WOOD-LJUNGDAHL微生物的基因敲除的制作方法
相关申请的交叉引用本申请要求于2017年9月28日提交的美国临时专利申请第62/565,000号的权益,所述美国临时专利申请全部内容通过引用并入本文。
背景技术:
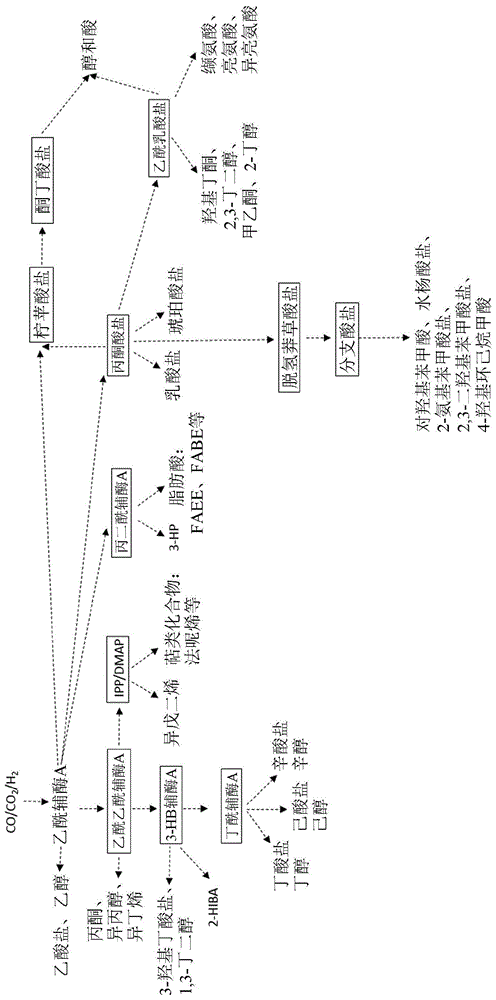
:早已认识到催化工艺(如费托工艺(fischer-tropschprocess))可以用于将含有二氧化碳(co2)、一氧化碳(co)和/或氢气(h2)的气体(例如工业废气或合成气)转化成各种燃料和化学品。然而,最近,气体发酵已经成为用于对此类气体进行生物固定的替代平台。特别是,已经证明c1固定微生物将含有co2、co和/或h2的气体转化成如乙醇和2,3-丁二醇的产物。然而,这些产物的有效生产可能受到微生物生长缓慢、气体吸收有限、对毒素敏感或碳底物转化为不需要的副产物的限制。因此,仍然需要特性改进的经基因工程化的微生物。附图说明图1是显示在wood-ljungdahl微生物中的关键生产途径和关键代谢节点(用方框表示)的图。通过这些节点提高碳通量(例如通过破坏某些基因的表达)能够提高下游产物的产量。具体实施方式本发明提供了包括至少一种被破坏的基因的非天然存在的微生物。在本发明的微生物中,碳通量被策略性地从非必需或不期望的产物转向关注的产物。在某些实施方案中,这些被破坏的基因使碳通量从非必需或不期望的代谢节点转移并转移通过目标代谢节点,以提高那些目标代谢节点下游的产物产量。本发明的微生物来源于亲本细菌,如伍氏醋酸杆菌(acetobacteriumwoodii)、巴氏嗜喊菌(alkalibaculumbacchii)、黏液真杆菌(blautiaproducta)、食甲基丁酸杆菌(butyribacteriummethylotrophicum)、醋酸梭菌(clostridiumaceticum)、产乙醇梭菌(clostridiumautoethanogenum)、食一氧化碳梭菌(clostridiumcarboxidivorans)、克氏梭菌(clostridiumcoskatii)、德氏梭菌(clostridiumdrakei)、乙酸醋酸梭菌(clostridiumformicoaceticum)、永达尔梭菌(clostridiumljungdahlii)、大梭菌(clostridiummagnum)、拉氏梭菌(clostridiumragsdalei)、粪味梭菌(clostridiumscatologenes)、粘液真杆菌(eubacteriumlimosum)、热自养穆尔氏菌(moorellathermautotrophica)、热醋穆尔氏菌(moorellathermoacetica)、普氏产醋杆菌(oxobacterpfennigii)、卵形鼠孢菌(sporomusaovata)、森林土壤醋酸鼠孢菌(sporomusasilvacetica)、球形鼠孢菌(sporomusasphaeroides)和凯伍热厌氧菌(thermoanaerobacterkiuvi)。在一个优选实施例中,所述亲本细菌是产乙醇梭菌、永达尔梭菌或拉氏梭菌。在一个特别优选的实施例中,亲本细菌是产乙醇梭菌。在一个实施例中,本发明提供了一种非天然存在的wood-ljungdahl细菌,所述wood-ljungdahl细菌包括异源性硫解酶和编码例如nad依赖性电子分叉[fefe]-氢化酶、谷氨酸盐合酶、柠苹酸合酶、乙酰乳酸盐脱羧酶、乳酸盐脱氢酶、乙酸盐激酶、磷酸转乙酰酶和乙醛脱氢酶中的一种或多种的一种或多种基因中的破坏性突变,其中与亲本细菌相比,所述非天然存在的细菌已经通过乙酰乙酰辅酶a提高碳通量。具体而言,与所述亲本细菌相比,所述一种或多种基因的表达减少或消除。这样的实施例中,所述非天然存在的细菌可产生产物,如丙酮、异丙醇、3-羟基异戊酰辅酶a、3-羟基异戊酸盐、异丁烯、异戊烯基焦磷酸盐、焦磷酸二甲基烯丙基焦磷酸盐、异戊二烯、法呢烯、3-羟基丁酰基辅酶a、巴豆酰基辅酶a、3-羟基丁酸盐、3-羟基丁醛、1,3-丁二醇、2-羟基异丁酰辅酶a、2-羟基异丁酸盐、丁酰辅酶a、丁酸盐、丁醇、己酸盐、己醇、辛酸盐、辛醇、1,3-己二醇、2-丁烯-1-醇、异戊酰辅酶a、异戊酸盐或异戊醇。例如,当亲本细菌是产乙醇梭菌时,(a)所述nad依赖性电子分叉[fefe]氢化酶可以选自由以下组成的组:caethg_1576、caethg_1578、caethg_3569、caethg_3570和caethg_3571,(b)所述谷氨酸合酶可以选自由以下组成的组:caethg_0477、caethg_1580、caethg_3850和caethg_3851,(c)所述柠苹酸合酶可以是caethg_2751,(d)所述乙酰乳酸脱羧酶可以是caethg_2932,(e)所述乳酸脱氢酶可以是caethg_1147;(f)所述乙酸激酶可以是caethg_3359,(g)所述磷酸转乙酰酶可以是caethg_3358,或者(h)所述乙醛脱氢酶可以选自由以下组成的组:caethg_1819,caethg_3287和caethg_1830。在另一个实施例中,本发明提供了一种非天然存在的wood-ljungdahl细菌,所述wood-ljungdahl细菌包括一种或多种基因中的破坏性突变,其中与亲本细菌相比,所述非天然存在的细菌已经通过分支酸盐提高碳通量。所述一种或多种基因编码例如以下中的一种或多种:嘌呤核苷磷酸化酶、乳酸通透酶、胱硫醚γ-裂解酶、腺嘌呤磷酸核糖基转移酶、5'-核苷酸酶/3'-核苷酸酶/外切聚磷酸酶、小电导机械敏感通道、精氨酸脱亚胺酶、ll-二氨基庚二酸氨基转移酶脱辅基酶和磷酸戊糖变位酶。具体而言,与所述亲本细菌相比,所述一种或多种基因的表达减少或消除。在这样的实施例中,所述非天然存在的细菌可以产生产物,如分支酸盐、对羟基苯甲酸、水杨酸盐、2-氨基苯甲酸盐、二羟基苯甲酸盐、4-羟基环己烷甲酸以及其盐和离子。例如,当所述亲本细菌是产乙醇梭菌时,所述一种或多种基因可以编码以下中的一种或多种:caethg_0160、caethg_0248、caethg_0498、caethg_1270、caethg_1371、caethg_2107、caethg_3021、caethg_3510和caethg_3924;当所述亲本细菌是永达尔梭菌时,所述一种或多种基因可以编码以下中的一种或多种:clju_c20750、clju_c21610、clju_c24380、clju_c33720、clju_c34740、clju_c42810、clju_c09270、clju_c14280和clju_c18150;并且当所述亲本细菌是拉氏梭菌,所述一种或多种基因可以编码以下中的一种或多种:clrag_19250、clrag_31200、clrag_25120、clrag_24560、clrag_14800、clrag_25620、clrag_09600或clrag_00520。本发明还提供了通过在底物(如包括co、co2和/或h2中的一种或多种的气体底物)存在下培养本发明的微生物来生产产物的方法。当用于提及微生物时,术语“非天然存在的”是指微生物具有至少一种未在提及物种的天然存在的菌株(包含所述提及物种的野生型菌株)中发现的遗传修饰。非天然存在的微生物通常在实验室或研究机构中培养。术语“遗传修饰”、“遗传改变”或“遗传工程化”广义上指人类对微生物基因组或核酸的操作。同样,术语“经基因修饰的”、“经基因改变的”或“经基因工程化的”是指含有这种基因修饰、基因改变或基因工程化的微生物。这些术语可用于区分实验室产生的微生物和自然产生的微生物。遗传修饰的方法包含例如异源性基因表达、基因或启动子插入或缺失、核酸突变、经改变基因表达或失活、酶工程化、定向进化、基于知识的设计、随机诱变方法、基因改组和密码子优化。“重组”表示核酸、蛋白质或微生物是遗传修饰、工程化或重组的产物。一般而言,术语“重组体”是指含有来自多种来源(如两种或多种不同的微生物菌株或物种)的遗传物质或由其编码的核酸、蛋白质或微生物。“野生型”指区别于突变体形式或变体形式的自然出现的有机体、菌株、基因或特征的典型形式。“内源性”是指存在于或表达于本发明微生物来源的野生型或亲本微生物中的核酸或蛋白质。例如,内源性基因是天然存在于本发明微生物来源的野生型或亲本微生物中的基因。在一个实施例中,内源性基因的表达可以由外源性调节元件(如外源性启动子)控制。“外源性”是指来源于本发明微生物之外的核酸或蛋白质。例如,外源性基因或酶可以通过人工或重组方式产生,并引入本发明的微生物中或在本发明的微生物中表达。外源性基因或酶也可以从异源性微生物中分离出来,并引入本发明的微生物或在本发明的微生物中表达。外源性核酸可以适于整合到本发明微生物的基因组中,或者在本发明微生物中保持染色体外状态,例如在质粒中。“异源性”是指来源于不同菌株或物种并被引入本发明的微生物中或在本发明的微生物中表达的核酸或蛋白质。术语“多核苷酸”、“核苷酸”、“核苷酸序列”、“核酸”和“寡核苷酸”可互换使用。这些术语指任何长度的核苷酸的聚合形式,包含脱氧核糖核苷酸或核糖核苷酸或其类似物。多核苷酸可以具有任何三维结构,并且可以执行任何已知或未知的功能。以下是多核苷酸的非限制性实例:基因或基因片段的编码或非编码区、由连锁分析定义的基因座(基因座)、外显子、内含子、信使rna(mrna)、转移rna、核糖体rna、短干扰rna(sirna)、短发夹rna(shrna)、微-rna(mirna)、核酶、cdna、重组多核苷酸、分支多核苷酸、质粒、载体、任何序列的分离dna、任何序列的分离rna、核酸探针和引物。多核苷酸可以包括一个或多个修饰的核苷酸,如甲基化核苷酸或核苷酸类似物。如果存在,对核苷酸结构的修饰可以在聚合物装配之前或之后进行。核苷酸序列可以被非核苷酸组分中断。多核苷酸可以在聚合后进一步进行修饰,如通过与标记组分缀合来修饰。如本文所用的,“表达”是指多核苷酸从dna模板转录的过程(如转录为mrna或其它rna转录物)和/或经转录mrna随后被转译成肽、多肽或蛋白质的过程。转录物和编码的多肽可以统称为“基因产物”。术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,是指任何长度的氨基酸聚合物。聚合物可以是直链或支链的,其可以包括经修饰氨基酸,并且可以被非氨基酸中断。该术语还包括已修饰的氨基酸聚合物;例如二硫键形成、糖基化、脂质化、乙酰化、磷酸化或任何其它操作,如与标记成分的缀合。如本文所用的,术语“氨基酸”包含天然和/或非天然或合成的氨基酸,包含甘氨酸和d或l光学异构体两者,以及氨基酸类似物和肽模拟物。“酶活性”,或简称“活性”,泛指酶活性,包含但不限于酶的活性、酶的量或酶催化反应的可用性。因此,“增加”酶活性包含增加酶的活性、增加酶的量或增加酶催化反应的可用性。类似地,“降低”酶活性包括降低酶的活性、降低酶的量或降低酶催化反应的可用性。“突变的”是指与衍生本发明微生物的野生型或亲本微生物相比在本发明微生物中已修饰的核酸或蛋白质。在一个实施例中,突变可以是编码酶的基因中的缺失、插入或取代。在另一个实施例中,突变可以是酶中一个或多个氨基酸的缺失、插入或取代。“被破坏的基因”是指以某种方式被修饰以减少或消除基因表达、基因调节活性或经编码蛋白或酶活性的基因。破坏可以部分灭活、完全灭活或删除基因或酶。破坏可以是完全消除基因、蛋白质或酶的表达或活性的敲除(ko)突变。这种破坏也可能是降低但不完全消除基因、蛋白质或酶的表达或活性的敲除。破坏可以是减少、阻止或阻断酶产生的产物的生物合成的任何东西。所述破坏可以包含例如编码蛋白质或酶的基因中的突变、参与编码酶的基因表达的基因调节元件中的突变、引入产生降低或抑制酶活性的蛋白质的核酸、或引入抑制蛋白质或酶表达的核酸(例如,反义核rna、rnai、talen、sirna、crispr或crispri)。可以使用本领结构域已知的任何方法引入破坏。为了实现本发明的目的,破坏是实验室产生的,而不是自然发生的。“密码子优化”是指核酸(如基因)的突变,用于优化或改进特定菌株或物种中核酸的转译。密码子优化可能产生更快的转译速度或更高的转译准确度。在一个优选的实施方案中,本发明的基因经密码子优化以在梭状芽孢杆菌中(特别是在产乙醇梭菌、永达尔梭菌或拉氏梭菌中)表达。在进一步优选的实施例中,本发明的基因经密码子优化以在产乙醇梭菌lz1561(其保藏在dsmz保藏号dsm23693下)中表达。“过表达”是指与衍生本发明微生物的野生型或亲本微生物相比本发明微生物中核酸或蛋白质的表达增加。过表达可以通过本领结构域已知的任何方式实现,包含改变基因拷贝数、基因转录速率、基因转译速率或酶降解速率。术语“变体”包含其序列不同于参考核酸和蛋白质序列(如现有技术中公开的或本文举例说明的参考核酸和蛋白质序列)的核酸和蛋白质。本发明可以使用执行与参考核酸或蛋白质基本相同的功能的变体核酸或蛋白质实践。例如,变体蛋白质可以执行与参考蛋白质基本相同的功能或催化与参考蛋白质基本相同的反应。变体基因可以编码与参考基因相同或基本相同的蛋白质。变体启动子可以具有与参考启动子基本相同的促进一种或多种基因表达的能力。此类核酸或蛋白质在本文中可称为“功能等同的变体”。举例来说,核酸的功能等同的变体可以包含等位基因变体、基因片段、突变基因、多态性等。来自其它微生物的同源基因也是功能等同变体的实例。这些基因包含如丙酮丁醇梭菌(clostridiumacetobutylicum)、贝氏梭菌(clostridiumbeijerinckii)或永达尔梭菌(clostridiumljungdahlii)等物种的同源基因,其详细信息可在如genbank或ncbi等网站上公开获得。功能等同的变体还包含核酸,其序列因特定微生物的密码子优化而变化。核酸的功能等同变体优选与参考核酸的核酸序列同一性(同源性百分比)为至少约70%、约80%、约85%、约90%、约95%、约98%或更高。蛋白质的功能等同变体优选与氨基酸同一性(同源性百分比)为参考蛋白质具有至少约70%、约80%、约85%、约90%、约95%、约98%或更高。变体核酸或蛋白质的功能等同物可以使用本领结构域已知的任何方法评估。“互补性”是指一个核酸通过传统的沃森-克里克(watson-crick)或其它非传统类型与另一个核酸序列形成一个或多个氢键的能力。互补性百分比表示核酸分子中能够与第二个核酸序列形成氢键(例如沃森-克里克碱基配对)的残基的百分比(例如,5/10、6/10、7/10、8/10、9/10、10/10是50%、60%、70%、80%、90%和100%互补)。“完全互补”是指核酸序列的所有连续残基将与第二核酸序列中相同数量的连续残基氢键结合。如本文所用的,“基本互补”是指在8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、30、35、40、45、50、或更多个核甘酸的区结构域内互补程度至少为60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%或100%,或指在严格条件下杂交的两种核酸。“杂交”是指一种一个或多个多核苷酸反应形成通过在核苷酸残基碱基之间进行氢键结合稳定的复合物的反应。氢键结合可以通过沃森克里克碱基配对(watsoncrickbasepairing)、胡格斯坦结合(hoogsteinbinding)或任何其它序列特异性方式发生。复合物可以包括形成双链结构的两条链、形成多链复合物的三条或多条链、单个自杂交链或这些的任意组合。杂交反应可以构成更广泛过程(如启动pcr,或用酶切割多核苷酸)中的一个步骤。能够与给定序列杂交的序列被称为给定序列的“互补序列”。可以将核酸使用本领结构域已知的任何方法递送到本发明的微生物。例如,核酸可以作为裸核酸递送,或者可以用一种或多种试剂(如脂质体)配制。适当时,核酸可以是dna、rna、cdna或其组合。在某些实施例中可以使用限制性抑制剂。另外的载体可以包含质粒、病毒、噬菌体、粘粒和人工染色体。在一个优选的实施例中,使用质粒将核酸递送到本发明的微生物。举例来说,转化(包含转导或转染)可以通过电穿孔、超声波处理、聚乙二醇介导的转化、化学或天然感受态、原生质体转化、前噬菌体诱导或缀合来实现。在某些具有活性限制性酶系统的实施例中,在将核酸引入微生物之前,可能有必要将核酸甲基化。此外,核酸可被设计成包括调节元件(如启动子),以增加或控制特定核酸的表达。启动子可以是组成型启动子或诱导型启动子。理想情况下,启动子是wood-ljungdahl途径启动子、铁氧还蛋白启动子、丙酮酸盐:铁氧还蛋白氧化还原酶启动子、rnf复合操纵子启动子,atp合酶操纵子启动子或磷酸转乙酰酶/乙酸盐激酶操纵子启动子。“微生物”是一种微小的有机体,尤其是细菌、太古代、病毒或真菌。本发明的微生物通常是细菌。在本文中,表述“微生物”应该被理解为包括“细菌”。“亲本微生物”是用于产生本发明的微生物的微生物。亲本微生物可以是天然存在的微生物(即野生型微生物)或先前已经修饰的微生物(即突变体微生物或重组微生物)。本发明的微生物可以被修饰以表达或过表达一种或多种在亲本微生物中未表达或过表达的酶。类似地,本发明的微生物可以被修饰以含有亲本微生物不包含的一种或多种基因。本发明的微生物也可以被修m饰以不表达或表达在亲本微生物中表达的较低量的一种或多种酶。在一个实施例中,亲本微生物是产乙醇梭菌、永达尔梭菌或拉氏梭菌。在一个优选实施例中,亲本微生物是产自乙醇梭菌(clostridiumautoethanogenum)lz1561,其于2010年6月7日根据《布达佩斯条约(budapesttreaty)》的条款保藏在德国布伦瑞克省d-38124inhoffenstraβ7b的deutschesammlungvonmikroorganismenundzellkulturengmbh(dsmz),保藏号为dsm23693。国际专利申请第pct/nz2011/000144号(其被公开为wo2012/015317)中对该菌株进行了描述。术语“来源于”表示核酸、蛋白质或微生物从不同的(如亲本或野生型)核酸、蛋白质或微生物修饰或改造,从而产生新的核酸、蛋白质或微生物。这种修饰或改造通常包含核酸或基因的插入、缺失、突变或替换。通常,本发明的微生物衍生自亲本微生物。在一个实施例中,本发明的微生物衍生自产乙醇梭菌、永达尔梭菌或拉氏梭菌。在优选实施例中,本发明的微生物衍生自产乙醇梭菌lz1561(其保藏在dsmz保藏号dsm23693下)中表达。本发明的微生物可以根据功能特性进一步分类。例如,本发明的微生物可以是或可以衍生自c1固定微生物、厌氧菌、产乙酸菌、产乙酸菌、产羧酸菌和/或甲烷氧化菌。表1提供了微生物的代表性列表,并标识了其功能特性。1木质纤维素杆菌(acetobacteriumwoodi)可以从果糖中产生乙醇,但不能从气体中产生乙醇。2尚未研究马氏梭菌(clostridiummagnum)是否可以依靠co生长。3已报道,穆尔氏菌属热醋穆尔氏菌(moorellathermoacetica)中的一种菌株—moorellasp.huc22-1能够从气体中产生乙醇。4尚未研究卵形鼠孢菌(sporomusaovata)是否可以依靠co生长。5尚未研究森林土壤醋酸鼠孢菌(sporomusasilvacetica)是否可以依靠co生长。6尚未研究球形鼠孢菌(sporomusasphaeroides)是否可以依靠co生长。“wood-ljungdahl”是指如例如ragsdale,《生化与生物物理学报(biochimbiophysacta)》,1784:1873-1898,2008所述的wood-ljungdahl固碳途径。“wood-ljungdahl微生物”可以预见地是指含有wood-ljungdahl途径的微生物。本发明的微生物是一种wood-ljungdahl微生物,通常是一种wood-ljungdahl细菌。一般来说,本发明的微生物含有天然的wood-ljungdahl途径。在本文中,wood-ljungdahl途径可以是天然的、未经修饰的wood-ljungdahl途径,或者可以是具有一定程度的遗传修饰(例如,过表达、异源表达、敲除等)的wood-ljungdahl途径途径,只要其仍具有将co、co2和/或h2转化为乙酰辅酶a的功能。“c1”指一个碳分子,例如co、co2、ch4或ch3oh。“c1含氧有机物”是指也包括至少一个氧原子的单碳分子,例如co、co2或ch3oh。“c1碳源”是指作为本发明微生物的部分或唯一碳源的一个碳分子。例如,c1碳源可以包括以下中的一种或多种:co、co2、ch4、ch3oh或ch2o2。优选地,c1碳源包括co和co2中的一种或两种。“c1固定微生物”是指能够从c1-碳源生产一种或多种产物的微生物。典型地,本发明的微生物是c1固定细菌。在一个优选的实施例中,本发明的微生物衍生自表1中标识的c1固定微生物。“厌氧菌”是一种生长不需要氧的微生物。如果氧含量超过某一阈值,厌氧菌可能会产生负面反应,甚至死亡。然而,一些厌氧菌能够耐受低水平的氧(例如,0.000001-5%的氧)。通常,本发明的微生物是厌氧菌。在一个优选的实施例中,本发明的微生物衍生自表1中标识的厌氧菌。“乙酰原”是一种微生物t,它产生或能够产生作为厌氧呼吸的产物的乙酸盐(或乙酸)。通常,乙酰原是专性厌氧菌,其使用wood-ljungdahl途径作为其能量保存和合成乙酰辅酶a和乙酰辅酶a衍生产物(如乙酸盐)的主要机制(ragsdale,《生化与生物物理学报》,1784:1873-1898,2008)。乙酰原在细胞碳合成中使用乙酰辅酶a途径作为(1)从co2还原合成乙酰辅酶a的机制,(2)末端电子接受和能量保存过程,(3)co2固定(同化)的机制(drake,“乙酰原原核生物(acetogenicprokaryotes)”,见:《原核生物(theprokaryotes)》,第3版,第354页,纽约,纽约州,2006)。所有天然存在的产乙酸菌都是c1固定型、厌氧型、自养型和非甲烷营养型。通常,本发明的微生物是乙酰原。在一个优选的实施例中,本发明的微生物衍生自表1中标识的乙酰原。“乙醇原”是一种产生或能够产生乙醇的微生物。通常,本发明的微生物是乙醇。在一个优选的实施方案中,本发明的微生物衍生自表1中标识的乙醇。“自养菌”是一种能够在没有有机碳的情况下生长的微生物。相反,自养菌使用无机碳源,如co和/或co2。通常,本发明的微生物是自养菌。在一个优选的实施方案中,本发明的微生物衍生自表1中标识的自养菌。“产羧酸菌”是一种能够利用co作为唯一碳源和能源的微生物。通常,本发明的微生物是一种产羧酸菌。在一个优选的实施例中,本发明的微生物衍生自表1中标识的产羧酸菌。“甲烷氧化菌”是一种能够利用甲烷作为唯一碳源和能源的微生物。在某些实施例中,本发明的微生物是甲烷氧化菌或衍生自甲烷氧化菌。在其它实施例中,本发明的微生物不是甲烷氧化菌或不是衍生自甲烷氧化菌。更广泛地说,本发明的微生物可以衍生自表1中标识的任何属或种。例如,所述微生物可以是选自由以下组成的组的成员:醋酸菌属(acetobacterium)、碱杆菌属(alkalibaculum)、布拉氏菌属(blautia)、丁酸菌属(butyribacterium)、梭菌属(clostridium)、真细菌属(eubacterium)、摩尔氏菌属(moorella)、氧代杆菌属(oxobacter)、孢子菌属(sporomusa)和嗜热厌氧菌属(thermoanaerobacter)。特别地,微生物可以衍生自选自由以下组成的组的亲本细菌:伍氏醋酸杆菌(acetobacteriumwoodii)、巴氏嗜喊菌(alkalibaculumbacchii)、黏液真杆菌、食甲基丁酸杆菌(butyribacteriummethylotrophicum)、醋酸梭菌(clostridiumaceticum)、产乙醇梭菌(clostridiumautoethanogenum)、食一氧化碳梭菌(clostridiumcarboxidivorans)、克氏梭菌(clostridiumcoskatii)、德氏梭菌(clostridiumdrakei)、乙酸醋酸梭菌(clostridiumformicoaceticum)、永达尔梭菌(clostridiumljungdahlii)、马氏梭菌(clostridiummagnum)、拉氏梭菌(clostridiumragsdalei)、粪味梭菌(clostridiumscatologenes)、粘液真杆菌(eubacteriumlimosum)、热自养穆尔氏菌(moorellathermautotrophica)、热醋穆尔氏菌(moorellathermoacetica)、普氏产醋杆菌(oxobacterpfennigii)、卵形鼠孢菌(sporomusaovata)、森林土壤醋酸鼠孢菌(sporomusasilvacetica)、球形鼠孢菌(sporomusasphaeroides)和凯伍热厌氧菌(thermoanaerobacterkiuvi)。在一个优选实施例中,本发明的微生物衍生自梭状芽胞杆菌(clostridia)的簇,所述簇包括产乙醇梭菌物种、永达尔梭菌物种和拉氏梭菌物种。这些物种最初由abrini,《微生物学文献集(archmicrobiol)》,161:345-351,1994(产乙醇梭菌);tanner,《国际系统细菌学杂志(intjsystembacteriol)》,43:232-236,1993(永达尔梭菌);以及huhnke,wo2008/028055(拉氏梭菌)报导和表征。这三个物种有许多相似之处。具体地说,这些物种都是梭状芽胞杆菌属的c1-固定、厌氧性、产乙酸性、产乙醇性和羧基营养性成员。这些物种具有相似的基因型和表型以及相似的能量保存和发酵代谢模式。此外,这些物种簇集于16srrnadna超过99%相同的梭菌rrna同源组i中,具有约22-30摩尔%的dnag+c含量,是革兰氏阳性的,具有相似形态和大小(对数生长期细胞介于0.5-0.7×3-5μm之间),是嗜温性的(在30-37℃下生长最佳),具有约4-7.5的相似ph范围(最佳ph为约5.5-6),缺乏细胞色素,并且通过rnf复合体保存能量。另外,在这些物种中已证明羧酸还原为其对应的醇(perez,《生物技术与生物工程(biotechnolbioeng)》,110:1066-1077,2012)。重要的是,这些物种还均示出依靠含co气体的强自养性生长,产生乙醇和乙酸盐(或醋酸)作为主要发酵产物,并且在一定条件下产生少量的2,3-丁二醇和乳酸。然而,这三种物种也有许多不同之处。这些物种分离自不同的来源:来自兔子肠道的自产乙醇梭菌(clostridiumautoethanogenum)、来自养鸡场废物的永达尔梭菌(clostridiumljungdahlii),以及来自淡水沉积物的拉氏梭菌(clostridiumragsdalei)。这些物种对各种糖(例如,鼠李糖、阿拉伯糖)、酸(例如,葡糖酸盐、柠檬酸盐)、氨基酸(例如,精氨酸、组氨酸)以及其它底物(例如,甜菜碱、丁醇)的利用不同。此外,这些物种对某些维生素(例如,硫胺素、生物素)的营养缺陷型不同。这些物种在wood-ljungdahl途径基因和蛋白质的核酸和氨基酸序列上存在差异,不过已经发现所有物种中的这些基因和蛋白质的一般结构和数量相同(《生物技术最新观点(curropinbiotechnol)》,22:320-325,2011)。因此,总的来说,产乙醇梭菌、永达尔梭菌或拉氏梭菌的多种特性并不对这些物种具有特异性,但是梭状芽胞杆菌属的c1-固定、厌氧性、产乙酸性、产乙醇性和羧基营养性成员的这一簇的一般特性却对这些物种具有特异性。然而,由于这些物种实际上是不同的,所以这些物种中的一种的基因修饰或操纵在这些物种的另一种中可能不具有相同的作用。举例来说,可观察到生长、性能或产物产生的不同。本发明的微生物也可以来源于产乙醇梭菌、永达尔梭菌或拉氏梭菌的分离株或突变体。产乙醇梭菌的分离株和突变体包含ja1-1(dsm10061)(abrini,《微生物学文献集》,161:345-351,1994)、lbs1560(dsm19630)(wo2009/064200)和lz1561(dsm23693)(wo2012/015317)。永达尔梭菌的分离物和突变体包含atcc49587(tanner,《国际系统细菌学杂志(intjsystbacteriol)》,43:232-236,1993)、petct(dsm13528,atcc55383)、eri-2(atcc55380)(us5,593,886)、c-01(atcc55988)(us6,368,819)、o-52(atcc55989)(us6,368,819)和ota-1(tirado-acevedo,《使用永达尔梭菌由合成气体产生生物乙醇(productionofbioethanolfromsynthesisgasusingclostridiumljungdahlii)》,博士论文,北卡罗莱纳州立大学(northcarolinastateuniversity),2010)。拉氏梭菌的分离株和突变体包含pi1(atccbaa-622、atccpta-7826)(wo2008/028055)。“底物”是指本发明微生物的碳源和/或能量来源。通常,底物是气态的并且包括c1碳源,例如co、co2和/或ch4。优选地,底物包括co或co+co2的c1碳源。底物可进一步包括其它非碳组分,如h2、n2或电子。所述底物通常包括至少一定量的co,如约1、2、5、10、20、30、40、50、60、70、80、90或100mol%的co。底物可包括一定范围的co,如约20-80、30-70或40-60mol%的co。优选地,底物包括约40-70mol%的co(例如,钢厂或高炉煤气)、约20-30mol%的co(例如,碱性氧气炉气)或约15-45mol%的co(例如,合成气)。在一些实施例中,底物可包括相对较低量的co,如约1-10或1-20mol%的co。本发明的微生物通常将底物中的至少一部分co转化为产物。在一些实施例中,底物不包括或基本上不包括(<1mol%)co。底物可包括一定量的h2。举例来说,底物可包括约1、2、5、10、15、20或30mol%的h2。在一些实施例中,底物可包括相对较高量的h2,如约60、70、80或90mol%的h2。在其它实施例中,底物不包括或基本上不包括(<1mol%)h2。底物可包括一定量的co2。举例来说,底物可包括约1-80或1-30mol%的co2。在一些实施例中,底物可包括小于约20、15、10或5mol%的co2。在另一实施方式中,底物不包括或基本上不包含(<1mol%)co2。尽管底物通常是气态的,但底物也可以替代形式提供。举例来说,可以使用微泡分散发生器将底物溶解在用含co气体饱和的液体中。通过另一实例,底物可以吸附到固体载体上。底物和/或c1碳源可以是作为工业过程的副产物或从一些其它来源获得的废气,例如来自汽车废气或生物质气化。在某些实施例中,工业工艺选自由以下组成的组:含铁金属产品制造(例如钢厂制造)、非铁产品制造、石油精炼、煤炭气化、电力生产、碳黑生产、氨生产、甲醇生产以及焦炭制造。在这些实施例中,底物和/或c1碳源可在其被排放到大气中之前使用任何适宜方法从工业工艺中捕获。底物和/或c1碳源可以是合成气,如通过煤炭或精炼残余物的气化、生物质或木质纤维素材料的气化或天然气的重整而获得的合成气。在另一实施例中,合成气可以从城市固体废物或工业固体废物的气化中获得。底物的成分可能对反应的效率和/或成本有显著影响。举例来说,氧气(o2)的存在可能降低厌氧发酵工艺的效率。取决于底物的组成,可能需要处理、净化或过滤底物以除去任何不希望的杂质(如毒素、不需要的组分或灰尘颗粒),和/或增加所需组分的浓度。在某些实施例中,发酵在不存在碳水化合物底物(如糖、淀粉、木质素、纤维素或半纤维素)的情况下进行。本发明的微生物可以进行培养以产生一种或多种产物。例如,本发明的微生物可以产生或可以被工程化以产生乙醇(wo2007/117157)、乙酸盐(wo2007/117157)、丁醇(wo2008/115080和wo2012/053905)、丁酸盐(wo2008/115080)、2,3-丁二醇(wo2009/151342和wo2016/094334)、乳酸盐(wo2011/112103)、丁烯(wo2012/024522)、丁二烯(wo2012/024522)、甲基乙基酮(2-丁酮)(wo2012/024522和wo2013/185123)、乙烯(wo2012/026833)、丙酮(wo2012/115527)、异丙醇(wo2012/115527)、脂质(wo2013/036147)、3-羟基丙酸盐(3-hp)(wo2013/180581)、异戊二烯(wo2013/180584)、脂肪酸(wo2013/191567)、2-丁醇(wo2013/185123)、1,2-丙二醇(wo2014/036152)、1-丙醇(wo2014/0369152)、分支酸盐衍生产物(wo2016/191625)、3-羟基丁酸盐(wo2017/066498)和1,3-丁二醇(wo2017/0066498)。除了一种或多种这些目标产物以外,本发明的微生物还可产生乙醇、乙酸盐和/或2,3-丁二醇。在某些实施例中,微生物生物质本身就可以被视为是一种产物。“天然产物”是由未经过基因修饰的微生物产生的产物。举例来说,乙醇、乙酸盐和2,3-丁二醇是产乙醇梭菌、永达尔梭菌和拉氏梭菌的天然产物。“非天然产物”是由经基因修饰的微生物产生的,而不是由经基因修饰的微生物来源于的未经过基因修饰的微生物产生的产物。在本文中,提及酸(例如,乙酸或2-羟基异丁酸)应被理解为也包含对应的盐(例如,乙酸盐或2-羟基异丁酸盐)。“选择率”是指所需产物的产量与由微生物产生的全部发酵产物的产量的比率。本发明微生物可以被工程化来以特定选择率或最低选择率产生产物。在一个实施例中,目标产物占由本发明微生物产生的所有发酵产物的至少约5%、10%、15%、20%、30%、50%或75%。在一个实施例中,目标产物占由本发明微生物产生的全部发酵产物的至少10%,使得本发明微生物具有至少10%的目标产物选择率。在另一个实施例中,目标产物占由本发明微生物产生的全部发酵产物的至少30%,使得本发明微生物具有至少30%的目标产物选择率。“提高效率(increasingtheefficiency)”、“提高的效率(increasedefficiency)”等包含但不限于提高生长速率、产物产生速率或体积、消耗每体积底物的产物体积或产物选择率。可以相对于本发明微生物来源于的亲本微生物的性能来测量效率。通常,培养在生物反应器中进行。术语“生物反应器”包含由一个或多个容器、塔或管道布置组成的培养/发酵装置,如连续搅拌槽反应器(cstr)、固定化细胞反应器(icr)、滴流床反应器(tbr)、鼓泡塔、气升式发酵罐、静态混合器或适合气-液接触的其它容器或其它装置。在一些实施例中,生物反应器可包括第一生长反应器和第二培养/发酵反应器。可以向这些反应器中的一个或两个提供底物。如本文所用的,术语“培养”和“发酵”可互换使用。这些术语涵盖培养/发酵过程的生长期和产物生物合成期。培养通常在含有足以允许微生物生长的营养物、维生素和/或矿物质的水性培养基中维持。优选地,水性培养基是厌氧微生物生长培养基,如基本厌氧微生物生长培养基。合适的培养基是所属领结构域中众所周知的。培养/发酵应该理想地在产生目标产物的适当条件下进行。通常,培养/发酵在厌氧条件下进行。要考虑的反应条件包括压力(或分压)、温度、气体流速、液体流速、培养基ph、培养基氧化还原电势、搅拌速率(如果使用连续搅拌槽反应器)、接种物水平、确保液相中的气体不会变成限制因素的最大气体底物浓度和避免产物抑制的最大产物浓度。具体来说,可以控制底物的引入速率来确保液相中的气体的浓度不会变成限制因素,因为在气体限制条件下培养会消耗产物。在高压下操作生物反应器允许增加气体从气相到液相的传质速率。因此,在高于大气压力的压力下进行培养/发酵通常是优选的。此外,由于给定的气体转化率在某种程度上随底物保留时间而变并且保留时间指示生物反应器的所需体积,所以使用加压系统可以大大减小所需生物反应器的体积,并且因此降低培养/发酵设备的资金成本。这又意味着当在升高的压力而不是大气压力下维持生物反应器时,能够缩短保留时间,保留时间被定义为是生物反应器中的液体体积除以输入气体流速。最佳反应条件将部分取决于所用的特定微生物。然而,一般来说,在高于大气压力的压力下进行发酵是优选的。此外,由于给定的气体转化速率在某种程度上随底物保留时间而变,并且获得所需保留时间又指示生物反应器的所需体积,所以使用加压系统可以大大减小所需生物反应器的体积,并且因此降低发酵设备的资金成本。在某些实施例中,发酵在没有光的情况下或在不足以满足光合微生物的能量需求的光量存在下进行。在某些实施例中,本发明的微生物是非光合微生物。可使用任何方法或所属领结构域中已知方法的组合从发酵液中分离或纯化产物,所述方法包含例如分馏、蒸发、渗透蒸发、气提、相分离和萃取发酵(包含例如液-液萃取)。在某些实施例中,通过从生物反应器中不断除去培养液的一部分、从培养液分离微生物细胞(宜通过过滤)和从培养液中回收一种或多种目标产物而从发酵液中回收目标产物。可以例如通过蒸馏回收醇类和/或丙酮。可以例如通过吸附于活性炭来回收酸类。优选使分离的微生物细胞返回到生物反应器。同样优选使除去目标产物之后剩余的无细胞渗透物返回到生物反应器。额外营养物(例如维生素b)可以添加到无细胞渗透物中以补给培养基,随后使其返回到生物反应器中。本发明的微生物含有至少一个被破坏的基因。在一些实施例中,本发明的微生物含有一个以上的被破坏的基因,例如,2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、25、30、35、40、45、50、75、100或200个被破坏的基因。例如,被破坏的基因可以选自表2。尽管提供了产乙醇梭菌、永达尔梭菌和拉氏梭菌的代表性的保藏号,但是本领结构域的普通技术人员能够容易地识别其它wood-ljungdhli微生物中的同源物。诸位发明人进一步确定了wood-ljungdahl微生物中的关键代谢途径和关键代谢节点(图1)。本发明进一步提供了具有被破坏基因以战略性地将碳通量从非必要或不期望的代谢节点转移并转移通过目标代谢节点的微生物。这些菌株提高了那些目标代谢节点下游的产物产量。本发明最后提供了通过在底物(如包括co、co2和/或h2中的一种或多种的气体底物)存在下培养本发明的微生物来生产产物的方法。实例2-19描述了用于优化特定产物产生的被破坏基因的可能组合。如本申请中别处所述,这些产物可以包含wood-ljungdahl微生物的天然产物或非天然产物。例如,此类产物包含但不限于:乙酰辅酶a、乙醇、乙酸盐、丁醇、丁酸盐、丁酰辅酶a、2,3-丁二醇、乳酸盐、丁烯、丁二烯、甲乙酮、乙烯、丙酮、异丙醇、脂质、3-羟基丙酸盐(3-hp)、二甲丙烯焦磷酸盐、法呢烯、脂肪酸(脂肪酸乙酯、脂肪酸丁酯)、2-丁醇、1,2-丙二醇、1-丙醇、分支酸盐衍生产物、3--羟基丁酸盐、1,3-丁二醇、c6-c8醇(己醇、庚醇、辛醇)、己酸盐、辛酸盐、异戊烯基焦磷酸盐(ipp)、二甲基烯丙基焦磷酸盐(dmapp)、乙酰乙酰辅酶a、3-羟基丁酸辅酶a(3-hb辅酶a)、丙二酰辅酶a、丙酮酸盐、脱氢莽草酸甲酯、分支酸盐、对羟基苯甲酸、水杨酸盐、2-氨基苯甲酸盐、2,3-二羟基苯甲酸盐、2-羟基环己烷甲酸、柠苹酸盐、酮丁酸盐、乙酰乳酸盐、乙偶姻、缬氨酸、亮氨酸和异亮氨酸。实例以下实例进一步说明了本发明,但是,当然,不应该被解释为以任何方式限制本发明的范围。实例1本实施例描述了wood-ljungdahl微生物的代谢模型。利用了梭状芽孢杆菌产乙醇梭菌(clostridiumautoethanogenum)的基因组级代谢模型,如marcellin,《绿色化学(greenchem)》,18:3020-3028,2016所述。该模型用于模拟设计、构建、电子生长和筛选具有破坏性基因突变的菌株,以预测那些能产生更高产量的天然化合物的菌株。此外,为许多非天然化合物生产菌株建立了新的基因组规模模型。为此,将异源性基因和代谢反应添加到野生型产乙醇梭菌模型结构中,以表示非天然化合物生产途径的结合。尽管本文描述的实验工作所使用的模型是基于产乙醇梭菌,但考虑到代谢的相似性,这一结果也可合理地预期应用于其它wood-ljungdahl微生物。对于每一种化学生产菌株,数以百万计的结合不同破坏性基因突变组合的突变菌株是在计算机上构建的。布尔型基因-蛋白质-反应关联用于确定在基因破坏后哪些代谢反应被灭活(thiele,《自然方案(natureprotocols)》,5:93-121,2010)。使用cameo版本0.11.2(sonneschein,biosustain/cameo:0.11.0,doi:10.5281/zenodo.835730,2017)和通过inspyred版本1.0.1实施的进化算法进行突变菌株的设计、构建和筛选。使用两种基于限制的计算建模技术来模拟这些突变菌株的生长:通量平衡分析(fba)和代谢调节的线性最小化(lmoma)。这些生长模拟技术用于在之后捕获两种可能的代谢表型(maia,《gecco'17遗传和进化计算会议论文集(proceedingsofthegeneticandevolutionarycomputationconferencecompanionon–gecco'17)》,纽约,纽约州,acm出版社(acmpress),1661-1668,2017)。构建了实验代谢通量分布图,并将其用作lmoma模拟的参考状态。使用cobrapy版本0.8.2(ebrahim.,cobrapy:“基于限制的python重构和分析(constraints-basedreconstructionandanalysisforpython)”,《bmc系统生物学(bmcsystbiol)》,7:74,2013)的脚本运行增长模拟,使用optlang版本1.2.3(jensen,optlang:“用于数学优化的代数建模语言(algebraicmodelinglanguageformathematicaloptimization)”,《开源软件杂志(thejournalofopensourcesoftware)》,2,doi:10.21105/joss.00139,2017)作为求解器接口,使用gurobi优化器版本7.0.2作为优化求解器。对生长速率和关键代谢通量(包含发酵产物的生长速率和关键代谢通量)进行了记录,并将其用于筛选菌株。对于每个菌株模拟,计算了生物质-产物偶联产率(bpcy)和碳摩尔产率。这些产率用来确定适合度得分。此外,进行了通量可变性分析(fva),以确定突变菌株是否需要产生用于生长的目标化合物(生长偶联菌株设计)。如果关注化合物的最小边界通量在生长期间大于零,则所述菌株被分类为生长偶联。这些生长偶联菌株设计应该允许在连续发酵过程中有更大的发酵稳定性。该最小通量被转化为碳产率(fva最小产率),并用于比较菌株之间的生长偶联水平。实例2此实例描述了用于提高wood-ljungdahl微生物中乙酸盐的产量的破坏。如实例1所述进行代谢建模实验。乙酸盐是wood-ljungdahl微生物的天然产物。实例3此实例描述了在wood-ljungdahl微生物中乙醇产量提高的破坏。如实例1所述进行代谢建模实验。乙醇是wood-ljungdahl微生物的天然产物。实例4此实例描述了用于提高wood-ljungdahl微生物中丙酮产量的破坏。如实例1所述进行代谢建模实验。例如在wo2012/115527中描述了在wood-ljungdahl微生物中产生丙酮。本文使用以下途径模拟丙酮产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;乙酸+乙酰乙酰辅酶a-->乙酰辅酶a+乙酰乙酸盐;乙酰乙酸盐-->co2+丙酮;丙酮-->丙酮_ext。实例5此实例描述了用于提高wood-ljungdahl微生物中异丙醇产量的破坏。如实例1所述进行代谢建模实验。例如在wo2012/115527中描述了在wood-ljungdahl微生物中产生异丙醇。本文使用以下途径模拟异丙醇产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;乙酸+乙酰乙酰辅酶a-->乙酰辅酶a+乙酰乙酸盐;乙酰乙酸盐-->co2+丙酮;异丙醇-->异丙_ext。实例6此实例描述了用于提高wood-ljungdahl微生物中乳酸盐的产量的破坏。如实例1所述进行代谢建模实验。乳酸盐是wood-ljungdahl微生物的天然产物。实例7此实例描述了用于提高wood-ljungdahl微生物中1,3-丁二醇的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2017/0066498中描述了在wood-ljungdahl微生物中产生1,3-丁二醇。本文使用以下途径模拟1,3-丁二醇产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;nadph+h++乙酰乙酰辅酶a-->nadp+(r)-3-羟基丁酰辅酶a;磷酸盐+(r)-3-羟基丁酰基辅酶a-->辅酶a+(r)-3-羟基丁酰基-磷酸盐;adp+(r)-3-羟基丁酰-磷酸盐-->atp+(r)-3-羟基丁酸盐;(r)-3-羟基丁酸盐+还原铁氧还蛋白-->氧化铁氧还蛋白+(r)-3-羟基丁醛;nadph+h++(r)-3-羟基丁醛-->nadp+13bdo;nadh+h++(r)-3-羟基丁醛-->nad+13bdo;13bdo-->13bdo_ext。实例8此实例描述了用于提高wood-ljungdahl微生物中2,3-丁二醇的产量的破坏。如实例1所述进行代谢建模实验。2,3-丁二醇是至少一些wood-ljungdahl微生物的天然产物。实例9此实例描述了用于提高wood-ljungdahl微生物中2-丁醇的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2013/185123中描述了在wood-ljungdahl微生物中产生2-丁醇。本文使用以下途径模拟2-丁醇产生:nadh+h++(r)-乙偶姻-->nad+内消旋-2,3-丁二醇;内消旋-2,3-丁二醇-->h2o+mek;mek+nadph+h+-->2-丁醇+nadp;2-丁醇-->2-丁醇_ext。实例10此实例描述了用于提高wood-ljungdahl微生物中2-羟基异丁酸的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2017/0066498中描述了在wood-ljungdahl微生物中产生2-羟基异丁酸。以下途径用于模拟以下第1-40行中的2-羟基异丁酸产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;nadh+h++乙酰乙酰辅酶a-->nad+(s)-3-羟基丁酰辅酶a;(s)-3-羟基丁酰基辅酶a-->2hib辅酶a;2hib辅酶a+h2o-->2hib+辅酶a;2hib-->2hib_ext。以下途径用于模拟以下第41-93行中的2-羟基异丁酸产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;nadh+h++乙酰乙酰辅酶a-->nad+(s)-3-羟基丁酰辅酶a;(s)-3-羟基丁酰基辅酶a-->2hib辅酶a;2hib辅酶a+adp+磷酸盐-->2hib+atp+辅酶a;2hib-->2hib_ext。以下途径用于模拟以下第94-103行中的2-羟基异丁酸产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;nadh+h++乙酰乙酰辅酶a-->nad+(s)-3-羟基丁酰辅酶a;(s)-3-羟基丁酰基辅酶a-->2hib辅酶a;2hib辅酶a+醋酸盐-->2hib+乙酰辅酶a;2hib-->2hib_ext。实例11此实例描述了用于提高wood-ljungdahl微生物中3-以及其盐和离子的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2017/066498中描述了在wood-ljungdahl微生物中产生3-羟基丁酸盐。本文使用以下途径模拟3-羟基丁酸盐以及其盐和离子产生:3-羟基丁酸盐-->3-羟基丁酸盐_ext;乙酸+乙酰乙酰辅酶a-->乙酰辅酶a+乙酰乙酸盐;nadh+h++乙酰乙酸盐-->nad+3-羟基丁酸盐;2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a。实例12此实例描述了用于提高wood-ljungdahl微生物中甲基乙基酮(mek)(即2-丁酮)的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2012/024522和wo2013/185123中描述了在wood-ljungdahl微生物中产生mek。本文使用以下途径模拟甲基乙基酮产生:nadh+h++(r)-乙偶姻-->nad+内消旋-2,3-丁二醇;内消旋-2,3-丁二醇-->h2o+mek;h++mek-->h+_ext+mek_ext。实例13此实例描述了用于提高wood-ljungdahl微生物中乙酰乳酸盐的产量的破坏。如实例1所述进行代谢建模实验。乙酰乳酸盐是至少一些wood-ljungdahl微生物的天然产物。实例14此实例描述了用于提高wood-ljungdahl微生物中二甲丙烯焦磷酸盐的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2013/180584中描述了在wood-ljungdahl微生物中产生二甲丙烯焦磷酸盐。本文使用以下途径模拟二甲丙烯焦磷酸盐产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;2.0nadh+2.0h++(s)-3-羟基-3-甲基戊二酰辅酶a-->2.0nad+辅酶a+(r)-甲羟戊酸盐;atp+(r)-甲羟戊酸盐-->adp+(r)-5-磷酸甲戊酸盐;atp+(r)-5-磷酸戊戊酸盐-->adp+(r)-5-磷酸二戊戊酸盐;atp+(r)-5-二磷酸戊二酸盐-->adp+磷酸盐+co2+异戊烯基二磷酸盐;异戊烯基二磷酸酯-->dmapp;dmapp-->ppi+二甲丙烯焦磷酸盐;h2o+乙酰辅酶a+乙酰乙酰辅酶a-->辅酶a+(s)-3-羟基-3-甲基戊二酰辅酶a;二甲丙烯焦磷酸盐-->二甲丙烯焦磷酸盐_ext。这些破坏也有助于提高ipp/dmapp和/或ipp/dmapp下游产物(如法呢烯和其它萜类化合物)的产量。实例15此实例描述了用于提高wood-ljungdahl微生物中己二酸的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2017/0066498中描述了在wood-ljungdahl微生物中产生己二酸。本文使用以下途径模拟己二酸产生:atp+辅酶a+琥珀酸酯-->adp+磷酸+琥珀酰辅酶a;nad+辅酶a+2-氧化戊二酸盐-->nadh+co2+琥珀酰辅酶a;乙酰辅酶a+琥珀酰辅酶a-->辅酶a+3-氧代己二酰辅酶a;nadh+h++3-羟基己二酰辅酶a-->nad+3-羟基己二酰辅酶a;3-羟基己二酰辅酶a-->2,3-脱氢己二酰辅酶a;nadh+h++2,3-脱氢己二酰辅酶a-->nad+己二酰辅酶a;磷酸盐+己二酰辅酶a-->辅酶a+己二酰-p;adp+己二酰-p-->atp+己二酸;己二酸-->己二酸_ext。实例16此实例描述了用于提高wood-ljungdahl微生物中2-氨基苯甲酸盐的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2016/191625中描述了在wood-ljungdahl微生物中产生2-氨基苯甲酸盐。本文使用以下途径模拟2-氨基苯甲酸盐产生:nh3+分支酸盐=>h2o+丙酮酸盐+h++2-氨基苯甲酸盐。实例17此实例描述了用于提高wood-ljungdahl微生物中水杨酸盐的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2016/191625中描述了在wood-ljungdahl微生物中产生水杨酸盐。本文使用以下途径模拟水杨酸盐产生:分支酸盐-->异分支酸盐异分支酸盐-->水杨酸+丙酮酸盐。实例18此实例描述了用于提高wood-ljungdahl微生物中分支酸盐的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2016/191625中描述了在wood-ljungdahl微生物中产生分支酸盐。分支酸盐是至少一些wood-ljungdahl微生物的天然产物。这些相同的破坏预计会类似地增加通过分支酸盐上游的代谢节点—脱氢莽草酸盐的流量。实例19此实例描述了用于提高wood-ljungdahl微生物中法尼烯的产量的破坏。如实例1所述进行代谢建模实验。例如在wo2013/180584中描述了在wood-ljungdahl微生物中产生法呢烯。本文使用以下途径模拟法呢烯产生:2.0乙酰辅酶a-->辅酶a+乙酰乙酰辅酶a;h2o+乙酰辅酶a+乙酰乙酰辅酶a-->辅酶a+(s)-3-羟基-3-甲基戊二酰辅酶a;2.0nadh+2.0h++(s)-3-羟基-3-甲基戊二酰辅酶a-->2.0nad+辅酶a+(r)-甲羟戊酸盐;atp+(r)-甲羟戊酸盐-->adp+(r)-5-磷酸甲戊酸盐;atp+(r)-5-二磷酸戊二酸盐-->adp+磷酸盐+co2+异戊烯基二磷酸盐;atp+(r)-5-二磷酸戊二酸盐-->adp+磷酸盐+co2+异戊烯基二磷酸;异戊烯基二磷酸酯-->dmapp;2.0异戊烯基二磷酸+dmapp-->ppi+反式,反式-法呢基-二磷酸;反式,反式-法呢基二磷酸酯-->法呢烯+ppi;法呢烯-->法呢烯_ext。实例20此实例描述了不同产物途径中常见的基因破坏目标。在444个途径上使用进化算法进行优化。基于获得的产率(非生长偶联设计)和生物量-产物偶联产率(生长偶联设计),对每个菌株设计进行评分。每个基因都是根据其在菌株设计中出现的频率排序的。在优化的菌株中,通常识别出19个基因进行破坏。实例21此实例描述了在自养生长过程中增加目标化合物产量的基因破坏目标。该策略涉及消除或减少其它发酵副产物的产生,使目标化合物成为所需的生长副产物。如实例1所述进行代谢建模实验。模拟证据表明,这一策略适用于产生具有最小atp负担的目标化合物。这一策略不太适合自养生长过程中具有大量atp负担的产物。这是因为这一策略通过消除乙酸激酶催化的底物水平磷酸化来降低细胞atp产率。具体地,如丙酮、异丙醇、1,3-丁二醇、3-羟基异丁酸盐、2-羟基异戊酸盐、3-羟基异戊酸盐和己二酸等产物的产量可以通过将破坏性突变引入编码乙酸激酶和/或磷酸酯的基因中来提高,并任选地可以进一步将破坏性突变引入编码乙酰乳酸脱羧酶、乳酸脱氢酶、乙醛脱氢酶或柠苹酸合酶的一种或多种基因中来提高。使用通量可变性分析来评估每个模型,以确定正常生长期间朝向目标化合物的最小所需通量。然后,提出的一组破坏性基因突变被应用于每个模型。再次进行通量可变性分析,以确定化合物产生和生长之间是否存在偶联。使用cobrapy版本0.13.4进行了模拟。产物细胞生长期间最低产率目标基因被破坏细胞生长期间最低产量丙酮0.000.3011,3-丁二醇0.000.3233-羟基丁酸盐0.000.3952-羟基异丁酸盐0.000.3953-羟基异戊酸盐0.000.368硫辛酸0.000.428实例22此实例描述了通过减少所需的乙酸盐共产量,在自养生长期间在含低比例co的气体混合物上增加目标化合物产量。如实例1所述进行代谢建模实验。这一策略涉及调节氧化还原辅因子的平衡,以使nadph过量。为了维持氧化还原稳态,细胞必须产生产生途径需要nadph的产物。由于醋酸盐产生不能满足这一要求,将需要细胞产生用于实现最大生长速率的其它产物。建模证据表明,这一策略适用于具有需要协同产生乙酸盐的atp负担的目标化合物。这一策略也适用于产生乙醇作为主要产物的菌株。这种策略预计在低co气体下有效,其中细胞可以利用氢化酶来减少铁氧还蛋白和nad(p)+。在一些情况下,目标化合物的最大可能产率会降低,因为这一策略降低了细胞能量代谢的效率。具体地,如乙醇、丙酮、异丙醇、1,3-丁二醇、2-丁醇、2-羟基异丁酸盐、3-羟基异戊酸盐、己二酸、甲基乙基酮、二甲丙烯焦磷酸盐、水杨酸盐、分支酸盐和法呢烯等产物的产量可以通过将破坏性突变引入编码nad依赖性电子分叉[fefe]-氢化酶(例如,hyd)的基因中来提高,并任选地可以进一步将破坏性突变引入编码谷氨酸合酶、柠苹酸合酶、乙酰乳酸脱羧酶或乳酸盐的一种或多种基因中来提高。使用通量可变性分析对每个模型进行了评估以确定在高生长速率下乙酸盐所需的最小通量。然后,提出的一组破坏性基因突变被应用于每个模型。nad依赖性氢化酶(hyd)被从化学计量基质中移除,以表示该酶的敲除。通过谷氨酸合酶反应的通量被减少了30%,以表示该酶的破坏。再次进行了通量可变性分析以确定实现最大生长所需的最低乙酸盐产量。使用cobrapy版本0.13.4进行了模拟。实例23此实例描述了通过中枢代谢节点—乙酰乙酰辅酶a,流量不断增加。通过此节点增加通量将增加如以下的下游产物的产量:丙酮、异丙醇、3-羟基异戊酰辅酶a、3-羟基异戊酸盐、异丁烯、异戊烯焦磷酸盐(ipp)、二甲基烯丙基焦磷酸盐(dmapp)、二甲丙烯焦磷酸盐、萜类化合物(如法呢烯)、3-羟基丁酰基辅酶a、巴豆酰基辅酶a、3-羟基丁酸盐、3-羟基丁醛、1,3-丁二醇、2-羟基异丁酰基辅酶a、2-羟基异丁酸盐、丁酰基辅酶a、丁酸盐、丁醇、己酸盐、己醇、辛酸盐、辛醇、1,3-己二醇、2-丁烯-1-醇、异戊酰辅酶a、异戊酸盐或异戊醇。如实例1所述进行代谢建模实验。大多数wood-ljungdahl微生物本身不能将乙酰辅酶a转化为乙酰乙酰辅酶a,因此该步骤可能需要引入异源性酶,如硫解酶(即乙酰辅酶a乙酰转移酶)(酶代码ec2.3.1.9)。硫解酶可以是例如来自丙酮丁醇梭菌(clostridiumacetobutylicum)的thla(wp_010966157.1)、来自铜胞杆菌(cupriavidusnecator)的phaa(wp_013956452.1)、来自铜胞杆菌(cupriavidusnecator)的bktb(wp_011615089.1)、来自大肠杆菌(escherichiacoli)的atob(np_416728.1)或类似物。具体地,通过乙酰乙酰辅酶a的流量可以通过将破坏性突变引入编码nad依赖性电子分叉[fefe]-氢化酶(例如hyd)、谷氨酸合酶、柠苹酸合酶、乙酰乳酸脱羧酶、乳酸脱氢酶、乙酸激酶、磷酸转乙酰酶或乙醛脱氢酶中的一个或多个、两个或多个、三个或多个、四个或多个或五个或多个的一种或多种基因来提高。本文引用的所有参考文献(包含出版物、专利申请和专利)均通过引用的方式并入本文,其程度如同每篇参考文献被单独并且具体地指出通过引用的方式并入并且在本文中被整体阐述。本说明书中对任何现有技术的提及不是该被理解为承认该现有技术形成了何国家的研究领结构域中的公知常识的一部分。除非本文中另有所指或明显与上下文相矛盾,否则在描述本发明的上下文中(特别是在以下权利要求的上下文中)使用的术语“一个/一种(a/an)”和“所述(the)”以及类似的指代词应被解释为涵盖单数和复数两者。除非另外指出,否则术语“包括”、“具有”、“包含”和“含有”应解释为开放式术语(即,意味着“包含但不限于”)。术语“基本上由……组成”将组合物、工艺或方法的范围限制在特定的材料或步骤,或者限制在那些对组合物、工艺或方法的基本和新颖特性没有实质性影响的材料或步骤。替代方案(例如,“或”)的使用应该理解为意指替代方案中的一个、两个或其任何组合。如本文中所使用的,术语“约”是指所指定范围、值或结构的±20%,除非另外指明。除非在此另外指示,否则在此叙述的数值范围仅仅旨在充当单独地提及每个落入该范围内的单独数值的简写方法,并且将每个单独值并入说明书中,就如同单独在本文中对其进行叙述一样。例如,本文中提供的任何浓度范围、百分比范围、比值范围、整数范围、尺寸范围或厚度范围应被理解为包含所陈述范围内的任何整数的值并且在适当的情况下包含其分数(如整数的十分之一和百分之一),除非另外指明。在此描述的所有方法能以任何适合的顺序进行,除非在此另外指示或明显地与上下文矛盾。在此提供的任何和所有实例或示例性语言(例如,“如”)的使用仅旨在更好地描述本发明并且不对本发明的范围构成限制,除非另外指示。说明书中的任何语言都不应当解释为指示任何未要求保护的要素为实践本发明所必需的。本文描述了本发明的优选实施例。在阅读前述描述后,对本领结构域的普通技术人员而言,这些优选实施例的变化可以变得显而易见。本发明人预期本领结构域的技术人员可以在适当时采用这些变化,并且本发明人旨在使本发明以与在此具体描述的方式不同的方式来进行实践。因此,在适用法律允许的情况下,本发明包含对所附权利要求书所叙述的标的物的所有修改和等效物。此外,除非本文另有说明或者与上下文明显矛盾,否则本发明涵盖上述要素在其所有可能变化中的任何组合。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1