被置换到衣壳蛋白中。在另一个代表性实施方案中,aav衣壳蛋白可以通过将如cleves(currentbiology7:r318(1997))所述的“非经典”输入/输出信号肽(例如,成纤维细胞生长因子-1和-2、白细胞介素1、hiv-1tat蛋白、疱疹病毒vp22蛋白等)置换到aav衣壳蛋白中来进行修饰。还涵盖指导特定细胞摄取的肽基序,例如fvflp肽基序触发肝细胞摄取。
噬菌体展示技术以及本领域已知的其他技术可用于鉴定识别任何目标细胞类型的肽。
靶向序列可以编码靶向至细胞表面结合位点(包括受体(例如蛋白、碳水化合物、糖蛋白或蛋白多糖))的任何肽。细胞表面结合位点的实例包括但不限于硫酸乙酰肝素、硫酸软骨素和其他糖胺聚糖、唾液酸部分、聚唾液酸部分、糖蛋白和神经节苷脂、mhci糖蛋白、膜糖蛋白上发现的碳水化合物组分,包括甘露糖、n-乙酰基-半乳糖胺、n-乙酰基-葡糖胺、岩藻糖、半乳糖等。
作为又一个替代方案,靶向序列可以是能够用于与另一靶向对细胞的进入的分子化学偶联的肽(例如,可包含能够通过其r基团化学偶联的精氨酸和/或赖氨酸残基)。
本发明的前述实施方案可用于将异源核酸递送至如本文所述的细胞或受试者。因此,在一个实施方案中,本发明提供了将核酸分子引入细胞的方法,其包括使细胞与本发明的病毒载体和/或组合物接触。
本文进一步提供了将核酸分子递送至受试者的方法,其包括向受试者施用本发明的病毒载体和/或本发明的组合物。在一些实施方案中,将病毒载体或组合物施用于受试者的中枢神经系统。
本文另外提供了选择性转导表面上具有硫酸乙酰肝素的细胞的方法,其包括使细胞与本发明的病毒载体和/或本发明的组合物接触。
本发明进一步提供了将目标核酸分子递送至视网膜细胞和/或视网膜色素上皮细胞的方法,其包括使细胞与本发明的病毒载体接触,其中病毒载体包含目标核酸分子。在该方法的一些实施方案中,目标核酸分子编码治疗性蛋白或治疗性rna。
在上述方法的一些实施方案中,视网膜细胞和/或视网膜色素上皮细胞可以在受试者中,并且在一些实施方案中,受试者可以是人受试者。
本发明进一步提供了治疗受试者眼睛病症或缺陷的方法,其包括向受试者玻璃体内施用本发明的病毒载体,其中病毒载体包含编码有效治疗受试者眼睛病症或缺陷的治疗性蛋白或治疗性rna的核酸分子。
在进一步的实施方案中,本发明提供了选择性转导视网膜细胞和/或视网膜色素上皮细胞的方法,其包括使细胞与包含如本文所述的aav衣壳蛋白的病毒载体接触。
本领域技术人员将理解,对于一些aav衣壳蛋白,相应的修饰将是插入和/或置换,这取决于是否相应的氨基酸位置是部分或完全存在于病毒中,或者可替代地完全不存在。同样地,当修饰除aav4之外的aav时,特定氨基酸位置可以不同于aav4中的位置(参见例如表4,其显示了对应于aav4中s257的氨基酸残基的代表性实例)。如本文其他部分所讨论的,使用熟知的技术,相应的氨基酸位置对于本领域技术人员而言是显而易见的。
上述修饰可以彼此组合和/或与现在已知或以后发现的任何其他修饰组合并入本发明的衣壳蛋白或衣壳中。
本发明还涵盖包含本发明的修饰的衣壳蛋白和衣壳的病毒载体。在具体的实施方案中,病毒载体是细小病毒载体(例如,包含细小病毒衣壳和/或载体基因组),例如aav载体(例如,包含aav衣壳和/或载体基因组)。在代表性实施方案中,病毒载体包含修饰的aav衣壳,其包含本发明的修饰的衣壳亚基和载体基因组。
例如,在代表性实施方案中,病毒载体包含:(a)包含本发明的修饰的衣壳蛋白的修饰的病毒衣壳(例如,修饰的aav衣壳);(b)包含末端重复序列(例如aavtr)的核酸,其中包含末端重复序列的核酸被修饰的病毒衣壳包裹。核酸可任选地包含两个末端重复序列(例如,两个aavtr)。
在代表性实施方案中,病毒载体是重组病毒载体,其包含编码目标蛋白、肽和/或功能性rna的异源核酸分子。
本领域技术人员将理解,本发明的修饰的衣壳蛋白、病毒衣壳和病毒载体不包括在其天然状态下具有特定位置处的指定氨基酸的那些衣壳蛋白、衣壳和病毒载体(即,不是突变体)。
重组病毒载体
本发明的病毒载体可用于体外、离体和体内将核酸递送至细胞。具体地,病毒载体可以有利地用于将核酸递送或转移至动物细胞,包括哺乳动物细胞。
可以在本发明的病毒载体中递送任意目标异源核酸序列。目标核酸包括编码多肽、包括治疗性(例如,用于医学或兽医用途)或免疫原性(例如用于疫苗)蛋白和/或功能性或治疗性rna分子的核酸。
编码多肽的异源核酸序列包括编码报告多肽(例如酶)的那些。报道多肽是本领域已知的,并且包括但不限于绿色荧光蛋白(gfp)、β-半乳糖苷酶、碱性磷酸酶、荧光素酶和氯霉素乙酰转移酶基因。
任选地,异源核酸编码分泌的多肽(例如多肽,其为其天然状态的分泌的多肽或已被工程化以分泌,例如,通过与本领域已知的分泌信号序列可操作地结合而分泌)。
或者,在本发明的具体实施方案中,异源核酸可以编码反义核酸、核酶(例如,如美国专利no.5,877,022中所述)、影响剪接体介导的反式剪接的rna(参见puttaraju等人(1999)naturebiotech.17:246;美国专利no.6,013,487;美国专利no.6,083,702)、介导基因沉默的干扰rna(rnai)、包括sirna、shrna或mirna(参见sharp等人(2000)science287:2431)和其他非翻译rna例如“指导”rna(gorman等人(1998)proc.nat.acad.sci.usa95:4929;yuan等人的美国专利no.5,869,248)等。示例性的非翻译rna包括针对多重耐药性(mdr)基因产物的rnai(例如,用于治疗和/或预防肿瘤和/或用于施用于心脏以防止通过化疗的损伤)、针对肌肉生长抑制素的rnai(例如,用于杜兴肌营养不良症)、针对vegf的rnai(例如,用于治疗和/或预防肿瘤)、针对受磷蛋白的rnai(例如,用于治疗心血管疾病,参见例如andino等人j.genemed.10:132-142(2008)和li等人actapharmacolsin.26:51-55(2005))、受磷蛋白抑制性或显性阴性分子如受磷蛋白s16e(例如,用于治疗心血管疾病,参见例如hoshijima等人nat.med.8:864-871(2002))、对腺苷激酶的rnai(例如,用于癫痫)、以及针对病原生物和病毒(例如,乙型肝炎和/或丙型肝炎病毒、人类免疫缺陷病毒、cmv、单纯疱疹病毒、人乳头瘤病毒等)的rnai。
此外,可以递送指导可变剪接的核酸序列。为了说明,与肌营养不良蛋白外显子51的5'和/或3'剪接位点互补的反义序列(或其他抑制序列)可以与u1或u7小核(sn)rna启动子一起递送以诱导跳过该外显子。例如,包含位于反义/抑制序列5'的u1或u7snrna启动子的dna序列可以在本发明的修饰的衣壳中包装并递送。
病毒载体还可以包含异源核酸,其与宿主染色体上的基因座具有同源性并与之重组。例如,该方法可用于校正宿主细胞中的遗传缺陷。
作为进一步替代方案,异源核酸可以编码在体外、离体或体内在细胞中理想地产生的任意多肽。例如,可以将病毒载体引入培养的细胞中并由其分离表达的基因产物。
本领域技术人员将理解,目标异源核酸可以与适当的控制序列可操作地结合。例如,异源核酸可以与表达控制元件如转录/翻译控制信号、复制起点、多腺苷酸化信号、内部核糖体进入位点(ires)、启动子和/或增强子等可操作地结合。
此外,例如,通过存在或不存在选择性地阻断特定位点处的剪接活性的寡核苷酸、小分子和/或其他化合物来调控不同内含子的选择性剪接,可以在转录后水平实现目标异源核酸的受调控的表达(例如,如wo2006/119137中所述)。
本领域技术人员将理解,取决于所需的水平和组织特异性表达,可以使用各种启动子/增强子元件。取决于所需的表达模式,启动子/增强子可以是组成型或诱导型的。启动子/增强子可以是天然的或外源的,并且可以是天然的或合成的序列。通过外源,意欲表示在引入转录起始区的野生型宿主中不存在转录起始区。
在具体的实施方案中,启动子/增强子元件对于待治疗的靶细胞或受试者可以是天然的。在代表性实施方案中,启动子/增强子元件相对于异源核酸序列可以是天然的。通常选择启动子/增强子元件,使得其在目标靶细胞中起作用。此外,在具体实施方案中,启动子/增强子元件是哺乳动物启动子/增强子元件。启动子/增强子元件可以是组成型或诱导型的。
诱导型表达控制元件通常在那些需要提供针对异源核酸序列表达的调控的应用中是有利的。用于基因递送的诱导型启动子/增强子元件可以是组织特异性或优选的启动子/增强子元件,并且包括肌肉特异性或优选的(包括心脏、骨骼和/或平滑肌特异性或优选的),神经组织特异性或优选的(包括脑特异性或优选的),眼特异性或优选的(包括视网膜特异性或角膜特异性),肝特异性或优选的,骨髓特异性或优选的,胰腺特异性或优选的,脾特异性或优选的,以及肺特异性或优选的启动子/增强子元件。其他诱导型启动子/增强子元件包括激素诱导型和金属诱导型元件。示例性诱导型启动子/增强子元件包括但不限于tet开/关元件、ru486诱导型启动子、蜕皮激素诱导型启动子、雷帕霉素诱导型启动子和金属硫蛋白启动子。
在其中异源核酸序列被转录且然后在靶细胞中被翻译的实施方案中,通常包括特异性起始信号以有效翻译插入的蛋白编码序列。可以包括atg起始密码子和相邻序列的这些外源翻译控制序列可以具有天然的和合成的各种起源。
根据本发明的病毒载体提供了将异源核酸递送到包括分裂和非分裂细胞的范围宽广的细胞中的手段。病毒载体可用于在体外将目标核酸递送至细胞,例如,以在体外产生多肽或用于离体基因治疗。病毒载体还可用于将核酸递送至需要其的受试者(例如,以表达免疫原性或治疗性多肽或功能性rna)的方法。以这种方式,多肽或功能rna可以在受试者中体内产生。受试者可因该受试者缺乏所述多肽而需要所述多肽。此外,可以实施该方法,因为在受试者中产生多肽或功能性rna可以赋予一些有益效果。
病毒载体还可用于在培养的细胞或受试者中产生目标多肽或功能性rna(例如,使用受试者作为生物反应器产生多肽或观察功能性rna对受试者的作用,例如,结合筛选方法)。
通常,本发明的病毒载体可用于递送编码多肽或功能性rna的异源核酸,以治疗和/或预防递送治疗性多肽或功能性rna对其有益的任何疾病状态。本发明的说明性疾病状态包括但不限于年龄相关性黄斑变性、莱伯氏先天性黑蒙症1型、莱伯氏先天性黑蒙症2型、色素性视网膜炎、视网膜劈裂、失色症、色盲、先天性静止性夜盲症或其任意组合。
基因转移具有用于理解和提供疾病状态治疗的巨大潜在用途。存在许多缺陷基因是已知的并且已被克隆的遗传疾病。一般而言,上述疾病状态分为两类:缺乏状态,通常是酶的缺乏状态,其通常以隐性方式遗传;和不平衡状态,其可涉及调控蛋白或结构蛋白,并且通常以显性方式遗传。对于缺乏状态疾病,基因转移可用于将正常基因带入受影响的组织中以进行替代治疗,以及使用反义突变产生疾病的动物模型。对于不平衡疾病状态,基因转移可用于在模型系统中产生疾病状态,然后可将其用于抵抗疾病状态。因此,根据本发明的病毒载体允许治疗和/或预防遗传疾病。
根据本发明的病毒载体也可用于在体外或体内向细胞提供功能性rna。功能性rna在细胞中的表达例如可以减少细胞对具体靶蛋白的表达。因此,可施用功能性rna以降低有需要的受试者中的特定蛋白的表达。功能性rna也可以在体外施用于细胞以调控基因表达和/或细胞生理学,例如,以优化细胞或组织培养系统或筛选方法。
此外,根据本发明的病毒载体可用于诊断和筛选方法,藉此目标核酸在细胞培养系统或可替代地转基因动物模型中瞬时或稳定表达。
在一些实施方案中,本发明的病毒载体可用于在受试者中诱导免疫应答,并且病毒载体可包含编码免疫原的核苷酸序列。
本发明的病毒载体还可用于各种非治疗目的,包括但不限于用于评估基因靶向、清除、转录、翻译等的方案中,如本领域技术人员所显而易见。病毒载体也可用于评估安全性(扩散、毒性、免疫原性等)的目的。例如,这样的数据在评估临床疗效之前被美国食品和药物管理局视为监管批准过程的一部分。
或者,可以将病毒载体离体施用于细胞,并将改变的细胞施用于受试者。将包含异源核酸的病毒载体引入细胞,并将该细胞施用于受试者,其中异源核酸可在受试者中表达。
受试者、药物制剂和施用模式
根据本发明的病毒载体和衣壳可用于兽医和医学应用两者中。合适的受试者包括禽类和哺乳动物两者。如本文所用,术语“禽类”包括但不限于鸡、鸭、鹅、鹌鹑、火鸡、野鸡、鹦鹉、长尾小鹦鹉等。如本文所用,术语“哺乳动物”包括但不限于人类、非人灵长类动物、啮齿动物、牛类、绵羊类、山羊类、马类、猫类、犬类、兔类动物等。人类受试者包括新生儿、婴儿、青少年、成人和老年受试者。
在代表性实施方案中,受试者“需要”本发明的方法。
在具体的实施方案中,本发明提供药物组合物,其在药学上可接受的载体,以及任选其他医疗剂、药剂、稳定剂、缓冲剂、载体、佐剂、稀释剂等中包含本发明的病毒载体和/或衣壳。对于注射,载体通常是液体。对于其他施用方法,载体可以是固体或液体。对于吸入施用,载体是可吸入的,并且任选地可以是固体或液体颗粒形式。
“药学上可接受的”是指并非有毒或以其他方式不合需要的材料,即该材料可以施用于受试者而不会引起任何不希望的生物学效应。
本发明的一个方面是在体外将核酸转移至细胞的方法。可以根据适合于具体靶细胞的标准转导方法,以适当的感染复数将病毒载体引入细胞中。施用的病毒载体的滴度可以根据靶细胞类型和数量以及特定的病毒载体而改变,并且可以由本领域技术人员确定而无需过多的实验。在代表性实施方案中,将至少约103个感染单位,任选至少约105个感染单位引入细胞。
引入病毒载体的细胞可以是任何类型,包括但不限于眼睛细胞(包括视网膜细胞、视网膜色素上皮细胞和角膜细胞)。
可以将病毒载体体外引入细胞中,用于将修饰的细胞施用于受试者的目的。在具体的实施方案中,已从受试者中取出细胞,将病毒载体引入其中,且然后将该细胞施用回受试者中。从受试者中取出细胞以便离体操作、然后引入回受试者的方法是本领域已知的(参见例如美国专利no.5,399,346)。或者,可以将重组病毒载体引入来自供体受试者的细胞、引入培养的细胞、或引入来自任何其他合适来源的细胞,并将该细胞施用于需要其的受试者(即“受体”受试者)。
用于离体核酸递送的合适细胞如上所述。施用于受试者的细胞的剂量将根据受试者的年龄、状况和物种,细胞类型,细胞所表达的核酸,施用方式等而改变。通常,每个剂量,在药学上可接受的载体中施用至少约102至约108个细胞或至少约103至约106个细胞。在具体的实施方案中,用病毒载体转导的细胞以治疗有效量或预防有效量与药物载体组合施用于受试者。
本发明的另一方面是将病毒载体和/或病毒衣壳施用于受试者的方法。根据本发明的病毒载体和/或衣壳至需要其的人类受试者或动物的施用可以通过本领域已知的任何手段进行,但在特定的实施方案中,施用是玻璃体内施用。任选地,病毒载体和/或衣壳以治疗有效剂量或预防有效剂量在药学上可接受的载体中递送。
待施用于受试者的病毒载体和/或衣壳的剂量取决于施用模式、待治疗和/或预防的疾病或病症、个体受试者的病症、具体病毒载体或衣壳、以及待递送的核酸等,并且可以以常规方式确定。用于实现治疗效果的示例性剂量是至少约105、106、107、108、109、1010、1011、1012、103、1014、1015个转导单位,任选约108–1013个转导单位的滴度。
在具体的实施方案中,可以使用多于一次的施用(例如两次、三次、四次或更多次施用)以在不同间隔例如每天、每周、每月、每年等的时间段内实现期望水平的基因表达。
可注射剂可以以常规形式制备为液体溶液或悬浮液、适用于临注射前的液体中的溶液或悬浮液的固体形式,或制备为乳液。或者,可以以局部而非全身方式施用本发明的病毒载体和/或病毒衣壳,例如,在长效制剂(depot)或缓释制剂中。此外,病毒载体和/或病毒衣壳可以粘附至外科手术可植入的基质而递送(例如,如美国专利公开no.us-2004-0013645-a1中所述)。
cns的病症包括涉及视网膜、后路(posteriortract)和视神经的眼科病症(例如,色素性视网膜炎、糖尿病性视网膜病和其他视网膜退行性疾病、葡萄膜炎、年龄相关性黄斑变性、青光眼)。
大多数(如果不是全部)眼科疾病和病症与三种类型的适应症中的一种或多种相关:(1)血管生成、(2)炎症和(3)变性。本发明的递送载体可用于递送抗血管生成因子;抗炎因子;延缓细胞变性、促进细胞保留(cellsparing)或促进细胞生长和前述的组合的因子。
例如,糖尿病性视网膜病的特征在于血管生成。糖尿病性视网膜病可以通过眼内(例如,在玻璃体内)或眼周(例如,在眼球筋膜下区域)递送一种或多种抗血管生成因子来治疗。一种或多种神经营养因子也可以眼内(例如,玻璃体内)或眼周地共同递送。
葡萄膜炎涉及炎症。一种或多种抗炎因子可以通过眼内(例如,玻璃体或前房)施用本发明的递送载体来施用。
相比之下,色素性视网膜炎的特征在于视网膜变性。在代表性实施方案中,色素性视网膜炎可以通过眼内施用(例如,玻璃体施用)编码一种或多种神经营养因子的递送载体来治疗。
年龄相关性黄斑变性涉及血管生成和视网膜变性。可以通过眼内(例如玻璃体)施用编码一种或多种神经营养因子的本发明的递送载体和/或眼内或眼周(例如,在眼球筋膜下区域)施用编码一种或多种抗血管生成因子的本发明的递送载体来治疗该病症。
青光眼的特征在于眼压升高和视网膜神经节细胞丧失。青光眼的治疗包括使用本发明的递送载体施用一种或多种保护细胞免受兴奋毒性损伤的神经保护剂。这样的药剂包括眼内递送、任选玻璃体内递送的n-甲基-d-天冬氨酸(nmda)拮抗剂、细胞因子和神经营养因子。
在具体实施方案中,载体可包含分泌信号,如美国专利no.7,071,172中所述。
病毒载体和/或衣壳也可以施用于眼睛的不同区域如视网膜、角膜和/或视神经。
在具体的实施方案中,病毒载体和/或衣壳在液体制剂中通过直接注射(例如,立体定向注射)施用于cns中的所需区域或区室。在其他实施方案中,病毒载体和/或衣壳可以通过局部施用于所需区域或通过鼻内施用气溶胶制剂来提供。向眼睛的施用可以通过局部施用液滴来实现。作为进一步的替代方案,病毒载体和/或衣壳可以作为固体缓释制剂施用(参见例如,美国专利no.7,201,898)。
已经描述了本发明,将在以下实施例中更详细地解释本发明,所述实施例仅出于说明目的而包括在本文中,并且并不意欲限制本发明。
实施例
实施例1.
重组腺相关病毒(raav)已成为视网膜基因转移的优选载体。通过玻璃体将raav递送至视网膜导致少数血清型有效转导视网膜。这些少数血清型具有结合至硫酸乙酰肝素蛋白多糖(hspg)的衣壳蛋白。使用具有修饰的受体相互作用的raav衣壳评估衣壳和受体之间的相互作用。在成年小鼠中玻璃体内递送病毒,并在八周后进行评估。raav2的硫酸乙酰肝素(hs)结合残基中的突变导致内部视网膜转导的显著降低。使用设计者嵌合衣壳raav2.5将非hs结合raav1的元件添加至raav2。raav2.5沿视网膜血管转导并在muller神经胶质细胞中显示更大表达。加入结合至raav1的hs显示出荧光增加,其类似于用raav2.5观察到的表达模式。使用最近设计的嵌合衣壳raav2g9评估hs和半乳糖的双受体相互作用,以确定添加半乳糖结合是否会增强衣壳转导。相比于raav2,raav2g9显示眼底的荧光模式改变。raav2g9的转导概况显示趋向性向muller神经胶质细胞的转变。总之,hs结合对于成功的玻璃体内转导是必需的,并且这种转导可以通过添加半乳糖结合而偏向于特定的视网膜细胞。
实施例2.硫酸乙酰肝素结合促进视网膜处玻璃体内递送的腺相关病毒载体的累积以增强转导
许多腺相关病毒(aav)血清型在递送到视网膜下空间时有效地转导视网膜,但由于内界膜(ilm)的缘故递送到玻璃体时显示出有限的成功。视网膜下递送raav2及其hs结合缺陷衣壳表明外部视网膜的raav2转导通过hs非依赖性机制发生。然而,相比于raav2,玻璃体内递送hs-消融的raav2导致转导减少300倍。aav视网膜输送的荧光原位杂交(fish)揭示了ilm处aav2累积的机制受hs结合所影响。该机制在人离体视网膜上进行了测试,并且显示了仅与hs结合aav2衣壳相似的累积。为了评估hs结合是否可以应用于其他aav血清型以增强其转导,用单个氨基酸突变修饰aav1和aav8以结合hs并在小鼠中进行测试。aav1和aav8的hs结合突变体均具有比其非hs结合亲本衣壳更高的玻璃体内转导。为了理解hs结合对aav2趋向性的影响,在小鼠中玻璃体内测试具有双重聚糖使用的嵌合衣壳。相比于单独的hs结合,这些嵌合衣壳显示出增强的转导,其与趋向性的变化相关。总之,这表明hs结合用于将aav衣壳从玻璃体隔离到ilm,但不影响趋向性。增强的hs结合衣壳的视网膜转导提供了合理的设计策略,用于跨物种地对衣壳进行工程化而用于玻璃体内递送。
本研究显示hspg结合与视网膜中更大的累积和转导相关。我们验证了这种累积在小鼠和人类视网膜中是保守的。对任何aav衣壳添加hspg结合可以增加显示有效玻璃体内转导的血清型的数量。
腺相关病毒(aav)是一种小的(25nm)非致病性病毒,已被广泛研究作为基因转移应用的载体。该病毒由两部分组成:病毒基因组和蛋白衣壳。病毒基因组可以在很大程度上被所需的转基因替代,以产生用于基因递送的重组aav(raav)载体。蛋白衣壳通过各种聚糖和细胞表面受体而负责细胞附着和进入。存在11种天然存在的aav血清型,表示为aav1至aav11。已经阐明了几种aav血清型的聚糖和受体。硫酸乙酰肝素蛋白多糖(hspg)已被显示用于raav2和raav3细胞进入。raav6显示与hspg和唾液酸的双重聚糖相互作用;然而,单独的hspg结合对于细胞进入并不足够。唾液酸的各种连接对于raav1、raav4和raav5血清型的转导是重要的。n-连接的半乳糖用于转导raav9血清型。在细胞表面上表达的聚糖决定了用各种aav衣壳观察到的组织和细胞趋向性。除了与这些聚糖的附着之外,aav血清型与用于进入的细胞受体(包括人生长因子受体、整联蛋白和层粘连蛋白受体)相互作用。
raav已显示出视网膜基因转移的前景。除了如上所讨论的衣壳的影响之外,施用于视网膜的途径决定了转基因表达概况和功效。视网膜下(sr)递送在外核层(onl)和视网膜色素上皮(rpe)之间沉积载体,这导致这两层的分离以容纳注射的溶液。许多血清型在onl和rpe层中显示转导,其中一些血清型显示出受限的趋向性。在天然血清型中,基于其快速转基因表达和所有视网膜层的转导,raav8是sr递送的最佳者之一。除了sr递送之外,可以将载体施用于玻璃体。由于若干原因,玻璃体内递送raav载体已成为达到视网膜下的优选途径,所述原因包括1)注射的技术容易性,2)将载体递送至视网膜更大区域的潜力,以及3)其对视网膜的损害较小。对于临床应用,玻璃体内递送可作为门诊手术进行,并且避免视网膜破裂,所述视网膜破裂可以排除适用于患有严重视网膜变性的患者。然而,很少血清型通过这种途径表现出有效的转导。raav2是在多种动物模型中测试的少数血清型之一,通常导致视网膜神经节细胞(rgc)的转导。在啮齿动物模型中,在偶尔的müller神经胶质细胞、无长突神经细胞和水平细胞中已看到用这种血清型的转导。此外,在啮齿动物模型中已看到rgc和内核层(inl)中的raav6表达。理解病毒输送和有效玻璃体内转导的障碍为合理设计衣壳以克服当前限制的提供了机会。
在玻璃体视网膜连接处,内界膜(ilm)已被暗示为导致大多数raav载体转导视网膜的低效率的屏障。尽管转导有限,但几种aav血清型能够在递送后于玻璃体视网膜连接处累积。将荧光标记的衣壳(raav1、2、5、8和9)注射到成年啮齿动物的玻璃体中显示raav2、raav8和raav9在ilm处累积,但仅raav2导致转导。对于变性的ilm,所有这些aav血清型都能够转导视网膜。ilm由müller神经胶质末端的细胞外基质组成,所述细胞外基质显示与其他基底膜类似的聚糖阵列并且阻止接近aav转导所需的细胞。raav2、raav8和raav9的结合可能通过层粘连蛋白相互作用来解释;然而,层粘连蛋白的累积对于raav8和raav9的转导并不足够。hspg似乎解释了raav2转导,但hspg的酶促消化增加了raav2在视网膜中的转导和穿透。因为raav2在其他组织中显示出hspg非依赖性转导,所以raav2可能不需要hspg结合用于视网膜转导,并且hspg可以防止raav2颗粒扩散到外部视网膜。为此,在ilm处raav2衣壳与hspg的相互作用构成了对视网膜的有效玻璃体内转导的限速步骤,并且理解这些相互作用将有助于指导载体的合理设计以更有效地进行玻璃体内递送。
我们使用自身互补的cbh-gfp盒来优化转基因性能。与cmv或cba启动子活性相比,cbh启动子在其他神经元组织中显示出格外的活性而没有潜在的沉默问题,并且其小尺寸有利于最大化raav的有限转基因能力。转基因的自身互补形式促进更快的表达,其比经典的单链形式更牢固。这种自身互补形式还可以促进在不提供第二链合成的细胞中产生转基因产物。除了优化gfp产生外,我们还使用fish跟踪玻璃体内递送后的raav衣壳,并获得准确的输送图。基因衣壳突变用于理解hs结合在不改变ilm结构的情况下对于小鼠视网膜的raav转导的作用。我们在raav2的hs结合足迹中使用已知的衣壳突变来消融hs结合。raav2衣壳上的基序由三重刺突基底处的基本残基碎片(r484、r487、k532、r585和r588)组成。衣壳突变体如raav2i8置换残基585q和588t以改变趋向性远离富含hs的肝组织,并且当静脉内递送时变得更加全身性。使用这种衣壳,我们研究了hs结合对于视网膜转导的单方面必要性。
载体生产和纯化。通过使用聚乙烯亚胺的三重转染方法产生在普遍存在的cbh启动子控制下携带gfp基因的自身互补raav。收获病毒。将裂解物通过以6200xg离心澄清,并通过以402,000xg碘克沙醇梯度超速离心1小时来纯化。从40%/60%界面取出病毒,通过在1-mlqhyperdf柱(pall)上的离子交换层析纯化,并用200mmnacl,25mmtris[ph9.0]洗脱。通过碘克沙醇难以以显著的产率产生aav8-e533k载体。因此,aav8和aav8-e533k通过cscl纯化,且然后通过蔗糖纯化,以获得纯载体。将病毒针对1xpbs中的350mmnacl,5%山梨糖醇透析,然后等分并在-80℃下冷冻。相对于病毒标准,通过qpcr针对dnase抗性载体基因组的野生型itr测定病毒滴度。病毒在1%bis-tris凝胶(novex)上进行电泳并银染(lifetechnologies)以评估纯度。
动物注射。成年c57bl/6小鼠用于该研究。所有动物在北卡罗来纳大学实验动物医学设施部门的12/12小时光照/黑暗循环中饲养,并根据北卡罗来纳大学的机构动物护理和使用委员会的指南进行处理。在载体递送之前,用氯胺酮(75mg/kg)、甲苯噻嗪(10mg/kg)、乙酰丙嗪(1.5mg/kg)麻醉动物,并用1%托吡卡胺和2.5%苯肾上腺素散瞳。将丙美卡因-hcl作为局部麻醉剂施用于眼睛。使用通过填充有水的管连接到hamilton注射器的32g套管构建眼内针。气泡将水与病毒悬浮液分离。将新鲜解冻的病毒稀释成工作原液,并在注射前在室温下在眼内针中温育10分钟。将针抽空并装入新鲜悬浮液。将病毒悬浮液与荧光素钠盐(sigma)混合以确认注射成功。所有注射均由同一外科医生进行。对于玻璃体内注射,在眼睛的角膜缘后大约0.5mm的上部处,用斜面30g针的尖端制作导孔。在通过显微镜直接观察下将眼内针穿过该孔插入玻璃体内。使用注射泵在30秒内以恒定速率递送1微升的体积。将针保持在适当位置20秒以允许在移除之前进行眼内压平衡。对于视网膜下注射,将眼内针相切于眼睛插入。立即递送液体并通过光学相干断层扫描(oct)和使用microniv(phoenixresearchlaboratories)的眼底镜检查表征成功。将genteal滴眼剂施用于眼睛以防止角膜干燥,并使小鼠在加热垫上恢复。
体内成像。如本文所述,通过散瞳和镇静动物进行眼底成像和oct。使用microniv在相同的设置和类似的视网膜位置拍摄所有荧光图像。分离荧光眼底图像的绿色通道,转换为灰度,并使用imagej软件(nationalinstitutesofhealth)通过积分密度测量进行定量。
摘除术和组织学。将动物镇静并用含有1单位肝素/ml的pbs灌注,然后用pbs中的4%多聚甲醛(pfa)灌注。摘除眼睛,并用18g针头在角膜缘前进行穿刺,然后在4%pfa中温育10分钟。取出前段、肌肉组织和晶状体,并将眼罩置于4℃的10%蔗糖中过夜,然后进行20%和30%蔗糖温育。将眼罩包埋于oct切割介质(sakura)中,在-20℃冷冻,并储存在-80℃。在预清洁的superfrostplus载玻片(fisher)上收集10微米横切片,并储存在-80℃直至进一步加工。
免疫组织化学和分析。将切片在含有0.3%tween-20的tbs(tbs-t)中洗涤,并在封闭缓冲液(10%正常山羊血清,0.1%triton-x100,在pbs中)中在潮湿腔室(humidchamber)中温育1小时。将载玻片在抗体溶液(3%ngs,0.1%triton-x100,在pbs中)与一抗在室温下在潮湿腔室中温育过夜。使用的一抗是兔抗gfp(1:500;millipore)、小鼠抗谷氨酰胺合成酶(1:100;abcam)、小鼠抗硫酸乙酰肝素10e4表位(1:70;amsbio)、小鼠抗视紫红质(1:100;rockland)和小鼠抗pkcα[h-7](1:250;santacruz)。用tbs-t洗涤三次后,将二级荧光抗体在潮湿腔室中施用于抗体溶液中2小时。二抗是alexa-fluor488山羊抗兔(1:1000;molecularprobes)、alexa-fluor568山羊抗小鼠(1:1000;molecularprobes)、或alexafluor568兔抗山羊(1:1000;molecularprobes)。如制造商的方案所述,将载玻片封固在具有dapi(molecularprobes)的prolonggoldantifade中。在leicasp2aobs立式激光扫描共聚焦显微镜或olympusix83荧光显微镜上拍摄图像。
可溶性hs模拟测定。对于体外研究,将hek293细胞以每孔105个细胞的密度铺板在24孔培养皿中,并使其在37℃、5%co2下粘附过夜。将病毒与可溶性肝素以规定浓度预温育1小时,然后添加至细胞,浓度为每个细胞10,000vg。48小时后收获细胞并通过流式细胞术定量。
荧光原位杂交。在hindiii和ecori位点处将gfp基因克隆到pspt18载体(rocherna体外转录试剂盒)中并测序确认。用这些限制酶使质粒线性化,并通过苯酚-氯仿提取/乙醇沉淀纯化,并重悬于水中。线性化的质粒通过分光光度法定量并通过凝胶电泳验证,然后如制造商(roche)所述进行反义和正义核糖核酸探针的体外转录。将等份的核糖核酸探针在水中冷冻,并如制造商所述进行定量,并通过凝胶电泳和sybrgold染色(invitrogen)进行分析。通过病毒对照至带正电荷的硝酸纤维素膜(roche)的斑点印迹针对灵敏度和选择性测定核糖核酸探针功能性。正义和反义探针都能够同等地检测病毒gfp转基因。
将冷冻载玻片加热至55℃保持10分钟并进行预处理。然后将载玻片在杂交缓冲液(50%甲酰胺,10mmtris[ph7.6],200μg/ml酵母trna,1xdenhardt溶液,10%硫酸葡聚糖,600mmnacl,0.25%sds,1mmedta[ph8])中在没有探针的情况下在65℃的杂交温度下温育2-4小时。将载玻片转移到含有50ng/ml正义核糖核酸探针的预杂交缓冲液中,以特异性地检测dna而不是mrna转录物。将载玻片加热至80℃保持20分钟,在冰上快速冷却,并在65℃下温育过夜。将载玻片在50%甲酰胺/2×ssc中在65℃下洗涤30分钟,2×ssc在55℃下洗涤20分钟,且0.2×ssc在55℃下洗涤20分钟2次。将载玻片在1x洗涤缓冲液(roche)中洗涤,然后在潮湿腔室中在1x封闭缓冲液中温育10%绵羊血清1小时。施用绵羊抗dig-ap抗体(1:1000;roche)并在潮湿腔室中温育2-3小时。将载玻片在洗涤缓冲液中洗涤三次,轻轻搅拌10分钟,然后在检测缓冲液(100mmtris,100mmnacl,10mmmgcl2[ph8.0])中两次温育10分钟。按照制造商(roche)所指示制备和施用hnpp/fastred检测底物,用于2-3次施用。在检测反应之后,将载玻片在蒸馏水中漂洗并使用具有dapi的prolonggold抗褪色试剂进行盖玻片。在leicasp2aobs立式激光扫描共聚焦显微镜或olympusix83荧光显微镜上拍摄图像。
离体人视网膜结合测定。死亡后立即采集人的整个眼球,并置于冰上的潮湿室中4天。移除前房、虹膜和晶状体,并将眼球四等分,其中一些玻璃体保持附着于视网膜。将10μl载体以2×109vg/ml的滴度施用于玻璃体中,并使其在4℃下与视网膜结合2小时。将四等分的视网膜保持在介质外,以防止载体溶液的分散。温育后,将pbs对组织进行洗涤,收集并储存在-80℃。分别收集玻璃体、视网膜、脉络膜和巩膜并储存在-80℃。使用dneasyblood&tissue试剂盒(qiagen)消化和纯化组织样品。使用针对gfp和hgapdh管家基因的引物,通过qpcr定量收集的样品中的病毒。
视网膜下转导不需要raav2上结合hs。当在视网膜下递送时,各种aav血清型(hs结合和非hs结合两者)在视网膜转导中有效地起作用。为了确定在外部视网膜的转导中raav2是否需要hs结合,视网膜下递送hs缺陷型raav2i8和raav2。通过眼底镜检查,它们之间的转导是相似的。在分离区域内看到具有raav2i8的gfp荧光的最强信号,但是在水泡区域外可以看到表达。类似地,注射raav2的眼睛显示转导主要发生在分离区域。两种载体都造成大的转导区域,其似乎是rpe。对于raav2,通过从注射部位通向光学头的荧光轴突证实了神经节细胞的转导。
免疫组织化学(ihc)用于评估两种载体的细胞趋向性。rpe和onl是两种载体转导的主要细胞层。可以看到发生高rpe转导但低onl转导的区域,表明rpe可能是待转导的主要细胞类型。对于raav2和raav2i8衣壳两者,onl的转导主要发生在视杆中。rlav2i8在inl中的细胞的转导被鉴定为视杆双极细胞和müller神经胶质细胞。这些结果证实hs缺陷型raav2衣壳通过视网膜下递送在视网膜中具有感染性。
玻璃体内转导需要raav2的hs结合。为了评估hs结合是否是玻璃体内转导所必需的,将aav2和aav2i8衣壳以108vg的滴度递送至成年小鼠。注射raav2的眼睛在两周的第一个成像时间点发荧光,而raav2i8显示没有表达。由于较慢的表达动力学的可能性,评估眼睛长达12周。在该时间期间,raav2荧光继续增加,但用raav2i8未检测到荧光。到12周时,raav2衣壳导致神经视网膜上的荧光扩散模式,如眼底成像所见。raav2i8衣壳通过眼底镜检查未产生可观察到的gfp荧光,并导致gfp荧光减少300倍(图1)。我们通过在视网膜下注射之前混合玻璃体和病毒证实了玻璃体没有抑制hs-消融raav2衣壳的转导。同样,raav2和raav2i8在眼底镜检查中观察到的大部分视网膜中具有强烈表达。两种衣壳的转导细胞似乎均是rpe,但是用raav2可以看到rgc的添加。
通过ihc进一步评估玻璃体内注射raav的眼睛。raav2转导主要在rgc和inl中检测到,并且在一些切片中,可以观察到光感受器的转导。hs-消融的raav2衣壳的组织学揭示极少有gfp阳性视杆,但大多数视网膜保持阴性。我们为raav2及其hs结合突变体尝试了2x109vg的更高滴度,以最大化观察表达的机会。与用低滴度注射的眼所观察到的相比,具有更高滴度的raav2病毒的表达高得多,但转导模式未改变。
荧光原位杂交(fish)用于确定玻璃体内递送后两种衣壳之间的转基因分布。对视网膜下注射病毒的研究已经显示,病毒分布和转基因表达并不同义;因此,我们想确定hs结合如何影响raav2分布而不管表达如何。与ihc表达类似,raav2递送的转基因的fish信号主要在rgc和inl中检测到,其中onl中的转基因较少。注射raav2i8的眼睛显示onl中存在gfp转基因,但在rgc或inl中未检测到。因为组织学在注射后数月进行,所以这些转基因最可能代表通过玻璃体内递送进入视网膜后稳定的附加体。
hs结合对于小鼠中的ilm处raav2的玻璃体累积是必需的。为了更好地理解玻璃体内递送后衣壳之间的输送差异,在注射后不久将眼摘出用于fish分析。因为我们已经确定衣壳表面上带电荷残基的暴露在转导中是重要的,所以我们使用fish作为用荧光颗粒修饰衣壳用于输送实验的替代方案。使用我们的fish方案证实了aav颗粒在注射后24小时在ilm处累积,并且这表明我们可以检测由仍然完整的衣壳所携带的转基因。将时间点延长至注射后3天以允许足够的时间在ilm处的衣壳累积并观察raav2衣壳和其hs结合突变体之间的输送差异。使用一系列剂量来捕获累积中的任何浓度效应,并且缩短fish信号的酶促时间以给出更动态的范围。注射pbs的眼睛作为阴性对照,其具有最小的背景标记。随着检测时间的缩短,对于raav2和hs缺陷型raav2-r585e衣壳两者,1x108vg的剂量仅在视网膜中具有弱信号。在5×108vg的剂量下,由raav2递送的转基因显示在ilm处的累积,并且存在于所有视网膜层中。没有hs结合的情况下,raav2-r585e仅具有最小信号。在2x109vg的测试的最高剂量下,raav2导致在ilm处甚至更大的信号强度,其中在多个视网膜层中检测到散发性转基因。在相同剂量下,在视网膜中检测到很少的raav2-r585e递送的转基因,但是在ilm处没有导致任何累积。
总之,这些结果表明1)raav2上的hs结合有助于在ilm处累积载体,2)这种累积增加了驻留在视网膜中的转基因的数量,3)衣壳可以从玻璃体穿透视网膜而不用结合hs,但程度要小得多,和4)当载体被视网膜下递送时,视网膜的raav2转导不需要hs结合。fish数据表明,在快速通过ilm屏障的衣壳中,它们似乎能够快速输送到外部视网膜的远端层(图2)。这突出了raav对视网膜的有效玻璃体内转导的限速步骤在于衣壳和ilm之间的相互作用。
hs结合对于raav2在人视网膜上的玻璃体累积是必需的。ilm处丰富的hspg染色存在于包括人类的许多动物模型中。这表明该机制可以跨物种转化用于人类临床应用。通过将眼四等分并留下少量玻璃体附着于视网膜以维持ilm,对离体人视网膜进行病毒结合测定。将载体施用于玻璃体中,且然后收获各种视网膜层。与raav2i8衣壳的那些不同,raav2所携带的转基因与视网膜结合。hs缺陷型raav2i8在任何收集的组织中均具有相对低的载体结合,但与raav2相比确实显示出与脉络膜和巩膜的结合的显著增加(图3)。结合小鼠数据,这些结果佐证了hs结合促进aav载体离开玻璃体累积到ilm上并随后用于增强视网膜中的转导的机制。
hs结合增加了其他raav血清型的玻璃体内转导。其他血清型的玻璃体内转导可受益于hs结合基序的添加并在小鼠中进行了该选择。raav1和raav6血清型仅相差6个氨基酸,其中单个残基负责其hspg结合的差异。为了评估raav1和raav6视网膜转导之间的hs结合对玻璃体内转导的影响,玻璃体内测试了单残基突变体衣壳。尽管raav1和hs结合raav1-e531k具有相似的表达模式,但与raav1相比,raav1-e531k具有3倍更高的gfp荧光(图4)。使用raav6-k531e衣壳去除raav6中的hs结合导致通过眼底镜检查的视网膜荧光减少。raav1和raav6衣壳两者均在视网膜血管周围显示点状表达模式。由于raav1和raav6之间的同源性,仅进一步评估raav1和raav1-e531k的细胞趋向性的可能差异。免疫组织化学通过gfp和谷氨酰胺合成酶的共定位显示两种衣壳的主要müller胶质细胞的转导,尽管inl的额外细胞似乎被转导。此外,raav1和raav1-e531k两者均显示少数rgc和光感受器的转导。raav1和raav1-e531k的相似的转导模式表明hs结合并未改变raav1的趋向性。
为了证实raav1上的hs结合突变不传达hspg用于转导的用途,将可溶性肝素与衣壳混合并施用于细胞进行体外竞争测定。raav2需要hspg用于体外转导并且显示出转导的剂量依赖性降低。在任何肝素剂量下,raav1和raav1-e531k转导均未受影响,表明raav1-e531k衣壳不依赖于hs结合进行转导(图6)。平均而言,相比于raav1,用raav1-e531k转导导致更少的gfp阳性细胞,表明单独的单个氨基酸改变不提供转导的增强。已经在raav8上鉴定了单个氨基酸突变体,其提供hs结合能力。将raav8和raav8-e533k的滴度匹配至1x108vg并玻璃体内注射。在施用后8周,对注射眼睛拍摄眼底图像。raav8的转导非常低。注射raav8-e533k的眼睛在视网膜上产生模糊的荧光,当定量时,其与非结合血清型相比更高。我们使用fish分析观察注射后不久的输送差异,并再次发现hs结合促进了ilm处和视网膜内的载体累积。这些结果表明单独的hs结合足以通过增加累积到视网膜上的载体的量来增强玻璃体内递送的aav衣壳的转导。
使用raav2.5g9衣壳的双嵌合体来确定raav1或raav9衣壳元件的哪一种是优势的。当通过眼底镜检查成像时,raav2.5g9的玻璃体内递送导致表达类似于raav2.5亲本。视网膜血管周围的荧光非常明显,其中在血管周围发现点状表达。当全部在一起比较时,该表达的定量显示raav2.5和raav2.5g9具有最高的转导和显示出最低转导的非hs结合衣壳(图5)。
总之,在ilm处与hspg的结合促进raav从玻璃体累积到视网膜上,但来自其他非hs结合衣壳的基序可用于影响视网膜转导模式。这将解释raav1-e531k如何维持raav1的转导概况,尽管具有hs结合。将raav2与raav1-e531k精确比较将是困难的,因为这两种衣壳的残基组成和对hs的亲和力不同。因此,为了评估raav1可对玻璃体内转导具有的影响,我们使用了raav2.5嵌合衣壳。该衣壳显示出比任一亲本更大的转导,并且可以通过添加半乳糖结合而偏向于转导müller神经胶质细胞。尽管视网膜聚糖染色不容易表明趋向性,但这些嵌合衣壳是用于视网膜中的靶向转导的试剂。由于hspg在数种动物模型中的ilm处是丰富的,因此该机制可跨多个物种(包括人类)应用。一旦在ilm处,其他聚糖与raav衣壳的相互作用促进了用各种衣壳突变体观察到的趋向性概况。
raav衣壳与ilm的相互作用为视网膜输送提供了限速步骤。通过使用fish,少量衣壳通过ilm并快速输送至onl和外部区段。一旦在外部视网膜中,这些载体可以转导光感受器,但这种转导非常罕见。也许进入视网膜的大多数衣壳在细胞中一次也不能成功地输送至细胞核以建立潜伏。增加病毒浓度更可能引发免疫反应,因为玻璃体空间不像视网膜下空间那样具有免疫特权。通过使用更高的滴度可以增加使其进入细胞核的转基因的数量。这种增强的细胞内输送可能有助于增加玻璃体内递送的raav2i8和raav2-r585e衣壳的转基因表达。ilm处的累积可以是载体浓度的函数,因为高剂量的载体可导致在ilm处和视网膜中发现的转基因增加。为了利用视网膜结构,可以使用与ilm的结合在视网膜处累积载体用于转导。这可能是raav2成功进行玻璃体内视网膜转导的原因。
带电荷的硫酸基团促进hspg与raav2衣壳的相互作用以进行转导。通过fish观察到的转基因表明hs缺陷型raav2颗粒能够向视网膜的远端层输送,这表明衣壳上的这些带电荷残基变化不会防止载体进入视网膜,而只是限制载体累积在视网膜上的数量。这也通过用hs缺陷衣壳对视杆的罕见转导得到证明。为了测试衣壳与硫酸的相互作用是否必要,我们将raav2与硫酸化和未硫酸化的hs混合并进行玻璃体内测试。我们发现这两种形式均可以抑制硫酸化肝素的转导,导致更大的抑制。该实验仅表明与乙酰肝素链相互作用对玻璃体内转导的重要性。遗传衣壳突变体似乎验证了这种化学抑制测定,但再次,破坏hspg结合中涉及的基本残基碎片的单和双氨基酸修饰依赖于电荷相互作用。涂覆有特定带电荷制剂的纳米颗粒表明阳离子电荷(碱性)在促进从玻璃体渗透到视网膜中是成功的。这可能有助于解释onl中存在的聚糖如何可以影响玻璃体内递送的raav载体的转导。
sr递送的raav2和raav2i8的相似转导概况表明,raav2不需要hs结合用于视网膜转导。可能是视网膜下递送有效地浓缩载体,从而化学计量地使衣壳朝向表达倾斜。除了通过受体介导的内吞作用的转导之外,sr还为吞噬性rpe提供了丰富的raav载体,并且可以解释为何rpe似乎是raav2和raav2i8两者的主要细胞靶标。无论对任何特定细胞类型的亲和力如何,raav2和raav2i8载体导致rpe、视杆细胞、视锥细胞、视杆双极细胞和müller神经胶质细胞的转导。这些转导细胞中的大多数位于注射水泡内,但可以在分离区域之外远处看到rpe的转导。fish可用于绘制sr递送的载体的输送。
跨多个物种发现ilm结构,并且其可用于从玻璃体吸引和浓缩raav衣壳。实际上,当注射后不久通过fish评估时,与亲本衣壳相比,hs结合导致视网膜中转基因的存在更多。仍然可以在视网膜中检测到非hs结合的raav1和raav8的转基因,但程度较小,类似于在raav2和raav2-r585e衣壳之间观察到的数据。由于该血清型在大多数细胞类型的转导中效率低,并且可能遇到细胞有效转导的额外障碍,因此预期在玻璃体内注射时用hs结合raav3的表达缺乏。raav6是视网膜转导目标血清型,并且已被修饰以增加使用shh10衣壳的müller神经胶质细胞的特异性转导。在没有hs结合的情况下,shh10载体可能揭示弱得多的荧光。因为raav6和raav1-e531k两者均不依赖于hspg介导的转导,所以对任意衣壳血清型简单地添加结合hs的能力可以增强其转基因表达。我们通过使用raav8-e533k突变体衣壳测试了这一点。hs结合的添加可以应用于其他血清型,用于玻璃体内递送所用的aav的宽度更大。
尽管hspg在ilm处是丰富的,但是其他受体可以在从玻璃体转导视网膜中起作用。raav1和raav6的转导表明在ilm处存在2,3-或2,6-n-连接的唾液酸,尽管在该区域缺乏染色。通过眼底观察到的转导模式可以指示通过组织学并不可见的视网膜中的这种唾液酸的独特模式,其通过铺片(flatmount)可见。已知与raav4和raav5相互作用的其他形式的唾液酸可能不会在ilm处大量表达或可能被其他聚糖掩盖。这将解释当在正常小鼠视网膜中比较时通过这些血清型的转导的缺乏。除唾液酸外,层粘连蛋白染色在ilm处丰富并且限于血管。已知层粘连蛋白受体与raav2、raav3、raav8和raav9血清型相互作用。尽管这些衣壳可以与玻璃体视网膜连接处的层粘连蛋白受体相互作用,但这种相互作用似乎不足以促进有效的玻璃体内视网膜转导。
重要的是记住,在小鼠模型中观察到的转导可能不指示其他模型。虽然小鼠已成为视网膜基因转移的标准模型,但它们和灵长类动物之间存在一定的大小和解剖学的差异。与灵长类动物相比,其他模型如兔子或猪,具有相似大小的眼球和玻璃体体积。此外,小鼠和灵长类动物之间ilm的厚度差异可能导致选择跨物种不有效的衣壳。在众多动物物种(包括禽类、啮齿动物、兔、灵长类动物和人)中ilm处的过度hspg富集使得研究衣壳与hs相互作用的影响对于玻璃体内视网膜基因转移的有效载体的合理设计是重要的。
前述内容说明本发明,而不应解释为限制本发明。本发明由以下权利要求所限定,权利要求的等同物也包括在其中。
本文提及的所有出版物、专利申请、专利、序列和其他参考文献均通过引用全文并入本文。
- 还没有人留言评论。精彩留言会获得点赞!