一种二酰甘油酰基转移酶1及其在生产甘油三酯中的应用的制作方法
本发明涉及一种二酰甘油酰基转移酶1及其在生产甘油三酯中的应用,属于酶工程和微生物工程技术领域。
背景技术:
多不饱和脂肪酸(pufas)是指含有两个或两个以上双键且碳链长度为18~22个碳原子的直链脂肪酸。由于其具有降低心脑血管疾病的功能且在维持身体健康和预防疾病等方面发挥着十分重要的作用,因此,多不饱和脂肪酸(pufas)作为一种保健品,在国内、国际均拥有巨大的市场。
目前,市场上售卖的多不饱和脂肪酸(pufas)多来源于水生浮游植物,食此类植物为生的野鳕鱼、鲱鱼、鲑鱼等深海鱼类的内脏中也富含多不饱和脂肪酸(pufas)。但是,由于动植物的生长周期过于长且动植物的培育成本相对高,仅靠从这些动、植物中提取获得多不饱和脂肪酸(pufas)已无法满足日益增长的市场需求,因此,急需找到可以提升多不饱和脂肪酸(pufas)产量的方法。
研究表明,有些细菌、真菌等微生物也具有生产脂质的功能,并且,这些细菌、真菌等微生物均具有生长周期短、繁殖快、培养成本低、不受地理环境及气候条件影响、环境友好等诸多优势,另外,这些细菌、真菌等微生物还具有油脂产量高、产得的油脂种类丰富等特点,因此,通过微生物产油脂提升多不饱和脂肪酸(pufas)的产量是一种极具潜力的方法。
微生物来源的脂质包括甘油酯、类脂(磷脂、固醇等)和游离脂肪酸,其中,甘油三酯(tag)是微生物脂质最主要的储存形式。而微生物中的甘油三酯(tag)主要通过内质网上的肯尼迪途径合成,酰基辅酶a:二酰甘油酰基转移酶(dgat,ec2.3.1.20)则是肯尼迪途径的关键限速酶,可催化在二酰甘油sn-3位置上加一个脂肪酸酰基的过程。
现在,已经被报道的dgat分为四种类型:dgat1、dgat2、dgat3和dgat/ws,其中,dgat1和dgat2在真核生物中普遍存在,二者虽然在dna层面同源性低,是两种不同的酶,但均对生物体中tag的合成有重要作用,例如,bouvier-navép等在酵母中表达拟南芥dgat基因,使得重组酵母菌中dgat活性增加200~600倍,tag积累量增加3~9倍;并且,二者虽然具有广泛的脂肪酸底物偏好性,但在某些情况下也会优先选择特定的脂肪酸底物,例如,taylordc等发现甘蓝型油菜的dgat1偏好棕榈酸,shockey,j.m等发现油桐的dgat2优先选择α-桐酸。
鉴于dgat在脂肪酸合成过程中的重要调控作用,进一步研究其功能对定向强化产油微生物脂质积累,进而提升产油微生物多不饱和脂肪酸(pufas)的产量具有重要指导意义。
但是,并非所有dgat都能对产油微生物的脂质积累起到促进作用,例如,文章《卷枝毛霉二酰甘油酰基转移酶的基因鉴定与功能分析》中将卷枝毛霉的dgat1基因通过载体pyes2在酿酒酵母中异源表达,结果表明两种dgat1酶均没有表现出合成tag的能力(参考文献:张鲁宁.卷枝毛霉二酰甘油酰基转移酶的基因鉴定与功能分析.江南大学,2015);文章《蒺藜苜蓿dgat1基因的克隆和功能鉴定》中将蒺藜苜蓿的dgat1基因在酿酒酵母中异源表达,发现其mtdgat1-1没有表现出合成tag的能力(参考文献:徐荣华,崔涛,王建才.蒺藜苜蓿dgat1基因的克隆和功能鉴定.西北植物学报,2016,36(10):1941-1947);文章《产油酵母圆红冬孢酵母二酰甘油酰基转移酶的鉴定和性质研究》中将圆红冬孢酵母中的dgat1通过载体pyes2在酿酒酵母中异源表达,结果表明其dgat1酶没有表现出合成tag的能力(参考文献:王珍.产油酵母圆红冬孢酵母二酰甘油酰基转移酶的鉴定和性质研究.江南大学,2016)。因此,选择合适的dgat格外重要。
高山被孢霉(mortierellaalpina,m.alpina)是一种具有强脂质合成能力的产油丝状真菌,其总脂含量可达菌体干重的50%以上,其中,ω-6多不饱和脂肪酸:花生四烯酸(ara)的含量占总脂的30%~40%,因此,高山被孢霉(mortierellaalpina,m.alpina)可以说是一种非常优质的ara生产菌,高山被孢霉(mortierellaalpina,m.alpina)来源的dgat也相对更具对产油微生物的脂质积累起到促进作用的潜力。
然而,目前,关于m.alpina中dgat的研究报道很少,且这些报道集中于对dgat2的研究,例如,jeennors、luoxa等发现m.alpina中dgat2能提高tag含量并具有特定的脂肪酸偏好性,至今为止,还未有关于m.alpina中dgat1的相关研究报道。
若能够成功获取m.alpina中的dgat1,并对其进行生物信息学分析并在微生物中表达以研究该酶的功能,一定能够为进一步提高微生物中tag和特定的pufas的产量提供更充实的理论依据。
技术实现要素:
[技术问题]
本发明要解决的技术问题是提供一种高山被孢霉(mortierellaalpina,m.alpina)来源的二酰甘油酰基转移酶1(dgat1)以提高微生物产甘油三酯(tag)的能力。
[技术方案]
为解决上述问题,本发明提供了一种二酰甘油酰基转移酶1(diacylglycerolacyltransferase,dgat;ec2.3.1.20),所述二酰甘油酰基转移酶1的氨基酸序列如seqidno.1所示。
本发明还提供了编码上述二酰甘油酰基转移酶1的基因。
在本发明的一种实施方式中,所述基因的核苷酸序列如seqidno.2所示。
本发明还提供了携带上述基因的重组质粒。
在本发明的一种实施方式中,所述重组质粒的载体为pyes2载体。
本发明还提供了携带上述基因或上述重组质粒的宿主细胞。
在本发明的一种实施方式中,所述宿主细胞为酿酒酵母或大肠杆菌。
在本发明的一种实施方式中,所述宿主细胞为酿酒酵母。
本发明还提供了上述二酰甘油酰基转移酶1的制备方法,所述方法为使用上述宿主细胞,将上述宿主细胞加入到不含尿嘧啶的培养基中培养36~48h后收集菌体,将收集的菌体加入到不含葡萄糖而含有半乳糖的培养基中至od600=0.3~0.5后继续诱导培养48~96h,得到二酰甘油酰基转移酶1。
在本发明的一种实施方式中,所述培养基包含为sc-u培养基。
在本发明的一种实施方式中,所述培养的温度为28~30℃、转速为200~250rpm。
本发明还提供了应用上述方法制备得到的二酰甘油酰基转移酶1。
本发明还提供了上述二酰甘油酰基转移酶1或上述基因或上述重组质粒或上述宿主细胞或上述制备方法或上述制备得到的二酰甘油酰基转移酶1在生产甘油三酯方面的应用。
本发明还提供了一种生产甘油三酯的方法,所述方法为使用上述宿主细胞,将上述宿主细胞加入到不含尿嘧啶的培养基中培养36~48h后收集菌体,将收集的菌体加入到不含葡萄糖而含有半乳糖的培养基中至od600=0.3~0.5后继续诱导培养48~96h,得到甘油三酯。
在本发明的一种实施方式中,所述培养基包含为sc-u培养基。
在本发明的一种实施方式中,所述培养的温度为28~30℃、转速为200~250rpm。
[有益效果]
本发明的二酰甘油酰基转移酶1具有催化甘油三酯(tag)合成的功能,将含有本发明二酰甘油酰基转移酶1的重组酿酒酵母诱导培养48h,可使含有本发明二酰甘油酰基转移酶1的重组酿酒酵母中的总脂肪酸含量以及甘油三酯含量分别较不含本发明二酰甘油酰基转移酶1的酿酒酵母提升1.94倍以及12.09倍,甘油三酯中的单不饱和脂肪酸16:1、18:1的含量较不含本发明二酰甘油酰基转移酶1的酿酒酵母分别提升10.36、9.14倍,该结果为通过基因工程手段进一步提升高山被孢霉等产油微生物产甘油三酯(tag)的能力,进而提高微生物产多不饱和脂肪酸(pufas)的能力提供了充实的理论支持。
附图说明
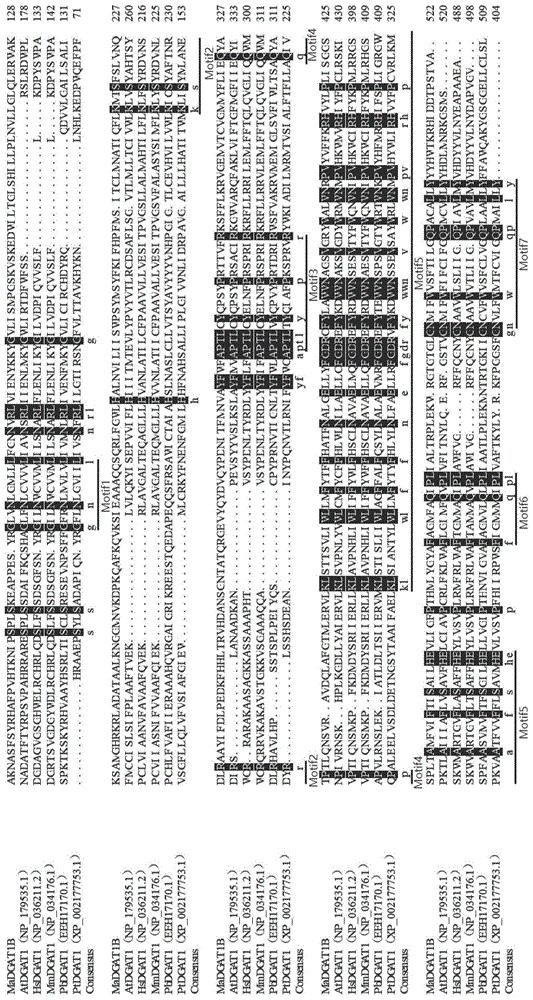
图1:madgat1b与已知二酰甘油酰基转移酶1的氨基酸序列比对结果图。
图2:madgat1b的跨膜结构预测图。
图3:重组大肠杆菌e.coli-pyes2-madgat1b的阳性转化子验证图;其中,m表示marker,1~20分别表示转化子编号。
图4:重组酿酒酵母invsc1-pyes2-madgat1b的阳性转化子验证图;其中,m表示marker,1~9分别表示转化子编号。
图5:重组酿酒酵母invsc1-pyes2-madgat1b的目的蛋白madgat1b的表达情况验证图;其中,m表示marker,“-”表示阴性对照(含空载体的invsc1),“+”表示阳性对照(其他带his标签蛋白的invsc1),1表示invsc1-pyes2-madgat1b。
图6:重组酿酒酵母invsc1-pyes2-madgat1b全细胞蛋白和全细胞蛋白沉淀物中的目的蛋白madgat1b的表达情况的westernblot结果;其中,m表示marker,“-”表示阴性对照(含空载体的invsc1),“+”表示阳性对照(其他带his标签蛋白的invsc1),1a表示invsc1-pyes2-madgat1b全细胞蛋白,1b表示invsc1-pyes2-madgat1b全细胞蛋白沉淀物。
图7:重组酿酒酵母invsc1-pyes2-madgat1b全细胞蛋白和全细胞蛋白沉淀物中的目的蛋白madgat1b的表达情况的灰度分析估算结果;其中,1a表示invsc1-pyes2-madgat1b全细胞蛋白,1b表示invsc1-pyes2-madgat1b全细胞蛋白沉淀物。
图8:重组酿酒酵母invsc1-pyes2-madgat1b的总脂质薄层色谱图;其中,c表示invsc1-pyes2,1①、1②表示invsc1-pyes2-madgat1b的两个平行组,sta表示三软脂酸甘油三酯标准品。
图9:重组酿酒酵母invsc1-pyes2-madgat1b的总脂(tfa)中tag的含量;其中,c表示invsc1-pyes2,1表示invsc1-pyes2-madgat1b。
图10:重组酿酒酵母invsc1-pyes2-madgat1b的tag中脂肪酸的组成和含量;其中,c表示invsc1-pyes2,1表示invsc1-pyes2-madgat1b,other表示c16:0、c16:1、c18:0、c18:1外的脂肪酸含量之和。
图11:重组酿酒酵母invsc1-pyes2-madgat1b的tag中主要脂肪酸的含量变化;其中,纵坐标表示invsc1-pyes2-madgat1b与invsc1-pyes2的比值。
图12:重组酿酒酵母invsc1-pyes2-madgat1b的总脂肪酸含量;其中,c表示invsc1-pyes2,1表示invsc1-pyes2-madgat1b。
图13:重组酿酒酵母invsc1-pyes2-madgat1b的生物量(菌体干重);其中,c表示invsc1-pyes2,1表示invsc1-pyes2-madgat1b,dcw表示菌体干重。
具体实施方式
下面结合具体实施例,对本发明进行进一步的阐述。
下述实施例中涉及的高山被孢霉(mortierellaalpina)atcc32222购自美国标准生物品收藏中心(atcc);大肠杆菌(e.coli)dh5α、酿酒酵母(saccharomycescerevisiae,s.cerevisiae)营养缺陷型菌株invsc1、表达载体pyes2/ntc购自invitrogen公司;kodplus高保真dna聚合酶购自日本toyobo公司;taqdna聚合酶购自cwbio公司;反转录试剂盒(primescriptrtregentkitwithgdnaeraserrr047a&r6110a)购自日本takara公司;质粒提取试剂盒购自北京天根生化科技有限公司;真菌基因组dna提取试剂盒购自bioflux公司;限制性内切酶(ecorⅰ、xbaⅰ)、t4连接酶、trizol、pcr产物纯化试剂盒、凝胶回收试剂盒、generulerdnaladdermix、pagerulerprestainedproteinladder、抗体(6×his-tagmonoclonalantibody、peroxidaseaffinipuremouseanti-goatlgg[h+l])购自thermo公司;正十五烷酸(c15:0)、三软脂酸甘油酯、20%(w/w)盐酸甲醇、酸洗玻璃珠、鲑鱼精购自sigma公司;丙烯酰胺/甲叉双丙烯酰胺40%溶液(29:1)、temed、depc水、氨苄(ampicillin)抗生素、无氨基酵母氮源(ynb)和各类氨基酸购自上海生物工程有限公司;酵母提取物、胰蛋白胨购自oxoid公司;薄层层析硅胶板购自乳山太阳干燥剂有限公司;牛血清蛋白购自麦克林公司;低吸附型无酶枪头、无酶离心管、无酶pcr管、2ml棕色气相瓶及瓶盖购自苏州科晴生物公司;bca蛋白测定试剂盒购于上海碧云天公司;玻璃点样毛细管购自上海申迪玻璃仪器有限公司;脱脂乳粉购自美国bd公司;pvdf膜、plusecl显影液购自millipore公司;其他试剂购自国药集团。
下述实施例中涉及的酿酒酵母转化相关实验用到的试剂及配方均参考invitrogen公司的pyes2质粒表达手册,其中,将peg-3350替换为peg-4000。
下述实施例中涉及的引物由上海桑尼公司合成,测序工作由上海华大基因公司完成。
(高山被孢霉(mortierellaalpina)atcc32222、大肠杆菌(e.coli)dh5α、酿酒酵母(saccharomycescerevisiae,s.cerevisiae)营养缺陷型菌株invsc1均可以购买得到,不需要进行用于专利程序的保藏)
下述实施例中涉及的培养基如下:
lb液体培养基:10g/l胰蛋白胨、5g/l酵母提取物、10g/lnacl。
lb固体培养基:10g/l胰蛋白胨、5g/l酵母提取物、10g/lnacl、20g/l琼脂。
soc复苏培养基:20g/l胰蛋白胨、5g/l酵母提取物、0.5g/lnacl、0.186g/lkcl、0.95g/lmgcl2、3.6g/l葡萄糖。
ypd液体培养基:10g/l酵母提取物、20g/l胰蛋白胨、20g/l葡萄糖。
ypd固体培养基:10g/l酵母提取物、20g/l胰蛋白胨、20g/l葡萄糖、20g/l琼脂。
sc-u液体培养基:20g/l葡萄糖、6.7g/lynb、各0.1g/l的氨基酸(腺嘌呤、精氨酸、半胱氨酸、亮氨酸、赖氨酸、苏氨酸、色氨酸),各0.05g/l的氨基酸(天冬氨酸、组氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、酪氨酸、缬氨酸)。
sc-u固体培养基:20g/l葡萄糖、6.7g/lynb、各0.1g/l的氨基酸(腺嘌呤、精氨酸、半胱氨酸、亮氨酸、赖氨酸、苏氨酸、色氨酸),各0.05g/l的氨基酸(天冬氨酸、组氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、酪氨酸、缬氨酸)、20g/l琼脂。
sc-u诱导培养基:10g/l棉籽糖、6.7g/lynb、各0.1g/l的氨基酸(腺嘌呤、精氨酸、半胱氨酸、亮氨酸、赖氨酸、苏氨酸、色氨酸),各0.05g/l的氨基酸(天冬氨酸、组氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、酪氨酸、缬氨酸)、20g/l半乳糖。
实施例1:编码本发明二酰甘油酰基转移酶1的基因的筛选与生物信息学分析
具体步骤如下:
在ncbi中选择不同物种中已鉴定功能的dgat1基因序列为模板(表1),在已完成测序的m.alpinaatcc3222菌株的基因库中进行blast比对,获得备选的目的基因;然后将备选基因在ncbi库中进行二次比对筛选,将最终得到的目的基因命名为madgat1b,对应蛋白命名为madgat1b。
为了判断筛选出的madgat1b是否属于dgat1类酶,将其与表1的5种来自动物、高等植物、微生物、微藻中典型dgat1的氨基酸序列进行比对,通过dnaman软件进行氨基酸同源性和保守结构分析,结果见图1(黑色标注部分表示相似度100%的氨基酸);通过在线网站tmhmm预测madgat1b的跨膜结构,结果见图2;并且,通过在线网站expasy-protparamtool预测madgat1b的蛋白大小和等电点。
研究结果表明,通常情况下,dgat1中有7个保守区域,分别标记为motif1-7,对应的保守区域如下:motif1(gl区段)、motif2(ks区段)、motif3(ptr区段)、motif4(qp区段)、motif5(lwlffefdrfywwnwwnppfshp区段)、motif6(fql区段)和motif7(ngqpy区段),并且,一般而言,dgat1包含500多个氨基酸残基,分子量为50~60kd,蛋白三维结构中包含6~9个假定的跨膜区域。
由图1可知,madgat1b中含有dgat1酶通常应有的7个motif保守结构(划线部分),并且,madgat1b与表1中5种不同来源的dgat1蛋白比较发现相似度为37.48%,madgat1b对应cdna的全长为1569bp,编码522个氨基酸;由图2可知,madgat1b含有7个跨膜结构;并且,由expasy-protparamtool预测结果可知,madgat1b蛋白大小约为59.49kd,等电点为9.43,可见,madgat1b具有dgat1蛋白结构的基本特征。
综合以上结果,madgat1b与不同物种的dgat1氨基酸序列相似性具有生物学意义,基因长度、氨基酸个数、保守区域和跨膜结构均符合dgat1酶的特征,由此认为筛选得到的madgat1b具有dgat1蛋白的功能。
表1比对模板dgat1基因的相关信息
实施例2:madgat1b的克隆
具体步骤如下:
使用trizol法提取高山被孢霉(mortierellaalpina)atcc32222的总rna,按照takara反转录试剂盒说明书进行反转录获得cdna,在高山被孢霉(mortierellaalpina)atcc32222的cdna文库中通过pcr反应扩增madgat1b,扩增madgat1b所用引物见表2。
所用pcr仪为bio-radt100thermalcycler,使用kodplus高保真dna聚合酶,反应体系为50μl,体系内容按照该dna聚合酶说明书内容进行;反应过程如下:94℃预变性3min,然后94℃变性30s,60℃退火30s,68℃延伸100s,重复上述三步骤30次,然后68℃充分延伸7min,最后降到12℃保持5min后停止。
反应结束后,通过1%琼脂糖凝胶电泳验证扩增产物条带大小正确后,对该pcr产物进行纯化,获得madgat1b。
表2引物序列及其用途
注:酶切位点用下划线标出。
实施例3:madgat1b在酿酒酵母中的表达
具体步骤如下:
(1)酵母表达载体的构建
使用限制性内切酶ecorⅰ、xbaⅰ对madgat1b进行酶切后,利用t4连接酶将酶切产物连接到pyes2/ntc表达载体中β-半乳糖苷酶(gal1)启动子下游的对应多克隆位点中,通过化学转化法(42℃、90s转化条件)将连接产物(不超过5%)导入大肠杆菌(e.coli)dh5α中,通过添加有0.1g/lampicillin的lb固体培养基平板初步筛选得到大肠杆菌阳性转化子,然后提取质粒作为模板,通过使用表3的通用引物t7/t7terminator进行pcr扩增,将产物用1%琼脂糖凝胶电泳验证,初步筛选出条带大小正确的大肠杆菌阳性转化子并进行测序分析,测序成功即获得重组质粒,命名为pyes2-madgat1b(e.colidh5α阳性转化子筛选验证的结果见图3所示,用通用引物扩增后的madgat1b由于从pyes2载体上开始复制,因而长度应为1917bp,图3中显示e.coli-pyes2-madgat1b的转化子5、12在约2000bp处有目的条带,将转化子e.coli-pyes2-madgat1b-12进一步测序验证,测序结果与理论一致)。
(2)酿酒酵母的转化筛选
根据invitrogen公司的pyes2质粒手册内容,通过peg/liac方法将(1)中获得的测序正确的重组质粒pyes2-madgat1b和空载体pyes2/ntc(用作阴性对照)分别转化到酿酒酵母菌株invsc1中,通过不含尿嘧啶(ura-)的sc-u固体培养基平板初步筛选得到酿酒酵母阳性转化子,然后提取酵母基因组作为模板,使用表3的t7/t7terminator通用引物进行pcr扩增,将产物用1%琼脂糖凝胶电泳验证,初步筛选出条带大小正确的阳性转化子并进行测序分析,得到重组酿酒酵母invsc1-pyes2-madgat1b以及重组酿酒酵母invsc1-pyes2(酿酒酵母阳性转化子筛选验证结果见图4,由图4可知,invsc1-pyes2-madgat1b的1~9号转化子pcr验证结果与理论值一致,在约2000bp附近有目的条带,将invsc1-pyes2-madgat1b的1~9号转化子进一步测序验证,测序结果与理论一致)。
(3)诱导培养
将重组酿酒酵母invsc1-pyes2-madgat1b以及重组酿酒酵母invsc1-pyes2在sc-u液体培养基中培养48小时,培养条件为28℃、200r/min,然后6000rpm离心5min后弃去培养基,接入sc-u诱导培养基(起始od600=0.4)中使madgat1b被半乳糖诱导表达,在同样条件下培养48小时,得到发酵液,将发酵液在12000rpm、1min条件下离心收集菌体,取部分湿菌体用于蛋白表达分析,将其余菌体冷冻干燥后测定生物量以及脂质组分与含量。
(4)目的蛋白表达水平测定
收集诱导48h后的发酵液2ml,在12000rpm、4℃条件下离心1min收集菌体,基于invitrogen公司的pyes2质粒手册内容提取酿酒酵母的全细胞蛋白质,在菌体中加入裂解液使od600值达到50~100,然后加入与裂解液等体积的0.5mm酸洗玻璃珠,置于漩涡振荡器振荡30s后置于冰上30s,重复4次后,通过bca试剂盒测定全细胞蛋白质的浓度,通过sds-page(10%分离胶)分离样品,电转移(200ma、1h)到pvdf膜上,使用稀释度1:2000的his抗体作为第一抗体,以稀释度1:5000鼠二抗与之结合,然后与化学发光溶液反应并在成像仪上显影拍照。
利用westernblot对酵母重组菌株中目的蛋白madgat1b的表达情况进行分析,选用pyes2/ntc载体n端6×his标签作为抗原,使用在线软件expasy-protparamtool预测带有6×his标签的madgat1b蛋白质的分子量约为64kd,westernblot结果见图5所示。
由图5可知,invsc1-pyes2-madgat1b的全蛋白提取物中有与his抗体结合并且大小在55~72kd之间的蛋白,与目的蛋白的预期大小64kd基本一致,因而该条带为带有6×his标签的madgat1b,表明外源蛋白madgat1b在酿酒酵母中能正常表达。
将invsc1-pyes2-madgat1b的全细胞蛋白提取液在20000g条件下离心15min后取上清液和沉淀测定蛋白浓度,最终分别取20μg的全细胞蛋白样品和沉淀样品以进行westernblot实验(上清液蛋白浓度较低不进行实验),通过imagej软件进行蛋白表达量估算,结果如图6~图7所示。
由图6~图7可知,在沉淀物中检测到较强的蛋白信号,而等质量的菌体全细胞蛋白样品中madgat1b表达量明显降低,相差11倍左右,推测该现象可能是由于madgat1b为内质网上膜蛋白,在离心时与细胞器碎片一同沉淀。
(5)甘油三酯中脂肪酸组成及含量测定
薄层色谱法(tlc)是一种基于混合物组分在固定相和流动相之间的不均匀分配或保留而将其分离的方法,组分经展开、显色,形成色谱谱带,可以通过适当技术对谱带进行处理得到定性和定量的检测结果,因此,用薄层色谱从(3)获得的菌体的总脂质中分离tag,利用gc-ms技术检测其tag部分的脂肪酸组成及含量,具体步骤如下:
称取20mg菌体于提脂玻璃瓶中,用氯仿:甲醇:水=2:2:1(v/v/v)的混合有机试剂提取总脂质,以正己烷:乙醚:乙酸=80:20:1(v/v/v)的混合溶剂作为展开剂在薄层色谱板上分离各个脂质,碘蒸气熏染使各个斑点可视化,并使用imagej软件进行灰度分析估算tag在总脂中的比例,随后,立即刮下tag部分的斑点置于提脂玻璃瓶中并加入十五烷酸标品作为内标,使用盐酸:甲醇=1:9(w/w)溶剂在60℃水浴条件下甲酯化3小时,用正己烷提取脂肪酸甲酯,通过岛津公司单四级杆气相色谱质谱联用仪gcms-qp2010ultra进行分析tag中的脂肪酸成分和含量;
操作条件如下:使用restek的rtx-wax气相色谱柱(30m×0.25mm×0.25μm),氦气用作载气,进样口温度为240℃,进样量为1μl,分流比为10;柱温箱温度从150℃开始,保持2min,然后以10℃/min的速率升温至190℃,保持5min,后以5℃/min速率升温至220℃,保持16min。质谱扫描范围为50~550m/z;离子源和传输线接口温度分别为220℃和250℃。
利用tlc对重组菌中总脂进行分离的展开图及分析结果如图8~图9所示。
由图8~图9可知,对照组invsc1-pyes2中tag含量占总脂的42%;与对照组相比,invsc1-pyes2-madgat1b中tag/tfa比例显著变化(p<0.05),比对照组高19.63%。可见,madgat1b表现出较强的提高tag在总脂质中的含量的能力。
刮取tlc板上tag部分的样品并通过gc-ms对脂肪酸进行定性、定量分析后,tag中脂肪酸的组成和含量结果见图10。
由图10可知,酿酒酵母中tag的主要脂肪酸成分为十六烷酸(c16:0)、十六碳烯酸(c16:1)、十八烷酸(c18:0)、十八碳烯酸(c18:1),且与对照组相比,转化子中各脂肪酸成分间比例无明显变化。
将invsc1-pyes2-madgat1b转化子的tag中的4种主要脂肪酸与对照组相比,探究含量的变化情况,结果见图11。
由图11可知,与对照组相比,invsc1-pyes2-madgat1b的tag中4种主要脂肪酸含量均有较大提升,c16:0、c16:1、c18:0、c18:1含量的提升倍数分别为15.24、10.36、15.21、9.14倍,导致tag总提升量为对照组的12.09倍。可见,madgat1b在增加tag含量方面有明显效果,在提升产油微生物的脂质含量方面有巨大潜力。
(6)菌体总脂肪酸的组成及含量测定
分别称取20mg冻干的invsc1-pyes2、invsc1-pyes2-madgat1b菌体于提脂玻璃瓶中,加入2ml浓度为4mol/l的盐酸,于80℃条件下水浴1h后,充分振荡,然后在-80℃冰箱中静置15min,重复上述反复冻融操作3次,然后加入100μl2.096mg/ml的十五烷酸标准品作为内标,1ml氯仿、1ml甲醇提取脂质,旋转振荡1h后2000rpm离心吸取下层氯仿层到新瓶,补加1ml氯仿并充分振荡后,同上操作,混合两次的氯仿层并氮吹吹干,后续甲酯化和测定方法同(5)中所述。
重组酵母菌株中总脂肪酸含量如图12所示,菌体的生物量(每升培养基中的菌体干重)如图13所示,总脂肪酸的组成和含量如表4所示。
由图12~图13可知,与对照组invsc1-pyes2相比,在酿酒酵母中表达外源madgat1b对菌体生长无显著影响,对照组酿酒酵母中总脂含量为菌体干重的7.79%,invsc1-pyes2-madgat1b中总脂含量为菌体干重的15.15%,与对照组相比显着增加1.94倍(p<0.05)。
由表4可知,invsc1-pyes2-madgat1b中各脂肪酸的含量均有大幅度提升,其中c14:0提升最大,为5.79倍,但含量较低,对总脂含量影响较小;c16:0、c16:1、c18:0、c18:1的含量分别提升2.40、1.61、2.91、1.63倍,分别占总脂的25.33%、31.55%、13.50%、23.58%,是导致总脂含量提高的主要脂肪酸成分。
可见,madgat1b在提升总脂含量方面有明显的优势,说明高山被孢霉来源的dgat1蛋白可以更好地促进脂质积累,具有很好的应用价值。
表3引物序列及其用途
表4酿酒酵母总脂肪酸的组成和含量
注:c为invsc1-pyes2,1为invsc1-pyes2-madgat1b。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
序列表
<110>江南大学
<120>一种二酰甘油酰基转移酶1及其在生产甘油三酯中的应用
<160>6
<170>patentinversion3.3
<210>1
<211>522
<212>prt
<213>人工序列
<400>1
metthrlysasngluprovalleualaalaaspvalalaglualathr
151015
leuprogluileproprothrvalgluaspleulyslysalalysasn
202530
alaserphesertyrarghisalapheprovalhisthrlysasnile
354045
proserproleuserlysglualaproproglusertyrargglyleu
505560
valasnleuglymetleuleuleupheglyasnasnvalargleuval
65707580
ilegluasntyrlyslystyrglyvalleuilesermetproglyser
859095
lysvalserlysgluasptrpileleuthrglyleuserhisileleu
100105110
leuproleuasnvalleuleuglyleuglnleugluargtrpalalys
115120125
lysseralametglyhisarglysargleualaaspalathralaala
130135140
leulysasnglyglymetasnvallysaspprolysglnalaphelys
145150155160
glnvallysserileglualaalaalaglnglnserglnargleuphe
165170175
glytrpleuhisalaleuasnvalileleuileilesertrpproser
180185190
tyrmetsertyrphelysilephehisprophemetserilethrcys
195200205
leumetasnalathrileglnpheleulysmetthrserpheserleu
210215220
valasnglnaspleuargalaalatyrilepheaspleuprogluasp
225230235240
lysphehishisleuthrargvalhisaspalaasnserglyasnthr
245250255
alathrglnargglygluvaltyrglntyraspvalglntyrproasp
260265270
asnilethrphemetasnvalalatyrphetrpphealaprothrleu
275280285
cystyrglnprosertyrproargthrthrvalphearglysserphe
290295300
pheleulysargvalglyglumetvalthrcysvalglymetmettyr
305310315320
pheleuilegluglntyralathrprothrleuglnasnservalarg
325330335
alavalaspglnleualapheglythrmetleugluargvalleulys
340345350
leuserthrthrservalleuiletrpleuleumetphetyrthrphe
355360365
phehisalathrpheasnalaleuglygluleuleutyrpheglyasp
370375380
argcysphetyrleualatrptrpasnalaglyservalglyargtyr
385390395400
trpalaleutrpasnargprovaltyrvalphephelysarghisval
405410415
tyrleuproleuileserglyglyserserproleuthralametphe
420425430
valilephethrileseralaileleuhisgluvalleuileglyphe
435440445
prothrhismetleutyrglytyralaphealaglymetphealagln
450455460
ileproleuilealaleuthrargproleuglulystrpargglythr
465470475480
glythrglyleuglyasnmetilephetrpvalserphethrileleu
485490495
glyglnproalacysalaleuleutyrtyrtyrhistrpthrlysarg
500505510
hisileaspaspthrproserthrvalala
515520
<210>2
<211>1569
<212>dna
<213>人工序列
<400>2
atgaccaagaacgagcccgttttggcagcagacgtcgctgaagctaccctccccgagatc60
ccgccaacagtcgaggatctcaaaaaagctaaaaacgcaagcttctcgtatcggcatgct120
ttccccgtccataccaagaacatcccgagccccctgtcgaaggaagcgccgccagagagt180
tatagaggcctcgtcaacctcgggatgctgcttctattcggcaacaacgttcggttggtg240
atagagaactataagaagtatggagttctgatctccatgcctggatcaaaggtttcgaag300
gaggactggatcctgacaggtctctcccacattctgttgcccttgaatgttcttttgggg360
cttcagttggagcgctgggcaaagaagagtgccatgggtcatcgcaaacgcctcgctgac420
gctactgcagctctcaagaacggaggcatgaatgtcaaggaccccaagcaggctttcaag480
caggttaaaagcatcgaagccgccgctcaacaatcgcagaggctgttcggatggctgcat540
gcgctcaatgtgatcctgatcatctcttggccctcatacatgtcctacttcaagatcttc600
catcctttcatgtcgattacctgcctgatgaacgccaccattcaattcctaaagatgaca660
tccttctcgcttgtcaatcaagacctcagggctgcatacatctttgaccttccagaggac720
aaattccaccatttgactcgggtgcacgatgccaactcgggcaacacagcaacccaacga780
ggagaggtataccagtacgatgttcaataccctgacaacatcacctttatgaacgtggcc840
tatttttggttcgcaccaacgctctgttaccagccttcgtaccctcgaacgacggttttt900
cgcaagtcgttcttccttaagcgcgtgggcgagatggttacctgtgtcggaatgatgtac960
tttctaatcgagcagtacgcgacaccaaccctgcaaaattctgttcgggctgtggaccag1020
ctcgcattcgggacgatgcttgaaagagttcttaagctgagcacaaccagtgttctcatt1080
tggctcctcatgttctacacgttcttccatgccacctttaatgctctgggtgagcttttg1140
tattttggcgatcgctgcttttatctcgcttggtggaatgcaggttctgttggccgatac1200
tgggcgctctggaacaggcccgtgtatgtgttttttaaaagacacgtttacctgccactg1260
attagcggaggctcgtcgcccttgacggccatgtttgtgatctttacgatttccgccatt1320
ctgcacgaggtcttgattggattcccgacgcacatgctctatggttatgcattcgcaggc1380
atgtttgcgcagatcccgctcattgccttgacaagaccgctcgagaagtggcgaggtacc1440
ggaactggactcggcaacatgattttctgggtatcgttcacgatcctgggccagccggcc1500
tgtgcgctgctctactattatcactggaccaagcgccatatcgatgacacgccctcgaca1560
gtagcctga1569
<210>3
<211>27
<212>dna
<213>人工序列
<400>3
ccggaattcatgaccaagaacgagccc27
<210>4
<211>28
<212>dna
<213>人工序列
<400>4
ctagtctagatcaggctactgtcgaggg28
<210>5
<211>20
<212>dna
<213>人工序列
<400>5
taatacgactcactataggg20
<210>6
<211>18
<212>dna
<213>人工序列
<400>6
tcggttagagcggatgtg18
- 还没有人留言评论。精彩留言会获得点赞!
- 用于检测α-甲酰基-CoA消旋酶(AMACR)和前列腺癌的系统和方法
- 与赖氨酰氧化酶样蛋白2(loxl2)相关的治疗和诊断方法
- 包含3-甲酰基利福霉素sv和3-甲酰基利福霉素s的3-(4-肉桂基-1-哌嗪基)氨基衍生物的 ...的制作方法
- 用于检测赖氨酰氧化酶样蛋白-2(loxl2)和乳癌的系统和方法
- 一种用固定化脂质酰基转移酶对大豆毛油脱胶的方法
- 作为组织转谷氨酰胺酶的抑制剂的吡啶酮衍生物的制作方法
- 用于转移在具有空位团的基片中形成的薄层的改进方法
- 一种用于人血清谷氨酰基转移酶测定的缓冲液的制作方法
- 使用甘氨酰-tRNA合成酶和钙粘蛋白筛选用于预防或治疗癌症的试剂的方法
- 一种精浆γ-L-谷氨酰基转肽酶活性定量检测试剂盒及应用的制作方法