一种检测神经管畸形的分子标志物及其应用的制作方法
本发明涉及生物领域,具体涉及一种检测神经管畸形的分子标志物及其应用。
背景技术:
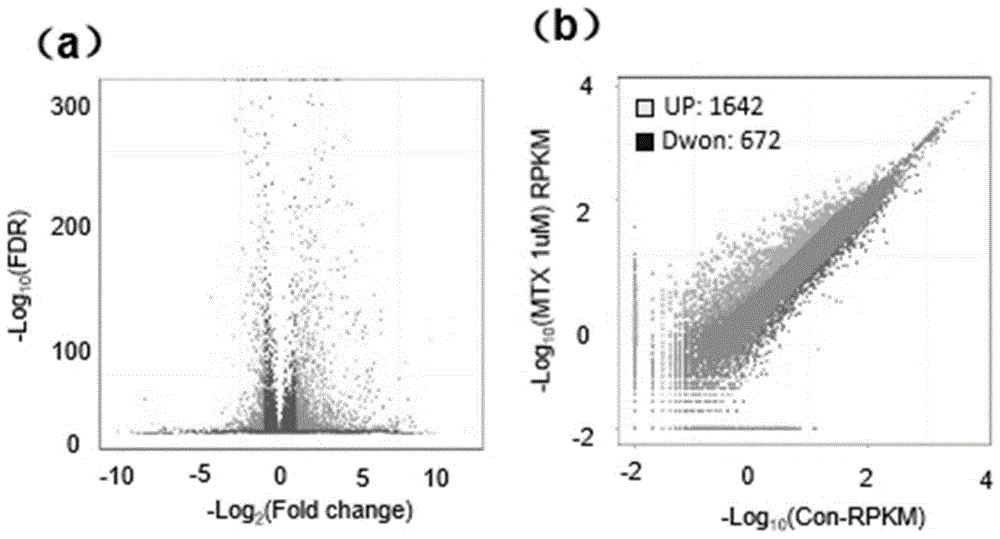
:神经管畸形(neuraltubedefects,ntds)是一种高发的出生缺陷性疾病,是出生缺陷中发病率和死亡率最高的类型,也是影响我国乃至世界人口素质最常见和最严重的出生缺陷性疾病。ntds通常发生于怀孕的早期,是由于神经管不闭合或闭合不全而造成的头部至脊柱部位畸形。从无脑到轻度脊柱裂不同程度的畸形:包括无脑、露脑、小头畸形、大头畸形、脑积水、脑脊膜膨出、颅脊柱裂和脊柱裂。新生儿中ntds畸形率高达0.1-0.5%,其病因和发病机理至今仍不十分清楚。神经管的发育是一个复杂的多步骤过程,是基因-基因,基因-环境和基因-营养共同作用的结果,受基因严格控制,环境因素也起重要作用。神经管是体内形成结构最复杂的一个系统,发育最早,结束最晚。在发育过程当中,若受到遗传因素或环境因素的干扰,超过胚胎的代偿能力就可能会发生ntds。这一时期作为神经管发育的关键时期,对孕妇体内生化代谢中物质的增加或少是极为敏感的。根据动物实验、临床观察和流行病学研究认为遗传因素(多基因遗传)和环境因素(叶酸维生素缺乏、高热、酒精及药物致畸等)都会干扰神经管的闭合。近年来对于人类ntds的发病机理的研究主要包括基因表达调控的改变(启动子和调控元件的突变)和表观遗传调控的分析,大多关注于基因及特定位点的基因多态性与ntds的关系。然而,对叶酸通路相关基因的突变位点所进行的关联分析并未找到造成ntds的一致风险因子,同一基因的同一位点在某些人群中与ntds相关,在另外一些人群中却不相关。神经胚发育过程中有多种信号的参与,作为网状相连的信号通路,如果有某一环节出问题就可能导致机体代谢失调,基因调控紊乱,进而表现出疾病状况。近年来,研究人员一直在试图寻找ntds的候选基因,研究数据主要来源于生物化学和发育学途径、动物模型以及定位候选基因三方面,由于小鼠神经管发育机制与人类神经管发育过程十分接近,因而成为了人们研究ntds发生机制最主要的一种模式动物。近年来,在小鼠模型中已有超过200个基因经研究发现与ntds相关,ntds相关基因主要编码参与细胞信号转导、调节转录、抑制肿瘤生长及染色质重塑相关的蛋白质。目前,研究发现有几条信号通路无疑是ntds发生发展过程的重要的调控环节,主要涉及:wnt/β-catenin信号通路(参与调控胚胎神经管的闭合)、pcp信号通路(参与神经管起始闭合)、hh/shh信号通路(过激活导致ntds,导致神经管闭合不全,形成低位脊柱裂)、bmp信号通路(调控中胚层形成、神经诱导、神经管背腹化的形成)、notch信号通路(通过调节神经发生参与神经管闭合)。这些通路相关基因可以作为人类ntds,尤其是颅脊柱裂研究的候选基因。例如,刘阳(刘阳.wnt/pcp-jnk信号通路在ntds发生及牛磺酸预防ntds的介导作用.宁夏医科大学,2014.)研究发现,wnt/pcp-jnk信号蛋白通路关键蛋白分子dvl表达增高激活下游rhoa同步增高,活化jnk,使jnk发生磷酸化,参与了神经管畸形的发生。上官少方(上官少方,王理,吴丽华等.pcp通路在神经管畸形发生中的作用和研究进展[j].中国优生与遗传杂志,2010(12):140-142.)等研究发现,pcp信号通路中基因在不同种属间的高度保守性,及其在汇聚延伸这一细胞极性变化过程中有重要作用,并且动物模型中已经证实,pcp途径中几种核心基因的突变引发神经管闭合缺陷,导致严重ntds的发生;因此,研究pcp通路相关基因在ntds中的变化和作用,对于揭示ntds的发病机制,了解神经管形成的调控通路,是十分有意义的。虽然现有技术已经发现wnt、pcp、notch、hh/shh、bmp等信号通路与人类ntds都有关系,但并没有对单一特定的蛋白修饰的功能实验研究,尤其是组蛋白及相关基因与ntds是否有关的报道,更没有获得明确的人类ntds的分子标志物,以用于人类ntds的临床诊断和检测。技术实现要素:因此,本发明要解决的技术问题在于提供一种新的检测神经管畸形的分子标志物及其应用。为此,本发明技术方案如下:一种用于检测神经管畸形的分子标志物,所述分子标志物包括特异性标志基因和/或蛋白质;其中,所述特异性标志基因为pax6、nestin和bmp4中的至少一种,所述蛋白质为h2ak119、mdm2和/或编码h2ak119和mdm2中至少一种蛋白质的基因。本发明还提供上述用于检测神经管畸形的分子标志物在制备诊断或检测神经管畸形的产品中的用途。进一步地,所述产品包括基于所述的用于检测神经管畸形的分子标志物设计的基因芯片、蛋白芯片或检测试剂。进一步地,所述基因芯片为包括用于扩增检测所述的用于检测神经管畸形的分子标志物的引物或与所述的用于检测神经管畸形的分子标志物杂交的探针。进一步地,所述蛋白芯片包括与所述的用于检测神经管畸形的分子标志物特异性结合的抗体。进一步地,所述检测试剂包括采用bradford法定量分析蛋白h2ak119、mdm2含量的试剂。进一步地,所述检测试剂还包括采用荧光定量pcr法分析特异性标志基因pax6、nestin和bmp4含量的试剂。本发明还提供一种检测神经管畸形的试剂盒,包括上述用于检测神经管畸形的分子标志物设计的基因芯片、蛋白芯片或检测试剂。进一步地,所述试剂盒,包括采用bradford法定量分析蛋白h2ak119、mdm2含量的试剂;优选的,还包括采用荧光定量pcr法分析特异性标志基因pax6、nestin和bmp4含量的试剂;优选的,还包括提取h2ak119、mdm2、的试剂。本发明技术方案,具有如下优点:1、本发明提供一种用于检测神经管畸形的分子标志物,该分子标志物包括特异性标志基因和/或蛋白质;其中,所述特异性标志基因为pax6、nestin和bmp4中的至少一种,所述蛋白质为h2ak119、mdm2和/或编码h2ak119和mdm2中至少一种蛋白质的基因。mdm2能够通过促进h2ak119的泛素-蛋白酶体途径的上调来调控其蛋白稳定性,h2ak119有抑制下游基因的功能,动物模型研究发现,甲氨蝶呤可以引起dna双链断裂发生atm介导的磷酸化,从而将mdm2招募到h2ak119位点,发生单泛素化,mdm2水平上调、促进h2ak119单泛素化从而抑制特异性标志基因的现象,并且人体样本研究也发现神经管畸形的人体内h2ak119和mdm2水平上调,特异性标志基因pax6、nestin和bmp4全部下调;因此,通过检测h2ak119、mdm2的表达,和特异性标志基因pax6、nestin和bmp4的变化情况,可以实现神经管畸形的早期诊断和干预。2、本发明还提供一种检测神经管畸形的试剂盒,包含上述分子标志物试剂盒制作工艺简单、操作安全、实用性强,易于推广使用。附图说明为了更清楚地说明本发明具体实施方式或现有技术中的技术方案,下面将对具体实施方式或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图是本发明的一些实施方式,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。图1为本发明实验例1中mtx为1um浓度时小鼠胚胎干细胞rnasequencing差异基因的表达情况;图2为本发明实验例1中mtx诱导后小鼠胚胎干细胞内特异性标志基因mrna水平的表达;图3为本发明实验例1中mtx诱导的小鼠胚胎干细胞的蛋白表达变化情况结果图;图4为本发明实验例1中小鼠胚胎干细胞h2ak119的chipqpcr的结果图;图5为本发明实验例1中小鼠胚胎干细胞mdm2的chipqpcr的结果图;图6为本发明实验例1中敲除mdm2后mtx诱导的小鼠胚胎干细胞的蛋白情况;图7为本发明实验例1中敲除mdm2后mtx诱导的小鼠胚胎干细胞的特异性标志基因的变化情况;图8为本发明实验例1中敲除mdm2后h2ak119的chipqpcr的结果图;图9为本发明实验例1中mtx诱导后小鼠胚胎干细胞的atm磷酸化的蛋白情况;图10为本发明实验例1中mtx诱导后小鼠胚胎干细胞的atm被抑制前后的特异性标志基因情况;图11为本发明实验例2中mtx诱导后小鼠神经组织内h2ak119及mdm2的变化情况;图12为本发明实验例2中的神经管畸形小鼠神经组织中特异性标志基因的变化情况;图13为本发明实施例1中人脑组织中特异性标志基因的变化情况;图14为本发明实施例1中人脑组织中蛋白h2ak119的变化情况;图15为本发明实施例1中人脑组织中mdm2的变化情况。具体实施方式提供下述实施例是为了更好地进一步理解本发明,并不局限于所述最佳实施方式,不对本发明的内容和保护范围构成限制,任何人在本发明的启示下或是将本发明与其他现有技术的特征进行组合而得出的任何与本发明相同或相近似的产品,均落在本发明的保护范围之内。实施例中未注明具体实验步骤或条件者,按照本领域内的文献所描述的常规实验步骤的操作或条件即可进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规试剂产品。人体正常与畸形组样本均取自山西省吕梁市家属知晓并同意由干冰运输至北京首都儿科研究所保存在-80冰箱。实验仪器:thermoforma37℃、二氧化碳培养箱(thermofisherscientific),tcssp8共聚焦显微镜(germanyleica),低温高速冷冻离心机(eppendorf),超低温冰箱(thermofisherscientific),水平电泳仪(bio-rad),垂直电泳槽(bio-rad),全湿转膜仪(bio-rad),quantstudiotm7flexpcr仪(termofisherscientific),nanodrop1000分光光度计(thermofisherscientific),nanostringprepstation(nanostringtechnology),nanostringncounter(nanostringtech.),veriti96wellthermalcycler(abi)。特异性标志基因(pax6,nestin,bmp4)采用nanostring方法定量检测或是采用mrna提取法,用rt-qpcr方法定量。rna提取法:采用trizol提取法提取细胞或组织总rna,按加拿大abm逆转录试剂盒操作将总rna逆转录成cdna,然后用qpcr法检测特异性标志基因的表达,其中使用的forwardprimer和reverseprimer使用常规的引物设计软件设计而得;qpcr的反应体系为:总反应体系为20μl/反应evagreen2×qpcrmastermix,10ul;forwardprimer(10μm),0.6ul;reverseprimer(10μm),0.6ul;cdna模板(100ng/μl),2ul;去离子水,6.8ul;反应条件如下:步骤温度时间循环enzymeactivation95℃10min1cycledenaturation95℃15s35cycleannealing/extension60℃60s35cyclemeltingcurve95℃15s1cycle60℃60s1cycle95℃15s1cycle60℃15s1cycle反应结束后,根据溶解曲线分析和琼脂糖凝胶电泳验证引物特异性。每对引物的反应均包括一个无模板阴性对照,并且每份模板的每个基因都设置3个重复孔,反应完成后,根据系统软件自动推算出每个反应扩增的thresholdcycle(ct值),计算目的基因和管家基因的ct平均值,以2-△△ct法进行比较,计算出目的基因的相对表达量,至少3次生物学重复,实验结果以mean±sd表示。rh2ax和h2ak119蛋白的表达水平,由于rh2ax是作为组蛋白dna双链断裂的靶蛋白,存在于细胞核里,而h2ak119蛋白作为组蛋白h2a的泛素位点修饰,提取出组蛋白即得到rh2ax和h2ak119蛋白的表达水平,为此,提供了组蛋白的提取方法为:采用epigentek试剂盒epiquiktmtotalhistoneextractionkit。具体步骤为:以1x107细胞量或100mg组织量为例。将试剂盒内10xpre-lysisbuffer以蒸馏水稀释成1×pre-lysisbuffer。重悬打碎或匀浆好的细胞置于冰上10分钟并在每三分钟轻轻晃动,然后3000rpm、4℃离心5分钟。如果细胞量和样本较少可放置于1.5-2mlep管中在10000rpm、4℃离心1分钟,弃上清,再用三倍体积的lysisbuffer重悬组织或细胞碎片,冰上孵育30分钟,12000rpm、4℃离心5分钟,将上清转入新的管子中。用dtt溶液和balancebuffer配置1:500溶液,以300ul体积立即加入到上一步转入管中的上清,测蛋白浓度。所提取的组蛋白可保存于-80℃。mdm2可以采用nanostring方法相对定量检测mdm2在mrna水平的表达,也可以采用ripa裂解液全蛋白提取法,然后用bradford法给蛋白定量检测mdm2。所述的ripa裂解液全蛋白提取法,具体步骤为:以1×107细胞量或100mg组织量为例,加入溶解ripa裂解液,混匀,取出适量细胞裂解液,在使用前2分钟加入pmsf,使pmsf的最终浓度为1mm;用移液枪将细胞培养基吸取扔掉,加入3ml预冷pbs清洗两次。以防止酚红以及培养基中的残留物质对免疫沉淀,蛋白纯化,酶活性分析等实验的影响。加入500ul裂解液,用枪吹打数下,使裂解液和细胞充分接触,冰上孵育5分钟。充分裂解后,4℃离心,14000g,5分钟,取上清置于新的管中,保存于-80℃备用。所述的用bradford法给蛋白定量,具体步骤为:将已知浓度的bsa标准品蛋白按照浓度梯度2mg/ml、1mg/ml、0.5mg/ml、0.25mg/ml、0.125mg/ml、0mg/ml稀释;每个ep管中加入245μl考马斯亮蓝溶液及5μl标准品或样品;使用酶标仪测每个标准品及样品在595nm波长处的吸光度;根据标准品的吸光度通过excel软件得出标准曲线,最后得到每个样品的蛋白浓度。蛋白质sds-page电泳:用蒸馏水清洗玻璃板,晾干后将其放入玻璃夹中,底部对齐卡紧,垂直卡在夹子上准备灌胶;配胶:依据目的蛋白分子量来确定分离胶浓度,按照试剂配方配置8%或者12%的分离胶。灌胶时胶一定要沿一侧玻璃板流下,避免产生气泡。加入7ml分离胶后,缓慢加入蒸馏水封。静置30min,待分离胶凝固后,将蒸馏水弃去,并用滤纸将残余水分吸干,将配制好的5%的浓缩胶灌入玻璃板,轻轻插入点样梳,梳齿底部不可有气泡。然后在梳子上方再多加一些浓缩胶,静置30min;上样:待浓缩胶凝固后,用水冲洗浓缩胶残渣,将其放入电泳槽中,加入足够电泳液后,两手分别捏住梳子的两边竖直向上轻轻将其拔出,根据蛋白浓度吸取样品,一般全蛋白上样20-30μg,不要吸进气泡,将加样器针头插至加样孔中缓慢加入样品和5μl蛋白上样指示剂电泳:电泳选择恒压,初始为80v,观察上样指示剂越过浓缩胶时,电压改为110v,当上样指示剂将要越过分离胶底部时,停止电泳。转膜:将nc膜浸泡于转膜缓冲液中平衡5min,将胶剥出,去除浓缩胶,根据胶的大小准备两张海绵,六张滤纸和一张nc膜,滤纸、膜和胶三者的大小相同,并将其置于转膜缓冲液中平衡10min,将含有目的蛋白的胶转移到nc膜上,确保与膜之间没有气泡。按照电泳槽中从负极到正极按照海绵+滤纸+凝胶+nc膜+滤纸+海绵“三明治”夹心法放置,再将三者移至转膜装置。安装好湿转仪,调整电流为恒流250ma,转膜时间根据目的蛋白大小调整。将整个装置放置在冰浴中。转膜完成后,可用丽春红染色,观察蛋白条带,评判转膜效率,然后用蒸馏水将丽春红洗净。免疫反应封闭:将膜移至含5%脱脂牛奶中,室温摇床上封闭1小时;孵育一抗:将一抗用5%脱脂牛奶稀释至适当浓度(按照抗体说明书),将膜转移至含有一抗的孵育盒中,4℃孵育过夜;3)洗膜:室温,1×tbst洗膜,水平摇床洗3次,每次10min;孵育二抗:根据一抗的种属来源选择相应的二抗用5%脱脂牛奶进行稀释(按抗体说明书),室温孵育45-60min;5)洗膜:室温,1×tbst洗膜,水平摇床洗3次,每次10min;6)ecl发光显色:将a液和b液按照1:1等体积混合,将膜浸泡在发光液中约2min,在全自动曝光仪上进行曝光,用凝胶成像系统对曝光后条带的灰度值进行分析。在本发明中,当样品为细胞和小鼠组织时采用的是westernblot方法检测mdm2蛋白表达量的变化,而当样品为人组织时通过nanostring技术检测mdm2在mrna的水平来相对定量mdm2在人神经组织中的表达量。实验例1胚胎干细胞为小鼠胚胎干细胞sv/129,来自于北京宣武医院,冻存于北京首都儿科研究所细胞库,甲氨蝶呤(mtx)购自pfizer(perth)ptylimited,贮存液浓度为500mg/20ml。所述小鼠胚胎干细胞于37℃、二氧化碳培养箱中在dmem高糖培养基中培养,培养基配方(小鼠胚胎干细胞(mesc)sv/129维持在dulbecco改良的eagle培养基(dmem,gibco,usa)中,培养基中含有0.1mmβ-巯基乙醇(invitrogen,carlsbad,usa),0.1mmnon-essentialaminoacid(neaa)(invitrogen,carlsbad)美国),0.1mm谷氨酸(invitrogen,carlsbad,usa),15%胎牛血清(gibco,usa)和1000u/ml小鼠白血病抑制因子(millipore,billerica,usa),0.2%明胶(invitrogen,carlsbad,usa))。将细胞置于加湿培养箱中,37℃、5%co2条件下培养,每2-3天传代一次。培养后,分为实验组和对照组,实验组培养基中加入mtx(甲氨蝶呤),使得培养基中mtx浓度为1μm;对照组添加等量生理盐水;培养12个小时,检测实验组和对照组胚胎干细胞的蛋白和特异性标志基因的变化情况。(1)mtx诱导的胚胎干细胞的rnasequencing使用trizol试剂(invitrogen)提取实验组和对照组胚胎干细胞总rna。通过bioanalyzer2200(aligent)检查rna质量并将rna于-80℃下保存。检测rna质量rnaintegritynumberrin>8.0的rna可以用于后期rnasequencing。同样rnaintegritynumberrin>8.0的rna适用于mirna纯化。使用mirneasyminikit(qiagen)纯化mirna,并通过凝胶电泳验证纯化结果。cdna文库构建根据制造商的说明书,使用vahtstmtotalrna-seq(h/m/r)为每个合并的rna样品构建cdna文库。通常,该方案由以下步骤组成:rrna耗尽并使用二价阳离子在94℃下片段化为150-200bp,持续8分钟。将切割的rna片段逆转录成第一链cdna,第二链cdna合成,末端修复片段,a尾并用索引衔接子连接。通过vahtstmdnacleanbeads获得目标条带。通过pcr纯化和富集产物以产生最终的cdna文库并通过agilent2200定量。将标记的cdna文库以相等比例合并,并在novelbiocorp.laboratory,shanghai的illuminahiseqxten的单个泳道中用于150bp配对末端测序。使用iontotalrna-seqkitv2.0(lifetechnologies)根据制造商的说明制备用于单端测序的互补dna(cdna)文库。通过page凝胶电泳对cdna文库进行大小选择以进行mirna测序。然后根据市售方案处理cdna文库用于质子测序过程。将样品稀释并混合,将混合物在onetouch2仪器(lifetechnologies)上处理并在onetouch2es工作站(lifetechnologies)上富集以根据ionpitm制备模板阳性离子pitm离子spheretm颗粒(lifetechnologies)模板ot2200kitv2.0(lifetechnologies)。富集后,加载混合的模板阳性离子pitm离子球粒子样品颗粒到1个p1v2质子芯片上根据来自novelbiocorp.laboratory,shanghai的ionpisequencing200kitv2.0(lifetechnologies)进行测序并在质子测序仪上测序。如图1所示为不同条件下胚胎干细胞的rnasequencing,图1(a)表示实验组和对照组两组之间的差异倍数,而图1(b)显示了实验组和对照组相比上调有差异的基因是1642个,而下调的是有672个,贴近于45度深色部分为差异不足的取对数后2倍的基因。(2)mtx诱导后小鼠胚胎干细胞后特异性标志基因的变化情况实验结果如图2所示,可以看出,mtx诱导后小鼠胚胎干细胞内特异性标志基因pax6、nestin和bmp4相对于正常胚胎干细胞,表达下调。(3)mtx诱导的小鼠胚胎干细胞的蛋白表达变化情况用western方法检测实验组和对照组胚胎干细胞蛋白含量的变化,检测h2ak119蛋白以h2a为内参,检测mdm2蛋白以gapdh为内参,如图3所示,可以看出,mtx诱导后,h2ak119和mdm2蛋白表达上调。如图4所示,为h2ak119chipqpcr,可以看出,特异性标志基因与h2ak119的结合明显加强。图4中的igg为negativecontrol。如图5所示,为mdm2chipqpcr,可以看出,特异性标志基因与mdm2的结合明显加强。(4)敲除mdm2,mtx诱导的胚胎干细胞的蛋白表达情况用westernblot检测敲除mdm2后h2ak119的变化,如图6所示,可以看出mtx诱导前后,以h2a作为内参,蛋白h2ak119变化不明显,说明蛋白h2ak119的上调依赖于mdm2。用qpcr方法检测特异性标志基因的变化情况,如图7所示,可以看出,敲除mdm2基因,mtx诱导前后,特异性标志基因的变化不明显。h2ak119chipqpcr,如图8所示,可以看出,mtx诱导的小鼠胚胎干细胞,敲除mdm2基因,特异性标志基因变化不明显。在上述附图6-8中,si-mdm2表示敲除mdm2基因;mtx+表示实验组,con-表示对照组。nc是negativecontrol,在反应中不会被敲除.因此可以比对mdm2的敲除效率。(5)mtx诱导的胚胎干细胞的atm磷酸化的检测如图9-10所示,在低叶酸的状态下dna发生双链断裂,mdm2被招募到dna受损位点并发生了atm激酶诱导的磷酸化,atm激酶的活性与被招募到受损位点的mdm2一起诱导了h2ak119泛素化发生。图9表示dna双链断裂是倚赖于atm激酶介导的磷酸化而并非atr激酶。图10表示在atm被抑制后,特异性标志基因没有下调,因此,特异性标志基因也是通过atm磷酸化的发生而被抑制的。综上,mtx诱导的小鼠胚胎干细胞,会导致h2ak119和mdm2的表达上调,特异性标志基因nestin、pax6和bmp4下调。mtx诱导前后,敲除mdm2基因,蛋白h2ak119表达水平变化不明显;特异性标志基因nestin、pax6和bmp4变化不明显,与蛋白h2ak119结合程度变化不明显。可以得出,mtx诱导小鼠胚胎干细胞,招募mdm2到h2ak119位点,发生单泛素化,从而抑制特异性标志基因表达。实验例21、实验动物和材料spf级成年c57bl/6j小鼠,雌雄各半,7~8周龄,体重17~19g。购自北京金牧羊实验动物养殖责任公司。甲氨蝶呤(mtx)购自pfizer(perth)ptylimited,贮存液浓度为500mg/20ml。2、小鼠模型制备购入的小鼠适应性喂养2天后,随机分组。分组后的雌鼠与同一批雄鼠按2:1的比例合笼过夜,第二天早晨发现阴道栓被定为孕0.5天。实验组在怀孕7.5天给予不同剂量的mtx腹腔注射;对照组给予等体积的生理盐水。怀孕13.5天时,对孕鼠进行摘眼球采血。采血后的小鼠断颈处死,腹部向上置于手术台上,75%酒精消毒。沿腹中线剖腹取出妊娠子宫,将子宫成串取下置于盛有pbs的培养皿中,体视显微镜下小心取出胚胎,并仔细观察每窝胚胎,观察体视显微镜下可见的畸形情况,并对胚胎进行拍照。取部分正常胚胎和畸形胚胎固定于体积浓度为10%中性甲醛中,用于做组织切片。取畸形胚胎,测小鼠体内基因和蛋白含量的变化。3、实验方法和实验结果(1)mtx诱导后小鼠体内特异性标志基因的变化情况实验结果如图12所示,可以看出,神经管畸形小鼠特异性标志基因pax6、nestin和bmp4相对于正常小鼠,全部下调。(2)mtx诱导的神经管畸形小鼠神经组织中蛋白含量的变化实验结果如图11所示,可以看出,小鼠脑组织中mdm2、h2ak119和rh2ax上调,rh2ax的上调说明小鼠神经组织中发生dna双链断裂,图中con表示正常小鼠,ntd表示mtx诱导产生的神经管畸形小鼠;brain表示脑组织,spine表示脊髓组织。综上,可知,神经管畸形小鼠的发生是由于mtx诱导的dna双链断裂发生了atm介导的磷酸化招募mdm2到dna受损位点发生了h2ak119单泛素化从而抑制分化基因的表达而致畸;即神经管畸形小鼠的mdm2、h2ak119水平上调,特异性标志基因pax6、nestin和bmp4下调。实施例1本实施例提供了一种检测神经管畸形的方法,包括以下步骤:1)提取待测样本的h2ak119、mdm2;2)定量分析h2ak119、mdm2,特异性标志基因pax6、nestin、bmp4的含量;3)与对照样本比对,若h2ak119、mdm2水平上调,且特异性标志基因pax6、nestin、bmp4全部下调,则神经管畸形发生。其中待测样本为神经管畸形患者的脑组织,对照样本为正常人体的脑组织。上述组织取自山西省吕梁市家属知晓并同意由干冰运输至北京首都儿科研究所保存在-80冰箱。h2ak119的提取方法为:采用epigentek试剂盒epiquiktmtotalhistoneextractionkit。采用nanostring方法定量检测mdm2水平变化情况。特异性标志基因(cdx2,gata4,bmp4)是采用mrna提取法,采用nanostring方法定量检测。结果如图13所示,为人脑组织中特异性标志基因的变化情况,可以看出,ntds患者体内特异性标志基因明显低于正常人体。如图14所示,为人脑组织中蛋白h2ak119的变化情况,可以看出,ntds患者体内h2ak119明显高于正常人体,图中的ntds1-ntds8表示神经管畸形患者的脑组织样本1-8,图中的normal1-normal8表示正常人体的脑组织样本1-8。如图15所示,为人脑组织中mdm2变化情况,可以看出,ntds患者体内mdm2明显高于正常人体。实施例2本实施例提供了一种检测神经管畸形的试剂盒以特异性标志基因pax6、nestin和bmp4,蛋白质h2ak119和mdm2为分子标志物,包括提取h2ak119、mdm2的试剂,h2ak119、mdm2的定量分析试剂,和荧光定量pcr反应试剂。实施例3本实施例提供了一种检测神经管畸形的试剂盒的使用方法(1)提取待测样品的h2ak119、mdm2;(2)检测h2ak119、mdm2中及特异性标志基因pax6、nestin和bmp4的含量;(3)与对照样本比对,若h2ak119、mdm2水平上调,且特异性标志基因pax6、nestin和bmp4全部下调,则神经管畸形发生。显然,上述实施例仅仅是为清楚地说明所作的举例,而并非对实施方式的限定。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式的变化或变动。这里无需也无法对所有的实施方式予以穷举。而由此所引伸出的显而易见的变化或变动仍处于本发明创造的保护范围之中。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1