去岩藻醣基化抗体及其制造的制作方法
发明领域
本发明系有关于一种去岩藻醣基化蛋白,包括有改善的活性及治疗特性之去岩藻醣基化免疫功能分子,以及制备去岩藻醣基化蛋白的方法。
发明背景
醣蛋白涉及人类的许多基本功能,包括催化、信号传导、细胞间的讯息传递以及分子辨识别与结合。许多醣蛋白已被用于治疗的目的,且在过去的二十年中,天然存在的分泌醣蛋白重组形式一直是生技产业的主要产物。范例包括促红血球生成素(erythropoietin,epo)、治疗性单株抗体(therapeuticmonoclonalantibody,治疗性mab)、组织纤维溶酶原活化剂(tissueplasminogenactivator,tpa)、干扰素-β(interferon-β,ifn-β)、颗粒细胞-噬细胞集落刺激因子(granulocyte-macrophagecolonystimulatingfactor,gm-csf)和人类绒毛膜促性腺激素(humanchorionicgonadotrophin,hcg)。
哺乳动物中存在5类抗体,即igm、igd、igg、iga及ige。人类igg抗体由于其血液中半衰期长与功能特性,如各种效应子功能等,主要用于各种人类疾病的诊断、预防和治疗。人类igg抗体更可分成以下4种亚类:igg1、igg2、igg3及igg4。已有许多针对作为igg类抗体的效应子功能之抗体依赖性细胞毒性(antibody-dependentcellularcytotoxicity,adcc)活性与补体依赖性细胞毒性(complement-dependentcytotoxicity,cdc)的研究,且已报导igg1亚类的抗体在人类igg抗体中具有最大的adcc活性和cdc活性。
人类igg1亚类抗体的adcc活性和cdc活性的表现需要抗体fc区与效应细胞,如杀手细胞、自然杀手细胞、活化的巨噬细胞或其类似细胞及各种补体的表面上存在的抗体受体(以下称为「fcγr」)结合。已显示抗体铰链区及c区(以下称为「cγ2结构域(domain)」)中第2结构域之许多胺基酸残基,以及链接至cγ2结构域的醣链对于此结合反应是重要的。
降低或抑制抗体或fc融合蛋白的n-多醣的岩藻醣基化可增强adcc活性。adcc一般参与自然杀手(naturalkiller,nk)细胞的活化,且依赖抗体包覆细胞经nk细胞表面上fc受体的辨识。fc区域与nk细胞上fc受体的结合会受fc区域醣基化状态的影响。此外,在fc区域的n-多醣类型也会影响adcc活性。因此,对于抗体组成物或fc融合蛋白组成物,可藉由增加去岩藻醣基之n-多醣的相对量来增加对fcγriii结合亲和力,或组成物的adcc活性。
许多可影响醣基化的因子,包含物种、组织及细胞类型,均已显示对于醣化的发生方式非常重要。此外,细胞外环境,通过改变培养条件,如血清浓度,可直接对醣基化产生影响。已提出各种方法,来改变在特定宿主生物体中达到的醣基化模式,包括导入或过度表现与产生寡醣有关之某些酵素(u.s.pat.no.5,047,335;u.s.pat.no.5,510,261)。这些方案并不局限于胞内的方法((u.s.pat.no.5,278,299)。
wo98/58964描述抗体组成物,其中大致上所有的n端-连接的寡醣皆为g2寡醣。g2系指具有两个终端gal,且没有neuacs的分歧(biantennary)结构。wo99/22764提到大致上没有在ch2结构域中具有n端-连接之g1、g0或g-1寡醣的醣蛋白的抗体组成物。g1系指具有一个gal,且不具有neuacs的分歧结构,g0系指其中不具有末端neuacs或gals的分歧结构,且g-1系指核心单元少一个glcnac。
wo00/61739报导的是,与73%的由nso(小鼠骨髓瘤)细胞表现的抗体相比,47%的由yb2/0(大鼠骨髓瘤)细胞表现的抗-hil-5r抗体具有α1-6岩藻醣连接的醣链。由不同宿主细胞表现的αhil-5r抗体的岩藻醣相对比例为yb2/0<cho/d<nso。
wo02/31140与wo03/85118显示可藉由利用rnai抑制α1,6-岩藻醣基转移酶的功能,来控制与醣链结合之岩藻醣的修饰。使用细胞生产抗体组成物的方法,其包括使用抗凝集素的细胞,凝集素辨识醣链,其中岩藻醣的第1位置透过复合的n-醣苷连接的醣链中的α-键,结合至还原端中n-乙酰葡萄醣胺的第6位置。
醣链的结构在人类igg1亚类抗体的效应功能上扮演非常重要的角色,且藉由改变醣链结构,来制备具有更强效应功能的抗体可以是可能的。然而,醣链的结构多样且复杂,且醣链生理角色解决方案仍不足且昂贵。因此,需要一种生产去岩藻醣基化抗体的方法。
参考文献
1.paulson,james,etal.,“processforcontrollingintracellularglycosylationofproteins”uspatentno.5,047,335(1991)
2.goochee,charlesf.,etal.,“methodofcontrollingthedegradationofglycoproteinoligosaccharidesproducedbyculturedchinesehamsterovarycells”uspatentno.5,510,261(1996)
3.wong,chi-huey,etal.,“methodandcompositionforsynthesizingsialylatedglycosylcompounds”uspatentno.5,278,299(1994)
4.raju,t.,shantha,“methodsandcompositionsforgalactosylatedglycoproteins”wo/1998/058964(1998)
5.raju,t.,shantha,“methodsandcompositionscomprisinggalactosylatedglycoproteins”wo/1999/022764(1999)
6.hanai,nobuo,etal.,“methodforcontrollingtheactivityofimmunologicallyfunctionalmolecule”wo/2000/061739(2000)
7.kanda,yutaka,etal.,“cellsproducingantibodycompositions”wo/2002/031140(2002)
8.blumberg,r.s.,etal.,“centralairwayadministrationforsystemicdeliveryoftherapeutics”wo03/077834(2002)
9.blumberg,r.s.,etal.,“centralairwayadministrationforsystemicdeliveryoftherapeutics”uspatentapplicationpublication2003-0235536(2003)
10.
11.ferrara,c.,etal.,“uniquecarbohydrate-carbohydrateinteractionsarerequiredforhighaffinitybindingbetweenfcγriiiandantibodieslackingcorefucose”procnatlacadsciu.s.a.108,12669-12674
发明概述
本发明系有关于产生具加强活性之去岩藻醣化蛋白,包括去岩藻醣化抗体的新颖方法。本发明也有关于以本发明方法所产生之去岩藻醣基化蛋白,以及产生去岩藻醣基化蛋白的细胞。本发明的去岩藻醣基化抗体,相较于天然存在的岩藻醣基化抗体,具有增加之抗体依赖性细胞毒性(adcc)活性。
本发明的一范畴系有关于一种在宿主细胞产生去岩藻醣基化蛋白,包括去岩藻醣基化抗体的方法。本发明之方法一般包括将编码岩藻醣基化途径中经修饰之酵素的核酸导入至宿主细胞中,以抑制抗体在此宿主细胞中的岩藻醣基化。经修饰的酵素可源自于岩藻醣基化途径中的酵素。在某些实施例中,经修饰的酵素可源自于gdp-甘露醣4,6-脱水酶(gdp-mannonse4,6-dehydratase,gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(gdp-4-keto-6-deoxy-d-mannoseepinierase-reductase,fx)及/或岩藻醣转移酶(fut1至fut12、pofut1及pofut2)。在一些实施例中,经修饰的酵素可源自于gmd或fut。在特定实施例中,经修饰的酵素可源自于α-1,6-岩藻醣转移酶(fut8)。经修饰的酵素可抑制宿主细胞在岩藻醣化途径中天然存在的酵素的功能,进而抑制宿主细胞中抗体的岩藻醣化。
在一些实施例中,生产去岩藻醣基化蛋白(包括去岩藻醣基化抗体)的方法,包括(a)提供宿主细胞,(b)将编码岩藻醣基化途径之经修饰的酵素的核酸导入至此宿主细胞中,以及(c)于此宿主细胞中产生去岩藻醣基化蛋白。
本发明另一范畴系有关于以本发明方法所产生的去岩藻醣基化蛋白,包括去岩藻醣基化抗体。相较于天然存在的岩藻醣基化抗体,去岩藻醣化抗体具有增加及改善的活性。在一些实施例中,抗体具有增加及改善的adcc。
本发明另有关于生产去岩藻醣基化蛋白,包括去岩藻醣基化抗体的细胞。
附图简述
于以下实施例提供本发明之实施方式,并搭配所附图式。
第1图为于rc79细胞(表现
第2图为表现f83m突变蛋白之rc79重组细胞和rc79亲源细胞的流式细胞分析。长虚线之波峰代表经罗丹明-lca染色之表现f83m的rc79重组细胞。灰色填满之波峰代表未经罗丹明-lca染色之表现f83m的rc79重组细胞(阴性对照组)。短虚线之波峰代表经罗丹明-lca染色的rc79细胞(未表现f83m的亲源细胞)(阳性对照组)。
第3a及3b图为显示adcc分析结果的图。第3a及3b图分别显示经由来自捐赠者1(第3a图)及捐赠者2(第3b图)之pbmc细胞的
第4a至4c图为显示用spr生物传感器(biacoretmx100)的fcγriiia亲和力分析的spr感应图的图。将his-标签fcγriiia(1μg/ml)及5-80nm去岩藻醣基化抗-cd20单株抗体(第4a图)、20-320nm
第5图为显示
第6图为显示以食盐水(载体)、
第7图为显示自以食盐水(载体)、
第8图为显示以食盐水(载体)、
发明详述
本发明系有关于生产具改善活性之去岩藻醣基化抗体的新颖方法。本发明亦有关于以此发明方法所产生的去岩藻醣基化抗体以及产生此去岩藻醣基化抗体的细胞。相较于天然存在的岩藻醣基化抗体,本发明之去岩藻醣基化抗体具有增加的抗体依赖性细胞毒性(adcc)活性。
以下为提供的实施方式,以协助本发明所属技术领域中具有通常知识者实施本发明。本发明所属技术领域中具有通常知识者可理解本发明说明明确描述的实施例的修改或变化并不会偏离本发明所含有的信息的精神或范围,仍属于本发明之范畴。于描述中所使用的术语仅用于描述特定实施例,并不限制本发明。以下所使用的标题仅为组织各章节的目的,不应被视为限制所描述的主题。
本发明说明中所提及的所有文献、申请专利、专利、图式及其他参考文献整体(包括部分)可完全并入,如在发明说明中所公开和叙述。
除另外说明,本发明说明所使用的所有技术和科学术语与本发明所属技术领域中具有通常知识者通常理解具有相同的意义。除非上下文另有说明,发明说明中所述单数「一」和「该」包括复数的意思。类似地,除非上下文另有说明,词语「或」意图包括「及」的意思。因此,「包括a或b」系指包括a或b、或a及b。更应了解的是,多胜肽的所有胺基酸大小,所有分子量或分子质量值为近似值,且仅供描述。尽管可使用与本发明说明所述类似或相同的方法和材料,但适合的方法及材料如下所述。本发明说明中所述所有文献、专利说明书、专利及其它文献皆可并入本发明中。如果发生冲突,会受到本发说明(包括术语解释)限制。此外,材料、方法和实施例仅是说明性质,不意图限制本发明。
1.细胞中岩藻醣基化的抑制
本发明之一范畴系有关于抑制或减少细胞中岩藻醣基化的方法。
a.宿主细胞
任何适当的宿主细胞可用于生产去岩醣基化抗体,包括源自于酵母菌、昆虫、两栖类、鱼类、爬虫动物、鸟类、哺乳动物或人类,或融合瘤的宿主细胞。宿主细胞可以是未经修饰的细胞或细胞株,或经遗传修饰的细胞株(例如,以促进生物产品的生产)。在一些实施例中,宿主细胞为细胞株,其已经修饰,以允许在期望的条件下,例如在无血清培养基中、在细胞悬浮式培养中或在贴壁式细胞培养中生长。
使用哺乳动物宿主细胞有助于给予至人类的抗体。在一些实施例中,宿主细胞为中国仓鼠卵巢(chinesehamsterovary,cho)细胞,其为用于表现许多重组蛋白的细胞株。一般用于表现重组蛋白的额外哺乳动物细胞株包括293hek细胞、hela细胞、cos细胞、nih/3t3细胞、jurkat细胞、nso细胞及huvec细胞。在另一些实施例中,宿主细胞为表现抗体的重组细胞。
于本发明说明提供的方法中有用的人类细胞株的范例包括293t(胚胎肾)、786-0(肾)、a498(肾)、a549(肺泡上皮)、achn(肾)、bt-549(乳房)、bxpc-3(胰)、caki-1(肾)、capan-i(胰)、ccrf-cem(白血病)、colo205(结肠)、dld-1(结肠)、dms114(小细胞肺)、du145(前列腺)、ekvx(非小细胞肺)、hcc-2998(结肠)、hct-15(结肠)、hct-116(结肠)、ht29(结肠)、sit-1080(纤维肉瘤)、hek293(胚胎肾)、hela(子宫颈癌)、hepg2(肝细胞癌)、hl-60(tb)(白血病)、hop-62(非小细胞肺)、hop-92(非小细胞肺)、hs578t(乳房)、ht-29(结肠腺癌)、ig-ov1(卵巢)、imr32(神经母细胞瘤)、jurkat(t淋巴细胞)、k-562(白血病)、km12(结肠)、km20l2(结肠)、lans(神经母细胞瘤)、lncap.fgc(高加索前列腺腺癌)、loximv1(黑色素瘤)、lxfl529(非小细胞肺)、m14(黑色素瘤)、m19-mel(黑色素瘤)、malme-3m(黑色素瘤)、mcfioa(乳房上皮)、mci′7(乳腺)、mda-mb-453(乳房上皮)、mda-mb-468(乳房)、mda-mb-231(乳房)、mda-n(乳房)、molt-4(白血病)、ncl/adr-res(卵巢)、nci-1122.0(非小细胞肺)、nci-h23(非小细胞肺)、nc1-h322m(非小细胞肺)、nci-h460(非小细胞肺)、nci-h522(非小细胞肺)、ovcar-3(卵巢)、qvcar-4(卵巢)、ovcar-5(卵巢)、ovcar-8(卵巢)、p388(白血病)、p388/adr(白血病)、pc-3(前列腺)、
于本发明说明提供的方法中有用之非人类灵长类细胞株的范例包括经猴肾(cvi-76)、非洲绿猴肾(vero-76)、绿猴纤维细胞(cos-1)及经sv40转化之猴肾细胞(cos-7)之细胞株。额外的哺乳动物细胞株为本发明所属技术领域中具有通常知识者已知,且为编录在美国典型培养物保藏中心(americantypeculturecollection,atcc)目录(manassas,va)中。
b.修饰岩藻醣基化途径中的酵素
本发明之去岩藻醣基化抗体可在宿主细胞中产生,此宿主细胞中的岩藻醣基化途径已经以减少或抑制蛋白的岩藻醣基化的方式被改变。
i.经修饰的酵素
本发明说明中所使用之词组「经修饰的酵素」系指衍生自在岩藻醣基化途径中天然存在或野生型酵素的蛋白,其以在修饰后改变或破坏蛋白质的天然酵素活性的方式被改变。经修饰的酵素能够抑制或干扰其对应的野生型酵素,以改变、抑制或降低宿主细胞中野生型酵素的活性。
经修饰的酵素可藉由改变天然存在的酵素来生产,例如,藉由改变总蛋白质电荷、共价连接化学或蛋白质部分(moiety)、导入胺基酸置换、插入及/或删除,及/或其任何组合。在一些实施例中,相较于天然存在的酵素对应物,经修饰的酵素具有胺基酸的取代、添加、及/或删除。在一些实施例中,相较于天然存在的酵素对应物,经修饰的酵素有介于1至约20个胺基酸取代、添加、及/或删除。胺基酸的取代、添加及插入可利用天然或非天然胺基酸来完成。非天然存在之胺基酸包括,但不限于,ε-n离胺酸、β-丙胺酸、鸟胺酸、正白胺酸、正缬胺酸、羟脯胺酸、甲状腺素、γ-胺基丁酸、高丝胺酸、瓜胺酸、胺基苯甲酸、6-氨基己酸(aca;6-aminohexanoicacid)、羟脯胺酸、硫醇丙酸(mpa)、3-硝基-酪胺酸、焦谷胺酸及其类似。天然存在之胺基酸包括丙胺酸、精胺酸、天门冬酰胺酸、天冬胺酸、半胱胺酸、麸胺酸、麸酰胺酸、甘胺酸、组胺酸、异白胺酸、白胺酸、离胺酸、甲硫胺酸、苯丙胺酸、脯胺酸、丝胺酸、苏胺酸、色胺酸、酪胺酸及缬胺酸。
经修饰的酵素可源自于岩藻醣基化途径中的任何天然存在之酵素。例如,经修饰的酵素可源自于gdp-甘露醣4,6-脱水酶(gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(fx)及/或任何岩藻醣转移酶,包含:2-α-l-岩藻醣转移酶1(fut1)、半乳醣苷2-α-l-岩藻醣转移酶2(fut2)、半乳醣苷3(4)-l-岩藻醣转移酶(fut3)、α(1,3)岩藻醣转移酶,骨髓特异性(fut4)、α-(1,3)-岩藻醣转移酶(fut5)、α-(1,3)-岩藻醣转移酶(fut6)、α-(1,3)-岩藻醣转移酶(fut7)、α-(1,6)-岩藻醣转移酶(fut8)、α-(1,3)-岩藻醣转移酶(fut9)、蛋白o-岩藻醣转移酶1(pofut1)、蛋白o-岩藻醣转移酶2(pofut2)。
在一些实施例中,岩藻醣基化途径中之一个以上的酵素被修饰。在某些实施例,经修饰的酵素源自于gmd、fx及/或fut8。
ii.编码经修饰的酵素的核酸
本发明的去岩藻醣基化抗体可于宿主细胞中产生,在此宿主细胞中,岩藻醣基化途径中酵素已经以降低或抑制蛋白的岩藻醣基化的方式被改变。
在一些实施例中,藉由将编码岩藻醣基化途径中经修饰之酵素的核酸导入至宿主细胞中,以改变宿主细胞的岩藻醣基化途径。例如,将编码经修饰的酵素的核酸分子插入至表现载体中,并转染至宿主细胞中。编码经修饰的酵素的核酸分子可瞬时导入宿主细胞中,或稳定地融合至宿主细胞的基因体中。可使用标准的重组dna方法,来产生编码经修饰的酵素的核酸,将核酸并入至表现载体中,以及将载体导入至宿主细胞中。
在一些实施例中,宿主细胞可表现2个或以上的经修饰的酵素。例如,可用编码2个或以上经修饰的酵素的核酸,来转染宿主细胞。或者,可用一个以上的核酸,来转染宿主细胞,各个核酸可编码一个或以上的经修饰之酵素。
编码经修饰的酵素的核酸可含有额外的核酸序列。例如,核酸可含有蛋白卷标、筛选标志、或控制宿主细胞中蛋白表现的调控序列,例如启动子、增强子或其它控制核酸(如,聚腺苷酸化信号)的转录或转译的表现控制片段。此种调控序列已为习知。本发明所属技术领域中具有通常知识者可理解的是,可依据数个因素,包括要转形的宿主细胞的选择、所需蛋白质的表现量等,来选择表现载体,包括调控片段的选择。用于哺乳动物宿主细胞表现的示例性调控序列,包括哺乳动物细胞中指引高蛋白质表现量的病毒片段,例如,源自于巨细胞病毒(cytomegalovirus,cmv)(例如,cmv启动子及/或增强子)、猿猴病毒40(simianvirus,sv40)(例如,sv40启动子及/或增强子)、腺病毒(例如,腺病毒主要晚期启动子(adenovirusmajorlatepromoter,admlp))与多瘤性病毒的启动子及/或增强子。
在某些实施例中,将含有源自于gmd、fx及/或fut的经修饰的酵素之核酸序列导入宿主细胞。在表现经修饰的酵素的宿主细胞中,岩藻醣基化途径会被改变、抑制、或降低。
c.表现经修饰的酵素的宿主细胞
本发明另一范畴有关于表现岩藻醣基化途径中经修饰的酵素的宿主细胞。宿主细胞中经修饰的酵素的表现会干扰野生型酵素的活性,导致抑制或降低岩藻醣基化途径。因此,于表现经修饰的酵素之宿主细细胞中所产生的蛋白(如,抗体)被去岩藻醣基化(无岩醣基化)。
如本发明所使用的词组「低岩藻醣基化细胞」或「低岩藻醣基化宿主细胞」系指,因为细胞表现岩藻醣基化途径中经修饰的酵素,所以岩藻醣基化途径已受到抑制或减少的细胞。
可藉由将含有编码岩藻醣基化途径中经修饰之酵素的核酸序列的表现载体转染至宿主细胞,以制备低岩藻醣基化细胞。可使用本领域已知的技术进行转染。例如,可使用以化学为基础的方法(如,脂质、磷酸钙、阳离子聚合物、deae-葡聚醣、活化的树枝形大分子,磁珠等)、藉由以仪器为基础的方法(如,电穿孔、生物喷射技术、显微注射、雷射/光注射等),或藉由以病毒为基础的方法,来进行转染。
可使用存在于表现载体上的筛选标志,自未转染细胞筛选并分离出转染细胞。此外,可进一步藉由各种技术,自具正常岩醣基化途径的细胞筛选并分离出具有岩藻醣基化途径被抑制或降低的转染细胞,。例如,可使用抗体、凝集素、代谢标志或化学酵素策略来确定岩藻醣基化。此外,可藉由使转染细胞暴露于小扁豆凝集素(lensculinarisagglutinin,lca,vectorlaboratoriesl-1040),来筛选出具有岩藻醣基化途径被抑制或降低的细胞。lca辨识n端连接的寡醣的α-1,6-岩藻醣基化三甘露醣核心结构,并使表现此结构的细胞进入细胞死亡途径。因此,在接触lca后存活的细胞具有经抑制或降低的岩藻醣化途径,且被视为低岩藻醣基化细胞。
2.去岩藻醣基化蛋白
本发明另一范畴系有关于一种生产去岩藻醣基化蛋白的方法。在一些实施例中,去岩藻醣基化蛋白为去岩藻醣基化抗体。
a.蛋白
可被产生为去岩藻醣基化蛋白的蛋白的非限制范例包括,gp-73、凝血酶、hbsag、b型肝炎病毒颗粒、α-酸-醣蛋白、α-1-糜蛋白酶、α-1-糜蛋白酶、α-1-糜蛋白酶组胺酸-脯胺酸-较少(his-pro-less)、α-1-抗胰蛋白酶、血清转铁蛋白(serotransferrin)、血浆铜蓝蛋白、α-2-巨球蛋白、α-2-hs-醣蛋白、α-甲胎蛋白、结合珠蛋白、纤维蛋白原γ链前驱物、免疫球蛋白(包括igg、iga、igm、igd、ige及其类似)、apo-d、激肽原、富含组胺酸的醣蛋白、补体因子1前驱物、补体因子i重链、补体因子i轻链、补体c1s、补体因子b前驱物、补体因子bba片段、补体因子bbb片段、补体c3前驱物、补体c3β链、补体c3α链、c3a过敏毒素、补体、c3bα’链、补体c3c片段、补体c3dg片段、补体c3g片段、补体c3d片段、补体c3f片段、补体c5、补体c5β链、补体c5α链、c5a过敏毒素、补体c5α'链、补体c7、α-1b醣蛋白、b-2-醣蛋白、维生素d-结合蛋白、inter-α-胰蛋白酶抑制剂重链h2、α-1b-醣蛋白、血管收缩素原前驱物、血管收缩素-1、血管收缩素-2、血管收缩素-3、garp蛋白、β-2-醣蛋白、聚集素(clusterin)(apoj)、整合素α-8前驱物醣蛋白、整合素α-8重链、整合素α-8轻链、c型肝炎病毒颗粒、elf-5、激肽原、hsp33-同源、离胺酰基内肽酶及富含重复亮胺酸的蛋白质32前驱物。
b.抗体
本发明说明所使用的术语「抗体」广泛地涵盖了能被岩藻醣基化的完整抗体及其片段。例如,抗体包括完全组装的免疫球蛋白(例如多株、单株、单特异性、多特异性、嵌合、去免疫、人源化、人类、灵长类化、单链、单结构域、合成及重组抗体);具所需活性或功能之完整抗体的部分(例如,抗体的免疫片段,其含有fab、fab’、f(ab’)2、fv、scfv、单域片段);以及含有能被岩藻醣基化之fc结构域的胜肽和蛋白(例如,fc-融合蛋白)。
本发明说明所使用的术语「去岩藻醣基化抗体」系指在相较于在天然条件下所产生的抗体,岩藻醣基化被抑制或显著降低的条件下所产生的抗体或其片段。以本发明方法所产生的去岩藻醣基化抗体可为完全(100%)去岩藻醣基化,或者可包括岩藻醣基化及去岩藻醣基化的分子的混合。例如,在一些实施例中,由本发明方法所产生之抗体可含有约20%至约100%的去岩藻醣基化分子。在另一些实施例中,由本发明方法所产生之抗体可含有约40%至约100%的去岩藻醣基化分子。在某些实施例中,由本发明方法所产生的抗体可含有约至少20%、30%、40%、50%、60%、70%、80%、90%、91%、92%、93%、94%、95%、96%、97、98%、99%或100%的去岩藻醣基化分子。并非所有的n-醣化抗体或其片段(例如,fc-融合蛋白)皆需为去岩藻醣基化。
b.抗体种类
可使用本发明方法可将任何抗体生产为去岩藻醣基化抗体。对于可使用本发明方法产生的抗体的种类并无限制。以下为可产生的抗体的非详尽列表。
辨识肿瘤相关抗原的抗体的范例包括抗-gd2抗体、抗-gd3抗体、抗-gm2抗体、抗-her2抗体、抗-cd52抗体、抗-mage抗体、抗-hm124抗体、抗-甲状旁腺激素相关蛋白(parathyroidhormone-relatedprotein,pthrp)抗体、抗-碱性纤维细胞生长因子抗体与抗-fgf8抗体、抗-碱性纤维细胞生长因子受体抗体与抗-fgfs受体抗体、抗-类胰岛素生长因子抗体、抗-类胰岛素生长因子受体抗体、抗-pmsa抗体、抗-血管内皮细胞生长因子抗体、抗-血管内皮细胞生长因子受体抗体及其类似物。
辨识过敏或发炎相关抗原的抗体的范例包括抗-il-6抗体、抗-il-6受体抗体、抗-il-5抗体、抗-il-5受体抗体与抗-il-4抗体、抗-肿瘤坏死因子抗体、抗-肿瘤坏死因子受体抗体、抗-ccr4抗体、抗-趋化因子抗体、抗-趋化因子受体抗体及其类似物。
辨识循环器官疾病相关抗原的抗体的范例包括抗-gpiib/iiia抗体、抗-血小板衍生生长因子抗体、抗-血小板衍生生长因子受体抗体及抗-血液凝固因子抗体及其类似物。
辨识病毒或细菌感染相关抗原的抗体的范例包括抗-gpl20抗体、抗-cd4抗体、抗-ccr4抗体与抗-维罗毒素抗体及其类似物。
许多治疗性抗体为市售可得的,例如,结合vegf(如,bevacizumab
3.制备去岩藻醣基化蛋白的方法
在低岩藻醣基化细胞中产生本发明去岩藻醣基化蛋白,其包括去岩藻醣基化抗体。使用本领域习知技术,例如以编码蛋白之表现载体转染低岩藻醣基化细胞,可在低岩藻醣基化细胞中表现去岩藻醣基化蛋白。
可使用本领域习知技术,来制备编码蛋白的表现载体。例如,将胺基酸序列反转译成核酸序列,以构筑表现载体,较佳使用对于在其中表现蛋白质之生物体而言优化的密码子。然后,将编码蛋白的核酸和其任何调控片段组装并插入至所欲的表现载体中。表现载体可含有额外的核酸序列,例如蛋白卷标、筛选标志、或控制蛋白表现的调控序列,如同上述含有经修饰的酵素的表现载体。表现载体接着可利用转染导入宿主细胞。可使用已知本领域习知技术进行转染。例如,可使用以化学为基础的方法(如,脂质、磷酸钙、阳离子聚合物、deae-葡聚醣、活化的树枝形大分子、磁珠等),藉由以仪器为基础的方法(如,电穿孔、生物喷射技术、显微注射、雷射/光注射等),或藉由以病毒为基础的方法进行转染。之后在适合所选表现系统及宿主的条件下,可在经转染的细胞中表现蛋白。可接着使用亲和性管柱或其它本领域已知技术纯化表现的蛋白。
将编码经修饰的酵素(欲成为一低岩藻醣基化细胞)的核酸及编码蛋白(欲表现此蛋白)的核酸,以任意的顺序转染至宿主细胞,以产生去岩藻醣基化蛋白。例如,可先将编码经修饰的酵素(欲成为低岩藻醣基化细胞)的核酸转染至宿主细胞中,再以编码一蛋白(欲表现此蛋白)的核酸转染。或者,可先将编码蛋白(欲表现此蛋白)的核酸转染至宿主细胞中,再以编码经修饰的酵素(欲成为一低岩藻醣基化细胞)的核酸转染。在另一变化中,可同时将编码经修饰的酵素(欲成为一低岩藻醣基化细胞)的核酸与编码蛋白(欲表现此蛋白)的核酸转染至宿主细胞中。
在一特定实施例中,依照下列步骤,藉由先制备低岩藻醣基化细胞,然后将编码蛋白质的核酸转染至低岩藻醣基化细胞中,以产生去岩藻醣基化蛋白:
a)获得适合表现蛋白的宿主细胞;
b)将编码经修饰的酵素的核酸转染至此宿主细胞中;
c)筛选及/或分离具低岩藻醣基化的转染细胞;
d)将编码蛋白的核酸转染至低岩藻醣基化细胞中;
e)筛选及/或分离经以编码蛋白之核酸转染的低岩藻醣基化细胞;
f)诱导蛋白在低岩藻醣基化细胞中的表现。
在另一实施例中,依照下列步骤,藉由先将编码蛋白的核酸转染至宿主细胞中,再以编码经修饰的酵素的核酸转染此细胞,以产生去岩藻醣基化蛋白:
a)获得适合表现蛋白的宿主细胞;
b)将编码蛋白的核酸转染至宿主细胞中;
c)筛选及/或分离经以编码蛋白的核酸转染的细胞;
d)将编码经修饰的酵素的核酸转染至步骤(c)的细胞中;
e)筛选及/或分离步骤(d)中之具低岩藻醣基化的转染细胞;
f)诱导蛋白在低岩藻醣基化细胞中的表现。
在上述实施例的变化中,去岩藻醣基化蛋白可依下述步骤产生:
a)获得表现或过度表现蛋白的宿主细胞;
b)将编码经修饰的酵素的核酸转染至此细胞中;
c)筛选及/或分离具低岩藻醣基化的转染细胞;
d)诱导蛋白在低岩藻醣基化细胞中的表现。
在又一实施例中,如下,藉由同时将编码经修饰的酵素(欲成为一低岩藻醣基化细胞)的核酸和编码蛋白(欲表现此蛋白)的核酸转染至宿主细胞,以产生去岩藻醣基化蛋白:
a)获得适合表现一蛋白的宿主细胞;
b)将编码蛋白的第一核酸及编码经修饰的酵素的第二核酸转染至宿主细胞;
c)筛选及/或分离表现蛋白且具有低岩藻醣基化的转染细胞;
d)诱导蛋白在低岩藻醣基化细胞中的表现。
使用上述的本发明方法产生去岩藻醣基化蛋白,包括抗体,可以本领域习知方法纯化。例如,藉由本发明方法所产生的去岩藻醣基化蛋白,包括抗体,可以生理化学分馏、抗体类特异性亲和力、抗原特异性亲和力等方式纯化。
4.去岩藻醣基化抗体改善的特性
相较于使用标准方式所产生的抗体,以本发明方法生产的去岩藻醣基化抗体具有改善的特性。
可藉由elisa、荧光法及其类似方式,来测量经纯化之去岩藻醣基化抗体的活性。可藉由测量其adcc和cdc等方式,来评估抗原阳性的培养细胞株的细胞毒性活性。可使用相对接近人类的动物种类的适当模型,来评估抗体在人体内的安全性和治疗效果。
a.增加的adcc活性
相较于使用标准方式所产生的抗体,本发明的去岩藻醣基化抗体具有增加的adcc活性。
本发明方法所用之「adcc活性」系指抗体引起抗体依赖性细胞毒性(adcc)反应的能力。adcc为细胞媒介的反应,其中表现fcrs之抗原非特异性细胞毒性细胞(如,自然杀手(nk)细胞、嗜中性细胞与巨噬细胞)辨识结合至目标细胞表面的抗体,接着造成目标细胞的裂解(即,杀死)。adcc中的主要媒介细胞是自然杀手(nk)细胞。nk细胞表现fcγriii,其中fcγriiia为活化受体而fcγriiib为抑制受体。单核细胞表现fcγri、fcγrii与fcγriii。可使用体外分析法,如实施例3中描述的分析法,来直接评定adcc活性。
可使用体外分析法,来直接评定adcc活性。在一些实施例中,本发明的去岩藻醣基化抗体的adcc活性至少是其野生型对照抗体的0.5、1、2、3、5、10、20、50、100倍。
由于去岩藻醣基化抗体具有增加的adcc活性,相较于其岩藻醣基化抗体,可以较低的量或浓度,来给予去岩藻醣基化的治疗抗体。在一些实施例中,本发明的去岩藻醣基化抗体的浓度可比其岩藻醣基化抗体的浓度低至少2、3、5、10、20、30、50或100倍。在一些实施例中,相较于其野生型抗体,本发明的去岩藻醣基化抗体可展现更高的最大目标细胞裂解(maximaltargetcelllysis)。例如,本发明的去岩藻醣基化抗体的最大目标细胞裂解(maximaltargecelllysis)比其野生型抗体的最大目标细胞裂解高10%、15%、20%、25%、30%、40%、50%或更多。
b.增加的cdc活性
相较于使用标准方法产生的抗体,本发明的去岩藻醣基化抗体具有增加的补体依赖性细胞毒性(cdc)活性。
本发明方法所使用的「cdc活性」系指辨识在目标细胞上结合的抗体,接着使目标细胞分解的补体系统中的一或多个成分的反应。本发明的去岩藻醣基化抗体不会降低或抑制cdc活性,但相反地,其维持类似或高于其岩藻醣基化抗体的cdc活性。
本发明更提供具有增强的cdc功能的去岩藻醣基化抗体。在一实施例中,本发明的fc变异体具有增加的cdc活性。在另一实施例中,所述去岩藻醣基化抗体具有比相当的分子高至少2倍,或至少3倍,或至少5倍,或至少10倍,或至少50倍,或至少100倍的cdc活性。
4.使用去岩藻醣基化抗体
可以静脉内(intravenously,i.v.)、皮下(subcutaneously,s.c.)、肌内(intra-muscularly,i.m.)、皮内(intradermal,i.d.)、腹膜内(intraperitoneal,i.p.)或经由任何黏膜表面,如口服(orally,p.o.)、舌下(sublingually,s.l.)、脸颊、鼻、直肠、阴道或经由肺部途径,来给予本发明的去岩藻醣基化抗体。
对于治疗或预防各种疾病,包括癌症、发炎性疾病、免疫及自体免疫疾病、过敏、循环器官疾病(如,动脉硬化)以及病毒或细菌感染,去岩藻醣基化抗体是有用。
本发明的去岩藻醣基化抗体的剂量会依据给予的对象及特定模式而改变。可根据本发明所属技术领域中具有通常知识者所熟知的许多因子,包括但不限于,抗体目标、对象的种类以及对象的大小/体重,而改变所需的剂量。剂量可为0.1至100,000μg/kg体重。可单剂或多剂给予去岩藻醣基化抗体。去岩藻醣基化抗体可在24小时期间中给予一次,在24小时期间给予多次,或持续注入。可持续地或以特定时程给予去岩藻醣基化抗体。可由获自动物模型的剂量-反应曲线推断有效剂量。
4.特定实施例
本发明的特定实施例包括,但不限于以下:
(1)一种在宿主细胞中生产去岩藻醣基化蛋白的方法,包括将至少一个编码岩藻醣基化途径(fucosylationpathway)之经修饰之酵素的核酸导入至宿主细胞中,以产生去岩藻醣基化蛋白。
(2)如(1)之方法,其中此经修饰的酵素源自于gdp-甘露醣4,6-脱水酶(gdp-mannose4,6-dehydrogenase,gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(gdp-4-keto-6-deoxy-d-mannoseepinierase-reductase,fx)、或岩藻醣转移酶(fucosyltransferase,fut)。
(3)如(1)之方法,其中经修饰的酵素源自于岩藻醣转移酶(fut)。
(4)如(1)之方法,其中经修饰的酵素源自于α-1,6-岩藻醣转移酶(fut8)。
(5)如(1)之方法,其中经修饰的酵素于宿主细胞中降低或抑制经修饰之酵素所源自的野生型酵素的活性。
(6)如(1)之方法,其中修饰酵素抑制或降低宿主细胞中的岩藻醣基化。
(7)如(1)之方法,其中于宿主细胞中所产生的去岩藻醣基化蛋白为去岩藻醣基化抗体或其片段。
(8)如(7)之方法,其中去岩藻醣基化抗体或其片段至少90%去岩藻醣基化。
(9)如(7)之方法,其中去岩藻醣基化抗体,相较于其岩藻醣基化抗体,具有增加的抗体依赖性细胞毒性(antibody-dependentcellularcytotoxicity,adcc)活性。
(10)如(7)之方法,其中去岩藻醣基化抗体的补体依赖性细胞毒性(cdc)相较于其岩藻醣基化抗体,未被降低或抑制。
(11)一种在宿主细胞中产生去岩藻醣基化蛋白的方法,包括将编码岩藻醣基化途径之经修饰之酵素的第一核酸序列,以及编码欲产生的蛋白之第二核酸序列转染至宿主细胞中。
(12)如(11)之方法,其中宿主细胞同时转染第一核酸与第二核酸。
(13)如(12)之方法,其中所产生的去岩藻醣基化蛋白为抗体和/或其片段。
(14)如(11)之方法,其中第一核酸编码源自于gdp-甘露醣4,6-脱水酶(gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(fx)、及/或岩藻醣转移酶(fut)的经修饰的酵素。
(15)如(11)之方法,其中第一核酸编码源自于α-1,6-岩藻醣转移酶(fut8)的经修饰的酵素。
(16)如(11)之方法,包括下列步骤:
a)将编码经修饰的酵素的第一核酸转染至宿主细胞中;
b)筛选及分离低岩藻醣基化的转染细胞;
c)将编码欲产生之蛋白的第二核酸转染至低岩藻醣基化细胞中;
d)在此低岩藻醣基化细胞中表现欲产生之蛋白。
(17)如(11)之方法,其中所产生的去岩藻醣基化蛋白为抗体及/或其片段。
(18)如(11)之方法,包括以下步骤:
a)将编码欲产生的蛋白的第二核酸转染至宿主细胞;
b)筛选及分离转染第二核酸的细胞;
c)将编码经修饰的酵素之第一核酸转染至于步骤(b)中的细胞;
d)筛选及分离低岩藻醣基化的转染细胞;
e)在此低岩藻醣基化细胞中表现欲产生之蛋白。
(19)如(18)之方法,其中所产生之去岩藻醣基化蛋白为抗体及/或其片段。
5.额外实施例
本发明的额外实施例包括,但不限于以下:
(1)一种产生去岩藻醣基化抗体的方法,包括:
将编码至少一经修饰的酵素的核酸导入至宿主细胞中,以在宿主细胞中产生去岩藻醣基化抗体。
(2)如(1)之方法,其中经修饰的酵素源自于gdp-甘露醣4,6-脱水酶(gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(fx)、或岩藻醣转移酶(fut)。
(3)如(1)之方法,其中经修饰的酵素源自于岩藻醣转移酶(fut)。
(4)如(1)之方法,其中经修饰的酵素源自于α-1,6-岩藻醣转移酶(fut8)。
(5)如(1)之方法,其中经修饰的酵素抑制宿主细胞中的野生型岩藻醣基化酵素的活性。
(6)如(1)之方法,其中经修饰的酵素抑制及/或降低宿主细胞中抗体的岩藻醣基化。
(7)如(1)之方法,其中去岩藻醣基化抗体具有增加的adcc。
(8)如(1)之方法,其中去岩藻醣基化抗体的cdc活性未被降低或抑制。
(9)一种产生去岩藻醣基化抗体的方法,包括:
a)提供宿主细胞,
b)将编码至少一经修饰的酵素的核酸导入至宿主细胞中,以及
c)在宿主细胞中产生去岩藻醣基化抗体。
(10)如(9)之方法,其中于步骤(a)中,宿主细胞包括至少一编码抗体的核酸。
(11)如(9)之方法,更将编码抗体的核酸导入至步骤(b)后的宿主细胞中。
(12)如(9)之方法,其中经修饰的酵素源自于gdp-甘露醣4,6-脱水酶(gmd)、gdp-4-酮基-6-脱氧-d-甘露醣异构酶-还原酶(fx)、及/或岩藻醣转移酶(fut)。
(13)如(9)之方法,其中经修饰的酵素源自于岩藻醣转移酶(fut)。
(14)如(9)之方法,其中经修饰的酵素源自于α-1,6-岩藻醣转移酶(fut8)。
(15)如(9)之方法,其中经修饰的酵素抑制宿主细胞中岩藻醣基化酵素的活性。
(16)如(9)之方法,其中经修饰的酵素抑制及降低宿主细胞中抗体的醣基化。
(17)如(9)之方法,其中去岩藻醣基化抗体具有增加的adcc。
(18)如(9)之方法,其中去岩藻醣基化抗体的cdc活性未被降低或抑制。
(19)一种以(1)或(9)之方法所产生的去岩藻醣基化抗体,其中去岩藻醣基化抗体具有增加的adcc活性。
(20)如(19)之方法,其中去岩藻醣基化抗体为人类抗体或其片段。
(21)如(19)之方法,其中去岩藻醣基化抗体维持原有的cdc活性。
(22)一种医药组成物,包括(19)之去岩藻醣基化抗体及药学上可接受之载体或赋形剂。
(23)一种无或具低岩藻醣基化的细胞,包括编码至少一经修饰的酵素的核酸。
额外的本发明特定实施例包括,但不限于以下实施例。
实施例
【实施例1】
岩藻醣基化途径中经修饰之酵素的制备及表现此经修饰之酵素的稳定
细胞株
1.细胞株
市售chodhfr(-)细胞株(atcccrl-9096),其为缺乏二氢叶酸还原酶活性的cho细胞突变株,购自于菌种保存及研究中心(culturecollectionandresearchcenter(ccrc),中国台湾)。将chodhfr(-)细胞株分成3个分开的培养物,且如下所示进行处理:
将第一培养物转染编码
将第二培养物转染编码
第三培养物为未经处理的细胞,且维持为chodhfr(-)细胞株。
2.编码fut8与gmd之经修饰的酵素之表现载体的构筑
构筑许多编码经修饰之酵素fut8与gmd的表现载体。
f83m、f8m1、f8m2、f8m3及f8d1突变型分别表示对α-1,6-岩藻醣转移酶,野生型fut8蛋白(genbankno.np_058589.2),的不同修饰。表1总结针对各fut8载体对于野生型核酸序列所做的修饰,及在所表现的酵素中所产生的胺基酸改变。特别是,f83m代表在野生型fut8蛋白中于r365a、d409a与d453a有3个修饰的突变型。f8m1、f8m2与f8m3分别代表野生型fut8蛋白中各自在k369e、d409k与s469v有一个修饰的突变型。f8d1代表于野生型fut8蛋白中,具有于第365至386位之胺基酸残基删除的突变型。
表2总结gmd载体对于野生型核酸序列所做的修饰,及在所表现的酵素中所产生的胺基酸改变。特别是,突变型gmd4m表示对gdp-甘露醣4,6-脱水酶,野生型gmd蛋白(genbankno.np_001233625.1,的修饰,其在野生型gmd蛋白中有4个突变,在t155a、e157a、y179a与k183a。
所有编码f83m、f8m1、f8m2、f8m3、f8d1及gmd4m的核酸序列由genedirex公司合成,并次选殖至phd表现载体(pcdna3.1hygro,invitrogen,carlsbad,ca,cat.no.v870-20,具dhfr基因)的paci/ecorv或bamhi/ecorv位置,以形成phd/f83m、phd/f8m1、phd/f8m2、phd/f8m3、phd/f8d1及phd/gmd4m质体。
3.表现经修饰的酵素之稳定重组细胞株的制备
将phd/f83m、phd/f8m1、phd/f8m2、phd/f8m3、phd/f8d1及phd/gmd4m质体藉由电穿孔(pa4000
a.rc79细胞
转染的rc79细胞株先培养于含0.1至0.25mg/ml潮霉素(hygromycin)的rc79培养基中(含有0.4μmmtx、0.5mg/ml遗传霉素(geneticin)、0.05mg/ml博来霉素(zeocin)、4mmglutamax-i和0.01%f-68的
b.hc59细胞
转染的hc59细胞株先培养于含0.1至0.25mg/ml潮霉素的hc59培养基(含有0.8μmmtx、0.5mg/ml遗传霉素、0.05mg/ml博来霉素与4mmglutamax-i的
c.chodhfr(-)细胞
chodhfr(-)细胞先培养于含4mmglutamax-i与0.1至0.25mg/ml潮霉素的
4.具低岩藻醣基化细胞的分离
在此实施例中使用罗丹明(rhodamine)标记的扁豆凝集素(lensculinarisagglutinin,lca)(vectorlaboratories,cat.rl-1042),来筛选具低岩藻醣基化的细胞。
所有的rc79、hc59及cho转染株受到含有潮霉素的初步筛选培养基作为筛选压力,接着使用可辨识n-连接寡醣之α-1,6-岩藻醣基化三甘露醣核心结构,并使表现此结构的细胞往细胞死亡途径的lca,来进行最终筛选。先以1.2x105细胞/ml,将rc79、hc59或cho的转染株种于含有0.4mg/mllca的2.5ml新鲜培养基中,且在第3或4天时计数细胞存活率。将细胞培养于此初步筛选培养基中,直到细胞存活率达到80%。在细胞存活率达到80%后,以1.2x105cells/ml,将细胞重新悬浮于含有梯度浓度之lca的新鲜筛选培养基中。lca筛选重复数次,直到lca最终浓度达到0.6至1.2mg/ml。
为分析细胞表面上的岩藻醣的量,以lca标志细胞,且以流式细胞术进行分析。首先,将细胞种于不含lca的完全培养基14天,以去除来自筛选试剂lca的讯号干扰。接着,以1ml冰冷pbs清洗3x105细胞2次,且重新悬浮于含1%牛血清白蛋白与5μg/mllca的200μl冰pbs中。在冰上培养30分钟后,以1ml冰冷pbs清洗细胞2次。将细胞重新悬浮于350μl冷的pbs中,并使用facscaliburtm流式细胞仪(bdbiosciences,sanjose,ca)进行分析。
接着,以10ml的冰冷pbs清洗1x107细胞2次,且重新悬浮于6.5ml含1%牛血清白蛋白与5μg/mllca的冰冷pbs中。在冰上培养30分钟后,以10ml冷的pbs清洗细胞2次。将细胞重新悬浮于1ml含1%热灭活的胎牛血清(gibco,cat.10091-148)与抗生素-抗霉菌剂(invitrogen,cat.15240062)的冰冷pbs中。
以facsariatm或influxtmcellsorter(bdbiosciences,sanjose,ca)分析及分类细胞。对于不同的细胞株,需要1-3次的分类,以产生具低岩藻醣基化细胞的同质族群。此外,使用clonepixtm2系统(molecular
5.表现
在以lca分离低岩藻醣基化chodhfr(-)细胞(c109f83m细胞)后,将编码
所获得的细胞为表现
【实施例2】
去岩藻醣基化抗体的表现及分析
1.抗体的表现与纯化
将实施例1中所获得的具低岩藻醣基化活性的细胞以批次或馈料批次培养,来表现抗体。由细胞所纯化的抗体进行单醣分析,以对fc区中的醣链进行定量分析。
将重组rc79细胞培养于含有4mmglutamax与0.01%f-68的
将重组hc59细胞培养于含有0.8μmmtx、0.5mg/ml遗传霉素、0.05mg/ml博来霉素、4mmglutamax-i与0.25mg/ml潮霉素的
每天常规地监测细胞培养的参数。使用血球计数器(hemocytometer)以锥蓝质(trypanblue)排除,来判定细胞密度及存活率。当细胞存活率低于60%时,藉由离心来收集条件培养基并以蛋白a树脂来纯化所表现的抗体。以5倍管柱体积的0.1mtris,ph8.3,来平衡蛋白a管柱,接着将样本添加至管柱中。以0.1mtris,ph8.3(2倍管柱体积)及pbs,ph6.5(10倍管柱体积)清洗掉未结合的蛋白。进一步以0.1m醋酸钠,ph6.5(10倍管柱体积)清洗管柱。最后,以0.1m甘胺酸,ph2.8冲提抗体,并以相同冲提体积的0.1mtris,ph8.3进行中和。
2.抗体的n-多醣图谱分析的判定
以acquity
流式细胞仪的结果显示,在所有细胞类型中,过度表现f83m蛋白的细胞表面上,lca的结合极低。类似地,在过度表现f8m1、f8m2、f8m3、f8d1或gmd4m蛋白的rc79细胞上,未侦测到lca结合(数据未显示)。
表3显示,具有未修饰的岩藻醣基化途径之rc79与hc59细胞以及藉由过度表现f83m经修饰的酵素,来修饰其岩藻醣基化途径的rc79与hc59细胞中所产生之抗体的n-多醣图谱。表3中的数据显示,具有未修饰岩藻醣基化途径之细胞中所产生的抗-cd20与抗-erbb2抗体大部分被重度岩藻醣基化。特别是,在这些细胞中,仅3.67%的抗-cd20与3.64%的抗-erbb2抗体被去岩藻醣基化。相较之下,过度表现f83m经修饰之酵素的细胞中所产生之抗体具有非常低的岩藻醣基化程度。特别是,在过度表现f83m经修饰之酵素的细胞中,约98.86-98.91%的抗-cd20与约92.12-96.52%的抗-erbb2抗体被去岩藻醣基化。
此外,表4显示具有未修饰的岩藻基化途径之rc79细胞以及藉由过度表现f8m1、f8m2、f8m3、f8d1或gmd4m经修饰的酵素之一,来修饰其岩藻醣基化途径的rc79细胞株中所产生之抗体的n-多醣图谱。表4的数据显示,具有未修饰的岩藻醣基化途径的rc79细胞中所产生之抗-cd20抗体大部分被重岩藻醣基化。特别是,在这些细胞中,仅3.67%的抗-cd20抗体为去岩藻醣基化。相较之下,过度表现经修饰的酵素之rc79细胞中所产生的抗体具有非常低的岩藻醣基化程度。特别是,由过度表现f8m1、f8m2、f8m3、f8d1或gmd4m经修饰的酵素之细胞所产生之抗-cd20抗体的去岩藻醣基化程度为约92.78%至约97.16%,如表4所示。
表4亦显示由过度表现fut8经修饰的酵素(f8m1、f8m2,f8m3,f8d1)之一的细胞所产生之抗体的去岩藻醣基化程度为95.70至97.16%,且由过度表现gmd经修饰的酵素(gmd4m)所生产之抗体的去岩藻醣基化程度为92.78%。这些结果证实,在过度表现fut8经修饰的酵素之细胞中所产生之抗体的去岩醣基化程度高于在过度表现gmd突变型蛋白的细胞中所生产之抗体。
表3及4的结果证实,已经设计来表现抗体的宿主细胞可转染表现岩藻醣基化途径中经修饰的酵素(fut8或gmd)的载体。此结果也显示,在这些转染细胞中所产生的抗体是去岩藻醣基化抗体。
此外,评估af97细胞株中所产生之抗体的岩藻醣基化程度。表5的结果显示,过度表现f83m经修饰的酵素之af97细胞中所产生之抗体具有非常低岩藻醣基化程度。特别是,97.83%的在af97细胞株中所产生之抗-cd20抗体被去岩藻醣基化。相较之下,市售
表5的结果显示,可在先转染编码经修饰的酵素之核酸,接着第二次转染编码抗体之核酸的细胞中,产生去岩藻醣基化抗体。因此,可使用本发明方法,将细胞修饰以产生去岩藻醣基化抗体。
3.重组细胞中fut8蛋白的表现
将rc79细胞与表现fut8经修饰的酵素(即,f8m1、f8m2、f8m3、或f8d1)的重组细胞的沉淀物,以含有磷酸酶抑制剂鸡尾酒(sigma-aldrich,cat.s8820)的1%tritonx-100裂解。以dctm(与清洁剂兼容)蛋白质分析(bio-rad)试剂,来确认裂解细胞的上清液中的蛋白浓度。将含有30μg蛋白的上清液的每个样品,以12.5%十二烷基磺酸钠-聚丙烯酰胺胶体电泳(sodiumdodecylsulfate-polyacrylamidegelelectrophoresis,sds-page)来分离并移至硝化纤维素膜上。于室温下,藉由使用含有120mmnacl、0.1%明胶(w/w)与0.1%
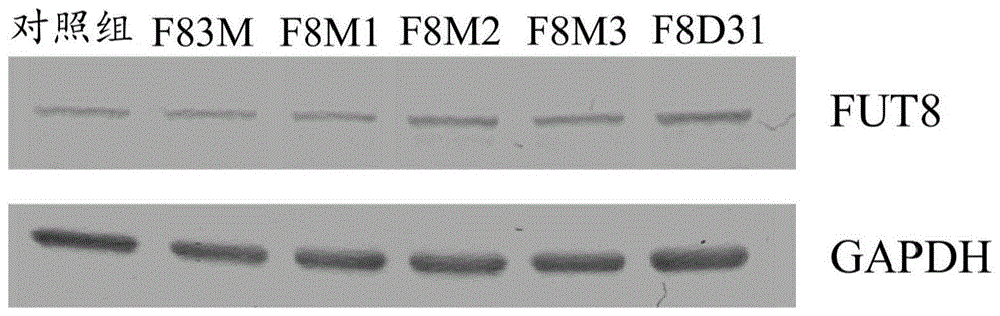
第1图为西方墨点法,其显示rc79亲源细胞中及表现经修饰的酵素之rc79重组细胞中的fut8蛋白的表现。重组细胞及rc79亲源细胞中fut8蛋白的表现量类似。这些结果表示,表现经修饰的酵素之rc79细胞中之去岩藻醣基化抗体的产生与fut8蛋白的表现量无关。这些结果显示fut8经修饰的酵素会干扰细胞中的野生型fut8蛋白,以抑制及/或降低岩藻醣基化途径,使重组细胞有效率地产生去岩藻醣基化抗体。
相较于其它依靠抑制或向下调控野生型fut8基因、或使用rna干扰,来减少fut8蛋白表现的方式,使用本发明方法产生去岩藻醣基化抗体的机制为新颖且独特的。
4.重组细胞的稳定性
评估表现f83m经修饰的酵素之rc79重组细胞的稳定性。
将rc79重组细胞培养在不含筛选试剂的培养基3个月。每周以流式细胞术分析,来监测细胞的岩藻醣基化,且每月以acquity
此外,在72天的期间,评估在5个稳定的rc79f83m细胞株(r4-r8)中所产生之抗-cd20抗体的去岩藻醣基化程度。如表6所示,所有rc79f83m细胞株在72天的研究中,皆产生高度去岩藻醣基化抗体。
此研究的这些结果证明,藉由本发明方法制备的表现经修饰的酵素之重组细胞株为稳定的且长时间产生高度去岩藻醣基化的抗体。
【实施例3】
去岩藻醣基化抗体的adcc活性
为了评估实施例2所获得之经纯化的抗-cd20抗体的体外细胞毒性,依照下述方法测量adcc活性。
1.效应细胞溶液的制备
将自健康捐赠者的人类外围血液(100ml)加至含有肝素钠的
2.目标细胞溶液的制备
将来自75t烧瓶的细胞悬浮液以1,000rpm离心5分钟,去除上清液,然后以10ml1xpbs清洗。将经清洗的细胞以1,200rpm离心5分钟,去除上清液。以rpmi分析培养基,将细胞重新悬浮,以制备5x105细胞/ml的目标细胞溶液。将目标细胞溶液(40μl的5x105细胞/ml)加至v底的96孔细胞培养盘的孔洞中。接着,分别将20μl的经制备的市售
3.adcc活性分析
将效应细胞溶液(40μl的8x105效应细胞/孔洞)或40μl的rpmi分析培养基加至盘中,以与目标细胞溶液反应。将盘子以300xg离心4分钟。将盘子培养于37℃,5%co24小时。在收集上清液前,将cytotox
将乳酸脱氢酶(lactatedehydrogenase,ldh)(2μl)加至10ml之ldh阳性对照稀释剂中,以制备ldh阳性对照溶液。将制备的ldh阳性对照组溶液(50μl)添加至96孔平底分析盘的孔洞中。
将ldh重建基质混合物(50μl)添加至分析盘的每个测试孔洞。将盘子覆盖,并于室温下于黑暗中培养30分钟。将终止溶液(50μl)添加至盘子之每个测试孔洞。在加入终止溶液后,立即记录波长490nm的吸光值。使用各组(s、pbmc、t、e和tmax)空白移除吸光值,以下列算式计算adcc活性。
其中s为样本(目标细胞+pbmc+抗-cd20抗体)的ldh释放的吸光值;pbmc为目标细胞及pbmc的ldh释放的吸光值;e为pbmc的ldh释放的吸光值;t为目标细胞的自发性ldh释放的吸光值;且tmax为目标细胞的最大ldh释放的吸光值。
相较于市售
如表7所示,来自rc79f83mr1细胞株之去岩藻醣基化抗-cd20抗体的ec50显著低于市售
此研究的这些结果证明去岩藻醣基化抗-cd20抗体(r1细胞株)展现的adcc活性为岩藻醣基化抗-cd20抗体
【实施例4】
去岩藻醣基化抗体的结合亲和性
使用偶联至具有胺偶联套组之
以10μl/分钟的速率,将his标记之fcγriiia重组蛋白(1μg/ml),注入至抗-his抗体固定之cm5芯片上,持续20秒。
将来自细胞株1的去岩藻醣基化抗-cd20抗体(5、10、20、40及80nm)、市售岩藻醣基化抗-cd20抗体
利用
第4a至4c图显示经测试之三种抗体的典型spr应感图谱。典型的spr应感图谱显示,此分析所使用的条件(例如,结合时间、解离时间和抗体浓度范围)是适当的。此外,三种抗体之chi2值低于1/10xrmax值,表示1∶1langmuir模型适合作为所有三种抗体的应感图谱适配。
如表8所示,去岩藻醣基化抗-cd20抗体(r1细胞株)对fcγriiia的结合亲和性比
此研究的这些结果证实,根据本发明制备的去岩藻醣基化抗-cd20抗体(r1细胞株)具有比市售岩藻醣基化抗-cd20抗体
【实施例5】
去岩藻醣基化抗体的cdc活性
评估以本发明方法所产生之去岩藻醣基化抗体的cdc活性。
以rpmi培养基培养daudi细胞,且当细胞密度达到1×106细胞/ml时,进行继代培养(继代培养密度:2-3x105细胞/ml)。收集daudi细胞,并以300rpm离心5分钟。以rpmi培养基重新悬浮细胞,以制备浓度为1x105细胞/ml的细胞悬浮液。在重新悬浮后,将100μl的细胞悬浮液或100μl的rpmi培养基种至白色96孔盘的孔洞中。
以120μg/ml至0.234μg/ml的浓度,将市售可得
第5图显示去岩藻醣基化抗-cd20抗体(r1细胞株)之cdc活性相当于
【实施例6】
去岩藻醣基化抗体在动物模型中的功效证据
在此实施例中,使用b细胞淋巴瘤皮下异种移植模型,来证明本发明的去岩藻醣基化抗体的抗肿瘤功效。su-dhl-4为b细胞淋巴瘤细胞株,其在细胞膜上表现大量的cd20,且可成长并皮下形成固态肿瘤。因此,开发scid/beige小鼠中的异种移植模型,以比较去岩藻醣基化抗体(r1细胞株)与市售可得
以rpmi培养基(cm),将su-dhl-4细胞培养于烧瓶中。当细胞浓度达0.8-1.0x106细胞/ml时,收集细胞悬浮液并以300g离心5分钟,以去除上清液。以含有一些条件培养基的新培养基(新鲜cm∶条件cm=9∶1),将细胞重新悬浮。细胞以1∶2至1∶10的比例(接种的细胞数量:总获得细胞数量)继代培养,且细胞浓度为至少1x105细胞/ml。将培养盘培养于37℃。su-dhl-4细胞培养于5个150t的烧瓶。当细胞浓度到达0.8至1.0x106细胞/ml时,将细胞悬浮液收集至50ml试管,之后以1,200rpm离心5分钟,以移除上清液。使用无血清rpmi培养基,将细胞浓度调整至1x108细胞/ml。在冰上,使用具18g针头之预冷针筒,于50ml试管中混合细胞悬浮液与等体积的
当肿瘤体积达约200mm3(198.25±55.53mm3)时,其在接种肿瘤后约20天发生,将小鼠分成三组,每组5只,之后以食盐水(载体),市售可得
如第6图所示,去岩藻醣基化抗-cd20抗体(r1细胞株)显示比
去岩藻醣基化抗-cd20抗体(r1细胞株)治疗组的肿瘤重量显著小于仅载体组的肿瘤重量(p<0.001),如第7图中所示。然而,在相同剂量下,相较于载体组,
在治疗期间,所有组的小鼠体重逐渐增加,如第8图所示。
这些研究的结果显示去岩藻醣基化抗-cd20抗体(r1细胞株)是安全的。
表1、于分析中所使用的fut8酵素的修饰
1基于野生型fut8dna序列的seqidno:1的dna序列位置
2基于野生型fut8蛋白质序列的seqidno:2的胺基酸位置
3n/a=不适用
表6、过度表现f83m经修饰的酵素之细胞所产生之去岩藻醣基化抗
-cd20抗体的稳定性
表7、岩藻醣基化及去岩藻醣基化抗-cd20抗体的ec50值及adcc活性
表8、
表9、各组中经治疗之小鼠的肿瘤体积、肿瘤重量与体重
数值为:平均±s.d.(s.c.)。
序列表
<110>联亚药业
联合生物制药
彭文君
陈惠蓉
<120>去岩藻醣基化抗体及其制造
<130>ubip1004a-us
<140>tbd
<141>2018-05-08
<160>17
<170>patentinversion3.5
<210>1
<211>1728
<212>dna
<213>小鼠
<400>1
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtgcacgtgaggaggaccgacaaggtgggcaccgaggccgccttccaccccatcgag1140
gagtacatggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggac1200
aagaagagggtgtacctggccaccgacgaccccaccctgctgaaggaggccaagaccaag1260
tactccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaac1320
aggtacaccgagaactccctgaggggcgtgatcctggacatccacttcctgtcccaggcc1380
gacttcctggtgtgcaccttctcctcccaggtgtgcagggtggcctacgagatcatgcag1440
accctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggc1500
ggccagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggag1560
atccccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactcc1620
aagggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggag1680
aagatcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1728
<210>2
<211>575
<212>prt
<213>人类
<400>2
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyvalhisvalargargthrasp
355360365
lysvalglythrglualaalaphehisproilegluglutyrmetval
370375380
hisvalglugluhispheglnleuleualaargargmetglnvalasp
385390395400
lyslysargvaltyrleualathraspaspprothrleuleulysglu
405410415
alalysthrlystyrserasntyrglupheileseraspasnserile
420425430
sertrpseralaglyleuhisasnargtyrthrgluasnserleuarg
435440445
glyvalileleuaspilehispheleuserglnalaasppheleuval
450455460
cysthrpheserserglnvalcysargvalalatyrgluilemetgln
465470475480
thrleuhisproaspalaseralaasnphehisserleuaspaspile
485490495
tyrtyrpheglyglyglnasnalahisasnglnilealavaltyrpro
500505510
hislysproargthrgluglugluileprometgluproglyaspile
515520525
ileglyvalalaglyasnhistrpaspglytyrserlysglyileasn
530535540
arglysleuglylysthrglyleutyrprosertyrlysvalargglu
545550555560
lysilegluthrvallystyrprothrtyrproglualaglulys
565570575
<210>3
<211>1728
<212>dna
<213>人工序列
<220>
<223>f83m经修饰之酵素的核酸序列,野生型fut8基因第1093-1095、1225-1227与
1357-1359位置被改变
<400>3
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtgcacgtggccaggaccgacaaggtgggcaccgaggccgccttccaccccatcgag1140
gagtacatggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggac1200
aagaagagggtgtacctggccaccgccgaccccaccctgctgaaggaggccaagaccaag1260
tactccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaac1320
aggtacaccgagaactccctgaggggcgtgatcctggccatccacttcctgtcccaggcc1380
gacttcctggtgtgcaccttctcctcccaggtgtgcagggtggcctacgagatcatgcag1440
accctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggc1500
ggccagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggag1560
atccccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactcc1620
aagggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggag1680
aagatcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1728
<210>4
<211>575
<212>prt
<213>人工序列
<220>
<223>f83m经修饰之酵素的胺基酸序列,野生型fut8蛋白第365、409与453位置的残基被改变
<400>4
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyvalhisvalalaargthrasp
355360365
lysvalglythrglualaalaphehisproilegluglutyrmetval
370375380
hisvalglugluhispheglnleuleualaargargmetglnvalasp
385390395400
lyslysargvaltyrleualathralaaspprothrleuleulysglu
405410415
alalysthrlystyrserasntyrglupheileseraspasnserile
420425430
sertrpseralaglyleuhisasnargtyrthrgluasnserleuarg
435440445
glyvalileleualailehispheleuserglnalaasppheleuval
450455460
cysthrpheserserglnvalcysargvalalatyrgluilemetgln
465470475480
thrleuhisproaspalaseralaasnphehisserleuaspaspile
485490495
tyrtyrpheglyglyglnasnalahisasnglnilealavaltyrpro
500505510
hislysproargthrgluglugluileprometgluproglyaspile
515520525
ileglyvalalaglyasnhistrpaspglytyrserlysglyileasn
530535540
arglysleuglylysthrglyleutyrprosertyrlysvalargglu
545550555560
lysilegluthrvallystyrprothrtyrproglualaglulys
565570575
<210>5
<211>1728
<212>dna
<213>人工序列
<220>
<223>f8m1经修饰之酵素的核酸序列,野生型fut8基因第1105-1107位置被改变
<400>5
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtgcacgtgaggaggaccgacgaggtgggcaccgaggccgccttccaccccatcgag1140
gagtacatggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggac1200
aagaagagggtgtacctggccaccgacgaccccaccctgctgaaggaggccaagaccaag1260
tactccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaac1320
aggtacaccgagaactccctgaggggcgtgatcctggacatccacttcctgtcccaggcc1380
gacttcctggtgtgcaccttctcctcccaggtgtgcagggtggcctacgagatcatgcag1440
accctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggc1500
ggccagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggag1560
atccccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactcc1620
aagggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggag1680
aagatcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1728
<210>6
<211>575
<212>prt
<213>人工序列
<220>
<223>f8m1经修饰之酵素的胺基酸序列,野生型fut8蛋白第369位置的残基被改变
<400>6
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyvalhisvalargargthrasp
355360365
gluvalglythrglualaalaphehisproilegluglutyrmetval
370375380
hisvalglugluhispheglnleuleualaargargmetglnvalasp
385390395400
lyslysargvaltyrleualathraspaspprothrleuleulysglu
405410415
alalysthrlystyrserasntyrglupheileseraspasnserile
420425430
sertrpseralaglyleuhisasnargtyrthrgluasnserleuarg
435440445
glyvalileleuaspilehispheleuserglnalaasppheleuval
450455460
cysthrpheserserglnvalcysargvalalatyrgluilemetgln
465470475480
thrleuhisproaspalaseralaasnphehisserleuaspaspile
485490495
tyrtyrpheglyglyglnasnalahisasnglnilealavaltyrpro
500505510
hislysproargthrgluglugluileprometgluproglyaspile
515520525
ileglyvalalaglyasnhistrpaspglytyrserlysglyileasn
530535540
arglysleuglylysthrglyleutyrprosertyrlysvalargglu
545550555560
lysilegluthrvallystyrprothrtyrproglualaglulys
565570575
<210>7
<211>1728
<212>dna
<213>人工序列
<220>
<223>f8m2经修饰之酵素的核酸序列,野生型fut8基因第1225-1227位置被改变
<400>7
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtgcacgtgaggaggaccgacaaggtgggcaccgaggccgccttccaccccatcgag1140
gagtacatggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggac1200
aagaagagggtgtacctggccaccaaggaccccaccctgctgaaggaggccaagaccaag1260
tactccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaac1320
aggtacaccgagaactccctgaggggcgtgatcctggacatccacttcctgtcccaggcc1380
gacttcctggtgtgcaccttctcctcccaggtgtgcagggtggcctacgagatcatgcag1440
accctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggc1500
ggccagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggag1560
atccccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactcc1620
aagggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggag1680
aagatcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1728
<210>8
<211>575
<212>prt
<213>人工序列
<220>
<223>f8m2经修饰之酵素的胺基酸序列,野生型fut8蛋白第409位置的残基被改变
<400>8
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyvalhisvalargargthrasp
355360365
lysvalglythrglualaalaphehisproilegluglutyrmetval
370375380
hisvalglugluhispheglnleuleualaargargmetglnvalasp
385390395400
lyslysargvaltyrleualathrlysaspprothrleuleulysglu
405410415
alalysthrlystyrserasntyrglupheileseraspasnserile
420425430
sertrpseralaglyleuhisasnargtyrthrgluasnserleuarg
435440445
glyvalileleuaspilehispheleuserglnalaasppheleuval
450455460
cysthrpheserserglnvalcysargvalalatyrgluilemetgln
465470475480
thrleuhisproaspalaseralaasnphehisserleuaspaspile
485490495
tyrtyrpheglyglyglnasnalahisasnglnilealavaltyrpro
500505510
hislysproargthrgluglugluileprometgluproglyaspile
515520525
ileglyvalalaglyasnhistrpaspglytyrserlysglyileasn
530535540
arglysleuglylysthrglyleutyrprosertyrlysvalargglu
545550555560
lysilegluthrvallystyrprothrtyrproglualaglulys
565570575
<210>9
<211>1728
<212>dna
<213>人工序列
<220>
<223>f8m3经修饰之酵素的核酸序列,野生型fut8基因第1405-1407位置被改变
<400>9
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtgcacgtgaggaggaccgacaaggtgggcaccgaggccgccttccaccccatcgag1140
gagtacatggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggac1200
aagaagagggtgtacctggccaccgacgaccccaccctgctgaaggaggccaagaccaag1260
tactccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaac1320
aggtacaccgagaactccctgaggggcgtgatcctggacatccacttcctgtcccaggcc1380
gacttcctggtgtgcaccttcgtgtcccaggtgtgcagggtggcctacgagatcatgcag1440
accctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggc1500
ggccagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggag1560
atccccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactcc1620
aagggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggag1680
aagatcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1728
<210>10
<211>575
<212>prt
<213>人工序列
<220>
<223>f8m3经修饰之酵素的胺基酸序列,野生型fut8蛋白第469位置的残基被改变
<400>10
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyvalhisvalargargthrasp
355360365
lysvalglythrglualaalaphehisproilegluglutyrmetval
370375380
hisvalglugluhispheglnleuleualaargargmetglnvalasp
385390395400
lyslysargvaltyrleualathraspaspprothrleuleulysglu
405410415
alalysthrlystyrserasntyrglupheileseraspasnserile
420425430
sertrpseralaglyleuhisasnargtyrthrgluasnserleuarg
435440445
glyvalileleuaspilehispheleuserglnalaasppheleuval
450455460
cysthrpheservalglnvalcysargvalalatyrgluilemetgln
465470475480
thrleuhisproaspalaseralaasnphehisserleuaspaspile
485490495
tyrtyrpheglyglyglnasnalahisasnglnilealavaltyrpro
500505510
hislysproargthrgluglugluileprometgluproglyaspile
515520525
ileglyvalalaglyasnhistrpaspglytyrserlysglyileasn
530535540
arglysleuglylysthrglyleutyrprosertyrlysvalargglu
545550555560
lysilegluthrvallystyrprothrtyrproglualaglulys
565570575
<210>11
<211>1665
<212>dna
<213>人工序列
<220>
<223>f8d1经修饰之酵素的核酸序列,野生型fut8基因第1087-1149位置被删除
<400>11
atgagggcctggaccggctcctggaggtggatcatgctgatcctgttcgcctggggcacc60
ctgctgttctacatcggcggccacctggtgagggacaacgaccaccccgaccactcctcc120
agggagctgtccaagatcctggccaagctggagaggctgaagcagcagaacgaggacctg180
aggaggatggccgagtccctgaggatccccgagggccccatcgaccagggcaccgccacc240
ggcagggtgagggtgctggaggagcagctggtgaaggccaaggagcagatcgagaactac300
aagaagcaggccaggaacggcctgggcaaggaccacgagatcctgaggaggaggatcgag360
aacggcgccaaggagctgtggttcttcctgcagtccgagctgaagaagctgaagcacctg420
gagggcaacgagctgcagaggcacgccgacgagatcctgctggacctgggccaccacgag480
aggtccatcatgaccgacctgtactacctgtcccagaccgacggcgccggcgactggagg540
gagaaggaggccaaggacctgaccgagctggtgcagaggaggatcacctacctgcagaac600
cccaaggactgctccaaggccaggaagctggtgtgcaacatcaacaagggctgcggctac660
ggctgccagctgcaccacgtggtgtactgcttcatgatcgcctacggcacccagaggacc720
ctgatcctggagtcccagaactggaggtacgccaccggcggctgggagaccgtgttcagg780
cccgtgtccgagacctgcaccgacaggtccggcctgtccaccggccactggtccggcgag840
gtgaacgacaagaacatccaggtggtggagctgcccatcgtggactccctgcaccccagg900
cccccctacctgcccctggccgtgcccgaggacctggccgacaggctgctgagggtgcac960
ggcgaccccgccgtgtggtgggtgtcccagttcgtgaagtacctgatcaggccccagccc1020
tggctggagaaggagatcgaggaggccaccaagaagctgggcttcaagcaccccgtgatc1080
ggcgtggtgcacgtggaggagcacttccagctgctggccaggaggatgcaggtggacaag1140
aagagggtgtacctggccaccgacgaccccaccctgctgaaggaggccaagaccaagtac1200
tccaactacgagttcatctccgacaactccatctcctggtccgccggcctgcacaacagg1260
tacaccgagaactccctgaggggcgtgatcctggacatccacttcctgtcccaggccgac1320
ttcctggtgtgcaccttctcctcccaggtgtgcagggtggcctacgagatcatgcagacc1380
ctgcaccccgacgcctccgccaacttccactccctggacgacatctactacttcggcggc1440
cagaacgcccacaaccagatcgccgtgtacccccacaagcccaggaccgaggaggagatc1500
cccatggagcccggcgacatcatcggcgtggccggcaaccactgggacggctactccaag1560
ggcatcaacaggaagctgggcaagaccggcctgtacccctcctacaaggtgagggagaag1620
atcgagaccgtgaagtaccccacctaccccgaggccgagaagtga1665
<210>12
<211>548
<212>prt
<213>人工序列
<220>
<223>f8d1经修饰之酵素的胺基酸序列,野生型fut8蛋白第365-386位置的残基被删
除
<400>12
metargalatrpthrglysertrpargtrpilemetleuileleuphe
151015
alatrpglythrleuleuphetyrileglyglyhisleuvalargasp
202530
asnasphisproasphisserserarggluleuserlysileleuala
354045
lysleugluargleulysglnglnasngluaspleuargargmetala
505560
gluserleuargileprogluglyproileaspglnglythralathr
65707580
glyargvalargvalleuglugluglnleuvallysalalysglugln
859095
ilegluasntyrlyslysglnalaargasnglyleuglylysasphis
100105110
gluileleuargargargilegluasnglyalalysgluleutrpphe
115120125
pheleuglnsergluleulyslysleulyshisleugluglyasnglu
130135140
leuglnarghisalaaspgluileleuleuaspleuglyhishisglu
145150155160
argserilemetthraspleutyrtyrleuserglnthraspglyala
165170175
glyasptrpargglulysglualalysaspleuthrgluleuvalgln
180185190
argargilethrtyrleuglnasnprolysaspcysserlysalaarg
195200205
lysleuvalcysasnileasnlysglycysglytyrglycysglnleu
210215220
hishisvalvaltyrcysphemetilealatyrglythrglnargthr
225230235240
leuileleugluserglnasntrpargtyralathrglyglytrpglu
245250255
thrvalpheargprovalsergluthrcysthraspargserglyleu
260265270
serthrglyhistrpserglygluvalasnasplysasnileglnval
275280285
valgluleuproilevalaspserleuhisproargproprotyrleu
290295300
proleualavalprogluaspleualaaspargleuleuargvalhis
305310315320
glyaspproalavaltrptrpvalserglnphevallystyrleuile
325330335
argproglnprotrpleuglulysgluilegluglualathrlyslys
340345350
leuglyphelyshisprovalileglyhispheglnleuleualaarg
355360365
argmetglnvalasplyslysargvaltyrleualathraspasppro
370375380
thrleuleulysglualalysthrlystyrserasntyrglupheile
385390395400
seraspasnserilesertrpseralaglyleuhisasnargtyrthr
405410415
gluasnserleuargglyvalileleuaspilehispheleusergln
420425430
alaasppheleuvalcysthrpheserserglnvalcysargvalala
435440445
tyrgluilemetglnthrleuhisproaspalaseralaasnphehis
450455460
serleuaspaspiletyrtyrpheglyglyglnasnalahisasngln
465470475480
ilealavaltyrprohislysproargthrgluglugluilepromet
485490495
gluproglyaspileileglyvalalaglyasnhistrpaspglytyr
500505510
serlysglyileasnarglysleuglylysthrglyleutyrproser
515520525
tyrlysvalargglulysilegluthrvallystyrprothrtyrpro
530535540
glualaglulys
545
<210>13
<211>1119
<212>dna
<213>小鼠
<400>13
atggcccacgcccccgcctcctgcccctcctccaggaactccggcgacggcgacaagggc60
aagcccaggaaggtggccctgatcaccggcatcaccggccaggacggctcctacctggcc120
gagttcctgctggagaagggctacgaggtgcacggcatcgtgaggaggtcctcctccttc180
aacaccggcaggatcgagcacctgtacaagaacccccaggcccacatcgagggcaacatg240
aagctgcactacggcgacctgaccgactccacctgcctggtgaagatcatcaacgaggtg300
aagcccaccgagatctacaacctgggcgcccagtcccacgtgaagatctccttcgacctg360
gccgagtacaccgccgacgtggacggcgtgggcaccctgaggctgctggacgccatcaag420
acctgcggcctgatcaactccgtgaagttctaccaggcctccacctccgagctgtacggc480
aaggtgcaggagatcccccagaaggagaccacccccttctaccccaggtccccctacggc540
gccgccaagctgtacgcctactggatcgtggtgaacttcagggaggcctacaacctgttc600
gccgtgaacggcatcctgttcaaccacgagtcccccaggaggggcgccaacttcgtgacc660
aggaagatctccaggtccgtggccaagatctacctgggccagctggagtgcttctccctg720
ggcaacctggacgccaagagggactggggccacgccaaggactacgtggaggccatgtgg780
ctgatgctgcagaacgacgagcccgaggacttcgtgatcgccaccggcgaggtgcactcc840
gtgagggagttcgtggagaagtccttcatgcacatcggcaagaccatcgtgtgggagggc900
aagaacgagaacgaggtgggcaggtgcaaggagaccggcaagatccacgtgaccgtggac960
ctgaagtactacaggcccaccgaggtggacttcctgcagggcgactgctccaaggcccag1020
cagaagctgaactggaagcccagggtggccttcgacgagctggtgagggagatggtgcag1080
gccgacgtggagctgatgaggaccaaccccaacgcctga1119
<210>14
<211>372
<212>prt
<213>小鼠
<400>14
metalahisalaproalasercysproserserargasnserglyasp
151015
glyasplysglylysproarglysvalalaleuilethrglyilethr
202530
glyglnaspglysertyrleualaglupheleuleuglulysglytyr
354045
gluvalhisglyilevalargargserserserpheasnthrglyarg
505560
ilegluhisleutyrlysasnproglnalahisilegluglyasnmet
65707580
lysleuhistyrglyaspleuthraspserthrcysleuvallysile
859095
ileasngluvallysprothrgluiletyrasnleuglyalaglnser
100105110
hisvallysileserpheaspleualaglutyrthralaaspvalasp
115120125
glyvalglythrleuargleuleuaspalailelysthrcysglyleu
130135140
ileasnservallysphetyrglnalaserthrsergluleutyrgly
145150155160
lysvalglngluileproglnlysgluthrthrprophetyrproarg
165170175
serprotyrglyalaalalysleutyralatyrtrpilevalvalasn
180185190
pheargglualatyrasnleuphealavalasnglyileleupheasn
195200205
hisgluserproargargglyalaasnphevalthrarglysileser
210215220
argservalalalysiletyrleuglyglnleuglucyspheserleu
225230235240
glyasnleuaspalalysargasptrpglyhisalalysasptyrval
245250255
glualamettrpleumetleuglnasnaspgluprogluasppheval
260265270
ilealathrglygluvalhisservalarggluphevalglulysser
275280285
phemethisileglylysthrilevaltrpgluglylysasngluasn
290295300
gluvalglyargcyslysgluthrglylysilehisvalthrvalasp
305310315320
leulystyrtyrargprothrgluvalasppheleuglnglyaspcys
325330335
serlysalaglnglnlysleuasntrplysproargvalalapheasp
340345350
gluleuvalargglumetvalglnalaaspvalgluleumetargthr
355360365
asnproasnala
370
<210>15
<211>1119
<212>dna
<213>人工序列
<220>
<223>gmd4m经修饰之酵素的核酸序列,野生型gmd基因第463-465、469-471、535-537
与547-549位置被改变
<400>15
atggcccacgcccccgcctcctgcccctcctccaggaactccggcgacggcgacaagggc60
aagcccaggaaggtggccctgatcaccggcatcaccggccaggacggctcctacctggcc120
gagttcctgctggagaagggctacgaggtgcacggcatcgtgaggaggtcctcctccttc180
aacaccggcaggatcgagcacctgtacaagaacccccaggcccacatcgagggcaacatg240
aagctgcactacggcgacctgaccgactccacctgcctggtgaagatcatcaacgaggtg300
aagcccaccgagatctacaacctgggcgcccagtcccacgtgaagatctccttcgacctg360
gccgagtacaccgccgacgtggacggcgtgggcaccctgaggctgctggacgccatcaag420
acctgcggcctgatcaactccgtgaagttctaccaggcctccgcctccgccctgtacggc480
aaggtgcaggagatcccccagaaggagaccacccccttctaccccaggtcccccgccggc540
gccgccgccctgtacgcctactggatcgtggtgaacttcagggaggcctacaacctgttc600
gccgtgaacggcatcctgttcaaccacgagtcccccaggaggggcgccaacttcgtgacc660
aggaagatctccaggtccgtggccaagatctacctgggccagctggagtgcttctccctg720
ggcaacctggacgccaagagggactggggccacgccaaggactacgtggaggccatgtgg780
ctgatgctgcagaacgacgagcccgaggacttcgtgatcgccaccggcgaggtgcactcc840
gtgagggagttcgtggagaagtccttcatgcacatcggcaagaccatcgtgtgggagggc900
aagaacgagaacgaggtgggcaggtgcaaggagaccggcaagatccacgtgaccgtggac960
ctgaagtactacaggcccaccgaggtggacttcctgcagggcgactgctccaaggcccag1020
cagaagctgaactggaagcccagggtggccttcgacgagctggtgagggagatggtgcag1080
gccgacgtggagctgatgaggaccaaccccaacgcctga1119
<210>16
<211>372
<212>prt
<213>人工序列
<220>
<223>gmd4m经修饰之酵素的胺基酸序列,野生型gmd蛋白第155、157、179与183
位置的残基被改变
<400>16
metalahisalaproalasercysproserserargasnserglyasp
151015
glyasplysglylysproarglysvalalaleuilethrglyilethr
202530
glyglnaspglysertyrleualaglupheleuleuglulysglytyr
354045
gluvalhisglyilevalargargserserserpheasnthrglyarg
505560
ilegluhisleutyrlysasnproglnalahisilegluglyasnmet
65707580
lysleuhistyrglyaspleuthraspserthrcysleuvallysile
859095
ileasngluvallysprothrgluiletyrasnleuglyalaglnser
100105110
hisvallysileserpheaspleualaglutyrthralaaspvalasp
115120125
glyvalglythrleuargleuleuaspalailelysthrcysglyleu
130135140
ileasnservallysphetyrglnalaseralaseralaleutyrgly
145150155160
lysvalglngluileproglnlysgluthrthrprophetyrproarg
165170175
serproalaglyalaalaalaleutyralatyrtrpilevalvalasn
180185190
pheargglualatyrasnleuphealavalasnglyileleupheasn
195200205
hisgluserproargargglyalaasnphevalthrarglysileser
210215220
argservalalalysiletyrleuglyglnleuglucyspheserleu
225230235240
glyasnleuaspalalysargasptrpglyhisalalysasptyrval
245250255
glualamettrpleumetleuglnasnaspgluprogluasppheval
260265270
ilealathrglygluvalhisservalarggluphevalglulysser
275280285
phemethisileglylysthrilevaltrpgluglylysasngluasn
290295300
gluvalglyargcyslysgluthrglylysilehisvalthrvalasp
305310315320
leulystyrtyrargprothrgluvalasppheleuglnglyaspcys
325330335
serlysalaglnglnlysleuasntrplysproargvalalapheasp
340345350
gluleuvalargglumetvalglnalaaspvalgluleumetargthr
355360365
asnproasnala
370
<210>17
<211>63
<212>dna
<213>人工序列
<220>
<223>f8d1突变基因中被删除的核酸序列
<400>17
cacgtgaggaggaccgacaaggtgggcaccgaggccgccttccaccccatcgaggagtac60
atg63
- 还没有人留言评论。精彩留言会获得点赞!