在杆状病毒/Sf9系统中大规模生产rAAV的表达载体的制作方法
相关申请的交叉引用
本申请要求享有以下的权益:2018年10月15日提交的标题为在杆状病毒/sf9系统中大规模生产raav的表达载体(expressionvectorsforlarge-scaleproductionofraavinthebaculovirus/sf9system)的美国临时专利申请号62/745,703;其内容通过引用整体并入本文。
参考序列表
本申请与电子格式的序列表一起提交。序列表以标题为20571514pctsl.txt的文件形式提供,该文件创建于2019年10月15日,大小为25,091字节。序列表的电子格式中的信息通过引用整体并入本文。
发明领域
本公开描述了用于生产腺相关病毒(aav)颗粒(包括重组腺相关病毒(raav)颗粒)的方法和系统。在某些实施方案中,生产方法和系统在aav颗粒的生产中使用杆状病毒表达载体(bev)。在某些实施方案中,生产方法和系统使用草地贪夜蛾(spodopterafrugiperda)昆虫细胞(例如sf9或sf21)作为病毒生产细胞。在某些实施方案中,生产方法和系统使用缺少v-cath基因或包含v-cath基因的突变失活形式的bev。
背景
aav已经成为将基因转移到哺乳动物细胞中最广泛研究和利用的病毒载体之一。参见,例如,tratschin等人,mol.cellbiol.,5(11):3251-3260(1985)和grimm等人,hum.genether.,10(15):2445-2450(1999),其内容通过引用整体并入本文。腺相关病毒(aav)载体是治疗性基因递送的有希望的候选者,并且在临床试验中已被证明是安全有效的。为此目的设计和生产改进的aav颗粒是一个活跃的研究领域。
随着aav领域的发展,仍然需要用于生产aav载体(例如aav颗粒)和相应的基因疗法生产材料如杆状病毒感染的昆虫细胞(biic)的改进的系统和方法。
发明概述
本公开提出了用于生产重组腺相关病毒(raav)载体的工程化核酸构建体。本公开描述了用于生产重组腺相关病毒(raav)载体的病毒生产系统和病毒生产细胞(例如昆虫细胞),其包括工程化核酸构建体。本公开描述了生产本公开的病毒生产细胞的方法,以及使用本公开的工程化核酸构建体、病毒生产系统和病毒生产细胞来生产重组腺相关病毒(raav)载体的方法。
本公开描述了用于生产腺相关病毒(aav)载体(例如重组腺相关病毒(raav)载体)的核酸构建体。在某些实施方案中,核酸构建体是aav表达构建体。
本公开描述了生产本公开的病毒生产细胞的方法。在某些实施方案中,该方法包括以下步骤:提供本公开的一种或多种表达构建体;提供本公开的有效载荷构建体;将一种或多种表达构建体和有效载荷构建体转染到病毒生产细胞中。在某些实施方案,病毒生产细胞是哺乳动物细胞。在某些实施方案中,病毒生产细胞是昆虫细胞。在某些实施方案中,病毒生产细胞是aav病毒生产细胞。在某些实施方案中,该方法包括以下步骤:提供本公开的一种或多种aav表达构建体;提供本公开的aav有效载荷构建体;和将一种或多种aav表达构建体和aav有效载荷构建体转染到aav病毒生产细胞中。在某些实施方案中,该方法包括以下步骤:提供本公开的病毒生产系统;和将病毒生产系统转染到病毒生产细胞中。
本公开提出了用于改进aav颗粒生产的方法和策略。这样的方法包括使用一种或多种重组杆状病毒表达载体(bev)在昆虫细胞系中生产aav颗粒的方法。在某些实施方案中,bev缺少v-cath基因的一部分或全部,或包括v-cath基因的突变失活形式。在某些实施方案中,bev缺少v-cath基因的一部分或全部。在某些实施方案中,bev包括v-cath基因的突变失活形式。
本公开提出了用于改进aav颗粒生产的方法和策略,其导致aav生产蛋白,特别是aav衣壳蛋白(vp1、vp2和vp3)的更高产量和更少的蛋白质降解。
在某些实施方案中,本公开的方法可以用于采用病毒生产细胞的生产系统,所述病毒生产细胞包括昆虫细胞,包括(但不限于)sf9、sf21和highfivetmbti-tn-5b1-4细胞系。在某些实施方案中,本公开的方法可用于采用重组bev系统,例如bacmagictm(millipore)、flashbactm(oxfordexpressiontechnologies)和bestbac2.0tm(expressionsystems)的生产系统中。
在某些实施方案中,本公开提出了腺相关病毒(aav)表达构建体,其包括vp编码区,该vp编码区包含编码选自vp1、vp2和vp3的一种或多种aav衣壳蛋白的核苷酸序列。在某些实施方案中,aav表达构建体是杆状病毒表达载体(bev)。在某些实施方案中,bev不包括功能性杆状病毒v-cath基因。在某些实施方案中,bev缺少杆状病毒v-cath基因的一部分或全部。在某些实施方案中,bev包含杆状病毒v-cath基因的突变失活形式。
在某些实施方案中,一种或多种aav衣壳蛋白的血清型选自voy101、voy201、aavphp.b(php.b)、aavphp.a(php.a)、aavg2b-26、aavg2b-13、aavth1.1-32、aavth1.1-35、aavphp.b2(php.b2)、aavphp.b3(php.b3)、aavphp.n/php.b-dgt、aavphp.b-est、aavphp.b-ggt、aavphp.b-atp、aavphp.b-att-t、aavphp.b-dgt-t、aavphp.b-ggt-t、aavphp.b-sgs、aavphp.b-aqp、aavphp.b-qqp、aavphp.b-snp(3)、aavphp.b-snp、aavphp.b-qgt、aavphp.b-nqt、aavphp.b-egs、aavphp.b-sgn、aavphp.b-egt、aavphp.b-dst、aavphp.b-dst、aavphp.b-stp、aavphp.b-pqp、aavphp.b-sqp、aavphp.b-qlp、aavphp.b-tmp、aavphp.b-ttp、aavphp.s/g2a12、aavg2a15/g2a3(g2a3)、aavg2b4(g2b4)、aavg2b5(g2b5)、php.s、aav1、aav2、aav2g9、aav3、aav3a、aav3b、aav3-3、aav4、aav4-4、aav5、aav6、aav6.1、aav6.2、aav6.1.2、aav7、aav7.2、aav8、aav9、aav9.11、aav9.13、aav9.16、aav9.24、aav9.45、aav9.47、aav9.61、aav9.68、aav9.84、aav9.9、aav10、aav11、aav12、aav16.3、aav24.1、aav27.3、aav42.12、aav42-1b、aav42-2、aav42-3a、aav42-3b、aav42-4、aav42-5a、aav42-5b、aav42-6b、aav42-8、aav42-10、aav42-11、aav42-12、aav42-13、aav42-15、aav42-aa、aav43-1、aav43-12、aav43-20、aav43-21、aav43-23、aav43-25、aav43-5、aav44.1、aav44.2、aav44.5、aav223.1、aav223.2、aav223.4、aav223.5、aav223.6、aav223.7、aav1-7/rh.48、aav1-8/rh.49、aav2-15/rh.62、aav2-3/rh.61、aav2-4/rh.50、aav2-5/rh.51、aav3.1/hu.6、aav3.1/hu.9、aav3-9/rh.52、aav3-11/rh.53、aav4-8/r11.64、aav4-9/rh.54、aav4-19/rh.55、aav5-3/rh.57、aav5-22/rh.58、aav7.3/hu.7、aav16.8/hu.10、aav16.12/hu.11、aav29.3/bb.1、aav29.5/bb.2、aav106.1/hu.37、aav114.3/hu.40、aav127.2/hu.41、aav127.5/hu.42、aav128.3/hu.44、aav130.4/hu.48、aav145.1/hu.53、aav145.5/hu.54、aav145.6/hu.55、aav161.10/hu.60、aav161.6/hu.61、aav33.12/hu.17、aav33.4/hu.15、aav33.8/hu.16、aav52/hu.19、aav52.1/hu.20、aav58.2/hu.25、aava3.3、aava3.4、aava3.5、aava3.7、aavc1、aavc2、aavc5、aav-dj、aav-dj8、aavf3、aavf5、aavh2、aavrh.72、aavhu.8、aavrh.68、aavrh.70、aavpi.1、aavpi.3、aavpi.2、aavrh.60、aavrh.44、aavrh.65、aavrh.55、aavrh.47、aavrh.69、aavrh.45、aavrh.59、aavhu.12、aavh6、aavlk03、aavh-1/hu.1、aavh-5/hu.3、aavlg-10/rh.40、aavlg-4/rh.38、aavlg-9/hu.39、aavn721-8/rh.43、aavch.5、aavch.5r1、aavcy.2、aavcy.3、aavcy.4、aavcy.5、aavcy.5r1、aavcy.5r2、aavcy.5r3、aavcy.5r4、aavcy.6、aavhu.1、aavhu.2、aavhu.3、aavhu.4、aavhu.5、aavhu.6、aavhu.7、aavhu.9、aavhu.10、aavhu.11、aavhu.13、aavhu.15、aavhu.16、aavhu.17、aavhu.18、aavhu.20、aavhu.21、aavhu.22、aavhu.23.2、aavhu.24、aavhu.25、aavhu.27、aavhu.28、aavhu.29、aavhu.29r、aavhu.31、aavhu.32、aavhu.34、aavhu.35、aavhu.37、aavhu.39、aavhu.40、aavhu.41、aavhu.42、aavhu.43、aavhu.44、aavhu.44r1、aavhu.44r2、aavhu.44r3、aavhu.45、aavhu.46、aavhu.47、aavhu.48、aavhu.48r1、aavhu.48r2、aavhu.48r3、aavhu.49、aavhu.51、aavhu.52、aavhu.54、aavhu.55、aavhu.56、aavhu.57、aavhu.58、aavhu.60、aavhu.61、aavhu.63、aavhu.64、aavhu.66、aavhu.67、aavhu.14/9、aavhu.t19、aavrh.2、aavrh.2r、aavrh.8、aavrh.8r、aavrh.10、aavrh.12、aavrh.13、aavrh.13r、aavrh.14、aavrh.17、aavrh.18、aavrh.19、aavrh.20、aavrh.21、aavrh.22、aavrh.23、aavrh.24、aavrh.25、aavrh.31、aavrh.32、aavrh.33、aavrh.34、aavrh.35、aavrh.36、aavrh.37、aavrh.37r2、aavrh.38、aavrh.39、aavrh.40、aavrh.46、aavrh.48、aavrh.48.1、aavrh.48.1.2、aavrh.48.2、aavrh.49、aavrh.51、aavrh.52、aavrh.53、aavrh.54、aavrh.56、aavrh.57、aavrh.58、aavrh.61、aavrh.64、aavrh.64r1、aavrh.64r2、aavrh.67、aavrh.73、aavrh.74、aavrh8r、aavrh8ra586r突变体、aavrh8rr533a突变体、aaav、baav、山羊aav、牛aav、aavhe1.1、aavher1.5、aavher1.14、aavher1.8、aavher1.16、aavher1.18、aavher1.35、aavher1.7、aavher1.36、aavher2.29、aavher2.4、aavher2.16、aavher2.30、aavher2.31、aavher2.36、aavher1.23、aavher3.1、aav2.5t、aav-paec、aav-lk01、aav-lk02、aav-lk03、aav-lk04、aav-lk05、aav-lk06、aav-lk07、aav-lk08、aav-lk09、aav-lk10、aav-lk11、aav-lk12、aav-lk13、aav-lk14、aav-lk15、aav-lk16、aav-lk17、aav-lk18、aav-lk19、aav-paec2、aav-paec4、aav-paec6、aav-paec7、aav-paec8、aav-paec11、aav-paec12、aav-2-pre-mirna-101、aav-8h、aav-8b、aav-h、aav-b、aavsm10-2、aav改组100-1、aav改组100-3、aav改组100-7、aav改组10-2、aav改组10-6、aav改组10-8、aav改组100-2、aavsm10-1、aavsm10-8、aavsm100-3、aavsm100-10、bnp61aav、bnp62aav、bnp63aav、aavrh.50、aavrh.43、aavrh.62、aavrh.48、aavhu.19、aavhu.11、aavhu.53、aav4-8/rh.64、aavlg-9/hu.39、aav54.5/hu.23、aav54.2/hu.22、aav54.7/hu.24、aav54.1/hu.21、aav54.4r/hu.27、aav46.2/hu.28、aav46.6/hu.29、aav128.1/hu.43、真型aav(ttaav)、upennaav10、日本aav10血清型、aavcbr-7.1、aavcbr-7.10、aavcbr-7.2、aavcbr-7.3、aavcbr-7.4、aavcbr-7.5、aavcbr-7.7、aavcbr-7.8、aavcbr-b7.3、aavcbr-b7.4、aavcbr-e1、aavcbr-e2、aavcbr-e3、aavcbr-e4、aavcbr-e5、aavcbr-e5、aavcbr-e6、aavcbr-e7、aavcbr-e8、aavcht-1、aavcht-2、aavcht-3、aavcht-6.1、aavcht-6.10、aavcht-6.5、aavcht-6.6、aavcht-6.7、aavcht-6.8、aavcht-p1、aavcht-p2、aavcht-p5、aavcht-p6、aavcht-p8、aavcht-p9、aavckd-1、aavckd-10、aavckd-2、aavckd-3、aavckd-4、aavckd-6、aavckd-7、aavckd-8、aavckd-b1、aavckd-b2、aavckd-b3、aavckd-b4、aavckd-b5、aavckd-b6、aavckd-b7、aavckd-b8、aavckd-h1、aavckd-h2、aavckd-h3、aavckd-h4、aavckd-h5、aavckd-h6、aavckd-n3、aavckd-n4、aavckd-n9、aavclg-f1、aavclg-f2、aavclg-f3、aavclg-f4、aavclg-f5、aavclg-f6、aavclg-f7、aavclg-f8、aavclv-1、aavclv1-1、aavclv1-10、aavclv1-2、aavclv-12、aavclv1-3、aavclv-13、aavclv1-4、aavclv1-7、aavclv1-8、aavclv1-9、aavclv-2、aavclv-3、aavclv-4、aavclv-6、aavclv-8、aavclv-d1、aavclv-d2、aavclv-d3、aavclv-d4、aavclv-d5、aavclv-d6、aavclv-d7、aavclv-d8、aavclv-e1、aavclv-k1、aavclv-k3、aavclv-k6、aavclv-l4、aavclv-l5、aavclv-l6、aavclv-m1、aavclv-m11、aavclv-m2、aavclv-m5、aavclv-m6、aavclv-m7、aavclv-m8、aavclv-m9、aavclv-r1、aavclv-r2、aavclv-r3、aavclv-r4、aavclv-r5、aavclv-r6、aavclv-r7、aavclv-r8、aavclv-r9、aavcsp-1、aavcsp-10、aavcsp-11、aavcsp-2、aavcsp-3、aavcsp-4、aavcsp-6、aavcsp-7、aavcsp-8、aavcsp-8.10、aavcsp-8.2、aavcsp-8.4、aavcsp-8.5、aavcsp-8.6、aavcsp-8.7、aavcsp-8.8、aavcsp-8.9、aavcsp-9、aav.hu.48r3、aav.vr-355、aav3b、aav4、aav5、aavf1/hsc1、aavf11/hsc11、aavf12/hsc12、aavf13/hsc13、aavf14/hsc14、aavf15/hsc15、aavf16/hsc16、aavf17/hsc17、aavf2/hsc2、aavf3/hsc3、aavf4/hsc4、aavf5/hsc5、aavf6/hsc6、aavf7/hsc7、aavf8/hsc8、aavf9/hsc9、aavrh20、aavrh32/33、aavrh39、aavrh46、aavrh73、aavrh74、aavhu.26,或其变体或嵌合体。在某些实施方案中,一种或多种aav衣壳蛋白的血清型是voy101。
在某些实施方案中,本公开提出了一种aav病毒生产系统,其包括本公开的aav病毒生产细胞和aav表达构建体。在某些实施方案中,aav病毒生产系统包括aav有效载荷构建体。在某些实施方案中,aav病毒生产细胞是昆虫细胞。在某些实施方案中,aav病毒生产细胞是sf9细胞。在某些实施方案中,aav病毒生产细胞是sf21细胞。
在某些实施方案中,本公开提出了在aav病毒生产细胞中生产重组腺相关病毒(raav)载体的方法。在某些实施方案中,该方法包括:提供一种aav病毒生产系统,其包括本公开的aav表达构建体和aav有效载荷构建体;将aav病毒生产系统转染到aav病毒生产细胞中;使aav病毒生产细胞暴露于允许aav病毒生产细胞将aav表达构建体和aav有效载荷构建体加工成raav颗粒的条件;以及从aav病毒生产细胞中收集raav颗粒。在某些实施方案中,aav病毒生产细胞是昆虫细胞。在某些实施方案中,aav病毒生产细胞是sf9细胞或sf21细胞。
在某些实施方案中,本公开提出了在使用杆状病毒表达载体(bev)的杆状病毒表达系统中减少aav蛋白的蛋白酶降解的方法。在某些实施方案中,该方法包括从bev中删除杆状病毒v-cath基因的一部分或全部。在某些实施方案中,该方法包括使bev的杆状病毒v-cath基因突变失活。
在以下描述中阐述了本公开的各种实施方案的细节。通过说明书、附图以及权利要求书,本公开的其他特征、目的和优点将变得显而易见。在说明书中,单数形式也包括复数,除非上下文另外明确指出。除非另有定义,否则本文使用的所有技术和科学术语具有与本公开所属领域的普通技术人员通常所理解的相同含义。在有冲突的情况下,以本说明书为准。
附图简要说明
根据以下本公开的特定实施方案的描述,前述和其他目的、特征和优点将是显而易见的,如附图中所示。附图不必然按比例或全面绘制,而是将重点放在说明本公开的各种实施方案的原理上。
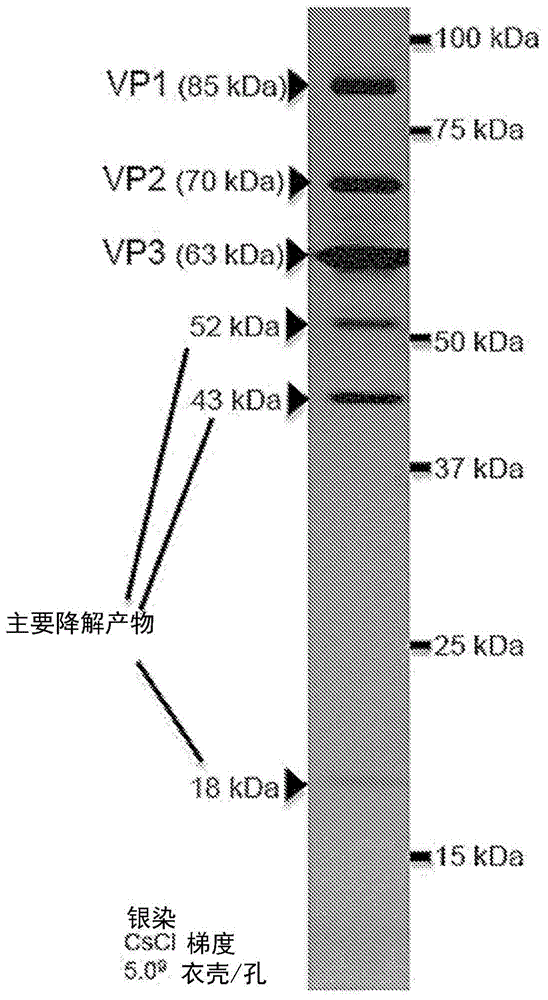
图1是使用bev/sf9系统从aav9变体衣壳生产的纯化的aav衣壳蛋白的银染凝胶。
图2是vp1、vp2和vp3衣壳蛋白与鉴定的裂解产物(包括18kda产物)比对的比对结果。
图3是v-cath杆状病毒基因保守性的比较。摘自handbookofproteolyticenzymes2003(a.j.barrett,rawlings和j.fwoesnered.)academicpress,2ndedition,[557]viralcathepsin,j.slack。
图4a示出了v-cath基因的三个pcr筛选。
图4b是三个pcr筛选的pcr产物的凝胶,显示如预期的那样v-cath基因座出现。
图5是显示v-cath缺失对细胞活力的影响的图。
图6是显示v-cath缺失对总细胞计数的影响的图。
图7是显示v-cath缺失对细胞直径的影响的图。
图8是显示v-cath缺失对蛋白酶活性的影响的图。
图9是显示在替代缓冲液中v-cath缺失对蛋白酶活性的影响的图。
图10a是显示v-cath缺失对精氨酸缓冲液中降解产物的出现的影响的凝胶。
图10b是显示v-cath缺失对精氨酸缓冲液中降解产物的出现的影响的凝胶。
发明详述
i.腺相关病毒(aav)
概述
腺相关病毒(aav)是细小病毒科(parvoviridae)的小的非包膜二十面体衣壳病毒,其特征在于单链dna病毒基因组。细小病毒科病毒由两个亚科组成:感染脊椎动物的细小病毒亚科(parvovirinae)和感染无脊椎动物的浓核病毒亚科(densovirinae)。细小病毒科包括能够在脊椎动物宿主(包括但不限于人类、灵长类、牛、犬、马和绵羊物种)中复制的依赖病毒属(dependovirus),其包括aav。
细小病毒和细小病毒科的其他成员在kennethi.berns,“parvoviridae:thevirusesandtheirreplication,”fieldsvirology(1996年底3版)第69章中概述,因为涉及细小病毒其内容通过引用整体并入本文。
aav由于其结构相对简单、具有感染多种细胞(包括静止和分裂细胞)而无需整合到宿主基因组中且无需复制的能力以及相对温和的免疫原性,已被证明可用作生物学工具。可以操纵病毒的基因组以使其含有用于组装功能性重组病毒或病毒颗粒的最少组分,所述重组病毒或病毒颗粒装载有或工程化以靶向特定组织并表达或递送所需的有效载荷。
aav病毒基因组
野生型aav病毒基因组是线性、单链dna(ssdna)分子,长度约为5,000个核苷酸(nt)。反向末端重复序列(itr)传统上在5'和3'端都给病毒基因组加帽,为病毒基因组提供了复制起点。尽管不希望受到理论的束缚,但aav病毒基因组通常包括两个itr序列。这些itr具有特征性t形发夹结构,该结构由形成能量稳定的双链区域的ssdna的5'和3'端的自我互补区域(在野生型aav中为145nt)定义。双链发夹结构包括多种功能,包括但不限于通过充当宿主病毒复制细胞的内源性dna聚合酶复合物的引物来用作dna复制的起点。
野生型aav病毒基因组还包括两个开放阅读框的核苷酸序列,一个为四个非结构rep蛋白(rep78、rep68、rep52、rep40,由rep基因编码),且一个为三个衣壳或结构蛋白(vp1、vp2、vp3,由衣壳基因或cap基因编码)。rep蛋白对于复制和包装重要,而衣壳蛋白则组装形成aav的蛋白壳或aav衣壳。rep78和rep68可以从p5启动子转录,而rep52和rep40可以从p19启动子转录。可变剪接和可变起始密码子和启动子导致从单个开放阅读框产生四种不同的rep蛋白以及从单个开放阅读框产生三种衣壳蛋白。尽管其随aav血清型而变化,但作为非限制性实例,对于aav9/hu.14(us7,906,111中的seqidno:123,由于涉及aav9/hu.14,其内容通过引用整体并入本文),vp1是指氨基酸1-736,vp2是指氨基酸138-736,而vp3是指氨基酸203-736。换句话说,vp1是全长衣壳序列,而vp2和vp3是完整的较短组成部分。结果,vp3区域中序列的变化也是vp1和vp2的变化,但是,与亲本序列相比的百分比差异对于vp3将是最大的,因为它是三个序列中最短的序列。尽管在此关于氨基酸序列进行了描述,但是可以类似地描述编码这些蛋白的核酸序列。这三种衣壳蛋白合起来组装形成aav衣壳蛋白。尽管不希望受到理论的束缚,但aav衣壳蛋白通常包括摩尔比为1:1:10的vp1:vp2:vp3。如本文所用,“aav血清型”主要由aav衣壳定义。在一些情况下,还通过aav血清型(例如,aav2/9)具体描述itr。
为了用作生物学工具,可以修饰野生型aav病毒基因组,以用包含具有至少一个itr区域的有效载荷区域的核酸序列替换rep/cap序列。通常,在重组aav病毒基因组中有两个itr区域。rep/cap序列可以在生产过程中反式提供以生产aav颗粒。
除编码的异源有效载荷外,aav载体还可包含全部或部分的任何天然存在的和/或重组的aav血清型核苷酸序列或变体的病毒基因组。aav变体可在核酸(基因组或衣壳)和氨基酸水平(衣壳)上具有显著同源性的序列,以产生通常为物质和功能等同物的构建体,通过相似的机制复制,并通过相似的机制组装。参见chiorini等人,j.vir.71:6823-33(1997);srivastava等人,j.vir.45:555-64(1983);chiorini等人,j.vir.73:1309-1319(1999);rutledge等人,j.vir.72:309-319(1998);和wu等人,j.vir.74:8635-47(2000),由于涉及aav变体和等同物,其各自内容通过引用整体并入本文。
在某些实施方案中,本公开的aav颗粒、病毒基因组和/或有效载荷及其使用方法可以如wo2017189963中所述,由于涉及aav颗粒、病毒基因组和/或有效载荷,其内容通过引用整体并入本文。
本公开的aav颗粒可以配制成本公开的任何基因治疗制剂,包括对本领域技术人员显而易见的此类制剂的任何变化形式。在本申请中提及的“aav颗粒”、“aav颗粒制剂”和“配制的aav颗粒”是指可以配制的aav颗粒和被配制而没有限制的那些。
在某些实施方案中,本公开的aav颗粒是复制缺陷的重组aav(raav)病毒颗粒,在其病毒基因组内缺乏编码功能性rep和cap蛋白的序列。这些缺陷的aav颗粒可能缺乏大多数或全部亲本编码序列,并且基本上仅携带一个或两个aavitr序列和用于递送至细胞、组织、器官或生物体的目标核酸(即有效载荷)。
在某些实施方案中,本公开的aav颗粒的病毒基因组包括至少一种控制元件,该控制元件提供了其中编码的编码序列的复制、转录和翻译。并非总是需要存在所有的控制元件,只要编码序列能够在适当的宿主细胞中复制、转录和/或翻译即可。表达控制元件的非限制性实例包括用于转录起始和/或终止的序列、启动子和/或增强子序列、有效的rna加工信号(例如剪接和聚腺苷酸化信号)、稳定细胞质mrna的序列、增强翻译效率的序列(例如,kozak共有序列)、增强蛋白稳定性的序列和/或增强蛋白加工和/或分泌的序列。
根据本公开,用于治疗和/或诊断的aav颗粒包括已被提取(distilled)或减少至转导目标核酸有效载荷或货物所需的最小组分的病毒。以这种方式,aav颗粒被工程化为用于特异性递送的载体,同时缺少在野生型病毒中发现的有害复制和/或整合特征。
本公开的aav颗粒可以重组产生,并且可以基于腺相关病毒(aav)亲本或参考序列。如本文所用,“载体”是转运、转导或以其他方式充当异源分子(例如本文所述的核酸)的载体的任何分子或实体。
除了单链aav病毒基因组(例如ssaav)之外,本公开还提供了自互补aav(scaav)病毒基因组。scaav载体基因组包含dna链,它们退火在一起形成双链dna。通过跳过第二链合成,scaav可在细胞中快速表达。
在某些实施方案中,本公开的aav病毒基因组是scaav。在某些实施方案中,本公开的aav病毒基因组是ssaav。
本领域公开了产生和/或修饰aav颗粒的方法,例如假型aav颗粒(pct专利公开号wo200028004;wo200123001;wo2004112727;wo2005005610和wo2005072364,由于涉及产生和/或修饰aav颗粒,其各自的内容通过引用整体并入本文)。
可以修饰aav颗粒以增强递送效率。这样的修饰的aav颗粒可以被有效地包装,并被用于以高频率和最小毒性成功地感染靶细胞。在某些实施方案中,根据美国公开号us20130195801中描述的方法工程化aav颗粒的衣壳,由于涉及修饰aav颗粒以增强递送效率,其内容通过引用整体并入本文。
在某些实施方案中,可以将包含编码本公开的多肽或蛋白的有效载荷区的aav颗粒引入到哺乳动物细胞中。
反向末端重复序列(itr)
本公开的aav颗粒包含具有至少一个itr区和一个有效载荷区的病毒基因组。在某些实施方案中,病毒基因组具有两个itr。这两个itr在5'和3'端位于有效载荷区的侧翼。itr用作包含复制识别位点的复制的起点。itr包含可以互补和对称排列的序列区。引入本公开的病毒基因组中的itr可以由天然存在的多核苷酸序列或重组衍生的多核苷酸序列组成。
itr可以衍生自与衣壳相同的血清型或其衍生物。itr可以具有与衣壳不同的血清型。在某些实施方案中,aav颗粒具有多于一个itr。在非限制性实例中,aav颗粒具有包含两个itr的病毒基因组。在某些实施方案中,itr具有彼此相同的血清型。在另一个实施方案中,itr具有不同的血清型。非限制性实例包括具有与衣壳相同血清型的零个、一个或两个itr。在某些实施方案中,aav颗粒的病毒基因组的两个itr均为aav2itr。
独立地,每个itr可以具有约100-约150个核苷酸长度。itr可以具有约100-105个核苷酸长度、106-110个核苷酸长度、111-115个核苷酸长度、116-120个核苷酸长度、121-125个核苷酸长度、126-130个核苷酸长度、131-135个核苷酸长度、136-140个核苷酸长度、141-145个核苷酸长度或146-150个核苷酸长度。在某些实施方案中,itr具有140-142个核苷酸长度。itr长度的非限制性实例是102、130、140、141、142、145个核苷酸长度,以及与其具有至少95%同一性的那些核苷酸。
在某些实施方案中,每个itr的长度可以是141个核苷酸。在某些实施方案中,每个itr的长度可以是130个核苷酸。在某些实施方案中,每个itr的长度可以是119个核苷酸。
在某些实施方案中,aav颗粒包括两个itr,并且一个itr的长度为141个核苷酸,而另一个itr的长度为130个核苷酸。在某些实施方案中,aav颗粒包括两个itr,并且这两个itr的长度均为141个核苷酸。
启动子
在某些实施方案中,病毒基因组的有效载荷区包含至少一种增强转基因靶特异性和表达的元件(参见例如powell等人,viralexpressioncassetteelementstoenhancetransgenetargetspecificityandexpressioningenetherapy,2015;由于涉及有效载荷/转基因增强子元件,其内容通过引用整体并入本文)。增强转基因靶特异性和表达的元件的非限制性实例包括启动子、内源性mirna、转录后调控元件(pre)、聚腺苷酸化(polya)信号序列和上游增强子(use)、cmv增强子和内含子。
本领域技术人员可以认识到,本公开的多肽在靶细胞中的表达可能需要特异性启动子,包括但不限于物种特异性、诱导型、组织特异性或细胞周期特异性的启动子(参见parr等人,nat.med.3:1145-9(1997);由于涉及多肽表达启动子,其内容通过引用整体并入本文)。
在某些实施方案中,当启动子驱动在aav颗粒的病毒基因组的有效载荷区中编码的多肽的表达时,该启动子被认为是有效的。在某些实施方案中,所述启动子是当其在待靶向的细胞中驱动表达时被认为有效的启动子。在某些实施方案中,启动子是对被靶向的细胞具有嗜性的启动子。在某些实施方案中,启动子是对于病毒生产细胞具有嗜性的启动子。
在某些实施方案中,启动子驱动有效载荷在靶细胞或组织中表达一段时间。由启动子驱动的表达可以持续1-31天(或其中的任何值或范围)、1-23个月(或其中的任何值或范围)、2-10年(或其中的任何值或范围)或超过10年。表达可以持续1-5小时、1-12小时、1-2天、1-5天、1-2周、1-3周、1-4周、1-2个月、1-4个月、1-6个月、2-6个月、3-6个月、3-9个月、4-8个月、6-12个月、1-2年、1-5年、2-5年、3-6年、3-8年、4-8年或5-10年。作为非限制性实例,启动子可以是用于在神经(例如cns)细胞或组织中持续表达有效载荷的弱启动子。
在某些实施方案中,启动子驱动本公开的多肽的表达持续至少1-11个月(或其中任何个体值)、2-65年(或其中任何个体值)或超过65年。
启动子可以是天然存在的或非天然存在的。启动子的非限制性实例包括病毒启动子、植物启动子和哺乳动物启动子。在某些实施方案中,启动子可以是人启动子。在某些实施方案中,启动子可以被截短或突变。
在大多数组织中驱动或启动表达的启动子包括但不限于人延伸因子1α亚基(ef1α)、巨细胞病毒(cmv)立即早期增强子和/或启动子、鸡β-肌动蛋白(cba)及其衍生物cag、β葡萄糖醛酸糖苷酶(gusb)或泛素c(ubc)。组织特异性表达元件可用于将表达限制于某些细胞类型,例如但不限于肌肉特异性启动子、b细胞启动子、单核细胞启动子、白细胞启动子、巨噬细胞启动子、胰腺腺泡细胞启动子、内皮细胞启动子、肺组织启动子、星形胶质细胞启动子或神经系统启动子,其可用于将表达限制于神经元或神经元的亚型、星形胶质细胞或少突胶质细胞。
肌肉特异性启动子的非限制性实例包括哺乳动物肌肉肌酸激酶(mck)启动子、哺乳动物结蛋白(des)启动子、哺乳动物肌钙蛋白i(tnni2)启动子和哺乳动物骨骼α-肌动蛋白(aska)启动子(参见例如美国专利公开us20110212529,由于涉及肌肉特异性启动子,其内容通过引用整体并入本文)。
神经元的组织特异性表达元件的非限制性实例包括神经元特异性烯醇化酶(nse)、血小板衍生生长因子(pdgf)、血小板衍生生长因子b链(pdgf-β)、突触蛋白(syn)、甲基-cpg结合蛋白2(mecp2)、ca2+/钙调蛋白依赖性蛋白激酶ii(camkii)、代谢型谷氨酸受体2(mglur2)、神经丝轻肽(neurofilamentlight,nfl)或神经丝重肽(neurofilamentheavy,nfh)、β-珠蛋白小基因nβ2、前脑啡肽原(ppe)、脑啡肽(enk)和兴奋性氨基酸转运蛋白2(eaat2)启动子。星形胶质细胞的组织特异性表达元件的非限制性实例包括神经胶质原纤维酸性蛋白(gfap)和eaat2启动子。少突胶质细胞的组织特异性表达元件的非限制性实例包括髓鞘碱性蛋白(mbp)启动子。
在某些实施方案中,启动子可以小于1kb。启动子的长度可以为200-800个核苷酸(或其中的任何值或范围)或大于800个核苷酸。启动子的长度可以是200-300、200-400、200-500、200-600、200-700、200-800、300-400、300-500、300-600、300-700、300-800、400-500、400-600、400-700、400-800、500-600、500-700、500-800、600-700、600-800或700-800。
在某些实施方案中,启动子可以是相同或不同的起始或亲本启动子的两个或更多个组分的组合,例如但不限于cmv和cba。每个组分的长度可以是200-800个核苷酸(或其中的任何值或范围),或超过800个核苷酸。每个组分的长度可以是200-300、200-400、200-500、200-600、200-700、200-800、300-400、300-500、300-600、300-700、300-800、400-500、400-600、400-700、400-800、500-600、500-700、500-800、600-700、600-800或700-800。在某些实施方案中,启动子是382个核苷酸的cmv-增强子序列和260个核苷酸的cba-启动子序列的组合。
在某些实施方案中,病毒基因组包括遍在启动子。遍在启动子的非限制性实例包括cmv、cba(包括衍生物cag、cbh等)、ef-1α、pgk、ubc、gusb(hgbp)和ucoe(hnrpa2b1-cbx3的启动子)。
yu等人(molecularpain2011,7:63;其内容通过引用整体并入本文)使用慢病毒载体评价了egfp在大鼠drg细胞和原代drg细胞中在cag、efiα、pgk和ubc启动子下的表达,并发现ubc显示出比其他3种启动子更弱的表达,并且所有启动子仅观察到10-12%的神经胶质表达。soderblom等人(e.neuro2015;其内容通过引用整体并入本文)评价了在运动皮层注射后,egfp在具有cmv和ubc启动子的aav8中和在具有cmv启动子的aav2中的表达。鼻内施用含有ubc或efiα启动子的质粒显示比用cmv启动子表达更强的持续气道表达(参见,例如,gill等人,genetherapy2001,vol.8,1539-1546;其内容通过引用整体并入本文)。husain等人(genetherapy2009;其内容通过引用整体并入本文)评价了具有hgusb启动子、hsv-1lat启动子和nse启动子的hβh构建体,并且发现hβh构建体在小鼠脑中显示出比nse弱的表达。passini和wolfe(j.virol.2001,12382-12392,其内容通过引用整体并入本文)评价了在新生小鼠脑室内注射后hβh载体的长期作用,并发现其持续表达至少一年。xu等人(genetherapy2001,8,1323-1332;其内容通过引用整体并入本文)发现,与cmv-lacz、cmv-luc、ef、gfap、henk、nachr、ppe、ppe+wpre、nse(0.3kb)、nse(1.8kb)和nse(1.8kb+wpre)相比,当使用nfl和nfh启动子时,在所有脑区域中表达低。xu等人发现启动子活性按照降序是nse(1.8kb)、ef、nse(0.3kb)、gfap、cmv、henk、ppe、nfl和nfh。nfl是一个650个核苷酸的启动子,而nfh是一个920个核苷酸的启动子,它们在肝脏中都不存在,但是nfh在感觉本体感受神经元、大脑和脊髓中含量丰富,并且nfh存在于心脏中。scn8a是一个470个核苷酸的启动子,其在整个drg、脊髓和脑中表达,并在海马神经元和小脑浦肯野细胞、皮层、丘脑和下丘脑中表现出特别高的表达(参见例如drews等人,identificationofevolutionaryconserved,functionalnoncodingelementsinthepromoterregionofthesodiumchannelgenescn8a,mammgenome(2007)18:723-731;和raymond等人,expressionofalternativelysplicedsodiumchannelα-subunitgenes,journalofbiologicalchemistry(2004)279(44)46234-46241;其各自内容通过引用整体并入本文)。
由前述yu、soderblom、gill、husain、passini、xu、drews或raymond教导的任何启动子都可以用于本公开。
在某些实施方案中,启动子不是细胞特异性的。
在某些实施方案中,该启动子是泛素c(ubc)启动子。ubc启动子可以具有300-350个核苷酸的大小。作为非限制性实例,ubc启动子是332个核苷酸。在某些实施方案中,启动子是β-葡糖醛酸糖苷酶(gusb)启动子。gusb启动子的大小可以是350-400个核苷酸。作为非限制性实例,gusb启动子是378个核苷酸。在某些实施方案中,启动子是神经丝轻肽(nfl)启动子。nfl启动子可以具有600-700个核苷酸的大小。作为非限制性实例,nfl启动子是650个核苷酸。在某些实施方案中,启动子是神经丝重肽(nfh)启动子。nfh启动子可以具有900-950个核苷酸的大小。作为非限制性实例,nfh启动子为920个核苷酸。在某些实施方案中,启动子是scn8a启动子。scn8a启动子的大小可以是450-500个核苷酸。作为非限制性实例,scn8a启动子为470个核苷酸。
在某些实施方案中,启动子是共济蛋白(fxn)启动子。在某些实施方案中,启动子是磷酸甘油酸激酶1(pgk)启动子。在某些实施方案中,启动子是鸡β-肌动蛋白(cba)启动子或其变体。在某些实施方案中,启动子是cb6启动子。在某些实施方案中,启动子是最小cb启动子。在某些实施方案中,启动子是巨细胞病毒(cmv)启动子。在某些实施方案中,启动子是h1启动子。在某些实施方案中,启动子是cag启动子。在某些实施方案中,启动子是gfap启动子。在某些实施方案中,启动子是突触蛋白启动子。在某些实施方案中,启动子是工程化的启动子。在某些实施方案中,启动子是肝或骨骼肌启动子。肝启动子的非限制性实例包括人α-1-抗胰蛋白酶(haat)和甲状腺素结合球蛋白(tbg)。骨骼肌启动子的非限制性实例包括结蛋白、mck或合成c5-12。在某些实施方案中,启动子是rnapoliii启动子。作为非限制性实例,rnapoliii启动子是u6。作为非限制性实例,rnapoliii启动子是h1。在某些实施方案中,启动子是心肌细胞特异性启动子。心肌细胞特异性启动子的非限制性实例包括αmhc、ctnt和cmv-mlc2k。在某些实施方案中,病毒基因组包含两个启动子。作为非限制性实例,启动子是ef1α启动子和cmv启动子。
在某些实施方案中,病毒基因组包括增强子元件、启动子和/或5'utr内含子。增强子元件,在本文中也称为“增强子”,可以是但不限于cmv增强子,启动子可以是但不限于cmv、cba、ubc、gusb、nse、突触蛋白、mecp2和gfap启动子,并且5'utr/内含子可以是但不限于sv40和cba-mvm。作为非限制性实例,组合使用的增强子、启动子和/或内含子可以是:(1)cmv增强子、cmv启动子、sv405'utr内含子;(2)cmv增强子、cba启动子、sv405'utr内含子;(3)cmv增强子、cba启动子、cba-mvm5'utr内含子;(4)ubc启动子;(5)gusb启动子;(6)nse启动子;(7)突触蛋白启动子;(8)mecp2启动子和(9)gfap启动子。
在某些实施方案中,病毒基因组包括工程化的启动子。
在另一个实施方案中,病毒基因组包括来自天然表达的蛋白的启动子。
非翻译区(utr)
根据定义,基因的野生型非翻译区(utr)被转录但未被翻译。通常,5'utr在转录起始位点开始并在起始密码子处终止,3'utr在终止密码子后立即开始,并一直持续到转录终止信号为止。
通常在特定靶器官的大量表达基因中发现的特征可以被工程化为utr,以增强稳定性和蛋白产生。作为非限制性实例,来自正常在肝脏中表达的mrna(例如白蛋白、血清淀粉样蛋白a、载脂蛋白a/b/e、转铁蛋白、甲胎蛋白、促红细胞生成素或因子viii)的5'utr可以用在本公开的aav颗粒的病毒基因组中,以增强在肝细胞系或肝脏中的表达。
尽管不希望受到理论的束缚,野生型5'非翻译区(utr)包含在翻译起始中起作用的特征。通常已知参与核糖体启动许多基因翻译的过程的kozak序列通常包含在5'utr中。kozak序列具有共有ccr(a/g)ccaugg,其中r是起始密码子(atg)上游3个碱基的嘌呤(腺嘌呤或鸟嘌呤),其后是另一个'g'。在某些实施方案中,病毒基因组中的5'utr包含kozak序列。在某些实施方案中,病毒基因组中的5'utr不包含kozak序列。
尽管不希望受到理论的束缚,已知野生型3'utr具有嵌入其中的一段腺苷和尿苷。这些富含au的标记在具有高周转率的基因中特别普遍。基于它们的序列特征和功能特性,富含au的元件(are)可以分为三类(chen等人,1995,由于涉及富含au的元件,其内容通过引用整体并入本文):i类are,例如但不限于c-myc和myod,在富含u的区域内包含数个auuua基序的分散拷贝。ii类are,例如但不限于gm-csf和tnf-a,具有两个或多个重叠的uuauuua(u/a)(u/a)九聚体。iii类are的定义不太明确,例如但不限于c-jun和生肌蛋白(myogenin)。这些富含u的区域不包含auuua基序。已知大多数与are结合的蛋白都会使信使不稳定,而elav家族的成员(最值得注意的是hur)已被证明可以增加mrna的稳定性。hur结合到所有三类的are。将hur特异性结合位点工程化到核酸分子的3'utr中将导致hur结合,并且从而使体内信使稳定。
可以使用3'utr富含au的元件(are)的引入、去除或修饰来调节多核苷酸的稳定性。当工程化特定多核苷酸(例如病毒基因组的有效载荷区)时,可以引入一个或多个are拷贝,以使多核苷酸的稳定性降低,从而减少翻译并降低所得蛋白的生产。同样,可以识别并去除或突变are,以增加细胞内稳定性,并且从而增加所得蛋白的翻译和生产。
在某些实施方案中,病毒基因组的3'utr可以包括寡聚(dt)序列,用于模板化添加poly-a尾。
在某些实施方案中,病毒基因组可以包括至少一个mirna种子、结合位点或完整序列。microrna(或mirna或mir)是19-25个核苷酸的非编码rna,可与核酸靶位点结合并通过降低核酸分子的稳定性或抑制翻译来下调基因表达。microrna序列包括“种子”区域,即成熟microrna的2-8位区域中的序列,该序列与核酸的mirna靶序列具有完美的watson-crick互补性。
在某些实施方案中,可以对病毒基因组工程化以包括、改变或去除至少一个mirna结合位点、序列或种子区域。
可将来自本领域已知的任何基因的任何utr引入到aav颗粒的病毒基因组中。这些utr或其部分可以以与由其选择它们的基因中相同的方向放置,或者可以改变方向或位置。在某些实施方案中,可以将aav颗粒的病毒基因组中使用的utr反转、缩短、延长、用本领域已知的一种或多种其他5'utr或3'utr制成。如本文所使用的,术语“改变”在涉及utr时,表示相对于参考序列已经以某种方式改变了utr。例如,可以通过如上所述的方向或位置的改变来相对于野生型或天然utr改变3'或5'utr,或者可以通过包括额外的核苷酸、核苷酸的缺失、核苷酸的交换或转座来改变3'或5'utr。
在某些实施方案中,aav颗粒的病毒基因组包括至少一种人工utr,其不是野生型utr的变体。
在某些实施方案中,aav颗粒的病毒基因组包括utr,所述utr选自其蛋白具有共同功能、结构、特征或特性的转录物家族。
聚腺苷酸化序列
在某些实施方案中,本公开的aav颗粒的病毒基因组包括至少一个聚腺苷酸化序列。aav颗粒的病毒基因组可以在有效载荷编码序列的3'端与3'itr的5'端之间包括聚腺苷酸化序列。
在某些实施方案中,聚腺苷酸化序列或“polya序列”的长度范围可以从不存在到约500个核苷酸。聚腺苷酸化序列的长度可以是但不限于1-500个核苷酸(或其中的任何值或范围)。
在某些实施方案中,聚腺苷酸化序列的长度为127个核苷酸。在某些实施方案中,聚腺苷酸化序列的长度为477个核苷酸。在某些实施方案中,聚腺苷酸化序列的长度为552个核苷酸。
内含子
在某些实施方案中,载体基因组包含至少一种增强转基因靶标特异性和表达的元件(参见例如powell等人,viralexpressioncassetteelementstoenhancetransgenetargetspecificityandexpressioningenetherapy,2015,由于涉及转基因靶向增强子,其内容通过引用整体并入本文),例如内含子。内含子的非限制性实例包括mvm(67-97bp)、f.ix截短的内含子1(300bp)、β-珠蛋白sd/免疫球蛋白重链剪接受体(250bp)、腺病毒剪接供体/免疫球蛋白剪接受体(500bp)、sv40晚期剪接供体/剪接受体(19s/16s)(180bp)和杂种腺病毒剪接供体/igg剪接受体(230bp)。
在某些实施方案中,内含子或内含子部分的长度可以是100-500个核苷酸。内含子的长度可以是80、90、100、110、120、130、140、150、160、170、171、172、173、174、175、176、177、178、179、180、190、200、210、220、230、240、250、260、270、280、290、300、310、320、330、340、350、360、370、380、390、400、410、420、430、440、450、460、470、480、490或500。内含子的长度可以是80-100、80-120、80-140、80-160、80-180、80-200、80-250、80-300、80-350、80-400、80-450、80-500、200-300、200-400、200-500、300-400、300-500或400-500。
填充物序列
在某些实施方案中,病毒基因组包含至少一种改善包装效率和表达的元件,例如填充物序列或填充序列。填充物序列的非限制性实例包括白蛋白和/或α-1抗胰蛋白酶。可以操控任何已知的病毒、哺乳动物或植物序列以用作填充物序列。
在某些实施方案中,填充物或填充序列的长度可以是约100-3500个核苷酸。填充物序列的长度可以是大约100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800、1900、2000、2100、2200、2300、2400、2500、2600、2700、2800、2900或3000。
mirna
在某些实施方案中,病毒基因组包含至少一个编码mirna以减少转基因在特定组织中表达的序列。mirna及其所靶向的组织是本领域众所周知的。作为非限制性实例,可以在病毒基因组中编码mir-122mirna以减少病毒基因组在肝脏中的表达。
有效载荷
本公开的aav颗粒可以包含含有至少一个有效载荷区的至少一种有效载荷构建体,或使用其生产。在某些实施方案中,有效载荷区可以位于病毒基因组内,例如有效载荷构建体的病毒基因组内。在有效载荷区的5’和/或3’端,可能存在至少一个反向末端重复序列(itr)。在有效载荷区内可以存在启动子区、内含子区和编码区。
在某些实施方案中,本公开的有效载荷构建体可以是杆粒,也称为杆状病毒质粒或重组杆状病毒基因组。
在某些实施方案中,aav颗粒的有效载荷区包含一个或多个编码目标多肽或蛋白的核酸序列。
在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码多于一种目标多肽的核酸序列。在某些实施方案中,可以复制编码一种或多种多肽的病毒基因组,并将其包装到病毒颗粒中。用包含载体基因组的病毒颗粒转导的靶细胞可以在单个靶细胞中表达一种或多种多肽中的每一种。
如果aav颗粒有效载荷区编码多肽,该多肽可以是肽、多肽或蛋白。作为非限制性实例,有效载荷区可以编码至少一种目标治疗性蛋白。编码本文描述的多肽的aav病毒基因组可用于人类疾病、病毒、感染、兽医应用和各种体内和体外环境的领域。
在某些实施方案中,将配制的aav颗粒(其包含病毒基因组)施用于受试者将增加蛋白在受试者中的表达。在某些实施方案中,蛋白表达的增加将减少与有效载荷编码的多肽相关的疾病或病症的作用和/或症状。
在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码目标蛋白(即有效载荷蛋白、治疗性蛋白)的核酸序列。
在某些实施方案中,有效载荷区包含编码蛋白的核酸序列,所述蛋白包括但不限于抗体、芳香族l-氨基酸脱羧酶(aadc)、apoe2、共济蛋白、存活的运动神经元(smn)蛋白、葡萄糖脑苷脂酶、n-磺基葡糖胺磺基水解酶、n-乙酰基-α-氨基葡糖苷酶、艾杜糖2-硫酸酯酶、α-l-艾杜糖醛酸酶(alpha-l-iduronidase)、棕榈酰蛋白硫酯酶1、三肽基肽酶1、battenin、cln5、cln6(linclin)、mfsd8、cln8、天冬氨酸酰积分转移酶(aspa)、颗粒蛋白前体(progranulin)(grn)、mecp2、β-半乳糖苷酶(glb1)和/或巨轴索神经病蛋白(gigaxonin)(gan)。
在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码在以下国际公开中的任一个中描述的任何疾病相关性蛋白(及其片段或变体)的核酸序列:wo2016073693、wo2017023724、wo2018232055、wo2016077687、wo2016077689、wo2018204786、wo2017201258、wo2017201248、wo2018204803、wo2018204797、wo2017189959、wo2017189963、wo2017189964、wo2015191508、wo2016094783、wo20160137949、wo2017075335;其各自内容通过引用整体并入本文,只要他们不与本公开冲突。
由本公开的病毒基因组的有效载荷区编码的氨基酸序列可以翻译为完整的多肽、多个多肽或多肽片段,其可以独立地由一个或多个核酸、核酸片段或上述任何物质的变体编码。如本文所用,“多肽”是指最通常通过肽键连接在一起的氨基酸残基(天然或非天然)的聚合物。如本文所用,该术语是指任何大小、结构或功能的蛋白、多肽和肽。在某些情况下,编码的多肽小于约50个氨基酸,则将该多肽称为肽。如果多肽是肽,则其长度将是至少约2、3、4或至少5个氨基酸残基。因此,多肽包括基因产物、天然存在的多肽、合成多肽、同系物、直系同源物、旁系同源物、片段和前述的其他等同物、变体和类似物。多肽可以是单分子,也可以是多分子复合物,例如二聚体、三聚体或四聚体。它们还可以包含单链或多链多肽,并且可以被结合或连接。术语多肽还可以应用于氨基酸聚合物,其中一个或多个氨基酸残基是相应的天然存在的氨基酸的人工化学类似物。
在某些实施方案中,提供了“多肽变体”。术语“多肽变体”是指其氨基酸序列与天然序列或参考序列不同的分子。与天然序列或参考序列相比,氨基酸序列变体可在氨基酸序列内的某些位置具有取代、缺失和/或插入。通常,变体与天然序列或参考序列将具有至少约50%的同一性(同源性),并且在某些实施方案中,它们与天然序列或参考序列具有至少约80%或至少约90%的同一性(同源性)。
本公开包括配制的aav颗粒作为治疗剂的用途,所述aav颗粒的载体基因组编码调节性多核苷酸,例如rna或dna分子。因此,本公开提供了编码多核苷酸的载体基因组,所述多核苷酸被加工成靶向目标基因的小双链rna(dsrna)分子(小干扰rna、sirna、mirna、pre-mirna)。本公开还提供了其用于抑制目标基因的等位基因的基因表达和蛋白生产,用于治疗疾病、病症和/或病状的方法。
在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码或包含一种或多种调节性多核苷酸的核酸序列。在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码目标调节性多核苷酸的核酸序列。在本公开的某些实施方案中,调节性多核苷酸,例如rna或dna分子,作为治疗剂提供。rna干扰介导的基因沉默可以特异性抑制所靶向的基因的表达。
在某些实施方案中,有效载荷区包含编码调节性多核苷酸的核酸序列,所述调节性多核苷酸干扰靶基因表达和/或靶蛋白产生。在某些实施方案中,待抑制/修饰的基因表达或蛋白产生可包括但不限于超氧化物歧化酶1(sod1)、染色体9开放阅读框72(c9orf72)、tardna结合蛋白(tardbp)、共济失调蛋白-3(atxn3)、亨廷顿蛋白(htt)、淀粉样前体蛋白(app)、载脂蛋白e(apoe)、微管相关蛋白τ(mapt)、α-突触核蛋白(snca)、电压门控钠通道α亚基9(scn9a)和/或电压门控钠通道α亚基10(scn10a)。
在某些实施方案中,aav颗粒包含具有有效载荷区的病毒基因组,该有效载荷区包含编码以下国际公开任何一个中描述的任何调节性多核苷酸、rnai分子、sirna分子、dsrna分子和/或rna双链体的核酸序列:wo2016073693、wo2017023724、wo2018232055、wo2016077687、wo2016077689、wo2018204786、wo2017201258、wo2017201248、wo2018204803、wo2018204797、wo2017189959、wo2017189963、wo2017189964、wo2015191508、wo2016094783、wo20160137949、wo2017075335;其各自内容通过引用整体并入本文,只要它们不与本公开冲突。
在某些实施方案中,将编码此类sirna分子或sirna分子的单链的核酸序列插入到腺相关病毒载体中,并引入到细胞(特别是中枢神经系统中的细胞)中。
因为几个独特的特征,已经针对sirna递送研究了aav载体。所述特征的非限制性实例包括(i)感染分裂的细胞和非分裂的细胞的能力;(ii)侵染性的广宿主范围,包括人细胞;(iii)野生型aav尚未与任何疾病相关联,且尚未被证实在受感染的细胞中复制;(iv)细胞介导的针对载体的免疫应答的缺乏,和(v)在宿主染色体中的不整合性质,由此减少长期表达的可能。此外,aav颗粒感染对改变细胞基因表达模式均仅具有微小影响(stilwell和samulski等人,biotechniques,2003,34,148)。
在某些实施方案中,本公开的编码的sirna双链体包含杂交在一起形成双链体结构的反义链和有义链,其中所述反义链与所述目标所靶向基因的核酸序列互补,并且其中有义链与目标所靶向基因的核酸序列同源。在其他方面,在每条链的3'端具有0、1或2个核苷酸突出端。
本公开的配制的aav颗粒的有效载荷可以编码一种或多种物质,所述物质经受rna干扰(rnai)诱导的基因表达抑制。本文提供了靶向目标基因的编码的sirna双链体或编码的dsrna(本文统称为“sirna分子”)。此类sirna分子,例如编码的sirna双链体、编码的dsrna或编码的sirna或dsrna前体,可以降低或沉默细胞(例如星形胶质细胞或小胶质细胞神经元、皮层神经元、海马神经元、内嗅神经元、丘脑神经元、感觉神经元或运动神经元)中的基因表达。
rnai(也称为转录后基因沉默(ptgs)、压抑或共抑制)是转录后基因沉默的过程,其中rna分子以序列特异性方式抑制基因表达,通常是通过导致破坏特异性的mrna分子。rnai的活性组分是短/小双链rna(dsrna),称为小干扰rna(sirna),其通常包含15-30个核苷酸(例如19至25、19至24或19-21个核苷酸)和2个核苷酸的3’突出端,且其与靶基因的核酸序列匹配。这些短rna物质可由dicer介导的较大dsrna的裂解在体内自然产生,并且它们在哺乳动物细胞中具有功能。
天然表达的小rna分子,称为microrna(mirna),通过调节mrna的表达引起基因沉默。包含rna诱导沉默复合物(risc)的mirna靶向mrna,所述mrna与在mirna的5’区中的核苷酸2-7(称为种子区)和其他具有其3’区的碱基对呈完美序列互补性。mirna介导的基因表达下调可能是由靶标mrna的切割、靶标mrna的翻译抑制或mrna降解引起的。mirna靶向序列通常位于靶mrna的3’-utr中。单个mirna可以靶向来自多种基因的超过100种转录物,而一种mrna可以被不同的mirna靶向。
可以将靶向特定mrna的sirna双链体或dsrna设计成aav颗粒的有效载荷,并引入细胞中以激活rnai过程。elbashir等人证明了21个核苷酸的sirna双链体(称为小干扰rna)能够在不诱导哺乳动物细胞的免疫应答的情况下有效地强烈且特异性地敲低基因(elbashirsm等人,nature,2001,411,494-498)。自从这份初步报告以来,通过sirna进行转录后基因沉默迅速成为哺乳动物细胞遗传分析的有力工具,并具有产生新疗法的潜力。
sirna双链体由与靶mrna同源的有义链和与靶mrna互补的反义链组成,与单链(ss)-sirna(例如反义链rna或反义寡核苷酸)的使用相比,在破坏靶rna的效率方面,sirna双链体具有更多优势。在许多情况下,需要更高浓度的ss-sirna才能实现对应双链体的有效基因沉默效力。
在某些实施方案中,可以在还包含分子支架的调节性多核苷酸中编码sirna分子。如本文所用,“分子支架”是框架或起始分子,其形成设计或制备后续分子所依据的序列或结构基础。
在某些实施方案中,包含有效载荷(例如本文所述的sirna、mirna或其他rnai剂)的调节性多核苷酸包括分子支架,其包含前导5’侧翼序列,该序列可以具有任何长度,并且可以全部或部分衍生自野生型microrna序列或是完全人工的。3’侧翼序列可以在大小和来源上与5’侧翼序列镜像。在某些实施方案中,5’和3’侧翼序列中的一个或两个都不存在。
在某些实施方案中,分子支架可包含一个或多个本领域已知的接头。接头可以将多个区域或将一个分子支架与另一个分开。作为非限制性实例,分子支架可以是多顺反子的。
在某些实施方案中,使用以下特性中的至少一种来设计调节性多核苷酸:环变体、种子错配/凸起/摆动变体、茎错配、环变体和基茎错配变体、种子错配和基茎错配变体、茎错配和基础茎错配变体、种子摆动和基础茎摆动变体或茎序列变体。
基因组大小
在某些实施方案中,包含本文所述的有效载荷的aav颗粒可以是单链或双链载体基因组。载体基因组的大小可以是小、中等、大或最大大小。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文描述的有效载荷的载体基因组可以是小的单链载体基因组。小的单链载体基因组的大小可以为2.1至3.5kb,例如大小为约2.1、2.2、2.3、2.4、2.5、2.6、2.7、2.8、2.9、3.0、3.1、3.2、3.3、3.4和3.5kb。作为非限制性实例,小的单链载体基因组的大小可以是3.2kb。作为另一个非限制性实例,小的单链载体基因组的大小可以是2.2kb。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文描述的有效载荷的载体基因组可以是小的双链载体基因组。小的双链载体基因组的大小可以为1.3至1.7kb,例如大小为约1.3、1.4、1.5、1.6和1.7kb。作为非限制性实例,小的双链载体基因组的大小可以为1.6kb。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文所述的有效载荷例如多核苷酸、sirna或dsrna的载体基因组可以是中等单链载体基因组。中等单链载体基因组的大小可以为3.6至4.3kb,例如大小为约3.6、3.7、3.8、3.9、4.0、4.1、4.2和4.3kb。作为非限制性实例,中等单链载体基因组的大小可以为4.0kb。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文描述的有效载荷的载体基因组可以是中等双链载体基因组。中等双链载体基因组的大小可以为1.8至2.1kb,例如大小为约1.8、1.9、2.0和2.1kb。作为非限制性实例,中等双链载体基因组的大小可以为2.0kb。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文所述的有效载荷的载体基因组可以是大的单链载体基因组。大的单链载体基因组的大小可以是4.4至6.0kb,例如大小为约4.4、4.5、4.6、4.7、4.8、4.9、5.0、5.1、5.2、5.3、5.4、5.5、5.6、5.7、5.8、5.9和6.0kb。作为非限制性实例,大的单链载体基因组的大小可以是4.7kb。作为另一个非限制性实例,大的单链载体基因组的大小可以是4.8kb。作为又一个非限制性实例,大的单链载体基因组的大小可以是6.0kb。另外,载体基因组可以包含启动子和polya尾。
在某些实施方案中,包含本文所述的有效载荷的载体基因组可以是大的双链载体基因组。大的双链载体基因组的大小可以为2.2至3.0kb,例如大小为约2.2、2.3、2.4、2.5、2.6、2.7、2.8、2.9和3.0kb。作为非限制性实例,大的双链载体基因组的大小可以是2.4kb。另外,载体基因组可以包含启动子和polya尾。
aav血清型
本公开的aav颗粒可以包括或衍生自任何天然或重组aav血清型。根据本公开,aav颗粒可以利用或基于血清型或包括选自以下任何一种的肽:voy101、voy201、aavphp.b(php.b)、aavphp.a(php.a)、aavg2b-26、aavg2b-13、aavth1.1-32、aavth1.1-35、aavphp.b2(php.b2)、aavphp.b3(php.b3)、aavphp.n/php.b-dgt、aavphp.b-est、aavphp.b-ggt、aavphp.b-atp、aavphp.b-att-t、aavphp.b-dgt-t、aavphp.b-ggt-t、aavphp.b-sgs、aavphp.b-aqp、aavphp.b-qqp、aavphp.b-snp(3)、aavphp.b-snp、aavphp.b-qgt、aavphp.b-nqt、aavphp.b-egs、aavphp.b-sgn、aavphp.b-egt、aavphp.b-dst、aavphp.b-dst、aavphp.b-stp、aavphp.b-pqp、aavphp.b-sqp、aavphp.b-qlp、aavphp.b-tmp、aavphp.b-ttp、aavphp.s/g2a12、aavg2a15/g2a3(g2a3)、aavg2b4(g2b4)、aavg2b5(g2b5)、php.s、aav1、aav2、aav2g9、aav3、aav3a、aav3b、aav3-3、aav4、aav4-4、aav5、aav6、aav6.1、aav6.2、aav6.1.2、aav7、aav7.2、aav8、aav9、aav9.11、aav9.13、aav9.16、aav9.24、aav9.45、aav9.47、aav9.61、aav9.68、aav9.84、aav9.9、aav10、aav11、aav12、aav16.3、aav24.1、aav27.3、aav42.12、aav42-1b、aav42-2、aav42-3a、aav42-3b、aav42-4、aav42-5a、aav42-5b、aav42-6b、aav42-8、aav42-10、aav42-11、aav42-12、aav42-13、aav42-15、aav42-aa、aav43-1、aav43-12、aav43-20、aav43-21、aav43-23、aav43-25、aav43-5、aav44.1、aav44.2、aav44.5、aav223.1、aav223.2、aav223.4、aav223.5、aav223.6、aav223.7、aav1-7/rh.48、aav1-8/rh.49、aav2-15/rh.62、aav2-3/rh.61、aav2-4/rh.50、aav2-5/rh.51、aav3.1/hu.6、aav3.1/hu.9、aav3-9/rh.52、aav3-11/rh.53、aav4-8/r11.64、aav4-9/rh.54、aav4-19/rh.55、aav5-3/rh.57、aav5-22/rh.58、aav7.3/hu.7、aav16.8/hu.10、aav16.12/hu.11、aav29.3/bb.1、aav29.5/bb.2、aav106.1/hu.37、aav114.3/hu.40、aav127.2/hu.41、aav127.5/hu.42、aav128.3/hu.44、aav130.4/hu.48、aav145.1/hu.53、aav145.5/hu.54、aav145.6/hu.55、aav161.10/hu.60、aav161.6/hu.61、aav33.12/hu.17、aav33.4/hu.15、aav33.8/hu.16、aav52/hu.19、aav52.1/hu.20、aav58.2/hu.25、aava3.3、aava3.4、aava3.5、aava3.7、aavc1、aavc2、aavc5、aav-dj、aav-dj8、aavf3、aavf5、aavh2、aavrh.72、aavhu.8、aavrh.68、aavrh.70、aavpi.1、aavpi.3、aavpi.2、aavrh.60、aavrh.44、aavrh.65、aavrh.55、aavrh.47、aavrh.69、aavrh.45、aavrh.59、aavhu.12、aavh6、aavlk03、aavh-1/hu.1、aavh-5/hu.3、aavlg-10/rh.40、aavlg-4/rh.38、aavlg-9/hu.39、aavn721-8/rh.43、aavch.5、aavch.5r1、aavcy.2、aavcy.3、aavcy.4、aavcy.5、aavcy.5r1、aavcy.5r2、aavcy.5r3、aavcy.5r4、aavcy.6、aavhu.1、aavhu.2、aavhu.3、aavhu.4、aavhu.5、aavhu.6、aavhu.7、aavhu.9、aavhu.10、aavhu.11、aavhu.13、aavhu.15、aavhu.16、aavhu.17、aavhu.18、aavhu.20、aavhu.21、aavhu.22、aavhu.23.2、aavhu.24、aavhu.25、aavhu.27、aavhu.28、aavhu.29、aavhu.29r、aavhu.31、aavhu.32、aavhu.34、aavhu.35、aavhu.37、aavhu.39、aavhu.40、aavhu.41、aavhu.42、aavhu.43、aavhu.44、aavhu.44r1、aavhu.44r2、aavhu.44r3、aavhu.45、aavhu.46、aavhu.47、aavhu.48、aavhu.48r1、aavhu.48r2、aavhu.48r3、aavhu.49、aavhu.51、aavhu.52、aavhu.54、aavhu.55、aavhu.56、aavhu.57、aavhu.58、aavhu.60、aavhu.61、aavhu.63、aavhu.64、aavhu.66、aavhu.67、aavhu.14/9、aavhu.t19、aavrh.2、aavrh.2r、aavrh.8、aavrh.8r、aavrh.10、aavrh.12、aavrh.13、aavrh.13r、aavrh.14、aavrh.17、aavrh.18、aavrh.19、aavrh.20、aavrh.21、aavrh.22、aavrh.23、aavrh.24、aavrh.25、aavrh.31、aavrh.32、aavrh.33、aavrh.34、aavrh.35、aavrh.36、aavrh.37、aavrh.37r2、aavrh.38、aavrh.39、aavrh.40、aavrh.46、aavrh.48、aavrh.48.1、aavrh.48.1.2、aavrh.48.2、aavrh.49、aavrh.51、aavrh.52、aavrh.53、aavrh.54、aavrh.56、aavrh.57、aavrh.58、aavrh.61、aavrh.64、aavrh.64r1、aavrh.64r2、aavrh.67、aavrh.73、aavrh.74、aavrh8r、aavrh8ra586r突变体、aavrh8rr533a突变体、aaav、baav、山羊aav、牛aav、aavhe1.1、aavher1.5、aavher1.14、aavher1.8、aavher1.16、aavher1.18、aavher1.35、aavher1.7、aavher1.36、aavher2.29、aavher2.4、aavher2.16、aavher2.30、aavher2.31、aavher2.36、aavher1.23、aavher3.1、aav2.5t、aav-paec、aav-lk01、aav-lk02、aav-lk03、aav-lk04、aav-lk05、aav-lk06、aav-lk07、aav-lk08、aav-lk09、aav-lk10、aav-lk11、aav-lk12、aav-lk13、aav-lk14、aav-lk15、aav-lk16、aav-lk17、aav-lk18、aav-lk19、aav-paec2、aav-paec4、aav-paec6、aav-paec7、aav-paec8、aav-paec11、aav-paec12、aav-2-pre-mirna-101、aav-8h、aav-8b、aav-h、aav-b、aavsm10-2、aav改组100-1、aav改组100-3、aav改组100-7、aav改组10-2、aav改组10-6、aav改组10-8、aav改组100-2、aavsm10-1、aavsm10-8、aavsm100-3、aavsm100-10、bnp61aav、bnp62aav、bnp63aav、aavrh.50、aavrh.43、aavrh.62、aavrh.48、aavhu.19、aavhu.11、aavhu.53、aav4-8/rh.64、aavlg-9/hu.39、aav54.5/hu.23、aav54.2/hu.22、aav54.7/hu.24、aav54.1/hu.21、aav54.4r/hu.27、aav46.2/hu.28、aav46.6/hu.29、aav128.1/hu.43、真型aav(ttaav)、upennaav10、日本aav10血清型、aavcbr-7.1、aavcbr-7.10、aavcbr-7.2、aavcbr-7.3、aavcbr-7.4、aavcbr-7.5、aavcbr-7.7、aavcbr-7.8、aavcbr-b7.3、aavcbr-b7.4、aavcbr-e1、aavcbr-e2、aavcbr-e3、aavcbr-e4、aavcbr-e5、aavcbr-e5、aavcbr-e6、aavcbr-e7、aavcbr-e8、aavcht-1、aavcht-2、aavcht-3、aavcht-6.1、aavcht-6.10、aavcht-6.5、aavcht-6.6、aavcht-6.7、aavcht-6.8、aavcht-p1、aavcht-p2、aavcht-p5、aavcht-p6、aavcht-p8、aavcht-p9、aavckd-1、aavckd-10、aavckd-2、aavckd-3、aavckd-4、aavckd-6、aavckd-7、aavckd-8、aavckd-b1、aavckd-b2、aavckd-b3、aavckd-b4、aavckd-b5、aavckd-b6、aavckd-b7、aavckd-b8、aavckd-h1、aavckd-h2、aavckd-h3、aavckd-h4、aavckd-h5、aavckd-h6、aavckd-n3、aavckd-n4、aavckd-n9、aavclg-f1、aavclg-f2、aavclg-f3、aavclg-f4、aavclg-f5、aavclg-f6、aavclg-f7、aavclg-f8、aavclv-1、aavclv1-1、aavclv1-10、aavclv1-2、aavclv-12、aavclv1-3、aavclv-13、aavclv1-4、aavclv1-7、aavclv1-8、aavclv1-9、aavclv-2、aavclv-3、aavclv-4、aavclv-6、aavclv-8、aavclv-d1、aavclv-d2、aavclv-d3、aavclv-d4、aavclv-d5、aavclv-d6、aavclv-d7、aavclv-d8、aavclv-e1、aavclv-k1、aavclv-k3、aavclv-k6、aavclv-l4、aavclv-l5、aavclv-l6、aavclv-m1、aavclv-m11、aavclv-m2、aavclv-m5、aavclv-m6、aavclv-m7、aavclv-m8、aavclv-m9、aavclv-r1、aavclv-r2、aavclv-r3、aavclv-r4、aavclv-r5、aavclv-r6、aavclv-r7、aavclv-r8、aavclv-r9、aavcsp-1、aavcsp-10、aavcsp-11、aavcsp-2、aavcsp-3、aavcsp-4、aavcsp-6、aavcsp-7、aavcsp-8、aavcsp-8.10、aavcsp-8.2、aavcsp-8.4、aavcsp-8.5、aavcsp-8.6、aavcsp-8.7、aavcsp-8.8、aavcsp-8.9、aavcsp-9、aav.hu.48r3、aav.vr-355、aav3b、aav4、aav5、aavf1/hsc1、aavf11/hsc11、aavf12/hsc12、aavf13/hsc13、aavf14/hsc14、aavf15/hsc15、aavf16/hsc16、aavf17/hsc17、aavf2/hsc2、aavf3/hsc3、aavf4/hsc4、aavf5/hsc5、aavf6/hsc6、aavf7/hsc7、aavf8/hsc8、aavf9/hsc9、aavrh20、aavrh32/33、aavrh39、aavrh46、aavrh73、aavrh74、aavhu.26,或其变体或衍生物。
aav-dj序列可以包含两个突变:(1)r587q,其中氨基酸587处的精氨酸(r;arg)变为谷氨酰胺(q;gln)和(2)r590t,其中氨基酸590处的精氨酸(r;arg)变为苏氨酸(t;thr)。作为另一个非限制性实例,可以包含3个突变:(1)k406r,其中氨基酸406处的赖氨酸(k;lys)变为精氨酸(r;arg),(2)r587q,其中氨基酸587处的精氨酸(r;arg)变为谷氨酰胺(q;gln)和(3)r590t,其中氨基酸590处的精氨酸(r;arg)变为苏氨酸(t;thr)。
在某些实施方案中,aav可以是由aav9衣壳文库产生的具有390-627位氨基酸(vp1编号)中的突变的血清型。血清型以及相应的核苷酸和氨基酸取代可以是但不限于aav9.1(g1594c;d532h)、aav6.2(t1418a和t1436x;v473d和i479k)、aav9.3(t1238a;f413y)、aav9.4(t1250c和a1617t;f417s)、aav9.5(a1235g、a1314t、a1642g、c1760t;q412r、t548a、a587v)、aav9.6(t1231a;f411i)、aav9.9(g1203a、g1785t;w595c)、aav9.10(a1500g、t1676c;m559t)、aav9.11(a1425t、a1702c、a1769t;t568p、q590l)、aav9.13(a1369c、a1720t;n457h、t574s)、aav9.14(t1340a、t1362c、t1560c、g1713a;l447h)、aav9.16(a1775t;q592l)、aav9.24(t1507c、t1521g;w503r)、aav9.26(a1337g、a1769c;y446c、q590p)、aav9.33(a1667c;d556a)、aav9.34(a1534g、c1794t;n512d)、aav9.35(a1289t、t1450a、c1494t、a1515t、c1794a、g1816a;q430l、y484n、n98k、v606i)、aav9.40(a1694t、e565v)、aav9.41(a1348t、t1362c;t450s)、aav9.44(a1684c、a1701t、a1737g;n562h、k567n)、aav9.45(a1492t、c1804t;n498y、l602f)、aav9.46(g1441c、t1525c、t1549g;g481r、w509r、l517v)、9.47(g1241a、g1358a、a1669g、c1745t;s414n、g453d、k557e、t582i)、aav9.48(c1445t、a1736t;p482l、q579l)、aav9.50(a1638t、c1683t、t1805a;q546h、l602h)、aav9.53(g1301a、a1405c、c1664t、g1811t;r134q、s469r、a555v、g604v)、aav9.54(c1531a、t1609a;l511i、l537m)、aav9.55(t1605a;f535l)、aav9.58(c1475t、c1579a;t492i、h527n)、aav.59(t1336c;y446h)、aav9.61(a1493t;n498i)、aav9.64(c1531a、a1617t;l511i)、aav9.65(c1335t、t1530c、c1568a;a523d)、aav9.68(c1510a;p504t)、aav9.80(g1441a,;g481r)、aav9.83(c1402a、a1500t;p468t、e500d)、aav9.87(t1464c、t1468c;s490p)、aav9.90(a1196t;y399f)、aav9.91(t1316g、a1583t、c1782g、t1806c;l439r、k528i)、aav9.93(a1273g、a1421g、a1638c、c1712t、g1732a、a1744t、a1832t;s425g、q474r、q546h、p571l、g578r、t582s、d611v)、aav9.94(a1675t;m559l)和aav9.95(t1605a;f535l)。
在本文引用和/或描述的任何dna和rna序列中,单个字母符号具有以下描述:a代表腺嘌呤;c代表胞嘧啶;g代表鸟嘌呤;t代表胸腺嘧啶;u代表尿嘧啶;w代表弱碱基,例如腺嘌呤或胸腺嘧啶;s代表强核苷酸,例如胞嘧啶和鸟嘌呤;m代表氨基核苷酸,如腺嘌呤和胞嘧啶;k代表酮核苷酸,如鸟嘌呤和胸腺嘧啶;r代表嘌呤腺嘌呤和鸟嘌呤;y代表嘧啶胞嘧啶和胸腺嘧啶;b代表任何不是a的碱基(例如胞嘧啶、鸟嘌呤和胸腺嘧啶);d代表任何不是c的碱基(例如,腺嘌呤、鸟嘌呤和胸腺嘧啶);h代表任何不是g的碱基(例如,腺嘌呤、胞嘧啶和胸腺嘧啶);v代表任何不是t的碱基(例如,腺嘌呤、胞嘧啶和鸟嘌呤);n代表任何核苷酸(其不是空位);并且z代表零。
在本文提及和/或描述的任何氨基酸序列中,单字母符号具有以下描述:g(gly)代表甘氨酸;a(ala)代表丙氨酸;l(leu)代表亮氨酸;m(met)代表甲硫氨酸;f(phe)代表苯丙氨酸;w(trp)代表色氨酸;k(lys)代表赖氨酸;q(gln)代表谷氨酰胺;e(glu)代表谷氨酸;s(ser)代表丝氨酸;p(pro)代表脯氨酸;v(val)代表缬氨酸;i(ile)代表异亮氨酸;c(cys)代表半胱氨酸;y(tyr)代表酪氨酸;h(his)代表组氨酸;r(arg)代表精氨酸;n(asn)代表天冬酰胺;d(asp)代表天冬氨酸;t(thr)代表苏氨酸;b(asx)代表天冬氨酸或天冬酰胺;j(xle)代表亮氨酸或异亮氨酸;o(pyl)为吡咯赖氨酸;u(sec)代表硒代胱氨酸;x(xaa)代表任何氨基酸;z(glx)代表谷氨酰胺或谷氨酸。
在某些实施方案中,aav血清型可以是或可以包括如以下专利公开中所述的序列、插入、修饰或突变:wo2015038958、wo2017100671、wo2016134375、wo2017083722、wo2017015102、wo2017058892、wo2017066764、us9624274、us9475845、us20160369298、us20170145405,由于涉及aav血清型和修饰,其内容通过引用整体并入本文。
在某些实施方案中,aav可以是如deverman等人(naturebiotechnology34(2):204-209(2016))所述的基于cre重组的aav靶向进化(create)生成的血清型,其内容通过引用整体并入本文。在某些实施方案中,aav血清型可以如jackson等人(frontiersinmolecularneuroscience9:154(2016))所述,由于涉及aav血清型和修饰,该文献的内容通过引用整体并入本文。在某些实施方案中,选择aav血清型进行使用是由于其对中枢神经系统细胞的嗜性。在某些实施方案中,中枢神经系统的细胞是神经元。在另一个实施方案中,中枢神经系统的细胞是星形胶质细胞。
在某些实施方案中,选择aav血清型进行使用是由于其对肌肉细胞的嗜性。
在某些实施方案中,用于aavvp1衣壳蛋白翻译的起始密码子可以是ctg、ttg或gtg,如美国专利号us8163543中所述,由于涉及aav血清型和修饰,其内容通过引用整体并入本文。
本公开涉及由衣壳(cap)基因编码的结构衣壳蛋白(包括vp1、vp2和vp3)。这些衣壳蛋白形成病毒载体例如aav的外部蛋白结构壳(即衣壳)。由cap多核苷酸合成的vp衣壳蛋白通常包含甲硫氨酸作为肽序列中的第一个氨基酸(met1),其与相应的cap核苷酸序列中的起始密码子(aug或atg)相关。然而,常见的是在蛋白合成之后或期间通过蛋白加工酶例如met-氨基肽酶将第一甲硫氨酸(met1)残基或通常任何第一氨基酸(aa1)裂解掉。这种“met/aa-剪切”过程通常与多肽序列中的第二个氨基酸(例如,丙氨酸、缬氨酸、丝氨酸、苏氨酸等)的相应乙酰化相关。met-剪切通常发生在vp1和vp3衣壳蛋白上,但也可发生在vp2衣壳蛋白上。
当met/aa剪切不完全时,可能产生包含病毒衣壳在内的一种或多种(一个、两个或三个)vp衣壳蛋白的混合物,其中一些可能包含met1/aa1氨基酸(met+/aa+),并且其中一些由于met/aa剪切可能缺少met1/aa1氨基酸(met-/aa-)。有关衣壳蛋白中met/aa剪切的更多讨论,参见以下文献:jin等人,directliquidchromatography/massspectrometryanalysisforcompletecharacterizationofrecombinantadeno-associatedviruscapsidproteins.humgenethermethods.2017oct.28(5):255-267;hwang等人,n-terminalacetylationofcellularproteinscreatesspecificdegradationsignals.science.2010february19.327(5968):973–977;由于涉及壳蛋白修饰和剪切,其内容各自通过引用整体并入本文。
根据本公开,对衣壳蛋白的提及不限于剪切的(met-/aa-)或未剪切的(met+/aa+),并且在上下文中可以是指独立的衣壳蛋白、包含衣壳蛋白混合物的病毒衣壳和/或编码、描述、产生或导致本公开的衣壳蛋白的多核苷酸序列(或其片段)。直接提及“衣壳蛋白”或“衣壳多肽”(例如vp1、vp2或vp2)还可以包括:包含met1/aa1氨基酸(met+/aa+)的vp衣壳蛋白以及相应的由于met/aa剪切而缺少met1/aa1氨基酸(met-/aa-)的vp衣壳蛋白。
进一步根据本公开,提及分别包含或编码一种或多种包含met1/aa1氨基酸的衣壳蛋白(met+/aa+)的特定seqidno:(无论是蛋白还是核酸)应被理解为教导了检查序列时缺少met1/aa1氨基酸的vp衣壳蛋白,很明显其是任何仅缺少第一列出的氨基酸的序列(无论是否是met1/aa1)。
作为非限制性实例,提及长度为736个氨基酸且包括由aug/atg起始密码子编码的“met1”氨基酸(met+)的vp1多肽序列也可以被理解为教导了长度为735个氨基酸且不包含736个氨基酸met+序列中的“met1”氨基酸(met-)的vp1多肽序列。作为第二非限制性实例,提及的长度为736个氨基酸且包含由任何nnn起始密码子编码的“aa1”氨基酸(aa1+)的vp1多肽序列也可以被理解为教导了长度为735个氨基酸且不包含736个氨基酸aa1+序列中的“aa1”氨基酸(aa1-)的vp1多肽序列。
提及由vp衣壳蛋白形成的病毒衣壳(例如提及特定的aav衣壳血清型)可以并入包含met1/aa1氨基酸的vp衣壳蛋白(met+/aa1+)、相应的由于met/aa1剪切而缺少met1/aa1氨基酸的vp衣壳蛋白(met-/aa1-)及其组合(met+/aa1+和met-/aa1-)。
作为非限制性实例,aav衣壳血清型可包括vp1(met+/aa1+)、vp1(met-/aa1-)或vp1(met+/aa1+)和vp1(met-/aa1-)的组合。aav衣壳血清型还可以包括vp3(met+/aa1+)、vp3(met-/aa1-)或vp3(met+/aa1+)和vp3(met-/aa1-)的组合;并且还可以包括vp2(met+/aa1+)和vp2(met-/aa1-)的类似可选组合。
引入细胞
本公开的编码的sirna分子(例如,sirna双链体)可以通过被aav颗粒的载体基因组编码而引入到细胞中。对这些aav颗粒进行了工程化和优化,以帮助其进入到不易被转染/转导修饰的细胞。而且,一些合成的病毒载体具有将shrna整合到细胞基因组中的能力,从而导致稳定的sirna表达和靶基因的长期敲低。以这种方式,将病毒载体工程化为用于特定递送的载体,同时缺乏在野生型病毒中发现的有害复制和/或整合特征。
在某些实施方案中,通过用aav颗粒转染、感染或转导细胞,将编码的sirna分子引入到细胞中,所述aav颗粒包含当在细胞中转录时能够产生sirna分子的核酸序列。在某些实施方案中,通过将aav颗粒注射到细胞或组织中来将sirna分子引入到细胞中,所述aav颗粒包含当在细胞中转录时能够产生sirna分子的核酸序列。
在某些实施方案中,在转染/转导之前,可以将包含编码本公开的sirna分子的核酸序列的aav颗粒转染到细胞中。
引入包含本文所述的sirna分子的核酸序列的aav颗粒的其他方法可以包括如美国专利公开号20120264807中所述的光化学内在化,由于涉及光化学内在化,该专利的内容通过引用整体并入本文。
在某些实施方案中,本文所述的制剂可以包含至少一个aav颗粒,其包含编码本文所述的sirna分子的核酸序列。在某些实施方案中,sirna分子可以在一个靶位点靶向目标基因。在另一个实施方案中,制剂包含多个aav颗粒,每个aav颗粒均包含编码在不同靶位点靶向目标基因的sirna分子的核酸序列。可以在2、3、4、5或超过5个位点靶向目标基因。
在某些实施方案中,可以将来自任何相关物种(例如但不限于人、狗、猪、小鼠、大鼠或猴)的aav颗粒引入到细胞中。
在某些实施方案中,可以将配制的aav颗粒引入到与待治疗的疾病相关的细胞或组织中。
在某些实施方案中,可以将配制的aav颗粒引入到具有高水平靶序列内源性表达的细胞中。
在另一个实施方案中,可以将配制的aav颗粒引入到具有低水平靶序列内源性表达的细胞中。
在某些实施方案中,细胞可以是具有高aav转导效率的细胞。
在某些实施方案中,配制的包含编码本公开的sirna分子的核酸序列的aav颗粒可以用于将sirna分子递送至中枢神经系统(例如,美国专利号6,180,613;由于涉及sirna分子和aav颗粒的递送和治疗用途,其内容通过引用整体并入本文)。
在某些实施方案中,配制的包含编码本公开的sirna分子的核酸序列的aav颗粒可以进一步包含修饰的衣壳,其包含来自非病毒来源的肽。在其他方面,aav颗粒可以包含cns特异性嵌合衣壳,以促进将编码的sirna双链体递送至脑和脊髓。例如,可以构建来自显示cns嗜性的aav变体的cap核苷酸序列的比对,以识别可变区(vr)序列和结构。
在某些实施方案中,配制的包含编码本公开的sirna分子的核酸序列的aav颗粒可以编码作为多顺反子分子的sirna分子。sirna分子可以在sirna分子的区域之间另外包含一个或多个接头。
在某些实施方案中,配制的aav颗粒可以包含至少一个编码本文所述的sirna序列或双链体中的至少一个的调节性多核苷酸。
在某些实施方案中,表达载体从itr至itr可以包含(从5’至3’陈述)itr、启动子、内含子、调节性多核苷酸、polya序列和itr。
在某些实施方案中,编码的sirna分子在表达载体中可位于启动子的下游,该启动子例如但不限于cmv、u6、h1、cba或具有sv40内含子的cba启动子。此外,编码的sirna分子在表达载体中还可位于聚腺苷酸化序列的上游。作为非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或多于30个氨基酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的1-5、1-10、1-15、1-20、1-25、1-30、5-10、5-15、5-20、5-25、5-30、10-15、10-20、10-25、10-30、15-20、15-25、15-30、20-25、20-30或25-30个核苷酸内。作为非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的前1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%或多于25%的核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的前1-5%、1-10%、1-15%、1-20%、1-25%、5-10%、5-15%、5-20%、5-25%、10-15%、10-20%、10-25%、15-20%、15-25%或20-25%的核苷酸内。
在某些实施方案中,编码的sirna分子在表达载体中可位于聚腺苷酸化序列的上游。此外,编码的sirna分子在表达载体中可位于启动子的下游,该启动子例如但不限于cmv、u6、cba或具有sv40内含子的cba启动子。作为非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或多于30个核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的1-5、1-10、1-15、1-20、1-25、1-30、5-10、5-15、5-20、5-25、5-30、10-15、10-20、10-25、10-30、15-20、15-25、15-30、20-25、20-30或25-30个核苷酸内。作为非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的前1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%或多于25%的核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可位于启动子下游和/或聚腺苷酸化序列上游的前1-5%、1-10%、1-15%、1-20%、1-25%、5-10%、5-15%、5-20%、5-25%、10-15%、10-20%、10-25%、15-20%、15-25%或20-25%的核苷酸内。
在某些实施方案中,编码的sirna分子可以位于scaav中。
在某些实施方案中,编码的sirna分子可以位于ssaav中。
在某些实施方案中,编码的sirna分子在表达载体中可以位于flipitr的5’端附近。在另一个实施方案中,编码的sirna分子在表达载体中可以位于flipitr的3’端附近。在又一个实施方案中,编码的sirna分子在表达载体中可以位于flopitr的5’端附近。在又一个实施方案中,编码的sirna分子在表达载体中可以位于flopitr的3’端附近。在某些实施方案中,编码的sirna分子在表达载体中可以位于flipitr的5’端和flopitr的3’端之间。在某些实施方案中,编码的sirna分子在表达载体中可以位于flipitr的3’端和flopitr的5’端之间(例如,在flipitr的5’端与flopitr的3’端或在flopitr的3’端和flipitr的5’端之间的中间)。作为非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端下游的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或多于30个核苷酸内。作为非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端上游的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30或多于30个核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端下游的1-5、1-10、1-15、1-20、1-25、1-30、5-10、5-15、5-20、5-25、5-30、10-15、10-20、10-25、10-30、15-20、15-25、15-30、20-25、20-30或25-30个核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端上游的1-5、1-10、1-15、1-20、1-25、1-30、5-10、5-15、5-20、5-25、5-30、10-15、10-20、10-25、10-30、15-20、15-25、15-30、20-25、20-30或25-30个核苷酸内。作为非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端上游的前1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%或大于25%的核苷酸内。作为另一个非限制性实例,编码的sirna分子在表达载体中可以位于itr(例如flip或flopitr)的5’或3’端下游的前1-5%、1-10%、1-15%、1-20%、1-25%、5-10%、5-15%、5-20%、5-25%、10-15%、10-20%、10-25%、15-20%、15-25%或20-25%内。
在某些实施方案中,可以配制包含本公开的sirna分子的核酸序列的aav颗粒用于cns递送。可以使用穿过大脑血液屏障的药物。例如,可以将sirna分子靶向脑血屏障内皮的一些细胞穿透肽可用于配制靶向目标基因的sirna双链体。
在某些实施方案中,可以将配制的包含编码本公开的sirna分子的核酸序列的aav颗粒直接施用于cns。作为非限制性实例,载体包含编码靶向目标基因的sirna分子的核酸序列。
在特定实施方案中,可以以促进载体或sirna分子进入中枢神经系统并渗透进运动神经元的方式施用配制的包含编码本公开的sirna分子的核酸序列的aav颗粒的组合物。
在某些实施方案中,可以以治疗有效量将配制的aav颗粒施用于受试者(例如,通过鞘内施用施用于受试者的cns),以使sirna双链体或dsrna靶向脊髓和/或脑干中的运动神经元和星形胶质细胞。作为非限制性实例,sirna双链体或dsrna可减少蛋白或mrna的表达。
ii.aav生产
通用病毒生产方法
用于生产raav颗粒的病毒生产细胞通常包括哺乳动物细胞类型。然而,哺乳动物细胞给raav颗粒的大规模生产带来了一些问题,包括每个复制细胞的病毒颗粒的普遍低产率,以及病毒生产细胞中存在不希望的来自其他哺乳动物生物材料的污染的高风险。因此,昆虫细胞已经成为大规模生产raav颗粒的替代载体。
使用昆虫细胞的aav生产系统也存在一系列问题。例如,高产量生产raav颗粒通常要求与rep52相比更低的rep78的表达。因此,控制rep78和rep52在昆虫细胞中的相对表达要求在rep操纵子内精心设计的控制机制。这些控制机制可包括单独工程化的昆虫细胞启动子,例如用于rep78的δie1启动子和用于rep52的polh启动子,或将编码rep的核苷酸序列划分为独立工程化的序列或构建体。但是,实施这些控制机制通常导致raav颗粒产量降低或结构不稳定的病毒体。
在另一个实例中,raav颗粒的生产需要vp1、vp2和vp3蛋白,它们组装以形成aav衣壳。raav颗粒的高产量生产需要调节的vp1、vp2和vp3比例,其通常应分别为1:1:10左右,但相对于10个vp3拷贝,vp1可以为1-2,和/或vp2可以为1-2。该比例对于衣壳的质量很重要,因为太多的vp1使破坏衣壳的稳定性,而太少的vp1将降低病毒的感染性。
野生型aav使用缺陷剪接方法来控制vp1的表达;使用具有特殊周围环境(“kozak”序列)的弱起始密码子(acg)来控制vp2;以及对于vp3的表达使用标准起始密码子(atg)。但是,在一些杆状病毒系统中,哺乳动物剪接序列并不总是被识别,并且不能适当地控制vp1、vp2和vp3的产生。因此,来自vp2的相邻核苷酸和acg起始序列可用于驱动衣壳蛋白的产生。不幸的是,对于大多数aav血清型,这种方法产生的衣壳具有较低的vp1比vp2的比(相对于10个vp3拷贝,<1)。为了更有效地控制vp蛋白的生产,已使用了非经典或起始密码子,例如ttg、gtg或ctg。然而,相对于野生型atg或acg起始密码子,这些起始密码子被本领域技术人员认为是次优的(参见wo2007046703和wo2007148971,由于涉及aav衣壳蛋白的生产,其内容通过引用整体并入本文)。
在另一个实例中,使用杆状病毒/sf9系统生产raav颗粒通常需要广泛使用的基于杆粒的杆状病毒表达载体系统(bev),该系统未针对大规模aav生产进行优化。在基于杆粒的bev中病毒蛋白的异常蛋白水解降解是一个出于意料的问题,这妨碍了使用杆状病毒/sf9系统可靠地大规模生产aav衣壳蛋白的过程。
仍然需要允许在哺乳动物和昆虫细胞中有效且高效地大规模(商业)生产raav颗粒的方法和系统。
在以下描述中阐述了本公开的一个或多个实施方案的细节。根据说明书、附图和权利要求书,本发明的其他特征、目的和优点将变得显而易见。在说明书中,单数形式也包括复数,除非上下文另外明确指出。除非另有定义,否则本文使用的所有技术和科学术语具有与本公开所属领域的普通技术人员通常所理解的相同含义。在与通过引用并入的公开内容有冲突的情况下,以本文的明确描述为准。
在某些实施方案中,本公开的构建体、多核苷酸、多肽、载体、血清型、衣壳制剂或颗粒可以是、可以包括以下国际公开之一中所述的任何序列、元件、构建体、系统、靶标或方法,可以被以下国际公布之一所述的任何序列、元件、构建体、系统、靶标或方法修饰、使用,可以被以下国际公布之一所述的任何序列、元件、构建体、系统、靶标或方法使用,可以用于以下国际公布之一所述的任何序列、元件、构建体、系统、靶标或方法,可以与以下国际公布之一所述的任何序列、元件、构建体、系统、靶标或方法一起使用,或可以用以下国际公布之一所述的任何序列、元件、构建体、系统、靶标或方法生产:wo2016073693、wo2017023724、wo2018232055、wo2016077687、wo2016077689、wo2018204786、wo2017201258、wo2017201248、wo2018204803、wo2018204797、wo2017189959、wo2017189963、wo2017189964、wo2015191508、wo2016094783、wo20160137949、wo2017075335,;其内容均通过引用整体并入本文,只要其不与本公开冲突。
本公开的aav生产包括生产aav颗粒和病毒载体的过程和方法,所述aav颗粒和病毒载体可接触靶细胞以递送有效载荷,例如重组病毒构建体,其包含编码有效载荷分子的核苷酸。在某些实施方案中,病毒载体是腺相关病毒(aav)载体,例如重组腺相关病毒(raav)载体。在某些实施方案中,aav颗粒是腺相关病毒(aav)颗粒,例如重组腺相关病毒(raav)颗粒。
本公开提供了通过以下方式生产aav颗粒或病毒载体的方法:(a)使病毒生产细胞与编码至少一种嵌合衣壳蛋白的一种或多种病毒表达构建体以及一种或多种有效载荷构建体载体接触,其中所述有效载荷构建体载体包括编码有效载荷分子的有效载荷构建体,所述有效载荷分子选自转基因、编码蛋白的多核苷酸和调节性核酸;(b)在能够生产至少一种aav颗粒或病毒载体的条件下培养所述病毒生产细胞,和(c)分离所述至少一种aav颗粒或病毒载体。
在这些方法中,病毒表达构建体可以编码至少一种结构蛋白和/或至少一种非结构蛋白。结构蛋白可以包括任何天然或野生型衣壳蛋白vp1、vp2和/或vp3或嵌合蛋白。非结构蛋白可以包括任何天然或野生型rep78、rep68、rep52和/或rep40蛋白或嵌合蛋白。
在某些实施方案中,接触通过瞬时转染、病毒转导和/或电穿孔发生。
在某些实施方案中,病毒生产细胞选自哺乳动物细胞和昆虫细胞。在某些实施方案中,昆虫细胞包括草地贪夜蛾(spodopterafrugiperda)昆虫细胞。在某些实施方案中,昆虫细胞包括sf9昆虫细胞。在某些实施方案中,昆虫细胞包括sf21昆虫细胞。
本公开的有效载荷构建体载体可以包含至少一个反向末端重复序列(itr),并且可以包含哺乳动物dna。
还提供了根据本文描述的方法生产的aav颗粒和病毒载体。
本公开的aav颗粒可以被配制为具有一种或多种可接受的赋形剂的药物组合物。
在某些实施方案中,可以通过本文描述的方法生产aav颗粒或病毒载体。
在某些实施方案中,可以通过使病毒生产细胞(例如昆虫细胞或哺乳动物细胞)与编码至少一种衣壳蛋白的至少一种病毒表达构建体和至少一种有效载荷构建体载体接触来生产aav颗粒。病毒生产细胞可以通过瞬时转染、病毒转导和/或电穿孔来接触。有效载荷构建体载体可以包括编码有效载荷分子的有效载荷构建体,所述有效载荷分子例如但不限于转基因、编码蛋白的多核苷酸和调节性核酸。在生产、分离(例如,使用温度诱导的裂解、机械裂解和/或化学裂解)和/或纯化(例如,使用过滤、色谱法和/或免疫亲和纯化)至少一种aav颗粒或病毒载体的条件下培养所述病毒生产细胞。作为非限制性实例,有效载荷构建体载体可以包含哺乳动物dna。
在某些实施方案中,使用本文所述的方法在昆虫细胞(例如,草地贪夜蛾(sf9)细胞)中生产aav颗粒。作为非限制性实例,使用病毒转导接触昆虫细胞,所述病毒转导可以包括杆状病毒转导。
在另一个实施方案中,使用本文描述的方法在哺乳动物细胞中生产aav颗粒。作为非限制性实例,使用瞬时转染接触哺乳动物细胞。
在某些实施方案中,病毒表达构建体可以编码至少一种结构蛋白和至少一种非结构蛋白。作为非限制性实例,结构蛋白包括vp1、vp2和/或vp3。作为另一个非限制性实例,非结构蛋白包括rep78、rep68、rep52和/或rep40。
在某些实施方案中,本文所述的aav颗粒生产方法在病毒生产细胞中生产大于101、大于102、大于103、大于104或大于105个aav颗粒。
在某些实施方案中,本公开的方法包括使用病毒生产系统在病毒生产细胞中生产病毒颗粒,所述病毒生产系统包括至少一种病毒表达构建体和至少一种有效载荷构建体。可以将至少一种病毒表达构建体和至少一种有效载荷构建体共转染(例如双重转染、三重转染)到病毒生产细胞中。使用本领域技术人员已知和常规进行的标准分子生物学技术完成转染。病毒生产细胞提供表达生产aav颗粒所必需的蛋白和其他生物材料所必需的细胞机制,包括复制有效载荷构建体的rep蛋白和组装形成包围复制的有效载荷构建体的衣壳的cap蛋白。从病毒生产细胞中提取所得的aav颗粒,并将其加工成用于施用的药物制剂。
一旦施用,aav颗粒接触靶细胞并以内含体进入细胞。aav颗粒从内含体释放,然后接触靶细胞的核以递送有效载荷构建体。有效载荷构建体(例如重组病毒构建体)被递送至靶细胞的核,其中可以表达由有效载荷构建体编码的有效载荷分子。
在某些实施方案中,生产病毒颗粒的方法利用病毒生产细胞的种子培养物,其包括一种或多种杆状病毒(例如,杆状病毒表达载体(bev)或已经被病毒表达构建体和有效载荷构建体载体转染的杆状病毒感染的昆虫细胞(biic))。在某些实施方案中,收获种子培养物,将其分成等分试样并冷冻,并可以在稍后的时间点用于引发生产细胞未经处理群体的感染。
aav颗粒的大规模生产可以利用生物反应器。使用生物反应器允许精确测量和/或控制支持病毒生产细胞的生长和活性的变量,例如质量、温度、混合条件(喷射器rpm或波振荡)、co2浓度、o2浓度、气体喷射速率和体积、气体覆盖率和体积、ph、活细胞密度(vcd)、细胞活力、细胞直径和/或光密度(od)。在某些实施方案中,生物反应器用于批量生产,其中在实验确定的时间点收获整个培养物,并纯化aav颗粒。在另一个实施方案中,使用生物反应器进行连续生产,其中在实验确定的时间点收获一部分培养物以纯化aav颗粒,并且用另外的生长培养基组分更新生物反应器中的剩余培养物。
可以在包括细胞裂解、澄清、灭菌和纯化的过程中从病毒生产细胞中提取aav病毒颗粒。细胞裂解包括破坏病毒生产细胞的结构从而释放aav颗粒的任何过程。在某些实施方案中,细胞裂解可以包含热激、化学或机械裂解方法。澄清可以包括裂解细胞、培养基组分和aav颗粒的混合物的粗纯化。在某些实施方案中,澄清包括离心和/或过滤,包括但不限于深度末端过滤、切向流过滤和/或中空纤维过滤。
病毒生产的最终结果是纯化的aav颗粒收集物,其中包括两种组分:(1)有效载荷构建体(例如重组病毒基因组构建体)和(2)病毒衣壳。
在某些实施方案中,本公开的病毒生产系统或方法包括使用病毒生产细胞(vpc)和质粒构建体生产杆状病毒感染的昆虫细胞(biic)的步骤。将来自细胞库(cb)的病毒生产细胞(vpc)解冻并扩增,以提供目标工作体积和vpc浓度。将得到的vpc汇聚物分成rep/capvpc汇聚物和有效载荷vpc汇聚物。将一种或多种rep/cap质粒构建体(病毒表达构建体)加工成rep/cap杆粒多核苷酸,并转染到rep/capvpc汇聚物中。将一种或多种有效载荷质粒构建体(有效载荷构建体)加工成有效载荷杆粒多核苷酸并转染到有效载荷vpc汇聚物中。将两个vpc汇聚物温育以生产p1rep/cap杆状病毒表达载体(bev)和p1有效载荷bev。将两个bev汇聚物扩增为斑块收集物,并选择一个斑块进行克隆斑块(cp)纯化(也称为单一斑块扩增)。该过程可以包括单个cp纯化步骤,或者可以包括串联的或被其他处理步骤分开的多个cp纯化步骤。一个或多个cp纯化步骤提供cprep/capbev汇聚物和一个cp有效载荷bev汇聚物。然后可以储存这两个bev汇聚物并将其用于将来的生产步骤,或者可以将它们转染到vpc中以生产rep/capbiic汇聚物和有效载荷biic汇聚物。
在某些实施方案中,本公开的病毒生产系统或方法包括使用病毒生产细胞(vpc)和杆状病毒感染的昆虫细胞(biic)来生产aav颗粒的步骤。将来自细胞库(cb)的病毒生产细胞(vpc)解冻并扩增,以提供目标工作体积和vpc浓度。将工作体积的病毒生产细胞接种到生产生物反应器中,并可以进一步扩增至200-2000l的工作体积及以及用于biic感染的目标vpc浓度。然后,将生产生物反应器中的工作体积的vpc与rep/capbiic和有效载荷biic,以目标vpc:biic比例和目标biic:biic比例共感染。vcd感染也可以利用bev。在生产生物反应器中温育并扩增共感染的vpc,以生产aav颗粒和vpc的大量收获物。
病毒表达构建体
本公开的病毒生产系统包括一种或多种可以被转染/转导到病毒生产细胞中的病毒表达构建体。在某些实施方案中,本公开的病毒表达构建体或有效载荷构建体可以是杆粒,也被称为杆状病毒质粒或重组杆状病毒基因组。在某些实施方案中,病毒表达包括编码蛋白的核苷酸序列和用于在病毒生产细胞中表达的至少一个表达控制序列。在某些实施方案中,病毒表达包括有效地连接至至少一个用于在病毒生产细胞中表达的表达控制序列的编码蛋白的核苷酸序列。在某些实施方案中,病毒表达构建体包含在一个或多个启动子控制下的细小病毒基因。细小病毒基因可以包括编码非结构性aav复制蛋白的核苷酸序列,例如编码rep52、rep40、rep68或rep78蛋白的rep基因。细小病毒基因可以包括编码结构性aav蛋白的核苷酸序列,例如编码vp1、vp2和vp3蛋白的cap基因。
在某些实施方案中,病毒表达构建体可以包含rep52编码区;rep52编码区是包含编码rep52蛋白的rep52核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含rep78编码区;rep78编码区是包含编码rep78蛋白的rep78核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含rep40编码区;rep40编码区是包含编码rep40蛋白的rep40核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含rep68编码区;rep68编码区是包含编码rep68蛋白的rep68核苷酸序列的核苷酸序列。
在某些实施方案中,病毒表达构建体可以包含vp编码区;vp编码区是包含编码vp1、vp2、vp3或其组合的vp核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含vp1编码区;vp1编码区是包含编码vp1蛋白的vp1核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含vp2编码区;vp2编码区是包含编码vp2蛋白的vp2核苷酸序列的核苷酸序列。在某些实施方案中,病毒表达构建体可以包含vp3编码区;vp3编码区是包含编码vp3蛋白的vp3核苷酸序列的核苷酸序列。
病毒表达构建体的结构性vp蛋白vp1、vp2和vp3以及非结构性蛋白rep52和rep78可以在通过利用可变剪接受体和非规范翻译起始密码子二者调节的单个开放阅读框中编码。rep78和rep52均可从单个转录物进行翻译:rep78翻译在第一个起始密码子(aug或非aug)处启动,而rep52翻译从rep78序列内的rep52起始密码子(例如aug)启动。rep78和rep52也可以从具有独立起始密码子的单独的转录物进行翻译。可以对rep78序列内的rep52起始密码子进行突变、修饰或去除,以使对修饰的rep78序列进行的加工将不生产rep52蛋白。
可以从单个转录物转录和翻译vp1、vp2和vp3,其中,对框内和/或框外起始密码子都进行工程化以控制由核苷酸转录物产生的vp1:vp2:vp3比例。在某些实施方案中,vp1可以由仅对vp1进行编码的序列产生。如本文所用,术语“仅对vp1”或“仅vp1”是指核苷酸序列或转录物,其编码vp1衣壳蛋白并且:(i)在vp1序列内缺少用于从同一序列完全转录或翻译vp2和vp3的必需起始密码子(即缺失或突变);(ii)在vp1序列内包括防止从同一序列转录或翻译vp2和vp3的其他密码子;或(iii)包含vp1的起始密码子(例如atg),使得vp1是核苷酸转录物产生的主要vp蛋白。
在某些实施方案中,vp2可以由仅对vp2进行编码的序列生产。如本文所用,术语“仅对vp2”或“仅vp2”是指编码vp2衣壳蛋白的核苷酸序列或转录物,并且:(i)该核苷酸转录物是编码仅vp2和vp3衣壳蛋白的完整vp衣壳序列的截短变体,和(ii)其包含vp2的起始密码子(例如atg),使得vp2是该核苷酸转录物产生的主要vp蛋白。
在某些实施方案中,vp1和vp2可以由仅对vp1和vp2进行编码的序列生产。如本文所用,术语“仅对vp1和vp2”或“仅vp1和vp2”是指编码vp1和vp2衣壳蛋白的核苷酸序列或转录物,并且其:(i)在vp序列内缺少用于从同一序列完全转录或翻译vp3的必需起始密码子(即删除或突变);(ii)在vp序列内包含防止从同一序列转录或翻译vp3的其他密码子;(iii)包含vp1的起始密码子(例如atg)和vp2的起始密码子(例如atg),使得vp1和vp2是该核苷酸转录物生产的主要vp蛋白;或(iv)包含通过接头(例如ires区域)连接的仅vp1的核苷酸转录物和仅vp2的核苷酸转录物。
本公开的病毒生产系统不受用于将细小病毒功能引入到病毒复制细胞中的病毒表达载体的限制。病毒复制细胞中病毒表达构建体的存在不需是永久性的。可以通过任何已知的方式引入病毒表达构建体,例如通过细胞的化学处理、电穿孔或感染。
本公开的病毒表达构建体可以包括任何化合物或制剂(生物的或化学的),其有助于用核酸转化、转染或转导细胞。示例性的生物病毒表达构建体包括质粒、线性核酸分子和包括杆状病毒的重组病毒。示例性的化学载体包括脂质复合物。根据本公开,病毒表达构建体用于将核酸序列引入到病毒复制细胞中。(o'reilly,davidr.,loisk.miller和vernea.luckow.baculovirusexpressionvectors:alaboratorymanual.oxforduniversitypress,1994.);maniatis等人,eds.molecularcloning.cshlaboratory,ny,n.y.(1982);和philiport和scluber,eds.liposoesastoolsinbasicresearchandindustry.crcpress,annarbor,mich.(1995),由于涉及病毒表达构建体及其用途,其各自内容通过引用整体并入本文。
在某些实施方案中,病毒表达构建体是aav表达构建体,其包含一个或多个编码非结构性aav复制蛋白、结构性aav衣壳蛋白或其组合的核苷酸序列。
在某些实施方案中,本公开的病毒表达构建体可以是质粒载体。在某些实施方案中,本公开的病毒表达构建体可以是杆状病毒构建体。
本公开不受用于生产aav颗粒或病毒载体的病毒表达构建体的数量的限制。在某些实施方案中,根据本公开,可以采用一种、两种、三种、四种、五种、六种或更多种病毒表达构建体在病毒生产细胞中生产aav颗粒。在一个非限制性实例中,五种表达构建体可以分别编码aavvp1、aavvp2、aavvp3、rep52、rep78,以及伴随的包含有效载荷多核苷酸和至少一个aavitr的有效载荷构建体。在另一个实施方案中,表达构建体可以用于表达例如rep52和rep40,或rep78和rep68。表达构建体可以包含vp1、vp2、vp3、rep52/rep40和rep78/rep68编码序列的任何组合。
在本公开的某些实施方案中,病毒表达构建体可以用于在昆虫细胞中生产aav颗粒。在某些实施方案中,可以对衣壳和/或rep基因的野生型aav序列进行修饰,例如以改善病毒颗粒的特性,例如增加的感染性或特异性,或提高产产量。
在某些实施方案中,病毒表达构建体可在蛋白编码核苷酸序列之间包含一个或多个表达控制序列。在某些实施方案中,表达控制区可包含ires序列区,其包含编码内部核糖体进入位点(ires)的ires核苷酸序列。内部核糖体进入位点(ires)可以选自:来自口蹄疫病毒的fmdv-ires、来自脑心肌炎病毒的emcv-ires及其组合。
在某些实施方案中,表达控制区可包含2a序列区,其包含编码病毒2a肽的2a核苷酸序列。该序列允许在单个开放阅读框(orf)内共翻译多个多肽。随着orf的翻译,带有2a序列的甘氨酸和脯氨酸残基阻止正常肽键的形成,这导致多肽链内的核糖体“跳跃(skipping)”和“自我切割”。病毒2a肽可以选自:来自口蹄疫病毒的f2a、来自明脉扁刺蛾(thoseaasigna)病毒的t2a、来自马甲型鼻炎(equinerhinitisa)病毒的e2a、来自猪捷申病毒1(porcineteschovirus-1)的p2a、来自细胞质多角体病病毒的bmcpv2a、来自家蚕(b.mori)软化病病毒的bmifv2a,及其组合。
在某些实施方案中,病毒表达构建体可以包含含有起始密码子区的核苷酸序列,例如编码包含一个或多个起始密码子区的aav衣壳蛋白的序列。在某些实施方案中,起始密码子区可以在表达控制序列内。起始密码子可以是atg或非atg密码子(即次佳起始密码子,其中aavvp1衣壳蛋白的起始密码子是非atg)。
在某些实施方案中,用于aav生产的病毒表达构建体可以包含编码aav衣壳蛋白的核苷酸序列,其中aavvp1衣壳蛋白的起始密码子是非atg,即次佳的起始密码子,从而允许以修饰比例在生产系统中表达病毒衣壳蛋白,以提供改善的宿主细胞的感染性。在非限制性实例中,病毒构建体载体可以包含核酸构建体,该核酸构建体包含编码aavvp1、vp2和vp3衣壳蛋白的核苷酸序列,其中用于翻译aavvp1衣壳蛋白的起始密码子是ctg、ttg或gtg,如美国专利号us8,163,543中所述的,由于涉及aav衣壳蛋白及其生产,其内容通过引用整体并入本文。
在某些实施方案中,本公开的病毒表达构建体可以是质粒载体或杆状病毒构建体,其编码细小病毒rep蛋白以在昆虫细胞中表达。在某些实施方案中,针对rep78和rep52蛋白使用单一编码序列,其中用于翻译rep78蛋白的起始密码子是次佳起始密码子,其选自acg、ttg、ctg和gtg,其在昆虫细胞中表达时影响部分外显子跳跃,如美国专利号8,512,981中所述,其内容通过引用整体并入本文,例如,促进与rep52相比更不丰富的rep78的表达,这可能是因为它促进了高载体收率。
在某些实施方案中,病毒表达构建体可以是用于在昆虫细胞中表达的质粒载体或杆状病毒构建体,其包含具有不同密码子偏倚的重复密码子,例如以达到改善的rep蛋白(例如rep78和rep52)的比例从而改善了在昆虫细胞中大规模(商业)生产病毒表达构建体和/或有效载荷构建体载体,如美国专利号8,697,417中所教导的,由于涉及aav复制蛋白及其生产,其内容通过引用整体并入本文。
在另一个实施方案中,可以使用美国专利号8,642,314中描述的方法和构建体来实现改善的rep蛋白的比例,由于涉及aav复制蛋白及其生产,该专利的内容通过引用整体并入本文。
在某些实施方案中,病毒表达构建体可以编码突变的细小病毒rep多肽,其与它们相应的野生型rep多肽相比具有一种或多种改善的特性,例如制备更高的病毒滴度以进行大规模生产。或者,它们可能能够产生质量更好的病毒颗粒或维持更稳定的病毒生产。在非限制性实例中,病毒表达构建体可以编码具有突变的核定位序列或锌指结构域的突变rep多肽,如专利申请us20130023034中所述,由于涉及aav复制蛋白及其生产,其内容通过引用整体并入本文。
在某些实施方案中,病毒表达构建体可以编码具有引入的gly-ala重复区的细小病毒衣壳的组分,其可以起到免疫侵袭序列的作用,如美国专利申请20110171262中所述,由于涉及细小病毒衣壳蛋白,其内容通过引用并入本文。
在本公开的某些实施方案中,病毒表达构建体可以用于在昆虫细胞中生产aav颗粒。在某些实施方案中,可以对衣壳和/或rep基因的野生型aav序列进行修饰,例如以改善病毒颗粒的特性,例如增加的感染性或特异性,或提高产量。
在某些实施方案中,vp编码区编码特定aav血清型的一种或多种aav衣壳蛋白。vp编码区的aav血清型可以相同或不同。在某些实施方案中,可以对vp编码区进行密码子优化。在某些实施方案中,可以针对哺乳动物细胞对vp编码区或核苷酸序列进行密码子优化。在某些实施方案中,可以针对昆虫细胞对vp编码区或核苷酸序列进行密码子优化。在某些实施方案中,可以针对草地贪夜蛾对vp编码区或核苷酸序列进行密码子优化。在某些实施方案中,可以针对sf9或sf21细胞系对vp编码区或核苷酸序列进行密码子优化。
在某些实施方案中,可以对编码一种或多种vp衣壳蛋白的核苷酸序列进行密码子优化,以使其与参考核苷酸序列的核苷酸同源性小于100%。在某些实施方案中,密码子优化的vp核苷酸序列与参考vp核苷酸序列之间的核苷酸同源性小于100%、小于99%、小于98%、小于97%、小于96%、小于95%、小于94%、小于93%、小于92%、小于91%、小于90%、小于89%、小于88%、小于87%、小于86%、小于85%、小于84%、小于83%、小于82%、小于81%、小于80%、小于78%、小于76%、小于74%、小于72%、小于70%、小于68%、小于66%、小于64%、小于62%、小于60%、小于55%、小于50%和小于40%。
在某些实施方案中,本公开的病毒表达构建体或有效载荷构建体可以是杆粒,也被称为杆状病毒质粒或重组杆状病毒基因组。在某些实施方案中,本公开的病毒表达构建体或有效载荷构建体(例如杆粒)可以包含通过本领域技术人员已知和实施的标准分子生物学技术通过同源重组(转座子供体/受体系统)引入到杆粒中的多核苷酸。
在某些实施方案中,引入到杆粒中的多核苷酸(即多核苷酸插入物)可包含与编码蛋白的核苷酸序列有效地连接的表达控制序列。在某些实施方案中,引入到杆粒中的多核苷酸可包含表达控制序列,其包含启动子,例如p10或polh,并且其有效地连接至编码结构性aav衣壳蛋白(例如,vp1、vp2、vp3或其组合)的核苷酸序列。在某些实施方案中,引入到杆粒中的多核苷酸可包含表达控制序列,其包含启动子,例如p10或polh,并且其有效地连接至编码非结构性aav衣壳蛋白(例如,rep78、rep52或其组合)的核苷酸序列。
在某些实施方案中,可将多核苷酸插入物在杆状病毒基因的位置引入到杆粒中。在某些实施方案中,可将多核苷酸插入物在非必需杆状病毒基因的位置引入到杆粒中。在某些实施方案中,可通过用多核苷酸插入物替换杆状病毒基因或杆状病毒基因的一部分将多核苷酸插入物引入到杆粒中。在某些实施方案中,可通过用融合多核苷酸替换杆状病毒基因或杆状病毒基因的一部分将多核苷酸插入物引入到杆粒中,所述融合多核苷酸包含多核苷酸插入物和被替换的杆状病毒基因(或其部分)。
在某些实施方案中,可通过用多核苷酸插入物分裂杆状病毒基因将多核苷酸插入物引入到杆粒中(即,将多核苷酸插入物引入到基因的中间,将杆粒基因的5'部分与3'部分分开)。在某些实施方案中,可通过用融合多核苷酸分裂杆状病毒基因将多核苷酸插入物引入到杆粒中,所述融合多核苷酸包含多核苷酸插入物和被分裂的杆状病毒基因的一部分。在某些实施方案中,融合多核苷酸的3'端包含被分裂的基因的5'部分,使得融合多核苷酸中的基因的5'部分和保留在杆粒中的基因的3'部分形成杆状病毒基因的完整或功能部分。在某些实施方案中,融合多核苷酸的5'端包含被分裂的基因的3'部分,使得融合多核苷酸中的基因的3'部分和保留在杆粒中的基因的5'部分形成杆状病毒基因的完整或功能部分。实施例13和14提供了非限制性实例,其中将融合多核苷酸工程化并生产为包含来自gta基因orf(完全/部分ac-lef12启动子、完全/部分ac-gta基因)的组分。
在某些实施方案中,可将多核苷酸在与杆状病毒基因相关的限制性核酸内切酶(ren)切割位点的位置(即,ren接入点)引入到杆粒中。在某些实施方案中,杆粒中的ren接入点是fsei(对应于gta杆状病毒基因)(ggccggcc)。在某些实施方案中,杆粒中的ren接入点是sdai(对应于dna聚合酶杆状病毒基因)(cctgcagg)。在某些实施方案中,杆粒中的ren接入点是maubi(对应于lef-4杆状病毒基因)(cgcgcgcg)。在某些实施方案中,杆粒中的ren接入点是sbfi(对应于gp64/gp67杆状病毒基因)(cctgcagg)。在某些实施方案中,杆粒中的ren接入点是i-ceui(对应于v-cath杆状病毒基因)(seqidno:1)。在某些实施方案中,杆粒中的ren接入点是avrii(对应于egt杆状病毒基因)(cctagg)。在某些实施方案中,杆粒中的ren接入点是nhei(gctagc)。在某些实施方案中,杆粒中的ren接入点是spei(actagt)。在某些实施方案中,杆粒中的ren接入点是bstz17i(gtatac)。在某些实施方案中,杆粒中的ren接入点是ncoi(ccatgg)。在某些实施方案中,杆粒中的ren接入点是mlui(acgcgt)。
在其中杆粒为双链构建体的某些实施方案中,ren切割位点可在一条链中包含裂解序列以及在另一条链中包含裂解序列的反向互补序列(其也起裂解序列的作用)。因此,多核苷酸插入物(或其链)可包含ren裂解序列或反向互补ren裂解序列(其通常在功能上可互换)。作为非限制性实例,多核苷酸插入物的链可包含fsei裂解序列(ggccggcc)或其反向互补ren裂解序列(ccggccgg)。
可以通过以下方式将多核苷酸引入到这些ren接入点中:(i)提供已被工程化以包含目标ren裂解序列的多核苷酸插入物(例如,已被工程化以在多核苷酸的两端均包含fseiren序列的多核苷酸插入物);(ii)提供包含用于多核苷酸插入的目标ren接入点的杆粒(例如,包含fsei裂解位点的acmnpv杆粒bmon14272的变体(ii)用适当的ren酶消化ren工程化的多核苷酸(例如,使用fsei酶消化在两端均包含fsei区的ren工程多核苷酸,以产生多核苷酸-fsei插入物);(iii)用相同的ren酶消化杆粒,以在ren接入点产生单切杆粒(例如使用fsei酶在fsei位置产生单切杆粒);和(iv)使用适当的连接酶,例如t4连接酶,将多核苷酸插入物连接到单切杆粒中。结果是工程化的杆粒dna,其包含在目标ren接入点处工程化的多核苷酸插入物。
插入过程可以重复一次或多次,以在不同的ren接入点将其他工程化的多核苷酸插入物引入到同一杆粒中(例如,在egt中的avriiren接入点插入第一工程化的多核苷酸插入物,然后在cath基因的i-ceuiren接入点插入第二工程化的多核苷酸插入物,然后在gta基因的fseiren接入点插入第三工程化的多核苷酸插入物)。
在某些实施方案中,可使用限制性核酸内切酶(ren)裂解从杆粒中去除一个或多个野生型基因。在某些实施方案中,可使用限制性核酸内切酶(ren)裂解去除先前已插入到杆粒中的一个或多个工程化的多核苷酸插入物。在某些实施方案中,可使用限制性核酸内切酶(ren)裂解用包含相同ren裂解序列的不同工程化的多核苷酸插入物替换一个或多个工程化的多核苷酸插入物(例如,在fseiren接入点处的工程化的多核苷酸插入物可用包含fseiren裂解序列的不同的工程化多核苷酸插入物替换)。
在某些实施方案中,可以使用在以下文献中教导的的病毒表达构建体:美国专利号us8,512,981、us8,163,543、us8,697,417、us8,642,314,美国专利公开号us20130296532、us20110119777、us20110136227、us20110171262、us20130023034,国际专利申请号pct/nl2008/050613、pct/nl2009/050076、pct/nl2009/050352、pct/nl2011/050170、pct/nl2012/050619和美国专利申请号14/149,953,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以源自以下文献中教导的病毒表达构建体:美国专利号us6,468,524、us6,984,517、us7,479,554、us6,855,314、us7,271,002、us6,723,551,美国专利公开号20140107186,美国专利申请号us09/717,789、us11/936,394、us14/004,379,欧洲专利申请号ep1082413、ep2500434、ep2683829、ep1572893以及国际专利申请pct/us99/11958、pct/us01/09123、pct/ep2012/054303和pct/us2002/035829,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,病毒表达构建体可以包含来自猴物种的序列。在某些实施方案中,病毒表达构建体可以包含序列,其包括但不限于来自以下文献的衣壳和rep序列:国际专利申请pct/us1997/015694、pct/us2000/033256、pct/us2002/019735、pct/us2002/033645、pct/us2008/013067、pct/us2008/013066、pct/us2008/013065、pct/us2009/062548、pct/us2009/001344、pct/us2010/036332、pct/us2011/061632、pct/us2013/041565,美国申请号us13/475535、us13/896722、us10/739096、us14/073979,美国专利公开号us20010049144、us20120093853、us20090215871、us20040136963、us20080219954、us20040171807、us20120093778、us20080090281、us20050069866、us20100260799、us20100247490、us20140044680、us20100254947、us20110223135、us20130309205、us20120189582、us20130004461、us20130315871,美国专利号us6083716、us7838277、us7344872、us8603459、us8105574、us7247472、us8231880、us8524219、us8470310,欧洲专利申请号ep2301582、ep2286841、ep1944043、ep1453543、ep1409748、ep2463362、ep2220217、ep2220241、ep2220242、ep2350269、ep2250255、ep2435559、ep2643465、ep1409748、ep2325298、ep1240345,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以包含来自以下文献中所述的一种或多种病毒构建体的一个或多个核苷酸序列:国际申请号pct/us2002/025096、pct/us2002/033629、pct/us2003/012405,美国申请号us10/291583、us10/420284、us7,319,002,美国专利公开号us20040191762、us20130045186、us20110263027、us20110151434、us20030138772、us20030207259,欧洲申请号ep2338900、ep1456419、ep1310571、ep1359217、ep1427835、ep2338900、ep1456419、ep1310571、ep1359217以及美国专利号us7,235,393和us8,524,446,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以包含在以下文献中描述的序列或组合物:国际专利申请号pct/us1999/025694、pct/us1999/010096、pct/us2001/013000、pct/us2002/25976、pct/us2002/033631、pct/us2002/033630、pct/us2009/041606、pct/us2012/025550,美国专利号us8637255、us8637255、us7186552、us7105345、us6759237、us7056502、us7198951、us8318480、us7790449、us7282199,美国专利公开号us20130059289、us20040057933、us20040057932、us20100278791、us20080050345、us20080050343、us20080008684、us20060204479、us20040057931、us20040052764、us20030013189、us20090227030、us20080075740、us20080075737、us20030228282、us20130323226、us20050014262,美国专利申请号us14/136331、us09/076369、us10/738609,欧洲申请号ep2573170、ep1127150、ep2341068、ep1845163、ep1127150、ep1078096、ep1285078、ep1463805、ep2010178940、us20140004143、ep2359869、ep1453547、ep2341068和ep2675902,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以包含来自以下文献中所述的那些中的一种或多种核苷酸序列的一个或多个核苷酸序列:美国专利号us7186552、us7105345、us6759237、us7056502、us7198951、us8318480、us7790449、us7282199,美国专利公开号us20130059289、us20040057933、us20040057932、us20100278791、us20080050345、us20080050343、us20080008684、us20060204479、us20040057931、us20140004143、us20090227030、us20080075740、us20080075737、us20030228282、us20040052764、us20030013189、us20050014262、us20130323226,美国专利申请号us14/136331、us10/738609,欧洲专利申请号ep1127150、ep2341068、ep1845163、ep1127150、ep1078096、ep1285078、ep2573170、ep1463805、ep2675902、ep2359869、ep1453547、ep2341068,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以包含修饰的aav的构建体,如以下文献中所述:国际专利申请号pct/us1995/014018、pct/us2000/026449、pct/us2004/028817、pct/us2006/013375、pct/us2007/010056、pct/us2010/032158、pct/us2010/050135、pct/us2011/033596,美国专利申请号12/473917、us08/331384、us09/670277,美国专利号us5871982、us5856152、us6251677、us6387368、us6399385、us7906111,欧洲专利申请号ep2000103600,欧洲专利公开号ep797678、ep1046711、ep1668143、ep2359866、ep2359865、ep2357010、ep1046711、ep1218035、ep2345731、ep2298926、ep2292780、ep2292779、ep1668143、us20090197338、ep2383346、ep2359867、ep2359866、ep2359865、ep2357010、ep1866422、us20090317417、ep2016174,美国专利公开号us20110236353、us20070036760、us20100186103、us20120137379和us20130281516,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,本公开的病毒表达构建体可以包含以下文献中描述的一种或多种构建体:国际申请号pct/us1999/004367、pct/us2004/010965、pct/us2005/014556、pct/us2006/009699、pct/us2010/032943、pct/us2011/033628、pct/us2011/033616、pct/us2012/034355,美国专利号us8394386、ep1742668,美国专利公开号us20080241189、us20120046349、us20130195801、us20140031418、ep2425000、us20130101558、ep1742668、ep2561075、ep2561073、ep2699688,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。
表达控制
表达控制区
本公开的病毒表达构建体可以包含一个或多个由表达控制序列编码的表达控制区。在某些实施方案中,表达控制序列用于在病毒生产细胞例如昆虫细胞中表达。在某些实施方案中,表达控制序列有效地连接至编码蛋白的核苷酸序列。在某些实施方案中,表达控制序列有效地连接至vp编码核苷酸序列或rep编码核苷酸序列。
在本文中,术语“编码核苷酸序列”、“编码蛋白的基因”或“编码蛋白的核苷酸序列”是指编码或翻译蛋白产物例如vp蛋白或rep蛋白的核苷酸序列。“有效地连接”是指表达控制序列相对于编码序列定位,使得其可促进编码的基因产物的表达。
“表达控制序列”是指调节与其有效地连接的核苷酸序列的表达的核酸序列。当表达控制序列控制和调节核苷酸序列的转录和/或翻译时,表达控制序列“有效地连接”至该核苷酸序列。因此,表达控制序列可包括启动子、增强子、非翻译区(utr)、内部核糖体进入位点(ires)、转录终止子、编码蛋白的基因前面的起始密码子、内含子的剪接信号和终止密码子。术语“表达控制序列”旨在至少包括其存在被设计为影响表达的序列,并且还可以包括其他有利的组分。例如,前导序列和融合伴侣序列是表达控制序列。该术语还可包括核酸序列的设计,以使得从序列中去除框内和框外的不希望的潜在的起始密码子。它还可以包括核酸序列的设计,以使得去除不希望的潜在剪接位点。它包括指导polya尾的添加的序列或聚腺苷酸化序列(pa),即在mrna的3'端的一串腺嘌呤残基,该序列称为polya序列。它也可以设计为增强mrna的稳定性。在昆虫细胞中已知影响转录和翻译稳定性的表达控制序列,例如启动子,以及影响翻译的序列,例如kozak序列。表达控制序列可以具有调节与之有效地连接的核苷酸序列从而获得更低的表达水平或更高的表达水平的性质。
在某些实施方案中,表达控制序列可包括一个或多个启动子。启动子可包括但不限于杆状病毒主要晚期启动子、昆虫病毒启动子、非昆虫病毒启动子、脊椎动物病毒启动子、核基因启动子、来自一种或多种物种(包括病毒和非病毒元件)的嵌合启动子,和/或合成启动子。在某些实施方案中,启动子可以是ctx、op-ei、ei、δei、ei-1、ph、pio、polh(多面体)、δpolh、dmhsp70、hr1、hsp70、4xhsp27ecre+最小hsp70、ie、ie-1、δie-1、δie、p10、δp10(p10的修饰变体或衍生物)、p5、p19、p35、p40、p6.9及其变体或衍生物。在某些实施方案中,启动子是ctx启动子。在某些实施方案中,启动子是p10启动子。在某些实施方案中,启动子是polh启动子。在某些实施方案中,启动子可选自组织特异性启动子、细胞类型特异性启动子、细胞周期特异性启动子,及其变体或衍生物。在某些实施方案中,启动子可以是cmv启动子、α1-抗胰蛋白酶(α1-at)启动子、甲状腺激素结合球蛋白启动子、甲状腺素结合球蛋白(lps)启动子、hcr-apocii杂种启动子、hcr-haat杂种启动子、白蛋白启动子、载脂蛋白e启动子、α1-at+eaib启动子、肿瘤选择性e2f启动子、单核血液il-2启动子,及其变体或衍生物。在某些实施方案中,启动子是低表达启动子序列。在某些实施方案中,启动子是表达增强的启动子序列。在某些实施方案中,启动子可包括如美国专利申请20110136227中所述的rep或cap启动子,由于涉及表达启动子,该申请的内容通过引用整体并入本文。
在某些实施方案中,病毒表达构建体可以在所有核苷酸序列中包含相同的启动子。在某些实施方案中,病毒表达构建体可以在两个或更多个核苷酸序列中包含相同的启动子。在某些实施方案中,病毒表达构建体可以在两个或更多个核苷酸序列中包含不同的启动子。在某些实施方案中,病毒表达构建体可以在所有核苷酸序列中包含不同的启动子。
在某些实施方案中,病毒表达构建体编码元件以改善在某些细胞类型中的表达。在另一个实施方案中,表达构建体可以包含polh和/或δie-1昆虫转录启动子、cmv哺乳动物转录启动子和/或p10昆虫特异性启动子,用于在哺乳动物或昆虫细胞中表达所需基因。
可以将多于一个的表达控制序列有效地连接至给定的核苷酸序列。例如,启动子序列、翻译起始序列和终止密码子可以有效地连接至核苷酸序列。
在某些实施方案中,病毒表达构建体可以包含含有起始密码子区域的核苷酸序列,例如包含一个或多个起始密码子区的编码aav衣壳蛋白的序列。在某些实施方案中,起始密码子区可以在表达控制序列内。
真核mrna的翻译起始位点部分地由称为kozak序列的核苷酸序列控制,如以下文献中所述:kozak,mcell.1986jan31;44(2):283-92和kozak,m.jcellbiol.1989feb;108(2):229-41,由于涉及kozak序列及其用途,其各自的内容通过引用整体并入本文。通过分子遗传技术,kozak,m.mammgenome.1996aug;7(8):563-74(由于涉及kozak序列及其用途,其内容通过引用整体并入本文),可以将kozak形式的天然存在的和合成的翻译起始位点都用于多肽的生产中。剪接位点是mrna上的序列,其促进在mrna的转录(形成)后去除部分的mrna序列。通常,剪接发生在细胞核中,然后将mrna转运到细胞的细胞质中。
本公开的方法不受特定表达控制序列的使用的限制。但是,当达到一定化学计量的vp产物时(vp1、vp2和vp3分别接近于1:1:10)并且还在rep52或rep40(也称为p19rep)的水平明显高于rep78或rep68(也称为p5rep)时,可以提高生产细胞(例如昆虫细胞)中aav的产量。在某些实施方案中,p5/p19比例低于0.6或更多、低于0.4或低于0.3,但总是至少0.03。这些比例可以在蛋白质水平上测量,或者可以从特定mrna的相对水平暗示。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为1:1:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为2:2:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为2:0:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为1-2:0-2:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为1-2:1-2:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为2-3:0-3:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为2-3:2-3:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为3:3:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为3-5:0-5:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,在病毒生产细胞(例如哺乳动物或昆虫细胞)中生产aav颗粒,其中所有三种vp蛋白均以接近、约或为3-5:3-5:10(vp1:vp2:vp3)的化学计量表达。
在某些实施方案中,将表达控制区工程化以生产选自以下的vp1:vp2:vp3的比例:大约或恰好1:0:10;大约或恰好1:1:10;大约或恰好2:1:10;大约或恰好2:1:10;大约或恰好2:2:10;大约或恰好3:0:10;大约或恰好3:1:10;大约或恰好3:2:10;大约或恰好3:3:10;大约或恰好4:0:10;大约或恰好4:1:10;大约或恰好4:2:10;大约或恰好4:3:10;大约或恰好4:4:10;大约或恰好5:5:10;大约或恰好是l-2:0-2:10;大约或恰好1-2:1-2:10;大约或恰好1-3:0-3:10;大约或恰好1-3:1-3:10;大约或恰好是l-4:0-4:10;大约或恰好1-4:1-4:10;大约或恰好1-5:1-5:10;大约或恰好2-3:0-3:10;大约或恰好2-3:2-3:10;大约或恰好2-4:2-4:10;大约或恰好2-5:2-5:10;大约或恰好3-4:3-4:10;大约或恰好3-5:3-5:10;大约或恰好4-5:4-5:10。
在本公开的某些实施方案中,从杆状病毒衍生的多面体启动子(polh)转录rep52或rep78。rep52或rep78也可以从较弱的启动子转录,例如ie-1启动子的缺失突变体(δie-1启动子),其具有该ie-1启动子的转录活性的约20%。可以使用与δie-1启动子基本上同源的启动子。关于启动子,至少50%、60%、70%、80%、90%或更高的同源性被认为是基本上同源的启动子。
病毒生产细胞和载体
哺乳动物细胞
本文公开的本公开的病毒生产描述了生产aav颗粒或病毒载体的过程和方法,所述aav颗粒或病毒载体与靶细胞接触以递送有效载荷构建体,例如重组aav颗粒或病毒构建体,其包含编码有效载荷分子的核苷酸。病毒生产细胞可以选自任何生物,包括原核(例如细菌)细胞和真核细胞,包括昆虫细胞、酵母细胞和哺乳动物细胞。
在某些实施方案中,本公开的aav颗粒可以在包括哺乳动物细胞的病毒生产细胞中生产。病毒生产细胞可以包括哺乳动物细胞,例如a549、weh1、3t3、10t1/2、bhk、mdck、cos1、cos7、bsc1、bsc40、bmt10、vero.w138、hela、hek293、hek293t(293t)、saos、c2c12、l细胞、ht1080、hepg2和来源于哺乳动物的原代成纤维细胞、肝细胞和成肌细胞。病毒生产细胞可包括来源于哺乳动物物种的细胞,包括但不限于人、猴、小鼠、大鼠、兔和仓鼠,或细胞类型,包括但不限于成纤维细胞、肝细胞、肿瘤细胞、细胞系转化细胞等。
通常用于生产重组aav颗粒的aav病毒生产细胞包括但不限于hek293细胞、cos细胞、c127细胞、3t3细胞、cho细胞、hela细胞、kb细胞、bhk和其他哺乳动物细胞系,如以下文献中所描述的:美国专利号6,156,303、5,387,484、5,741,683、5,691,176、6,428,988和5,688,676;美国专利公开2002/0081721,和国际专利公开号wo00/47757、wo00/24916和wo96/17947,所述文献各自的内容通过引用整体并入本文,只要其与本公开不冲突。在某些实施方案中,aav病毒生产细胞是反式-补充包装细胞系,其提供从复制缺陷性辅助病毒缺失的功能,例如hek293细胞或其他ea反式-补充细胞。
在某些实施方案中,可以使用包装细胞系293-10-3(atcc登记号pta-2361)来生产aav颗粒,如美国专利号us6,281,010中所述,由于涉及293-10-3包装细胞系及其用途,该专利的内容通过引用整体并入本文。
在本公开的某些实施方案中,用于反式-补充e1缺失的腺病毒载体的细胞系(例如hela细胞系,其可在磷酸甘油酸激酶(pgk)启动子的控制下编码腺病毒e1a和腺病毒e1b)可用于aav颗粒的生产,如美国专利号6365394号中所述的,由于涉及hela细胞系及其用途,该专利的内容通过引用整体并入本文。
在某些实施方案中,使用三重转染方法在哺乳动物细胞中生产aav颗粒,其中有效载荷构建体、细小病毒rep和细小病毒cap以及辅助构建体包含在三个不同的构建体中。aav颗粒生产的三个组分的三重转染方法可以用于生产少量病毒,用于包括转导效率、靶组织(嗜性)评估和稳定性的测定。
可以通过三重转染或杆状病毒介导的病毒生产或本领域已知的任何其他方法生产待配制的aav颗粒。可以采用本领域已知的任何合适的允许或包装细胞来生产载体。在某些实施方案中,使用反式-补充包装细胞系,其提供从复制缺陷性辅助病毒缺失的功能,例如293细胞或其他e1a反式-补充细胞。
基因盒可以包含一些或全部的细小病毒(例如,aav)cap和rep基因。在某些实施方案中,通过将编码衣壳和/或rep蛋白的包装载体引入到细胞中,反式提供了一些或全部的cap和rep功能。在某些实施方案中,基因盒不编码衣壳或rep蛋白。或者,使用包装细胞系,其被稳定转化以表达cap和/或rep基因。
在某些实施方案中,根据如us2016/0032254中所描述的程序从培养上清液中生产和纯化重组aav病毒颗粒,由于涉及重组aav病毒颗粒的生产和加工,该文献的内容通过引用整体并入。生产还可以涉及本领域已知的方法,包括使用293t细胞的方法、三重转染或任何合适的生产方法。
在某些实施方案中,哺乳动物病毒生产细胞(例如293t细胞)可处于贴壁/粘附状态(例如,采用磷酸钙)或悬浮状态(例如采用聚乙烯亚胺(pei))。用产生aav所需的质粒(即aavrep/cap构建体、腺病毒辅助构建体和/或itr侧翼有效载荷构建体)转染哺乳动物病毒生产细胞。在某些实施方案中,转染过程可包括任选的培养基改变(例如,对于贴壁形式的细胞,培养基改变;对于悬浮形式的细胞,无培养基改变;如果需要,对于悬浮形式的细胞,培养基改变)。在某些实施方案中,转染过程可包括转染培养基,例如dmem或f17。在某些实施方案中,转染培养基可包括血清或可以是无血清的(例如,贴壁状态的细胞采用磷酸钙和血清,悬浮状态的细胞采用pei而没有血清)。
随后可以通过刮擦(贴壁形式)和/或沉淀(悬浮形式和刮擦的贴壁形式)收集细胞,并将其转移到容器中。为了完全收集生产的细胞,必要时可重复收集步骤。接下来,可通过连续的冻融循环(-80℃至37℃)、化学裂解(例如添加去污剂triton)、机械裂解或通过使细胞培养物在达到~0%的活力后降解来实现细胞裂解。通过离心和/或深度过滤除去细胞碎片。通过dnaqpcr通过dna酶抗性基因组滴定对样品的aav颗粒进行定量。
根据基因组拷贝数(每毫升基因组颗粒数)测量aav颗粒滴度。如先前所报道(clark等人(1999)hum.genether.,10:1031-1039;veldwijk等人(2002)mol.ther.,6:272-278,由于涉及颗粒浓度的测量,其各自内容通过引用整体并入本文中),基因组颗粒浓度基于载体dna的dnaqpcr。
昆虫细胞
本公开的病毒生产包括用于生产与靶细胞接触以递送有效载荷构建体例如重组病毒构建体的aav颗粒或病毒载体的过程和方法,所述有效载荷构建体包含编码有效载荷分子的核苷酸。在某些实施方案中,本公开的aav颗粒或病毒载体可以在病毒生产细胞(包括昆虫细胞)中生产。
培养中昆虫细胞的生长条件以及培养中昆虫细胞中异源产物的生产在本领域中是众所周知的,参见美国专利号6,204,059,由于涉及昆虫细胞在病毒生产中的生长和用途,其内容通过引用整体并入本文。
根据本公开,可以使用允许细小病毒复制并且可以在培养物中保持的任何昆虫细胞。通常用于生产重组aav颗粒的aav病毒生产细胞包括但不限于草地贪夜蛾,包括但不限于sf9或sf21细胞系、果蝇(drosophila)细胞系或蚊子细胞系,例如白纹伊蚊(aedesalbopictus)起源的细胞系。昆虫细胞用于表达异源蛋白的用途已得到充分证明,将核酸例如载体(例如昆虫细胞相容性载体)引入到此类细胞中的方法以及在培养中维持此类细胞的方法也已得到充分证明。参见,例如,methodsinmolecularbiology,ed.richard,humanapress,nj(1995);o'reilly等人,baculovirusexpressionvectors,alaboratorymanual,oxforduniv.press(1994);samulski等人,j.vir.63:3822-8(1989);kajigaya等人,proc.nat'l.acad.sci.usa88:4646-50(1991);ruffing等人,j.vir.66:6922-30(1992);kimbauer等人,vir.219:37-44(1996);zhao等人,vir.272:382-93(2000);和samulski等人,u.s.pat.no.6,204,059,由于涉及昆虫细胞在病毒生产中的用途,其各自的内容通过引用整体并入本文。
在一个实施方案中,使用在wo2015/191508中描述的方法制备aav颗粒,该文献的内容通过引用整体并入本文,只要其与本公开不冲突。
在某些实施方案中,可以使用昆虫宿主细胞系统与杆状病毒系统的组合(例如,如luckow等人,bio/technology6:47(1988)所述的)。在某些实施方案中,用于制备嵌合肽的表达系统是粉纹夜蛾(trichoplusiani)tn5b1-4昆虫细胞/杆状病毒系统,其可以用于高水平的蛋白,如美国专利号6660521中所述由于涉及病毒颗粒的生产,该文献的内容通过引用整体并入本文。
昆虫细胞的扩增、培养、转染、感染和储存可以在本领域已知的任何细胞培养基、细胞转染培养基或储存培养基中进行,包括hyclonesfx昆虫细胞培养基(hyclonesfxinsectcellculturemedia)、表达系统esfaf昆虫细胞培养基(expressionsystemesfafinsectcellculturemedium)、thermofishersf900ii培养基、thermofishersf900iii培养基或thermofishergrace昆虫培养基(thermofishergrace’sinsectmedia)。本公开的昆虫细胞混合物还可包括本公开中描述的任何制剂添加剂或成分,包括(但不限于)盐、酸、碱、缓冲剂、表面活性剂(例如泊洛沙姆188/普朗尼克f-68),以及其他已知的培养基成分。制剂添加剂可以逐渐或“快加(spikes)”引入(短时间内大量引入)。
杆状病毒生产系统
在某些实施方案中,本公开的方法可包括使用病毒表达构建体和有效载荷构建体载体在杆状病毒系统中生产aav颗粒或病毒载体。在某些实施方案中,杆状病毒系统包括杆状病毒表达载体(bev)和/或杆状病毒感染的昆虫细胞(biic)。在某些实施方案中,本公开的病毒表达构建体或有效载荷构建体可以是杆粒,也被称为杆状病毒质粒或重组杆状病毒基因组。在某些实施方案中,本公开的病毒表达构建体或有效载荷构建体可以是通过本领域技术人员已知和执行的标准分子生物学技术通过同源重组(转座子供体/受体系统)引入到杆粒中的多核苷酸。分离的病毒复制细胞群的转染产生两组或更多组(例如,两组、三组)杆状病毒(bev)、一个或多个可以包含病毒表达构建体的组(表达bev)以及一个或多个可以包含有效载荷构建体的组(有效载荷bev)。杆状病毒可以用于感染病毒生产细胞以生产aav颗粒或病毒载体。
在某些实施方案中,该方法包括转染单个病毒复制细胞群以产生单个杆状病毒(bev)组,其包括病毒表达构建体和有效载荷构建体二者。这些杆状病毒可以用于感染病毒生产细胞,以生产aav颗粒或病毒载体。
在某些实施方案中,使用杆粒转染剂如promegafugenehd、wfi水或thermofishercellfectinii试剂产生bev。在某些实施方案中,在病毒生产细胞例如昆虫细胞中产生并扩增bev。
在某些实施方案中,该方法利用包括一种或多种bev的病毒生产细胞(包括杆状病毒感染的昆虫细胞(biic))的种子培养物。种子biic已经用包括病毒表达构建体的表达bev以及包括有效载荷构建体的有效载荷bev转染/转导/感染。在某些实施方案中,收获种子培养物,将其分成等分试样并冷冻,并可以在以后的时间用于起始初始生产细胞群的转染/转导/感染。在某些实施方案中,将种子biic库储存在-80℃或ln2蒸气中。
杆状病毒由几种对杆状病毒的功能和复制必不可少的必需蛋白组成,例如复制蛋白、包膜蛋白和衣壳蛋白。因此,杆状病毒基因组包括几个编码必需蛋白的必需基因核苷酸序列。作为非限制性实例,基因组可包括必需基因区,该必需基因区包括编码杆状病毒构建体的必需蛋白的必需基因核苷酸序列。必需蛋白可包括:gp64杆状病毒包膜蛋白、vp39杆状病毒衣壳蛋白或杆状病毒构建体的其他类似必需蛋白。
用于在昆虫细胞(包括但不限于草地贪夜蛾(sf9)细胞)中产生aav颗粒的杆状病毒表达载体(bev),可提供高滴度的病毒载体产物。编码病毒表达构建体和有效载荷构建体的重组杆状病毒引发病毒载体复制细胞的生产性感染。从初次感染释放的感染性杆状病毒颗粒继发感染培养物中的其他细胞,并在多个感染周期内以指数方式感染整个细胞培养群,其是初始感染复数的函数,参见urabe,m.等人jvirol.2006feb;80(4):1874-85,由于涉及bev和病毒颗粒的生产和使用,其内容通过引用整体并入本文。
用杆状病毒在昆虫细胞系统中生产aav颗粒可以解决了已知的杆状病毒的遗传和物理不稳定性。
在某些实施方案中,本公开的生产系统通过利用无滴度的感染细胞保存和放大系统来解决杆状病毒经过多次传代的不稳定性。用编码aav颗粒的结构和/或非结构组分的病毒表达构建体转染病毒生产细胞的小规模种子培养物。收获杆状病毒感染的病毒生产细胞,分成等分试样,然后将其冷冻保存在液氮中;等分试样保留了感染大规模病毒生产细胞培养物的活力和感染力,wasilkodj等人proteinexprpurif.2009jun;65(2):122-32,由于涉及bev和病毒颗粒的生产和使用,其内容通过引用整体并入本文。
可以使用遗传稳定的杆状病毒来产生用于在无脊椎动物细胞中生产aav颗粒的一种或多种组分源。在某些实施方案中,缺陷型杆状病毒表达载体可以在昆虫细胞中以游离体维持。在这样的实施方案中,相应的杆粒载体用复制控制元件工程化,所述复制控制元件包括但不限于启动子、增强子和/或细胞周期调节的复制元件。
在某些实施方案中,杆状病毒可以用标记物工程化以重组到几丁质酶/组织蛋白酶基因座中。chia/v-cath基因座对于在组织培养中传播杆状病毒不是必需的,并且v-cath(ec3.4.22.50)是半胱氨酸内切蛋白酶,其对含arg-arg二肽的底物最活跃。arg-arg二肽存在于浓核病毒和细小病毒衣壳结构蛋白中,但在依赖病毒vp1中则很少出现。
在某些实施方案中,将允许杆状病毒感染的稳定的病毒生产细胞用aav复制和载体产生所必需的任何元件的至少一个稳定的整合拷贝进行工程化,所述元件包括但不限于整个aav基因组、rep和cap基因、rep基因、cap基因、作为单独转录盒的每个rep蛋白、作为单独转录盒的每个vp蛋白、aap(装配激活蛋白)或至少一个带有天然或非天然启动子的杆状病毒辅助基因。
在某些实施方案中,杆状病毒表达载体(bev)基于acmnpv杆状病毒或bmnpv杆状病毒bmnpv。在某些实施方案中,本公开的杆粒是基于acmnpv杆粒(例如bmon14272、vace25ko或vaclef11ko)(即,其工程化变体)。
通常,bev并未针对aav生产进行优化。在基于杆粒的bev中病毒蛋白的异常蛋白水解降解通常可导致预料不到的问题,这不包括使用杆状病毒/sf9系统可靠地大规模生产aav衣壳蛋白。
本公开确定了蛋白降解的源头和途径。本公开提出了一种相应的系统,其可包括经修饰的bev(mbev),其在杆状病毒/sf9系统中生产的过程中消除了raav衣壳和衣壳变体的降解。这种新系统不仅能够减轻蛋白水解降解,而且还导致功能性aav衣壳蛋白的提高的产量,从而提供了更广阔的收获窗口以支持大规模aav生产,从而支持临床试验。
在某些实施方案中,本公开提出了经修饰的杆状病毒表达载体(mbev)及其在杆状病毒/sf9系统中大规模生产重组腺相关病毒(raav)中的使用方法。
在某些实施方案中,杆状病毒表达载体(bev)是其中杆状病毒v-cath基因已部分或完全缺失(“v-cath缺失的bev”)或突变的bev。在某些实施方案中,bev缺少v-cath基因或包含v-cath基因的突变失活形式。在某些实施方案中,bev缺少v-cath基因。在某些实施方案中,bev包含v-cath基因的突变失活形式。
其中杆状病毒v-cath基因已部分缺失、完全缺失或突变失活的bev构建体的非限制性实例包括bev,其包含选自seqidno:2、seqidno:3或seqidno:4的核苷酸序列。在某些实施方案中,本公开的bev构建体包含seqidno:2、seqidno:3或seqidno:4。在某些实施方案中,本公开的bev构建体包含seqidno:2。在某些实施方案中,本公开的bev构建体包含seqidno:3。在某些实施方案中,本公开的bev构建体包含seqidno:4。
本公开的病毒生产杆粒可包括某些杆状病毒基因或基因座的缺失。
其他
在某些实施方案中,表达宿主包括但不限于埃希氏菌属(escherichia)、芽孢杆菌属(bacillus)、假单胞菌属(pseudomonas)、沙门氏菌属(salmonella)内的细菌物种。
在某些实施方案中,可以使用包含稳定整合在细胞染色体内的aavrep和cap基因的宿主细胞进行aav颗粒的生产。在非限制性实例中,根据美国专利号7238526中所述的方法和构建体,可以将在其染色体中稳定整合了至少两个拷贝的aavrep基因和aavcap基因的宿主细胞用于生产aav颗粒,由于涉及病毒颗粒的生产,该专利的内容通过引用整体并入本文。
在某些实施方案中,可以在用包含核酸序列的分子稳定转化的宿主细胞中生产aav颗粒,所述核酸序列允许在宿主细胞中调节罕见限制酶的表达,如us20030092161和ep1183380中所述,由于涉及病毒颗粒的生产,其内容通过引用整体并入本文。
在某些实施方案中,生产aav颗粒的生产方法和细胞系可以包括但不限于以下文献中教导的那些:pct/us1996/010245、pct/us1997/015716、pct/us1997/015691、pct/us1998/019479、pct/us1998/019463、pct/us2000/000415、pct/us2000/040872、pct/us2004/016614、pct/us2007/010055、pct/us1999/005870、pct/us2000/004755;美国专利申请号us08/549489、us08/462014、us09/659203、us10/246447、us10/465302;美国专利号us6281010、us6270996、us6261551、us5756283(转让给nih)、us6428988、us6274354、us6943019、us6482634、(转让给nih:us7238526、us6475769)、us6365394(转让给nih)、us7491508、us7291498、us7022519、us6485966、us6953690、us6258595、ep2018421、ep1064393、ep1163354、ep835321、ep931158、ep950111、ep1015619、ep1183380、ep2018421、ep1226264、ep1636370、ep1163354、ep1064393、us20030032613、us20020102714、us20030073232、us20030040101(转让给nih)、us20060003451、us20020090717、us20030092161、us20070231303、us20060211115、us20090275107、us2007004042、us20030119191、us20020019050,其各自的内容通过引用整体并入本文,只要其与本公开不冲突。
iii.定义
在本公开的各个地方,以组或范围公开了本公开的化合物的取代基或特性。特别意图表示的是,本公开包括这种组和范围的成员的每个单独或子组合。
除非另外说明,否则以下术语和短语具有下面描述的含义。所述定义无意在性质上成为限制性的,而是用于提供本公开的某些方面的更清楚的理解。
约:如本文所用,术语“约”是指所述值的+/-10%。
腺相关病毒:本文所用的术语“腺相关病毒”或“aav”是指包含任何颗粒、序列、基因、蛋白或衍生自其的组分的依赖病毒属的成员。
aav颗粒:如本文所用,“aav颗粒”是包含衣壳和病毒基因组的病毒,所述病毒基因组具有至少一个有效载荷区和至少一个itr区。本公开的aav颗粒可以重组产生并且可以基于腺相关病毒(aav)的亲本或参考序列。aav颗粒可衍生自本文所述或本领域已知的任何血清型,包括血清型的组合(即“假型”aav)或多种基因组(例如单链或自互补型)。另外,aav颗粒可能是复制缺陷的和/或靶向的。
活性:如本文所用,术语“活性”是指事物正在发生或正在完成的状况。本公开的组合物可以具有活性,并且该活性可以涉及一种或多种生物学事件。
施用:如本文所用,术语“施用”是指向受试者提供药剂或组合物。
联合施用:如本文所用,术语“联合施用(administeredincombination)”或“组合施用(combinedadministration)”是指将两种或更多种药剂同时或在一定间隔内施用于受试者,从而可能存在每种药剂对患者的作用的重叠。在某些实施方案中,它们在彼此的约60、30、15、10、5或1分钟内施用。在某些实施方案中,药剂的施用间隔足够地紧密地在一起,从而实现组合(例如,协同)作用。
改善:如本文所用,术语“改善(amelioration)”或“改善(ameliorating)”是指减轻至少一种病症或疾病指标的严重性。例如,在神经退行性疾病的背景下,改善包括神经元损失的减少。
动物:如本文所用,术语“动物”是指动物界的任何成员。在某些实施方案中,“动物”是指处于任何发育阶段的人类。在某些实施方案中,“动物”是指处于任何发育阶段的非人类动物。在某些实施方案中,非人类动物是哺乳动物(例如,啮齿动物、小鼠、大鼠、兔、猴、狗、猫、绵羊、牛、灵长类动物或猪)。在某些实施方案中,动物包括但不限于哺乳动物、禽类、爬行动物、两栖动物、鱼类和蠕虫。在某些实施方案中,动物是转基因动物、基因工程动物或克隆。
反义链:如本文所用,术语sirna分子的“反义链”或“第一链”或“引导链”是指与靶向沉默基因的mrna的约10-50个核苷酸(例如约15-30、16-25、18-23或19-22个核苷酸)的区段基本互补的链。所述反义链或第一链具有与期望的靶mrna序列足够互补的序列以指导靶标特异性的沉默,例如,足以触发rnai机制或过程对期望的靶mrna的破坏的互补性。
大约:如本文所用,如应用于一个或多个值,术语“大约”或“约”是指类似于所述参考值的值。在某些实施方案中,术语“大约”是指落入所述参考值的任一方向(大于或小于)的25%、20%、19%、18%、17%、16%、15%、14%、13%、12%、11%、10%、9%、8%、7%、6%、5%、4%、3%、2%、1%或更少内的值的范围,除非另有说明或从上下文中可以明显看出(除非该数字超过可能值的100%)。
结合:如本文所用,术语“结合”、“共轭”、“连接”、“附接”和“拴系”,当针对两个或多个部分使用时,是指这些部分直接地或通过一个或多个另外的用作连接剂的部分物理上彼此结合或连接,以形成足够稳定的结构,从而在使用该结构的条件(例如生理条件)下,这些部分保持物理结合。“结合”不必严格地通过直接共价化学键来实现。也可能暗示离子或氢键或基于杂交的连接性足够稳定,以使“结合的”实体保持物理结合。
杆状病毒表达载体(bev):如本文所用,bev是杆状病毒表达载体,即杆状病毒来源的多核苷酸载体。使用bev的系统称为杆状病毒表达载体系统(bevs)。
mbev或修饰的bev:如本文所用,修饰的bev是杆状病毒来源的表达载体,其已通过添加和/或缺失和/或复制和/或倒置以下的一个或多个从起始bev(无论是野生型还是人工型)改变而来:基因;基因片段;切割位点;限制位点;序列区;编码有效载荷或目标基因的序列;或前述各项的组合。
双功能:如本文所用,术语“双功能”是指能够或维持至少两种功能的任何物质、分子或部分。这些功能可能会实现相同或不同的结果。产生功能的结构可以相同或不同。
biic:如本文所用,biic是杆状病毒感染的昆虫细胞。
生物相容的:如本文所用,术语“生物相容的”是指与活细胞、组织、器官或系统相容,所述活细胞、组织、器官或系统具有很少的或没有免疫系统的伤害、毒性或排斥风险。
可生物降解的:如本文所用,术语“可生物降解的”是指能够通过生物的作用分解成无害的产品。
生物学活性的:如本文所用,短语“生物学活性的”是指任何物质在生物系统和/或生物中具有活性的特征。例如,当施用于生物时对该生物具有生物学作用的物质被认为具有生物学活性。在特定的实施方案中,即使编码的有效载荷的一部分具有生物学活性或模拟被认为与生物学相关的活性,则本公开的aav颗粒也可以被认为具有生物学活性。
衣壳:如本文所用,术语“衣壳”是指病毒颗粒的蛋白壳。
密码子优化的:如本文所用,术语“密码子优化的(codonoptimized)”或“密码子优化(codonoptimization)”是指修饰的核酸序列,其编码与亲本/参考序列相同的氨基酸序列,但是其已经被改变使得对修饰的核酸序列的密码子进行优化或改进以在特定系统(例如特定物种或物种组)中表达。作为非限制性实例,可以对包含aav衣壳蛋白的核酸序列进行密码子优化以在昆虫细胞或特定的昆虫细胞如草地贪夜蛾(spodopterafrugiperda)细胞中表达。密码子优化可以使用本领域技术人员已知的方法和数据库来完成。
互补的和基本上互补的:本文使用的术语“互补的”表示多核苷酸彼此形成碱基对的能力。碱基对通常由反向平行的多核苷酸链中的核苷酸单元之间的氢键形成。互补的多核苷酸链可以以沃森-克里克方式(例如,a至t、a至u、c至g)或以允许形成双链体的任意其它方式形成碱基对。本领域技术人员知晓,当使用rna而不是dna时,尿嘧啶(而不是胸腺嘧啶)是被认为与腺苷互补的碱基。但是,当在本公开的上下文中提及u时,暗示置换t的能力,除非另有说明。完美互补性或100%互补性表示这样的情形:其中一个多核苷酸链的每个核苷酸单元都可以与第二个多核苷酸链的核苷酸单元形成氢键。小于完美互补性表示这样的情形:其中两个链的一些(但是并非全部)核苷酸单元可以彼此形成氢键。例如,对于两个20聚体,如果在每个链上的仅两个碱基对可以彼此形成氢键,那么所述多核苷酸链表现出10%互补性。在相同的实例中,如果在每个链上的18个碱基对可以彼此形成氢键,那么所述多核苷酸链表现出90%互补性。本文使用的术语“基本上互补的”是指,所述sirna具有足以结合期望的靶mrna和触发靶mrna的rna沉默的序列(例如,在反义链中)。
化合物:本公开的化合物包括存在于中间或最终化合物中的原子的所有同位素。“同位素”是指由原子核中中子数量不同产生的具有相同原子序数但具有不同质量数的原子。例如,氢的同位素包括氚和氘。
本公开的化合物和盐可以通过常规方法与溶剂或水分子组合制备以形成溶剂化物和水合物。
有条件的活性:如本文所用,术语“有条件的活性”是指野生型多肽的突变体或变体,其中该突变体或变体在生理条件下比亲本多肽具有或多或少的活性。此外,与亲本多肽相比,有条件的活性多肽在异常条件下的活性可能增加或降低。有条件的活性多肽可以在正常生理条件或异常条件下可逆地或不可逆地失活。
保守的:如本文所用,术语“保守的”分别是指多核苷酸序列或多肽序列的核苷酸或氨基酸残基,其是在比较的两个或更多个序列的相同位置上未发生改变的那些。相对保守的核苷酸或氨基酸是比序列中其他地方出现的核苷酸或氨基酸更相关的序列中保守的核苷酸或氨基酸。
在某些实施方案中,如果两个或更多个序列彼此100%相同,则称它们为“完全保守的”。在某些实施方案中,如果两个或更多个序列彼此至少70%同一、至少80%同一、至少90%同一或至少95%同一,则被认为是“高度保守的”。在某些实施方案中,如果两个或更多个序列彼此约70%同一、约80%同一、约90%同一、约95%、约98%或约99%同一,则被称为“高度保守”。在某些实施方案中,如果两个或更多个序列彼此至少30%同一、至少40%同一、至少50%同一、至少60%同一、至少70%同一、至少80%同一,至少90%同一或至少95%同一,则被认为是“保守的”。在某些实施方案中,如果两个或更多个序列彼此约30%同一、约40%同一、约50%同一、约60%同一、约70%同一同、约80%同一、约90%同一、约95%同一,约98%同一或约99%同一,则被称为“保守的”。序列的保守性可以应用于多核苷酸或多肽的全长,或者可以应用于其一部分、区域或特征。
控制元件:如本文所用,“控制元件”、“调控控制元件”或“调控序列”是指启动子区域、聚腺苷酸化信号、转录终止序列、上游调节域、复制起点、内部核糖体进入位点(“ires”)、增强子等,其在受体细胞中提供编码序列的复制、转录和翻译。并非总是需要所有这些控制元件,只要选择的编码序列能够在适当的宿主细胞中复制、转录和/或翻译即可。
控制释放:如本文所用,术语“控制释放”是指符合释放的特定样式以影响治疗结果的药物组合物或化合物释放曲线。
细胞抑制:如本文所用,“细胞抑制”是指抑制、减少、阻抑细胞(例如,哺乳动物细胞(例如,人细胞))、细菌、病毒、真菌、原生动物、寄生虫、朊病毒或其组合的生长、分裂或繁殖。
细胞毒性的:如本文所用,“细胞毒性的”是指杀死细胞(例如,哺乳动物细胞(例如,人细胞))、细菌、病毒、真菌、原生动物、寄生虫、朊病毒或其组合或引起对其的伤害、毒性或致命作用。
递送:如本文所用,“递送”是指递送aav颗粒、化合物、物质、实体、部分、货物(cargo)或有效载荷。
递送剂:如本文所用,“递送剂”是指至少部分促进aav颗粒向靶细胞的体内递送的任何物质。
不稳定的:如本文所用,术语“不稳定的”、“不稳定”或“不稳定区域”是指比相同区域或分子的起始、野生型或初始形式更不稳定的区域或分子。
可检测的标记:如本文所用,“可检测的标记”是指一种或多种标志物、信号或部分,其与易于通过本领域已知的方法检测的另一实体连接、相连或结合,所述方法包括放射线照相、荧光、化学发光、酶促活性、吸光度等。可检测的标记包括放射性同位素、荧光团、发色团、酶、染料、金属离子、诸如生物素、亲和素、链霉亲和素和半抗原之类的配体、量子点等。可检测的标记可以位于本文公开的肽或蛋白中的任何位置。它们可以在氨基酸、肽或蛋白内,或位于n或c末端。
消化:如本文所用,术语“消化”是指分解成较小的碎片或组分。当涉及多肽或蛋白时,消化导致肽的生产。
远端:如本文所用,术语“远端”是指位于远离中心或远离目标点或区域。
给药方案:如本文所用,“给药方案”是施用的时间表,或医生确定的治疗、预防或姑息治疗方案。
包封:如本文所用,术语“包封”是指封入、包围或围住。
工程化的:如本文所用,当本公开的实施方案被设计为具有不同于起始点、野生型或初始分子的特征或特性时,则其是被“工程化的”。
有效量:如本文所用,术语药剂的“有效量”是这样的量:所述量足以实现有益的或期望的结果,例如,临床结果,且这样,“有效量”取决于它所应用的上下文。例如,在施用治疗癌症的药剂的上下文中,与不施用该药剂获得的应答相比,该药剂的有效量例如是足以实现如本文所定义的治疗的量。
表达:如本文所用,核酸序列的“表达”是指以下事件中的一个或多个:(1)从dna序列产生rna模板(例如,通过转录);(2)加工rna转录物(例如,通过剪接、编辑、5'帽形成和/或3'末端加工);(3)将rna翻译成多肽或蛋白;(4)多肽或蛋白的翻译后修饰。
特征:如本文所用,“特征”是指特点、特性或独特要素。
制剂:如本文所用,“制剂”包括至少一种aav颗粒和递送剂或赋形剂。
片段:如本文所用,“片段”是指一部分。例如,蛋白片段可以包含通过消化从培养的细胞分离的全长蛋白而获得的多肽。
功能性的:如本文所用,“功能性的”生物分子是以表现出表征其的性质和/或活性的形式的生物分子。
基因表达:术语“基因表达”表示这样的过程:核酸序列通过该过程发生成功转录和(在大多数情况下)翻译以生产蛋白或肽。为了清楚起见,当提及“基因表达”的测量时,这应当被理解为是指,可以测量转录的核酸产物(例如,rna或mrna)或翻译的氨基酸产物(例如,多肽或肽)。测量rna、mrna、多肽和肽的量或水平的方法是本领域众所周知的。
同源性:如本文所用,术语“同源性”是指聚合物分子之间例如多核苷酸分子(例如dna分子和/或rna分子)之间和/或多肽分子之间的整体相关性。在某些实施方案中,如果聚合物分子的序列为至少25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或99%相同或相似,则认为它们彼此“同源”。术语“同源的”必定是指至少两个序列(多核苷酸或多肽序列)之间的比较。根据本公开,如果两个多核苷酸序列编码的多肽是至少一个至少约20个氨基酸片段中的至少约50%、60%、70%、80%、90%、95%或至少99%,则它们被认为是同源的。在某些实施方案中,同源多核苷酸序列的特征在于能够编码至少4-5个独特指定的氨基酸片段的能力。对于长度少于60个核苷酸的多核苷酸序列,同源性由编码至少4-5个独特指定的氨基酸的片段能力决定。根据本公开,如果蛋白对于至少一个约20个氨基酸的片段至少约50%、60%、70%、80%或90%相同,则认为两个蛋白序列是同源的。
异源区域:如本文所用,术语“异源区域”是指将不被视为同源区域的区域。
同源区域:如本文所用,术语“同源区域”是指位置、结构、进化起源、特征、形式或功能相似的区域。
同一性:如本文所用,术语“同一性”是指聚合物分子之间例如多核苷酸分子(例如dna分子和/或rna分子)之间和/或多肽分子之间的整体相关性。例如,可以通过比对两个序列以达到最佳比对的目的来进行两个多核苷酸序列的同一性百分比的计算(例如,可以在第一和第二核酸序列中的一个或两个中引入缺口以实现最佳比对,以及出于比对的目的,可以忽略非同一的序列)。在某些实施方案中,为了比对的目的而比对的序列的长度为参考序列长度的至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少95%或100%。然后比较相应核苷酸位置上的核苷酸。当第一序列中的位置被与第二序列中的相应位置相同的核苷酸占据时,则分子在该位置是相同的。考虑到缺口的数目和每个缺口的长度,两个序列之间的同一性百分比是序列共享的相同位置数的函数,需要引入所述缺口以实现两个序列的最佳比对。序列的比对和两个序列之间同一性百分比的确定可以使用数学算法来完成。例如,两个核苷酸序列之间的同一性百分数可以使用诸如以下描述的那些:computationalmolecularbiology,lesk,a.m.,ed.,oxforduniversitypress,newyork,1988;biocomputing:informaticsandgenomeprojects,smith,d.w.,ed.,academicpress,newyork,1993;sequenceanalysisinmolecularbiology,vonheinje,g.,academicpress,1987;computeranalysisofsequencedata,parti,griffin,a.m.,和griffin,h.g.,eds.,humanapress,newjersey,1994;和sequenceanalysisprimer,gribskov,m.anddevereux,j.,eds.,mstocktonpress,newyork,1991;其各自均通过引用并入本文,只要其与本公开不冲突。例如,可以使用meyers和miller(cabios,1989,4:11-17)的算法确定两个核苷酸序列之间的同一性百分比,该算法已使用pam120权重残基表、缺口长度罚分12和缺口罚分4,该算法已经合并到align程序(2.0版)中。或者,可以使用nwsgapdna.cmp矩阵的gcg软件包中的gap程序确定两个核苷酸序列之间的同一性百分比。通常用于确定序列之间的同一性百分比的方法包括但不限于在carillo,h.和lipman,d.,siamjappliedmath.,48:1073(1988)中公开的那些方法,该文献通过引用并入本文,只要其与本公开不冲突。确定同一性的技术已编入公共可用的计算机程序中。确定两个序列之间的同源性的示例性计算机软件包括但不限于gcg程序包,devereux,j.,等人,nucleicacidsresearch,12(1),387(1984)),blastp,blastn和fastaaltschul,s.f.等人,j.molec.biol.,215,403(1990))。
抑制基因的表达:本文中使用的短语“抑制基因的表达”是指造成基因的表达产物的量的减少。所述表达产物可以是从基因转录的rna(例如mrna)或从基因转录的mrna所翻译成的多肽。通常,mrna水平的降低会导致从其翻译的多肽的水平的降低。通过使用用于测量mrna或蛋白的标准技术,可以确定表达的水平。
体外:本文使用的术语“体外”表示在人工环境中,例如在试管或反应容器中、在细胞培养物中、在培养皿等中,发生的事件,而不是在生物(例如,动物、植物或微生物)内发生的事件。
体内:本文使用的术语“体内”表示在生物(例如,动物、植物或微生物或其细胞或组织)内发生的事件。
分离的:如本文所用,术语“分离的”是指已经从与之结合的至少一些组分中分离的物质或实体(无论是在自然界还是在实验环境中)。分离出的物质相对于与它们结合的物质可能具有不同的纯度水平。分离的物质和/或实体可以从最初与它们相连的至少约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%或更多的其他组分分开。在某些实施方案中,分离的试剂大于约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、98%、约99%或大于约99%的纯度。如本文所用,如果物质基本上不含其他组分,则该物质是“纯的”。
基本上分离的:“基本上分离的”是指物质与形成或检测出物质的环境实质上分离。部分分离可以包括例如富含本发明的物质或aav颗粒的组合物。基本上分离可包括含有按重量计至少约50%、至少约60%、至少约70%、至少约80%、至少约90%、至少约95%、至少约97%或至少约99%的本公开的化合物或其盐的组合物。分离化合物及其盐的方法是本领域常规的。
接头:如本文所用,“接头”是指连接两个分子的一个分子或一组分子。接头可以是连接编码两个不同多肽的两个核酸序列的核酸序列。接头可能会被翻译,也可能不会被翻译。接头可以是可裂解的接头。
microrna(mirna)结合位点:如本文所用,microrna(mirna)结合位点代表mirna的“种子”区域至少与其结合的核酸转录物的核苷酸位置或区域。
修饰的:如本文所用,“修饰的”是指本公开的分子的改变的状态或结构。分子可以多种方式被修饰,包括化学、结构和功能上的修饰。如本文所用,当本公开的实施方案有或具有自起始点、野生型或天然分子变化的特征或特性(无论是结构还是化学性质)时,其均被“修饰”。
突变:如本文所用,术语“突变”是指基因结构的任何改变,从而形成可以传递至后代的变体(也称为“突变体”)形式。基因中的突变可能是由于dna中单个碱基的交替,或基因或染色体较大部分的缺失、插入或重排引起的。
天然存在的:如本文所用,“天然存在的”或“野生型”是指自然存在而没有人工辅助或人的介入。
非人类脊椎动物:如本文所用,“非人类脊椎动物”包括智人以外的所有脊椎动物,包括野生和驯养的物种。非人类脊椎动物的实例包括但不限于哺乳动物,例如羊驼、白臀野牛、野牛、骆驼、猫、牛、鹿、狗、驴、大额牛、山羊、豚鼠、马、美洲驼、骡、猪、兔、驯鹿、绵羊、水牛和牦牛。
脱靶:本文中使用的“脱靶”表示对任意一种或多种靶标、基因和/或细胞转录物的任何非目的的效果。
开放阅读框:如本文所用,“开放阅读框”或“orf”是指在阅读框末端之外的给定阅读框内不包含终止密码子的序列。
有效地连接:如本文所用,短语“有效地连接”是指两个或更多个分子、构建体、转录物、实体、部分等之间的功能连接。
患者:如本文所用,“患者”是指可能寻求治疗或需要治疗、要求治疗、正在接受治疗、将要接受治疗的受试者,或由受过训练的专业人员针对特定疾病或疾病进行护理的受试者。
有效载荷:如本文所用,“有效载荷”或“有效载荷区”是指由病毒基因组编码或在其中的一个或多个多核苷酸或多核苷酸区,或此类多核苷酸或多核苷酸区的表达产物,例如转基因、编码多肽或多个多肽的多核苷酸,或调节核酸或调节性核酸。
有效载荷构建体:如本文所用,“有效载荷构建体”是一种或多种载体构建体,其包含编码或包含在一侧或两侧侧接有反向末端重复(itr)序列的有效载荷的多核苷酸区域。有效载荷构建体提供了在病毒生产细胞中复制以生产治疗性病毒基因组的模板。
有效载荷构建体载体:如本文所用,“有效载荷构建体载体”是编码或包含有效载荷构建体以及用于在细菌细胞中复制和表达有效载荷构建体的调节区的载体。
有效载荷构建体表达载体:如本文所用,“有效载荷构建体表达载体”是编码或包含有效载荷构建体的载体,并且其还包含一个或多个编码或包含用于在病毒复制细胞中病毒表达的组分的多核苷酸区。
肽:如本文所用,“肽”的长度小于或等于50个氨基酸,例如约5、10、15、20、25、30、35、40、45或50个氨基酸长。
药学上可接受的:本文中使用的短语“药学上可接受的”在本文中用于表示这样的化合物、物质、组合物和/或剂型:在合理的医学判断范围内,其适用于接触人类和动物的组织,而没有过度的毒性、刺激、变应性应答或其它问题或并发症,与合理的收益/风险比相称。
药学上可接受的赋形剂:如本文所用,短语“药学上可接受的赋形剂”是指除本文所述化合物以外的任何成分(例如,能够悬浮或溶解活性化合物的溶媒),并且具有在患者中基本上无毒且无炎症性的性质。赋形剂可以包括例如:抗粘剂、抗氧化剂、粘合剂、包衣、压缩助剂、崩解剂、染料(着色剂)、润肤剂、乳化剂、填充剂(稀释剂)、成膜剂或包衣剂、调味剂、香料、助流剂(流动增强剂)、润滑剂、防腐剂、印刷油墨、吸附剂、悬浮剂或分散剂、甜味剂和水合水。示例性的赋形剂包括但不限于:丁基羟基甲苯(bht)、碳酸钙、磷酸氢钙、硬脂酸钙、交联羧甲基纤维素、交联聚乙烯吡咯烷酮、柠檬酸、交聚维酮、半胱氨酸、乙基纤维素、明胶、羟丙基纤维素、羟丙基甲基纤维素、乳糖、硬脂酸镁、麦芽糖醇、甘露醇、甲硫氨酸、甲基纤维素、对羟基苯甲酸甲酯、微晶纤维素、聚乙二醇、聚乙烯吡咯烷酮、聚维酮、预胶化淀粉、对羟基苯甲酸丙酯、视黄醇棕榈酸酯、虫胶、二氧化硅、羧甲基纤维素钠、柠檬酸钠、淀粉羟乙酸钠、山梨醇、淀粉(玉米)、硬脂酸、蔗糖、滑石、二氧化钛、维生素a、维生素e、维生素c和木糖醇。
药学上可接受的盐:本公开还可包含本文所述的化合物的药学上可接受的盐。本文中使用的“药学上可接受的盐”表示公开的化合物的衍生物,其中通过将现有的酸或碱部分转化成其盐形式(例如,通过使游离碱基团与合适的有机酸反应)来修饰母体化合物。药学上可接受的盐的实例包括但不限于碱性残基如胺的无机酸盐或有机酸盐;酸性残基如羧酸的碱盐或有机盐;等。代表性的酸加成盐包括乙酸盐、乙酸、己二酸盐、海藻酸盐、抗坏血酸盐、天冬氨酸盐、苯磺酸盐、苯磺酸、苯甲酸盐、硫酸氢盐、硼酸盐、丁酸盐、樟脑酸盐、樟脑磺酸盐、柠檬酸盐、环戊烷丙酸盐、二葡萄糖酸盐、十二烷基硫酸盐、乙磺酸盐、富马酸盐、葡庚糖酸盐、甘油磷酸盐、半硫酸盐、庚糖酸盐、己酸盐、氢溴酸盐、盐酸盐、氢碘酸盐、2-羟基-乙磺酸盐、乳糖酸盐、乳酸盐、月桂酸盐、月桂基硫酸盐、苹果酸盐、马来酸盐、丙二酸盐、甲磺酸盐、2-萘磺酸盐、烟酸盐、硝酸盐、油酸盐、草酸盐、棕榈酸盐、双羟萘酸盐、果胶酸盐、过硫酸盐、3-苯基丙酸盐、磷酸盐、苦味酸盐、新戊酸盐、丙酸盐、硬脂酸盐、琥珀酸盐、硫酸盐、酒石酸盐、硫氰酸盐、甲苯磺酸盐、十一烷酸盐、戊酸盐等。代表性的碱金属或碱土金属盐包括钠、锂、钾、钙、镁等,以及无毒的铵、季铵和胺阳离子,包括但不限于铵、四甲基铵、四乙基铵、甲胺、二甲胺、三甲胺、三乙胺、乙胺等。本公开的药学上可接受的盐包括形成的母体化合物的常规无毒盐,例如,从无毒的无机或有机酸形成。本公开的药学上可接受的盐可以通过常规化学方法从含有碱性或酸性部分的母体化合物合成。通常,通过使这些化合物的游离酸或碱形式与化学计量的量的适当的碱或酸在水或有机溶剂或这两者的混合物中反应,可以制备这样的盐;通常,可使用非水性介质如醚、乙酸乙酯、乙醇、异丙醇或乙腈。合适的盐的列表可在以下文献中找到:remington’spharmaceuticalsciences,17thed.,mackpublishingcompany,easton,pa.,1985,p.1418,pharmaceuticalsalts:properties,selection,anduse,p.h.stahl和c.g.wermuth(eds.),wiley-vch,2008,以及berge等人,journalofpharmaceuticalscience,66,1-19(1977),其各自的内容通过引用整体并入本文,只要其与本公开不冲突。
药学上可接受的溶剂合物:本文中使用的术语“药学上可接受的溶剂合物”是指这样的本公开的化合物,其中合适的溶剂的分子掺入晶格中。合适的溶剂在施用的剂量是生理学上可耐受的。通过从包括有机溶剂、水或其混合物的溶液结晶、重结晶或沉淀,可以制备溶剂合物。合适的溶剂的实例是乙醇、水(例如,一水合物、二水合物和三水合物)、n-甲基吡咯烷酮(nmp)、二甲亚砜(dmso)、n,n’-二甲基甲酰胺(dmf)、n,n’-二甲基乙酰胺(dmac)、1,3-二甲基-2-咪唑啉酮(dmeu)、1,3-二甲基-3,4,5,6-四氢-2-(1h)-嘧啶酮(dmpu)、乙腈(acn)、丙二醇、乙酸乙酯、苯甲醇、2-吡咯烷酮、苯甲酸苄酯等。当水是溶剂时,溶剂合物被称作“水合物”。
药代动力学:如本文所用,“药代动力学”是指分子或化合物涉及确定施用于活生物体的物质的命运时的任何一种或多种性质。药代动力学分为几个方面,包括吸收、分布、代谢和排泄的程度和速率。通常将其称为adme,其中:(a)吸收是物质进入血液循环的过程;(d)分布是指物质在人体的所有液体和组织中的分散或扩散;(m)代谢(或生物转化)是母体化合物不可逆地转化为子代代谢物;和(e)排泄(或消除)是指物质从体内消除。在极少数情况下,某些药物会不可逆地积聚在人体组织中。
物理化学的:如本文所用,“物理化学的”是指物理或化学性质或与之有关。
预防:如本文所用,术语“预防(preventing)”或“预防(prevention)”是指部分或完全延迟感染、疾病、病症和/或病况的发作;部分或完全延迟特定感染、疾病、病症和/或病况的一种或多种症状、特征或临床表现的发作;部分或完全延迟特定感染、疾病、病症和/或病况的一种或多种症状、特征或表现的发作;部分或完全延迟感染、特定疾病、病症和/或病包括的进展;和/或降低发生与感染、疾病、病症和/或病况相关的病理的风险。
增殖:如本文所用,术语“增殖”是指生长、扩增或增加,或引起迅速生长、扩增或增加。“增殖性”是指具有增殖的能力。“抗增殖性”是指具有与增殖性质相反或不适当的性质。
预防的:如本文所用,“预防的”是指用于预防疾病传播的治疗或作用过程。
预防(prophylaxis):如本文所用,“预防”是指为维持健康和预防疾病传播而采取的措施。
目标蛋白:如本文所用,术语“目标蛋白”或“所需蛋白”包括本文提供的蛋白及其片段、突变体、变体和改变。
近端:如本文所用,术语“近端”是指位于更靠近中心或目标点或区域。
纯化的:如本文所用,“纯化”、“纯化的”、“纯化”是指从不需要的组分、材料杂质、混合物或混杂物(imperfection)中使之基本上纯化或澄清。“纯化的”是指纯净的状态。“纯化”是指进行纯化的过程。
区域(region):如本文所用,术语“区域”是指区(zone)或一般区域(generalarea)。在某些实施方案中,当涉及蛋白或蛋白模块时,区域可包含沿着蛋白或蛋白模块的氨基酸的线性序列,或可包含三维区域、表位和/或表位簇。在某些实施方案中,区域包含末端区域。如本文所用,术语“末端区域”是指位于给定试剂的端点或末端的区域。当涉及蛋白时,末端区域可包含n末端和/或c末端。n末端是指包含具有游离氨基的氨基酸的蛋白的末端。c末端是指包含具有游离羧基的氨基酸的蛋白的末端。n-和/或c-末端区域可因此包含n-和/或c-末端以及周围的氨基酸。在某些实施方案中,n-和/或c-末端区域包含约3个氨基酸至约30个氨基酸、约5个氨基酸至约40个氨基酸、约10个氨基酸至约50个氨基酸、约20个氨基酸至约100个氨基酸和/或至少100个氨基酸。在某些实施方案中,n末端区域可包含任何长度的氨基酸,其包含n末端,但不包含c末端。在某些实施方案中,c末端区域可包含任何长度的氨基酸,其包含c末端,但不包含n末端。
在某些实施方案中,当提及多核苷酸时,区域可以包含沿着多核苷酸的核酸的线性序列,或者可以包含三维区域、二级结构或三级结构。在某些实施方案中,区域包含末端区域。如本文所用,术语“末端区域”是指位于给定试剂的端点或末端的区域。当提及多核苷酸时,末端区域可以包含5'和3'末端。5'末端是指包含具有游离磷酸基团的核酸的多核苷酸的末端。3'末端是指包含具有游离羟基的核酸的多核苷酸的末端。5'和3'区域可因此包含5'和3'末端以及周围的核酸。在某些实施方案中,5'和3'末端区域包含约9个核酸至约90个核酸、约15个核酸至约120个核酸、约30个核酸至约150个核酸、约60个核酸至约300个核酸和/或至少300个核酸。在某些实施方案中,5'区域可以包含任何长度的核酸,其包含5'末端,但不包含3'末端。在某些实施方案中,3'区域可以包含任何长度的核酸,其包含3'末端,但不包含5'末端。
rna或rna分子:本文使用的术语“rna”或“rna分子”或“核糖核酸分子”表示核糖核苷酸的聚合物;术语“dna”或“dna分子”或“脱氧核糖核酸分子”表示脱氧核糖核苷酸的聚合物。可以天然地合成(例如,分别通过dna复制和dna的转录)或化学地合成dna和rna。dna和rna可以是单链的(即,分别是ssrna或ssdna)或多链的(例如,双链的,即,分别是dsrna和dsdna)。本文中使用的术语“mrna”或“信使rna”表示编码一个或多个多肽链的氨基酸序列的单链rna。
rna干扰或rnai:本文使用的术语“rna干扰”或“rnai”表示由rna分子介导的序列特异性的调节机制,其导致对应的蛋白编码基因的表达的抑制或干扰或“沉默”。rnai已经在许多类型的生物中观察到,包含植物、动物和真菌。rnai天然地发生在细胞中以除去外来rna(例如,病毒rna)。天然rnai通过从游离dsrna切割的片段进行,其将降解机制引导至其它类似的rna序列。rnai由rna诱导的沉默复合物(risc)控制,且由细胞质中的短/小dsrna分子起始,其中它们与催化性的risc组分argonaute相互作用。可以将dsrna分子外源地引入细胞中。外源dsrna通过激活核糖核酸酶蛋白dicer而起始rnai,所述dicer结合和切割dsrna以产生21-25个碱基对的双链片段,其在每个末端上具有几个未配对的突出端碱基。这些短双链片段被称作小干扰rna(sirna)。
样品:如本文所用,术语“样品”或“生物样品”是指其组织、细胞或组成部分(例如体液,包括但不限于血液、粘液、淋巴液、滑液、脑脊髓液、唾液、羊水、羊脐血、尿液、阴道液和精液)的子集。样品还可以包括从整个生物体或其组织、细胞或组成部分的子集或其一部分或部分制备的匀浆、裂解物或提取物,包括但不限于例如血浆、血清、脊髓液、淋巴液,皮肤、呼吸道、肠道和泌尿生殖道的外部区域、眼泪、唾液、乳汁、血细胞、肿瘤、器官。样品还指可能包含细胞成分(例如蛋白或核酸分子)的培养基,例如营养肉汤或胶。
自互补病毒颗粒:如本文所用,“自互补病毒颗粒”是由至少两种组分(蛋白衣壳和包封在所述衣壳内的编码自身互补基因组的多核苷酸序列)组成的颗粒。
有义链:本文使用的术语sirna分子的“有义链”或“第二条链”或“过客链”表示与反义链或第一链互补的链。sirna分子的反义链和有义链发生杂交以形成双链体结构。本文中使用的“sirna双链体”包括与为了沉默而靶向的基因的mrna的约10-50个核苷酸的段具有足够互补性的sirna链和具有足够互补性以与另一条sirna链形成双链体的sirna链。
短干扰rna或sirna:本文使用的术语“短干扰rna”、“小干扰rna”或“sirna”表示能够指导或介导rnai的包含约5-60个核苷酸(或核苷酸类似物)的rna分子(或rna类似物)。在某些实施方案中,sirna分子包含约15-30个核苷酸或核苷酸类似物,诸如约16-25个核苷酸(或核苷酸类似物),约18-23个核苷酸(或核苷酸类似物),约19-22个核苷酸(或核苷酸类似物)(例如,19、20、21或22个核苷酸或核苷酸类似物),约19-25个核苷酸(或核苷酸类似物),和约19-24个核苷酸(或核苷酸类似物)。术语“短”sirna表示包含5-23个核苷酸、例如21个核苷酸(或核苷酸类似物)(例如,19、20、21或22个核苷酸)的sirna。术语“长”sirna表示包含24-60个核苷酸、例如约24-25个核苷酸(例如,23、24、25或26个核苷酸)的sirna。在某些情况下,短sirna可以包含少于19个核苷酸,例如,16、17或18个核苷酸,或少至5个核苷酸,前提条件是,较短的sirna保留介导rnai的能力。同样地,在某些情况下,长sirna可以包含超过26个核苷酸,例如,27、28、29、30、35、40、45、50、55或甚至60个核苷酸,前提条件是,较长的sirna保留介导rnai或翻译抑制的能力,无需进一步加工(例如,酶促加工)成短sirna。sirna可以是单链rna分子(ss-sirna)或包含有义链和反义链的双链rna分子(ds-sirna),所述有义链和反义链杂交以形成被称作sirna双链体的双链体结构。
信号序列:如本文所用,短语“信号序列”是指可以指导蛋白的运输或定位的序列。
单个单位剂量:如本文所用,“单个单位剂量”是指以一剂/一次/单途径/单接触点(即单次施用事件)施用的任何治疗剂的剂量。在某些实施方案中,提供单个单位剂量作为离散剂型(例如,片剂、胶囊、贴剂、载注射器、小瓶等)。
相似性:如本文所用,术语“相似性”是指聚合物分子之间例如多核苷酸分子(例如dna分子和/或rna分子)之间和/或多肽分子之间的整体相关性。聚合物分子彼此的相似性百分比的计算可以与计算同一性百分比相同的方式进行,除了相似性百分比的计算考虑了本领域已知的保守取代。
分剂量:如本文所用,“分剂量”是将单个单位剂量或每日总剂量分成两个或更多个剂量。
稳定的:如本文所用,“稳定的”是指足够强健以从反应混合物中分离到有用纯度时存活,并且在某些实施方案中,能够配制成有效治疗剂的化合物。
稳定化的:如本文所用,术语“稳定化(stabilize)”、“稳定化的(stabilized)”、“稳定化的区域(stabilizedregion)”是指使得或变得稳定。
受试者:本文使用的术语“受试者”或“患者”表示可以给其施用根据本公开的组合物的任何生物,例如,为了实验、诊断、预防和/或治疗目的。典型受试者包括动物(例如,哺乳动物诸如小鼠、大鼠、兔、非人灵长类动物和人类)和/或植物。
基本上:如本文所用,术语“基本上”是指展现出目标特征或特性的全部或接近全部程度或度的定性条件。生物学领域的普通技术人员将理解,生物学和化学现象很少(如果有的话)完全完成和/或进行到完全或达到或避免绝对结果。因此,本文中使用术语“基本上”来捕获许多生物学和化学现象中固有的潜在的完整性缺乏。
基本上相等的:如在本文中所使用的,当它涉及剂量之间的倍数差时,该术语是指正/负2%。
基本上同时的:如本文所用并且在涉及多个剂量时,该术语表示在2秒内。
患有:“患有”疾病、病症和/或病况的个体已被诊断出或表现出疾病、病症和/或病况的一种或多种症状。
易患:“易患”疾病、病症和/或病状的个体尚未被诊断为和/或未表现出该疾病、病症和/或病况的症状但具有发展疾病或其症状的倾向。在某些实施方案中,易患疾病、病症和/或病况(例如,癌症)的个体可以以下一种或多种为特征:(1)与疾病、病症和/或病况的发展相关的遗传突变;(2)与疾病、病症和/或病况发展相关的遗传多态性;(3)与疾病、病症和/或病况相关的蛋白和/或核酸的表达和/或活性的增加和/或降低;(4)与疾病、病症和/或病况发展相关的习惯和/或生活方式;(5)疾病、病症和/或病况的家族病史;(6)暴露于和/或感染与疾病、病症和/或病况发展相关的微生物。在某些实施方案中,易患疾病、病症和/或病况的个体将发展疾病、病症和/或病况。在某些实施方案中,易患疾病、病症和/或病况的个体将不会发展疾病、病症和/或病况。
持续释放:如本文所用,术语“持续释放”是指符合特定时间段内的释放速率的药物组合物或化合物的释放曲线。
合成的:术语“合成的”是指通过人手生产、制备和/或制造。本公开的多核苷酸或多肽或其它分子的合成可以是化学的或酶促的。
靶向:如本文所用,“靶向”是指设计和选择将与靶核酸杂交并诱导所需作用的核酸序列的过程。
靶细胞:如本文所用,“靶细胞”是指任何一种或多种目标细胞。可以在体外、体内、原位或生物体的组织或器官中发现细胞。该生物可以是动物,例如哺乳动物、人或人类患者。
末端区:如本文所用,术语“末端区”是指连接的核苷或氨基酸(分别为多核苷酸或多肽)的区的5’或3’端上的区。
末端优化的:术语“末端优化的”在指核酸时表示核酸的末端区相对于天然或野生型末端区以某种方式得到改善,例如密码子优化。
治疗剂:术语“治疗剂”是指当施用于受试者时具有治疗、诊断和/或预防作用和/或引起期望的生物学和/或药理作用的任何药剂。
治疗有效量:本文使用的术语“治疗有效量”是指要递送的药剂(例如,核酸、药物、治疗剂、诊断剂、预防剂等)的量,当施用给患有或易患感染、疾病、病症和/或病况的受试者时,所述量足以治疗、诊断、预防所述感染、疾病、病症和/或病况,改善其症状,和/或延迟其发作。在某些实施方案中,以单剂量提供治疗有效量。在某些实施方案中,以包括多个剂量的剂量方案施用治疗有效量。本领域技术人员将理解,在某些实施方案中,如果单位剂型包含当作为这种剂量方案的一部分施用时有效的量,则可以认为该单位剂型包含治疗有效量的特定试剂或实体。
治疗有效的结果:如本文所用,术语“治疗有效的结果”是指在患有或易患感染、疾病、病症和/或病况的受试者中足以治疗、诊断、预防所述感染、疾病、病症和/或病况,改善其症状,和/或延迟其发作的结果。
总日剂量:如本文所用,“总日剂量”是在24小时内给定或处方的剂量。它可以单个单位剂量施用。
转染:如本文所用,术语“转染”是指将外源核酸引入细胞的方法。转染的方法包含但不限于化学方法、物理处理和阳离子脂质或混合物。
治疗:如本文所用,术语“治疗”是指部分或完全缓解、改善、改进、解除特定的感染、疾病、病症和/或病况的一种或多种症状或特征、延迟其发作、抑制其进展、降低其严重程度和/或降低其发生率。例如,“治疗”癌症可以指抑制肿瘤的存活、生长和/或扩散。出于降低发展成与疾病、病症和/或病况相关的病理的风险,治疗可施用于不表现出疾病、病症和/或病况的体征的受试者和/或施用于仅表现出疾病、病症和/或病况的早期体征的受试者。
未修饰的:如本文所用,“未修饰的”是指在以任何方式改变之前的任何物质、化合物或分子。未修饰的可能但并非总是指生物分子的野生型或初始形式。分子可以经历一系列修饰,由此每个修饰的分子可以用作随后修饰的“未修饰的”起始分子。
载体:如本文所用,“载体”是运输、转导或以其他方式充当异源分子的运载体的任何分子或部分。本公开的载体可以重组产生,并且可以基于和/或可以包含腺相关病毒(aav)亲本或参考序列。这样的亲本或参考aav序列可以用作工程化载体的原始、第二、第三或后续序列。在非限制性实例中,此类亲本或参考aav序列可以包含以下序列中的任何一个或多个:编码多肽或多个多肽的多核苷酸序列,该序列可以是野生型或从野生型修饰的,以及该序列可以编码蛋白、蛋白结构域或蛋白的一个或多个亚基的全长或部分序列;包含调节性或调控性核酸的多核苷酸,该序列可以是野生型或从野生型修饰的;以及可以从或不从野生型序列修饰的转基因。这些aav序列可以用作一个或多个密码子(在核酸水平)或氨基酸(在多肽水平)的“供体”序列或一个或多个密码子(在核酸水平)或氨基酸(在多肽水平上)的“受体”序列。
病毒基因组:如本文所用,“病毒基因组”或“载体基因组”是指包封在aav颗粒中的核酸序列。
iv.等同和范围
本领域技术人员会认识到或使用不超过例行实验能够确定根据本文描述的本公开的具体实施方案的许多等同方案。本公开的范围无意限于上面的描述,而是如所附权利要求所述。
在权利要求中,诸如“一个”、“一种”和“所述”的冠词可以是指一个/种或超过一个/种,除非指示相反情形或以其它方式从上下文显而易见。如果群体成员中的一个、超过一个或所有成员存在于指定的产品或过程中、在指定的产品或过程中使用或以其它方式与指定的产品或过程相关,则认为满足在一组的一个或多个成员之间包括“或”的权利要求或描述,除非指示相反情形或以其它方式从上下文显而易见。本公开包括这样的实施方案,其中所述组的刚好一个成员存在于给定产品或方法中、用于给定产品或方法中或者以其它方式与给定产品或方法相关。本公开包括这样的实施方案,其中所述组成员中的超过一个或全部都存在于给定产品或方法中、用于给定产品或方法中或者以其它方式与给定产品或方法相关。
还应当指出,术语“包含”意图是开放式的,并且允许但不要求包括额外的要素或步骤。因此,当在本文中使用术语“包含”时,还包括和公开术语“由……组成”。
在给出范围的情况下,包括端点。此外,应当理解,除非另外指示或以其它方式从上下文和本领域普通技术人员的理解显而易见,以范围表述的数值在本公开的不同实施方案中可呈现处于所述范围内的任何具体数值或子范围,其精确至该范围的下限单位的十分之一,除非上下文另外清楚地指明。
另外,应当理解,可以从任意一项或多项权利要求中明确地排除落在现有技术内的本公开的任何特定实施方案。因为这样的实施方案被视为本领域普通技术人员已知的,所以可以排除它们,即使所述排除并未在本文中明确地阐明。可以因为任何原因从任意一项或多项权利要求排除本公开的组合物的任何特定实施方案(例如,任何抗生素、治疗剂或活性成分;任何生产方法;任何使用方法等),无论是否与现有技术的存在相关。
应当理解,已经使用的词语是描述而不是限制的词语,并且可以在所附权利要求的范围内做出变化,而不背离在它的更宽方面的本公开的真实范围和精神。
尽管已经就几个描述的实施方案而言相当详尽地且相当特定地描述了本公开,但是无意将本发明限于任何这样的细节或实施方案或任何具体实施方案,而是应当参考所附权利要求来解释本发明,从而考虑到现有技术提供这类权利要求的最广泛的可能解释,并因此有效地包括本公开的预期范围。
本文提及的所有出版物、专利申请、专利和其它参考文献都通过引用整体并入本文。如果发生冲突,以本说明书(包括定义)为准。另外,章节标题、材料、方法和实施例仅是说明性的,而非意图限制。
实施例
实施例1.异常衣壳蛋白裂解产物的鉴定
使用商业杆状病毒表达载体(bev)系统在sf9昆虫细胞中表达具有7个氨基酸的肽插入物的重组合成aav9变体衣壳(seqidno:5)。
经过处理和cscl梯度纯化后,将aav衣壳蛋白在sds-page凝胶上分级,并对蛋白进行银染。除了vp1、vp2和vp3外,aav药物物质中还出现了意想不到的18kda、43kda和52kda蛋白,如图1所示。
这些额外的蛋白的丰度随着纯化批次以及储存时间的变化而变化。因此损害了源自bev/sf9表达的aav药物物质的稳定性。
当通过质粒转染在哺乳动物细胞(hek293)中表达相同的aav衣壳构建体时,未见到任何意想不到的蛋白。但是,如果基因疗法应用需要大量的aav载体,则哺乳动物细胞转染不是aav生产的理想选择。
最小的18kda蛋白的n端测序表明它是vp1、vp2和vp3共有的c末端肽片段,如图2所示。
在银染的sds-page凝胶中观察到的52kda和43kda的肽片段与具有从其共同的c末端裂解的18kda的vp2和vp3蛋白相关。
由vp1的类似裂解预测的67kda的肽片段是观察不到的,因为它将与70kda的未裂解的vp2蛋白共迁移。我们无法生产100%转化为裂解产物的aav样品,因此我们无法确认vp1被裂解。这些数据表明,蛋白酶从vp1、vp2和vp3的c末端切割18kda的肽。
当使用抗vp1、vp2和vp3的共同c末端区的抗体通过抗体亲和柱纯化时,western印记免疫检测显示仅形成了18kda的c末端裂解产物。然而在丝氨酸/半胱氨酸蛋白酶抑制剂混合物的存在下,不再检测到18kda的肽。这些数据表明丝氨酸和/或半胱氨酸蛋白酶参与18kda的c末端裂解产物的出现。
实施例2.蛋白酶鉴定
大多数商业bev是基于杆状病毒acmnpv或密切相关的杆状病毒bmnpv。杆状病毒v-cath基因编码组织蛋白酶样蛋白酶(病毒组织蛋白酶)。v-cath基因与杆状病毒几丁质酶基因chia相连。除少数例外,几乎所有~80个测序的杆状病毒基因组都具有v-cath和chia基因。见图3,v-cath基因的比对。
考虑到保守性,可以得出结论,v-cath和chia是必需的杆状病毒基因,并且应在用于制备重组蛋白的杆状病毒表达载体(bev)中完整保留。
在发现v-cath基因后不久证明了,v-cath蛋白酶在鳞翅目宿主中的杆状病毒感染的组织液化和变黑病理中起作用(slack等人,1995j.genvirol.1091-8)。
在细胞培养的杆状病毒感染的昆虫细胞(biic)中,v-cath的存在与细胞裂解和感染结束时多面体形包含体的释放相关。
然而,尽管在感染结束时v-cath在增强来自宿主的杆状病毒包含体的分散中起作用,但是认为在昆虫细胞培养物中通过bev的重组蛋白表达不需要v-cath。
进行筛选以鉴定被利用的bev系统中的v-cath基因。虽然用于在sf9细胞中表达aav衣壳蛋白的bev系统常见地用于重组蛋白表达,尚无法获得bev的全长序列来确定v-cath是否仍存在于bev系统中。
进行pcr筛选以确定在野生型acmnpv杆状病毒中是否发现了v-cath基因。三种不同的pcr筛选证实bev系统中的v-cath基因座与根据wtacmnpv预测的相比没有变化。见图4。
实施例3.bev系统中v-cath缺失的产生和影响
研究了缺失v-cath基因的bev构建体,包括v-cath基因缺失对生产重组的合成aav衣壳构建体的可能影响。还研究了相应的蛋白裂解、稳定性、细胞活力、细胞直径、细胞外观和生产效率。
用120ulaavbev1(标准bev表达载体)或aavbev2(v-cath缺失bev表达载体,其包含seqidno:2的构建体)的杆状病毒感染的昆虫细胞(biic)感染含120ml浓度为1.4x106个细胞/ml的sf9细胞的摇瓶。还包括未感染的sf9细胞的对照瓶。在27℃以130rpm温育摇瓶。每24小时收集细胞样品,持续6天。
细胞活力
通过将细胞与0.2%w/v锥虫蓝染料混合并在nexcelomcellometerautot4上计数来确定细胞活力。在感染过程中,标准bev1感染和v-cath缺失的bev2感染产生了非常相似的sf9细胞活力谱(图5)和细胞计数曲线(图6)。
细胞直径
使用nexcelomcellometerautot4测定细胞直径。直到感染后96h,标准bev1感染和v-cath缺失的bev2感染产生了非常相似的sf9细胞直径(图7)。在感染后很后期的时候直径才是不同的。
细胞外观
用锥虫蓝染色后,未感染的sf9细胞、aav-bev1和aav-bev2感染的sf9细胞的显微镜图像在bev1和bev2细胞之间显示出相似的形态。
蛋白酶活性
对病毒感染的sf9细胞裂解物进行了偶氮酪蛋白蛋白酶活性测定以检测v-cath,该测定改编自slack等人2004,jgenvirol.211-9进行的测定。将收集的sf9细胞在pbs(ph7.4)中稀释至5x108个细胞/ml,然后在pbs中以50ul的体积进行系列稀释。将稀释液与50ul蛋白酶测定缓冲液c(6.7mm柠檬酸;10mm柠檬酸钠;0.25%tritonx-100;46mmnacl;3.3mm磷酸盐;0.9mmkcl;3.3mmedta)合并。然后将该混合物与50ul0.17%w/v的偶氮酪蛋白合并。缓冲液c混合物的最终ph经测量为ph5.87。
将样品在37℃的96孔u型底板中温育14h,无需摇动。将50ul体积的20%v/v三氯乙酸加入到每个孔中,混合后,将96孔板以2100xg离心5分钟。将150ul的上清液体积转移到平底96孔板中,并在分光光度计上测量od405。
未感染的sf9细胞和v-cath缺失的bev2感染的细胞具有相似的抗偶氮酪蛋白底物的蛋白酶活性水平。相比之下,随着感染的进行,被标准bev1感染的sf9细胞显示出升高的抗偶氮酪蛋白底物的蛋白酶活性水平。见图8。
还用缓冲液a代替早前描述的缓冲液c进行偶氮酪蛋白蛋白酶测定。缓冲液a(200mm精氨酸;0.25%tritonx-100;46mmnacl;3.3mm磷酸盐;0.9mmkcl)。然后将该混合物与50ul的0.17%w/v偶氮酪蛋白合并。测量缓冲液a的ph为ph6.89。
发现缓冲液a增加了与标准bev1sf9细胞裂解物相关的蛋白酶活性。甚至在存在缓冲液a的情况下,v-cath缺失bev2sf9细胞裂解物也保留低水平的蛋白酶活性。见图9。
由于先前的研究表明v-cath偏爱酸性条件,因此在中性ph下见到v-cath相关的蛋白酶活性增加是预料不到的。
尽管不希望受到理论的束缚,但是胺基团的存在可能充当底物的溶剂。发明人先前已经表明,在缓冲液中存在尿素的情况下,v-cath对偶氮酪蛋白底物的活性显著增强(slack等人,1995)。因此,精氨酸缓冲液可能以类似的方式起作用,即,它改变了底物蛋白或衣壳的构象并使残基暴露于蛋白酶。
因此,我们确定精氨酸提高了aav纯化的产量。
western印迹-降解产物的消除
在缓冲液a裂解缓冲液中处理由标准bev1感染和v-cath缺失的bev2感染表达的aav,然后使用抗vp1、vp2、vp3抗体(americanresearchproducts,产品编号:03-61058)通过western免疫印迹来检测aav衣壳。v-cath的缺失消除了降解产物。见图10a和图10b。
结论
本公开提供了可用于解决在sf9/杆状病毒系统中生产的aav衣壳的杆状病毒编码的蛋白酶降解的构建体、系统和方法。添加蛋白酶抑制剂不是好的解决方案,因为它们通常很昂贵,并且残留物可能在下游治疗产品中具有毒性作用。
此处的证据表明,v-cath是从杆状病毒表达的aav药物物质中的vp1、vp2和vp3切割18kdac末端片段的蛋白酶。
在bev系统中存在v-cath的另一个潜在问题是对aav药物物质长期稳定性的潜在损害,因为蛋白酶也可与其他蛋白结合,并且v-cath可能与一些aav共纯化。
序列表
<110>沃雅戈治疗公司(voyagertherapeutics,inc.)
<120>在杆状病毒/sf9系统中大规模生产raav的表达载体
<130>2057.1514pct
<140>pct/us2019/xxxxxx
<141>2019-10-15
<150>62/745,703
<151>2018-10-15
<160>5
<170>patentinversion3.5
<210>1
<211>26
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>来源(source)
<223>/备注=“人工序列的描述:合成的寡核苷酸”
<400>1
taactataacggtcctaaggtagcga26
<210>2
<211>5781
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>来源(source)
<223>/备注=“人工序列的描述:合成的多核苷酸”
<400>2
atgatgtcatcatcacaaataatagtatgtaataaaataaacatatttgtgtgtaaatat60
aatttattacaaataaattttacattgaatcaatctgtcttcgtgtttgttgtaaggtct120
tcgaatcttgtgtttcagcccctcgggatggtcaaaatgcgccgtagtaattgttaatgg180
atctttcaacgattttttgcccatggcgagtgtgacaaacgcggccacgacaaacagcag240
gataatcagtttcatggtgttctatattcgacaatatatgggtcgcttctaaatcacctt300
gtccccaaaagcctcttttatagttttttagaacacgttgtgtattccaacagtaattgt360
tccatctctttcaacagccattcagcatccggtcgttgactgtaatcatgctgaattaat420
ttacaaacaatttcggtcaatttaggatggccttgggataaacttgccggcatttgctgt480
acattgtttctaaagttagttagcgtagtttcgcgttccaaagcagtcttgaagggcatt540
atcaattcgaataaaacaatgcccaaactatacatgtcatttttgggggtgtacactttt600
ttgatttgttctggtgcagcgtacaaagttatattttgagggttgtttttgataaacgtt660
ttgtatagactgccaaacatgccgcccacatacaaatcaaagtcgggcccagtcatgaaa720
atatcttcgggattaatattgtggtgcacgatatttacggaatgaatcgctttcacggcg780
ctcaccaaatcaacaaacttgctaatataaaagccaaaatccgccggaactttaatgttg840
gtctttgcaaaagtttgcaaattgcgttgtttcaaatagtcgctcaacatgtactcgttt900
agaggcgacgcaaaatatatgcggtgctgccgcggattcaaataaaccaattgttcgggt960
ttcatggtatacagttaagtgttaacgcgtcactaaattcagacacgagcgcacgcccta1020
tatacatacaatttatcgcacaagatgcttaacgcgatctgtttataaactaaaacgcac1080
tgcaataaattttagcaagcatttgtatttaatcaatcgaaccgtgcactgatataagaa1140
ttaaaaatgggtttgtttgcgtgttgcacaaaatacacaaggctgtcgaccgacacaaaa1200
atgaagtttccctatgttgcgttgtcgtacatcaacgtgacgctgtgcacctacaccgcc1260
atgttggtgggatacatggtaacattcaatgactccagcgaattgaaatatttacaatac1320
tggttgctgttgtcgtttttgatgtccgtggtgctaaacgctccgactctgtggacgatg1380
ctcaaaaccacagaagcccatgaagtaatttacgaaatgaagctgttccacgccatgtac1440
tttagtaacgtgctgttgaattatgtggtgtttttggacaatcaaatgggtacaaatttt1500
gtttttgttaacaatttaattcactgttgtgtactttttatgatatttgttgaattgctt1560
atcctgttgggccacacaatgggcacgtacacggattatcaatatgtcaaatcgtgttat1620
atggttatattgtttgtttcagttatgagtgttactattgttatgggtttagagtgtttg1680
aaaacgaaactaattgataacagtttgatgtttaacgcgtttgtgtgcgctttgtacatt1740
gtgattgcaataatgtggtctttaaaaaataatttgactagttattacgtttcaaattta1800
caaagtattcaagttgttccgttttcatacaacgatccgccgccaccgttctctaacatt1860
gtaatggatgacataaaaaataaaaaataatttataaaaatgttttttattctttcacaa1920
ttctgtaaattctaaacaaaaaatataaatacaaacttattatgttgtcgtctaaataaa1980
catcaatttgtaaatctggacacctattcatatcattgatattacagtctactatacaac2040
aattaaaactaaccaaattatctttacaacaattaaagcaattaaaacaatttaaataat2100
cttcattgtcgtcgtataagtttatttgcactgtagacggtgttacacagcgatccattc2160
gacgttcgtgttcgatcaactttctcgccaacttgtaccataaaaattgtttggacaaaa2220
agttttccaacaatggtaacggccaattcaacgtgacgatgcgcacgtcctcgggtatgc2280
atttgttaaaaaacacacagctcgctttaccaaacgaaagcaaaggtactaaatatggcg2340
ccattggctgatttgttattccaagataattacaaataaactgatccgtcgtggggtgat2400
aactggcaggtgtcagctttaaataatcttcaacgttgttgtcgcgcaaaagtctgcatt2460
ttacacgcgttgttaatcccacgacttttgcatgtaaaatcggatccaaatactgcagaa2520
tcgtgtctataatttctaatggtaaacgtatgcgttttgctcgtgggcgctttgtaacgc2580
tcgacatcctaataacaactaacacaaaactaaaatgatactcaatatattgcttttaca2640
gttcatctttaggtttaaactgtgcgtttatcgcgttgagcaagtcgccgttatcggcat2700
caatctcccaagcaaacaggccgcccaatttatttcggtcgacatatttaacttttccta2760
acacagagtcgacgctgtcaaacgaaatcaaatcacctttacttttatcgaaaacgtacg2820
acgcttgagcggcgctgtcaaacgtgtacacataattgttgagatctttttgaatttgac2880
gataatctacaacaccgtcctcccacgtgcccgaccccggcccgttgccagtgccggaaa2940
aatagttgtcattcgtataatttgttacgccggtccagccgcggccgtacatggcgacgc3000
ccacaattattttgttgggatcgacgccttgtttcagtaacgcatcgacagcgtagtgtg3060
tagtgtatagctcttccgagttccaacttggcgcgtagactgttgtttggtagcccaaat3120
ccgtgtttgaccaagcccctttaaaatcgtaactcatgagaaatattttgcctaatgact3180
tttgcgcttcggcgtagtttaccacggcaatcttgtcgtaacccgcgcttatagcgcttg3240
ttaattcgtaaaccctgccggtttgcgcttcgaggtcgtctagcattgcgcgcagctcct3300
ccaacaacaaaatgtatgttttggcgtcaccgtccgcatcgcccaacgacgggttagccc3360
ctttgccgcccggaaactcccaatcgatgtctacaccgtcaaagaatttccacacttgca3420
gaaattccttaaccgaatctacaaaaacgtttcttttttcaacatcgtgggtacctaact3480
ataacggtcctaaggtagcgagacctttaattcaacccaacacaatatattatagttaaa3540
taagaattattatcaaatcatttgtatattaattaaaatactatactgtaaattacattt3600
tatttacaatcaaattatcgcaagataaggcgcacgttgattgggtcacccgagtgtacg3660
ttgataaagtcacgtgggcacccaacgcgttgataagcatcggtatataagggcctacag3720
tgttctggtaaatcagttgcactgtgctcttcacaggaacactacaagacctacaagatg3780
gcttcactggttaaaaaagacatgtgcatcaaaatgacgatggaaggaacagtaaacggt3840
caccatttcaagtgtgtaggagaaggcgaaggcaaaccatttgaagggacccaggtggaa3900
aagatacgcatcactgaaggtgggcccttaccatttgcgtatgatattttggccccttgt3960
tgcatgtatggcagtaaaaccttcattaagcatgtgtcgggtattccggattactttaag4020
gagtcttttcctgagggctttacctgggaaagaacacaaatcttcgaggatggcggctat4080
ctcaccatacaccaggacacgagccttcagggtaataattttattttcaaagttaatgtc4140
atcggtgccaacttccctgcaaacggtcccgtgatgcagaaaaaaacagctggatgggaa4200
ccgtgcgttgagatgctttatccgcgggacggcgtcctgtgtggtcagagcctgatggcc4260
ctgaaatgcactgatggcaatcatctgacgtcccacctgcgcactacctatcgttctcgc4320
aagccatccaatgcagttaacatgccggaatttcattttggggatcatcgcattgagatt4380
ttgaaagctgaacaaggtaaattttatgaacaatacgagtcagcggtggcccgttactgt4440
gaggcggcaccgagtaaattagggcatcactaataacgtttattttattgcaataagatc4500
gacagaattaaacgcacggaataaagtgttgggtcgtttgttctaactataacggtccta4560
aggtagcgaggtaccgaattcctgcagtcatttattaatctcaacacactcgctatttgg4620
aacataatcatatcgtctcagtagctcaaggtagagcgtagcgctctggatcgtatagat4680
cttgctaaggttgtgagttcaagtctcgcctgagatattaaaaaactttgtaattttaaa4740
aattttattttataatatacaattaaaaactatacaattttttattattacattaataat4800
gatacaatttttattattacatttaatattgtctattacggtttctaatcatacagtaca4860
aaaataaaatcacaattaatataattacaaagttaactacatgaccaaacatgaacgaag4920
tcaatttagcggccaattcgccttcagccatggaagtgatgtcgctcagactggtgccga4980
cgccgccaaacttggtgttctccatggtggttatgaggttgcttttttgttgggcaataa5040
acgaccagccgctggcatctttccaactgtcgtgataggtcgtgttgccgatggtcggga5100
tccaaaactcgacgtcgtcgtcaattgctagttccttgtagttgctaaaatctatgcatt5160
gcgacgagtccgtgttggccacccaacgcccttctttgtagatgctgttgttgtagcaat5220
tactggtgtgtgccggcggattggtgcacggcatcagcaaaaacgtgtcgtccgacaaaa5280
atgttgaagaaacagagttgttcatgagattgccaatcaaacgctcgtccaccttggcca5340
cggagactatcaggtcgtgcagcatattgtttagcttgttgatgtgcgcatgcatcagct5400
caatgttcattttcagcaaatcgttttcgtacatcagctcctcttgaatatgcatcaggt5460
cgcctttggtggcagtgtctccctctgtgtacttggctctaacgttgtggcgccaagtgg5520
gcggccgcttcttgactcggtgctcgactttgcgtttaatgcatctgttaaacttgcagt5580
tccacgtgtttttagaaagatcatatatatcattgtcaatcaaacagtgttcgcgtgtca5640
ccgactcggggttatttttgtcatctttaatgagcagacacgcagcttttatttggcgcg5700
tggtgaacgtagacttttgtttgagaatcatactcacgccgtctcgatgaagcacagtgt5760
ccacggtcacgttgatggggt5781
<210>3
<211>2860
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>来源(source)
<223>/备注=“人工序列的描述:合成的多核苷酸”
<400>3
atgatgtcatcatcacaaataatagtatgtaataaaataaacatatttgtgtgtaaatat60
aatttattacaaataaattttacattgaatcaatctgtcttcgtgtttgttgtaaggtct120
tcgaatcttgtgtttcagcccctcgggatggtcaaaatgcgccgtagtaattgttaatgg180
atctttcaacgattttttgcccatggcgagtgtgacaaacgcggccacgacaaacagcag240
gataatcagtttcatggtgttctatattcgacaatatatgggtcgcttctaaatcacctt300
gtccccaaaagcctcttttatagttttttagaacacgttgtgtattccaacagtaattgt360
tccatctctttcaacaatttcacacaggaaacagctatgaccatgattacgaattcctcg420
ggtatgcatttgttaaaaaacacacagctcgctttaccaaacgaaagcaaaggtactaaa480
tatggcgccattggctgatttgttattccaagataattacaaataaactgatccgtcgtg540
gggtgataactggcaggtgtcagctttaaataatcttcaacgttgttgtcgcgcaaaagt600
ctgcattttacacgcgttgttaatcccacgacttttgcatgtaaaatcggatccaaatac660
tgcagaatcgtgtctataatttctaatggtaaacgtatgcgttttgctcgtgggcgcttt720
gtaacgctcgacatcctaataacaactaacacaaaactaaaatgatactcaatatattgc780
ttttacagttcatctttaggtttaaactgtgcgtttatcgcgttgagcaagtcgccgtta840
tcggcatcaatctcccaagcaaacaggccgcccaatttatttcggtcgacatatttaact900
tttcctaacacagagtcgacgctgtcaaacgaaatcaaatcacctttacttttatcgaaa960
acgtacgacgcttgagcggcgctgtcaaacgtgtacacataattgttgagatctttttga1020
atttgacgataatctacaacaccgtcctcccacgtgcccgaccccggcccgttgccagtg1080
ccggaaaaatagttgtcattcgtataatttgttacgccggtccagccgcggccgtacatg1140
gcgacgcccacaattattttgttgggatcgacgccttgtttcagtaacgcatcgacagcg1200
tagtgtgtagtgtatagctcttccgagttccaacttggcgcgtagactgttgtttggtag1260
cccaaatccgtgtttgaccaagcccctttaaaatcgtaactcatgagaaatattttgcct1320
aatgacttttgcgcttcggcgtagtttaccacggcaatcttgtcgtaacccgcgcttata1380
gcgcttgttaattcgtaaaccctgccggtttgcgcttcgaggtcgtctagcattgcgcgc1440
agctcctccaacaacaaaatgtatgttttggcgtcaccgtccgcatcgcccaacgacggg1500
ttagcccctttgccgcccggaaactcccaatcgatgtctacaccgtcaaagaatttccac1560
acttgcagaaattccttaaccgaatctacaaaaacgtttcttttttcaacatcgtgggta1620
cctaactataacggtcctaaggtagcgaggtaccgaattcctgcagtcatttattaatct1680
caacacactcgctatttggaacataatcatatcgtctcagtagctcaaggtagagcgtag1740
cgctctggatcgtatagatcttgctaaggttgtgagttcaagtctcgcctgagatattaa1800
aaaactttgtaattttaaaaattttattttataatatacaattaaaaactatacaatttt1860
ttattattacattaataatgatacaatttttattattacatttaatattgtctattacgg1920
tttctaatcatacagtacaaaaataaaatcacaattaatataattacaaagttaactaca1980
tgaccaaacatgaacgaagtcaatttagcggccaattcgccttcagccatggaagtgatg2040
tcgctcagactggtgccgacgccgccaaacttggtgttctccatggtggttatgaggttg2100
cttttttgttgggcaataaacgaccagccgctggcatctttccaactgtcgtgataggtc2160
gtgttgccgatggtcgggatccaaaactcgacgtcgtcgtcaattgctagttccttgtag2220
ttgctaaaatctatgcattgcgacgagtccgtgttggccacccaacgcccttctttgtag2280
atgctgttgttgtagcaattactggtgtgtgccggcggattggtgcacggcatcagcaaa2340
aacgtgtcgtccgacaaaaatgttgaagaaacagagttgttcatgagattgccaatcaaa2400
cgctcgtccaccttggccacggagactatcaggtcgtgcagcatattgtttagcttgttg2460
atgtgcgcatgcatcagctcaatgttcattttcagcaaatcgttttcgtacatcagctcc2520
tcttgaatatgcatcaggtcgcctttggtggcagtgtctccctctgtgtacttggctcta2580
acgttgtggcgccaagtgggcggccgcttcttgactcggtgctcgactttgcgtttaatg2640
catctgttaaacttgcagttccacgtgtttttagaaagatcatatatatcattgtcaatc2700
aaacagtgttcgcgtgtcaccgactcggggttatttttgtcatctttaatgagcagacac2760
gcagcttttatttggcgcgtggtgaacgtagacttttgtttgagaatcatactcacgccg2820
tctcgatgaagcacagtgtccacggtcacgttgatggggt2860
<210>4
<211>4713
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>来源(source)
<223>/备注=“人工序列的描述:合成的多核苷酸”
<400>4
atgatgtcatcatcacaaataatagtatgtaataaaataaacatatttgtgtgtaaatat60
aatttattacaaataaattttacattgaatcaatctgtcttcgtgtttgttgtaaggtct120
tcgaatcttgtgtttcagcccctcgggatggtcaaaatgcgccgtagtaattgttaatgg180
atctttcaacgattttttgcccatggcgagtgtgacaaacgcggccacgacaaacagcag240
gataatcagtttcatggtgttctatattcgacaatatatgggtcgcttctaaatcacctt300
gtccccaaaagcctcttttatagttttttagaacacgttgtgtattccaacagtaattgt360
tccatctctttcaacagccattcagcatccggtcgttgactgtaatcatgctgaattaat420
ttacaaacaatttcggtcaatttaggatggccttgggataaacttgccggcatttgctgt480
acattgtttctaaagttagttagcgtagtttcgcgttccaaagcagtcttgaagggcatt540
atcaattcgaataaaacaatgcccaaactatacatgtcatttttgggggtgtacactttt600
ttgatttgttctggtgcagcgtacaaagttatattttgagggttgtttttgataaacgtt660
ttgtatagactgccaaacatgccgcccacatacaaatcaaagtcgggcccagtcatgaaa720
atatcttcgggattaatattgtggtgcacgatatttacggaatgaatcgctttcacggcg780
ctcaccaaatcaacaaacttgctaatataaaagccaaaatccgccggaactttaatgttg840
gtctttgcaaaagtttgcaaattgcgttgtttcaaatagtcgctcaacatgtactcgttt900
agaggcgacgcaaaatatatgcggtgctgccgcggattcaaataaaccaattgttcgggt960
ttcatggtatacagttaagtgttaacgcgtcactaaattcagacacgagcgcacgcccta1020
tatacatacaatttatcgcacaagatgcttaacgcgatctgtttataaactaaaacgcac1080
tgcaataaattttagcaagcatttgtatttaatcaatcgaaccgtgcactgatataagaa1140
ttaaaaatgggtttgtttgcgtgttgcacaaaatacacaaggctgtcgaccgacacaaaa1200
atgaagtttccctatgttgcgttgtcgtacatcaacgtgacgctgtgcacctacaccgcc1260
atgttggtgggatacatggtaacattcaatgactccagcgaattgaaatatttacaatac1320
tggttgctgttgtcgtttttgatgtccgtggtgctaaacgctccgactctgtggacgatg1380
ctcaaaaccacagaagcccatgaagtaatttacgaaatgaagctgttccacgccatgtac1440
tttagtaacgtgctgttgaattatgtggtgtttttggacaatcaaatgggtacaaatttt1500
gtttttgttaacaatttaattcactgttgtgtactttttatgatatttgttgaattgctt1560
atcctgttgggccacacaatgggcacgtacacggattatcaatatgtcaaatcgtgttat1620
atggttatattgtttgtttcagttatgagtgttactattgttatgggtttagagtgtttg1680
aaaacgaaactaattgataacagtttgatgtttaacgcgtttgtgtgcgctttgtacatt1740
gtgattgcaataatgtggtctttaaaaaataatttgactagttattacgtttcaaattta1800
caaagtattcaagttgttccgttttcatacaacgatccgccgccaccgttctctaacatt1860
gtaatggatgacataaaaaataaaaaataatttataaaaatgttttttattctttcacaa1920
ttctgtaaattctaaacaaaaaatataaatacaaacttattatgttgtcgtctaaataaa1980
catcaatttgtaaatctggacacctattcatatcattgatattacagtctactatacaac2040
aattaaaactaaccaaattatctttacaacaattaaagcaattaaaacaatttaaataat2100
cttcattgtcgtcgtataagtttatttgcactgtagacggtgttacacagcgatccattc2160
gacgttcgtgttcgatcaactttctcgccaacttgtaccataaaaattgtttggacaaaa2220
agttttccaacaatggtaacggccaattcaacgtgacgatgcgcacgtcctcgggtatgc2280
atttgttaaaaaacacacagctcgctttaccaaacgaaagcaaaggtactaaatatggcg2340
ccattggctgatttgttattccaagataattacaaataaactgatccgtcgtggggtgat2400
aactggcaggtgtcagctttaaataatcttcaacgttgttgtcgcgcaaaagtctgcatt2460
ttacacgcgttgttaatcccacgacttttgcatgtaaaatcggatccaaatactgcagaa2520
tcgtgtctataatttctaatggtaaacgtatgcgttttgctcgtgggcgctttgtaacgc2580
tcgacatcctaataacaactaacacaaaactaaaatgatactcaatatattgcttttaca2640
gttcatctttaggtttaaactgtgcgtttatcgcgttgagcaagtcgccgttatcggcat2700
caatctcccaagcaaacaggccgcccaatttatttcggtcgacatatttaacttttccta2760
acacagagtcgacgctgtcaaacgaaatcaaatcacctttacttttatcgaaaacgtacg2820
acgcttgagcggcgctgtcaaacgtgtacacataattgttgagatctttttgaatttgac2880
gataatctacaacaccgtcctcccacgtgcccgaccccggcccgttgccagtgccggaaa2940
aatagttgtcattcgtataatttgttacgccggtccagccgcggccgtacatggcgacgc3000
ccacaattattttgttgggatcgacgccttgtttcagtaacgcatcgacagcgtagtgtg3060
tagtgtatagctcttccgagttccaacttggcgcgtagactgttgtttggtagcccaaat3120
ccgtgtttgaccaagcccctttaaaatcgtaactcatgagaaatattttgcctaatgact3180
tttgcgcttcggcgtagtttaccacggcaatcttgtcgtaacccgcgcttatagcgcttg3240
ttaattcgtaaaccctgccggtttgcgcttcgaggtcgtctagcattgcgcgcagctcct3300
ccaacaacaaaatgtatgttttggcgtcaccgtccgcatcgcccaacgacgggttagccc3360
ctttgccgcccggaaactcccaatcgatgtctacaccgtcaaagaatttccacacttgca3420
gaaattccttaaccgaatctacaaaaacgtttcttttttcaacatcgtgggtacctaact3480
ataacggtcctaaggtagcgaggtaccgaattcctgcagtcatttattaatctcaacaca3540
ctcgctatttggaacataatcatatcgtctcagtagctcaaggtagagcgtagcgctctg3600
gatcgtatagatcttgctaaggttgtgagttcaagtctcgcctgagatattaaaaaactt3660
tgtaattttaaaaattttattttataatatacaattaaaaactatacaattttttattat3720
tacattaataatgatacaatttttattattacatttaatattgtctattacggtttctaa3780
tcatacagtacaaaaataaaatcacaattaatataattacaaagttaactacatgaccaa3840
acatgaacgaagtcaatttagcggccaattcgccttcagccatggaagtgatgtcgctca3900
gactggtgccgacgccgccaaacttggtgttctccatggtggttatgaggttgctttttt3960
gttgggcaataaacgaccagccgctggcatctttccaactgtcgtgataggtcgtgttgc4020
cgatggtcgggatccaaaactcgacgtcgtcgtcaattgctagttccttgtagttgctaa4080
aatctatgcattgcgacgagtccgtgttggccacccaacgcccttctttgtagatgctgt4140
tgttgtagcaattactggtgtgtgccggcggattggtgcacggcatcagcaaaaacgtgt4200
cgtccgacaaaaatgttgaagaaacagagttgttcatgagattgccaatcaaacgctcgt4260
ccaccttggccacggagactatcaggtcgtgcagcatattgtttagcttgttgatgtgcg4320
catgcatcagctcaatgttcattttcagcaaatcgttttcgtacatcagctcctcttgaa4380
tatgcatcaggtcgcctttggtggcagtgtctccctctgtgtacttggctctaacgttgt4440
ggcgccaagtgggcggccgcttcttgactcggtgctcgactttgcgtttaatgcatctgt4500
taaacttgcagttccacgtgtttttagaaagatcatatatatcattgtcaatcaaacagt4560
gttcgcgtgtcaccgactcggggttatttttgtcatctttaatgagcagacacgcagctt4620
ttatttggcgcgtggtgaacgtagacttttgtttgagaatcatactcacgccgtctcgat4680
gaagcacagtgtccacggtcacgttgatggggt4713
<210>5
<211>743
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>来源(source)
<223>/备注=“人工序列的描述:合成的多肽”
<400>5
metalaalaaspglytyrleuproasptrpleugluaspasnleuser
151015
gluglyileargglutrptrpalaleulysproglyalaproglnpro
202530
lysalaasnglnglnhisglnaspasnalaargglyleuvalleupro
354045
glytyrlystyrleuglyproglyasnglyleuasplysglyglupro
505560
valasnalaalaaspalaalaalaleugluhisasplysalatyrasp
65707580
glnglnleulysalaglyaspasnprotyrleulystyrasnhisala
859095
aspalaglupheglngluargleulysgluaspthrserpheglygly
100105110
asnleuglyargalavalpheglnalalyslysargleuleuglupro
115120125
leuglyleuvalgluglualaalalysthralaproglylyslysarg
130135140
provalgluglnserproglngluproaspserseralaglyilegly
145150155160
lysserglyalaglnproalalyslysargleuasnpheglyglnthr
165170175
glyaspthrgluservalproaspproglnproileglyglupropro
180185190
alaalaproserglyvalglyserleuthrmetalaserglyglygly
195200205
alaprovalalaaspasnasngluglyalaaspglyvalglyserser
210215220
serglyasntrphiscysaspserglntrpleuglyaspargvalile
225230235240
thrthrserthrargthrtrpalaleuprothrtyrasnasnhisleu
245250255
tyrlysglnileserasnserthrserglyglyserserasnaspasn
260265270
alatyrpheglytyrserthrprotrpglytyrpheasppheasnarg
275280285
phehiscyshispheserproargasptrpglnargleuileasnasn
290295300
asntrpglypheargprolysargleuasnphelysleupheasnile
305310315320
glnvallysgluvalthraspasnasnglyvallysthrilealaasn
325330335
asnleuthrserthrvalglnvalphethraspserasptyrglnleu
340345350
protyrvalleuglyseralahisgluglycysleuproprophepro
355360365
alaaspvalphemetileproglntyrglytyrleuthrleuasnasp
370375380
glyserglnalavalglyargserserphetyrcysleuglutyrphe
385390395400
proserglnmetleuargthrglyasnasnpheglnphesertyrglu
405410415
phegluasnvalprophehissersertyralahisserglnserleu
420425430
aspargleumetasnproleuileaspglntyrleutyrtyrleuser
435440445
argthrileasnglyserglyglnasnglnglnthrleulyspheser
450455460
valalaglyproserasnmetalavalglnglyargasntyrilepro
465470475480
glyprosertyrargglnglnargvalserthrthrvalthrglnasn
485490495
asnasnsergluphealatrpproglyalasersertrpalaleuasn
500505510
glyargasnserleumetasnproglyproalametalaserhislys
515520525
gluglygluaspargphepheproleuserglyserleuilephegly
530535540
lysglnglythrglyargaspasnvalaspalaasplysvalmetile
545550555560
thrasngluglugluilelysthrthrasnprovalalathrgluser
565570575
tyrglyglnvalalathrasnhisglnseraspglythrleualaval
580585590
prophelysalaglnalaglnthrglytrpvalglnasnglnglyile
595600605
leuproglymetvaltrpglnaspargaspvaltyrleuglnglypro
610615620
iletrpalalysileprohisthraspglyasnphehisproserpro
625630635640
leumetglyglypheglymetlyshisproproproglnileleuile
645650655
lysasnthrprovalproalaaspproprothralapheasnlysasp
660665670
lysleuasnserpheilethrglntyrserthrglyglnvalserval
675680685
gluileglutrpgluleuglnlysgluasnserlysargtrpasnpro
690695700
gluileglntyrthrserasntyrtyrlysserasnasnvalgluphe
705710715720
alavalasnthrgluglyvaltyrsergluproargproileglythr
725730735
argtyrleuthrargasnleu
740
- 还没有人留言评论。精彩留言会获得点赞!