棉花高衣分相关的SSR核酸序列及其应用的制作方法
本发明涉及棉花育种领域,具体涉及棉花高衣分相关的ssr核酸序列及其应用。
背景技术:
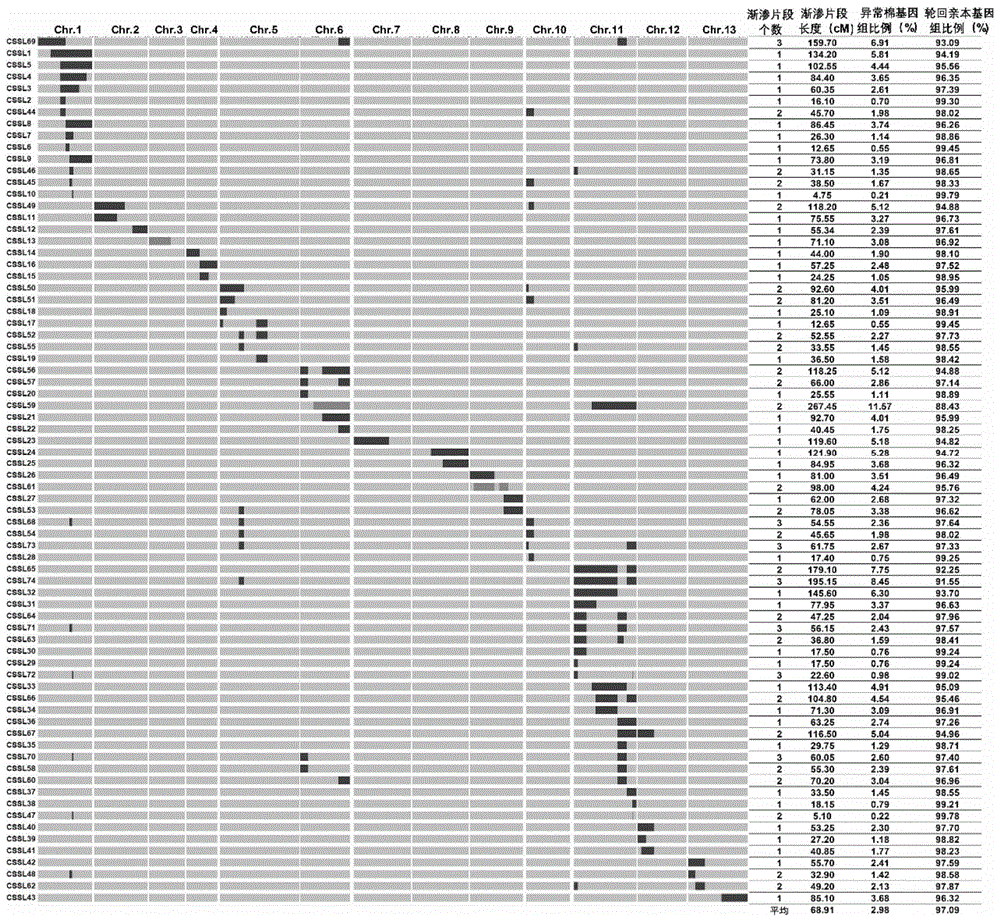
:棉花是一种重要的经济作物,天然棉纤维是世界纺织工业的主要原材料。棉属(gossypiumspp.)中有四个栽培种,包括异源四倍体的陆地棉(gossypiumhirsutuml.)和海岛棉(g.barbadensel.),二倍体的草棉(g.herbaceuml.)和亚洲棉(g.arboreuml.)可以生产纺织用天然纤维。其中,陆地棉因产量高、适应性广等特点主导了世界棉花生产。一直以来,提高产量和改良纤维品质一直是陆地棉品种改良的两个重要目标。随着纺织工业的快速发展,人们对纤维品质的要求越来越高,但在追求优异纤维品质的同时降低了衣分,所以在陆地棉遗传改良中如何协调好纤维品质和产量是目前一个重要的研究课题。棉花产量构成因素有:单株结铃数(numbersofperplant,nb)、单株籽棉产量(seed-cottonyield,sy)、单株皮棉产量(lintyield,ly)、单铃种子数(numberofseedsperboll,ns)、铃重(bollweight,w)、衣指(lintindex,li)、籽指(seedindex,si)和衣分(lintpercentage,lp)等,其中单株结铃数、铃重和衣分是棉花产量构成的三大因素,而衣分是决定棉纤维产量的重要因素,与纤维品质存在显著的负相关关系,所以通过传统的育种手段同步改良棉花纤维品质和提高棉花产量变得越来越困难。分子标记的发展,为育种家提供了一种快速、准确、有效的选择方法。技术实现要素:本发明的目的是提供一种棉花高衣分相关的ssr核酸序列及其应用。第一方面,本发明要求保护一个dna片段。本发明所要求保护的dna片段为位于异常棉5号染色体上至少含有自ssr分子标记jaas6365起至ssr分子标记jaas5604止的片段。其中,所述ssr分子标记jaas6365的核苷酸序列如seqidno.1所示;所述ssr分子标记jaas5604的核苷酸序列如seqidno.2所示。进一步地,所述dna片段为位于异常棉5号染色体上自所述ssr分子标记jaas6365起至所述ssr分子标记jaas5604止的片段。第二方面,本发明要求保护含有第一方面中所述dna片段的载体、表达盒、重组菌或转基因细胞系。所述载体可为人工染色体载体,如细菌人工染色体或酵母人工染色体等。所述重组菌和所述转基因细胞系含有所述人工染色体载体。第三方面,本发明要求保护第一方面中所述的dna片段或第二方面中所述的载体、表达盒、重组菌或转基因细胞系在提高棉花衣分或培育衣分提高的棉花品种中的应用。第四方面,本发明要求保护第一方面中所述dna片段上的ssr分子标记或成套ssr分子标记。本发明所要求保护的ssr分子标记为如下(a1)-(a3)中任一;所述成套ssr分子标记由如下(a1)-(a3)组成;(a1)ssr分子标记jaas6365:核苷酸序列如seqidno.1所示的dna分子;(a2)ssr分子标记jaas5604:核苷酸序列如seqidno.2所示的dna分子;(a3)ssr分子标记jaas0803:核苷酸序列如seqidno.3所示的dna分子。第五方面,本发明要求保护用于鉴定第四方面中所述ssr分子标记或成套ssr分子标记的引物对或成套引物对。本发明所要求保护的引物对为如下(b1)-(b3)中任一;所述成套引物对由如下(b1)-(b3)组成;(b1)用于鉴定所述ssr分子标记jaas6365的引物对1:根据seqidno.1设计得到;(b2)用于鉴定所述ssr分子标记jaas5604的引物对2:根据seqidno.2设计得到;(b3)用于鉴定所述ssr分子标记jaas0803的引物对3:根据seqidno.3设计得到。在本发明的具体实施方式中,所述引物对1由seqidno.4和seqidno.5所示的两条单链dna组成;所述引物对2由seqidno.6和seqidno.7所示的两条单链dna组成;所述引物对3由seqidno.8和seqidno.9所示的两条单链dna组成。第六方面,本发明要求保护含有第五方面中所述引物对或成套引物对的试剂盒。所述试剂盒中还可含有pcr扩增缓冲液、双蒸水、dna聚合酶、dntp等中的任一种或多种。第七方面,本发明要求保护第四方面中所述的ssr分子标记或成套ssr分子标记或第五方面中所述的引物对或成套引物对或第六方面中所述的试剂盒在筛选第一方面中所述dna片段以培育衣分提高的棉花品种中的应用。第八方面,本发明要求保护第四方面中所述的ssr分子标记或成套ssr分子标记或第五方面中所述的引物对或成套引物对或第六方面中所述的试剂盒在鉴定或者辅助鉴定棉花衣分性状中的应用。第九方面,本发明要求保护一种培育衣分提高的棉花品种的方法,包括如下步骤:用至少含有位于异常棉5号染色体上自ssr分子标记jaas6365起至ssr分子标记jaas5604止的片段(如位于异常棉5号染色体上自ssr分子标记jaas6365起至ssr分子标记jaas5604止的片段)替换受体亲本原有染色体片段,得到衣分提高的棉花品种。与现有技术相比,本发明的有益效果为:(1)本发明通过远缘杂交和分子标记辅助选择等技术相结合,将异常棉5号染色体上位于ssr分子标记jaas6365和jaas5604之间的片段置换于陆地棉苏8289遗传背景中,获得了具有染色体片段代换的高衣分棉花系。因此,本发明开发的染色体片段(异常棉5号染色体上位于ssr分子标记jaas6365和jaas5604之间的片段)在今后高衣分棉花育种中具有重要应用价值。(2)分子标记辅助目标片段选择,具有早期鉴定,快速鉴定以及准确性和稳定性高的特点,因此本发明提供的3个ssr分子标记及其引物对有望大大提高棉花育种的衣分性状的选择效率及育种速度,对于加快棉花高衣分新品种的培育进程具有重要意义。(3)本发明提供的高衣分棉花系,通过ssr分子标记及其引物对可用于高衣分基因进行精细定位及相关基因克隆与功能分析,不但可以阐明棉花高衣分性状的分子遗传机制,而且为培育能应用于生产的高衣分品种提供材料与分子基础。附图说明图1为本发明异常棉染色体片段代换系的图示基因型和渐渗片段的分布。浅灰色代表轮回亲本陆地棉苏8289的基因型,黑色代表异常棉的基因型,深灰色代表杂合区段。列代表染色体,从左到右依次为chr.1-chr.13;行从上到下依次代表74个染色体片段代换系,每个染色体片段代换系右侧对应了渐渗片段个数、长度、占异常棉基因组比例以及占轮回亲本基因组比例的信息。图2为渐渗系的异常棉染色体片段在13条染色体上的覆盖率。图3为本发明bc4f4胶图。a为ssr分子标记jaas6365;b为ssr分子标记jaas0803;c为ssr分子标记jaas5604。图4为本发明ssr分子标记jaas5604的bc4f3胶图。图5为本发明中异常棉5号染色体片段与衣分的分析图。图6为本发明中高衣分单片段代换系cssl18与轮回亲本苏8289的衣分差异柱形图。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。陆地棉86-1(g.hirsutumvar.86-1)、陆地棉苏8289(g.hirsutumvar.su8289)和异常棉(gossypiumanomalum):均记载于“caijiaozhai,pengxu,xiazhang等.developmentofgossypiumanomalumderivedmicrosatellitemarkersandtheiruseforgenome-wideidentificationofrecombinationbetweentheg.anomalumandg.hirsutumgenomes.theoreticalandappliedgenetics,2015,128(8):1531-1540”中,公众可从申请人处获得,仅可用于重复本发明实验使用,不得他用。实施例1、与棉花高衣分性状相关的染色体片段的确定及相关分子标记的开发本发明中的高衣分棉花,由以下步骤培育得到:以陆地棉86-1为供体,与父本异常棉杂交,获得三倍体f1(a1d1b1),通过0.15%的秋水仙素处理三倍体幼苗幼嫩的腋芽部位得到可育的六倍体f1(a1a1d1d1b1b1);以六倍体为母本,与亲本陆地棉苏8289进行杂交,得到五倍体a1a1d1d1b1,种植全部收到的种子,得到97棵单株;得到的五倍体与苏8289回交,得到bc2f1的种子,全部种植,获得384棵单株,重组类型50个,其中36个重组类型可育;将bc2f1的所有个体与苏8289回交,全部种植,共获得4331个bc3f1单株,40个重组类型;将bc3f1群体中的重组个体和附加系个体与苏8289继续回交,获得8540个bc4f1单株,56个重组类型;将bc4f1群体中的56个重组类型进行自交,获得自交的bc4f2种子,种植bc4f2群体,共获得4543个单株。本发明中重组类型采用ssr标记检测,ssr标记的来源:将异常棉特异ssr标记与已公布的棉花全基因组ssr物理图谱进行整合,筛选一套均匀覆盖基因组的异常棉特异ssr标记(230个)并分析该群体,获得异常棉渐渗片段的标记基因型。从bc2f1开始,选择重组个体继续回交两次,自交三次,每一世代根据bc2f1重组断裂点的标记信息,从覆盖基因组的230个标记中挑选130个均匀覆盖染色体组的关键ssr标记,利用pcr技术检测所有重组个体,每一世代群体大小见表1,最终获得了一套纯合的异常棉染色体片段代换系(渐渗系)。pcr扩增条件:以陆地棉苏8289、异常棉、六倍体f1和杂交后代为模板进行基因型检。pcr反应体系为10μl:dna模板2μl、10×buffer(mg2+)1μl、dntp0.2μl、taqdna聚合酶0.1μl、前后引物各0.5μl、ddh2o3.7μl。pcr热循环程序:第一步:95℃预变性5min;第二步:94℃变性30s;第三步:55℃退火45s;第四步:72℃延伸1min(第二步到第四步重复30个循环);第五步:72℃延伸10min;第六步:10℃保存。pcr扩增产物的检测:第一步:清洗玻璃板,晾干后备用;第二步:把成套的两块玻璃板对齐,平放,两边用夹子夹紧,至于平板上;第三步:用玻璃棒引流,缓慢倒入8%的聚丙烯酰胺凝胶(29:1),插入梳子。放置15min;第四步:待胶凝固后,松掉夹子,把有凝胶的玻璃板装入电泳槽中,倒入1×tbe电泳缓冲液,并用夹子两边加紧,继续加入电泳缓冲液,没过凝胶,拔梳子;第五步:点样(上样量为1μl),180v恒压环境下电泳1.5h。将2015年所获bc4f1群体中的所有重组个体进行自交,收获bc4f2种子,2016年夏种植bc4f2群体,总共获得4543个单株。根据遗传图谱和bc2f1统计数据重组断裂点的标记信息,挑选130对关键的均匀覆盖染色体组的ssr引物检测所有bc4f2群体,获得51个重组类型,其中45个为纯合基因型。2016年12月进一步种植bc4f3群体,共1533个单株。根据遗传图谱和bc2f1统计数据重组断裂点的标记信息,挑选130对关键的均匀覆盖染色体组的ssr引物检测所有bc4f3群体,获得53个重组类型,其中47个为纯合基因型。2017年种植bc4f4群体,共2225个单株,用本课题组开发筛选的均匀覆盖异常棉染色体组的230对ssr(包含130对引物在内)引物进行检测,共获得了74个重组类型,其中71个为纯合的基因型。每一世代群体大小组成详见表1,每一世代重组类型个数见表2。表1bc3f1、bc4f1、bc4f2、bc4f3、bc4f4世代用于分子标记辅助选择的群体大小注:第二、第三和第四列括号内数字为筛选到的附加系单株数。1、bc4f2、bc4f3和bc4f4世代重组类型的检测将2015年所获得的bc4f1群体中的所有重组个体进行自交,收获bc4f2种子,2016年夏种植bc4f2群体,共获得4543个单株,根据遗传图谱和bc2f1重组断裂点的标记信息,挑选130对关键的均匀覆盖染色体组的ssr引物进行检测,获得51个重组类型,其中45个为纯合基因型,详见表2。与bc4f1相比,出现了8个新的重组类型,其异常棉染色体片段所在的标记区间分别为chr.6:nau2714-jaas1095、nau2714-jaas1095&nau1272-jaas2480、jaas6227-jaas2480;chr.9:jaas1923-jaas0613;chr.10:jaas1256-jaas3294;chr.11:jaas4829-nau3703、nau3703-nau3234和chr.13:jaas4570-jaas2038。2016年12月的海南bc4f3群体,共1533个单株,根据遗传图谱和bc2f1重组断裂点的标记信息,挑选130对关键的均匀覆盖染色体组的ssr引物进行检测,获得了53个重组类型,其中有47个为纯合的代换系(表2)。与bc4f2相比,在bc4f3世代出现了2个新的重组类型,其异常棉染色体片段所在的标记区间分别是chr.5:dc40130-dc40130和chr.11:jaas0280-jaas3199。2017年种植bc4f4群体,共2225个单株,用本课题组开发筛选的均匀覆盖异常棉染色体组的230对ssr(包含130对引物在内)引物进行检测,共获得了74个重组类型,其中71个为纯合的基因型。与bc4f3相比较,在bc4f4中检测到了20种新的重组类型,其异常棉染色体片段所在的标记区间分别是chr.1:jaas0826-jaas1148、nau3615-jaas1148、nau3615-jaas5817、nau3615-jaas0392、nau5100-nau4045、nau2083-nau2083、nau2083-jaas2569、nau4045-nau4045;chr.4:jaas2022-jaas2076;chr.8:nau1037-jaas6420;chr.10:jaas3294-jaas3294、jaas1256-jaas1256;chr.11:jaas4829-jaas4829、jaas3088-jaas5224、jaas4259-jaas4259。由于所有个体均进行了全基因组水平的标记扫描,检测到的新重组类型中存在不同染色体渐渗区段的组合类型。异常棉染色体片段代换系群体的构建过程中连续六个世代检测重组类型的情况汇总详见表2。从bc2f1到bc4f4每一代均有新的重组类型产生,同时由于后代个体育性差和种子活力低等原因,每一代均存在重组类型丢失的现象。为了弥补重组类型丢失的情况,在bc3f1、bc4f1和bc4f2代进行重组个体与轮回亲本回交的同时也进行了附加系与轮回亲本的杂交,使得部分丢失的重组类型得以恢复,也产生了更多的来源于丢失类型的新重组类型。重组类型数目由最初的50个(bc2f1)增加到74个(bc4f4),并最终获得能够稳定遗传的异常棉染色体片段代换系71个(基因型纯合)。具体地,不同群体鉴定的重组类型个数如表2所示。表2bc2f1、bc3f1、bc4f1、bc4f2、bc4f3和bc4f4世代中鉴定的重组类型个数染色体bc2f1bc3f1bc4f1bc4f2bc4f3bc4f4chr.13498813chr.24563(2)3(2)3chr.31131(1)1(1)1(1)chr.41532(1)2(1)3chr.5466677chr.6223556(1)chr.7101111chr.8103112chr.93223(1)3(1)3(1)chr.10122113chr.1111111113(1)14(1)17chr.12323333chr.13104444合计36405651(6)53(6)74*(3)注:括号中的数字表示杂合重组类型个数。*由于在bc4f4代检测到同时包含两条以上染色体片段的重组类型,因此合计总数大于以染色体为单位累加的重组类型总数。2、异常棉染色体片段代换系的鉴定在bc4f4世代,用230对均匀覆盖异常棉基因组的特异ssr引物对所有重组个体进行全基因组前景与背景鉴定,最终确定74个稳定遗传的染色体片段代换系,以cssl1-cssl74依次编号,其中单片段代换系43个,两片段代换系24个,三片段代换系7个;有1个单片段代换系和2个两片段代换系的渐渗片段还保持杂合状态,其余71个为纯合染色体片段代换系。两片段和三片段代换系的染色体片段主要为chr.1、chr.2、chr.5、chr.6、chr.9、chr.10、chr.11、chr.12和chr.13上渐渗片段之间的组合。染色体片段代换系的种类及对应的渐渗片段类型具体见表3。表3bc4f4中检测到的异常棉染色体渐渗片段及获得的染色体片段代换系注:*该渐渗片段为杂合状态。下划线标注表示利用附加系回交获得。+标注的为两片段代换系,#标注的为三片段代换系,没有标注的为单片段代换系。通过ggt软件(vanberloor.ggt2.0:versatilesoftwareforvisualizationandanalysisofgeneticdata.journalofheredity,2008,99(2):232-236.https://doi.org/10.1093/jhered/esm109),按照young和tanksley(youngnd,tanksleysd.restrictionfragmentlengthpolymorphismmapsandtheconceptofgraphicalgenotypes.theoreticalandappliedgenetics,1989,77(1):95-101.https://doi.org/10.1007/bf00292322)的方法推断株系的染色体组成,界定供体片段的位置和大小等。对74个染色体片段代换系的染色体片段的分布进行作图(图1),可以发现,对角线上的异常棉片段(黑色)覆盖了绝大多数的基因组,多数染色体片段代换系的片段之间有重叠。染色体片段代换系中异常棉片段的长度在4.75cm(cssl10)到267.45cm(cssl59)之间,平均长度68.91cm,累加长度达4972.79cm,相当于异常棉基因组总长度的2.15倍。而由于渐渗片段之间有重叠,所有渐渗片段覆盖异常棉染色体的总长度为1607.1cm,覆盖率约为70%(图2)。具有chr.1和chr.11染色体片段的染色体片段代换系数目较多,分别为20个和27个,对这两条异常棉染色体的覆盖率均达到了100%。含有chr.3和chr.7染色体片段的染色体片段代换系数目最少,都仅有1个,但由于片段长度较长,染色体覆盖率并不低(分别为59.9%和61.2%)。而覆盖率最低的为chr.10,仅有17.9%。从图1中还可发现,对角线以外的浅灰色区域上的黑色和深灰色区段并不多,说明这套染色体片段代换系材料的陆地棉苏8289的遗传背景回复率较高,变幅从88.43%(cssl59)到99.79%(cssl10),平均回复率达97.09%。bc4f4株系的ssr检测胶图如图3所示。图3中a为ssr分子标记jaas6365的检测结果;第1泳道为marker,第2泳道为陆地棉苏8289的基因型,第3泳道为陆地棉86-1的基因型,第4泳道为异常棉的基因型,第5泳道为六倍体f1的基因型,第6泳道为cssl18的基因型,第7泳道为cssl50的基因型,第8泳道为cssl51的基因型,第9泳道为cssl19的基因型,第10泳道为cssl52的基因型,第11泳道为cssl53的基因型。图3中b为ssr分子标记jaas0803的检测结果;第1泳道为marker,第2泳道为苏8289的基因型,第3泳道为86-1的基因型,第4泳道为异常棉的基因型,第5泳道为六倍体f1的基因型,第6泳道为cssl18的基因型,第7泳道为cssl50的基因型,第8泳道为cssl51的基因型,第9泳道为cssl19的基因型,第10泳道为cssl52的基因型,第11泳道为cssl53的基因型。图3中c为ssr分子标记jaas5604的检测结果;第1泳道为marker,第2泳道为苏8289的基因型,第3泳道为86-1的基因型,第4泳道为异常棉的基因型,第5泳道为六倍体f1的基因型,第6泳道为cssl18的基因型,第7泳道为cssl50的基因型,第8泳道为cssl51的基因型,第9泳道为cssl19的基因型,第10泳道为cssl52的基因型,第11泳道为cssl53的基因型。cssl18、cssl50和cssl51的基因型与六倍体f1的基因型相同,原因如下:异常棉b1与陆地棉at亚组易发生重组。一般情况下,陆地棉dt亚组还存在一个与at亚组同源的未发生重组的ssr位点。pcr扩增时,引物与异常棉b1组ssr位点结合的同时,还与dt亚组的ssr位点结合,因此,cssl18、cssl50和cssl51的基因型与六倍体f1相同。为了验证这一结论,在cssl18的bc4f3群体中,选择自交的20个单株进行基因型检测,结果发现,其基因型不分离,表明三个染色体片段代换系已稳定遗传,为纯合的染色体片段代换系(图4)。图4为ssr分子标记jaas5604在cssl18的bc4f3世代检测结果。第1泳道为marker,第2泳道为苏8289的基因型,第3泳道为86-1的基因型,第4泳道为异常棉的基因型,第5泳道为六倍体f1的基因型,第6泳道-第25泳道为bc4f3群体中20个单株的基因型。ssr标记分析:在基因型数据收集时将与苏8289和86-1相同的带型记为“1”,与异常棉相同的带型记为“2”,与f1相同的带型记为“3”,带型缺失或模糊记为“-”。由图3所示的bc4f4株系的ssr检测胶图可见,ssr分子标记jaas6365和jaas0803的检测结果中,三个染色体片段代换系cssl50、cssl51和cssl18的带型均与异常棉带型一致(“2”)。ssr分子标记jaas5604在三个染色体片段代换系cssl50、cssl51和cssl18的带型虽与六倍体f1带型一致(“3”),但在cssl50、cssl51和cssl18的bc4f3世代不分离(图4),表明三个染色体片段代换系已稳定遗传,为纯合的染色体片段代换系。因此,cssl50、cssl51和cssl18均含有位于异常棉5号染色体上的ssr分子标记jaas6365、jaas5604和jaas0803。统计分析发现,高衣分棉花系均存在来自异常棉第5号染色体的渐渗片段。为了验证陆地棉chr.5渗入的异常棉片段对衣分的遗传贡献,我们分析了5号染色体的染色体片段代换系的基因型和衣分。结果见图5。图5中,左侧部分为5号染色体不同染色体片段代换系在5号染色体的遗传背景分析。浅灰色表示苏8289的基因片段,黑色表示异常棉基因片段。右侧部分为不同染色体片段代换系和轮回亲本苏8289的衣分表型统计。*表示差异显著。从图5可以看出,5号染色体的染色体片段代换系具有不同的代换片段,其中3个染色体片段代换系cssl50、cssl51和cssl18均含有特定的异常棉ssr分子标记jaas6365、jaas0803和jaas5604(即3个染色体片段代换系cssl50、cssl51和cssl18均含有位于异常棉5号染色体上自ssr分子标记jaas6365起至ssr分子标记jaas5604止的片段),并且其衣分与轮回亲本苏8289相比具有不同程度的提升。而其余3个染色体片段代换系不含这三个ssr分子标记的异常棉染色体片段,其衣分与轮回亲本苏8289相比差异不显著。这在一定程度上验证了这三个ssr分子标记jaas6365、jaas0803和jaas5604所在的异常棉染色体片段对高衣分的遗传贡献。采用winqtlcart2.5软件的单标记分析方法,将5号染色体的所有23个标记与衣分性状进行了相关性分析,结果显示:标记jaas6365、jaas0803和jaas5604与衣分存在显著相关性,其余标记与衣分无相关性(表4)。表4标记jaas6365、jaas0803和jaas5604与衣分的相关性分析性状染色体标记p值r2衣分chr.5jaas63650.008321950.111衣分chr.5jaas08030.008321950.111衣分chr.5jaas56040.008321950.111将株系cssl18在不同的环境种植,环境1:2017年江苏南京江苏省农业科学院溧水植物科学基地;环境2:2018年江苏南京江苏省农业科学院溧水植物科学基地;环境3:2018年新疆库尔勒。重复:各试验点均为随机区组排列,重复3-4次,种植密度和生产管理均与试验地点当地大田生产相同。统计衣分,如表5所示。表5株系cssl18与轮回亲本苏8289的衣分表型统计注:“-”表示未检测。从表5可以看出,第5号染色体一个单片段代换系cssl18(在构建一套异常棉染色体片段代换系群体的过程中,在每一世代通过ssr分子标记辅助选择鉴定,cssl18与轮回亲本苏8289相比,差别仅在于5号染色体上的相应片段被“位于异常棉5号染色体上自ssr分子标记jaas6365起至ssr分子标记jaas5604止的片段”所代换,基因组其余部分与轮回亲本苏8289完全一致),其衣分在不同环境下为41.97-46.57,显著高于轮回亲本苏8289(39.87-43.13),可作为棉花高产育种和衣分分子遗传机制研究的一个重要材料。图6对应于表5的数值。图6中,表示cssl18与苏8289的衣分差异显著。由上述实验结果,可见:含有位于异常棉5号染色体上自ssr分子标记jaas6365起至ssr分子标记jaas5604止的染色体片段可提高棉花衣分。该染色体片段上的3个ssr分子标记(jaas6365、jaas0803和jaas5604)以及开发的引物对可以用于筛选目标染色体片段以培育衣分提高的棉花品种。其中,各ssr分子标记及其检测引物的具体信息如下:ssr分子标记jaas6365的核苷酸序列如seqidno.1所示。ssr分子标记jaas5604的核苷酸序列如seqidno.2所示;ssr分子标记jaas0803的核苷酸序列如seqidno.3所示。用于检测ssr分子标记jaas6365的引物(即图3中a检测所用引物对)如下:jaas6365-f:5’-agcatccaaaacccatttgct-3’(seqidno.4);jaas6365-r:5’-accgcatcctaaggaaagct-3’(seqidno.5)。用于检测ssr分子标记jaas5604的引物(即图3中c以及图4检测所用引物对)如下:jaas5604-f:5’-tgacgtcgttgatccacctc-3’(seqidno.6);jaas5604-r:5’-tcccatgggtgtggtaaaacc-3’(seqidno.7)。用于检测ssr分子标记jaas0803的引物(即图3中b检测所用引物对)如下:jaas0803-f:5’-actttttgcattatctaaggttctgt-3’(seqidno.8);jaas0803-r:5’-accgatactctttttccctgca-3’(seqidno.9)。尽管已用具体实施例来说明和描述了本发明,然而应意识到,在不背离本发明的精神和范围的情况下可以做出许多其它的更改和修改。因此,这意味着在所附权利要求中包括属于本发明范围内的所有这些变化和修改。<110>江苏省农业科学院<120>棉花高衣分相关的ssr核酸序列及其应用<130>gncln190978<160>9<170>patentinversion3.5<210>1<211>180<212>dna<213>artificialsequence<400>1agcatccaaaacccatttgcttcacataaattgtcatgggaaagctcaatcttgcttcaa60ttttcactgcagtcaccattactgaaccactcttgaagaaagctttcacaccaccaccac120cacaagcgttttgattgttatttttctttccctttggcaaagctttccttaggatgcggt180<210>2<211>274<212>dna<213>artificialsequence<400>2tgacgtcgttgatccacctcgtaagttacttcaatcaacagagataaagacatggttgct60tggtgtagttattataataatttgaaaattatgaaattaaatatataatatcctttaatt120tcatctgaaatttggaattttagaagtaatttaagacatgtatgaaaaaaaaaaaagaag180gtatttgcatctatatgcatgtaagttatttatttatgcgttggtttatttttatggacc240ttgattagcttagggttttaccacacccatggga274<210>3<211>213<212>dna<213>artificialsequence<400>3actttttgcattatctaaggttctgtttcctctctctctctctctcacaaagtatttggg60tcattatgttatttgctaaaatggtactaaaatatcttttactcgaaagatttctcaaaa120agaacattttctttgagaagcatgtatctaacctagtaatgataccataaaaacagaacg180taaactaactttgcagggaaaaagagtatcggt213<210>4<211>21<212>dna<213>artificialsequence<400>4agcatccaaaacccatttgct21<210>5<211>20<212>dna<213>artificialsequence<400>5accgcatcctaaggaaagct20<210>6<211>20<212>dna<213>artificialsequence<400>6tgacgtcgttgatccacctc20<210>7<211>21<212>dna<213>artificialsequence<400>7tcccatgggtgtggtaaaacc21<210>8<211>26<212>dna<213>artificialsequence<400>8actttttgcattatctaaggttctgt26<210>9<211>22<212>dna<213>artificialsequence<400>9accgatactctttttccctgca22当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1