双特异性EGFR/c-Met抗体的制作方法
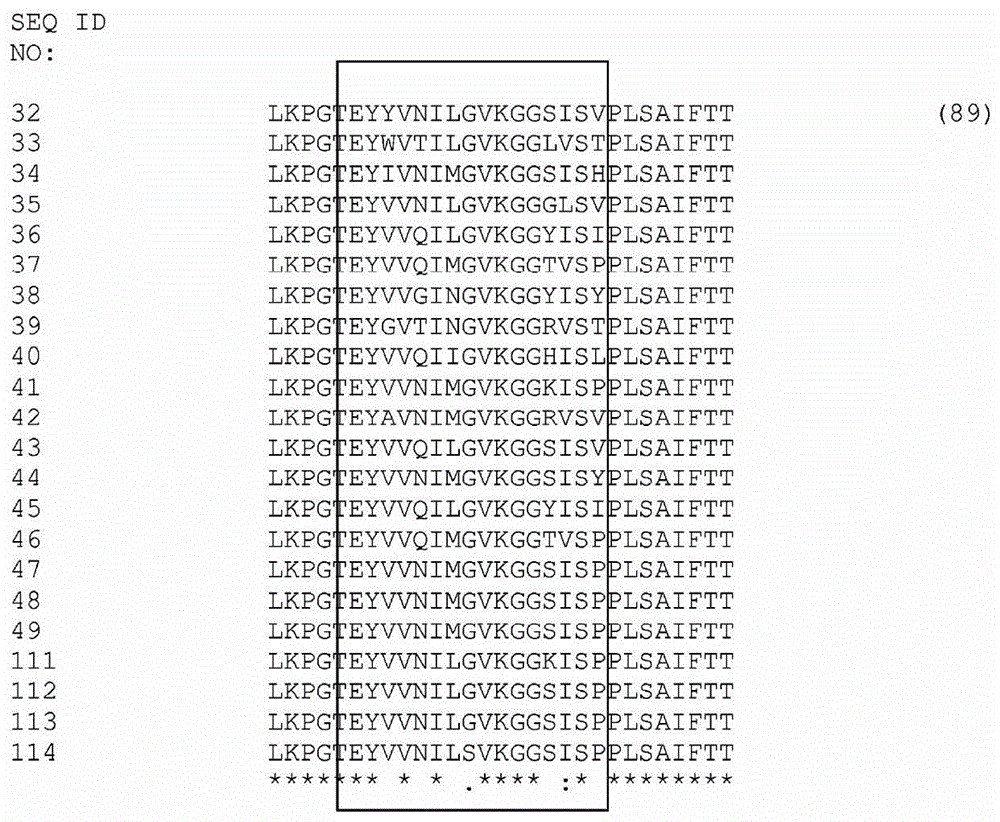
本申请是与母案发明名称相同的分案申请,母案的中国申请号是201380071081.8,国际申请号是pct/us2013/071288,申请日是2013年11月21日。本发明涉及双特异性egfr/c-met抗体及制备和使用所述分子的方法。
背景技术:
:表皮生长因子受体(egfr、erbbl或her1)是由c-erbbl原癌基因编码的170kdai型跨膜糖蛋白。egfr是受体酪氨酸激酶(rtk)的人表皮生长因子受体(her)家族的成员,该家族包括her2(erbb2)、her3(erbb3)和her4(erbb4)。egfr信号传导通过如下方式引发:配体结合、之后诱导构象变化、受体与其他erbb家族成员的同源二聚化或异源二聚化,以及受体的反式自磷酸化(ferguson等人,annurevbiophys,37:353-73,2008),这引发了信号转导级联,最终影响多种多样的细胞功能,包括细胞增殖和存活。egfr的表达或激酶活性的增加与一系列人癌症有关,从而使egfr成为治疗干预的有吸引力的靶标(mendelsohn等人,oncogene19:6550-6565,2000;grünwald等人,jnatlcancerinst95:851-67,2003;mendelsohn等人,seminoncol33:369-85,2006)。在非小细胞肺癌中,egfr基因拷贝数和蛋白表达两者的增加与对egfr酪氨酸激酶抑制剂iressatm(吉非替尼)的有利响应相关联(hirsch等人,annoncol18:752-60,2007)。egfr疗法包括小分子和抗egfr抗体两者,它们经批准用于治疗结肠直肠癌、胰腺癌、头颈癌和非小细胞肺癌(nsclc)(baselga和arteaga,jclinoncol23:2445-2459(20005;gill等人,jbiolchem,259:7755-7760,1984;goldstein等人,clincancerres,1:1311-1318;1995;prewett等人,clincancerres,4:2957-2966,1998)。抗egfr疗法的疗效可取决于瘤类型以及瘤中的egfr突变/扩增状态。当前疗法的副作用可包括皮肤毒性(deroock等人,lancetoncol11:753-762,2010;linardou等人,natrevclinoncol,6:352-366,2009;li和perez-soler,targoncol4:107-119,2009)。egfr酪氨酸激酶抑制剂(tki)通常用作非小细胞肺癌(nsclc)的二线疗法,但由于抗药途径的原因,通常会在十二个月内失效(riely等人,clincancerres12:839-44,2006)。c-met编码跨膜酪氨酸激酶受体。其在1984年在发现用致癌物处理产生了组成型活性融合蛋白tpr-met之后,首次被鉴定为原癌基因(cooper等人,nature311:29-33,1984)。c-met被其配体肝细胞生长因子(hgf)激活会刺激多个细胞过程,包括生长、运动、侵袭、转移、上皮细胞-间充质转化、血管生成/创伤愈合以及组织再生(christensen等人,cancerlett225:1-26,2005;peters和adjei,natrevclinoncol9:314-26,2012)。c-met被合成为单链蛋白质,该蛋白质被蛋白水解裂解成由二硫键连接的50kdaα-和140kdaβ-亚基(ma等人,cancerandmetastasisreviews,22:309-325,2003)。c-met在结构上类似于其他膜受体诸如ron和sea。hgf:c-met结合的准确化学计量尚不清楚,但普遍认为两个hgf分子结合于两个c-met分子,导致受体二聚化以及在1230、1234和1235位酪氨酸处自磷酸化(stamos等人,theembojournal23:2325-2335,2004)。非配体依赖性c-met自磷酸化也可由于基因扩增、突变或受体过表达而发生。c-met在多种类型的癌(包括胃癌、肺癌、结肠癌、乳腺癌、膀胱癌、头颈癌、卵巢癌、前列腺癌、甲状腺癌、胰腺癌和cns癌)中常常被扩增、突变或过表达。通常定位于激酶域的错义突变常见于遗传性乳头状肾细胞癌(prcc)和13%的散发性prcc(schmidt等人,oncogene18:2343-2350,1999)。定位于c-met的臂板蛋白或近膜域的c-met突变常见于胃癌、头颈癌、肝癌、卵巢癌、nsclc和甲状腺癌(ma等人,cancerandmetastasisreviews,22:309-325,2003;sakakura等人,chromosomesandcancer,1999,24:299-305)。已在脑癌、结肠直肠癌、胃癌和肺癌中检测到c-met扩增,这通常与疾病进展相关(ma等人,cancerandmetastasisreviews,22:309-325,2003)。分别多至4%和20%的非小细胞肺癌(nsclc)和胃癌表现出c-met扩增(sakakura等人,chromosomesandcancer,1999,24:299-305:sierra和tsao,therapeuticadvancesinmedicaloncology,3:s21-35,2011)。即使不存在基因扩增,肺癌中也常观察到c-met过表达(ichimura等人,jpnjcancerres,87:1063-9,1996)。此外,在临床样品中,将近一半的肺腺癌表现出高水平的c-met和hgf,这两者均与增强的瘤生长率、转移和不良预后相关(sierra和tsao,therapeuticadvancesinmedicaloncology,3:s21-35,2011;siegfried等人,annthoracsurg66:1915-8,1998)。抵抗egfr酪氨酸激酶抑制剂的所有瘤中的将近60%增加c-met表达、扩增c-met或增加c-met的唯一已知配体hgf(turke等人,cancercell,17:77-88,2010),表明存在通过c-met对egfr的补偿途径。在抵抗吉非替尼(一种egfr激酶抑制剂)并经her3途径表现出增强存活率的培养细胞中,c-met扩增被首次鉴定(engelman等人,science,316:1039-43,2007)。这进一步在临床样品中得到验证,其中具有对厄洛替尼或吉非替尼的获得性抗性的43名患者中有九名表现出c-met扩增,与之相比的是,62名未治疗患者中仅两名有此表现。九名治疗患者中有四名也获得了egfr激活突变t790m,证实为同步抗性途径(beat等人,procnatlacadsciusa,104:20932-7,2007)。egfr和c-met在癌中的单独作用已充分确立,使得这些靶标对于联合疗法具有吸引力。两个受体均通过相同的存活和抗凋亡途径(erk和akt)传导信号;因此,联合抑制该对受体可限制补偿途径激活的可能性,从而改善总体疗效。在(厄洛替尼)与nsclc的抗c-met单价抗体联用(spigel等人,2011ascoannualmeetingproceedings2011,journalofclinicaloncology:chicago,il.,第7505页)以及tarceva(厄洛替尼)与arq-197(一种c-met小分子抑制剂)联用(adjei等人,oncologist,16:788-99,2011)的临床试验中测试了靶向egfr和c-met的联合疗法。联合疗法或双特异性抗egfr/c-met分子已例如在如下专利中公开:国际专利公布wo2008/127710、wo2009/111691、wo2009/126834、wo2010/039248、wo2010/115551以及美国专利公布us2009/0042906。目前的拮抗egfr和/或c-met信号传导途径进行治疗的小分子和大分子治疗方法可能为次优的,这是由于可能缺少特异性、可能的脱靶活性以及小分子抑制剂可能遇到的剂量限制性毒性。典型的单特异性二价抗体可导致膜结合受体聚簇以及下游信号传导途径的非期望激活。具有全长重链(半臂)的单价抗体对制造工艺带来了相当大的复杂性和成本。因此,对另外的单特异性和双特异性egfr和/或c-met抑制剂存在需要,以用于治疗和诊断目的两者。技术实现要素:本发明的一个实施例是分离的双特异性表皮生长因子受体(egfr)/肝细胞生长因子受体(c-met)抗体,其包含:第一重链(hc1),其包含hc1恒定域3(hc1ch3)和hc1可变区1(vh1);第二重链(hc2),其包含hc2恒定域3(hc2ch3)和hc2可变区2(vh2);第一轻链(lc1),其包含轻链可变区1(vl1);和第二轻链(lc2),其包含轻链可变区2(vl2),其中vh1和vl1配对以形成特异性结合egfr的第一抗原结合位点,vh2和vl2配对以形成特异性结合c-met的第二抗原结合位点,hc1在hc1ch3中包含至少一个替换,并且hc2在hc2ch3中包含至少一个替换,并且当根据eu索引对残基进行编号时,hc1ch3中的替换和hc2ch3中的替换在不同氨基酸残基位置处发生。在其他实施例中,本发明提供双特异性egfr/c-met抗体,其中该抗体抑制nci-h292、nci-h1975或skmes-1细胞系中胞外信号相关激酶1和2(erk1/2)的磷酸化,并且与用包含重链3(hc3)和轻链3(lc3)的对照单价egfr抗体以及包含重链4(hc4)和轻链4(lc4)的对照单价c-met抗体的混合物抑制nci-h292、nci-h1975或skmes-1细胞系中erk1/2的磷酸化的ic50值相比,其ic50值小至少约10倍、小至少约20倍、小至少约30倍、小至少约40倍、小至少约50倍或小至少约60倍,其中hc3与hc1、lc3与lc1、hc4与hc2以及lc4与lc2分别具有相同的氨基酸序列,其中使用夹心免疫测定法,将抗磷酸化erk1/2抗体用作捕获抗体并将与电化学发光化合物缀合的结合于非磷酸化和磷酸化erk1/2的抗体用作检测抗体,在全细胞裂解物中测量erk1/2的磷酸化。在其他实施例中,本发明提供双特异性egfr/c-met抗体,其中该抗体抑制nci-h1975细胞系中蛋白激酶b(akt)在ser473处的磷酸化,并且与用包含hc3和lc3的对照单价egfr抗体以及包含hc4和lc4的对照单价c-met抗体的混合物抑制nci-h1975细胞系中akt在ser473处的磷酸化的ic50值相比,其ic50值小至少约70倍,其中hc3与hc1、lc3与lc1、hc4与hc2以及lc4与lc2分别具有相同的氨基酸序列,其中使用夹心免疫测定法,将结合于非磷酸化和磷酸化akt的抗体用作捕获抗体并将与电化学发光化合物缀合的抗磷酸化aktser473抗体用作检测抗体,在全细胞裂解物中测量akt在ser473处的磷酸化。在其他实施例中,本发明提供双特异性egfr/c-met抗体,该抗体在egfr残基k489、i491、k467和s492处结合seqidno:73的egfr,并且在残基pefrdsypikyvhaf(seqidno:238)和faqskpdsaepmdrsa(seqidno:239)处结合c-met。在其他实施例中,本发明提供双特异性egfr/c-met抗体,该抗体抑制nci-h292或nci-h1975细胞的生长,并且当nci-h292或nci-h1975细胞在低附着条件下生长时,与用西妥昔单抗抑制nci-h292或nci-h1975细胞的生长的ic50值相比,其ic50值小至少约300倍、小至少约400倍、小至少约500倍、小至少约600倍、小至少约700倍或小至少约800倍。在其他实施例中,本发明提供双特异性egfr/c-met抗体,该抗体抑制scidbeige小鼠中表达hgf的skmes-1细胞瘤的生长,并且当双特异性抗体和西妥昔单抗以20mg/kg剂量施用时,与西妥昔单抗相比,在第36天,其t/c百分比(%)值小至少500倍。在其他实施例中,本发明提供双特异性egfr/c-met抗体,其中hc1ch3包含k409r或f405l替换,并且hc2ch3包含k409r或f405l替换,其中根据eu索引对残基进行编号。在其他实施例中,本发明提供双特异性egfr/c-met抗体,该抗体包含特定重链和轻链cdr、vh1、vl1、vh2、vl2、hc1、lc1、hc2和lc2序列。本发明的另一个实施例是分离的合成多核苷酸,其编码本发明的hc1、hc2、lc1或lc2。本发明的另一个实施例是包含本发明多核苷酸的载体。本发明的另一个实施例是包含本发明载体的宿主细胞。本发明的另一个实施例是制备分离的双特异性egfr/c-met抗体的方法,所述方法包括:将包含seqidno:199的两条重链和seqidno:200的两条轻链的分离的单特异性二价抗egfr抗体与包含seqidno:201的两条重链和seqidno:202的两条轻链的分离的单特异性二价抗c-met抗体以约1:1摩尔比组合在混合物中;将还原剂引入混合物中;将混合物温育约九十分钟至约六小时;移除还原剂;以及纯化双特异性egfr/c-met抗体,其包含seqidno:199的第一重链和seqidno:201的第二重链、seqidno:200的第一轻链和seqidno:202的第二轻链,其中seqidno:199的第一重链与seqidno:200的第一轻链配对以形成特异性结合egfr的第一结合域,并且seqidno:201的第二重链与seqidno:202的第二轻链配对以形成特异性结合c-met的第二结合域。本发明的另一个实施例是包含本发明双特异性抗体和药学上可接受的载体的药物组合物。本发明的另一个实施例是治疗患有癌的受检者的方法,所述方法包括向对其有需要的患者施用治疗有效量的本发明双特异性egfr/c-met抗体足以治疗癌的时间。本发明的另一个实施例是抑制表达egfr和/或c-met的细胞的生长或增殖的方法,所述方法包括使细胞与本发明双特异性抗体接触。本发明的另一个实施例是抑制受检者中表达egfr和/或c-met的瘤或癌细胞的生长或转移的方法,所述方法包括向受检者施用有效量的本发明双特异性抗体以抑制表达egfr和/或c-met的瘤或癌细胞的生长或转移。附图说明图1a和1b.结合egfr的fn3域的氨基酸比对。在seqidno:18的第22-28位和第75-86位残基处对bc和fg环加框。一些变体包括改善热稳定性的l17a、n46k和e86i替换(残基根据tenconseqidno:1编号)。图2.tencon27支架(seqidno:99)与具有随机化c-cd-f-fg另选表面的tcl14文库(seqidno:100)的序列比对。对环残基加框。在序列的上方指示环和链。图3.结合c-met的fn3域的序列比对。c环和cd链及f环和fg链被加框并且跨越第29-43位和第65-81位残基。图4.示出了在用包含单特异性或双特异性fn3域的分子预处理并用hgf刺激的nci-h292细胞中c-met磷酸化的抑制。当与单独的单特异性的结合c-met的fn3域(p114ar5p74-a5,图中以a5示出)或联用结合egfr的fn3域(p54ar4-83v2,图中以83v2示出)相比时,观察到双特异性egfr/c-met分子(ecb1)的效力的大幅提高。图5.在用包含单特异性或双特异性fn3域的分子预处理的细胞中的egfr和c-met磷酸化的抑制。在表达高水平的egfr的细胞系nci-h292(图5a)和h596(图5b)中,包含抗egfr单特异性和双特异性fn3域的分子在减少egfr磷酸化方面效力相同。在表达相对于c-met而言低水平的egfr的细胞系nci-h441(图5c)中,与单独单特异性的结合egfr的fn3域相比,双特异性egfr/c-met分子改善了egfr磷酸化的抑制效力。在具有相对于egfr而言低水平的c-met的细胞系nci-h292(图5d)和h596(图5e)中,与仅单特异性的结合c-met的fn3域相比,双特异性egfr/c-met分子显著增强了c-met磷酸化的抑制。该研究所用的分子是:双特异性ecb5(图中以17-a3示出)、单特异性的结合egfr的fn3域p53a1r5-17(图中以“17”示出)、双特异性egfr/c-met分子ecb3(图中以83-h9示出)以及单特异性的结合c-met的fn3域p114ar7p93-h9(图中以h9示出)。图6.从用双特异性egfr/c-met分子给药6h或72h的小鼠分离的瘤中的药效信号传导。所有分子在6h和72h时显著减少了c-met、egfr和erk磷酸化,抑制的程度取决于fn3域对egfr和/或c-met的亲和力。双特异性分子通过如下方式生成:将具有高(图中的“83”是p54ar4-83v2)或中等(图中的“17v2”是p53a1r5-17v2)亲和力的结合egfr的fn3域连接到具有高(图中的“a3”是p114ar7p94-a3)或中等(图中的“a5”是p114ar5p74-a5)亲和力的结合c-met的fn3域。图7.示出了在ip给药后6h(左)和72h(右)连接到白蛋白结合域(abd)的可变亲和力的双特异性egfr/cmet分子的血浆(顶部)和瘤(底部)积聚。在用具有中等亲和力结合egfr的fn3域(17v2)或高亲和力egfr结合域(83v2)的双特异性分子给药的小鼠中,给药后六小时,瘤积聚达最大值。双特异性分子掺入了如下高或中等亲和力结合egfr或c-met的fn3域:83v2-a5-abd(ecb18;对egfr/cmet的亲和力为高/中等)83v2-a3-abd(ecb38;高/高)17v2-a5(ecb28;中等/中等)17v2-a3-abd(ecb39;中等/高)。在该图中,83v2是指p54ar4-83v2;17v2是指p53a1r5-17v2;a3是指p114ar7p94-a3,并且a5是指p114ar5p74-a5。图8.h292-hgf移植瘤被植入scidbeige小鼠中。当瘤达到大约80mm3的平均体积时,每周用双特异性egfr/c-met分子(25mg/kg)或pbs媒介物给予小鼠三次。所有双特异性分子降低了瘤生长,瘤生长抑制(tgi)取决于分子对c-met和egfr的亲和力(高egfr-高cmet是指p54ar4-83v2-p114ar7p94-a3(ecb38);高egfr-中等cmet是指p54ar4-83v2-p114ar5p74-a5(ecb18);中等egfr-高cmet是指p53a1r5-17v2-p114ar7p94-a3(ecb39);中等egfr-中等-cmet是指p53a1r5-17-p114ar5p74-a5(ecb28))。图9.h292-hgf移植瘤被植入scidbeige小鼠中并且它们用不同疗法治疗。示出了疗法的抗瘤活性(双特异性egfr/c-met分子是指p54ar4-83v2-p114ar7p94-a3-abd(ecb38);其他疗法是克唑替尼、厄洛替尼、西妥昔单抗,以及克唑替尼与厄洛替尼的组合)。图10.skmes-hgf移植瘤被植入scidbeige小鼠中并且这些小鼠用不同疗法治疗。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。以20mg/kg、5mg/kg或1mg/kg一周两次腹膜内(i.p.)给予双特异性egfr/c-met抗体em1-mab;以20mg/kg一周两次腹腔内给予西妥昔单抗。图中的箭头示出了施用天数。抗体后的数字指示施用剂量。图11.hcc827移植瘤被植入裸小鼠中并且这些小鼠用厄洛替尼或em1-mab以所指示的剂量治疗。em1-mab以两周一次给予并且厄洛替尼以一天一次给予,共给予四周。图中的箭头示出了施用天数。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。图12.snu-5移植瘤被植入cb17/scid小鼠中并且这些小鼠用10mg/kg西妥昔单抗或10mg/kg或1mg/kgem1-mab治疗。抗体以两周一次给予,共给予四周。图中的箭头示出了施用天数。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。图13.h1975-hgf移植瘤被植入裸小鼠中并且这些小鼠用10mg/kg西妥昔单抗、10mg/kgem1-mab、50mg/kg厄洛替尼、15mg/kg阿法替尼,或10mg/kgem1-mab与15mg/kg阿法替尼的组合治疗。抗体以两周一次给予并且小分子以一天一次给予,共给予三周。图中的箭头示出了施用天数。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。图14.hcc827-er1移植瘤被植入裸小鼠中并且这些小鼠用10mg/kgem1-mab、25mg/kg厄洛替尼,或这两种的组合治疗。em1-mab以两周一次给予并且厄洛替尼以一天一次给予,共给予19天。图中的箭头示出了施用天数。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。图15.在施用单剂量的20mg/kgem1-mab后从植入scidbeige小鼠中的h1975hgf移植瘤分离的瘤裂解物中的平均egfr和c-met水平。受体水平在所指示的治疗后时间以占pbs对照的百分比示出。图16.h1975-hgf移植瘤被植入裸小鼠中并且这些小鼠用10mg/kgem1-mab或10mg/kg不具有fc受体结合功能并缺少效应子功能的em1-mab变体igg2v234a/g237a/p238s/h268a/v309l/a330s/p331s治疗。抗体在所指示的天数以两周一次给予。疗法的抗瘤活性以瘤尺寸(mm3)随时间推移的变化示出。具体实施方式如本文所用,术语“iii型纤连蛋白(fn3)域”(fn3域)指在蛋白质中频繁出现的域,该蛋白质包括纤连蛋白、腱生蛋白、胞内细胞骨架蛋白、细胞因子受体和原核酶(bork和doolittle,procnatacadsciusa89:8990-8994,1992;meinke等人,jbacteriol175:1910-1918,1993;watanabe等人,jbiolchem265:15659-15665,1990)。示例性的fn3域是存在于人腱生蛋白c中的15种不同的fn3域、存在于人纤连蛋白(fn)中的15种不同的fn3域以及非天然的合成fn3域,如例如美国专利公布2010/0216708中所述。单独的fn3域通过域编号和蛋白质名称来表示,例如,腱生蛋白的第3fn3域(tn3),或纤连蛋白的第10fn3域(fn10)。如本文所用,术语“替换”或“替换的”或“突变”或“突变的”指在多肽或多核苷酸序列中改变、缺失或插入一个或多个氨基酸或核苷酸以产生该序列的变体。如本文所用,术语“使随机化”或“随机化的”或“多样化的”或“使多样化”指在多核苷酸或多肽序列中进行至少一处替换、插入或缺失。如本文所用,“变体”指与参考多肽或参考多核苷酸差别在于一种或多种修饰,例如替换、插入或缺失的多肽或多核苷酸。如本文所用,术语“特异性地结合”或“特异性结合”是指fn3域、特异性结合egfr和c-met的双特异性试剂,或本发明的双特异性egfr/c-met抗体以约1×10-6m或更小、例如约1×10-7m或更小、约1×10-8m或更小、约1×10-9m或更小、约1×10-10m或更小、约1×10-11m或更小、约1×10-12m或更小或约1×10-13m或更小的离解常数(kd)结合预定抗原的能力。通常,如通过表面等离子共振使用例如proteon仪器(biorad)所测量,fn3域、特异性结合egfr和c-met的双特异性试剂,或本发明的双特异性egfr/c-met抗体以比其对非特异性抗原(例如bsa或酪蛋白)的kd小至少十倍的kd结合预定抗原(即,egfr或c-met)。因此,包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体以至少约1×10-6m或更小、例如约1×10-7m或更小、约1×10-8m或更小、约1×10-9m或更小、约1×10-10m或更小、约1×10-11m或更小、约1×10-12m或更小或约1×10-13m或更小的结合亲和力(kd)特异性结合每个egfr和c-met。然而,特异性结合预定抗原的包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体可具有与其他相关抗原例如与来自其他种类的相同预定抗原(同源物)的交叉反应性。术语“文库”是指变体的集合。文库可由多肽或多核苷酸变体组成。如本文所用,术语“稳定性”指在生理条件下分子保持折叠状态以便其保持至少一种其正常功能活性例如结合于预定抗原诸如egfr或c-met的能力。如此处所用,“表皮生长因子受体”或“egfr”是指具有以seqidno:73和genbank登录号np_005219示出的氨基酸序列的人egfr(也称为her1或erbb1(ullrich等人,nature309:418-425,1984)以及其天然存在的变体。此类变体包括熟知的egfrviii和其他选择性剪接变体(例如,如通过如下swissprot登录号标识:p00533-1(野生型;与seqidno:73和np_005219相同)、p00533-2(f404l/l405s)、p00533-3(628-705:ctgpglegcp...geapnqallr→pgneslkaml...sviitassch且706-1210缺失)、p00533-4(c628s和629-1210缺失)、变体glnq98、r266、k521、i674、g962和p988(livingston等人,niehs-snps,环境基因组计划,niehses15478)、t790m、l858r/t790m和del(e746,a750)。如本文所用,“egfr配体”涵盖egfr的所有(例如,生理)配体,包括egf、tgfα、肝素结合egf(hb-egf)、双调蛋白(ar)和上皮调节蛋白(epi)。如本文所用,“表皮生长因子”(egf)是指具有以seqidno:74示出的氨基酸序列的熟知53氨基酸人egf。如本文所用,“肝细胞生长因子受体”或“c-met”是指具有以seqidno:101或genbank登录号np_001120972示出的氨基酸序列的人c-met以及其天然变体。如本文所用,“肝细胞生长因子”(hgf)是指具有以seqidno:102示出的氨基酸序列的熟知人hgf,其被裂解而形成由二硫键连接的α和β链的二聚体。如本文可互换使用的“阻碍结合”或“抑制结合”是指fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂,或本发明的双特异性egfr/c-met抗体阻碍或抑制egfr配体结合(诸如egf与egfr结合和/或hgf与c-met结合)的能力,并且涵盖部分和完全阻碍/抑制两者。fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体对egfr配体(诸如egf与egfr结合和/或hgf与c-met结合)的阻碍/抑制,与egfr配体结合于egfr和/或hgf结合于c-met而不受阻碍或抑制相比,部分或完全降低了egfr信号传导和/或c-met信号传导的正常水平。当抑制为至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%时,fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体使egfr配体(诸如egf与egfr结合和/或hgf与c-met结合)“结合受阻”。结合的抑制可使用熟知的方法测量,例如使用facs并且使用本文所述的方法测量暴露于fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体的表达egfr的a431细胞上生物素酰化egf的结合的抑制,或使用熟知的方法和本文所述的方法测量c-met胞外域上生物素酰化hgf的结合的抑制。术语“egfr信号传导”是指egfr配体结合于egfr所诱导的信号转导,导致egfr中的至少一个酪氨酸残基的自磷酸化。示例性的egfr配体是egf。如本文所用,“中和egfr信号传导”是指fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体将egfr配体诸如egf所诱导的egfr信号传导抑制至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的能力。术语“c-met信号传导”是指hgf结合于c-met所诱导的信号转导,导致c-met中的至少一个酪氨酸残基的自磷酸化。通常,在hgf结合时,在位置1230、1234、1235或1349的至少一个酪氨酸残基被自磷酸化。如本文所用,“中和c-met信号传导”是指fn3域、包含双特异性egfr/c-metfn3域的分子、特异性结合egfr和c-met的双特异性试剂或本发明的双特异性egfr/c-met抗体将hgf所诱导的c-met信号传导抑制至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的能力。如本文可互换使用的“过表达”和“过表达的”是指与相同组织类型的正常细胞相比,癌细胞或恶性细胞在表面上具有可测量更高水平的egfr和/或c-met。这种过表达可由基因扩增或由增加的转录或翻译引起。可使用熟知的测定法,利用例如elisa、免疫荧光、流式细胞术或放射免疫测定法对活细胞或裂解细胞测量egfr和/或c-met表达和过表达。另选地或除此以外,可例如使用荧光原位杂交、dna印迹或pcr技术在细胞中测量编码egfr和/或c-met的核酸分子的水平。当与正常细胞相比,细胞的表面上的egfr和/或c-met的水平高至少1.5倍时,egfr和/或c-met被过表达。如本文所用,“tencon”是指具有以seqidno:1示出的序列并且在美国专利公布us2010/0216708中描述的合成的iii型纤连蛋白(fn3)域。如本文所用,“癌细胞”或“瘤细胞”是指体内、离体或组织培养物中的癌变细胞、癌前细胞或转化细胞,其具有不一定涉及新遗传物质摄入的自发或诱导表型变化。虽然转化可由感染转化病毒和掺入新基因组核酸或摄入外源核酸引起,但也可自发引起或在暴露于致癌物之后引起,从而使内源基因突变。转化/癌的例子有例如合适动物宿主诸如裸小鼠体外、体内和离体的形态学变化、细胞永生化、异常细胞控制、病灶形成、增殖、恶性化、瘤特异性标记水平、侵袭性、瘤生长或抑制(freshney,cultureofanimalcells:amanualofbasictechnique(第3版1994))。术语“载体”是指能够在生物系统内复制或可以在此类系统间移动的多核苷酸。载体多核苷酸通常含有诸如复制起点、聚腺苷酸化信号或选择标记之类的元件,其功能是促进这些多核苷酸在生物系统中的复制或保持。此类生物系统的例子可包括细胞、病毒、动物、植物和用能够复制载体的生物组分再造的生物系统。包含载体的多核苷酸可以是dna或rna分子或这些分子的杂合分子。术语“表达载体”是指可用于在生物系统或再造生物系统中指导由存在于表达载体中的多核苷酸序列所编码的多肽的翻译的载体。术语“多核苷酸”是指包含由糖-磷酸主链或其它等同的共价化学方式所共价连接的核苷酸链的分子。双链dna和单链dna以及双链rna和单链rna是多核苷酸的典型例子。“互补dna”或“cdna”是指具有存在于天然成熟mrna种类中的序列元件的布置方式的熟知合成多核苷酸,其含有连续外显子,并且存在于基因组dna中的间插内含子被移除。编码起始子甲硫氨酸的密码子可存在于或可不存在于cdna中。cdna可例如通过逆转录或合成基因组装来合成。如本文所用,“合成的”或“非天然的”或“人工的”是指多核苷酸或多肽分子不存在于自然界。术语“多肽”或“蛋白质”是指包含至少两个由肽键连接以形成多肽的氨基酸残基的分子。少于约50个氨基酸的小多肽可以称作“肽”。如本文所用,术语“双特异性egfr/c-met分子”或“包含双特异性egfr/c-metfn3域的分子”是指包含结合egfr的fn3域和不同的结合c-met的fn3域的分子,这两个域直接或经由接头共价连接在一起。示例性的结合egfr/c-met的双特异性分子包含特异性结合egfr的第一fn3域以及特异性结合c-met的第二fn3域。如本文所用,“价”是指分子中存在对抗原具有特异性的指定数量的结合位点。因此,术语“单价”、“二价”、“四价”和“六价”是指分子中分别存在对抗原具有特异性的一个、两个、四个和六个结合位点。如本文所用,“混合物”是指未共价连接在一起的两个或更多个fn3域的样品或制备物。混合物可由两个或更多个相同的fn3域或不同的fn3域组成。如本文所用,混合物也是指单价指向egfr和/或单价指向c-met的两个或更多个单价抗体的样品或制备物。如本文所用,术语“特异性结合egfr和c-met的双特异性试剂”是指包含特异性结合egfr的第一域和特异性结合c-met的第二域的分子。示例性的特异性结合egfr和c-met的试剂是双特异性抗体。另一个示例性的特异性结合egfr和c-met的双特异性试剂是包含结合egfr的fn3域和不同的结合c-met的fn3域的分子。特异性结合egfr和c-met的双特异性试剂可由单种多肽或不止一种多肽构成。如本文所用,术语“双特异性抗egfr/c-met抗体”或“双特异性egfr/c-met抗体”是指具有特异性结合egfr的第一域和特异性结合c-met的第二域的双特异性抗体。特异性结合egfr和c-met的域通常为vh/vl对,并且双特异性抗egfr/c-met抗体就结合于egfr和c-met而言是单价的。如本文所用,术语“抗体”含义广泛且包括:免疫球蛋白分子,包括多克隆抗体;单克隆抗体,包括鼠、人、人适应性、人源化和嵌合单克隆抗体;抗体片段;双特异性或多特异性抗体;二聚体、四聚体或多聚体抗体;以及单链抗体。免疫球蛋白可根据重链恒定域氨基酸序列被指定为五种主要种类,即iga、igd、ige、igg和igm。iga和igg进一步亚分类为同种型iga1、iga2、igg1、igg2、igg3和igg4。任何脊椎动物种类的抗体轻链可基于其恒定域的氨基酸序列被指定至两种明显不同类型之一,即κ(κ)和λ(λ)。术语“抗体片段”是指免疫球蛋白分子的一部分,其保留重链和/或轻链抗原结合位点,诸如重链互补决定区(hcdr)1、2和3,轻链互补决定区(lcdr)1、2和3,重链可变区(vh)或轻链可变区(vl)。抗体片段包括fab片段,即由vl、vh、cl和chi域组成的单价片段;f(ab)2片段,即包含通过铰链区二硫桥连接的两个fab片段的二价片段;由vh和chi域组成的fd片段;由抗体的单条臂的vl和vh域组成的fv片段;由vh域组成的域抗体(dab)片段(ward等人,(1989)nature341:544-546)。vh和vl域可经工程改造并经由合成接头连接在一起以形成各种类型的单链抗体设计,其中在vh和vl域由单独的单链抗体构建体表达的情况下,vh/vl域在分子内或分子间配对,以形成单价抗原结合位点,诸如单链fv(scfv)或双价抗体(diabody);如例如在pct国际公布wo1998/44001、wo1988/01649、wo1994/13804和wo1992/01047中所述。使用本领域的技术人员已知的熟知技术获得这些抗体片段,并筛选出效用方式与全长抗体相同的片段。短语“分离的抗体”是指基本上不含具有不同抗原特异性的其他抗体的抗体或抗体片段(例如,分离的特异性结合egfr和c-met的双特异性抗体基本上不含特异性结合除人egfr和c-met之外的抗原的抗体)。然而,分离的特异性结合egfr和c-met的抗体可具有与其他抗原的交叉反应性,其他抗原诸如为人egfr和/或c-met的直系同源物,诸如食蟹猴(macacafascicularis)(猕猴)egfr和/或c-met。此外,分离的抗体可基本上不含其他细胞物质和/或化学物质。抗体可变区由插入有三个“抗原结合位点”的“框架”区组成。使用不同术语来定义抗原结合位点:(i)三个在vh(hcdr1、hcdr2、hcdr3)中且三个在vl(lcdr1、lcdr2、lcdr3)中的互补决定区(cdr)是基于序列变异性(wu和kabat,jexpmed132:211-50,1970;kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md.,1991)。(ii)三个在vh(h1、h2、h3)中且三个在vl(l1、l2、l3)中的“超变区”、“hvr”或“hv”是指抗体可变域中的在结构上超变的区域,如chothia和lesk所定义(chothia和lesk,molbiol196:901-17,1987)。其他术语包括“imgt-cdr”(lefranc等人,devcomparatimmunol27:55-77,2003)和“specificitydeterminingresidueusage”(sdru)(almagro,molrecognit17:132-43,2004)。internationalimmunogenetics(imgt)数据库(http://www_imgt_org)提供了抗原结合位点的标准化编号和定义。cdr、hv与imgt划分之间的对应性描述于lefranc等人,devcomparatimmunol27:55-77,2003。如本文所用,“chothia残基”是抗体vl和vh残基,其根据al-lazikani来编号(al-lazikani等人,jmolbiol273:927-48,1997)。“框架”或“框架序列”是可变区的除了被定义为抗原结合位点的那些序列之外的其余序列。因为抗原结合位点可以由如上所述的不同术语定义,所以框架的精确氨基酸序列取决于如何定义抗原结合位点。“人源化抗体”是指其中抗原结合位点来源于非人种类且可变区框架来源于人免疫球蛋白序列的抗体。人源化抗体在框架区中可包括替换,因此该框架可能不是表达的人免疫球蛋白或种系基因序列的精确拷贝。“人适应性”抗体或“人框架适应性(hfa)”抗体是指根据美国专利公布us2009/0118127所述的方法调整的人源化抗体。通过如下方式将人适应性抗体人源化:基于cdr1和cdr2环及一部分轻链cdr3环的最大cdr与fr相似性、长度相容性和序列相似性,来选择受体人框架。“人抗体”是指具有其中框架和抗原结合位点两者来源于人源序列的重链可变区和轻链可变区的抗体。如果抗体包含恒定区,则恒定区也来源于人源序列。如果抗体的可变区得自使用人胚系免疫球蛋白或重排免疫球蛋白基因的系统,则人抗体包含“来源于”人源序列的重链可变区或轻链可变区。此类系统包括噬菌体上展示的人免疫球蛋白基因文库,以及转基因非人动物诸如本文所述携带人免疫球蛋白基因文库的小鼠。当与人胚系或重排免疫球蛋白序列相比时,由于例如天然存在的体细胞突变或者框架或抗原结合位点中替换的有意引入,“人抗体”可包含氨基酸差异。通常,“人抗体”在氨基酸序列方面与由人胚系或重排免疫球蛋白基因所编码的氨基酸序列至少约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%相同。在一些情况下,“人抗体”可包含来源于人框架序列分析的共有框架序列,例如如knappik等人,jmolbiol296:57-86,2000所述,或包含掺入噬菌体上展示的人免疫球蛋白基因文库中的合成hcdr3,例如如shi等人,jmolbiol397:385-96,2010和国际专利公布wo2009/085462中所述。“人抗体”的定义并不包括抗原结合位点来源于非人种类的抗体。分离的人源化抗体可为合成的。人抗体虽然来源于人免疫球蛋白序列,但可使用系统诸如噬菌体展示掺入性合成cdr和/或合成框架生成,或可经受体外诱变以改善抗体性质,从而产生不天然存在于体内人抗体胚系库(repertoire)内的抗体。如本文所用,术语“重组抗体”包括通过重组方式制备、表达、产生或分离的所有抗体,诸如从对于人免疫球蛋白基因而言转基因或转染色体的动物(例如,小鼠)或由其制备的杂交瘤(下文将进一步描述)分离的抗体,从经转化而表达抗体的宿主细胞分离的抗体,从重组的组合抗体文库分离的抗体,以及通过涉及将人免疫球蛋白基因序列与其他dna序列剪接在一起的任何其他方式制备、表达、产生或分离的抗体,或使用fab臂交换体外生成的抗体。如本文所用,术语“单克隆抗体”是指单一分子组成的抗体分子的制备物。单克隆抗体组合物显示出对于特定表位的单一结合特异性和亲和力,或在双特异性单克隆抗体而言,显示出对于两个不同表位的双重结合特异性。如本文所用,术语“基本上相同”意指所比较的两个抗体可变区氨基酸序列是相同的或具有“非实质差异”。非实质差异是在抗体可变区序列中不会不利地影响抗体特性的1、2、3、4、5、6、7、8、9、10、11、12、13、14或15个氨基酸的替换。与本文所公开的可变区序列基本上相同的氨基酸序列在本发明范围内。在一些实施例中,序列同一性可以是约90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高。可使用vectorntiv.9.0.0(invitrogen,carlsbad,ca)的alignx模块的默认设置,例如通过逐对比对来测定百分比同一性。本发明的蛋白质序列可用作执行公共或专利数据库检索的查询序列,以例如识别相关序列。用于执行此类检索的示例性程序为使用默认设置的xblast或blastp程序(http_//www_ncbi_nlm/nih_gov)或genomequesttm(genomequest,westborough,ma)套件。如本文所用,术语“表位”是指抗体特异性结合的抗原部分。表位通常由化学活性的(诸如极性、非极性或疏水性)表面部分组而组成,诸如氨基酸或多糖侧链,并且可具有特定的三维结构特征,以及特定的电荷特征。表位可由形成构象空间单元的连续和/或不连续氨基酸构成。对于不连续表位而言,抗原线性序列不同部分的氨基酸,通过蛋白质分子的折叠,在三维空间中紧密靠近。如本文所用,术语“联用”是指两种或更多种治疗剂可以在混合物中、作为单一药剂同时地或作为单一药剂以任何顺序先后地共同向受检者施用。除非另有明确规定,否则在本说明书通篇中,抗体恒定区中的氨基酸残基的编号根据eu索引进行,如kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md,(1991)中所述。物质的组成本发明提供特异性结合egfr和c-met的双特异性试剂。本发明提供多肽和编码本发明双特异性试剂的多核苷酸或其互补核酸、载体、宿主细胞以及制备和使用它们的方法。包含单特异性和双特异性egfr和/或c-metfn3域的结合分子包含单特异性egfrfn3域的结合分子本发明提供iii型纤连蛋白(fn3)域,其特异性结合于表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr,从而可广泛用于治疗和诊断应用。本发明提供编码本发明fn3域的多核苷酸或其互补核酸、载体、宿主细胞以及制备和使用它们的方法。本发明的fn3域以高亲和力结合egfr并抑制egfr信号传导,并且可提供与小分子egfr抑制剂相比在特异性和降低的脱靶毒性方面的有益效果,以及与常规抗体治疗剂相比改善的组织渗透性。本发明的一个实施例是特异性结合表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr的分离的iii型纤连蛋白(fn3)域。在采用a431细胞并在与或不与本发明fn3域一起温育的情况下使用600nm链霉亲和素-藻红蛋白缀合物检测来自a431细胞上的结合生物素酰化egf的荧光量的竞争测定法中,本发明的fn3域可以小于约1×10-7m、小于约1×10-8m、小于约1×10-9m、小于约1×10-10m、小于约1×10-11m或小于约1×10-12m的ic50值阻碍egf结合于egfr。示例性的fn3域可以约1×10-9m至约1×10-7m之间的ic50值阻碍egf结合于egfr,诸如具有seqidno:18-29、107-110或122-137的氨基酸序列的结合egfr的fn3域。与在不存在本发明fn3域的情况下使用相同测定法条件时egf对egfr的结合相比,本发明的fn3域可将egf对egfr的结合阻碍至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。与在不存在本发明fn3域的情况下使用相同测定法条件时的信号传导水平相比,本发明的fn3域可将egfr信号传导抑制至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。配体诸如egf与egfr的结合刺激受体二聚化、自磷酸化、受体的内部细胞质酪氨酸激酶域的激活,以及多个参与dna合成(基因激活)和细胞周期进程或分裂的调控的信号转导和反式激活途径的引发。egfr信号传导的抑制可导致一个或多个egfr下游信号传导途径受抑制,因此中和egfr可具有各种效应,包括细胞增殖和分化、血管生成、细胞运动和转移的抑制。egfr信号传导可使用各种熟知的方法测量,例如测量受体在酪氨酸y1068、y1148和y1173任一者处的自磷酸化(downward等人,nature311:483-5,1984)和/或天然或合成底物的磷酸化。磷酸化可使用熟知的方法检测,诸如采用磷酸化酪氨酸特异性抗体的elisa测定法或蛋白质印迹。示例性的测定法可见于panek等人,jpharmacolexpthera283:1433-44,1997和batley等人,lifesci62:143-50,1998以及本文所述的测定法。在一个实施例中,当在a431细胞中使用50ng/ml人egf测量时,本发明的fn3域以小于约2.5×10-6m,例如小于约1×10-6m、小于约1×10-7m、小于约1×10-8m、小于约1×10-9m、小于约1×10-10m、小于约1×10-11m或小于约1×10-12m的ic50值抑制在egfr残基位置第1173位酪氨酸处egf诱导的egfr磷酸化。在一个实施例中,当在a431细胞中使用50ng/ml人egf测量时,本发明的fn3域以在约1.8×10-8m至约2.5×10-6m之间的ic50值抑制在egfr残基位置第1173位酪氨酸处egf诱导的egfr磷酸化。此类示例性的fn3域是具有seqidno:18-29、107-110或122-137的氨基酸序列的那些。在一个实施例中,如通过表面等离子共振或kinexa方法所测定,如本领域技术人员所实践,本发明的fn3域以小于约1×10-8m,例如小于约1×10-9m、小于约1×10-10m、小于约1×10-11m、小于约1×10-12m或小于约1×10-13m的离解常数(kd)结合人egfr。在一些实施例中,本发明的fn3域以约2×10-10至约1×10-8m之间的kd结合人egfr。fn3域对egfr的亲和力可使用任何合适的方法通过实验测定。(参见例如,berzofsky等人,“antibody-antigeninteractions”,载于fundamentalimmunology,paul,w.e.编辑,ravenpress:newyork,ny(1984);kuby,janisimmunology,w.h.freemanandcompany:newyork,ny(1992);以及本文所述的方法)。如果在不同的条件(例如,同渗容摩、ph)下测量,特定fn3域-抗原相互作用的测量亲和力可变化。因此,亲和力和其他抗原结合参数(例如,kd、kon、koff)的测量优选地用蛋白质支架和抗原的标准化溶液,以及标准化缓冲液,诸如本文所述的缓冲液进行。示例性的结合egfr的本发明fn3域包括seqidno:18-29、107-110或122-137的fn3域。在一个实施例中,特异性结合egfr的fn3域包含与seqidno:27的氨基酸序列至少87%相同的氨基酸序列。在一个实施例中,特异性结合egfr的fn3域包含:fg环,该fg环包含序列hnvykdtnx9rgl(seqidno:179)或序列lgsyvfehdvml(seqidno:180),其中x9为m或i;和bc环,该bc环包含序列x1x2x3x4x5x6x7x8(seqidno:181);其中x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;并且x8为y、f或l。特异性结合egfr并抑制egfr的自磷酸化的本发明fn3域可包含作为结构特征的fg环,该fg环包含序列hnvykdtnx9rgl(seqidno:179)或序列lgsyvfehdvml(seqidno:180),其中x9为m或i。此类fn3域还可包含8或9个氨基酸长并由序列x1x2x3x4x5x6x7x8(seqidno:181)限定的bc环,并且当在a431细胞中使用50ng/ml人egf测量时,以小于约2.5×10-6m的ic50值,或在约1.8×10-8m至约2.5×10-6m之间的ic50值抑制egfr自磷酸化。特异性结合egfr并抑制egfr的自磷酸化的本发明fn3域还包含如下序列:lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvhnvykdtnx9rglplsaeftt(seqidno:182),或序列lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvlgsyvfehdvmlplsaeftt(seqidno:183),其中x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;x8为y、f或l;并且x9为m或i可使用熟知的方法和本文所述的方法生成结合egfr的fn3域并测试其抑制egfr自磷酸化的能力。本发明的另一个实施例是特异性结合egfr的分离的fn3域,其中fn3域包含以seqidno:18-29、107-110或122-137示出的序列。在一些实施例中,结合egfr的fn3域包含连接到特定fn3域n-末端的起始子甲硫氨酸(met)或连接到c-末端的半胱氨酸(cys),例如以有利于半衰期延长分子的表达和/或缀合。本发明的另一个实施例是特异性结合egfr并阻碍egf结合于egfr的分离的iii型纤连蛋白(fn3)域,其中fn3域分离自基于seqidno:1的tencon序列而设计的文库。包含单特异性c-metfn3域的结合分子本发明提供iii型纤连蛋白(fn3)域,其特异性结合于肝细胞生长因子受体(c-met)并阻碍肝细胞生长因子(hgf)结合于c-met,从而可广泛用于治疗和诊断应用。本发明提供编码本发明fn3域的多核苷酸或其互补核酸、载体、宿主细胞以及制备和使用它们的方法。本发明的fn3域以高亲和力结合c-met并抑制c-met信号传导,并且可提供与小分子c-met抑制剂相比在特异性和降低的脱靶毒性方面的有益效果,以及与常规抗体治疗剂相比改善的组织渗透性。本发明的fn3域是单价的,因此防止了其他二价分子可能出现的不期望的受体聚簇和激活。本发明的一个实施例是特异性结合肝细胞生长因子受体(c-met)并阻碍肝细胞生长因子(hgf)结合于c-met的分离的iii型纤连蛋白(fn3)域。在检测在存在本发明fn3域的情况下对生物素酰化hgf结合于c-met-fc融合蛋白的抑制的测定法中,本发明的fn3域可以小于约1×10-7m、小于约1×10-8m、小于约1×10-9m、小于约1×10-10m、小于约1×10-11m或小于约1×10-12m的ic50值阻碍hgf结合于c-met。示例性的fn3域可以约2×10-10m至约6×10-8m之间的ic50值阻碍hgf结合于c-met。与在不存在本发明的fn3域的情况下使用相同测定法条件时hgf对c-met的结合相比,本发明的fn3域可将hgf结合于c-met阻碍至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。与在不存在本发明fn3域的情况下使用相同测定法条件时的信号传导水平相比,本发明的fn3域可将c-met信号传导抑制至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。hgf与c-met的结合刺激受体二聚化、自磷酸化、受体的内部细胞质酪氨酸激酶域的激活,以及多个参与dna合成(基因激活)和细胞周期进程或分裂的调控的信号转导和反式激活途径的引发。c-met信号传导的抑制可导致一个或多个c-met下游信号传导途径受抑制,因此中和c-met可具有各种效应,包括细胞增殖和分化、血管生成、细胞运动和转移的抑制。c-met信号传导可使用各种熟知的方法测量,例如测量受体在至少一个酪氨酸残基y1230、y1234、y1235或y1349上的自磷酸化和/或天然或合成底物的磷酸化。可例如在elisa测定法中或在蛋白质印迹上使用对磷酸化酪氨酸具有特异性的抗体来检测磷酸化。酪氨酸激酶活性的测定法描述于例如:panek等人,jpharmacolexpthera283:1433-44,1997和batley等人,lifesci62:143-50,1998以及本文所述的测定法。在一个实施例中,当在nci-h441细胞中使用100ng/ml重组人hgf测量时,本发明的fn3域以小于约1×10-6m、小于约1×10-7m、小于约1×10-8m、小于约1×10-9m、小于约1×10-10m、小于约1×10-11m或小于约1×10-12m的ic50值抑制在c-met残基位置1349处hgf诱导的c-met磷酸化。在一个实施例中,当在nci-h441细胞中使用100ng/ml重组人hgf测量时,本发明的fn3域以在约4×10-9m至约1×10-6m之间的ic50值抑制在c-met酪氨酸y1349处hgf诱导的c-met磷酸化。在一个实施例中,如通过表面等离子共振或kinexa方法所测定,如本领域技术人员所实践,本发明的fn3域以等于或小于约1×10-7m、1×10-8m、1×10-9m、1×10-10m、1×10-11m、1×10-12m、1×10-13m、1×10-14m或1×10-15m的离解常数(kd)结合人c-met。在一些实施例中,本发明的fn3域以约3×10-10m至约5×10-8m之间的kd结合人c-met。fn3域对c-met的亲和力可使用任何合适的方法通过实验测定。(参见例如,berzofsky等人,“antibody-antigeninteractions”,载于fundamentalimmunology,paul,w.e.编辑,ravenpress:newyork,ny(1984);kuby,janisimmunology,w.h.freemanandcompany:newyork,ny(1992);以及本文所述的方法)。如果在不同的条件(例如,同渗容摩、ph)下测量,特定fn3域-抗原相互作用的测量亲和力可变化。因此,亲和力和其他抗原结合参数(例如,kd、kon、koff)的测量优选地用蛋白质支架和抗原的标准化溶液,以及标准化缓冲液,诸如本文所述的缓冲液进行。示例性的结合c-met的本发明fn3域包括具有seqidno:32-49或111-114的氨基酸序列的fn3域。在一个实施例中,特异性结合c-met的fn3域包含与seqidno:41的氨基酸序列至少83%相同的氨基酸序列。在一个实施例中,特异性结合c-met的fn3域包含:c链和cd环,该c链和cd环包含序列dsfx10iryx11ex12x13x14x15gx16(seqidno:184),其中x10为w、f或v;x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;并且x16为e或d;和f链和fg环,该f链和fg环包含序列teyx17vx18ix19x20vkggx21x22sx23(seqidno:185),其中x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。特异性结合c-met并抑制c-met的自磷酸化的本发明fn3域还包含如下序列:lpapknlvvsrvtedsarlswtapdaafdsfx10iryx11ex12x13x14x15gx16aivltvpgsersydltglkpgteyx17vx18ix19x20vkggx21x22sx23plsaeftt(seqidno:186),其中x10为w、f或v;并且x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;x16为e或d;x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。本发明的另一个实施例是特异性结合c-met的分离的fn3域,其中fn3域包含以seqidno:32-49或111-114示出的序列。本发明的另一个实施例是特异性结合c-met并阻碍hgf结合于c-met的分离的iii型纤连蛋白(fn3)域,其中fn3域分离自基于seqidno:1的tencon序列而设计的文库。egfr或c-metfn3域从基于tencon序列的文库的分离tencon(seqidno:1)是由来自人腱生蛋白-c的十五个fn3域的共有序列而设计的非天然存在的iii型纤连蛋白(fn3)域(jacobs等人,proteinengineering,design,andselection,25:107-117,2012;美国专利公布2010/0216708)。tencon的晶体结构显示连接七条β-链的六个表面外露的环,这是fn3域所特有的,该β-链称为a、b、c、d、e、f和g,并且该环称为ab、bc、cd、de、ef和fg环(bork和doolittle,procnatlacadsciusa89:8990-8992,1992;美国专利6,673,901)。这些环或每个环内的所选残基可被随机化以便构建iii型纤连蛋白(fn3)域的文库,该文库可用于选择结合egfr或c-met的新型分子。表1示出了tencon(seqidno:1)中的每个环和β-链的位置和序列。基于tencon序列而设计的文库可因此具有随机化fg环或随机化bc和fg环,诸如如下所述的文库tcl1或tcl2。tenconbc环为7个氨基酸长,因此在bc环处多样化并基于tencon序列而设计的文库中,1、2、3、4、5、6或7个氨基酸可被随机化。tenconfg环为7个氨基酸长,因此在fg环处多样化并基于tencon序列而设计的文库中,1、2、3、4、5、6或7个氨基酸可被随机化。在tencon文库中的环处的进一步多样性可通过环处的残基的插入和/或缺失来实现。例如,fg和/或bc环可延伸1-22个氨基酸,或减少1-3个氨基酸。tencon中的fg环的长度为7个氨基酸,而抗体重链中相应环的范围为4-28个残基。为提供最大的多样性,fg环可在序列以及长度方面被多样化以对应于4-28个残基的抗体cdr3长度范围。例如,fg环可进一步通过将环延伸另外1、2、3、4或5个氨基酸而在长度方面被多样化。基于tencon序列而设计的文库也可具有随机化另选表面,该表面形成于fn3域的侧面上并且包含两条或更多条β链以及至少一个环。一种此类另选表面由c和fβ-链及cd和fg环中的氨基酸形成(c-cd-f-fg表面)。基于tencon另选c-cd-f-fg表面的文库设计示于图1中,并且此类文库的详细生成描述于美国专利公布us2013/0226834。基于tencon序列而设计的文库也包括基于tencon变体而设计的文库,该tencon变体诸如为在残基位置11、14、17、37、46、73或86(残基编号对应于seqidno:1)处具有替换的tencon变体,并且该变体表现出改善的热稳定性。示例性的tencon变体描述于美国专利公布2011/0274623,并且包括与seqidno:1的tencon相比具有替换e11r、l17a、n46v和e86i的tencon27(seqidno:99)。表1.fn3域tencon(seqidno:1)a链1-12ab环13-16b链17-21bc环22-28c链29-37cd环38-43d链44-50de环51-54e链55-59ef环60-64f链65-74fg环75-81g链82-89可使用随机或限定组的氨基酸在所选残基位置处将基于tencon和其他fn3序列的文库随机化。例如,可使用nnk密码子(其编码所有20种天然存在的氨基酸)产生具有随机替换的文库中的变体。在其它多样化方案中,dvk密码子可用于编码氨基酸ala、trp、tyr、lys、thr、asn、lys、ser、arg、asp、glu、gly和cys。另选地,nns密码子可用于产生所有20种氨基酸残基并同时降低终止密码子的频率。可例如使用technology(http:_//www_sloning_com)来合成在待多样化的位置具有偏向氨基酸分布的fn3域。这种技术使用预先制备的双链三体的文库,其用作对于成千上万的基因合成工艺而言为足够的通用构建模块。该三体文库代表对构建任何期望的dna分子所必需的所有可能的序列组合。密码子命名根据熟知的iub密码。本发明的特异性结合egfr或c-met的fn3域可通过如下方式产生fn3文库诸如tencon文库来分离:使用顺式展示将编码支架蛋白质的dna片段连接到编码repa的dna片段以生成体外翻译后形成的蛋白质-dna复合物集合(pool),其中每种蛋白质与编码其的dna稳定相关联(美国专利7,842,476;odegrip等人,procnatlacadsciusa101,2806-2810,2004),并且通过本领域已知及实例中所述的任何方法测定文库对egfr和/或c-met的特异性结合。可使用的示例性熟知的方法为elisa、夹心免疫测定法和竞争性和非竞争性测定法(参见例如,ausubel等人编辑,1994,currentprotocolsinmolecularbiology,第1卷,johnwiley&sons,inc.,newyork)。使用本文所述的方法,针对其阻碍egfr配体诸如egf结合于egfr或hgf结合于c-met的能力以及其抑制egfr和/或c-met信号传导的能力,进一步表征了所鉴定的特异性结合egfr或c-met的fn3域。可通过如下方式生成本发明的特异性结合于egfr或c-met的fn3域:使用任何fn3域作为模板来生成文库,以及使用本文中提供的方法从文库筛选特异性结合egfr或c-met的分子。可使用的示例性fn3域是腱生蛋白c的第三fn3域(tn3)(seqidno:75)、fibcon(seqidno:76)以及纤连蛋白的第十fn3域(fn10)(seqidno:77)。标准的克隆和表达技术用于将文库克隆进载体或合成文库的双链cdna盒以在体外表达或翻译文库。例如,可使用核糖体展示(hanes和pluckthun,procnatlacadsciusa,94,4937-4942,1997)、mrna展示(roberts和szostak,procnatlacadsciusa,94,12297-12302,1997)或其他无细胞系统(美国专利5,643,768)。fn3域变体的文库可表达为展示在例如任何合适的噬菌体的表面上的融合蛋白。用于在噬菌体的表面上展示融合多肽的方法是熟知的(美国专利公布2011/0118144;国际专利公布wo2009/085462;美国专利6,969,108;美国专利6,172,197;美国专利5,223,409;美国专利6,582,915;美国专利6,472,147)。本发明的特异性结合egfr或c-met的fn3域可被修饰以改善其性质,诸如改善热稳定性以及热折叠和解折叠的可逆性。多种方法已应用于增加蛋白质和酶的表观热稳定性,包括基于与高度相似的热稳定序列比较的合理设计、稳定二硫桥的设计、增加α-螺旋倾向的突变、对盐桥进行工程改造、蛋白质表面电荷的改变、定向进化和共有序列的构成(lehmann和wyss,curropinbiotechnol,12,371-375,2001)。高度的热稳定性可增加所表达蛋白的产量、改善溶解度或活性、降低免疫原性并且使生产中对冷链的需要最小化。可被替换而改善tencon(seqidno:1)的热稳定性的残基是残基位置11、14、17、37、46、73或86,并且描述于美国专利公布2011/0274623。对应于这些残基的替换可被掺入本发明的fn3域或包含双特异性fn3域的分子中。本发明的另一个实施例是特异性结合egfr并阻碍egf结合于egfr的分离的fn3域,其包含以seqidno:18-29、107-110、122-137示出的序列,还包含在与tencon(seqidno:1)的位置11、14、17、37、46、73和86对应的一个或多个残基位置处的替换。本发明的另一个实施例是特异性结合c-met并阻碍hgf结合于c-met的分离的fn3域,其包含以seqidno:32-49或111-114示出的序列,还包含在与tencon(seqidno:1)的位置11、14、17、37、46、73和86对应的一个或多个残基位置处的替换。示例性的替换是替换e11n、e14p、l17a、e37p、n46v、g73y和e86i(根据seqidno:1编号)。在一些实施例中,本发明的fn3域包含与tencon(seqidno:1)中的替换l17a、n46v和e86i对应的替换。特异性结合egfr的fn3域(图1)具有与tencon(seqidno:1)相比延伸的fg环。因此,与tencon(seqidno:1)中的残基11、14、17、37、46、73和86对应的残基是图1a和1b所示的egfrfn3域中的残基11、14、17、37、46、73和91,但例外的是seqidno:24的fn3域,其中对应残基是残基11、14、17、38、74和92,这是由于bc环中插入了一个氨基酸。本发明的另一个实施例是特异性结合egfr并阻碍egf结合于egfr的分离的fn3域,其包含以seqidno:18-29、107-110或122-137示出的氨基酸序列,任选地具有与tencon(seqidno:1)中的替换l17a、n46v和e86i对应的替换。本发明的另一个实施例是特异性结合c-met并阻碍hgf结合于c-met的分离的fn3域,其包含以seqidno:32-49或111-114示出的氨基酸序列,任选地具有与tencon(seqidno:1)中的替换l17a、n46v和e86i对应的替换。蛋白稳定性和蛋白易变性的测量可视为蛋白完整性的相同或不同方面。蛋白对由热、由紫外线或电离辐射、环境同渗容摩和ph(如果在液体溶液中)的变化、由小孔径过滤施加的机械剪切力、紫外线辐射、电离辐射(诸如由γ辐射)、化学或热脱水或任何其它可造成蛋白结构破坏的作用或力引起的变性敏感或“易变”。可使用标准方法测定分子的稳定性。例如,可通过使用标准方法测量热解链(“tm”)温度,即一半的分子变为解折叠的以摄氏度(℃)表示的温度来测定分子的稳定性。通常,tm越高,分子越稳定。除了热之外,化学环境也改变蛋白的稳定性以保持特定的三维结构。在一个实施例中,由tm的增加所测量,与工程改造之前的相同域相比,本发明的结合egfr或c-met的fn3域表现出稳定性增加至少5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%或95%或更多。可通过多种方法同样地测量化学变性。化学变性剂包括盐酸胍、硫氰酸胍、脲、丙酮、有机溶剂(dmf、苯、乙腈)、盐(硫酸铵、溴化锂、氯化锂、溴化钠、氯化钙、氯化钠);还原剂(例如二硫苏糖醇、β-巯基乙醇、二硝基硫苯和氢化物,诸如硼氢化钠)、非离子和离子型去污剂、酸(例如盐酸(hcl)、乙酸(ch3cooh)、卤代乙酸)、疏水分子(例如,磷脂)和靶向变性剂。变性程度的定量可依赖于功能特性的丧失,诸如结合靶分子的能力,或通过物理化学特性定量,诸如聚集的趋势、先前溶剂不可及残基的暴露或二硫键的破坏或形成。在一个实施例中,如通过使用盐酸胍作为化学变性剂所测量,与工程改造之前的相同支架相比,本发明的结合egfr或c-met的fn3域表现出稳定性增加至少5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%或95%或更多。使用熟知的方法,在用增加浓度的盐酸胍处理后,增加的稳定性可测量为降低的色氨酸荧光的函数。本发明的fn3域可作为单体、二聚体或多聚体产生,例如作为增加化合价和因此靶分子结合的亲和力的手段,或产生同时结合两种或多种不同的靶分子双特异性或多特异性支架的手段。二聚体和多聚体可通过连接单特异性、双特异性或多特异性蛋白质支架来产生,例如,通过纳入氨基酸接头,例如含有聚甘氨酸、甘氨酸和丝氨酸或丙氨酸和脯氨酸的接头。示例性的接头包括(gs)2、(seqidno:78)、(ggggs)5(seqidno:79)、(ap)2(seqidno:80)、(ap)5(seqidno:81)、(ap)10(seqidno:82)、(ap)20(seqidno:83)和a(eaaak)5aaa(seqidno:84)接头。二聚体和多聚体可彼此以n到c方向连接。使用天然存在的以及人工肽接头将多肽连接成新型连接的融合多肽在文献中为熟知的(hallewell等人,jbiolchem264,5260-5268,1989;alfthan等人,proteineng.8,725-731,1995;robinson&sauer,biochemistry35,109-116,1996;美国专利5,856,456)。特异性结合egfr和c-met的双特异性试剂本发明的特异性结合egfr和c-met的双特异性试剂可提供与小分子egfr和/或c-met抑制剂相比在特异性和降低的脱靶毒性方面的有益效果。本发明至少部分地基于这样令人惊讶的发现:当与结合egfr和结合c-met的单特异性试剂的混合物相比时,特异性结合egfr和c-met的双特异性试剂提供显著改善的协同抑制效应。分子可被调制为对于egfr和c-met两者具有特定亲和力,以使瘤渗透和潴留最大化。与西妥昔单抗相比,特异性结合egfr和c-met的双特异性试剂提供egfr和/或c-met信号传导途径的更有效抑制并更有效地抑制瘤生长。特异性结合egfr和c-met的双特异性试剂可由包含egfr结合域和c-met结合域的任何多肽或多聚多肽形成。egfr和c-met结合域可为抗体的抗原结合位点、抗体的vh/vl对,或另一种类型的结合分子,诸如基于iii型纤连蛋白(fn3)域、ix型纤连蛋白(fn9)域或它们的任何组合的域。egfr和c-met结合多肽可来源于既有的单特异性egfr和c-met结合多肽,或者可从头分离。包含双特异性egfr/c/metfn3域的分子本发明的一个实施例是分离的包含双特异性fn3域的分子,其包含第一iii型纤连蛋白(fn3)域和第二fn3域,其中第一fn3域特异性结合表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr,并且第二fn3域特异性结合肝细胞生长因子受体(c-met)并阻碍肝细胞生长因子(hgf)结合于c-met。本发明的包含双特异性egfr/c-metfn3域的分子可通过如下方式生成:将本发明的任何结合egfr的fn3域和任何结合c-met的fn3域直接地或经由接头共价连接。因此,双特异性分子的第一fn3域可具有如上对于结合egfr的fn3域所述的特性,并且双特异性分子的第二fn3域可具有如上对于结合c-met的fn3域所述的特性。在一个实施例中,当在a431细胞中使用50ng/ml人egf测量时,包含双特异性egfr/c-metfn3域的分子的第一fn3域以小于约2.5×10-6m的ic50值抑制在egfr残基第1173位酪氨酸处egf诱导的egfr磷酸化,并且当在nci-h441细胞中使用100ng/ml人hgf测量时,包含双特异性egfr/c-metfn3域的分子的第二fn3域以小于约1.5×10-6m的ic50值抑制在c-met残基第1349位酪氨酸处hgf诱导的c-met磷酸化。在另一个实施例中,当在a431细胞中使用50ng/ml人egf测量时,包含双特异性egfr/c-metfn3域的分子的第一fn3域以在约1.8×10-8m至约2.5×10-6m之间的ic50值抑制在egfr残基第1173位酪氨酸处egf诱导的egfr磷酸化,并且当在nci-h441细胞中使用100ng/ml人hgf测量时,包含双特异性egfr/c-metfn3域的分子的第二fn3域以在约4×10-9m至约1.5×10-6m之间的ic50值抑制在c-met残基第1349位酪氨酸处hgf诱导的c-met磷酸化。在另一个实施例中,包含双特异性egfr/c-metfn3域的分子的第一fn3域以小于约1×10-8m的离解常数(kd)结合人egfr,并且包含双特异性egfr/c-metfn3域的分子的第二fn3域以小于约5×10-8m的kd结合人c-met。在结合egfr和c-met两者的双特异性分子中,第一fn3域以约2×10-10至约1×10-8m之间的kd结合人egfr,并且第二fn3域以约3×10-10至约5×10-8m之间的kd结合人c-met。双特异性egfr/c-met分子对egfr和c-met的亲和力可如上对于单特异性分子所述而测定。在采用a431细胞并在与或不与第一fn3域一起温育的情况下使用600nm链霉亲和素-藻红蛋白缀合物检测来自a431细胞上的结合生物素酰化egf的荧光量的测定法中,本发明双特异性egfr/c-met分子中的第一fn3域可以在约1×10-9m至约1.5×10-7m之间的ic50值阻碍egf结合于egfr。与在不存在第一fn3域的情况下使用相同测定法条件时egf对egfr的结合相比,本发明的双特异性egfr/c-met分子中的第一fn3域可将egf对egfr的结合阻碍至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。在检测在存在第二fn3域的情况下对生物素酰化hgf结合于c-met-fc融合蛋白的抑制的测定法中,本发明的双特异性egfr/c-met分子中的第二fn3域可以在约2×10-10m至约6×10-8m之间的ic50值阻碍hgf结合于c-met。与在不存在第二fn3域的情况下使用相同测定法条件时hgf对c-met的结合相比,双特异性egfr/c-met分子中的第二fn3域可将hgf对c-met的结合阻碍至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。与在不存在本发明的双特异性egfr/c-met分子的情况下使用相同测定法条件时信号传导的水平相比,本发明的双特异性egfr/c-met分子可将egfr和/或c-met信号传导抑制至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。可使用如上对于单特异性分子所述的各种熟知方法测量egfr和c-met信号传导。与第一和第二fn3域的混合物所观察到的协同抑制相比,包含特异性结合egfr的第一fn3域和特异性结合c-met的第二fn3域的本发明双特异性egfr/c-met分子提供egfr和c-met信号传导及瘤细胞增殖的显著增强的协同抑制。可例如通过测量包含双特异性egfr/c-metfn3域的分子及两种单特异性分子(一种结合egfr,另一种结合c-met)的混合物对erk磷酸化的抑制,来评估协同抑制。本发明的双特异性egfr/c-met分子可以与两种单特异性fn3域的混合物的ic50值相比小至少约100倍、例如小至少500、1000、5000或10,000倍的ic50值抑制erk磷酸化,指示与两种单特异性fn3域的混合物相比,包含双特异性egfr/c-metfn3域的分子效力增强至少100倍。示例性的包含双特异性egfr-c-metfn3域的分子可以约5×10-9m或更小的ic50值抑制erk磷酸化。可使用标准方法和本文所述的方法测量erk磷酸化。本发明的包含双特异性egfr/c-metfn3域的分子可以与第一fn3域和第二fn3的混合物对nci-h292细胞生长的抑制的ic50值相比小至少30倍的ic50值抑制nci-h292细胞增殖,其中用包含10%的fbs且补充有7.5ng/ml的hgf的培养基诱导细胞增殖。本发明的双特异性分子可以与第一fn3域和第二fn3域的混合物对瘤细胞增殖的抑制的ic50值相比小约30、40、50、60、70、80、90、100、150、200、300、400、500、600、700、800或约1000倍的ic50值抑制瘤细胞增殖。可使用标准方法和本文所述的方法测量瘤细胞增殖的抑制。本发明的另一个实施例是包含双特异性fn3域的分子,其包含第一iii型纤连蛋白(fn3)域和第二fn3域,其中第一fn3域特异性结合表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr,并且第二fn3域特异性结合肝细胞生长因子受体(c-met),并阻碍肝细胞生长因子(hgf)结合于c-met,其中第一fn3域包含:fg环,该fg环包含序列hnvykdtnx9rgl(seqidno:179)或序列lgsyvfehdvml(seqidno:180),其中x9为m或i;和bc环,该bc环包含序列x1x2x3x4x5x6x7x8(seqidno:181),其中x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;并且x8为y、f或l;并且第二fn3域包含:c链和cd环,该c链和cd环包含序列dsfx10iryx11ex12x13x14x15gx16(seqidno:184),其中x10为w、f或v;x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;并且x16为e或d;和f链和fg环,该f链和fg环包含序列teyx17vx18ix19x20vkggx21x22sx23(seqidno:185),其中x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。在另一个实施例中,双特异性分子包含结合具有如下序列的egfr的第一fn3域:lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvhnvykdtnx9rglplsaeftt(seqidno:182),或序列lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvlgsyvfehdvmlplsaeftt(seqidno:183),其中在seqidno:182和183中;x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;x8为y、f或l;并且x9为m或i。在另一个实施例中,双特异性分子包含结合具有如下序列的c-met的第二fn3域:lpapknlvvsrvtedsarlswtapdaafdsfx10iryx11ex12x13x14x15gx16aivltvpgsersydltglkpgteyx17vx18ix19x20vkggx21x22sx23plsaeftt(seqidno:186),其中x10为w、f或v;并且x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;x16为e或d;x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。示例性的包含双特异性egfr/c-metfn3域的分子包含以seqidno:50-72、106、118-121或138-167示出的氨基酸序列。本发明的双特异性egfr/c-met分子具有与其功能特性(诸如抑制egfr自磷酸化)相关的某些结构特性,诸如结合egfr的第一fn3域的fg环,其包含序列hnvykdtnx9rgl(seqidno:179)或序列lgsyvfehdvml(seqidno:180),其中x9为m或i。在一个实施例中,本发明的包含双特异性egfr/c-metfn3域的分子。当在h292细胞中使用50ng/ml人egf测量时,以小于约8×10-7m的ic50值抑制在egfr残基第1173位酪氨酸处egf诱导的egfr磷酸化;当在nci-h441细胞中使用100ng/ml人hgf测量时,以小于约8.4×10-7m的ic50值抑制在c-met残基第1349位酪氨酸处hgf诱导的c-met磷酸化;以小于约9.5×10-6m的ic50值抑制hgf诱导的nci-h292细胞增殖,其中用包含7.5ng的hgf的10%的fbs诱导细胞增殖;以小于约2.0×10-8m的kd结合egfr;或以小于约2.0×10-8m的kd结合c-met。在另一个实施例中,本发明的包含双特异性egfr/c-metfn3域的分子当在h292细胞中使用50ng/ml人egf测量时,以在约4.2×10-9m与8×10-7m之间的ic50抑制egfr残基第1173位酪氨酸处egf诱导的egfr磷酸化;当在nci-h441细胞中使用100ng/ml人hgf测量时,以在约2.4×10-8m至约8.4×10-7m之间的ic50值抑制在c-met残基第1349位酪氨酸处hgf诱导的c-met磷酸化;以在约2.3×10-8m至约9.5×10-6m之间的ic50值抑制hgf诱导的nci-h292细胞增殖,其中用包含7.5ng的hgf的10%的fbs诱导细胞增殖;以在约2×10-10m至约2.0×10-8m之间的kd结合egfr;或以在约1×10-9m至约2.0×10-8m之间的kd结合c-met。在一个实施例中,双特异性egfr/c-met分子包含具有如下序列的结合egfr的fn3域:lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvhnvykdtnx9rglplsaeftt(seqidno:182),其中x1为d;x2为d;x3为p;x4不存在;x5为h或w;x6为a;x7为fx8为y;并且x9为m或i;和包含如下序列的结合c-met的fn3域:lpapknlvvsrvtedsarlswtapdaafdsfx10iryx11ex12x13x14x15gx16aivltvpgsersydltglkpgteyx17vx18ix19x20vkggx21x22sx23plsaeftt(seqidno:186),其中x10为w;x11为f;x12为f;x13为v或l;x14为g或s;x15为s或k;x16为e或d;x17为v;x18为n;x19为l或m;x20为g或s;x21为s或k;x22为i;并且x23为p。示例性的双特异性egfr/c-met分子是具有以seqidno:57、61、62、63、64、65、66、67和68示出的序列的那些。本发明的双特异性分子还可包含在第一fn3域和/或第二fn3域中的与如上所述的tencon(seqidno:1)中的位置11、14、17、37、46、73和86对应的一个或多个残基位置处的替换,以及位置29处的替换。示例性的替换是替换e11n、e14p、l17a、e37p、n46v、g73y、e86i和d29e(根据seqidno:1编号)。本领域的技术人员将认识到其他氨基酸可用于替换,诸如如下文所述在其侧链中相关的氨基酸的家族内的氨基酸。可使用本文的方法测试所生成的变体的稳定性及其对egfr和/或c-met的结合。在一个实施例中,包含双特异性egfr/c-metfn3域的分子包含特异性结合egfr的第一fn3域和特异性结合c-met的第二fn3域,其中第一fn3域包含如下序列:lpapknlvvsx24vtx25dsx26rlswddpx27afyx28sfliqyqx29sekvgeaix30ltvpgsersydltglkpgteytvsiyx31vhnvykdtnx32rglplsax33ftt(seqidno:187),其中x24为e、n或r;x25为e或p;x26为l或a;x27为h或w;x28为e或d;x29为e或p;x30为n或v;x31为g或y;x32为m或i;并且x33为e或i;并且第二fn3域包含如下序列:lpapknlvvsx34vtx35dsx36rlswtapdaafdsfwiryfx37fx38x39x40gx41aix42ltvpgsersydltglkpgteyvvnix43x44vkggx45ispplsax46ftt(seqidno:188);其中x34为e、n或r;x35为e或p;x36为l或a;x37为e或p;x38为v或l;x39为g或s;x40为s或k;x41为e或d;x42为n或v;x43为l或m;x44为g或s;x45为s或k;并且x46为e或i。在其他实施例中,包含双特异性egfr/c-metfn3域的分子包含具有与seqidno:27的氨基酸序列至少87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%相同的氨基酸序列的第一fn3域,以及具有与seqidno:41的氨基酸序列至少83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%相同的氨基酸序列的第二fn3域。本发明的包含双特异性egfr/c-metfn3域的分子可被调制为对于egfr和c-met具有特定亲和力以使瘤积聚最大化。本发明的另一个实施例是分离的包含双特异性fn3域的分子,其包含第一iii型纤连蛋白(fn3)域和第二fn3域,其中第一fn3域特异性结合表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr,并且第二fn3域特异性结合肝细胞生长因子受体(c-met)并阻碍肝细胞生长因子(hgf)结合于c-met,其中第一fn3域和第二fn3域分离自基于seqidno:1的tencon序列而设计的文库。本发明的包含双特异性egfr/c-metfn3域的分子可通过如下方式生成:使用熟知的方法将本发明的结合egfr的fn3域和结合c-met的fn3域共价偶联。fn3域可经由接头连接,例如包含聚甘氨酸、甘氨酸和丝氨酸或丙氨酸和脯氨酸的接头。示例性的接头包括(gs)2、(seqidno:78)、(ggggs)5(seqidno:79)、(ap)2(seqidno:80)、(ap)5(seqidno:81)、(ap)10(seqidno:82)、(ap)20(seqidno:83)、a(eaaak)5aaa(seqidno:84)接头。使用天然存在的以及人工肽接头将多肽连接成新型连接的融合多肽在文献中为熟知的(hallewell等人,jbiolchem264,5260-5268,1989;alfthan等人,proteineng.8,725-731,1995;robinson&sauer,biochemistry35,109-116,1996;美国专利5,856,456)。本发明的双特异性egfr/c-met分子可从第一fn3域的c-末端到第二fn3域的n-末端或从第二fn3域的c-末端到第一fn3域的n-末端连接在一起。任何结合egfr的fn3域可共价连接到结合c-met的fn3域。示例性的结合egfr的fn3域是具有以seqidno:18-29、107-110和122-137示出的氨基酸序列的域,并且示例性的结合c-met的fn3域是具有以seqidno:32-49和111-114示出的氨基酸序列的域。要偶联到双特异性分子的结合egfr的fn3域可另外在其n-末端包含起始子甲硫氨酸(met)。包含双特异性egfr/c-metfn3域的分子的变体在本发明的范围内。例如,可在包含双特异性egfr/c-metfn3域的分子中作出替换,以及当与亲本分子相比时,所得变体保留对于egfr和c-met的相似选择性和效力。示例性的修饰是例如将产生具有与亲本分子类似特性的变体的保守替换。保守替换是在其侧链中相关的氨基酸家族内发生的那些。基因编码的氨基酸可被分成四个家族:(1)酸性(天冬氨酸、谷氨酸);(2)碱性(赖氨酸、精氨酸、组氨酸);(3)非极性(丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸);以及(4)不带电极性(甘氨酸、天冬酰胺、谷氨酰胺、半胱氨酸、丝氨酸、苏氨酸、酪氨酸)。苯丙氨酸、色氨酸和酪氨酸有时被共同分类为芳族氨基酸。另选地,氨基酸库(repertoire)可被分组为(1)酸性(天冬氨酸、谷氨酸);(2)碱性(赖氨酸、精氨酸、组氨酸),(3)脂族(甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、丝氨酸、苏氨酸),并且丝氨酸和苏氨酸任选地被单独分组为脂族-羟基;(4)芳族(苯丙氨酸、酪氨酸、色氨酸);(5)酰胺(天冬酰胺、谷氨酰胺);以及(6)含硫(半胱氨酸和甲硫氨酸)(stryer编辑,biochemistry,第2版,whfreemanandco.,1981)。可对包含双特异性egfr/c-metfn3域的分子作出非保守替换,其涉及不同种类氨基酸之间的氨基酸残基的替换,以改善双特异性分子的性质。可以使用本文描述的测定通过评估修饰的多肽或片段以类似于未经修饰的多肽或片段的方式产生应答的能力,容易地测定多肽或其片段的氨基酸序列中的变化是否导致功能同源物。其中已发生超过一个替换的肽、多肽或蛋白质可以容易地以相同方式进行测试。本发明的包含双特异性egfr/c-metfn3域的分子可作为二聚体或多聚体生成,例如作为增加化合价和因此靶分子结合的亲合力的手段。多聚体可通过如下方式生成:例如通过使用熟知方法引入氨基酸接头,而将一个或多个结合egfr的fn3域和一个或多个结合c-met的fn3域连接,以形成包含对于egfr或c-met具有至少双特异性的至少三个单独的fn3域的分子。本发明的另一个实施例是包含双特异性fn3域的分子,其包含第一iii型纤连蛋白(fn3)域和第二fn3域,其中第一fn3域特异性结合表皮生长因子受体(egfr)并阻碍表皮生长因子(egf)结合于egfr,并且第二fn3域特异性结合肝细胞生长因子受体(c-met),并阻碍肝细胞生长因子(hgf)结合于c-met,并且包含以seqidno:50-72、106或138-165示出的氨基酸序列。半衰期延长部分本发明的包含双特异性egfr/c-metfn3域的分子或单特异性结合egfr或c-met的fn3域可例如经由共价相互作用掺入其他亚基。在本发明的一个方面,本发明的包含双特异性egfr/c-metfn3域的分子还包含半衰期延长部分。示例性的半衰期延长部分是白蛋白、白蛋白变体、白蛋白结合蛋白质和/或域、转铁蛋白及其片段和类似物,以及fc区域。示例性的白蛋白结合域以seqidno:117示出。抗体恒定区的全部或一部分可连接至本发明的分子以赋予抗体样特性,特别是与fc区域相关的那些特性,例如fc效应子功能诸如c1q结合、补体依赖性细胞毒性(cdc)、fc受体结合、抗体依赖性细胞介导的细胞毒性(adcc)、吞噬作用,细胞表面受体(例如,b细胞受体;bcr)的下调,并且可通过修饰促成这些活性的fc中的残基来进一步修饰(对于综述,参见strohl,curropinbiotechnol.20,685-691,2009)。另外的部分可掺入到本发明的双特异性分子,诸如用于期望特性的聚乙二醇(peg)分子,诸如peg5000或peg20,000;不同链长的脂肪酸和脂肪酸酯,例如,月桂酸酯、肉豆蔻酸酯,硬脂酸酯、花生酸酯、山嵛酸酯、油酸酯、花生四烯酸、辛二酸、十四烷二酸、十八烷二酸、二十二烷二酸等、聚赖氨酸、辛烷、糖类(葡聚糖、纤维素、寡糖或多糖)。这些部分可与蛋白质支架编码序列直接融合并且可通过标准的克隆和表达技术产生。另选地,熟知的化学偶联方法可用于将这些部分连接至本发明的重组制备的分子。聚乙二醇化(pegyl)部分可通过如下方式例如被添加到本发明的双特异性或单特异性分子:使用熟知的方法,将半胱氨酸残基掺入到分子的c-末端,以及将聚乙二醇化基团连接到半胱氨酸。具有c-末端半胱氨酸的示例性双特异性分子是具有以seqinno:170-178示出的氨基酸序列的那些分子。可通过若干熟知测定法比较掺入另外部分的本发明单特异性和双特异性分子的功能性。例如,由于掺入fc域和/或fc域变体而导致的单特异性和/或双特异性分子的改变的特性可在fc受体结合测定法中使用该受体的可溶形式来测定,该受体的可溶形式诸如fcγri、fcγrii、fcγriii或fcrn受体,或使用熟知的测量例如adcc或cdc的基于细胞的测定法,或在体内模型中评价本发明分子的药代动力学特性。多核苷酸、载体、宿主细胞本发明提供编码结合egfr或结合c-met的fn3域或本发明的包含双特异性egfr/c-metfn3域的分子的核酸,作为分离的多核苷酸或作为表达载体的部分或作为线性dna序列的部分,包括用于体外转录/翻译的线性dna序列、与其组合物或定向诱变剂的原核、真核或丝状噬菌体表达、分泌和/或展示相容的载体。本文公开了某些示例性的多核苷酸,然而,其他多核苷酸也在本发明的范围内,考虑到给定表达系统中遗传密码的简并性或密码子偏好,其他多核苷酸编码结合egfr或结合c-met的fn3域或本发明的包含双特异性egfr/c-metfn3域的分子。本发明的一个实施例是分离的多核苷酸,其编码具有seqidno:18-29、107-110或122-137的氨基酸序列的特异性结合egfr的fn3域。本发明的一个实施例是分离的多核苷酸,其包含seqidno:97-98或168-169的多核苷酸序列。本发明的一个实施例是分离的多核苷酸,其编码具有以seqidno:32-49或111-114示出的序列的氨基酸序列的特异性结合c-met的fn3域。本发明的一个实施例是分离的多核苷酸,其编码具有seqidno:50-72、106、118-121或138-165的氨基酸序列的包含双特异性egfr/-c-metfn3域的分子。本发明的一个实施例是分离的多核苷酸,其包含seqidno:115-116或166-167的多核苷酸序列。本发明的多核苷酸可以通过化学合成(诸如在自动化多核苷酸合成仪上的固相多核苷酸合成)来制备,并装配成完整的单链或双链分子。另选地,本发明的多核苷酸可以通过其他技术来制备,诸如pcr之后进行常规克隆。制备或获得具有给定的已知序列的多核苷酸的技术是本领域熟知的。本发明的多核苷酸可包含至少一个非编码序列,诸如启动子或增强子序列、内含子、聚腺苷酸化信号、促进repa结合的顺式序列等。多核苷酸序列还可包含编码另外的氨基酸的另外序列,其编码例如标记或标签序列,诸如组氨酸标签或ha标签以促进蛋白质的纯化或检测;信号序列;融合蛋白伴侣诸如repa、fc;或噬菌体外壳蛋白诸如pix或piii。本发明的另一个实施例是包含本发明的至少一种多核苷酸的载体。此类载体可以是质粒载体、病毒载体、杆状病毒表达载体、基于转座子的载体或任何其它适用于通过任何手段将本发明的多核苷酸引入给定生物体或遗传背景的载体。此类载体可以是包含可控制、调节、引起或允许由这种载体编码的多肽的表达的核酸序列元件的表达载体。此类元件可包含转录增强子结合位点、rna聚合酶起始位点、核糖体结合位点以及有利于被编码多肽在给定表达系统中的表达的其它位点。此类表达系统可以是本领域熟知的基于细胞的系统或无细胞系统。本发明的另一个实施例是包含本发明载体的宿主细胞。单特异性结合egfr或结合c-met的fn3域或本发明的包含双特异性egfr/c-metfn3域的分子可任选地由细胞系、混合细胞系、永生化细胞或永生化细胞的无性系群体产生,如本领域熟知的。参见例如,ausubel等人编辑,currentprotocolsinmolecularbiology,johnwiley&sons,inc.,ny,ny(1987-2001);sambrook等人,molecularcloning:alaboratorymanual,第2版,coldspringharbor,ny(1989);harlow和lane,antibodies,alaboratorymanual,coldspringharbor,ny(1989);colligan等人编辑,currentprotocolsinimmunology,johnwiley&sons,inc.,ny(1994-2001);colligan等人,currentprotocolsinproteinscience,johnwiley&sons,ny,ny(1997-2001)。选择用于表达的宿主细胞可以是哺乳动物起源的或可选自cos-1、cos-7、hek293、bhk21、cho、bsc-1、heg2、sp2/0、hela、骨髓瘤、淋巴瘤、酵母、昆虫或植物细胞,或它们的任何衍生物、永生化细胞或转化的细胞。另选地,宿主细胞可选自不能够使多肽糖基化的种类或生物体,例如原核细胞或生物体,诸如bl21、bl21(de3)、bl21-gold(de3)、xl1-blue、jm109、hms174、hms174(de3)以及任何天然或工程改造的大肠杆菌属(e.colispp)、克雷伯氏菌属(klebsiellaspp.)或假单胞菌属(pseudomonasspp)菌株。本发明的另一个实施例是产生分离的本发明的特异性结合egfr或c-met的fn3域或分离的本发明的包含双特异性egfr/c-metfn3域的分子的方法,所述方法包括在一定条件下培养分离的本发明的宿主细胞,使得分离的特异性结合egfr或c-met的fn3域或分离的包含双特异性egfr/c-metfn3域的分子被表达,以及纯化该域或分子。特异性结合egfr或c-met的fn3域或分离的本发明的包含双特异性egfr/c-metfn3域的分子可通过熟知的方法从重组细胞培养物中纯化,例如通过蛋白质a纯化、硫酸铵或乙醇沉淀、酸提取、阴离子或阳离子交换色谱法、磷酸纤维素色谱法、疏水相互作用色谱法、亲和色谱法、羟基磷灰石色谱法和凝集素色谱法,或高效液相色谱法(hplc)。双特异性egfr/c-met抗体双特异性egfr/c-met抗体可从头生成或可根据既有单特异性抗egfr和抗c-met抗体来工程改造。可用于工程改造双特异性分子的示例性抗egfr抗体是例如帕尼单抗(abx-egf)、尼妥珠单抗、奈昔木单抗(necitumumab)、马妥珠单抗以及例如如下专利中所述的那些:美国专利us7,595,378、美国专利us7,247,301、美国专利公布us2011/0256142、美国专利us5,891,996、美国专利us5,212,290、美国专利us5,558,864或美国专利us7,589,180。例如,可使用具有以seqidno:189或191示出的氨基酸序列的抗体vh域以及具有以seqidno:190或192示出的氨基酸序列的抗体vl域。可用于工程改造双特异性分子的示例性抗c-met抗体是例如利妥木单抗(rilotumumab)、奥纳珠单抗(onartuzumab)、非妥珠单抗(ficlatuzumab)以及例如如下专利中所述的那些:pct国际公布wo2011/110642、美国专利公布us2004/0166544、pct国际公布wo2005/016382或pct国际公布wo2006/015371。例如,可使用具有以seqidno:193或195示出的氨基酸序列的抗体vh域以及具有以seqidno:194或196示出的氨基酸序列的抗体vl域。由其美国采用药名(usan)标识的抗体的重链和轻链氨基酸序列可经由美国医学协会(americanmedicalassociation)在http://_www_ama-assn_org处获得或经由cas登记号获得。单特异性结合egfr和c-met的可变域可从例如噬菌体展示文库从头选择,其中噬菌体被工程改造以表达人免疫球蛋白或其部分,诸如fab、单链抗体(scfv)或未配对或配对的抗体可变区(knappik等人,jmolbiol296:57-86,2000;krebs等人,jimmunolmeth254:67-84,2001;vaughan等人,naturebiotechnology14:309-314,1996;sheets等人,pitas(usa)95:6157-6162,1998;hoogenboom和winter,jmolbiol227:381,1991;marks等人,jmolbiol222:581,1991)并且随后被工程改造成双特异性形式。单特异性结合egfr和c-met的可变域可例如从以与噬菌体pix外壳蛋白的融合蛋白的形式表达抗体重链和轻链可变区的噬菌体展示文库分离,如shi等人,(2010)j.mol.biol.397:385-96和pct国际公布wo09/085462所述。针对与人egfr或c-met胞外域的结合来筛选抗体文库,并进一步表征所得的阳性克隆,从克隆裂解物分离fab。此类用于分离人抗体的噬菌体展示法在本领域是成熟的。参见例如:美国专利5,223,409;美国专利5,403,484;以及美国专利5,571,698、美国专利5,427,908、美国专利5,580,717、美国专利5,969,108、美国专利6,172,197、美国专利5,885,793;美国专利6,521,404;美国专利6,544,731;美国专利6,555,313;美国专利6,582,915和美国专利6,593,081。使用本文所述的方法将从头获得的结合egfr或c-met的可变区工程改造成双特异性形式。双特异性抗体形式本发明的抗体具有两个或更多个抗原结合位点并且是双特异性的。本发明的双特异性抗体包括具有全长抗体结构的抗体。如本文所用,“全长抗体”是指具有两条全长抗体重链和两条全长抗体轻链的抗体。全长抗体重链(hc)由熟知的重链可变域vh和恒定域ch1、ch2和ch3组成。全长抗体轻链(lc)由熟知的轻链可变域vl和恒定域cl组成。全长抗体可能在一条或两条重链中缺少c-末端赖氨酸(k)。术语“fab臂”或“半分子”是指特异性结合抗原的一个重链-轻链对。本发明的全长双特异性抗体可例如使用两个单特异性二价抗体之间的fab臂交换(或半分子交换)通过如下方式生成:在每个半分子中的重链ch3界面处引入替换以有利于具有不同特异性的两个抗体半分子在体外无细胞环境中或使用共表达进行的异源二聚体形成。fab臂交换反应是ch3域的二硫键异构化反应和离解-缔合的结果。亲本单特异性抗体的铰链区中的重链二硫键被还原。亲本单特异性抗体之一的所得游离半胱氨酸与第二亲本单特异性抗体分子的半胱氨酸残基形成重链间二硫键,同时亲本抗体的ch3域通过离解-缔合而释放和重新形成。fab臂的ch3域可被工程改造以有利于异源二聚化而不利于同源二聚化。所得产物是具有各自结合不同表位(即egfr上的表位和c-met上的表位)的两个fab臂或半分子的双特异性抗体。如本文所用,“同源二聚化”是指具有相同ch3氨基酸序列的两条重链的相互作用。如本文所用,“同源二聚体”是指包含具有相同ch3氨基酸序列的两条重链的抗体。如本文所用,“异源二聚化”是指具有不同ch3氨基酸序列的两条重链的相互作用。如本文所用,“异源二聚体”是指包含具有不同ch3氨基酸序列的两条重链的抗体。“杵臼”策略(参见例如pct国际公布wo2006/028936)可用于生成全长双特异性抗体。简而言之,可在影响ch3域相互作用的位置处使形成人igg中ch3域的界面的所选氨基酸突变,以促进异源二聚体形成。将具有小侧链(臼)的氨基酸引入特异性结合第一抗原的抗体的重链中,并且将具有大侧链(杵)的氨基酸引入特异性结合第二抗原的抗体的重链中。在这两种抗体共表达之后,由于具有“臼”的重链与具有“杵”的重链的优先相互作用,而形成异源二聚体。形成杵和臼的示例性ch3替换对是(表达为第一重链的第一ch3域中的修饰位置/第二重链的第二ch3域中的修饰位置):t366y/f405a、t366w/f405w、f405w/y407a、t394w/y407t、t394s/y407a、t366w/t394s、f405w/t394s和t366w/t366s_l368a_y407v。可使用其他策略,诸如使用静电相互作用通过替换一个ch3表面处的带正电残基和第二ch3表面处的带负电残基来促进重链异源二聚化,如美国专利公布us2010/0015133;美国专利公布us2009/0182127;美国专利公布us2010/028637或美国专利公布us2011/0123532中所述。在其他策略中,可通过下列替换来促进异源二聚化(表达为第一重链的第一ch3域中的修饰位置/第二重链的第二ch3域中的修饰位置):l351y_f405a_y407v/t394w、t366i_k392m_t394w/f405a_y407v、t366l_k392m_t394w/f405a_y407v、l351y_y407a/t366a_k409f、l351y_y407a/t366v_k409f、y407a/t366a_k409f,或t350v_l351y_f405a_y407v/t350v_t366l_k392l_t394w,如美国专利公布us2012/0149876或美国专利公布us2013/0195849中所述。除上述方法之外,本发明的双特异性抗体还可通过如下方式在体外无细胞环境中生成:根据国际专利公布wo2011/131746中所述的方法,在两种单特异性同源二聚抗体的ch3区中引入非对称突变,并且在允许二硫键异构化的还原条件下由两种亲本单特异性同源二聚抗体形成双特异性异源二聚抗体。在所述方法中,第一单特异性二价抗体(例如,抗c-met抗体)和第二单特异性二价抗体(例如,抗egfr抗体)被工程改造以在ch3域处具有促进异源二聚体稳定性的某些替换;将这些抗体在足以允许铰链区中的半胱氨酸进行二硫键异构化的还原条件下一起温育;从而通过fab臂交换生成双特异性抗体。温育条件可优选地恢复为非还原条件。可使用的示例性还原剂是2-巯基乙胺(2-mea)、二硫苏糖醇(dtt)、二硫赤藓糖醇(dte)、谷胱甘肽、三(2-羰基乙基)膦(tcep)、l-半胱氨酸和β-巯基乙醇,优选地为选自如下的还原剂:2-巯基乙胺、二硫苏糖醇和三(2-羰基乙基)膦。例如,可使用在存在至少25mm的2-mea的情况下或在存在至少0.5mm的二硫苏糖醇的情况下在5-8的ph下例如在7.0的ph或7.4的ph下以至少20℃的温度温育至少90分钟。双特异性egfr/c-met抗体本发明的双特异性egfr/c-met抗体可提供与小分子egfr和/或c-met抑制剂相比在特异性和降低的脱靶毒性方面的有益效果。本发明至少部分地基于这样令人惊讶的发现:当与结合egfr和结合c-met的单特异性抗体的混合物或已公布的双特异性egfr/c-met抗体相比时,本发明的双特异性egfr/c-met抗体提供显著改善的协同抑制效应。根据该测定法,所观察到的协同效应在约14倍至超过约800倍之间变化。与西妥昔单抗相比,本发明的双特异性egfr/c-met抗体提供egfr和c-met信号传导途径的更有效抑制并更有效地抑制瘤生长。本发明的双特异性egfr/c-met抗体抑制具有egfr激活突变和/或具有egfr中已知产生对酪氨酸激酶抑制剂(诸如吉非替尼)治疗的抗性的突变的瘤和/或瘤细胞系中的egfr信号传导,并且抑制c-met信号传导途径,即在癌诸如nsclc中被鉴定为上调并且用egfr酪氨酸激酶抑制剂治疗时提供补偿信号传导的途径。本发明的双特异性egfr/c-met抗体除了直接抑制egfr和c-met信号传导外,还通过增强的抗体依赖性细胞毒性(adcc)以及egfr和c-met受体的降解而展示出抗瘤活性。与当前的egfr疗法(西妥昔单抗和帕尼单抗)相反,本发明的双特异性egfr/c-met抗体经由增强的adcc来诱导具有kras突变的瘤细胞的杀伤。国际专利公布wo2010/115551描述了使用结合egfr的西妥昔单抗vh/vl对及结合c-met的抗体5d5(metmab,奥纳珠单抗)vh/vl对以igg-scfv形式工程改造的、当前处于iii期试验的双特异性egfr/c-met抗体(bsab01)。bsab01展现出与亲本抗体相比时a431细胞增殖的大约两倍(加性)增强的抑制(wo2010/115551中的实例7、图8b),以及与两种亲本抗体的组合相比时ovarc-8细胞增殖的适度加性抑制(15%相对于10%抑制)(wo2010/115551中的图10a、实例16)。因此,令人惊讶且出人意料的是,本发明提供双特异性egfr/c-met抗体,其展现出egfr和c-met信号传导、癌细胞存活和瘤生长的抑制的显著协同效应。不希望受任何理论的约束,据信本发明的双特异性抗体的显著协同效应至少部分地由egfr和c-met结合臂两者的表位特异性所引起,从而可能不仅通过egfr和c-met同源二聚体而且通过egfr/herx异源二聚体来抑制信号传导。本发明的一个实施例是分离的双特异性表皮生长因子受体(egfr)/肝细胞生长因子受体(c-met)抗体,其包含:a)第一重链(hc1),该第一重链包含hc1恒定域3(hc1ch3)和hc1可变区1(vh1);b)第二重链(hc2),该第二重链包含hc2恒定域3(hc2ch3)和hc2可变区2(vh2);c)第一轻链(lc1),该第一轻链包含轻链可变区1(vl1);和第二轻链(lc2),该第二轻链包含轻链可变区2(vl2),其中vh1和vl1配对以形成特异性结合egfr的第一抗原结合位点,并且vh2和vl2配对以形成特异性结合c-met的第二抗原结合位点,其中hc1在hc1ch3中包含至少一个替换,并且hc2在hc2ch3中包含至少一个替换,其中当根据eu索引对残基进行编号时,hc1ch3中的替换和hc2ch3中的替换在不同氨基酸残基位置处发生。在本文所述的一些实施例中,双特异性egfr/c-met抗体抑制nci-h292、nci-h1975或skmes-1细胞系中的胞外信号相关激酶1和2(erk1/2)的磷酸化,并且与用包含重链3(hc3)和轻链3(lc3)的对照单价egfr抗体以及包含重链4(hc4)和轻链4(lc4)的对照单价c-met抗体的混合物抑制nci-h292、nci-h1975或skmes-1细胞系中erk1/2的磷酸化的ic50值相比,其ic50值小至少约10倍、小至少约20倍、小至少约30倍、小至少约40倍、小至少约50倍或小至少约60倍,其中hc3与hc1、lc3与lc1、hc4与hc2以及lc4与lc2分别具有相同的氨基酸序列,并且使用夹心免疫测定法,将抗磷酸化erk1/2抗体用作捕获抗体并将与电化学发光化合物缀合的结合于非磷酸化和磷酸化erk1/2的抗体用作检测抗体,在全细胞裂解物中测量erk1/2的磷酸化。当通过erk1/2磷酸化的抑制来评估抑制时,与单特异性egfr抗体和单特异性c-met抗体的组合相比,本发明的双特异性egfr/c-met抗体提供egfr和c-met信号传导的协同性更明显的抑制。此类示例性的双特异性egfr/c-met抗体是本发明的抗体em1-mab。如本文所用,“对照单特异性egfr抗体”是指这样的抗体,其具有结合egfr的第一fab臂,该第一fab臂在氨基酸序列方面与待测试的双特异性egfr/c-met抗体的结合egfr的fab臂相同,并且具有第二fab臂,该第二fab臂是“惰性的”并且结合无关/不相关抗原人免疫缺陷病毒(hiv)gp120。在待测试的双特异性egfr/c-met抗体中的结合egfr的fab臂包含f405l替换的情形中,第二fab臂包含具有seqidno:209序列的轻链和具有seqidno:198序列的重链。在待测试的双特异性egfr/c-met抗体中的结合egfr的fab臂包含k409r替换的情形中,第二fab臂包含具有seqidno:209序列的轻链和具有seqidno:197序列的重链。如本文所用,“对照单特异性c-met抗体”是指这样的抗体,其具有结合c-met的第一fab臂,该第一fab臂在氨基酸序列方面与待测试的双特异性egfr/c-met抗体的结合c-met的fab臂相同,并且具有第二fab臂,该第二fab臂是“惰性的”并且结合无关/不相关抗原hivgp120。在待测试的双特异性egfr/c-met抗体中的结合c-met的fab臂包含f405l替换的情形中,第二fab臂包含具有seqidno:209序列的轻链和具有seqidno:198序列的重链。在待测试的双特异性egfr/c-met抗体中的结合c-met的fab臂包含k409r替换的情形中,第二惰性fab臂包含具有seqidno:209序列的轻链和具有seqidno:197序列的重链。在本文所述的一些实施例中,双特异性egfr/c-met抗体以约2×10-9m或更小、约1×10-9m或更小,或约1×10-10m或更小的ic50值抑制erk1/2的磷酸化。在本文所述的一些实施例中,erk1在残基thr202和tyr204处被磷酸化,并且erk2在残基thr185和tyr197处被磷酸化。在本文所述的一些实施例中,双特异性egfr/c-met抗体抑制nci-h1975细胞系中蛋白激酶b(akt)在ser473处的磷酸化,并且与用包含hc3和lc3的对照单价egfr抗体以及包含hc4和lc4的对照单价c-met抗体的混合物抑制nci-h1975细胞系中akt在ser473处的磷酸化的ic50值相比,其ic50值小至少约70倍,其中hc3与hc1、lc3与lc1、hc4与hc2以及lc4与lc2分别具有相同的氨基酸序列,其中使用夹心免疫测定法,将结合于非磷酸化和磷酸化akt的抗体用作捕获抗体并将与电化学发光化合物缀合的抗磷酸化aktser473抗体用作检测抗体,在全细胞裂解物中测量akt在ser473处的磷酸化。在本文所述的一些实施例中,双特异性egfr/c-met抗体抑制nci-h1975细胞系中蛋白激酶b(akt)在thr308处的磷酸化,并且与用包含hc3和lc3的对照单价egfr抗体以及包含hc4和lc4的对照单价c-met抗体的混合物抑制nci-h1975细胞系中akt在thr308处的磷酸化的ic50值相比,其ic50值小至少约100倍,其中hc3与hc1、lc3与lc1、hc4与hc2以及lc4与lc2分别具有相同的氨基酸序列,其中使用夹心免疫测定法,将结合于非磷酸化和磷酸化akt的抗体用作捕获抗体并将与电化学发光化合物缀合的抗磷酸化aktthr308抗体用作检测抗体,在全细胞裂解物中测量akt在thr308处的磷酸化。当通过akt磷酸化的抑制来评估抑制时,与单特异性egfr抗体和单特异性c-met抗体的组合相比,本发明的双特异性egfr/c-met抗体提供egfr和c-met信号传导的协同性更明显的抑制。此类示例性的双特异性egfr/c-met抗体是本发明的抗体em1-mab。在本文所述的一些实施例中,双特异性egfr/c-met抗体以约1×10-9m或更小的ic50值抑制akt在ser473处或在thr308处的磷酸化。在本文所述的一些实施例中,双特异性egfr/c-met抗体在egfr残基k489、i491、k467和s492处结合seqidno:73的egfr,并且在残基pefrdsypikyvhaf(seqidno:238)和faqskpdsaepmdrsa(seqidno:239)处结合c-met。此类示例性的双特异性抗体是em1-mab。当与如上文和国际专利公布wo2010/115551中所述的抗体bsab01相比时,双特异性em-1抗体在不同表位处结合egfr和c-met。bsab01的亲本egfr结合臂(西妥昔单抗)结合成熟egfr中的egfr氨基酸残基r353、q384、q408、h409、f412、s418、s440、k443、k465、i467、s468和n473,对应于seqidno:73的全长egfr的残基r367、q408、q432、h433、f436、s442、s464、k467、k489、i491、s492和n497(li等人,cancercell7:301-311,2005)。bsab01(mab5d5)的亲本c-met结合臂结合c-met残基325-340pgaqlarqigaslndd(seqidno:240)。em1-mab的结合egfr的亲本抗体(2f8)的表位作图描述于美国专利公布us2011/0256142a1。西妥昔单抗和亲本2f8抗体结合部分重叠但不同的表位。可使用标准方法进行表位作图。例如,当两种各个组分的结构均已知时,可进行计算机芯片上的(insilico)蛋白质-蛋白质对接,以鉴定相互作用的相容位点。可使用抗原和抗体复合物进行氢-氘(h/d)交换以标测抗原上可能受到该抗体结合的区域。可使用抗原的片段和点诱变来定位对于抗体结合而言重要的氨基酸。在本文所述的一些实施例中,双特异性egfr/c-met抗体中和egfr和c-met信号传导。与在不存在本发明的双特异性egfr/c-met分子的情况下使用相同测定法条件时信号传导的水平相比,本发明的双特异性egfr/c-met抗体可将egfr和c-met信号传导中和至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。配体诸如egf与egfr的结合刺激受体二聚化、自磷酸化、受体的内部细胞质酪氨酸激酶域的激活,以及多个参与dna合成(基因激活)和细胞周期进程或分裂的调控的信号转导和反式激活途径的引发。egfr信号传导的中和可导致一个或多个egfr下游信号传导途径受抑制,因此中和egfr可具有各种效应,包括细胞增殖和分化、血管生成、细胞运动和转移的抑制,以及下游信号传导途径的抑制。egfr信号传导及egfr信号传导的中和可使用各种熟知的方法测量,例如测量受体在酪氨酸y1068、y1148和y1173任一者处的自磷酸化(downward等人,nature311:483-5,1984)和/或天然或合成底物的磷酸化,以及本发明的双特异性抗体对天然或合成底物的自磷酸化和/或磷酸化的抑制。磷酸化可使用熟知的方法检测,诸如采用磷酸化酪氨酸特异性抗体的elisa测定法或蛋白质印迹。示例性的测定法可见于panek等人,jpharmacolexpthera283:1433-44,1997和batley等人,lifesci62:143-50,1998并且如本文所述。hgf与c-met的结合刺激受体二聚化、自磷酸化、受体的细胞质酪氨酸激酶域的激活,以及多个参与dna合成(基因激活)和细胞周期进程或分裂的调控的信号转导和反式激活途径的引发。c-met信号传导的抑制可导致一个或多个c-met下游信号传导途径受抑制,因此中和c-met可具有各种效应,包括细胞增殖和分化、血管生成、细胞运动和转移的抑制。c-met信号传导及c-met信号传导的中和可使用各种熟知的方法测量,例如测量受体在至少一个酪氨酸残基y1230、y1234、y1235或y1349上的自磷酸化和/或天然或合成底物的磷酸化。可例如在elisa测定法中或在蛋白质印迹上使用对磷酸化酪氨酸具有特异性的抗体来检测磷酸化。示例性的测定法可见于panek等人,jpharmacolexpthera283:1433-44,1997和batley等人,lifesci62:143-50,1998并且如本文所述。egfr和c-met信号传导可使用如本文所述的各种熟知方法测量,诸如测量erk1/2和akt磷酸化的抑制。在thr202和tyr204处的erk1磷酸化及在thr185和tyr187处的erk2磷酸化的抑制以及akt在ser473或thr308处的抑制可例如在nci-h1975细胞裂解物中利用采用包被在固相载体上的捕获抗体和与电化学发光化合物诸如mesoscalediscover(msd)sulfo-tag标签缀合的检测抗体的夹心测定法测量,然后用读板机检测信号。在本文所述的一些实施例中,双特异性egfr/c-met抗体抑制nci-h292或nci-h1975细胞的生长,并且当nci-h292或nci-h1975细胞在低附着条件下生长时,与用西妥昔单抗抑制nci-h292或nci-h1975细胞的生长的ic50值相比,其ic50值小至少约300倍、小至少约400倍、小至少约500倍、小至少约600倍、小至少约700倍或小至少约800倍。细胞生长的抑制可通过已知的方法评估。例如,可将细胞接种在涂布有水凝胶或仿生聚合物的平板(corning制造的超低附着性平板(ultralowattachmentplate))中以防止或减少细胞附着,并且可在温育72小时后通过使用标准方法测量细胞活力百分比来评估抗体对7.5ng/mlhgf诱导的细胞生长的效应。与单特异性egfr抗体和单特异性c-met抗体的组合相比并与标准治疗西妥昔单抗相比,本发明的双特异性egfr/c-met抗体提供表达egfr和/或c-met的癌细胞的协同性更明显的抑制。此类示例性的双特异性egfr/c-met抗体是本发明的抗体em1-mab。本发明的双特异性egfr/c-met抗体抑制表达野生型egfr和野生型c-met的癌细胞,并且还抑制表达egfrl858r/t790m突变体的癌细胞,该突变被鉴定为起到抵抗小分子酪氨酸激酶抑制剂(tki)诸如吉非替尼治疗的作用。因此,与西妥昔单抗和tki相比,本发明的双特异性egfr/c-met抗体可在更广泛患者群体中提供有益效果。在本文所述的一些实施例中,双特异性egfr/c-met抗体抑制scidbeige小鼠中表达hgf的skmes-1细胞瘤的生长,并且当双特异性抗体和西妥昔单抗以20mg/kg剂量施用时,与西妥昔单抗相比,在第36天,其t/c百分比(%)值小至少500倍。使用scidbeige小鼠的移植瘤模型是熟知的。可使用标准方法来工程改造skmes-1细胞以表达人hgf。通常,scidbeige小鼠可皮下接种表达人hgf的skmes-1细胞,该细胞包埋于每只动物背侧的胞外基质诸如culturex中。植入后一周,小鼠可被分为具有相等瘤体积的组,之后以例如每周三次给予本发明的双特异性egfr/c-met抗体、对照或基准抗体或小分子。可每周记录两次瘤体积,并且可通过计算t/c百分比(%)值来观察瘤生长抑制(tgi)。t/c%值指示抗瘤疗效。t和c分别是在给定天的处理组和对照组的平均体积。与标准治疗西妥昔单抗相比,本发明的双特异性egfr/c-met抗体提供在体内瘤杀伤方面显著改善的疗效,因此与西妥昔单抗相比,可在患者群体中提供有益效果。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含igg1、igg2、igg3或igg4同种型的hc1和hc2。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含igg1同种型的hc1和hc2。在本文所述的一些实施例中,当根据eu索引对残基进行编号时,在残基位置350、366、368、370、399、405、407或409处,双特异性egfr/c-met抗体hc1ch3包含至少一、二、三、四、五、六、七或八个替换,并且hc2ch3包含至少一、二、三、四、五、六、七或八个替换。在本文所述的一些实施例中,当根据eu索引对残基进行编号时,在残基位置350、370、405或409处,双特异性egfr/c-met抗体hc1ch3包含至少一、二、三或四个替换,并且hc2ch3包含至少一、二、三或四个替换。抗体域和编号是熟知的。当两个ch3域(或ch3区)彼此相差至少一个氨基酸替换时,它们不相同。igg1ch3区通常由igg1上的第341-446位残基组成(根据eu索引对残基进行编号)。示例性的igg1恒定区以seqidno:203示出。ch3域跨越seqidno:203的第224-329位残基,并且对应于根据eu索引的第341-446位残基。在本文所述的一些实施例中,当根据eu索引对残基进行编号时,在残基位置405或409处,双特异性egfr/c-met抗体hc1ch3包含至少一个替换,并且hc2ch3包含至少一个替换。在本文所述的一些实施例中,双特异性egfr/c-met抗体hc1ch3包含k409r或f405l替换,并且hc2ch3包含k409r或f405l替换,其中根据eu索引对残基进行编号。在本文所述的一些实施例中,双特异性egfr/c-met抗体hc1ch3包含f405l替换,并且hc2ch3包含k409r替换。在本文所述的一些实施例中,hc1ch3和hc2ch3替换是在位置366、368、370、399、405、407或409(根据eu索引进行编号)处的替换。这些位置分别对应于seqidno:203和204的重链恒定区中的线性残基位置248、250、252、281、287、289和291。在本文所述的一些实施例中,hc1ch3位置409具有除lys、leu或met之外的氨基酸替换,并且hc2ch3位置405具有除phe之外的氨基酸替换。在本文所述的一些实施例中,hc1ch3位置405具有除phe之外的氨基酸替换,并且hc2ch3位置409具有除lys、leu或met之外的氨基酸替换。在本文所述的一些实施例中,hc1ch3位置409具有除lys、leu或met之外的氨基酸替换,并且hc2ch3位置405具有除phe、arg或gly之外的氨基酸替换。在本文所述的一些实施例中,hc1ch3位置405具有除phe、arg或gly之外的氨基酸替换,并且hc2ch3位置409具有除lys、leu或met之外的氨基酸替换。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置405处的除phe之外的氨基酸以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置405处的除phe之外的氨基酸以及在位置409处的lys,并且hc2ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置405处的除phe、arg或gly之外的替换以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置405处的除phe、arg或gly之外的替换以及在位置409处的lys,并且hc2ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置405处的leu以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置405处的leu以及在位置409处的lys,并且hc2ch3具有在位置405处的phe以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的arg,并且hc2ch3具有在位置405处的除phe、arg或gly之外的氨基酸以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置405处的除phe、arg或gly之外的氨基酸以及在位置409处的lys,并且hc2ch3具有在位置405处的phe以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的arg,并且hc2ch3具有在位置405处的leu以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置405处的leu以及在位置409处的lys,并且hc2ch3具有在位置405处的phe以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置405处的phe以及在位置409处的lys,并且hc2ch3具有在位置405处的leu以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置405处的leu以及在位置409处的arg,并且hc2ch3具有在位置405处的phe以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu。在本文所述的一些实施例中,hc1ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu,并且hc2ch3具有在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置409处的arg,并且hc2ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu。在本文所述的一些实施例中,hc1ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu,并且hc2ch3具有在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置370处的lys、在位置405处的phe以及在位置409处的arg,并且hc2ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu。在本文所述的一些实施例中,hc1ch3具有在位置409处的lys、在位置370处的thr以及在位置405处的leu,并且hc2ch3具有在位置370处的lys、在位置405处的phe以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸,并且hc2ch3具有在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp。在本文所述的一些实施例中,hc1ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp,并且hc2ch3具有在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的gly、leu、met、asn或trp。在本文所述的一些实施例中,hc1ch3具有在位置407处的gly、leu、met、asn或trp,并且hc2ch3具有在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有在位置407处的gly、leu、met、asn或trp以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的gly、leu、met、asn或trp以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的arg,并且hc2ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的除tyr、asp、glu、phe、lys、gln、arg、ser或thr之外的氨基酸以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的arg,并且hc2ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的ala、gly、his、ile、leu、met、asn、val或trp以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置407处的tyr以及在位置409处的arg,并且hc2ch3具有在位置407处的gly、leu、met、asn或trp以及在位置409处的lys。在本文所述的一些实施例中,hc1ch3具有在位置407处的gly、leu、met、asn或trp以及在位置409处的lys,并且hc2ch3具有在位置407处的tyr以及在位置409处的arg。在本文所述的一些实施例中,hc1ch3具有在位置409处的除lys、leu或met之外的氨基酸,并且hc2ch3具有(i)在位置368处的除phe、leu和met之外的氨基酸,或(ii)在位置370处的trp,或(iii)在位置399处的除asp、cys、pro、glu或gln之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有(i)在位置368处的除phe、leu和met之外的氨基酸,或(ii)在位置370处的trp,或(iii)在位置399处的除asp、cys、pro、glu或gln之外的氨基酸,并且hc2ch3具有在位置409处的除lys、leu或met之外的氨基酸。在本文所述的一些实施例中,hc1ch3具有在位置409处的arg、ala、his或gly,并且hc2ch3具有(i)在位置368处的lys、gln、ala、asp、glu、gly、his、ile、asn、arg、ser、thr、val或trp,或(ii)在位置370处的trp,或(iii)在位置399处的ala、gly、ile、leu、met、asn、ser、thr、trp、phe、his、lys、arg或tyr。在本文所述的一些实施例中,hc1ch3具有(i)在位置368处的lys、gln、ala、asp、glu、gly、his、ile、asn、arg、ser、thr、val或trp,或(ii)在位置370处的trp,或(iii)在位置399处的ala、gly、ile、leu、met、asn、ser、thr、trp、phe、his、lys、arg或tyr,并且hc2ch3具有在位置409处的arg、ala、his或gly。在本文所述的一些实施例中,hc1ch3具有在位置409处的arg,并且hc2ch3具有(i)在位置368处的asp、glu、gly、asn、arg、ser、thr、val或trp,或(ii)在位置370处的trp,或(iii)在位置399处的phe、his、lys、arg或tyr。在本文所述的一些实施例中,hc1ch3具有(i)在位置368处的asp、glu、gly、asn、arg、ser、thr、val或trp,或(ii)在位置370处的trp,或(iii)在位置399处的phe、his、lys、arg或tyr,并且hc2ch3具有在位置409处的arg。在本文所述的一些实施例中,hc1ch3包含k409r替换或f405l替换,并且hc2ch3包含k409r替换或f405l替换,其中根据eu索引对残基进行编号。在本文所述的一些实施例中,hc1ch3包含f405l替换,并且hc2ch3包含k409r替换。通常使用标准方法在dna水平对分子诸如抗体的恒定域作出替换。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含vh1和vl1,其中vh1包含分别为seqidno:210、211和212的重链互补决定区(hcdr)1(hcdr1)、hcdr2和hcdr3氨基酸序列;并且vl1包含分别为seqidno:213、214和215的轻链互补决定区(lcdr)1(lcdr1)、lcdr2和lcdr3氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含vh2和vl2,其中vh2包含分别为seqidno:216、217和218的hcdr1、hcdr2和hcdr3氨基酸序列;并且vl2包含分别为seqidno:219、220和221的lcdr1、lcdr2和lcdr3氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含分别为seqidno:189、190、193和194的vh1、vl1、vh2和vl2氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含分别为seqidno:199、200、201和202的hc1、lc1、hc2和lc2氨基酸序列,任选地从hc1、hc2、或hc1和hc2两者移除c-末端赖氨酸。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含vh1和vl1,其中vh1包含分别为seqidno:222、223和224的hcdr1、hcdr2和hcdr3氨基酸序列;并且vl1包含分别为seqidno:225、226和227的lcdr1、lcdr2和lcdr3氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含vh2和vl2,其中vh2包含分别为seqidno:228、229和230的hcdr1、hcdr2和hcdr3氨基酸序列;并且vl2包含分别为seqidno:231、232和233的lcdr1、lcdr2和lcdr3氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含分别为seqidno:191、192、195和196的vh1、vl1、vh2和vl2氨基酸序列。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含分别为seqidno:234、235、236和237的hc1、lc1、hc2和lc2氨基酸序列,任选地从hc1、hc2、或hc1和hc2两者移除c-末端赖氨酸。在本文所述的一些实施例中,在采用包被于平板上并且与或不与本发明的双特异性egfr/c-met抗体一起温育的重组人egfr或重组人c-met胞外域的竞争测定法中,双特异性egfr/c-met抗体可以小于约1×10-8m、小于约1×10-9m、小于约1×10-10m、小于约1×10-11m或小于约1×10-12m的ic50值阻碍egf结合于egfr以及hgf结合于c-met。与在不存在本文所述本发明的双特异性egfr/c-met抗体的情况下使用相同测定法条件时egf对egfr的结合以及hgf对c-met的结合相比,本文所述双特异性egfr/c-met抗体可将egf结合于egfr以及hgf结合于c-met阻碍至少30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。在本文所述的一些实施例中,双特异性egfr/c-met抗体包含hc1、lc1、hc2和lc2,其中hc1、lc1、hc2和lc2由分别包含seqidno:205、206、207和208的序列的合成多核苷酸编码。本发明的双特异性egfr/c-met抗体可使用本文所述的技术生成,诸如利用ch3工程改造并使用体外fab臂交换来生成抗体。示例性的双特异性抗体可通过如下方式由两种单特异性抗体生成:将约1-20mg/ml的每种抗体以1:1摩尔比组合在具有75mm的终浓度的2-巯基乙醇胺(2-mea)的缓冲液中的ph7.0-7.4的pbs中,在25-37℃下温育2-6小时,然后经由透析、透滤、切向流过滤和旋转细胞过滤来移除2-mea。双特异性抗体的收率可大于约80%、大于约90%、大于约91%、92%、93%、94%、95%、96%、97%、98%或99%。本文所述的一些实施例提供制备分离的双特异性egfr/c-met抗体的方法,所述方法包括:将包含seqidno:199的两条重链和seqidno:200的两条轻链的分离的单特异性二价抗egfr抗体与包含seqidno:201的两条重链和seqidno:202的两条轻链的分离的单特异性二价抗c-met抗体以约1:1摩尔比组合在混合物中;将还原剂引入混合物中;将混合物温育约九十分钟至约六小时;移除还原剂;以及纯化双特异性egfr/c-met抗体,该双特异性egfr/c-met抗体包含seqidno:199的第一重链和seqidno:201的第二重链、seqidno:200的第一轻链和seqidno:202的第二轻链,其中seqidno:199的第一重链与seqidno:200的第一轻链配对以形成特异性结合egfr的第一结合域,并且seqidno:201的第二重链与seqidno:202的第二轻链配对以形成特异性结合c-met的第二结合域。在本文所述的一些实施例中,还原剂是2-巯基乙醇胺(2-mea)。在本文所述的一些实施例中,2-mea以约25mm至约75mm的浓度存在。在本文所述的一些实施例中,温育步骤在约25℃至约37℃的温度下进行。本文所述的一些实施例提供分离的双特异性egfr/-c-met抗体,其包含hc1、lc1、hc2和lc2,其中hc1包含seqidno:199的序列,lc1包含seqidno:200的序列,hc2包含seqidno:201的序列,并且lc2包含seqidno:202的序列,其中hc1、lc1、hc2和/或lc2还包含1、2、3、4、5、6、7、8、9、10、11、12、13、14或15个保守氨基酸替换。本文所述的一些实施例提供分离的双特异性egfr/-c-met抗体,其包含hc1、lc1、hc2和lc2,其中hc1包含seqidno:234的序列,lc1包含seqidno:235的序列,hc2包含seqidno:236的序列,并且lc2包含seqidno:237的序列,其中hc1、lc1、hc2和/或lc2还包含1、2、3、4、5、6、7、8、9、10、11、12、13、14或15个保守氨基酸替换。其hc1、lc1、hc2和lc2氨基酸序列与本文所公开的那些抗体非实质性差异的双特异性egfr/c-met抗体涵盖在本发明的范围内。通常,这涉及采用在抗原结合位点或框架中具有类似电荷、疏水或立体化学特性的氨基酸的一个或多个保守氨基酸替换,而不会不利地改变抗体的性质。也可作出保守替换以改善抗体性质,例如稳定性或亲和力。可例如对vh1、vl1、vh2和/或vl2作出1、2、3、4、5、6、7、8、9、10、11、12、13、14或15个氨基酸替换。例如,“保守氨基酸替换”可涉及天然氨基酸残基受到非天然残基的替换,从而使该位置的氨基酸残基的极性或电荷几乎无影响或无影响。此外,多肽中的任何天然残基还可以用丙氨酸来替换,如此前对于丙氨酸扫描诱变所述(maclennan等人,actaphysiolscandsuppl643:55-67,1998;sasaki等人,advbiophys35:1-24,1998)。所期望的氨基酸替换可由本领域的技术人员在需要此类替换时来决定。例如,可使用氨基酸替换来鉴定分子序列的重要残基,或用于提高或降低本文所述分子的亲和性。示例性的保守氨基酸替换在上文描述。氨基酸替换可例如通过pcr诱变来进行(美国专利4,683,195)。变体的文库可使用熟知的方法生成,例如使用随机(nnk)或非随机密码子,例如dvk密码子,其编码11个氨基酸(ala、cys、asp、glu、gly、lys、asn、arg、ser、tyr、trp),并且筛选文库中具有所需性质的变体。在本文所述的一些实施例中,可对抗体的恒定区作出氨基酸替换。例如,可在本发明的双特异性egfr/c-met抗体中使用不同的igg1同种异型,诸如熟知的g1m17同种异型、g1m3同种异型或g1m1同种异型,或它们的组合。在本文所述的一些实施例中,双特异性egfr/c-met抗体的药代动力学性质可通过fc域中调节抗体半衰期的替换来增强。在本文所述的一些实施例中,双特异性egfr/c-met抗体在hc1和/或hc2中包含替换m252y/s254t/t256e,其中根据eu索引对残基进行编号。m252y/s254t/t256e替换已示出延长抗体半衰期(dall’acqua等人,jbiolchem281:23514-24,2006)。使用本文所述的方法,测试在其fc区中具有保守替换和/或另外的替换的双特异性egfr/c-met抗体的特性。在本文所述的某个实施例中,可通过本领域技术人员已知的技术进行的fc修饰来增强或沉默双特异性egfr/c-met抗体的免疫效应子性质。例如,fc效应子功能诸如c1q结合、补体依赖性细胞毒性(cdc)、抗体依赖性细胞介导的细胞毒性(adcc)、抗体依赖性细胞介导的吞噬作用(adcp)、细胞表面受体(例如b细胞受体;bcr)的下调等,可通过修饰促成这些活性的fc中的残基来提供和/或控制。“抗体依赖性细胞介导的细胞毒性”或“adcc”是指细胞介导的反应,其中表达fc受体(fcr)的非特异性细胞毒性细胞(例如,自然杀伤(nk)细胞、中性粒细胞和巨噬细胞)识别靶细胞上的结合抗体并且随后引起靶细胞的裂解。单克隆抗体诱导adcc的能力可通过工程改造其寡糖组分来增强。用熟知的二天线g0、g0f、g1、g1f、g2或g2f形式的大多数聚糖在asn297处对人igg1或igg3进行n-糖基化。由非工程改造的cho细胞所产生的抗体通常具有约至少85%的聚糖岩藻糖含量。核心岩藻糖从连接到fc区的二天线复合物型寡糖的移除经由改善的fcγriiia结合来增强抗体的adcc,而不改变抗原结合或cdc活性。此类mab可使用据报道引起具有二天线复合物型fc寡糖的相对较高去岩藻糖基化抗体的成功表达的不同方法实现,诸如培养渗透压度的控制(konno等人,cytotechnology64(:249-65,2012)、变体cho系lec13作为宿主细胞系的应用(shields等人,jbiolchem277:26733-26740,2002)、变体cho系eb66作为宿主细胞系的应用(olivier等人,mabs;2(4),2010;刊载之前的电子版;pmid:20562582)、大鼠杂交瘤细胞系yb2/0作为宿主细胞系的应用(shinkawa等人,jbiolchem278:3466-3473,2003)、特异性针对α1,6-岩藻糖基转移酶(fut8)基因的小干扰rna的引入(mori等人,biotechnolbioeng88:901-908,2004)或β-1,4-n-乙酰葡糖胺基转移酶iii和高尔基体α-甘露糖苷酶ii或强效α-甘露糖苷酶i抑制剂几夫碱的共表达(ferrara等人,jbiolchem281:5032-5036,2006,ferrara等人,biotechnolbioeng93:851-861,2006;xhou等人,biotechnolbioeng99:652-65,2008)。在本文所述的一些实施例中,由双特异性egfr/c-met抗体引起的adcc也可通过抗体fc中的某些替换来增强。示例性的替换是例如在氨基酸位置256、290、298、312、356、330、333、334、360、378或430(根据eu索引对残基进行编号)处的替换,如美国专利us6737056中所述。在本文所述的一些实施例中,本发明的双特异性egfr/c-met抗体具有岩藻糖含量约在1%至约15%之间,例如15%、14%、13%、12%、11%10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的二天线聚糖结构。在一些实施例中,双特异性egfr/c-met抗体具有岩藻糖含量为约50%、40%、45%、40%、35%、30%、25%或20%的聚糖结构。“岩藻糖含量”意指在asn297处的糖链内的岩藻糖单糖的量。岩藻糖的相对量是与所有糖基化结构相关的含岩藻糖的结构的百分比。这些可通过多种方法来表征和定量,例如:1)使用n-糖苷酶f处理的样品(例如,复合甘露糖、杂合甘露糖和寡聚甘露糖及高甘露糖结构)的maldi-tof,如国际专利公布wo2008/0775462所述;2)asn297聚糖的酶促释放,随后通过带荧光检测的hplc(uplc)和/或hplc-ms(uplc-ms)衍生化和检测/定量;3)在用endos或其他酶处理或不处理asn297聚糖的情况下天然或还原mab的完整蛋白分析,该endos或其他酶在第一与第二glcnac单糖之间切割,使岩藻糖连接到第一glcnac;4)通过酶消化(胰蛋白酶或肽链内切酶lys-c)将mab消化为成分肽,随后通过hplc-ms(uplc-ms)分离、检测和定量;5)使用png酶f通过在asn297处的特异性酶促去糖基化将mab寡糖从mab蛋白质分离。由此释放的寡糖可标记有荧光团,并通过各种配套技术分离和鉴定,这些技术允许:采用基质辅助激光解吸电离(maldi)质谱通过实验质量与理论质量的比较而对聚糖结构进行精细表征,通过离子交换hplc(glycosepc)来测定唾液酸化的程度,通过正相hplc(glycosepn)根据亲水性标准来分离和定量寡糖形式,以及通过高效毛细管电泳-激光诱导荧光(hpce-lif)来分离和定量寡糖。如本申请中所用,“低岩藻糖”或“低岩藻糖含量”是指岩藻糖含量为约1%-15%之间的抗体。如本文所用,“正常岩藻糖”或“正常岩藻糖含量”是指岩藻糖含量为约超过50%、通常约超过80%或超过85%的抗体。本发明的一些实施例提供编码如本文所述的本发明双特异性egfr/c-met结合抗体的重链和轻链的合成核酸,作为分离的多核苷酸或作为表达载体的部分或作为线性dna序列的部分,包括用于体外转录/翻译的线性dna序列、与其组合物或定向诱变剂的原核、真核或丝状噬菌体表达、分泌和/或展示相容的载体。本发明的一些实施例提供分离的多核苷酸,其包含seqidno:205、206、207或208的多核苷酸序列。本发明的多核苷酸可以通过化学合成(诸如在自动化多核苷酸合成仪上的固相多核苷酸合成)来制备,并装配成完整的单链或双链分子。另选地,本发明的多核苷酸可以通过其他技术来制备,诸如pcr之后进行常规克隆。制备或获得具有给定的已知序列的多核苷酸的技术是本领域熟知的。本发明的多核苷酸可包含至少一个非编码序列,诸如启动子或增强子序列、内含子、聚腺苷酸化信号、促进repa结合的顺式序列等。多核苷酸序列还可包含编码另外的氨基酸的另外序列,其编码例如标记或标签序列,诸如组氨酸标签或ha标签以促进蛋白质的纯化或检测;信号序列;融合蛋白伴侣诸如repa、fc;或噬菌体外壳蛋白诸如pix或piii。本文所述的一些实施例提供包含本发明的多核苷酸的载体。此类载体可以是质粒载体、病毒载体、杆状病毒表达载体、基于转座子的载体或任何其他适用于通过任何手段将本发明的多核苷酸引入给定生物体或遗传背景的载体。例如,可将编码本发明双特异性抗体的重链和轻链的多核苷酸插入表达载体中。轻链和重链可克隆在相同或不同的表达载体中。可将编码免疫球蛋白链的dna片段操作地连接到确保免疫球蛋白多肽表达的一个或多个表达载体中的控制序列。此类控制序列包括信号序列、启动子(例如,天然相关或异源启动子)、增强子元件和转录终止序列,并且可被选择为与选择用于表达抗体的宿主细胞相容。一旦载体已掺入到适当宿主中,则宿主可在适于由所掺入的合成多核苷酸编码的蛋白质的高水平表达的条件下维持。合适的表达载体通常可在宿主生物体中作为附加体或作为宿主染色体dna的组成部分进行复制。一般来讲,表达载体包含选择标记,诸如氨比西林抗性、潮霉素抗性、四环素抗性、卡那霉素抗性或新霉素抗性,以允许检测被所需dna序列转化的那些细胞。本文所述的一些实施例提供包含本发明的载体的宿主细胞。术语“宿主细胞”是指其中引入了载体的细胞。应当理解,术语宿主细胞旨在不仅指特定对象细胞,而且指此种细胞的后代。由于后续世代中可能因突变或环境影响而出现某些修饰,此类后代可能不同于亲本细胞,但仍然包括在如本文所用的术语“宿主细胞”的范围内。此类宿主细胞可以是真核细胞、原核细胞、植物细胞或古菌细胞。示例性的真核细胞可以是哺乳动物、昆虫、禽类或其他动物来源。哺乳动物真核细胞包括永生化细胞系,例如杂交瘤或骨髓瘤细胞系,例如sp2/0(americantypeculturecollection(atcc),manassas,va,crl-1581)、ns0(europeancollectionofcellcultures(ecacc),salisbury,wiltshire,uk,ecacc编号85110503)、fo(atcccrl-1646)和ag653(atcccrl-1580)鼠细胞系。示例性人骨髓瘤细胞系是u266(attccrl-tib-196)。其他可用的细胞系包括来源于中国仓鼠卵巢(cho)细胞的那些细胞系,诸如cho-k1sv(lonzabiologics,walkersville,md)、cho-k1(atcccrl-61)或dg44。本发明的包含双特异性egfr/c-metfn3域的分子、双特异性egfr/-c-met抗体以及结合egfr或结合c-met的fn3域的用途本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr-c-met抗体可用于诊断、监测、调节、治疗、缓解、帮助预防发生细胞、组织、器官、流体或通常宿主中的人疾病或特定病变或者减轻其症状。本发明的方法可用于治疗属于任何分类的动物患者。此类动物的例子包括哺乳动物,诸如人类、啮齿类、犬类、猫类和农畜/家畜。本发明的一个方面是用于抑制表达egfr和/或c-met的细胞的生长或增殖的方法,所述方法包括使细胞与本发明的分离的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体接触。本发明的另一个方面是用于抑制受检者中表达egfr和/或c-met的瘤或癌细胞的生长或转移的方法,所述方法包括向受检者施用有效量的本发明的分离的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体,使得表达egfr和/或c-met的瘤或癌细胞的生长或转移被抑制。本发明的另一个方面是治疗患有癌的受检者的方法,所述方法包括向对其有需要的患者施用治疗有效量的分离的本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体足以治疗癌的时间。本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体,可用于治疗特征在于egfr、c-met、egf、可溶性egfr、可溶性c-met或其他egfr配体或hgf的异常激活或产生的任何疾病或失调,或与egfr或c-met表达相关的失调,其可能涉及或可能不涉及恶性化或癌,其中egfr、c-met、egf或其他egfr配体或hgf的异常激活和/或产生会在患有或易患疾病或失调的受检者的细胞或组织中发生。本发明的特异性结合c-met并阻碍hgf结合于c-met的fn3域可用于治疗瘤,包括癌和良性瘤。适于通过本发明的结合c-met的fn3域来治疗的癌包括过表达c-met的那些癌。适于通过本发明的fn3域来治疗的示例性癌包括上皮细胞癌、乳腺癌、卵巢癌、肺癌、结肠直肠癌、肛门癌、前列腺癌、肾癌、膀胱癌、头颈癌、胃癌、卵巢癌、胰腺癌、皮肤癌、口腔癌、食道癌、阴道癌、宫颈癌、脾癌、睾丸癌和胸腺癌。本发明的特异性结合egfr并阻碍egf结合于egfr的fn3域可用于治疗瘤,包括癌和良性瘤。适于通过本发明的fn3域来治疗的癌包括过表达egfr或变体的那些癌。适于通过本发明的fn3域来治疗的示例性癌包括上皮细胞癌、乳腺癌、卵巢癌、肺癌、结肠直肠癌、肛门癌、前列腺癌、肾癌、膀胱癌、头颈癌、卵巢癌、胰腺癌、皮肤癌、口腔癌、食道癌、阴道癌、宫颈癌、脾癌、睾丸癌和胸腺癌。本发明的包含双特异性egfr/c-metfn3域的分子或双特异性egfr/c-met抗体可用于治疗瘤,包括癌和良性瘤。适于通过本发明的包含双特异性egfr/c-metfn3域的分子或双特异性egfr/c-met抗体来治疗的示例性癌包括过表达egfr和/或c-met的那些癌,与提高的egfr活性和/或表达水平(诸如,egfr激活突变、egfr基因扩增,或配体介导的egfr激活)和提高的c-met活性和/或表达水平(诸如,c-met激活突变、c-met基因扩增,或hgf介导的c-met激活)相关的癌。可能与癌相关的示例性egfr激活突变包括点突变、缺失突变、插入突变、倒位或基因扩增,它们引起egfr的至少一种生物活性的提高,诸如提高的酪氨酸激酶活性、受体同源二聚体和异源二聚体的形成、增强的配体结合等。突变可位于egfr基因的任何部分或与egfr基因相关的调控区中,并且包括第18、19、20或21位外显子中的突变或激酶域中的突变。示例性的激活egfr突变是g719a、l861x(x为任何氨基酸)、l858r、e746k、l747s、e749q、a750p、a755v、v765m、l858p或t790m替换,e746-a750的缺失,r748-p753的缺失,ala在m766与a767之间的插入,sva(ser,val,ala)在s768与v769之间的插入,以及ns(asn,ser)在p772与h773之间的插入。egfr激活突变的其他例子是本领域已知的(参见例如美国专利公布us2005/0272083)。关于egfr和其他erbb受体的信息(包括受体同源二聚体和异源二聚体、受体配体、自磷酸化位点以及参与erbb介导的信号传导的信号传导分子)是本领域已知的(参见例如hynes和lane,naturereviewscancer5:341-354,2005)。示例性的c-met激活突变包括点突变、缺失突变、插入突变、倒位或基因扩增,它们引起c-met蛋白质的至少一种生物活性的提高,诸如提高的酪氨酸激酶活性、受体同源二聚体和异源二聚体的形成、增强的配体结合等。突变可位于c-met基因的任何部分或与该基因相关的调控区中,诸如c-met的激酶域中的突变。示例性的c-met激活突变是在残基位置n375、v13、v923、r175、v136、l229、s323、r988、s1058/t1010和e168处的突变。用于检测egfr和c-met突变或基因扩增的方法是熟知的。适于通过本发明的双特异性分子诸如本发明的双特异性egfr/c-met抗体来治疗的示例性癌,包括上皮细胞癌、乳腺癌、卵巢癌、肺癌、非小细胞肺癌(nsclc)、肺腺癌、小细胞肺癌、结肠直肠癌、肛门癌、前列腺癌、肾癌、膀胱癌、头颈癌、咽癌、鼻癌、胰腺癌、皮肤癌、口腔癌、舌癌、食道癌、阴道癌、宫颈癌、脾癌、睾丸癌、胃癌、胸腺癌、结肠癌、甲状腺癌、肝癌(肝细胞癌(hcc))、或散发性或遗传性乳头状肾细胞癌(prcc)。本发明的另一个方面是治疗患有癌的受检者的方法,所述方法包括向对其有需要的患者施用治疗有效量的分离的本发明的双特异性egfr/c-met抗体足以治疗癌的时间,其中受检者对于cd16的位置158处的苯丙氨酸是纯合的(fcγriiia-158f/f基因型)或对于cd16的位置158处的缬氨酸和苯丙氨酸是杂合的(fcγriiia-158f/v基因型)。cd16又称为fcγ受体iiia(fcγriiia)或低亲和力免疫球蛋白γfc区受体iii-a同种型。在fcγriiia蛋白质残基位置158处的缬氨酸/苯丙氨酸(v/f)多态性已显示出能影响fcγriiia对人igg的亲和力。具有fcγriiia-158f/f或fcγriiia-158f/v多态性的受体展示出与fcγriiia-158v/v相比减少的fc接合以及因此减少的adcc。人n-连接寡糖上不含岩藻糖或含少量岩藻糖改善了抗体诱导adcc的能力,这是由于抗体对人fcγriiia(cd16)的结合得以改善(shields等人,jbiolchem277:26733-40,2002)。本发明的抗体具有减少的在1%至约10%之间的岩藻糖含量。在一些实施例中,双特异性egfr/c-met抗体具有岩藻糖含量为约50%、40%、45%、40%、35%、30%、25%、20%、15%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的聚糖结构。因此,本发明的抗体可能在治疗具有fcγriiia-158f/f或fcγriiia-158f/v基因型的患者方面更有效。可使用常规方法分析患者的fcγriiia多态性。在本文所述的一些方法中,本发明的抗体可用于治疗患有癌的受检者,该癌抵抗一种或多种egfr抑制剂的治疗或者已获得对一种或多种egfr抑制剂的治疗的抗性。癌可能获得对其的抗性的示例性egfr抑制剂是抗egfr抗体,即西妥昔单抗帕尼单抗马妥珠单抗、尼妥珠单抗;小分子egfr抑制剂,即(厄洛替尼)、iressa(吉非替尼)、ekb-569(培利替尼,不可逆egfrtki)、pan-erbb;以及其他受体酪氨酸激酶抑制剂,即拉帕替尼(egfr和her2抑制剂)、培利替尼(egfr和her2抑制剂)、凡德他尼(zd6474、zactimatm、egfr、vegfr2和rettki)、pf00299804(达克替尼,不可逆pan-erbbtki)、ci-1033(不可逆pan-erbbtki)、阿法替尼(bibw2992,不可逆pan-erbbtki)、av-412(双重egfr和erbb2抑制剂)、exel-7647(egfr、erbb2、gevgr和ephb4抑制剂)、co-1686(不可逆突变体-选择性egfrtki)、azd9291(不可逆突变体-选择性egfrtki)以及hki-272(来那替尼,不可逆egfr/erbb2抑制剂)。本文所述的方法可用于治疗抵抗吉非替尼、厄洛替尼、阿法替尼、co-1686、azd9291和/或西妥昔单抗治疗的癌。可使用的示例性抗体是em1-mab。本发明的另一个方面是治疗患有癌的受检者的方法,所述方法包括向对其有需要的患者施用治疗有效量的分离的本发明的双特异性egfr/c-met抗体足以治疗癌的时间,其中该受检者抵抗厄洛替尼、吉非替尼、阿法替尼、co-1686、azd9291或西妥昔单抗治疗,或已获得对治疗的抗性。各种定性和/或定量方法可用于确定受检者是否抵抗egfr抑制剂治疗,形成对egfr抑制剂治疗的抗性或易于形成对egfr抑制剂治疗的抗性。可能与对egfr抑制剂的抗性相关的症状包括例如患者健康状况的下降或平稳、瘤尺寸的增加、瘤生长的下降停滞或变缓,和/或体内的癌细胞从一个位置向其他器官、组织或细胞的扩散。与癌相关的各种症状的重建或恶化也可指示受检者已形成或易于形成对egfr抑制剂的抗性,诸如厌食、认知功能障碍、抑郁、呼吸困难、疲乏、激素紊乱、中性粒细胞减少、疼痛、周围神经病变和性功能障碍。与癌相关的症状可根据癌的类型而变化。例如,与宫颈癌相关的症状可包括异常出血、不寻常白带过多、与正常月经周期不相关的骨盆疼痛、膀胱疼痛或排尿时疼痛,以及规律月经期之间、性交、冲洗或骨盆检查后的出血。与肺癌相关的症状可包括持续性咳嗽、咳血、呼吸短促、哮喘性胸痛、食欲不振,在没有刻意情况下的体重减轻以及疲乏。肝癌的症状可包括食欲不振及体重减轻、腹痛(尤其是可延伸到背部和肩部中的腹部右上部)、恶心与呕吐、全身无力与疲乏、肝肿大、腹部肿胀(腹水)以及皮肤和眼白发黄(黄疸)。肿瘤学方面的技术人员可易于鉴定与特定癌类型相关的症状。用于确定受检者是否形成对egfr抑制剂的抗性的其他方式包括检查癌细胞中的egfr磷酸化、erk1/2磷酸化和/或akt磷酸化,其中增加的磷酸化可指示受检者已形成或易于形成对egfr抑制剂的抗性。测定egfr、erk1/2和/或akt磷酸化的方法是熟知的并且在本文中描述。已形成对egfr抑制剂的抗性的受检者的鉴定可涉及检测升高的c-met表达水平或提高的c-met活性,例如由升高水平的循环hgf、c-met基因的激活突变或c-met基因扩增引起。本发明的另一个实施例是治疗患者中的nsclc的方法,该患者具有带有激活egfr突变或egfr基因扩增的nsclc瘤或瘤转移,该方法包括向患者施用治疗有效量的本发明的双特异性egfr/c-met抗体。本发明的双特异性egfr/c-met抗体可用于治疗非小细胞肺癌(nsclc),其包括鳞状细胞癌、腺癌和大细胞癌。在一些实施例中,nsclc的细胞具有上皮表型。在一些实施例中,nsclc已获得对一种或多种egfr抑制剂治疗的抗性。在nsclc中,egfr基因中的特定突变与对egfr酪氨酸激酶抑制剂(egfr-tki)的高响应率(70-80%)相关。egfr中的第19位外显子的5氨基酸缺失或点突变l858r与egfr-tki敏感性相关(nakata和gotoh,expertopinthertargets16:771-781,2012)。这些突变导致egfr激酶活性的非配体依赖性激活。激活egfr突变在10-30%的nsclc患者中发生,并且在东亚人、女性、从未吸烟者以及具有腺癌组织学的患者中明显更常见(janne和johnson,clincancerres12(14增刊):4416s-4420s,2006)。egfr基因扩增也与egfr-tki治疗后的响应强烈相关(cappuzzo等人,jnatlcancerinst97:643-55,2005)。虽然具有egfr突变的大多数nsclc患者最初响应于egfrtki疗法,但几乎所有患者都获得防止持久响应的抗性。由于egfr的激酶域中的第二位点点突变(t790m),50-60%的患者获得抗性。抵抗egfr酪氨酸激酶抑制剂的所有瘤中的将近60%增加c-met表达、扩增c-met基因或增加其唯一已知配体hgf(turke等人,cancercell,17:77-88,2010)。本发明的另外实施例是治疗患有癌的患者的方法,所述方法包括向对其有需要的患者施用治疗有效量的本发明的双特异性egfr/c-met抗体足以治疗癌的时间,其中癌与egfr激活突变、egfr基因扩增、升高水平的循环hgf、c-met激活突变、c-met基因扩增或突变体kras相关。在一些实施例中,egfr激活突变是g719a、g719x(x为任何氨基酸)、l861x(x为任何氨基酸)、l858r、e746k、l747s、e749q、a750p、a755v、v765m、l858p或t790m替换,e746-a750的缺失,r748-p753的缺失,ala(a)在m766与a767之间的插入,ser、val和ala(sva)在s768与v769之间的插入,以及asn和ser(ns)在p772与h773之间的插入。本发明的另外实施例是治疗患有癌的患者的方法,所述方法包括向对其有需要的患者施用治疗有效量的本发明的双特异性egfr/c-met抗体足以治疗癌的时间,其中癌与egfr突变l858r、t790m或残基e746-a750的缺失(del(e746,a750))、egfr扩增或c-met扩增相关。在一些实施例中,癌与野生型egfr和野生型c-met相关。在一些实施例中,癌与野生型egfr和c-met扩增相关。在一些实施例中,癌与egfrl858r和t790m突变及野生型c-met相关。在一些实施例中,癌与egfr缺失del(e764,a750)和野生型c-met相关。在一些实施例中,癌与egfr缺失del(e764,a750)和c-met扩增相关。在一些实施例中,癌与egfr缺失del(e764,a750)、egfr扩增和c-met扩增相关。在一些实施例中,患者患有与egfrl858r和t790m突变及野生型c-met相关的nsclc。在一些实施例中,患者患有与egfr扩增和野生型c-met相关的nsclc。在一些实施例中,患者患有与egfr扩增和c-met扩增相关的nsclc。在一些实施例中,患者患有与egfr缺失del(e764,a750)和野生型c-met相关的nsclc。在一些实施例中,患者患有与egfr缺失del(e764,a750)和c-met扩增相关的nsclc。在一些实施例中,患者用本发明的em1-mab来治疗。当瘤与l858r、t790m、del(e746,a750)egfr、egfr扩增、野生型c-met和/或c-met扩增相关时,本发明的em1-mab在体内瘤动物模型中显示疗效。egfr或c-met的扩增可通过标准方法评价,例如通过dna印迹、fish或比较基因组杂交(cgh)测定egfr或c-met基因的拷贝数。本发明的另外实施例是治疗患有癌的患者的方法,所述方法包括向对其有需要的患者施用治疗有效量的本发明的双特异性egfr/c-met抗体足以治疗癌的时间,其中癌与egfr突变l858r、t790m或残基e746-a750的缺失(del(e746,a750))、egfr扩增或c-met扩增或突变体kras相关。在一些实施例中,突变体kras具有g12v替换。kras属于编码鸟苷三磷酸酶(gtp酶)的ras原癌基因的家族,并且介导受体下游的egfr信号转导。因此预计具有原癌kras突变诸如激活g12v或g12c突变的瘤不能通过egfr抗体来治疗。采用抗egfr抗体西妥昔单抗或帕尼单抗的临床研究证实,患有kras介导的结肠直肠瘤的患者不响应于这些药剂(vancutsem等人,nengjmed360:1408-1417,2009;lievre等人,jclinoncol26:374-379,2008;amado等人,jclinoncol26:1626-1634m2008)。本发明的双特异性egfr/c-met抗体介导经由有效adcc的kras突变体细胞系杀伤,因此与当前的抗egfr疗法相反,可有效治疗癌与kras激活突变相关的患者。此类示例性的抗体是em1-mab。术语“治疗”或“处理”是指治疗处理和预防或防御措施,其中目标是防止或减缓(减轻)非期望的生理变化或失调,诸如癌的发展或扩散。对于本发明而言,有利或所需的临床结果包括但不限于症状的减轻、疾病程度的削弱、疾病的稳定(即,未恶化)状态、疾病进展的延迟或减缓、疾病状态的改善或缓和,以及缓解(不论是部分缓解还是完全缓解),而不论是可检测的或是不可检测的。“治疗”也可意指与未接受治疗时的预期生存期相比生存期延长。需要治疗的个体包括已患有病症或失调的个体以及易患病症或失调的个体或者要预防病症或失调的个体。“治疗有效量”指在所需剂量和时间段有效实现所需治疗结果的量。治疗有效量的本发明的双特异性egfr/c-met抗体可根据多种因素变化,诸如个体的疾病状态、年龄、性别和体重,以及本发明的双特异性egfr/c-met抗体引起个体中的所需响应的能力。在与抗性的关联方面可降低或减少的有效egfr/c-met治疗剂的示例性指标包括例如改善的患者健康状况、瘤尺寸的减小或缩小、瘤生长停滞或减缓,和/或不存在癌细胞向体内其他位置的转移。施用/药物组合物本发明提供包含本发明的双特异性egfr/c-met抗体和药学上可接受的载体的药物组合物。就治疗用途而言,可将本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体制备为药物组合物,该药物组合物包含药学上可接受的载体中的有效量的域、分子或抗体作为活性成分。术语“载体”是指活性化合物与其一起施用的稀释剂、辅助剂、赋形剂或媒介物。此类媒介物可以是液体,例如水和油,包括那些来源于石油、动物、植物的油或合成的油,诸如花生油、大豆油、矿物油、芝麻油等。例如,可使用0.4%盐水溶液和0.3%甘氨酸。这些溶液是无菌的,并且通常不含颗粒物。它们可通过常规的已知的灭菌技术(例如过滤)进行灭菌。组合物可根据需要含有药学上可接受的辅助物质,以接近生理条件,例如ph调节剂和缓冲剂、稳定剂、增稠剂、润滑剂和着色剂等。在此类药物制剂中本发明的分子或抗体的浓度可广泛改变,即从按重量计小于约0.5%,通常为或至少约1%到多达15或20%,并且将根据所选择的具体施用方式,主要基于所需剂量、流体体积、粘度等进行选择。包含其它人蛋白如人血清白蛋白在内的合适的媒介物和制剂描述在例如,remington:thescienceandpracticeofpharmacy,第21版,troy,d.b.编辑,lipincottwilliamsandwilkins,philadelphia,pa2006,第5部分,pharmaceuticalmanufacturing第691-1092页,特别参见第958-989页。用于本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体的治疗用途的施用方式可以是将药剂递送至宿主的任何合适的途径,诸如肠胃外施用,例如,真皮内、肌肉内、腹膜内、静脉内或皮下、肺部;经粘膜(口腔、鼻内、阴道内、直肠);使用在片剂、胶囊、溶液、粉末、凝胶、颗粒中的制剂;以及包含在注射器、植入装置、渗透泵、盒、微型泵中的制剂;或技术人员所理解的本领域熟知的其它方式。可通过例如以下方式实现位点特异性施用:胃肠外、支气管内、腹内、囊内、软骨内、腔内、体腔内、小脑内、脑室内、结肠内、颈管内、胃内、肝内、心脏内、骨内、骨盆内、心包内、腹膜内、胸膜内、前列腺内、肺内、直肠内、肾内、视网膜内、脊柱内、滑膜内、胸内、子宫内、血管内、膀胱内、病灶内、阴道、直肠、口腔、舌下、鼻内或经皮递送。因此,可将用于肌内注射的本发明药物组合物制备成包含1ml的无菌缓冲水,及约1ng至约100mg/kg之间,例如约50ng至约30mg/kg或更优选地约5mg至约25mg/kg的本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域或结合c-met的fn3域。本发明的双特异性egfr/c-met抗体可通过任何合适的途径施用给患者,例如通过静脉内(iv)输注或弹丸式注射、肌内注射或皮下注射或腹膜内注射以肠胃外方式施用。可给予iv输注少至15分钟,但更常为30分钟、60分钟、90分钟或甚至2或3小时。也可将本发明的双特异性egfr/c-met抗体直接注射到病变部位(例如,瘤自身)。给予患有癌的患者的剂量足以缓解或至少部分地抑制正治疗的疾病(“治疗有效量”),并且有时可为0.1至10mg/kg体重,例如1、2、3、4、5、6、7、8、9或10mg/kg,但可甚至更高,例如15、20、30、40、50、60、70、80、90或100mg/kg。也可给予固定的单位剂量,例如50、100、200、500或1000mg,或剂量可基于患者的表面积,例如400、300、250、200或100mg/m2。通常可施用1至8剂量(例如,1、2、3、4、5、6、7或8)以治疗癌,但可给予10、12、20或更多剂量。可在一天、两天、三天、四天、五天、六天、一周、两周、三周、一个月、五周、六周、七周、两个月、三个月、四个月、五个月、六个月或更长时间后重复本发明的双特异性egfr/c-met抗体的施用。重复疗程也是可能的,长期施用同样可能。重复施用可采用相同剂量或采用不同剂量。例如,可将用于静脉内输注的包含本发明的双特异性egfr/c-met抗体的药物组合物制备成包含约200ml的无菌林格氏溶液,及约8mg至约2400mg、约400mg至约1600mg或约400mg至约800mg的双特异性egfr/c-met抗体以便施用给80kg患者。用于制备可肠胃外施用的组合物的方法是众所周知的,并且更详细地描述在,例如,“remington'spharmaceuticalscience”,第15版,mackpublishingcompany,easton,pa。可将本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体冻干保存,并且在使用前在合适载体中重构。已证实此技术对常规蛋白制备有效,并且可以采用熟知的冻干法和重构技术。本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体可联合第二治疗剂同时地、顺序地或单独地施用。第二治疗剂可为化学治疗剂或靶向抗癌疗法。双特异性egfr/c-met抗体可连同本领域技术人员已知的化学治疗药或其他抗癌治疗剂中的任何一者或多者一起施用。化学治疗剂是可用于治疗癌的化合物,并且包括生长抑制剂或其他细胞毒素剂,并且包括烷化剂、抗代谢药、抗微管抑制剂、拓扑异构酶抑制剂、受体酪氨酸激酶抑制剂、血管生成抑制剂等。化学治疗剂的例子包括烷化剂,诸如噻替哌和环磷酰胺烷基磺酸酯,诸如白消安、英丙舒凡和哌泊舒凡;氮丙啶类,诸如苯佐替哌(benzodopa)、卡波醌、美妥替哌(meturedopa)和乌瑞替哌(uredopa);乙撑亚胺和甲基蜜胺,包括六甲蜜胺、三乙撑蜜胺、三亚乙基磷酰胺、三亚乙基硫代磷酰胺和三羟甲基蜜胺;氮芥,诸如苯丁酸氮芥、萘氮芥、胆磷酰胺、雌氮芥、异环磷酰胺、双氯乙基甲胺、盐酸氧氮芥、美法仑、新恩比兴、苯芥胆甾醇、松龙苯芥、曲磷胺、尿嘧啶氮芥;亚硝基脲,诸如卡莫司汀、氯脲霉素、福莫司汀、洛莫司汀、尼莫司汀、雷莫司汀;抗生素,诸如阿克拉霉素、放线菌素、安曲霉素、重氮丝氨酸、博来霉素、放线菌素c、加利车霉素、卡柔比星(carabicin)、洋红霉素、嗜癌菌素、色霉素、放线菌素d、柔红菌素、6-重氮-5-氧-l-正亮氨酸、多柔比星、表柔比星、依索比星、伊达比星、麻西罗霉素、丝裂霉素、霉酚酸、诺加霉素、橄榄霉素、培洛霉素、泊非霉素(potfiromycin)、嘌呤霉素、三铁阿霉素、罗多比星、链黑霉素、链脲霉素、杀结核菌素、乌苯美司、净司他丁、佐柔比星;抗代谢药,诸如甲氨蝶呤和5-fu;叶酸类似物,诸如二甲叶酸、甲氨蝶呤、蝶罗呤、三甲曲沙;嘌呤类似物,诸如氟达拉滨、6-巯基嘌呤、硫咪嘌呤、硫鸟嘌呤;嘧啶类似物,诸如安西他滨、阿扎胞苷、6-氮尿苷、卡莫氟、阿糖胞苷、双脱氧尿苷、去氧氟尿苷、依诺他滨、氟尿苷;雄激素类,诸如二甲睾酮、丙酸甲雄烷酮、环硫雄醇、美雄烷、睾内酯;抗肾上腺类,诸如氨鲁米特、米托坦、曲洛司坦;叶酸补充剂,诸如亚叶酸;醋葡醛内酯;醛磷酰胺糖苷;氨基乙酰丙酸;安吖啶;bestrabucil;比生群;依达曲沙(edatraxate);地磷酰胺(defofamine);秋水仙胺;地吖醌;依氟鸟氨酸(elfornithine);依利醋铵;依托格鲁;硝酸镓;羟基脲;香菇多糖;氯尼达明;米托胍腙;米托蒽醌;莫哌达醇;硝氨丙吖啶;喷司他丁;苯来美特(phenamet);吡柔比星;足叶草酸;2-乙基酰肼;丙卡巴肼;雷佐生;西索菲兰;锗螺胺;细交链孢菌酮酸;三亚胺醌;2,2',2"-三氯三乙胺;乌拉坦;长春地辛;达卡巴嗪;甘露醇氮芥;二溴甘露醇;二溴卫矛醇;哌泊溴烷;加西托星(gacytosine);阿拉伯糖苷(“ara-c”);环磷酰胺;噻替哌;紫杉类或紫杉烷家族的成员,诸如紫杉醇(多西他赛及其类似物;苯丁酸氮芥;吉西他滨;6-硫鸟嘌呤;巯嘌呤;甲氨蝶呤;铂类似物,诸如顺铂和卡铂;长春碱;铂;依托泊苷(vp-16);异环磷酰胺;丝裂霉素c;米托蒽醌;长春新碱;长春瑞滨;诺维本;诺消灵;替尼泊苷;柔红霉素;氨基蝶呤;希罗达;伊班膦酸盐;cpt-11;拓扑异构酶抑制剂rfs2000;二氟甲基鸟氨酸(dmfo);维甲酸;艾司波霉素(esperamicins);卡培他滨;受体酪氨酸激酶和/或血管生成的抑制剂,包括索拉非尼舒尼替尼帕唑帕尼(votrienttm)、托西尼布(palladiatm)、凡德他尼(zactimatm)、西地尼布瑞格非尼(bay73-4506)、阿西替尼(ag013736)、来他替尼(cep-701)、厄洛替尼吉非替尼(iressatm)、bibw2992(tovoktm)、拉帕替尼来那替尼(hki-272)等,以及任何上述物质的药学上可接受的盐、酸或衍生物。还包括在该定义中的是抗激素剂,其用于调节或抑制激素对瘤的作用,诸如抗雌激素类,包括例如他莫昔芬、雷洛昔芬、抑制芳香化酶的4(5)-咪唑类、4-羟基他莫昔芬、曲沃昔芬、可莫昔芬(keoxifene)、ly117018、奥那司酮和托瑞米芬以及抗雄激素类,诸如氟他胺、尼鲁米特、比卡鲁胺、亮丙瑞林和戈舍瑞林;以及任何上述物质的药学上可接受的盐、酸或衍生物。其他常规细胞毒性化合物(如wiemann等人,1985,载于medicaloncology(calabresi等人编辑),第10章,mcmillanpublishing中所公开的那些化合物)也可应用于本发明的方法。可与本发明的包含双特异性egfr/c-metfn3域的分子、结合egfr的fn3域、结合c-met的fn3域或双特异性egfr/c-met抗体联合使用的示例性药剂包括酪氨酸激酶抑制剂和靶向抗癌疗法诸如(吉非替尼)和tarceva(厄洛替尼)及her2、her3、her4或vegf的其他拮抗剂。示例性的her2拮抗剂包括cp-724-714、herceptintm(曲妥珠单抗)、omnitargtm(帕妥珠单抗)、tak-165、拉帕替尼(egfr和her2抑制剂)和gw-282974。示例性的her3拮抗剂包括抗her3抗体(参见例如美国专利公布us2004/0197332)。示例性的her4拮抗剂包括抗her4sirna(参见例如maatta等人,molbiolcell17:67-79,2006)。示例性的vegf拮抗剂为贝伐单抗(avastintm)。当小分子与本发明的双特异性egfr/c-met抗体联合使用时,其通常更频繁地施用,优选地一天一次,但每天2、3、4或更多次也是可能的,每两天一次、每周一次或某个其他时间间隔同样可能。小分子药物通常以口服方式服用,但肠胃外施用也是可能的,例如通过iv输注或弹丸式注射或皮下或肌内注射。小分子药物的剂量可通常为10至1000mg,或约100、150、200或250mg。当本发明的双特异性egfr/c-met抗体与第二治疗剂联合施用时,该组合可在任何方便的时间段内发生。例如,可在同一天且甚至在相同静脉内输注中,将双特异性egfr/c-met抗体和第二治疗剂施用给患者。然而,也可在隔天或隔周、隔两周或隔月等施用双特异性egfr/c-met抗体和第二治疗剂。在一些方法中,双特异性egfr/c-met抗体和第二治疗剂以足够接近的时间施用,使得它们在正治疗的患者中同时以可检测水平存在(例如,存在于血清中)。在一些方法中,由一段时间内多剂量组成的双特异性egfr/c-met抗体的整个疗程之后或者之前是也由多剂量组成的第二治疗剂的疗程。在一些方法中,如果患者具有对最初施用的第二治疗剂的抗性或形成对该第二治疗剂的抗性,则开始接着施用双特异性egfr/c-met抗体进行治疗。患者可接受双特异性egfr/c-met抗体和第二治疗剂中一者或两者的仅单个疗程或多个疗程。双特异性egfr/c-met抗体和第二治疗剂施用期间可使用1、2或若干天或周的恢复期。当已确立第二治疗剂的合适治疗方案时,该方案可与本发明的双特异性egfr/c-met抗体联合使用。例如,(厄洛替尼)作为100mg或150mg丸剂每天服用一次,并且(吉非替尼)作为250mg片剂每天服用一次。双特异性egfr/c-met抗体,任选地与第二治疗剂联用,可连同任何形式的放射疗法和/或外科手术一起施用,该放射疗法包括外照射、调强放射疗法(imrt)和任何形式的放射外科手术,包括伽玛刀、射波刀、直线加速器和组织间放射(例如,植入的放射性粒子、gliasite球囊)。与放射疗法的联用可尤其适用于头颈癌和脑瘤。虽然以一般的术语描述了本发明,但是本发明的实施例还将在以下的实例中公开,该实例不应理解为对本权利要求所保护的范围的限制。实例1.tencon文库的构建tencon(seqidno:1)是一种免疫球蛋白样支架,即由来自人腱生蛋白-c的十五个fn3域的共有序列而设计的iii型纤连蛋白(fn3)域(jacobs等人,proteinengineering,design,andselection,25:107-117,2012;美国专利公布2010/0216708)。tencon的晶体结构显示连接七条β-链的六个表面外露的环。这些环或每个环内的所选残基可被随机化以便构建iii型纤连蛋白(fn3)域的文库,该文库可用于选择能结合于特定靶标的新型分子。tencon:lpapknlvvsevtedslrlswtapdaafdsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvkgghrsnplsaeftt(seqidno1):tcl1文库的构建构建被设计成仅使tencon(seqidno:1)的fg环随机化的文库tcl1,以与顺式展示系统一起使用(jacobs等人,proteinengineering,design,andselection,25:107-117,2012)。在该系统中,产生了掺入tac启动子、tencon文库编码序列、repa编码序列、顺式元件和复制起点元件的序列的单链dna。在体外转录/翻译系统中表达后,tencon-repa融合蛋白顺式结合于编码该融合蛋白的dna而产生了复合物。然后通过如下所述的聚合酶链反应(pcr)分离并扩增结合于靶分子的复合物。与顺式展示一起使用的tcl1文库的构建通过连续数轮pcr实现,以产生两个半部的最终线性的双链dna分子;5’片段包含启动子和tencon序列,而3’片段包含repa基因及顺式元件和复制起点元件。这两个半部通过限制酶切消化来组合,以产生完整构建体。tcl1文库被设计成仅在tencon的fg环中掺入随机氨基酸kgghrsn(seqidno:86)。nns密码子用于构建该文库,使得可能将所有20种氨基酸和一个终止密码子掺入到fg环中。tcl1文库包含六个单独的亚文库,各亚文库具有7至12个残基的不同随机化fg环长度,以进一步增加多样性。基于tencon的文库的设计示于表2中。表2.*tapdaafd:seqidno:1的第22-28位残基;**kgghrsn:seqidno:86x是指由nns密码子编码的简并氨基酸。#是指正文中所述的“氨基酸的设计分布”。为了构建tcl1文库,执行连续数轮pcr以附加tac启动子、在fg环中建立简并性以及添加最终组装所需的限制位点。首先,通过pcr在两个步骤中产生包含启动子序列和fg环的tencon序列5’的dna序列。对应于完全tencon基因序列的dna用作采用引物pop2220(seqidno:2)和tc5’tofg(seqidno:3)的pcr模板。从该反应所得的pcr产物用作采用引物130mer(seqidno:4)和tc5’tofg的下一轮pcr扩增的模板,以完成5’和启动子序列向tencon的附加。接下来,通过如下方式将多样性引入fg环中:采用正向引物pop2222(seqidno:5)以及包含简并核苷酸的反向引物tcf7(seqidno:6)、tcf8(seqidno:7)、tcf9(seqidno:8)、tcf10(seqidno:9)、tcf11(seqidnno:10)或tcf12(seqidno:11)扩增在第一步中产生的dna产物。每个亚文库执行至少八个100μl的pcr反应以使pcr循环最小化并使文库的多样性最大化。对至少5μg的该pcr产物进行凝胶纯化并用于采用引物pop2222(seqidno:5)和pop2234(seqidno:12)的随后pcr步骤,从而导致6xhis标签和noti限制位点连接到tencon序列的3’端。该pcr反应仅使用十五个pcr循环和至少500ng的模板dna来进行。对所得pcr产物进行凝胶纯化,用noti限制酶消化,并通过qiagen柱纯化。文库的3’片段是恒定dna序列,该序列包含用于展示的元件,包括pspomi限制位点、repa基因的编码区以及顺式元件和复制起点元件。使用包含该dna片段的质粒(pcr4blunt)(invitrogen),利用m13正向引物和m13反向引物执行pcr反应。通过pspomi将所得pcr产物消化过夜,并进行凝胶纯化。为了将文库dna的5’部分连接到包含repa基因的3’dna,在noti和pspomi酶及t4连接酶存在下,将2pmol的5’dna连接到等摩尔量的3’repadna。在37℃下过夜连接后,将小部分的连接dna在凝胶上电泳以检查连接效率。将连接文库产物分成十二次pcr扩增,并且采用引物对pop2250(seqidno:13)和didligrev(seqidno:14)运行12个循环的pcr反应。tcl1文库的每个亚文库的dna产量在32-34μg范围内。为了评估文库的质量,采用引物tcon5new2(seqidno:15)和tcon6(seqidno:16)扩增小部分的工作文库,并经由不依赖连接酶的克隆法克隆到修饰pet载体中。将质粒dna转化到bl21-gold(de3)感受态细胞(stratagene)中并且使用t7启动子引物对96个随机挑取的菌落进行测序。未发现重复的序列。总的说来,大约70-85%的克隆具有完整的启动子和tencon编码序列而无移码突变。功能序列比率(排除具有终止密码子的克隆)在59%与80%之间。tcl2文库的构建构建tcl2文库,其中tencon的bc和fg环被随机化并且每个位置处的氨基酸的分布受到严格控制。表3示出了tcl2文库中所需环位置处的氨基酸分布。所设计的氨基酸分布具有两个目标。首先,文库偏向于这样的残基,根据tencon晶体结构的分析和/或同源建模,预测这些残基在结构上对于tencon折叠和稳定性是重要的。例如,位置29固定为仅亚组的疏水氨基酸,因为该残基掩埋于tencon折叠的疏水核中。第二层设计包括使氨基酸分布偏向在抗体的重链hcdr3中优先发现的残基,以有效产生高亲和力结合物(birtalan等人,jmolbiol377:1518-28,2008;olson等人,proteinsci16:476-84,2007)。为了达到这个目的,表3的“所设计的分布”是指如下的分布:6%丙氨酸、6%精氨酸、3.9%天冬酰胺、7.5%天冬氨酸、2.5%谷氨酸、1.5%谷氨酰胺、15%甘氨酸、2.3%组氨酸、2.5%异亮氨酸、5%亮氨酸、1.5%赖氨酸、2.5%苯丙氨酸、4%脯氨酸、10%丝氨酸、4.5%苏氨酸、4%色氨酸、17.3%酪氨酸和4%缬氨酸。该分布没有甲硫氨酸、半胱氨酸和终止密码子。表3.*残基编号基于seqidno:1的tencon序列tcl2文库的5’片段包含启动子和tencon(seqidno:1)的编码区,其被化学合成为文库集合(librarypool)(sloningbiotechnology)。dna的该集合包含至少1×1011个不同成员。在片段的末端,bsai限制位点包括在设计中以用于连接到repa。文库的3’片段是恒定dna序列,该序列包含用于展示的元件,包括6xhis标签、repa基因的编码区以及顺式元件。使用既有dna模板(见上)以及引物ls1008(seqidno:17)和didligrev(seqidno:14),通过pcr反应制备dna。为了组装完整的tcl2文库,将总计1μg的经bsai消化的5’tencon文库dna连接到3.5μg的3’片段,该3’片段通过采用相同酶的限制性消化来制备。在过夜连接后,通过qiagen柱纯化dna,并通过测量260nm下的吸光度来定量dna。通过采用引物对pop2250(seqidno:13)和didligrev(seqidno:14)的12个循环的pcr反应来扩增连接文库产物。执行总计72个反应,每个反应包含50ng的连接dna产物作为模板。tcl2工作文库dna的总产量为约100μg。将小部分的工作文库亚克隆并测序,如上文针对文库tcl1所述。未发现重复的序列。约80%的序列包含完整的启动子和tencon编码序列而无移码突变。tcl14文库的构建顶部(bc、de和fg)和底部(ab、cd和ef)环,例如fn3域中报道的结合表面被形成fn3结构的中心的β-链分隔。存在于fn3域的两个“侧面”上且形状与仅由环形成的表面不同的另选表面,在fn3域的一个侧面处由两条反平行β-链即c和fβ-链以及cd和fg环形成,并且本文称为c-cd-f-fg表面。使tencon的另选表面随机化的文库通过如下方式产生:使所选c和f链的表面外露残基以及cd和fg环的部分随机化,如图1中所示。与tencon(seqidno:1)相比具有下列替换的tencon变体tencon27(seqidno:99)用于产生文库;e11rl17a、n46v、e86i。用于构建该文库的方法的完整描述描述于美国专利公布us2013/0226834。实例2:结合egfr并抑制egf结合的iii型纤连蛋白(fn3)域的选择文库筛选使用顺式展示从tcl1和tcl2文库选择egfr结合域。使用标准方法对融合到igg1fc的egfr(r&dsystems)的重组人胞外域进行生物素酰化,并用于淘选(seqidno:73的全长egfr的第25-645位残基)。为了体外转录和翻译(itt),将2-6μg的文库dna与0.1mm完整氨基酸、1xs30预混组分以及30μl的s30提取物(promega)一起在100μl的总体积中温育,并且在30℃下温育。1小时后,加入450μl的封闭溶液(pbsph7.4,补充有2%牛血清白蛋白、100μg/ml鲱鱼精dna和1mg/ml肝素),并且将反应在冰上温育15分钟。通过将重组人egf(r&dsystems)与生物素酰化重组egfr-fc在封闭溶液中在室温下混合1小时,以1:1和10:1摩尔比的egfr与egf来组装egfr-fc:egf复合物。为进行结合,将500μl经封闭的itt反应与100μl的egfr-fc:egf复合物混合,并且在室温下温育1小时,之后采用中性亲和素或链霉亲和素磁珠(seradyne)将结合复合物拉下。通过用pbst和pbs连续洗涤来移除未结合的文库成员。洗涤之后,通过加热到65℃持续10分钟将dna从结合复合物洗脱下来,通过pcr来扩增,并通过限制性消化和连接将其连接到编码repa的dna片段,以供随后几轮淘选所用。通过在每轮期间将靶标egfr-fc的浓度从200nm到50nm连续地降低并且增加洗涤严格性,来分离高亲和力结合物。在第4和5轮中,通过在10倍摩尔过量的非生物素酰化egfr-fc存在下在pbs中洗涤过夜,来移除未结合和弱结合的fn3域。在淘选后,使用标准分子生物学技术,采用寡核苷酸tcon5new2(seqidno:15)和tcon6(seqidno:16)通过pcr来扩增所选fn3域,亚克隆到经修饰而包括不依赖连接酶的克隆位点的pet载体中,并且转化到bl21-gold(de3)(stratagene)细胞中,以用于在大肠杆菌中进行可溶性表达。将编码c-末端聚组氨酸标签的基因序列添加到每个fn3域以实现纯化和检测。在37℃下使培养物在1ml的96孔板中补充有100μg/ml羧苄青霉素的2yt培养基中生长到0.6-0.8的光密度,然后添加终浓度为1mm的iptg,此时温度下降到30℃。大约16小时后通过离心收获细胞并在-20℃下冻存。通过如下方式实现细胞裂解:在摇动的情况下将各沉淀物在0.6ml的ht裂解缓冲液(novagenemdbiosciences)中在室温下温育45分钟。结合细胞上的egfr的fn3域的选择为了在更具生理性的环境中评估不同fn3域结合egfr的能力,测量了它们结合a431细胞的能力。a431细胞(americantypeculturecollection,目录号crl-1555)过表达egfr,每个细胞含约2×106个受体。将细胞以5,000/孔接种到不透明的黑色96孔板中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。将表达fn3域的细菌裂解物在facs染色缓冲液(bectondickinson)中稀释1,000倍,并且在三个平行测定板中在室温下温育1小时。移除裂解物,并且用150μl/孔的facs染色缓冲液将细胞洗涤3次。将细胞与50μl/孔的抗pentahis-alexa488抗体缀合物(qiagen)(以1:100稀释于facs染色缓冲液中)在室温下温育20分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次,之后在孔中充入100μl的facs染色缓冲液并且使用acumenex3读板机读取488nm下的荧光。针对其结合a431细胞的能力来筛选包含fn3域的细菌裂解物(用于tcl1和tcl2文库的1320个粗细菌裂解物),并且鉴定出516个阳性克隆,其中结合性超过背景信号≥10倍。针对结合来筛选来自tcl14文库的300个裂解物,得到结合于egfr的58个独特的fn3域序列。抑制egf结合于细胞上的egfr的fn3域的选择为了更好地表征egfr结合的机制,使用a431细胞测量了各种鉴定的fn3域克隆以egf竞争性方式结合egfr的能力,并且与a431结合测定筛选并行运行。将a431细胞以5,000/孔接种到不透明的黑色96孔板中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。将细胞与50μl/孔的1:1,000稀释的细菌裂解物在三个平行测定板中在室温下温育1小时。将生物素酰化egf(invitrogen,目录号e-3477)加入每个孔中达到30ng/ml的终浓度,并在室温下温育10分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次。将细胞与50μl/孔的链霉亲和素-藻红蛋白缀合物(invitrogen)(以1:100稀释于facs染色缓冲液中)在室温下温育20分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次,之后在孔中充入100μl的facs染色缓冲液并且使用acumenex3读板机读取600nm下的荧光。在上述egf竞争测定法中筛选包含fn3域的细菌裂解物。筛选来自tcl1和tcl2文库的1320个粗细菌裂解物,得到将egf结合抑制>50%的451个阳性克隆。鉴定的结合egfr的fn3域的表达和纯化用hismultitraptmhp板(gehealthcare)从澄清的大肠杆菌裂解物纯化带his标签的fn3域,并在ph7.4的包含20mm磷酸钠、500mm氯化钠和250mm咪唑的缓冲液中洗脱。使用pdmultitraptmg-25板(gehealthcare)将经纯化的样品交换到pbsph7.4中以用于分析。尺寸排阻色谱法分析使用尺寸排阻色谱法测定结合egfr的fn3域的聚集状态。将每个经纯化的fn3域的等分试样(10μl)在pbsph7.4的流动相中以0.3ml/min的流量注射到superdex755/150柱(gehealthcare)上。通过280nm处的吸光度监测来自柱的洗脱液。将通过sec显示出高水平聚集的fn3域排除,不进行进一步分析。所选结合egfr的fn3域从egfr-fc的解离速率在proteonxpr-36仪器(bio-rad)上筛选所选的结合egfr的fn3域,以鉴定在结合于egfr-fc时具有缓慢解离速率(koff)的那些,从而有利于选择高亲和力结合物。将5μg/ml浓度的山羊抗人fcigg(r&dsystems)以包含0.005%吐温-20的pbs中30μl/min的流量经由胺偶联(在ph5.0下)沿芯片上的水平取向直接固定在所有6个配体通道上。固定密度平均为约1500响应单位(ru),并且不同通道间的变化率小于5%。egfr-fc被捕获于抗人fcigg表面,在竖直配体取向上达到约600ru的密度。将所有经测试的fn3域归一化为1μm的浓度并且测试它们在水平取向上的结合。所有6个分析物通道用于fn3域以使筛选通量最大化。以100μl/min的流量监测离解相10分钟。点间结合信号用作参照以监测分析物与固定igg表面之间的非特异性结合,并且从所有结合响应扣除。经处理的结合数据局部拟合到1:1简单朗缪尔结合模型,以提取结合于捕获的egfr-fc的每个fn3域的koff。egf刺激的egfr磷酸化的抑制测试了经纯化的结合egfr的fn3域在单一浓度下抑制egf刺激的a431细胞中egfr的磷酸化的能力。使用egfr磷酸化(tyr1173)试剂盒(mesoscalediscovery)监测egfr磷酸化。将细胞以20,000/孔接种于透明96孔组织培养处理板(nunc)中含glutamaxtm与10%胎牛血清(fbs)(gibco)的100μl/孔rpmi培养基(gibco)中,并使之在37℃下于潮湿5%的co2气氛中附着过夜。将培养基完全移除,并且在100μl/孔的不含fbs的培养基中在37℃下于潮湿5%的co2气氛中对细胞进行饥饿处理过夜。然后用100μl/孔的包含2μm浓度的结合egfr的fn3域的预热(37℃)饥饿培养基,在37℃下于潮湿5%的co2气氛中将细胞处理1小时。对照仅用饥饿培养基处理。通过如下方式刺激细胞:加入100μl/孔的包含100ng/ml重组人egf(r&dsystems,目录号236-eg)(终浓度为50ng/mlegf和1μm结合egfr的fn3域)的预热(37℃)饥饿培养基并轻轻混合,并且在37℃、5%的co2下温育15分钟。一组对照孔保持不刺激,作为阴性对照。将培养基完全移除,并且根据制造商说明书,在摇动的情况下在室温下用100μl/孔的完全裂解缓冲液(mesoscalediscovery)裂解细胞10分钟。根据制造商说明书,用所提供的封闭溶液在室温下将被配置用于测量第1173位酪氨酸上磷酸化的egfr的测定板(mesoscalediscovery)封闭1.5-2小时。然后用200μl/孔的1xtris洗涤缓冲液(mesoscalediscovery)将板洗涤4次。将细胞裂解物的等分试样(30μl/孔)转移到测定板,将测定板用板密封膜(vwr)覆盖并且在摇动的情况下在室温下温育1小时。用200μl/孔的tris洗涤缓冲液将测定板洗涤4次,之后将25μl的冰冷检测抗体溶液(mesoscalediscovery)加入每孔中,应小心操作,避免引入气泡。在摇动的情况下在室温下将板温育1小时,然后用200μl/孔的tris洗涤缓冲液洗涤4次。通过如下方式检测信号:加入150μl/孔的读取缓冲液(readbuffer)(mesoscalediscovery)并且使用制造商安装的测定专用默认设置,在成像器6000仪器(mesoscalediscovery)上读数。对每个结合egfr的fn3域计算egf刺激的阳性对照信号的抑制百分比。对来自tcl1和tcl2文库的232个鉴定的克隆测量egf刺激的egfr磷酸化的抑制。这些克隆中的22个在1μm浓度下将egfr磷酸化抑制≥50%。在将弱表达或经尺寸排阻色谱法判定为多聚体的克隆移除之后,九个克隆接着进行进一步的生物表征。这些克隆的bc和fg环序列示于表4中。九个所选克隆中的八个具有共同的fg环序列(hnvykdtnmrgl;seqidno:95)并且在若干克隆之间在其bc环序列中可看到显著相似性的区域。表4.实例3:抑制egf结合的结合egfr的fn3域的表征大规模表达、纯化和内毒素移除表4中所示的fn3域按比例放大以提供用于详细表征的更多材料。使用包含每种结合egfr的fn3域变体的过夜培养物,以过夜培养物在新鲜培养基中的1/80稀释液接种0.8l补充有100μg/ml氨比西林的terrific肉汤培养基,并且在摇动的情况下在37℃下温育。当600nm下的光密度达到约1.2-1.5时,通过添加终浓度为1mm的iptg来诱导培养物,并且温度下降到30℃。在4小时后,通过离心收集细胞,并且将细胞沉淀物保存于-80℃下直到需要为止。为了进行细胞裂解,以每克沉淀物5ml的的比率,将解冻的沉淀物重悬于补充有25u/ml的(sigma-aldrich)和1ku/ml的rlysozymetm(novagenemdbiosciences)的1x中。在轻轻搅拌的情况下在室温下进行裂解1小时,然后在4℃下以56,000×g离心50分钟。收集上清液并通过0.2μm滤膜过滤,然后使用gehealthcare100s色谱系统将其上样于用缓冲液a(50mmtris-hclph7.5、500mmnacl、10mm咪唑)预先平衡的5ml的histrapff柱。将柱用20个柱体积的缓冲液a洗涤,并用6个柱体积的16%缓冲液b(50mmtris-hclph7.5、500mmnacl、250mm咪唑)进一步洗涤。将fn3域用10个柱体积的50%b、接着是超过6个柱体积的50-100%b的梯度进行洗脱。将包含fn3域蛋白的级分合并,使用millipore10kmwco浓缩器浓缩,并过滤,再上样于用pbs预先平衡的hiloadtm16/60superdextm75柱(gehealthcare)上。从尺寸排阻柱洗脱的蛋白单体峰得以保留。使用acticleanetox树脂(sterogenebioseparations),利用分批方法移除内毒素。在内毒素移除之前,将树脂用1nnaoh在37℃下预处理2小时(或4℃过夜)并且用pbs充分洗涤直到用ph试纸测得ph已稳定到约7。将经纯化的蛋白通过0.2μm滤膜过滤,再以10ml的蛋白与1ml的树脂的比率加入到1ml的etox树脂中。在轻轻旋转的情况下使内毒素对树脂的结合在室温下进行至少2小时。通过以500×g离心2分钟来移除树脂,并且保留蛋白上清液。使用-ptstm小柱测量内毒素水平,并且在-mcs读板机(charlesriver)分析。如果在第一次etox处理后内毒素水平高于5eu/mg,则重复以上程序直到内毒素水平降至≥5eu/mg。在用etox进行两次连续处理后内毒素水平高于5eu/mg并且稳定的情况下,确定蛋白质的阴离子交换或疏水相互作用色谱法条件,以移除剩余的内毒素。所选结合egfr的fn3域对egfr-fc的亲和力测定(egfr-fc亲和力)所选结合egfr的fn3域对重组egfr胞外域的结合亲和力进一步使用proteon仪器(biorad)通过表面等离子共振方法表征。测定装置(芯片制作,egfr-fc捕获)类似于以上针对解离速率分析所述的装置。以3倍稀释系列在1μm浓度下测试水平取向的所选结合egfr的fn3域。还注入缓冲液样品以监测基线稳定性。以100μl/min的流量监测所有浓度的每个结合egfr的fn3域的离解相30分钟(对于来自解离速率筛选的koff为约10-2s-1的那些)或1小时(对于koff为约10-3s-1或更慢的那些)。将两组参考数据从响应数据扣除:1)点间信号,以校正结合egfr的fn3域与固定化igg表面之间的非特异性相互作用;2)缓冲液通道信号,以校正因捕获的egfr-fc表面随时间推移的离解而引起的基线漂移。将在所有浓度下对每个fn3域处理的结合数据全局地拟合到1:1简单朗缪尔结合模型,以提取动力学(kon、koff)和亲和力(kd)常数的估计值。表5示出了每个构建体的动力学常数,并且亲和力从200pm变化到9.6nm。所选结合egfr的fn3域对细胞上的egfr的结合(“a431细胞结合测定”)将a431细胞以5,000/孔接种到不透明的黑色96孔板中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。在室温下将经纯化的结合egfr的fn3域(1.5nm至30μm)加入到三个平行测定板中的细胞(50ul)中持续1小时。移除上清液,并且用150μl/孔的facs染色缓冲液将细胞洗涤3次。将细胞与50μl/孔的抗pentahis-alexa488抗体缀合物(qiagen)(以1:100稀释于facs染色缓冲液中)在室温下温育20分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次,之后在孔中充入100μl的facs染色缓冲液并且使用acumenex3读板机读取488nm下的荧光。将数据作为原始荧光信号对fn3域摩尔浓度的对数作图,并且使用graphpadprism4(graphpadsoftware)拟合到具有可变斜率的s形剂量-响应曲线,以计算ec50值。表5报告了2.2nm至>20μm范围内的每个构建体的ec50。使用所选结合egfr的fn3域对egf结合于细胞上的egfr的抑制(a431细胞egf竞争测定法)将a431细胞以5,000/孔接种到不透明的黑色96孔板中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。在室温下将经纯化的结合egfr的fn3域(1.5nm至30μm)加入到三个平行测定板中的细胞(50μl/孔)中持续1小时。将生物素酰化egf(invitrogen,目录号:e-3477)加入每个孔中达到30ng/ml的终浓度,并在室温下温育10分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次。将细胞与50μl/孔的链霉亲和素-藻红蛋白缀合物(invitrogen)(以1:100稀释于facs染色缓冲液中)在室温下温育20分钟。用150μl/孔的facs染色缓冲液将细胞洗涤3次,之后在孔中充入100μl的facs染色缓冲液并且使用acumenex3读板机读取600nm下的荧光。将数据作为原始荧光信号对fn3域摩尔浓度的对数作图,并且使用graphpadprism4(graphpadsoftware)拟合到具有可变斜率的s形剂量-响应曲线,以计算ic50值。表5报告了1.8nm至121nm范围内的ic50值。egf刺激的egfr磷酸化的抑制(磷酸化egfr测定)通过测量抑制的ic50值,更全面地评估显著抑制egf刺激的egfr磷酸化的所选fn3域。在不同fn3域浓度(0.5nm至10μm)下评估egf刺激的egfr磷酸化的抑制,如以上在“egf刺激的egfr磷酸化的抑制”中所述。将数据作为电化学发光信号对fn3域摩尔浓度的对数作图,并且通过使用graphpadprism4(graphpadsoftware)将数据拟合到具有可变斜率的s形剂量响应,来测定ic50值。表5示出了在18nm至>2.5μm范围内的ic50值。人瘤细胞生长的抑制(nci-h292生长和nci-h322生长测定)通过如下方式评估egfr依赖性细胞生长的抑制:在暴露于结合egfr的fn3域之后,测量过表达egfr的人瘤细胞系nci-h292和nci-h322(americantypeculturecollection,分别为目录号crl-1848和目录号crl-5806)的活力。将细胞以500个细胞/孔(nci-h292)或1,000个细胞/孔(nci-h322)接种于不透明的白色96孔组织培养处理板(nunc)中含glutamaxtm和10mmhepes且补充有10%热灭活胎牛血清(gibco)和1%青霉素/链霉素(gibco)的100μl/孔rpmi培养基(gibco)中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。通过加入包含某浓度范围结合egfr的fn3域的5μl/孔的磷酸盐缓冲盐水(pbs)来处理细胞。对照仅用5μl/孔的pbs或pbs中的25mm乙二胺四乙酸处理。将细胞在37℃、5%的co2下温育120小时。通过如下方式检测活细胞:加入75μl/孔的celltiter-试剂(promega),然后在板摇动器上混合2分钟,并且在室温下在黑暗中再温育10分钟。在设定为发光模式的spectramaxm5读板机(moleculardevices)上读取板,对于仅空白介质的读取时间为0.5秒/孔。将数据作为经pbs处理的细胞生长的百分比对fn3域摩尔浓度的对数作图。使用graphpadprism4(graphpadsoftware)将数据拟合到具有可变斜率的s形剂量响应的方程,来测定ic50值。表5示出了分别使用nci-h292和nci-h322细胞时5.9nm至1.15μm以及9.2nm至>3.1μm范围内的ic50值。表5示出了每个测定法的结合egfr的fn3域的生物学特性的汇总。表5.实例4:结合egfr的fn3域的工程改造亚组的结合egfr的fn3域被工程改造为增加每个分子的构象稳定性。通过dna合成将已表明改善fn3域稳定性的突变l17a、n46v和e86i(描述于美国专利公布us2011/0274623)掺入到克隆p54ar4-83、p54cr4-31和p54ar4-37中。如上所述,将新突变体p54ar5-83v2、p54cr431-v2和p54ar4-37v2表达并纯化。使用pbs中的差示扫描量热法评估每个突变体的稳定性,以便将其与对应亲本分子的稳定性相比较。表6示出了每个变体分子被显著稳定,其中tm的平均增加值为18.5℃。表6.fn3域克隆seqidno:tm(℃)p54ar4-832550.6p54ar4-83v22769.8p54cr4-312660.9p54cr4-31v22878.9p54ar4-372245.9p54ar4-37v22964.2实例5:结合c-met并抑制hgf结合的iii型纤连蛋白(fn3)域的选择对人c-met的淘选针对生物素酰化人c-met胞外域(bt-c-met)来筛选tcl14文库,以鉴定能够特异性结合c-met的fn3域。对于选择,使3μg的tcl14文库在大肠杆菌s30线性提取物(promega,madison,wi)中体外转录并翻译(ivtt),并且表达的文库用cisblock(2%的bsa(sigma-aldrich,st.louis,mo)、100μg/ml鲱鱼精dna(promega)、1mg/ml肝素(sigma-aldrich))封闭。对于选择,bt-c-met以浓度为400nm(第1轮)、200nm(第2和3轮)和100nm(第4和5轮)添加。结合的文库成员使用中和亲和素磁珠(thermofisher,rockford,il)(第1、3和5轮)或链霉亲和素磁珠(promega)(第2和4轮)回收,并且未结合的文库成员通过用500ul的pbs-t洗涤珠粒5-14次接着用500μl的pbs洗涤2次来移除。进行另外的选择轮以便鉴定具有提高的亲和力的fn3域分子。简而言之,如上所述制备来自第5轮的输出并且使其经历另外重复轮的选择,除了以下变化:与bt-c-met的温育从1小时减少至15分钟,并且珠粒捕获从20分钟减少至15分钟,bt-c-met减少至25nm(第6和7轮)或2.5nm(第8和9轮),并且在过量的非生物素酰化c-met存在下进行另外1小时的洗涤。这些变化的目的是同时选择具有可能较快的结合速率和较慢的解离速率的结合物,产生大致较低的kd。使用标准方案,利用tcon6(seqidno.30)和tcon5e86ishort(seqidno.31)引物,将第5、7和9轮输出pcr克隆到包含不依赖连接酶的克隆位点的修饰pet15载体(emdbiosciences,gibbstown,nj)(pet15-lic)中,并且在转化和iptg诱导(1mm终浓度,30℃下持续16小时)后将蛋白质表达为c-末端带his6标签的蛋白质。通过离心收获细胞并随后用补充有0.2mg/ml鸡蛋清溶解酵素(sigma-aldrich)的bugbusterht(emdbiosciences)裂解。通过离心使细菌溶解物澄清并将上清液转移至新的96深孔板。筛选抑制hgf结合于c-met的fn3域在生物化学形式中针对它们抑制hgf结合于经纯化的c-met胞外域的能力来筛选存在于大肠杆菌裂解物中的fn3域。将重组人c-metfc嵌合体(pbs中的0.5μg/ml,100μl/孔)包被于96孔白色maxisorp板(nunc)上并在4℃下温育过夜。在biotek洗板机上使用300μl/孔含0.05%吐温20的tris缓冲盐水(tbs-t,sigma-aldrich)将板洗涤两次。在摇动的情况下在室温(rt)下用startingblockt20(200μl/孔,thermofisherscientific,rockland,il)封闭测定板1小时,并且再用300μl的tbs-t洗涤两次。使用hamiltonstarplus机器人系统将fn3域裂解物稀释于startingblockt20(从1:10到1:100,000)中。在摇动的情况下在室温下将裂解物(50μl/孔)在测定板上温育1小时。在不洗涤板的情况下,在室温下将bt-hgf(startingblockt20中的1μg/ml,50μl/孔,生物素酰化)加入板中持续30min,同时伴以摇动。包含tencon27裂解物的对照孔容纳有startingblockt20或稀释的bt-hgf。然后用300μl/孔的tbs-t将板洗涤四次,并且在摇动的情况下在室温下用100μl/孔的链霉亲和素-hrp(tbs-t中的1:2000,jacksonimmunoresearch,westgrove,pa)温育30-40分钟。再用tbs-t将板洗涤四次。为形成信号,将根据制造商说明书制备的pod化学发光底物(50μl/孔,rochediagnostics,indianapolis,in)加入板中,并且在大约3分钟内,使用softmaxpro在moleculardevicesm5上读取发光。使用下列计算来测定抑制百分比:100-((rlu样品–平均rlu无bt-hgf对照)/(平均rlubt-hgf对照-平均rlu无bt-hgf对照)*100)。50%或更大的抑制百分比值被视为活性化合物(hits)。fn3域的高通量表达和纯化用hismultitraptmhp板(gehealthcare)从澄清的大肠杆菌裂解物纯化带his标签的fn3域,并在ph7.4的包含20mm磷酸钠、500mm氯化钠和250mm咪唑的缓冲液中洗脱。使用pdmultitraptmg-25板(gehealthcare)将经纯化的样品交换到pbsph7.4中以用于分析。hgf结合于c-met的抑制的ic50测定在hgf竞争测定法中进一步表征所选fn3域。利用上述测定法建立经纯化的fn3域的剂量响应曲线(起始浓度为5μm)。计算抑制百分比值。将数据作为抑制%对fn3域摩尔浓度的对数作图,并且使用graphpadprism4将数据拟合到具有可变斜率的s形剂量响应来测定ic50值。从第5轮鉴定出在1:10稀释度下显示活性且ic50值在0.5至1500nm范围内的35个独特序列。第7轮得到在1:100稀释度下具有活性且ic50值在0.16至2.9nm范围内的39个独特序列。从第9轮鉴定出66个独特序列,其中活性化合物(hits)被定义为在1:1000的稀释度下具有活性。在第9轮中观察到低至0.2nm的ic50值(表8)。所选结合c-met的fn3域对c-met-fc的亲和力测定(egfr-fc亲和力)按照实例3中对于对所选结合egfr的fn3域的亲和力测定所述的方法,测定对所选结合c-met的fn3域的亲和力,不同的是在测定法中使用c-met-fc。实例6:结合c-met并抑制hgf结合的fn3域的表征按照以上在实例2中所述的方法表达并纯化fn3域。分别按照以上在实例1和2中所述的方法进行尺寸排阻色谱法和动力学分析。表7示出了c-链、cd环、f-链和fg环的序列以及每个域的完整氨基酸序列的seqidno。表7.c环残基对应于所指示seqidno的第28-37位残基cd链残基对应于所指示seqidno的第38-43位残基f环残基对应于所指示seqidno的第65-74位残基fg链残基对应于所指示seqidno的第75-81位残基所选结合c-met的fn3域对细胞上的c-met的结合(“h441细胞结合测定”)将nci-h441细胞(目录号htb-174,americantypeculturecollection,manassas,va)以每孔20,000个细胞接种于聚d-赖氨酸包被的黑色透明底96孔板(bdbiosciences,sanjose,ca)中,并使之在37℃、5%的co2下附着过夜。在4℃下将经纯化的fn3域(50μl/孔;0至1000nm)加入两个平行测定板中的细胞中持续1小时。移除上清液,并且用facs染色缓冲液(150μl/孔,bdbiosciences,目录号554657)将细胞洗涤三次。将细胞与生物素酰化抗his抗体(以1:160稀释于facs染色缓冲液中,50μl/孔,r&dsystems,目录号bam050)在4℃下温育30分钟。用facs染色缓冲液(150μl/孔)将细胞洗涤三次,之后将孔与抗小鼠igg1-alexa488缀合抗体(以1:80稀释于facs染色缓冲液中,50μl/孔,lifetechnologies,目录号a21121)在4℃下温育30分钟。用facs染色缓冲液(150μl/孔)将细胞洗涤三次并留在facs染色缓冲液(50μl/孔)中。使用acumenex3读板机测定总荧光。将数据作为原始荧光信号对fn3域摩尔浓度的对数作图,并且使用graphpadprism4(graphpadsoftware)拟合到具有可变斜率的s形剂量-响应曲线,以计算ec50值。发现fn3域表现出一系列结合活性,并且ec50值在1.4nm与22.0nm之间,如表8中所示。hgf刺激的c-met磷酸化的抑制使用得自mesoscalediscovery(gaithersburg,md)的c-met磷酸化(tyr1349)试剂盒,测试经纯化的fn3域抑制nci-h441中hgf刺激的c-met磷酸化的能力。将细胞以20,000/孔接种于透明96孔组织培养处理板中含10%胎牛血清(fbs;lifetechnologies)的100μl/孔rpmi培养基(包含glutamax和hepes,lifetechnologies)中,并使之在37℃、5%的co2下附着过夜。将培养基完全移除,并且在无血清rpmi培养基(100μl/孔)中在37℃、5%的co2下对细胞进行饥饿处理过夜。然后为细胞补充包含20μm及更低浓度的fn3域的新鲜无血清rpmi培养基(100μl/孔),在37℃、5%的co2下持续1小时。对照仅用培养基处理。用100ng/ml重组人hgf(100μl/孔,r&dsystems目录号294-hgn)刺激细胞并在37℃、5%的co2下温育15分钟。一组对照孔保持不刺激,作为阴性对照。然后将培养基完全移除,并且根据制造商说明书,在摇动的情况下在室温下用完全裂解缓冲液(50μl/孔,mesoscalediscovery)裂解细胞10分钟。根据制造商说明书,用所提供的封闭溶液在室温下将被配置用于测量磷酸化c-met的测定板封闭1小时。然后用tris洗涤缓冲液(200μl/孔,mesoscalediscovery)将板洗涤三次。将细胞裂解物(30μl/孔)转移到测定板,并且在摇动的情况下在室温下温育1小时。然后用tris洗涤缓冲液将测定板洗涤四次,之后将冰冷检测抗体溶液(25μl/孔,mesoscalediscovery)加入每孔中,在摇动的情况下在室温下持续1小时。再用tris洗涤缓冲液将板冲洗四次。通过如下方式检测信号:加入150读取缓冲液(150μl/孔,mesoscalediscovery)并且使用制造商安装的测定专用默认设置,在成像器6000仪器(mesoscalediscovery)上读数。将数据作为电化学发光信号对fn3域摩尔浓度的对数作图,并且使用graphpadprism4将数据拟合到具有可变斜率的s形剂量响应来测定ic50值。发现fn3域以4.6nm至1415nm范围内的ic50值抑制磷酸化c-met,如表8所示。人瘤细胞生长或活力的抑制通过如下方式评估c-met依赖性细胞生长的抑制:在暴露于结合c-met的fn3域后,测量u87-mg细胞(americantypeculturecollection,目录号htb-14)的活力。将细胞以每孔8000个细胞接种于不透明的白色96孔组织培养处理板(nunc)中补充有10%的fbs的100μl/孔rpmi培养基中,并使之在37℃、5%的co2下附着过夜。接种后二十四小时,吸出培养基并为细胞补充无血清rpmi培养基。血清饥饿后二十四小时,通过加入包含结合c-met的fn3域的无血清培养基(30μl/孔)来处理细胞。将细胞在37℃、5%的co2下温育72小时。通过如下方式检测活细胞:加入100μl/孔的celltiter-试剂(promega),然后在板摇动器上混合10分钟。在设定为发光模式的spectramaxm5读板机(moleculardevices)上读取板,读取时间为0.5秒/孔。将数据作为原始发光单位(rlu)对fn3域摩尔浓度的对数作图。使用graphpadprism4将数据拟合到具有可变斜率的s形剂量响应的方程,来测定ic50值。表8报告了1nm至>1000nm范围内的ic50值。结合c-met的fn3域的特征汇总于表8中。表8.结合c-met的fn3域的热稳定性使用pbs中的差示扫描量热法评估每个fn3域的稳定性。实验的结果在表9中示出。表9.实例7.双特异性抗egfr/c-met分子的构建和表征双特异性egfr/c-met分子的构建将实例1-6中所述的结合egfr和c-met的fn3域的多种组合连接成能够结合于egfr和c-met两者的双特异性分子。另外,制备具有以seqidno:107-110示出的氨基酸序列的结合egfr的fn3域以及具有以seqidno:111-114示出的氨基酸序列的结合c-met的fn3域,并连接成双特异性分子。形成合成基因以便编码以seqidno:50-72和106(表10)所述的氨基酸序列,使得下列形式得以保持:结合egfr的fn3域,之后是肽接头,之后是结合c-met的fn3域。将聚组氨酸标签掺入到c-末端以有助于纯化。除了表10中所述的那些分子之外,这两个fn3域之间的接头还根据如表11中所列的长度、序列组成和结构而变化。可以设想到多种其他接头可用于连接此类fn3域。使双特异性egfr/c-met分子表达,并且如针对单特异性egfr或c-metfn3域所述,使用imac和凝胶过滤色谱法步骤从大肠杆菌纯化。表10.表11.接头seqidno:以氨基酸计的接头长度结构gs782无序ggggs1055无序(ggggs)47920无序(ap)2804刚性(ap)5815刚性(ap)108220刚性(ap)208340刚性a(eaaak)5aaa8429α-螺旋与单独的单特异性分子相比,双特异性egfr/c-met分子增强了效力,表明有亲合力将nci-h292细胞接种于96孔板中含10%的fbs的rpmi培养基中。24小时后,将培养基更换为无血清rpmi。血清饥饿后24小时,用如下不同浓度的fn3域处理细胞:高亲和力单特异性egfrfn3域(p54ar4-83v2),弱亲和力单特异性c-metfn3域(p114ar5p74-a5),两种单特异性egfr和c-metfn3域的混合物,或由低亲和力c-metfn3域连接到高亲和力egfrfn3域(ecb1)构成的双特异性egfr/c-met分子。将细胞用单特异性或双特异性分子处理1小时,然后在37℃、5%的co2下用egf、hgf或egf与hgf的组合刺激15分钟。用msd裂解缓冲液裂解细胞,并且如上所述根据制造商说明书,使用适当msd测定板评估细胞信号传导。低亲和力c-metfn3域以610nm的ic50抑制c-met的磷酸化(图4)。如预期,egfrfn3域不能抑制c-met磷酸化,并且单特异性分子的混合物似乎与单独的c-metfn3域相同。然而,双特异性egfr/c-met分子以1nm的ic50抑制c-met的磷酸化(图4),相对于单独的c-met单特异性分子而言在改善效力方面提供了2-log的改变。在具有可变c-met和egfr密度和比率的多种细胞类型中评价了双特异性egfr/c-met分子通过亲合力效应增强对c-met和/或egfr磷酸化的抑制的潜能(图5)。将nci-h292、nci-h441或nci-h596细胞接种于96孔板中含10%的fbs的rpmi培养基中。24小时后,将培养基更换为无血清rpmi。血清饥饿后24小时,用不同浓度的单特异性结合egfr的fn3域、单特异性c-metfn3域或双特异性egfr/c-met分子(ecb5,由p53a1r5-17v2和p114ar7p94-a3构成)处理细胞。将细胞用单特异性或双特异性分子处理1小时,然后在37℃、5%的co2下用egf、hgf或egf与hgf的组合刺激15分钟。用msd裂解缓冲液裂解细胞,并且如上所述根据制造商说明书,使用适当msd测定板评估细胞信号传导。图5(a-c)示出了在三种不同细胞系中与双特异性egfr/cmet分子相比使用单特异性结合egfr的fn3域时egfr的抑制。为了在egfr磷酸化测定法中评估亲合力,将中等亲和力结合egfr的fn3域(1.9nm)(p53a1r5-17v2)与双特异性egfr/c-met分子(0.4nm)(p114ar7p94-a3)相比较,该双特异性egfr/c-met分子包含连接到高亲和力结合c-met的fn3域的相同结合egfr的fn3域。在nci-h292和h596细胞中,单特异性和双特异性分子对egfr的磷酸化的抑制是相当的(图5a和5b),可能是因为这些细胞系具有egfr与c-met受体的高比率。为了测试该理论,在显示c-met受体比egfr更多的nci-h441细胞中评价egfr磷酸化的抑制。与单特异性结合egfr的fn3域相比,用双特异性egfr/c-met分子处理nci-h441细胞将抑制egfr磷酸化的ic50降低了30倍(图5c)。使用具有对egfr(0.26nm)的高亲和力及对c-met(10.1nm)的中等亲和力的分子,在c-met磷酸化测定法中评价了双特异性egfr/c-met分子增强效力的可能性。在nci-h292和nci-h596细胞两者中,与单特异性结合c-met的fn3域相比,双特异性分子分别将c-met的磷酸化的抑制增强134倍和1012倍(图5d和5e)。已证实,双特异性egfr/c-met分子增强抑制egfr和c-met磷酸化的效力转化为增强抑制信号传导和增殖。对于这些实验,将结合fn3egfr和结合c-met的fn3域的混合物与双特异性egfr/c-met分子相比较。如表12和13中所述,当用双特异性egfr/c-met分子处理细胞时,与单特异性结合物的混合物相比,erk磷酸化(表12)和nci-h292细胞增殖(表13)的ic50值减小。相对于两种单特异性egfr和c-metfn3域的混合物,双特异性egfr/c-met分子对erk磷酸化抑制的ic50低143倍,在该测定法中显示了亲合力对分子效力的效应。在表12中,单特异性结合egfr和c-met的fn3域不会完全抑制活性,因此所示ic50值应视为下限。使用作为混合物或以双特异性形式连接的结合egfr和c-met的fn3域的不同组合,完成增殖测定。相对于单特异性亲本结合egfr或c-met的fn3域的混合物,双特异性egfr/c-met分子抑制增殖的ic50低34-236倍。这证实,在受体水平观察到的亲合力效应(图4和图5)转化为在抑制细胞信号传导(表12)和细胞增殖(表13)方面的改善。表12.表13.体内移植瘤:pk/pd为了测定体内单特异性和双特异性fn3域分子的疗效,对瘤细胞进行工程改造以分泌人hgf(鼠hgf不结合于人c-met)。使用慢病毒感染(表达人hgf的慢病毒dna载体(登录号x16322)和得自genecopoeia的慢病毒包装试剂盒)在nci-h292细胞中稳定表达人hgf。感染后,用4μg/ml嘌呤霉素(invitrogen)选择表达hgf的细胞。使用得自mesoscalediscovery的测定板在合并细胞的调理培养基中检测人hgf蛋白质。在每只动物的背侧上对scidbeige小鼠皮下接种表达人hgf的nci-h292细胞(200μl体积cultrex(trevigen)中的2.0×106个细胞)。每周进行两次瘤测量,直到瘤体积在150-250mm3之间的范围内。然后给予小鼠单一腹腔剂量的双特异性egfr/c-met分子(连接到白蛋白结合域以延长半衰期)或pbs媒介物。在给药后6h或72h,取出瘤并立即冷冻于液氮中。经由心脏穿刺将血液样品收集到含3.8%柠檬酸盐的蛋白酶抑制剂中。收集后立即将血液样品离心,并且将所得血浆转移到样品管中并保存于-80℃下。将瘤称重,切成小块,并在包含加有halt蛋白酶/磷酸酶抑制剂(pierce)、50mm氟化钠(sigma)、2mm活化原钒酸钠(sigma)和1mm的pmsf(mesoscalediscovery)的ripa缓冲液的裂解基质a试管(lma)中裂解。将裂解物从lma基质移除并且离心除去不溶蛋白。用bca蛋白测定法定量可溶瘤蛋白,并且在瘤裂解缓冲液中稀释到等同蛋白水平。使用得自mesoscalediscovery的测定板(根据制造商方案并如上文所述)测量磷酸化c-met、egfr和erk。图6示出了实验的结果。在6h和72h时,每个双特异性egfr/c-met分子显著降低了磷酸化c-met、egfr和erk的水平。图6中所示的数据显示同时抑制c-met和egfr两者的重要性以及双特异性egfr/c-met分子对每个受体的亲和力在抑制下游erk方面起到的作用。在6h和72h时,与包含中等亲和力结合egfr的fn3域的分子(p53a1r5-17v2;图中以“17”示出,kd=1.9nm)相比,包含高亲和力结合egfr的fn3域的分子(p54ar4-83v2;图中以“8”示出,kd=0.26nm)将egfr的磷酸化抑制到更大程度。在6小时的时间点,所测试的所有四种双特异性分子完全抑制了erk的磷酸化,而与亲和力无关。在72小时的时间点,与中等亲和力结合c-met的fn3域(p114ar5p74-a5;图中以“a5”示出;kd=10.1nm;图6)相比,包含紧密亲和力c-met结合域的分子(p114ar7p94-a3;图中以“a3”示出,kd=0.4nm)显著抑制了erk的磷酸化。在血液和瘤中给药后6和72小时,测量每个双特异性egfr/c-met分子的浓度(图7)。有趣的是,在6小时处,相对于其他分子而言,具有中等亲和力egfr结合域(p53a1r5-17v2;kd=1.9nm)但具有高亲和力结合c-met的fn3域(p114ar7p94-a3;kd=0.4nm)的双特异性分子具有明显更多的瘤积聚,但到72小时该差异减小。可假设瘤之外的细胞具有更低水平的egfr和c-met表面表达,因此与更高亲和力结合egfr的fn3域相比,中等亲和力egfr分子不紧密地结合于正常组织。因此,有更多游离的中等亲和力结合egfr的fn3域可供结合于瘤中。因此,鉴定对每个受体的适当亲和力可允许鉴定具有降低的系统性毒性和增加的瘤积聚的治疗剂。采用双特异性egfr/c-met分子的瘤疗效研究在每只动物的背侧对scidbeige小鼠皮下接种表达人hgf的nci-h292细胞(200μlcultrex(trevigen)中的2.0×106个细胞)。植入后一周,小鼠被分为具有相等瘤体积的组(平均瘤体积=77.9+/-1.7mm3)。用双特异性分子每周给予小鼠三次,并每周记录瘤体积两次。使用具有对c-met和egfr的可变亲和力的四种不同双特异性分子观察瘤生长抑制(tgi)。图8示出了抑制c-met和egfr两者的有益效果,因为当结合c-met的fn3域为中等亲和力时,与中等亲和力结合egfr的fn3域相比,在用包含高亲和力结合egfr的fn3域的分子治疗的小鼠中观察到瘤生长延迟(空心与实心三角形,p54ar4-83v2-p114ar5p74-a5与p53a1r5-17-p114ar5p74-a5相比)。此外,数据显示出具有高亲和力结合c-met的fn3域的重要性,因为包含高或中等亲和力结合egfr的fn3域但包含高亲和力结合c-met的fn3域的双特异性分子显示出最大疗效(灰色和黑色点线,p54ar4-83v2-p114ar7p94-a3和p53a1r5-17v2-p114ar7p94-a3)双特异性分子及egfr和c-met的其他抑制剂的疗效在scid/beige小鼠的皮下h292-hgf人肺癌移植瘤模型的治疗中评价了双特异性egfr/c-met分子(ecb38)及小分子抑制剂克唑替尼(c-met抑制剂)和厄洛替尼(egfr抑制剂)、西妥昔单抗(抗egfr抗体)(各作为单一药剂)以及克唑替尼与厄洛替尼联用的体内疗效。在37℃下在空气的5%的co2气氛中,在补充有胎牛血清(10%v/v)和l-谷氨酰胺(2mm)的rpmi1640培养基中体外维持h292-hgf细胞。每周通过胰蛋白酶-edta处理将细胞常规地继代培养两次。收获在指数生长期生长的细胞并且计数以进行瘤接种。每只小鼠在右侧区皮下接种在0.1ml的含用于瘤发展的cultrex(1:1)的pbs中的h292-hgf瘤细胞(2×106)。当平均瘤尺寸达到139mm3时开始处理。在下列实验设计表中示出了每个研究组中的测试制品施用和动物数目。瘤细胞接种的日期表示为第0天。表14示出了处理组。表14.n:动物数目;p.o.:口服;i.p.:腹膜内注射;3次/周:在一周的第1、3和5天给药。qd:每天一次;q4d:每四天一次;克唑替尼与厄洛替尼联用的时间间隔为0.5小时;基于体重调整给药量(10l/g);a:分组后第14天不给药。在开始处理之前,将所有动物称重并测量瘤体积。由于瘤体积可影响任何给定处理的有效性,使用基于它们的瘤体积的随机化区组设计,将小鼠分配到各组。这确保了在基线处所有组是相当的。随机化区组设计用于将实验动物分配到各组。首先,根据其初始瘤体积将实验动物分成均匀区组。其次,在每个区组内,进行实验动物对处理的随机化。使用随机化区组设计分配实验动物确保了每只动物有相同概率被分配到给定处理,从而减少系统误差。在常规监测时,检查动物瘤生长和处理对正常行为的任何效应,诸如可动性、食物和水消耗量的目测、体重增加/减轻(每周测量两次体重)、眼光/毛发暗淡和任何其他异常效应。终点是瘤生长是否可被延迟或荷瘤小鼠是否可被治愈。每周使用卡尺在两个维度上测量瘤尺寸两次,并且使用下式表示以mm3计的体积:v=0.5a×b2,其中a和b分别是瘤的长直径和短直径。然后使用瘤尺寸计算t-c值和t/c值两者。计算t-c,其中t是处理组的平均瘤尺寸达到1000mm3所需的时间(以天计),并且c是对照组的平均瘤尺寸达到相同尺寸的时间(以天计)。t/c值(以百分比计)是抗瘤疗效的指示;t和c分别是在给定天处理组和对照组的平均瘤体积。瘤完全消退(cr)被定义为瘤减小到低于触诊的极限(62.5mm3)。瘤部分消退(pr)被定义为瘤从初始瘤体积减小。需要在3次或更多次连续瘤测量中cr或pr的最短持续时间,cp或pr被视为持久的。对体重减轻超过20%或该组的平均瘤尺寸超过2000mm3的动物施行安乐死。在最后一剂量后观察两周之后终止研究。在每个时间点提供每组的瘤体积的汇总统计,包括平均值和平均值的标准误差(sem),并示于表15中。使用单因素方差分析,之后使用games-howell(未假定等方差)进行个体比较,以此来评价各组间瘤体积差异的统计分析。所有数据使用spss18.0分析。p<0.05被视为统计意义上的显著性。表15在瘤接种后第23天,媒介物处理组(第1组)的平均瘤尺寸达到1,758mm3。用25mg/kg剂量水平的双特异性egfr/c-met分子进行处理(第2组)导致所有小鼠中瘤完全消退(cr),这在>3次连续瘤测量中是持久的(在第23天,与媒介物组相比,平均tv=23mm3,t/c值=1%,p=0.004)。用50mg/kg剂量水平的作为单一药剂的克唑替尼进行处理(第3组)未显示抗瘤活性;在第23天,平均瘤尺寸为2,102mm3(与媒介物组相比,t/c值=120%,p=0.944)。用50mg/kg给药水平的作为单一药剂的厄洛替尼进行治疗(第4组)显示出微量抗瘤活性,但与媒介物组相比,未发现显著的差异;在第23天,平均瘤尺寸为1,122mm3(与媒介物组相比,t/c值=64%,p=0.429),与媒介物组相比,在瘤尺寸为1,000mm3时,瘤生长延迟4天。克唑替尼(50mg/kg,第5组)与厄洛替尼(50mg/kg,第5组)的联用显示出显著的抗瘤活性;在第23天,平均瘤尺寸为265mm3(t/c=15%;p=0.008),与媒介物组相比,在瘤尺寸为1,000mm3时,瘤生长延迟17天。1mg/小鼠给药水平的作为单一药剂的西妥昔单抗(第6组)显示出显著的抗瘤活性;在第23天,平均瘤尺寸为485mm3(t/c=28%;p=0.018),与媒介物组相比,在瘤尺寸为1,000mm3时,瘤生长延迟17天。图15和表16示出了各种疗法的抗瘤活性。表16在媒介物组中观察到中等到严重的体重减轻,这可能是由于增加的瘤负荷;到第23天,3只小鼠死亡,并且当bwl>20%时,对1只小鼠施行安乐死。在第2组中观察到双特异性egfr/c-met分子的轻微毒性;在处理期过程中当bwl>20%时,对3只小鼠施行安乐死。在2周观察期过程中,当撤去处理时,体重逐渐恢复。与媒介物组相比,在克唑替尼或厄洛替尼单一疗法组中观察到更严重的体重减轻,暗示处理相关的毒性。在给药期过程中大体能耐受克唑替尼与厄洛替尼联用,但在研究结束时观察到严重的体重减轻,这可能是由于在非处理期过程中恢复了快速瘤生长。在该研究中西妥昔单抗的单一疗法能被良好耐受;仅在研究结束时观察到体重减轻,这是由于瘤生长恢复。总之,25mg/kg的双特异性egfr/c-met分子(3次/周×3周)在scid/beige小鼠中的h292-hgf人肺癌移植瘤模型中产生了完全响应。该处理在10只小鼠中有7只耐受,并且导致10只小鼠中有3只严重体重减轻。图9示出了处理后各时间点期间各种疗法对瘤尺寸的影响。实例8:双特异性egfr/c-met分子的半衰期延长已描述了用于减少肾过滤并从而延长蛋白质的血清半衰期的多种方法,包括用聚乙二醇(peg)或其他聚合物修饰、结合于白蛋白、与结合于白蛋白或其他血清蛋白的蛋白域融合、与白蛋白的基因融合、与iggfc域的融合,以及与非结构化长氨基酸序列的融合。用peg修饰双特异性egfr/c-met分子,以便通过在分子c-末端掺入游离半胱氨酸来增加流体力学半径。最常见的是,半胱氨酸残基的游离巯基用于使用标准方法连接用马来酰亚胺或碘乙酰胺基团官能化的peg分子。各种形式的peg可用于修饰蛋白,包括1000、2000、5000、10,000、20,000或40,000kda的直链peg。这些分子量的支链peg分子也可用于修饰。在一些情况下,peg基团也可通过双特异性egfr/c-met分子中的伯胺连接。除聚乙二醇化之外,双特异性egfr/c-met分子的半衰期还可通过将这些蛋白制备成与天然存在的3-螺旋束血清白蛋白结合域(abd)或共有白蛋白结合域(abdcon)的融合分子来延长。这些蛋白域经由表12中所述的任何接头连接到结合c-met的fn3域的c-末端。abd或abdcon域也可在一级序列中设置在结合egfr的fn3域与结合c-met的fn3域之间。实例9:所选双特异性egfr/c-met分子的表征表征了所选双特异性egfr/c-met分子对egfr和c-met两者的亲和力、抑制egfr和c-met自磷酸化的能力以及对hgf细胞增殖的效应。还根据实例3中所述的方案,使用proteon仪器(biorad),通过表面等离子共振方法评价了双特异性egfr/c-met分子对重组egfr和/或c-met胞外域的结合亲和力。表征的结果在表17中示出。表17.实例10.双特异性egfr/cmet抗体的生成若干单特异性egfr和c-met抗体被表达为在其fc区中具有fc替换k409r或f405l(根据eu索引进行编号)的igg1,κ。单特异性抗体在两种cho细胞系中表达,一种细胞系具有降低的岩藻糖基化能力,从而产生在抗体多糖链中具有1-15%岩藻糖含量的抗体。单特异性抗体使用proteina柱(hitrapmabselectsure柱)以标准方法纯化。洗脱后,在d-pbs,ph7.2中透析合并液。通过在体外fab臂交换(如wo2011/131746中所述)中将单特异性egfrmab与单特异性c-metmab组合,来生成双特异性egfr/c-met抗体。简而言之,将pbs,ph7-7.4中的约1-20mg/ml的1:1摩尔比的每种抗体与75mm2-巯基乙醇胺(2-mea)混合在一起并且在25-37℃下温育2-6h,然后使用标准方法经由透析、透滤、切向流过滤和/或旋转细胞过滤来移除2-mea。在体外fab臂交换中将若干单特异性抗egfr抗体和抗c-met抗体组合于基质中以生成双特异性抗体,随后对该双特异性抗体进行进一步表征。使用如下测定法以四步策略对所生成的双特异性抗体进行评级:步骤1:facs测定法中对nci-h441、nci-h1975和a549细胞的结合。步骤2:a549细胞中pmet磷酸化的抑制。步骤3:nci-h1975、kp4和nci-h441细胞中增殖的抑制。步骤4:a549和snu-5细胞中egfr磷酸化的抑制。值得注意的是,双特异性抗体中未保留亲本抗体的特性。例如,双特异性抗体中存在某些egfr结合臂导致抑制丧失或减弱,或c-met磷酸化增强。基于表征研究来选择所选的对。生成了单特异性二价抗egfr抗体e1-k409r,其包含抗egfr抗体2f8的vh和vl区及具有k409r替换的igg1恒定区,抗egfr抗体2f8具有seqidno:189的vh和seqidno:190的vl(抗体2f8描述于国际专利公布wo2002/100348)。生成了单特异性二价抗egfr抗体e1-f405l,其包含抗egfr抗体2f8的vh和vl区及具有f405l替换的igg1恒定区,抗egfr抗体2f8具有seqidno:189的vh和seqidno:190的vl(抗体2f8描述于国际专利公布wo2002/100348)。生成了单特异性二价抗egfr抗体e2-k409r,其包含抗egfr抗体018的vh和vl区及具有k409r替换的igg1恒定区,抗egfr抗体018具有seqidno:191的vh和seqidno:192的vl(抗体018描述于国际专利公布wo2009/030239)。生成了单特异性二价抗egfr抗体e2-f405l,其包含抗egfr抗体018的vh和vl区及具有f405l替换的igg1恒定区,抗egfr抗体018具有seqidno:191的vh和seqidno:192的vl(抗体018描述于国际专利公布wo2009/030239)。生成了单特异性二价抗c-met抗体m1-k409r,其包含抗c-met抗体069的vh和vl区及具有k409r替换的igg1恒定区,抗c-met抗体069具有seqidno:193的vh和seqidno:194的vl(抗体069描述于wo2011/110642)。生成了单特异性二价抗c-met抗体m1-f405l,其包含抗c-met抗体069的vh和vl区及具有f405l替换的igg1恒定区,抗c-met抗体069具有seqidno:193的vh和seqidno:194的vl(抗体069描述于wo2011/110642)。生成了单特异性抗c-met抗体m2-k409r,其包含抗c-met抗体058的vh和vl区及具有k409r替换的igg1恒定区,抗c-met抗体058具有seqidno:195的vh和seqidno:196的vl(抗体058描述于wo2011/110642)。生成了单特异性抗c-met抗体m2-f405l,其包含抗c-met抗体058的vh和vl区及具有f405l替换的igg1恒定区,抗c-met抗体058具有seqidno:195的vh和seqidno:196的vl(抗体058描述于wo2011/110642)。以下示出了抗体的vh、vl、hc和lc序列:>seqidno:189egfrmabe1vhqvqlvesgggvvqpgrslrlscaasgftfstygmhwvrqapgkglewvaviwddgsykyygdsvkgrftisrdnskntlylqmnslraedtavyycardgitmvrgvmkdyfdywgqgtlvtvss>seqidno:190egfrmabe1vlaiqltqspsslsasvgdrvtitcrasqdissalvwyqqkpgkapklliydasslesgvpsrfsgsesgtdftltisslqpedfatyycqqfnsypltfgggtkveik>seqidno:191egfrmabe2vhevqlvesggglvqpggslrlscaasgftfssywmnwvrqapgkglewvanikkdgsekyyvdsvkgrftisrdnaknslylqmnslraedtavyycardlgwgwgwyfdlwgrgtlvtvss>seqidno:192egfrmabe2vleivltqspatlslspgeratlscrasqsvssylawyqqkpgqaprlliydasnratgiparfsgsgsgtdftltisslepedfavyycqqrsnwpptfgqgtkveik>seqidno:193cmetmabm1vhqvqlvqsgaevkkpgasvkvscetsgytftsygiswvrqapghglewmgwisayngytnyaqklqgrvtmttdtststaymelrslrsddtavyycardlrgtnyfdywgqgtlvtvss>seqidno:194cmetmabm1vldiqmtqspssvsasvgdrvtitcrasqgisnwlawfqhkpgkapklliyaassllsgvpsrfsgsgsgtdftltisslqpedfatyycqqansfp-itfgqgtrleik>seqidno:195cmetmabm2vhevqlvesggglvkpggslklscaasgftfsdyymywvrqtpekrlewvatisddgsytyypdsvkgrftisrdnaknnlylqmsslksedtamyycareglyyygsgsyynqdywgqgtlvtvss>seqidno:196cmetmabm2vlqltqspsslsasvgdrvtitcrasqglssalawyrqkpgkapklliydasslesgvpsrfsgsgsgtdftltisslqpedfatyycqqftsypqitfgqgtrleik>seqidno:199em1-mabh1(抗egfr,405l)qvqlvesgggvvqpgrslrlscaasgftfstygmhwvrqapgkglewvaviwddgsykyygdsvkgrftisrdnskntlylqmnslraedtavyycardgitmvrgvmkdyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:200em-1mabl1aiqltqspsslsasvgdrvtitcrasqdissalvwyqqkpgkapklliydasslesgvpsrfsgsesgtdftltisslqpedfatyycqqfnsypltfgggtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:201em-1mabh2(k409r,抗cmet)qvqlvqsgaevkkpgasvkvscetsgytftsygiswvrqapghglewmgwisayngytnyaqklqgrvtmttdtststaymelrslrsddtavyycardlrgtnyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:202em-1mabl2diqmtqspssvsasvgdrvtitcrasqgisnwlawfqhkpgkapklliyaassllsgvpsrfsgsgsgtdftltisslqpedfatyycqqansfpitfgqgtrleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:234e2mabhc1(egfr-f405l)evqlvesggglvqpggslrlscaasgftfssywmnwvrqapgkglewvanikkdgsekyyvdsvkgrftisrdnaknslylqmnslraedtavyycardlgwgwgwyfdlwgrgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:235e2mablc1(egfr)eivltqspatlslspgeratlscrasqsvssylawyqqkpgqaprlliydasnratgiparfsgsgsgtdftltisslepedfavyycqqrsnwpptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:236e2mabhc2(c-met-k409r)evqlvesggglvkpggslklscaasgftfsdyymywvrqtpekrlewvatisddgsytyypdsvkgrftisrdnaknnlylqmsslksedtamyycareglyyygsgsyynqdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:237e2mablc2(cmet)qltqspsslsasvgdrvtitcrasqglssalawyrqkpgkapklliydasslesgvpsrfsgsgsgtdftltisslqpedfatyycqqftsypqitfgqgtrleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec将所生成的单特异性抗egfr和c-met抗体混合以用于基质中的体外fab臂交换,并且在各种测定法中进行表征。双特异性抗体em1-mab包含mabe1-f405l的egfr结合臂和mabm1-k409r的c-met结合臂。双特异性抗体em2-mab包含mabe2-f405l的egfr结合臂和mabm2-k409r的c-met结合臂。双特异性抗体em3-mab包含mabe1-k409r的egfr结合臂和mabm1-f405l的c-met结合臂。双特异性抗体em4-mab包含mabe2-k409r的egfr结合臂和mabm2-f405l的c-met结合臂。em1-mab和em3-mab具有相当的特性。双特异性em1-mab培养于具有降低的糖蛋白岩藻糖基化能力的cho细胞系中,从而具有约1-15%的岩藻糖基含量。核心岩藻糖从连接到fc区的二天线复合物型寡糖的移除经由改善的fcγriiia结合来显著增强抗体的adcc,而不改变抗原结合或cdc活性。此类mab可使用据报道可引起具有二天线复合物型fc寡糖的相对较高去岩藻糖基化治疗抗体的成功表达的不同方法来实现,并且在上文有所描述。实例11.双特异性egfr/c-met抗体的纯化在体外fab臂交换后使用标准方法采用疏水相互作用色谱法进一步纯化双特异性em1-mab,以使残余亲本c-met和egfr抗体最小化。实例12.双特异性egfr/c-met抗体的表征在各种测定法中测试了egfr/c-met双特异性抗体em1-mab的特性,包括egf刺激的egfr磷酸化、hgf刺激的c-met磷酸化、erk1/2磷酸化、akt磷酸化的抑制,配体结合和细胞活力的抑制。将em1-mab的特性与对照单价egfr或c-met结合抗体比较,并与已知的egfr抑制剂诸如厄洛替尼(cas183321-74-6;酪氨酸激酶抑制剂)和西妥昔单抗(cas205923-56-4)比较。因为em-1mab抗体的亲本抗体是二价的,将对照单价egfr和c-met抗体以与结合于无关/不相关抗原的fab臂组合的双特异性形式生成,以便相比于相应对照单价分子的混合物来准确比较双特异性em-1mab的协同作用和亲合力。为了生成对照单价egfr和c-met抗体,生成了单特异性抗hivgp120抗体gp120-k409r,其包含seqidno:198的重链和seqidno:209的轻链。生成了单特异性抗hivgp120抗体gp120-f405l,其包含seqidno:197的重链和seqidno:209的轻链。通过e1-f405l与gp120-k409r之间的体外fab臂交换,生成了对照单价抗egfrmabe1-f405l-gp120-k409r,并且通过m1-k409r与gp120-f405l之间的体外fab交换,生成了对照单价抗cmetmabm1-k409r-gp120-f405l,并且按照先前所述进行纯化。在双特异性抗体的表征中使用了下列细胞系:nci-h292(americantypeculturecollection(atcc),目录号crl-1848)、nci-h1975(atcc目录号crl-5908)、skmes-1(atcc目录号htb-58)、a431(atcc目录号crl-1555)、nci-h441(atcc目录号htb-174)、nci-h3255(dctd瘤/细胞系储库,nci,frederick,nci-navymedicaloncologycellline增刊,jcellbiochemsuppl24,1996;tracys.,cancerres64:7241-4,2004;shimamurat.,cancerres65:6401-8,2005)和hcc-827(atcc目录号crl-2868)。nci-h292和skmes-1细胞表达野生型egfr和野生型c-met两者。nci-3255表达突变体l858regfr并且表现出egfr和c-met扩增。h1975表达突变体l858r/t790megfr和野生型c-met。hcc-827表达δ(e746,a750)egfr并且表现出egfr扩增。细胞系nci-h292、nci-h975、nci-h441和nci-h3255在本说明书中可互换地分别称为h292、h975、h441和h3255。双特异性egfr/cmet抗体对细胞上的egfr和c-met的结合(a431细胞结合测定)使用实例3(“a431细胞结合测定”)和实例6(“h441细胞结合测定”)中所述的方案测试了双特异性egfr/c-met抗体em1-mab对细胞上的egfr和c-met的结合。单价指向egfre1-f405l-gp120-k409r的西妥昔单抗和对照抗体用作a431细胞的对照。西妥昔单抗具有5.0nm的ec50值。表18示出了结合的ec50值。em1-mab表现出在结合方面与二价单特异性亲本对照抗体相比1.9倍(a431细胞)和2.3倍(h441细胞)的下降。西妥昔单抗与二价亲本对照抗体相当。由于egfr和c-met的单价结合,em1-mab表现出比亲本mab的值更高的ec50结合值。em1-mab具有与单臂e1/惰性臂和e2/惰性臂双特异性单价mab相似的结合ec50值。表18.配体结合于受体的抑制在elisa测定法中测试了双特异性抗体阻碍egf结合于egfr胞外域及hgf结合于c-met胞外域的能力。将重组人egfr-fc(r&dsystems,目录号:344-er-050)或人hgf(r&dsystems,目录号:294-hgn-025/cf)包被于msdhighbind平板(mesoscalediscovery,gaithersburg,md)在室温下持续2小时。将msd封闭剂a缓冲液(mesoscalediscovery,gaithersburg,md)加入每个孔并在室温下温育2小时。用0.1mhepes缓冲液(ph7.4)将平板洗涤三次,然后加入荧光染料标记(msd)egf或生物素酰化hgf蛋白与不同浓度抗体的混合物。将钌标记egf蛋白与从1nm递增到4μm浓度的不同抗体在室温下温育30分钟。在室温下在轻轻摇动的情况下温育2小时后,用0.1mhepes缓冲液(ph7.4)将平板洗涤3次。将msd读取缓冲液t稀释并分配,并且使用sector成像器6000分析信号。如egf/egfr抑制测定法进行hgf抑制测定法,不同的是将10nm的生物素酰化hgf与从1nm递增到2μm浓度的不同抗体在室温下温育30分钟。em1-mab在50nm的egf存在下以10.7nm±1.2的ic50值抑制egf结合于egfr,并且在80nmegf存在下以10.6±1.5nm的ic50值抑制egf结合于egfr。亲本二价抗体在50nm的egf存在下以0.14±1.5nm的ic50值抑制egf结合于egfr,并且在80nmegf存在下以1.7±1.4nm的ic50值抑制egf结合于egfr。em1mab具有对egf结合于egfr胞外域的较弱抑制,这是由于与亲本二价mab相比em1mab的单价结合。em1-mab以29.9±1.5nm的ic50值抑制hgf结合于c-met。亲本二价抗体以14.8±1.6nm的ic50抑制hgf结合于c-met。em1mab具有对hgf结合于cmet胞外域的较弱抑制,这是由于与亲本二价mab相比em1-mab的单价结合。egf刺激的egfr磷酸化和hgf刺激的c-met磷酸化的抑制对抗体进行测试以测定抑制egfr和c-met磷酸化的ic50值。如实例2(“egf刺激的egfr磷酸化的抑制”)和实例6(“hgf刺激的c-met磷酸化的抑制”)中所述以不同的抗体浓度(终浓度0.035-700nm)评估egf刺激的egfr磷酸化和hgf刺激的c-met磷酸化的抑制。在一些实验中,将egf和hgf两者加入细胞中,因此可使用相同的细胞裂解物检测egfr和c-met磷酸化两者。单价指向egfr的对照抗egfrmabe1-f405l-gp120-k409r以及具有低岩藻糖含量的亲本二价抗egfr抗体用作对照抗体。表19示出了测定法的ic50值。表19.*抗体具有低岩藻糖含量与单价抗体的混合物相比em1-mab增强了对perk和pakt的抑制(mabperk测定法)(mabpakt测定法)通过评估mab对perk和pakt下游信号传导的效应,评价了双特异性egfr/c-met抗体增强效力的潜能。对于这些实验,将单价对照egfr与单价对照c-met抗体的混合物与双特异性em1-mab比较。将细胞接种于透明96孔组织培养处理板(nunc)中含glutamax和25mmhepes(invitrogen)且补充有1mm丙酮酸钠(gibco)、0.1mmneaa(gibco)、10%热灭活胎牛血清(gibco)和7.5ng/mlhgf(r&dsystems目录号294-hgn)的100μl/孔rpmi培养基中,并使之在37℃下在潮湿5%的co2气氛中附着过夜。不对细胞进行血清饥饿处理。用不同浓度(终浓度0.11-700nm)的单价对照抗体或em1-mab处理细胞30分钟(perk测定法)或1小时(pakt测定法)。使用下列试剂盒并根据来自mesoscalediscovery的制造商说明书,评估细胞的perk或pakt水平:磷酸化erk1/2(thr202/tyr204;thr185/tyr187)测定法全细胞裂解物试剂盒(目录号k151dwd)、磷酸化akt(ser473)测定法全细胞裂解物试剂盒(目录号k151cad)、磷酸化akt(thr308)测定法全细胞裂解物试剂盒(目录号k151dyd)。对于perk测定法,将细胞裂解,并将全细胞裂解物加入包被有抗磷酸化erk1/2抗体(识别在残基thr202和tyr204处磷酸化的erk1以及在残基thr185和tyr187处磷酸化的erk2)的平板上,并且用与msdsulfo-tagtm试剂缀合的抗全erk1/2抗体检测磷酸化erk1/2。对于paktser473测定法,捕获抗体是抗全akt抗体并且检测抗体是与msdsulfo-tagtm试剂缀合的抗paktser473抗体。对于paktthr308测定法,捕获抗体是抗全akt抗体并且检测抗体是与msdsulfo-tagtm试剂缀合的抗paktthr308抗体。使用制造商安装的测定专用默认设置,在成像器6000仪器(mesoscalediscovery)上读取平板。将数据作为电化学发光信号对抗体浓度的对数作图,并且使用graphpadprism5软件将数据拟合到具有可变斜率的s形剂量响应来测定ic50值。nci-h292、h1975和skmes-1细胞系用于这些测定法。相对于两种单价对照抗体的混合物而言,双特异性em1-mab抑制erk磷酸化的ic50低约14-63倍,具体取决于所测试的细胞系(表20)。em1-mab与两种单价对照抗体的混合物相比改善的效力表明由于em1-mab对这些细胞系的增强结合而产生了协作或亲合力效应。与两种单价对照抗体的混合物相比,nci-h1975细胞系中ser475(pakts475)和thr308(paktt308)akt磷酸化的抑制的ic50分别低约75倍和122倍(表21)。em1-mab与两种单价对照抗体的混合物相比改善的效力表明由于em1-mab对这些细胞系的增强结合而产生了协作或亲合力效应。因此,em1-mab的双特异性质导致对下游信号传导效应子的增强效应。表20.表21.抗体对人瘤细胞生长或活力的抑制通过如下方式评估c-met依赖性细胞生长的抑制:在暴露于双特异性em1-mab后,测量各种瘤细胞的活力。nci-h292、skmes-1、nci-h1975和nci-h3255细胞用于研究。以标准2d和低附着形式培养细胞。厄洛替尼和西妥昔单抗用作对照。表22汇总了该测定法的ic50值。抗体对人瘤细胞生长或活力的抑制–标准2d形式通过如下方式以两种形式评估细胞生长的抑制:在暴露于抗体后,测量nci-h292和nci-h1975的活力。对于标准2d形式,将细胞接种于不透明的白色96孔组织培养处理板(perkinelmer)中含glutamax和25mmhepes(invitrogen)且补充有1mm丙酮酸钠(gibco)、0.1mmneaa(gibco)和10%热灭活胎牛血清(gibco)的rpmi培养基中,并使之在37℃、5%的co2下附着过夜。将细胞用不同浓度的抗体(终浓度0.035–700nm)以及hgf(7.5ng/ml,r&dsystems目录号294-hgf)处理,然后在37℃、5%的co2下温育72小时。一些孔不用hgf或抗体处理,作为对照。使用celltiter-试剂(promega)检测活细胞,并且如实例3在“人瘤细胞生长的抑制(nci-h292生长和nci-h322生长测定法)”中所述分析数据。抗体对人瘤细胞生长或活力的抑制–低附着形式为了评估低附着条件下的存活率,将细胞接种于超低附着性96孔板(corningcostar)中含glutamax和25mmhepes且补充有1mm丙酮酸钠(gibco)、0.1mmneaa(gibco)和10%热灭活胎牛血清(gibco)的50μl/孔rpmi培养基(invitrogen)中,并使之在37℃、5%的co2下附着过夜。将细胞用不同浓度的抗体(终浓度0.035–700nm)以及hgf(7.5ng/ml,r&dsystems目录号294-hgn)处理,然后在37℃、5%的co2下温育72小时。一些孔不用hgf或抗体处理,作为对照。使用celltiter-试剂(promega)检测活细胞,并且如以上在实例3的“人瘤细胞生长的抑制(nci-h292生长和nci-h322生长测定法)”中所述分析数据,不同的是在读取发光之前将裂解物转移到不透明的白色96孔组织培养处理板(perkinelmer)中。在标准2d培养中,em1-mab以31nm的ic50抑制nci-h292生长,并且在低附着条件下,以0.64nm的ic50抑制nci-h292生长。em-1mab在标准2d和低附着培养中分别以>700nm和0.8-1.35nm的ic50抑制nci-h1975细胞生长。在表达野生型egfr和cmet两者的nci-h292细胞中,与西妥昔单抗相比,em1-mab在标准2d培养条件下具有超过22倍改善的效力,并且在低附着培养条件下具有约330倍改善的效力。在表达l858r、t790megfr突变体和野生型cmet的nci-h1975细胞中,与西妥昔单抗相比,em-1mab在低附着培养条件下具有至少518倍改善的效力。表22示出了测定法的汇总。表22.厄洛替尼与em1-mab的组合有效抑制egfr突变体细胞系的生长在标准2d培养条件和低附着形式下评价了厄洛替尼加em1-mab的组合对细胞生长的抑制。如以上在“抗体对人瘤细胞生长或活力的抑制”中所述接种nci-h3255和hcc-827细胞。将hgf(7.5ng/ml,r&dsystems目录号294-hgn)加入细胞,同时用抗体进行处理。细胞使用不同浓度的抗体(终浓度0.11–700nm)或厄洛替尼(终浓度0.46–3000nm)或厄洛替尼加抗体的组合处理,它们各自以递增量以固定比率(例如,最低浓度的组合=最低浓度的抗体(0.11nm)+最低浓度的厄洛替尼(0.46nm))使用。一些孔不用hgf或抗体处理,作为对照。将细胞在37℃、5%的co2下温育72小时,然后使用celltiter-试剂(promega)检测活细胞,并且如以上在“人瘤细胞生长的抑制(nci-h292生长和nci-h322生长测定法)”中所述分析数据。表23汇总了该实验的结果。在该表中,各组合的ic50值是相对于抗体或厄洛替尼而言的,具体取决于括号中指示的内容。在nci-h3255和hcc-827细胞(egfr突变体细胞系)中,将em1-mab加入厄洛替尼中既提高了对细胞活力的抑制效力,又更有效产生了总共更少的活细胞。在使用标准2d条件的nci-h3255细胞中,单独厄洛替尼的ic50为122nm,而组合为49nm。相似地,在hcc-827细胞中,单独厄洛替尼的ic50为27nm,而组合为15nm。另外,厄洛替尼加em1-mab的组合比厄洛替尼加西妥昔单抗的组合更有效。因此,在hgf存在下,添加em1-mab提高了厄洛替尼在该测定法中的有效性。nci-h3255细胞表达l858r突变体egfr和扩增的cmet。hcc-827细胞表达在氨基酸位置746和750处具有缺失的egfr突变体和野生型c-met。在标准或低附着培养下,与单独厄洛替尼相比,em1-mab在厄洛替尼存在下对hcc-827和nci-3255的活力具有更强效应。表23.实例13.在体外细胞系中em1-mab的抗体介导的细胞毒性(adcc)adcc测定法如先前所述执行(scallon等人,molimmunol44:1524-15342007)。简言之,通过ficoll梯度从人血液中纯化pbmcs,并且用作用于adcc测定法的效应细胞。nci-h292、nci-h1975或nci-h441细胞用作靶细胞,采用1个靶细胞与50个效应细胞的比率。将靶细胞用batda(perkinelmer)在37℃预标记20分钟,洗涤两次并重悬于dmem、10%热灭活的fbs、2mml-谷氨酰胺(均来自invitrogen)中。组合靶细胞(1×104个细胞)和效应细胞(0.5×106个细胞),并且将100μl细胞加入96孔u形底部板的孔中。加入另外100μl连同或不连同野生型和蛋白酶抗性mab构建体。所有样品均一式两份执行。将板以200g离心3分钟,在37℃温育2小时,随后再次以200g离心3分钟。从每个孔中取出总共20μl的上清液,然后加入200μl基于delphiaeuropium的试剂(perkinelmer)并测定细胞溶解。使用envision2101多标记微孔板检测仪(perkinelmer)来测定荧光。使数据针对使用0.67%的tritonx-100(sigmaaldrich)的最大细胞毒性和使用下列公式通过在不存在任何抗体的情况下来自靶细胞的batda自发释放测定的最小对照标准化:(实验释放–自发释放)/(最大释放–自发释放)x100%。使用graphpadprismv5将数据拟合至s形剂量-响应模型。em1mab和对比物的adcc结果汇总于表24(nci-h292细胞)、表25(nci-h1975细胞)和表26(nci-h441细胞)及表27(nci-h1993细胞)中,并且列出了ec50值以及所实现的最大裂解。nci-h292细胞表达野生型(wt)egfr、wtc-met和wtkras;nci-h1975细胞表达突变体egfr(l858rt790m)、wtcmet和wtkras;nci-h441表达wtegfr、wtcmet和突变体kras(g12v),并且nci-h1993细胞表达wtegfr、扩增的cmet、wtkras。kras也称为gtp酶kras以及v-ki-ras2kirsten大鼠肉瘤病毒癌基因同源物。与西妥昔单抗和正常岩藻糖型式em1-mab相比,em1-mab具有更高效力的adcc响应,如具有更低ec50值所指示。在所实现的最大裂解方面,与西妥昔单抗和正常岩藻糖双特异性mab相比,em1mab具有更高的疗效。根据表24-27的概况,em-1mab具有对细胞的adcc活性,该细胞具有突变体和wtegfr、wt且水平正常和扩增的cmet,以及wt和突变体kras。表24.表25.表26.表27.实例14.使用em1-mab的瘤疗效研究如实例7“使用双特异性egfr/c-met分子的瘤疗效研究”中所述分析em1mab抵抗瘤生长的疗效。简而言之,使用cultrex将nci-h292-hgf细胞以2×106皮下(s.c.)植入雌性scidbeige小鼠中。植入后7天,按瘤体积将小鼠分为5组,每组10只小鼠。在每组起始的平均瘤体积在62-66mm3(小瘤)范围内之后,开始给药。每周腹腔内(i.p.)给予pbs或治疗剂2次。疗效的评价还采用了skmes-hgf,即一种用人hgf(肝细胞生长因子)转染的人鳞状细胞癌。将这些细胞以10×106皮下植入雌性scidbeige小鼠中。植入后12天,按瘤体积将这些小鼠分为5组,每组8只小鼠。每组起始的平均瘤体积在98-101mm3(大瘤)范围内时,开始第一研究。每周2次腹膜内给予pbs或治疗mab,共给予4周。在较大尺寸瘤研究中,在瘤体积为约200-300mm3后将小鼠分成2组。然后每周用西妥昔单抗(20mg/kg)或em1-mab(20mg/kg)腹膜内处理这些小鼠2次(3周)。数据的汇总示于表28中。图10示出了分子随时间推移的疗效。在h292-hgf小瘤模型以及skmes-hgf小瘤模型和大瘤模型中,与西妥昔单抗相比,em1-mab具有改善的瘤抑制概况。表28.表29示出了来自skmes-hgf瘤的处理组中的瘤尺寸,并且表30示出了抗瘤活性。在低至1mg/kg的多种剂量下,em1-mab将skmes-hgf模型中的瘤生长抑制97%或更多。虽然最初西妥昔单抗非常有效(20mg/kg下88%的tgi),但在给药结束后,经西妥昔单抗处理的瘤又重新长出。相反,用5或20mg/kg的em1-mab处理的瘤在研究过程中(>2个月)未重新长出。表29表30实例15.体外使用em1-mab时对细胞迁移的抑制方法在nih-1650细胞中评估了em-mab和对照单价抗体对抑制瘤细胞迁移的效应。将egfr突变体细胞系h1650(具有外显子19突变[缺失e746,a750]的肺细支气管肺泡癌细胞)在正常培养条件(37℃、5%的co2、95%湿度)下培养于组织培养瓶中。所有培养基和补充物按细胞供应商(americantypeculturecollection,manassas,va,usa)的建议采用。通过将h1650肺瘤细胞以10,000个细胞/孔及200μl/孔接种于“u”形底超低附着性(ula)96孔板(corning,tewksbury,usa)中来生成球体。这些平板刺激在24小时内自发形成单个细胞球体(在37℃、5%的co2下温育后)并且使球体在正常培养条件下生长四天。在37℃下用无菌水中的0.1%明胶(emdmillipore,billerica,usa)包被圆底96孔板(bdbioscience)1小时。对于化合物评价研究,将第4天的10,000个细胞瘤球体(h1650和nci-h1975)转移到包被的圆底板中,并且在具有20ng/ml的hgf(r&dsystems)的稀释系列中使用em1-mab、具有低岩藻糖含量的对照单价抗egfrmabe1-f405l-gp120-k409r、具有低岩藻糖含量的对照单价抗cmetmabm1-k409r-gp120-f405l,以及两种单价抗体e1-f405l-gp120-k409r和m1-k409r-gp120-f405l(以低岩藻糖形式制备)的组合进行处理。使用为igg1κ同种型对照(浓度等于最高药物处理的细胞)的媒介物处理对照。在48小时处,通过在具有2x物镜的全自动operetta高内涵成像系统(perkinelmer)中使用明视场图像测量迁移细胞所覆盖的面积,分析化合物的效应。通过除以仅培养基的对照以形成相对于对照的细胞迁移百分比而将数据归一化,以此来评估由于处理效应引起的对细胞迁移(总面积)的抑制。因此,小于1的值将抑制细胞迁移。结果与对照单价抗egrf和抗c-met抗体e1-f405l-gp120-k409r和m1-k409r-gp120-f405l的组合相比,em1-mab在h1650(l858regfr突变体)和h1975(l858r/t790megfr突变体)细胞中展现出细胞迁移的强效协同抑制。在h1650细胞中,与同种型对照相比,六种最高浓度的em1-mab显著抑制了细胞迁移(p<0.001)。em1-mab的ec50值为0.23nm,而单特异性对照抗体的组合的ec50值为4.39nm。因此,与单价对照抗体的组合相比,em1-mab在抑制h1650细胞迁移方面约19倍更有效。对于h1650和h1975细胞而言,em1-mab的细胞迁移抑制的水平优于单特异性对照mab的组合。表31示出了该测定法的ec50值。表31.*抗体具有低岩藻糖含量实例16.抗c-met抗体069和5d5的表位作图使用线性和限制性clips肽技术对抗c-metmab069结合表位作图。对人cmet的sema、psi和ig域进行肽扫描。使用上述cmet的氨基酸序列利用标准fmoc化学来合成线性和clips肽,并且使用三氟酸与清除剂进行去保护。使用支架化学连接肽(chemicallylinkedpeptidesonscaffold,clips)技术在化学支架上合成限制性肽以便重建构象表位(timmerman等人,jmolrecognition20:283,2007)。将线性和限制性肽偶联到pepscan卡并使用基于pepscan的elisa进行筛选(slootstra等人,moleculardiversity1,87-96,1996)。抗c-metmab069结合表位是由c-met第239-253位氨基酸pefrdsypikyvhaf(seqidno:238)和第346-361位氨基酸faqskpdsaepmdrsa(seqidno:239)组成的不连续表位。c-met氨基酸序列以seqidno:201示出。使用类似方法对mab5d5(metmab,奥纳珠单抗)表位作图。mab5d5结合c-met第325-340位残基pgaqlarqigaslndd(seqidno:240)。实例17.使用em1-mab的体内瘤疗效研究如实例7“使用双特异性egfr/c-met分子的瘤疗效研究”以及采用具有egfr突变或egfr和/或c-met扩增的另外瘤细胞系的实例14中所述分析em1mab抵抗瘤生长的疗效。简而言之,将表达人hgf的snu-5、h1975、hcc827细胞、h1975细胞,或者针对其对厄洛替尼增强的抗性所选择的hcc827细胞克隆(hcc827-er1细胞)皮下(s.c.)植入雌性裸小鼠中,不同的是将snu-5细胞植入cr17/scid小鼠中。以所指示的剂量和表32中所示的日程,为小鼠腹腔内给予pbs或em1-mab、西妥昔单抗(cas205923-56-4)、厄洛替尼(cas183321-74-6)、阿法替尼(cas439081-18-2),或em-1mab与阿法替尼的组合及em-1mab与厄洛替尼的组合。抗瘤疗效以%tgi(瘤生长抑制)测量,按照100-%t/c计算(在实例7中所述的给定天,t=处理组的平均瘤尺寸;c=对照组的平均瘤尺寸)。在具有原发egfr激活突变的瘤(不抵抗egfrtki)中:(hcc827瘤,egfrdel(e746,a750)),10mg/kg剂量的em1-mab将瘤生长抑制82%。厄洛替尼在该模型中效果相似,厄洛替尼与em1-mab的组合也是如此。图11示出了在hcc827瘤模型中治疗剂随时间推移的疗效。在具有野生型egfr和c-met基因扩增的瘤(胃癌模型snu-5)中,em1-mab显示出抗瘤活性且瘤完全消退(在第34天98%的tgi,使用单因素方差分析,之后使用games-howell进行个体比较得出,与媒介物相比,p<0.01)。在该模型中,抗egfrmab西妥昔单抗的抗瘤活性较低,在第34天为49%。图12示出了在snu-5模型中治疗剂随时间推移的疗效。在包含原发egfr激活突变和t790megfr突变的nsclc模型(h1975模型)中测试了em1-mab,该t790megfr突变使瘤保持抵抗第一代egfrtki。在植入裸小鼠中的h1975细胞系模型中em1-mab以57%的tgi抑制瘤生长(使用prism3.03进行对数秩分析得出,与pbs媒介物相比,p<0.0001)。如预期,厄洛替尼在具有t790m突变的该模型中没有效果。阿法替尼与em1-mab效果相同(57%的tgi)。西妥昔单抗以及em1-mab与阿法替尼的组合最有效,使瘤分别以91%和96%瘤生长抑制发生消退(使用prism3.03进行对数秩分析得出,西妥昔单抗与pbs和em1-mab+阿法替尼相比及与pbs+阿法替尼媒介物组相比,p<0.0001)。c-met信号传导途径在该模型中未被激活,因为小鼠hgf不结合于人c-met。在使用慢病毒转导系统进行工程改造而表达人hgf的若干模型中,测试了em1-mab。这允许对体内c-met途径的配体激活进行建模,因为小鼠hgf不会激活所植入人瘤细胞上的人c-met。skmes-hgf模型得出的结果示于实例14和图10中,并且%tgi汇总于表32中。在h1975-hgf模型中em1-mab将瘤生长抑制了71%(使用prism3.03进行对数秩分析得出,与pbs媒介物相比,p<0.0001)。阿法替尼、厄洛替尼和西妥昔单抗在该模型中不那么有效。em1-mab与阿法替尼的组合非常有效(96%tgi,使用prism3.03进行对数秩分析得出,与pbs+阿法替尼媒介物组相比,p<0.0001)。图13示出了在h1975-hgf模型中分子随时间推移的疗效。因此在c-met途径被激活的瘤模型中,厄洛替尼、阿法替尼和西妥昔单抗失去了其抗瘤疗效。在特征在于原发egfr激活突变和由于c-met基因扩增而对第一代egfrtki(厄洛替尼)抗性增强的瘤模型(hcc827-er1模型)中,测试了em1-mab。10mg/kg剂量的em1-mab使所植入的hcc827-er1瘤部分消退,在第25天达86%的tgi,并且比单独的厄洛替尼(在第25天达65%的tgi)更有效。em1-mab与厄洛替尼的组合未进一步改善疗效。图14示出了分子随时间推移的疗效。因此,在具有野生型egfr、具有原发激活egfr突变、具有与对egfr治疗剂的抗性相关的egfr突变t790m的瘤模型中,以及在c-met以配体依赖性(自分泌hgf表达)或非配体依赖性(c-met基因扩增)方式激活的模型中,em1-mab展示出疗效。在一些瘤模型中,em1-mab与厄洛替尼或阿法替尼的组合可改善疗效。表32.biw=两周一次qd=每天一次wt=野生型amp=扩增实例18.em1mab诱导的体内egfr和c-met的降解为了证实在瘤中egfr和c-met被em1-mab接合,在20mg/kgem1-mab的单次剂量后在不同时间从h1975-hgf瘤取样。制备瘤裂解物,归一化为总蛋白,并且将样品在sds-page凝胶上电泳。将凝胶转移到硝化纤维素并且针对egfr(小鼠(mab)抗人egfr(egf-r2);santacruzbiotechnology,目录号sc-73511)或c-met(小鼠(mab)抗人met(l41g3);cellsignalingtechnology,目录号3148)进行蛋白质印迹。将egfr水平归一化到gapdh;将c-met水平归一化到肌动蛋白。将来自经em1-mab处理的瘤的受体的水平与经pbs处理的瘤的受体水平比较,从而得到%总受体。em1-mab处理将h1975-hgf瘤中的总egfr和cmet受体水平减少到对照的20%至60%之间,具体取决于所分析的时间点。图15示出了与pbs相比随时间推移的平均受体水平。pegfr、pc-met和pakt也在em1的单次剂量后的72小时减少。实例19.egfr/c-met双特异性mab的igg1和igg2σ变体同种型的抗瘤活性比较为了更好地理解效应子功能对h1975-hgf模型中所观察到的疗效的贡献,在em1-mab与em1-mab变体之间进行比较,该em1-mab变体包含的igg2fc在igg2上具有效应子沉默替换v234a/g237a/p238s/h268a/v309l/a330s/p331s(国际专利申请wo2011/066501中所述的替换)(根据eu索引进行编号)。具有v234a/g237a/p238s/h268a/v309l/a330s/p331s替换的igg2抗体不与fc受体或效应子细胞(诸如nk细胞和巨噬细胞)相互作用。因此em1-mab的igg2v234a/g237a/p238s/h268a/v309l/a330s/p331s变体所观察到的任何活性丧失可表示效应子介导的机制诸如adcc和/或adcp所促成的抗瘤活性。在上述h1975-hgf模型中植入瘤细胞后32天,指示了与亲本em1-mab相比,em1-mab的igg2v234a/g237a/p238s/h268a/v309l/a330s/p331s变体的抗瘤活性丧失,表明效应子介导的机制促成了em-1mab的功能。图16示出了分子的抗瘤活性。序列表seqidno:73,prt,智人,egfr(包含24个氨基酸的信号序列,成熟蛋白质开始于第25位残基)seqidno:75,prt,智人,腱生蛋白-c>seqidno:101prt智人cmet>seqidno:179prt人工的结合egfr的fn3域的fg环hnvykdtnx9rgl;其中x9为m或i>seqidno:180prt人工的结合egfr的fn3域的fg环lgsyvfehdvml(seqidno:180),>seqidno:181prt人工的结合egfr的fn3域的bc环x1x2x3x4x5x6x7x8(seqidno:181);其中x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;并且x8为y、f或l。>seqidno:182prt人工的结合egf的rfn3域lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvhnvykdtnx9rglplsaeftt(seqidno:182),x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;x8为y、f或l;并且x9为m或i>seqidno:183prt人工的结合egfr的fn3域lpapknlvvsevtedslrlswx1x2x3x4x5x6x7x8dsfliqyqesekvgeainltvpgsersydltglkpgteytvsiygvlgsyvfehdvmlplsaeftt(seqidno:183),其中x1为a、t、g或d;x2为a、d、y或w;x3为p、d或n;x4为l或不存在;x5为d、h、r、g、y或w;x6为g、d或a;x7为a、f、g、h或d;并且x8为y、f或l。>seqidno:184prt人工的结合c-met的fn3域c链和cd环序列dsfx10iryx11ex12x13x14x15gx16(seqidno:184),其中x10为w、f或v;x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;并且x16为e或d;并且>seqidno:185prt人工的结合c-met的fn3域f链和fg环teyx17vx18ix19x20vkggx21x22sx23(seqidno:185),其中x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。>seqidno:186prt人工的结合c-met的fn3域lpapknlvvsrvtedsarlswtapdaafdsfx10iryx11ex12x13x14x15gx16aivltvpgsersydltglkpgteyx17vx18ix19x20vkggx21x22sx23plsaeftt(seqidno:186),其中x10为w、f或v;并且x11为d、f或l;x12为v、f或l;x13为v、l或t;x14为v、r、g、l、t或s;x15为g、s、a、t或k;x16为e或d;x17为y、w、i、v、g或a;x18为n、t、q或g;x19为l、m、n或i;x20为g或s;x21为s、l、g、y、t、r、h或k;x22为i、v或l;并且x23为v、t、h、i、p、y或l。>seqidno:187prt人工的包含双特异性egfr/c-metfn3域的egfrfn3域的分子lpapknlvvsx24vtx25dsx26rlswddpx27afyx28sfliqyqx29sekvgeaix30ltvpgsersydltglkpgteytvsiyx31vhnvykdtnx32rglplsax33ftt(seqidno:187),其中x24为e、n或r;x25为e或p;x26为l或a;x27为h或w;x28为e或d;x29为e或p;x30为n或v;x31为g或y;x32为m或i;并且x33为e或i;>seqidno:188包含双特异性egfr/c-metfn3域的c-metfn3域的分子lpapknlvvsx34vtx35dsx36rlswtapdaafdsfwiryfx37fx38x39x40gx41aix42ltvpgsersydltglkpgteyvvnix43x44vkggx45ispplsax46ftt(seqidno:188);其中x34为e、n或r;x35为e或p;x36为l或a;x37为e或p;x38为v或l;x39为g或s;x40为s或k;x41为e或d;x42为n或v;x43为l或m;x44为g或s;x45为s或k;并且x46为e或i。>seqidno:189egfrmabe1vhqvqlvesgggvvqpgrslrlscaasgftfstygmhwvrqapgkglewvaviwddgsykyygdsvkgrftisrdnskntlylqmnslraedtavyycardgitmvrgvmkdyfdywgqgtlvtvss>seqidno:190egfrmabe1vlaiqltqspsslsasvgdrvtitcrasqdissalvwyqqkpgkapklliydasslesgvpsrfsgsesgtdftltisslqpedfatyycqqfnsypltfgggtkveik>seqidno:191egfrmabe2vh1evqlvesggglvqpggslrlscaasgftfssywmnwvrqapgkglewvanikkdgsekyy61vdsvkgrftisrdnaknslylqmnslraedtavyycardlgwgwgwyfdlwgrgtlvtvs121s>seqidno:192egfrmabe2vl1eivltqspatlslspgeratlscrasqsvssylawyqqkpgqaprlliydasnratgipa61rfsgsgsgtdftltisslepedfavyycqqrsnwpptfgqgtkveik>seqidno:193cmetmabm1vhqvqlvqsgaevkkpgasvkvscetsgytftsygiswvrqapghglewmgwisayngytnyaqklqgrvtmttdtststaymelrslrsddtavyycardlrgtnyfdywgqgtlvtvss>seqidno:194cmetmabm1vldiqmtqspssvsasvgdrvtitcrasqgisnwlawfqhkpgkapklliyaassllsgvpsrfsgsgsgtdftltisslqpedfatyycqqansfp-itfgqgtrleik>seqidno:195cmetmabm2vhevqlvesggglvkpggslklscaasgftfsdyymywvrqtpekrlewvatisddgsytyypdsvkgrftisrdnaknnlylqmsslksedtamyycareglyyygsgsyynqdywgqgtlvtvss>seqidno:196cmetmabm2vlqltqspsslsasvgdrvtitcrasqglssalawyrqkpgkapklliydasslesgvpsrfsgsgsgtdftltisslqpedfatyycqqftsypqitfgqgtrleik>seqidno:197具有f405l的gp120重链qvqlvqsgaevkkpgasvkvscqasgyrfsnfvihwvrqapgqrfewmgwinpyngnkefsakfqdrvtftadtsantaymelrslrsadtavyycarvgpyswddspqdnyymdvwgkgttvivssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:198具有k409r的gp120重链qvqlvqsgaevkkpgasvkvscqasgyrfsnfvihwvrqapgqrfewmgwinpyngnkefsakfqdrvtftadtsantaymelrslrsadtavyycarvgpyswddspqdnyymdvwgkgttvivssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:199em1-mabh1(抗egfr,405l)qvqlvesgggvvqpgrslrlscaasgftfstygmhwvrqapgkglewvaviwddgsykyygdsvkgrftisrdnskntlylqmnslraedtavyycardgitmvrgvmkdyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:200em-1mabl1aiqltqspsslsasvgdrvtitcrasqdissalvwyqqkpgkapklliydasslesgvpsrfsgsesgtdftltisslqpedfatyycqqfnsypltfgggtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:201em-1mabh2(k409r,抗cmet)qvqlvqsgaevkkpgasvkvscetsgytftsygiswvrqapghglewmgwisayngytnyaqklqgrvtmttdtststaymelrslrsddtavyycardlrgtnyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:202em-1mabl2diqmtqspssvsasvgdrvtitcrasqgisnwlawfqhkpgkapklliyaassllsgvpsrfsgsgsgtdftltisslqpedfatyycqqansfpitfgqgtrleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:203h1恒定区astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:204h2恒定区astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:205em1-mabh1cdnapdr000015499caggtgcagctggtcgagagcggcggaggggtggtgcagcccggcagaagcctgaggctgtcctgcgccgccagcggcttcaccttcagcacctacggcatgcactgggtgcggcaggccccaggcaagggcctggagtgggtggccgtgatctgggacgacggcagctacaagtactacggcgacagcgtgaagggcaggttcaccatcagcagggacaacagcaagaacaccctgtacctgcagatgaacagcctgagggccgaggacaccgccgtgtactactgcgccagggacggcatcaccatggtgcggggcgtgatgaaggactacttcgactactggggccagggcaccctggtgaccgtgagcagcgccagcaccaagggcccaagcgtgttccccctggcccccagcagcaagagcaccagcggcggcacagccgccctgggctgcctggtgaaggactacttccccgagccagtgaccgtgtcctggaactctggcgccctgacctccggcgtgcacaccttccccgccgtgctgcagagcagcggcctgtacagcctgagcagcgtggtgaccgtgcccagcagcagcctgggcacccagacctacatctgcaacgtgaaccacaagcccagcaacaccaaggtggacaagagagtggagcccaagagctgcgacaagacccacacctgccccccctgcccagccccagagctgctgggcggacccagcgtgttcctgttcccccccaagcccaaggacaccctgatgatcagcaggacccccgaggtgacctgcgtggtggtggacgtgagccacgaggacccagaggtgaagttcaactggtacgtggacggcgtggaggtgcacaacgccaagaccaagcccagagaggagcagtacaacagcacctacagggtggtgtccgtgctgaccgtgctgcaccaggactggctgaacggcaaggaatacaagtgcaaggtctccaacaaggccctgccagcccccatcgaaaagaccatcagcaaggccaagggccagccacgggagccccaggtgtacaccctgccccccagccgggaggagatgaccaagaaccaggtgtccctgacctgtctggtgaagggcttctaccccagcgacatcgccgtggagtgggagagcaacggccagcccgagaacaactacaagaccacccccccagtgctggacagcgacggcagcttcctcctgtacagcaagctgaccgtggacaagtccaggtggcagcagggcaacgtgttcagctgcagcgtgatgcacgaggccctgcacaaccactacacccagaagtccctgagcctgagccccggcaaatga>seqidno:206em1-mabl1cdnapdr000015499atccagctgacccagagccccagcagcctgagcgccagcgtgggcgaccgggtgaccatcacctgccgggccagccaggacatcagcagcgccctggtctggtatcagcagaagcccggcaaggcccccaagctgctgatctacgacgccagctccctggaaagcggcgtgcccagccggttcagcggcagcgagagcggcaccgacttcaccctgaccatcagcagcctgcagcccgaggacttcgccacctactactgccagcagttcaacagctaccccctgacctttggcggcggaacaaaggtggagatcaagcgtacggtggccgctcccagcgtgttcatcttcccccccagcgacgagcagctgaagagcggcaccgccagcgtggtgtgcctgctgaacaacttctacccccgggaggccaaggtgcagtggaaggtggacaacgccctgcagagcggcaacagccaggagagcgtcaccgagcaggacagcaaggactccacctacagcctgagcagcaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaggtgacccaccagggcctgtccagccccgtgaccaagagcttcaacaggggcgagtgctga>seqidno:207em-1mabh2cdnapdr000016584caggttcagctggtgcagtctggagctgaggtgaagaagcctggggcctcagtgaaggtctcctgcgagacttctggttacacctttaccagctatggtatcagctgggtgcgacaggcccctggacacgggcttgagtggatgggatggatcagcgcttacaatggttacacaaactatgcacagaagctccagggcagggtcaccatgaccacagacacatccacgagcacagcctacatggagctgaggagcctgagatctgacgacacggccgtgtattactgtgcgagagatctgagaggaactaactactttgactactggggccagggaaccctggtcaccgtctcctcagcctccaccaagggcccaagcgtgttccctctggcccccagcagcaagagcacatctggcggaacagccgccctgggctgcctggtgaaagactacttccccgagcccgtgaccgtgtcctggaactctggcgccctgaccagcggcgtgcacacctttccagccgtgctgcagagcagcggcctgtacagcctgtccagcgtggtgaccgtgcccagcagctccctgggcacccagacctacatctgcaacgtgaaccacaagcccagcaacaccaaggtggacaagcgggtggaacccaagagctgcgacaagacccacacctgtcccccctgccctgcccctgaactgctgggcggaccctccgtgttcctgttccccccaaagcccaaggacaccctgatgatcagccggacccccgaagtgacctgcgtggtggtggacgtgtcccacgaggaccctgaagtgaagttcaattggtacgtggacggcgtggaagtgcacaacgccaagaccaagcccagagaggaacagtacaacagcacctaccgggtggtgtccgtgctgacagtgctgcaccaggactggctgaacggcaaagagtacaagtgcaaggtctccaacaaggccctgcctgctcccatcgagaaaaccatcagcaaggccaagggccagccccgcgagcctcaggtgtacacactgcctcccagccgggaagagatgaccaagaaccaggtgtccctgacctgtctggtgaaaggcttctaccccagcgatatcgccgtggaatgggagagcaacggacagcccgagaacaactacaagaccaccccccctgtgctggacagcgacggctccttcttcctgtactctcggctgaccgtggacaagagccggtggcagcagggaaacgtgttcagctgcagcgtgatgcacgaggccctgcacaaccactacacccagaagtccctgagcctgagccccgggaagtga>seqidno:208em-1mabl2cdnapdr000016584gacatccagatgacccagtccccctcctccgtgtccgcctctgtgggcgacagagtgaccatcacctgtcgggcctcccagggcatctccaactggctggcctggttccagcacaagcccggcaaggcccccaagctgctgatctacgccgcctcctccctgctgtccggcgtgccctccagattctccggctctggctccggcaccgacttcaccctgaccatctccagcctgcagcccgaggacttcgccacctactactgccagcaggccaactccttccccatcaccttcggccagggcacccggctggaaatcaagcgtacggtggccgctcccagcgtgttcatcttcccccccagcgacgagcagctgaagagcggcaccgccagcgtggtgtgcctgctgaacaacttctacccccgggaggccaaggtgcagtggaaggtggacaacgccctgcagagcggcaacagccaggagagcgtcaccgagcaggacagcaaggactccacctacagcctgagcagcaccctgaccctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaggtgacccaccagggcctgtccagccccgtgaccaagagcttcaacaggggcgagtgctga>seqidno:209gp120轻链eivltqspgtlslspgeratfscrsshsirsrrvawyqhkpgqaprlvihgvsnrasgisdrfsgsgsgtdftltitrvepedfalyycqvygassytfgqgtklerkrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:210e1hc1hcdr1tygmh>seqidno:211e1hc1hcdr2viwddgsykyygdsvkg>seqidno:212e1hc1hcdr3dgitmvrgvmkdyfdy>seqidno:213e1lc1lcdr1rasqdissalv>seqidno:214e1lc1lcdr2dassles>seqidno:215e1lc1lcdr3qqfnsyplt>seqidno:216e1hc2hcdr1sygis>seqidno:217e1hc2hcdr2wisayngytnyaqklqg>seqidno:218e1hc2hcdr3dlrgtnyfdy>seqidno:219e1lc2lcdr1rasqgisnwla>seqidno:220e1lc2lcdr2aasslls>seqidno:221e1lc2lcdr3qqansfpit>seqidno:222e2mabhc1hcdr1sywmn>seqidno:223e2mabhc1hcdr2nikkdgsekyyvdsvkg>seqidno:224e2mabhc1hcdr3dlgwgwgwyfdl>seqidno:225e2mablc1lcdr1rasqsvssyla>seqidno:226e2mablc1lcdr2dasnrat>seqidno:227e2mablc1lcdr3qqrsnwppt>seqidno:228e2mabhc2hcdr1dyymy>seqidno:229e2mabhc2hcdr2tisddgsytyypdsvkg>seqidno:230e2mabhc2hcdr3eglyyygsgsyynqdy>seqidno:231e2mablc2lcdr1rasqglssala>seqidno:232e2mablc2lcdr2dassles>seqidno:233e2mablc2lcdr3qqftsypqit>seqidno:234e2mabhc1(egfr-f405l)evqlvesggglvqpggslrlscaasgftfssywmnwvrqapgkglewvanikkdgsekyyvdsvkgrftisrdnaknslylqmnslraedtavyycardlgwgwgwyfdlwgrgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfllyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:235e2mablc1(egfr)eivltqspatlslspgeratlscrasqsvssylawyqqkpgqaprlliydasnratgiparfsgsgsgtdftltisslepedfavyycqqrsnwpptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:236e2mabhc2(c-met-k409r)evqlvesggglvkpggslklscaasgftfsdyymywvrqtpekrlewvatisddgsytyypdsvkgrftisrdnaknnlylqmsslksedtamyycareglyyygsgsyynqdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqqgnvfscsvmhealhnhytqkslslspgk>seqidno:237e2mablc2(cmet)qltqspsslsasvgdrvtitcrasqglssalawyrqkpgkapklliydasslesgvpsrfsgsgsgtdftltisslqpedfatyycqqftsypqitfgqgtrleikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec>seqidno:238mab069的c-met不连续表位pefrdsypikyvhaf>seqidno:239mab069的c-met不连续表位faqskpdsaepmdrsa>seqidno:2405d5mab表位pgaqlarqigaslndd序列表<110>詹森生物科技公司<120>双特异性egfr/c-met抗体<130>jbi5032wopct<150>61/728,912<151>2012-11-21<150>61/782,550<151>2013-03-14<150>61/809,541<151>2013-04-08<150>61/864,717<151>2013-08-12<150>61/892,797<151>2013-10-18<160>240<170>patentin版本3.5<210>1<211>89<212>prt<213>人工序列<220><223>tenconfn3支架<400>1leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpthralaproaspalaalapheaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvallysglyglyhisargser65707580asnproleuseralagluphethrthr85<210>2<211>58<212>dna<213>人工序列<220><223>引物<400>2ggaaacaggatctaccatgctgccggcgccgaaaaacctggttgtttctgaagttacc58<210>3<211>21<212>dna<213>人工序列<220><223>引物<400>3aacaccgtagatagaaacggt21<210>4<211>131<212>dna<213>人工序列<220><223>引物<400>4cggcggttagaacgcggctacaattaatacataaccccatccccctgttgacaattaatc60atcggctcgtataatgtgtggaattgtgagcggataacaatttcacacaggaaacaggat120ctaccatgctg131<210>5<211>21<212>dna<213>人工序列<220><223>引物<400>5cggcggttagaacgcggctac21<210>6<211>66<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(26)..(27)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(29)..(30)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(32)..(33)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(35)..(36)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(38)..(39)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(41)..(42)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(44)..(45)<223>n为a、c、g或t<400>6ggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnaacaccgtagataga60aacggt66<210>7<211>69<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(26)..(27)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(29)..(30)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(32)..(33)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(35)..(36)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(38)..(39)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(41)..(42)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(44)..(45)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(47)..(48)<223>n为a、c、g或t<400>7ggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnsnnaacaccgtagat60agaaacggt69<210>8<211>72<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(26)..(27)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(29)..(30)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(32)..(33)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(35)..(36)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(38)..(39)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(41)..(42)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(44)..(45)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(47)..(48)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(50)..(51)<223>n为a、c、g或t<400>8ggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnsnnsnnaacaccgta60gatagaaacggt72<210>9<211>75<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(26)..(27)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(29)..(30)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(32)..(33)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(35)..(36)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(38)..(39)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(41)..(42)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(44)..(45)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(47)..(48)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(50)..(51)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(53)..(54)<223>n为a、c、g或t<400>9ggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnsnnsnnsnnaacacc60gtagatagaaacggt75<210>10<211>80<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(28)..(29)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(31)..(32)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(34)..(35)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(37)..(38)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(40)..(41)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(43)..(44)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(46)..(47)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(49)..(50)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(52)..(53)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(55)..(56)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(58)..(59)<223>n为a、c、g或t<400>10rmggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnsnnsnnsnnsnna60acaccgtagatagaaacggt80<210>11<211>81<212>dna<213>人工序列<220><223>引物<220><221>尚未归类的特性<222>(26)..(27)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(29)..(30)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(32)..(33)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(35)..(36)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(38)..(39)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(41)..(42)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(44)..(45)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(47)..(48)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(50)..(51)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(53)..(54)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(56)..(57)<223>n为a、c、g或t<220><221>尚未归类的特性<222>(59)..(60)<223>n为a、c、g或t<400>11ggtggtgaattccgcagacagcggsnnsnnsnnsnnsnnsnnsnnsnnsnnsnnsnnsnn60aacaccgtagatagaaacggt81<210>12<211>81<212>dna<213>人工序列<220><223>引物<400>12aagatcagttgcggccgctagactagaaccgctgccatggtgatggtgatggtgaccgcc60ggtggtgaattccgcagacag81<210>13<211>30<212>dna<213>人工序列<220><223>引物<400>13cggcggttagaacgcggctacaattaatac30<210>14<211>22<212>dna<213>人工序列<220><223>引物<400>14catgattacgccaagctcagaa22<210>15<211>65<212>dna<213>人工序列<220><223>引物<400>15gagccgccgccaccggtttaatggtgatggtgatggtgaccaccggtggtgaattccgca60gacag65<210>16<211>37<212>dna<213>人工序列<220><223>引物<400>16aagaaggagaaccggtatgctgccggcgccgaaaaac37<210>17<211>89<212>dna<213>人工序列<220><223>引物<400>17tttgggaagcttctaggtctcggcggtcaccatcaccatcaccatggcagcggttctagt60ctagcggccccaactgatcttcaccaaac89<210>18<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>18leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>19<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>19leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpthrtyraspargaspglytyraspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>20<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>20leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpglytyrasnglyasphispheaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>21<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>21leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpaspaspproargglyphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>22<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>22leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpthrtrpprotyralaaspleuaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>23<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>23leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpglytyrasnglyasphispheaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>24<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>24leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpasptyraspleuglyvaltyrpheaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>25<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>25leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>26<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>26leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpthralaproaspalaalapheaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalleuglysertyrvalphe65707580gluhisaspvalmetleuproleuseralagluphethrthr8590<210>27<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>27leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthr8590<210>28<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>28leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalleuglysertyrvalphe65707580gluhisaspvalmetleuproleuseralailephethrthr8590<210>29<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>29leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpthrtrpprotyralaaspleuaspserpheleu202530ileglntyrglngluserglulysvalglyglualaileasnleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralagluphethrthr8590<210>30<211>37<212>prt<213>人工序列<220><223>引物<400>30alaalaglyalaalaglyglyalaglyalaalacyscysglyglythr151015alathrglycysthrglycyscysglyglycysglycyscysglyala202530alaalaalaalacys35<210>31<211>65<212>prt<213>人工序列<220><223>引物<400>31glyalaglycyscysglycyscysglycyscysalacyscysglygly151015thrthrthralaalathrglyglythrglyalathrglyglythrgly202530alathrglyglythrglyalacyscysalacyscysglyglythrgly354045glythrglyalaalaglyalathrcysglycysalaglyalacysala505560gly65<210>32<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>32leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyraspgluvalvalvalglyglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrtyrvalasnileleuglyvallysglyglyserileser65707580valproleuseralailephethrthr85<210>33<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>33leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphephe202530ileargtyraspglupheleuargserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrtrpvalthrileleuglyvallysglyglyleuvalser65707580thrproleuseralailephethrthr85<210>34<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>34leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrilevalasnilemetglyvallysglyglyserileser65707580hisproleuseralailephethrthr85<210>35<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>35leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnileleuglyvallysglyglyglyleuser65707580valproleuseralailephethrthr85<210>36<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>36leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserpheval202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnileleuglyvallysglyglytyrileser65707580ileproleuseralailephethrthr85<210>37<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>37leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrleuglupheleuleuglyglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnilemetglyvallysglyglythrvalser65707580proproleuseralailephethrthr85<210>38<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>38leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglyileasnglyvallysglyglytyrileser65707580tyrproleuseralailephethrthr85<210>39<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>39leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthraspleulysprogly505560thrglutyrglyvalthrileasnglyvallysglyglyargvalser65707580thrproleuseralailephethrthr85<210>40<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>40leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnileileglyvallysglyglyhisileser65707580leuproleuseralailephethrthr85<210>41<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>41leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilemetglyvallysglyglylysileser65707580proproleuseralailephethrthr85<210>42<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>42leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyralavalasnilemetglyvallysglyglyargvalser65707580valproleuseralailephethrthr85<210>43<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>43leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnileleuglyvallysglyglyserileser65707580valproleuseralailephethrthr85<210>44<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>44leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilemetglyvallysglyglyserileser65707580tyrproleuseralailephethrthr85<210>45<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>45leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnileleuglyvallysglyglytyrileser65707580ileproleuseralailephethrthr85<210>46<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>46leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalglnilemetglyvallysglyglythrvalser65707580proproleuseralailephethrthr85<210>47<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>47leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphegluphethrthralaglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilemetglyvallysglyglyserileser65707580proproleuseralailephethrthr85<210>48<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>48leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphegluleuleuserthrglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilemetglyvallysglyglyserileser65707580proproleuseralailephethrthr85<210>49<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>49leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphegluphevalserlysglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilemetglyvallysglyglyserileser65707580proproleuseralailephethrthr85<210>50<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>50metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphetrpileargtyraspgluvalvalvalglyglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrtyrvalasnileleuglyvallysgly180185190glyserileservalproleuseralailephethrthr195200205<210>51<211>204<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>51metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglyserleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyrpheglupheleuglyserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrvalvalasnilemetglyvallysglygly180185190lysileserproproleuseralailephethrthr195200<210>52<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>52metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphetrpileargtyrpheglupheleuglyserglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrvalvalglnileileglyvallysgly180185190glyhisileserleuproleuseralailephethrthr195200205<210>53<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>53metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphepheileargtyraspglupheleuargserglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrtrpvalthrileleuglyvallysgly180185190glyleuvalserthrproleuseralailephethrthr195200205<210>54<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>54metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphetrpileargtyrpheglupheleuglyserglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrvalvalasnilemetglyvallysgly180185190glylysileserproproleuseralailephethrthr195200205<210>55<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>55metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphetrpileargtyrpheglupheleuglyserglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrvalvalglnileileglyvallysgly180185190glyhisileserleuproleuseralailephethrthr195200205<210>56<211>205<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>56metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrgly859095glyglyglyserglyglyglyglyserglyglyglyglyserglygly100105110glyglysermetleuproalaprolysasnleuvalvalserargval115120125thrgluaspseralaargleusertrpthralaproaspalaalaphe130135140aspserphetrpileargtyrpheglupheleuglyserglygluala145150155160ilevalleuthrvalproglysergluargsertyraspleuthrgly165170175leulysproglythrglutyrvalvalglnileileglyvallysgly180185190glyhisileserleuproleuseralailephethrthr195200205<210>57<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>57metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175metglyvallysglyglylysileserproproleuseralailephe180185190thrthr<210>58<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>58metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyraspgluvalval130135140valglyglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrtyrvalasnile165170175leuglyvallysglyglyserileservalproleuseralailephe180185190thrthr<210>59<211>195<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>59metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaprometleuproalaprolysasn100105110leuvalvalserargvalthrgluaspseralaargleusertrpthr115120125alaproaspalaalapheaspserphetrpileargtyrphegluphe130135140leuglyserglyglualailevalleuthrvalproglysergluarg145150155160sertyraspleuthrglyleulysproglythrglutyrvalvalasn165170175ilemetglyvallysglyglylysileserproproleuseralaile180185190phethrthr195<210>60<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>60metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyraspgluvalval130135140valglyglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrtyrvalasnile165170175leuglyvallysglyglyserileservalproleuseralailephe180185190thrthr<210>61<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>61metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglylysileserproproleuseralailephe180185190thrthr<210>62<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>62metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>63<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>63metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140serlysglyaspalailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>64<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>64metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthr<210>65<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>65metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglylysileserproproleuseralailephe180185190thrthr<210>66<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>66metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>67<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>67metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140serlysglyaspalailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>68<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>68metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthr<210>69<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>69metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglylysileserproproleuseralailephe180185190thrthr<210>70<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>70metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>71<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>71metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140serlysglyaspalailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthr<210>72<211>194<212>prt<213>人工序列<220><223>结合egfr/cmet的双特异性分子<400>72metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthr<210>73<211>1210<212>prt<213>智人<400>73metargproserglythralaglyalaalaleuleualaleuleuala151015alaleucysproalaserargalaleugluglulyslysvalcysgln202530glythrserasnlysleuthrglnleuglythrphegluasphisphe354045leuserleuglnargmetpheasnasncysgluvalvalleuglyasn505560leugluilethrtyrvalglnargasntyraspleuserpheleulys65707580thrileglngluvalalaglytyrvalleuilealaleuasnthrval859095gluargileproleugluasnleuglnileileargglyasnmettyr100105110tyrgluasnsertyralaleualavalleuserasntyraspalaasn115120125lysthrglyleulysgluleuprometargasnleuglngluileleu130135140hisglyalavalargpheserasnasnproalaleucysasnvalglu145150155160serileglntrpargaspilevalserserasppheleuserasnmet165170175sermetasppheglnasnhisleuglysercysglnlyscysasppro180185190sercysproasnglysercystrpglyalaglyglugluasncysgln195200205lysleuthrlysileilecysalaglnglncysserglyargcysarg210215220glylysserproseraspcyscyshisasnglncysalaalaglycys225230235240thrglyproarggluseraspcysleuvalcysarglyspheargasp245250255glualathrcyslysaspthrcysproproleumetleutyrasnpro260265270thrthrtyrglnmetaspvalasnprogluglylystyrserphegly275280285alathrcysvallyslyscysproargasntyrvalvalthrasphis290295300glysercysvalargalacysglyalaaspsertyrglumetgluglu305310315320aspglyvalarglyscyslyslyscysgluglyprocysarglysval325330335cysasnglyileglyileglygluphelysaspserleuserileasn340345350alathrasnilelyshisphelysasncysthrserileserglyasp355360365leuhisileleuprovalalapheargglyaspserphethrhisthr370375380proproleuaspproglngluleuaspileleulysthrvallysglu385390395400ilethrglypheleuleuileglnalatrpprogluasnargthrasp405410415leuhisalaphegluasnleugluileileargglyargthrlysgln420425430hisglyglnpheserleualavalvalserleuasnilethrserleu435440445glyleuargserleulysgluileseraspglyaspvalileileser450455460glyasnlysasnleucystyralaasnthrileasntrplyslysleu465470475480pheglythrserglyglnlysthrlysileileserasnargglyglu485490495asnsercyslysalathrglyglnvalcyshisalaleucysserpro500505510gluglycystrpglyprogluproargaspcysvalsercysargasn515520525valserargglyargglucysvalasplyscysasnleuleuglugly530535540gluproarggluphevalgluasnserglucysileglncyshispro545550555560glucysleuproglnalametasnilethrcysthrglyargglypro565570575aspasncysileglncysalahistyrileaspglyprohiscysval580585590lysthrcysproalaglyvalmetglygluasnasnthrleuvaltrp595600605lystyralaaspalaglyhisvalcyshisleucyshisproasncys610615620thrtyrglycysthrglyproglyleugluglycysprothrasngly625630635640prolysileproserilealathrglymetvalglyalaleuleuleu645650655leuleuvalvalalaleuglyileglyleuphemetargargarghis660665670ilevalarglysargthrleuargargleuleuglngluarggluleu675680685valgluproleuthrproserglyglualaproasnglnalaleuleu690695700argileleulysgluthrgluphelyslysilelysvalleuglyser705710715720glyalapheglythrvaltyrlysglyleutrpileprogluglyglu725730735lysvallysileprovalalailelysgluleuargglualathrser740745750prolysalaasnlysgluileleuaspglualatyrvalmetalaser755760765valaspasnprohisvalcysargleuleuglyilecysleuthrser770775780thrvalglnleuilethrglnleumetpropheglycysleuleuasp785790795800tyrvalarggluhislysaspasnileglyserglntyrleuleuasn805810815trpcysvalglnilealalysglymetasntyrleugluaspargarg820825830leuvalhisargaspleualaalaargasnvalleuvallysthrpro835840845glnhisvallysilethrasppheglyleualalysleuleuglyala850855860gluglulysglutyrhisalagluglyglylysvalproilelystrp865870875880metalaleugluserileleuhisargiletyrthrhisglnserasp885890895valtrpsertyrglyvalthrvaltrpgluleumetthrpheglyser900905910lysprotyraspglyileproalasergluileserserileleuglu915920925lysglygluargleuproglnproproilecysthrileaspvaltyr930935940metilemetvallyscystrpmetileaspalaaspserargprolys945950955960phearggluleuileileglupheserlysmetalaargaspprogln965970975argtyrleuvalileglnglyaspgluargmethisleuproserpro980985990thraspserasnphetyrargalaleumetaspglugluaspmetasp99510001005aspvalvalaspalaaspglutyrleuileproglnglnglyphe101010151020pheserserproserthrserargthrproleuleuserserleu102510301035seralathrserasnasnserthrvalalacysileaspargasn104010451050glyleuglnsercysproilelysgluaspserpheleuglnarg105510601065tyrserseraspprothrglyalaleuthrgluaspserileasp107010751080aspthrpheleuprovalproglutyrileasnglnservalpro108510901095lysargproalaglyservalglnasnprovaltyrhisasngln110011051110proleuasnproalaproserargaspprohistyrglnasppro111511201125hisserthralavalglyasnproglutyrleuasnthrvalgln113011351140prothrcysvalasnserthrpheaspserproalahistrpala114511501155glnlysglyserhisglnileserleuaspasnproasptyrgln116011651170glnaspphepheprolysglualalysproasnglyilephelys117511801185glyserthralagluasnalaglutyrleuargvalalaprogln119011951200serserglupheileglyala12051210<210>74<211>53<212>prt<213>智人<400>74asnseraspserglucysproleuserhisaspglytyrcysleuhis151015aspglyvalcysmettyrileglualaleuasplystyralacysasn202530cysvalvalglytyrileglygluargcysglntyrargaspleulys354045trptrpgluleuarg50<210>75<211>2201<212>prt<213>智人<400>75metglyalametthrglnleuleualaglyvalpheleualapheleu151015alaleualathrgluglyglyvalleulyslysvalilearghislys202530argglnserglyvalasnalathrleuproglugluasnglnproval354045valpheasnhisvaltyrasnilelysleuprovalglyserglncys505560servalaspleugluseralaserglyglulysaspleualapropro65707580sergluprosergluserpheglngluhisthrvalaspglygluasn859095glnilevalphethrhisargileasnileproargargalacysgly100105110cysalaalaalaproaspvallysgluleuleuserargleugluglu115120125leugluasnleuvalserserleuarggluglncysthralaglyala130135140glycyscysleuglnproalathrglyargleuaspthrargprophe145150155160cysserglyargglyasnpheserthrgluglycysglycysvalcys165170175gluproglytrplysglyproasncyssergluproglucysprogly180185190asncyshisleuargglyargcysileaspglyglncysilecysasp195200205aspglyphethrglygluaspcysserglnleualacysproserasp210215220cysasnaspglnglylyscysvalasnglyvalcysilecyspheglu225230235240glytyralaglyalaaspcysserarggluilecysprovalprocys245250255serglugluhisglythrcysvalaspglyleucysvalcyshisasp260265270glyphealaglyaspaspcysasnlysproleucysleuasnasncys275280285tyrasnargglyargcysvalgluasnglucysvalcysaspglugly290295300phethrglygluaspcyssergluleuilecysproasnaspcysphe305310315320aspargglyargcysileasnglythrcystyrcysglugluglyphe325330335thrglygluaspcysglylysprothrcysprohisalacyshisthr340345350glnglyargcysglugluglyglncysvalcysaspgluglypheala355360365glyvalaspcysserglulysargcysproalaaspcyshisasnarg370375380glyargcysvalaspglyargcysglucysaspaspglyphethrgly385390395400alaaspcysglygluleulyscysproasnglycysserglyhisgly405410415argcysvalasnglyglncysvalcysaspgluglytyrthrglyglu420425430aspcysserglnleuargcysproasnaspcyshisserargglyarg435440445cysvalgluglylyscysvalcysgluglnglyphelysglytyrasp450455460cysseraspmetsercysproasnaspcyshisglnhisglyargcys465470475480valasnglymetcysvalcysaspaspglytyrthrglygluaspcys485490495argaspargglncysproargaspcysserasnargglyleucysval500505510aspglyglncysvalcysgluaspglyphethrglyproaspcysala515520525gluleusercysproasnaspcyshisglyglnglyargcysvalasn530535540glyglncysvalcyshisgluglyphemetglylysaspcyslysglu545550555560glnargcysproseraspcyshisglyglnglyargcysvalaspgly565570575glncysilecyshisgluglyphethrglyleuaspcysglyglnhis580585590sercysproseraspcysasnasnleuglyglncysvalserglyarg595600605cysilecysasngluglytyrserglygluaspcyssergluvalser610615620proprolysaspleuvalvalthrgluvalthrglugluthrvalasn625630635640leualatrpaspasnglumetargvalthrglutyrleuvalvaltyr645650655thrprothrhisgluglyglyleuglumetglnpheargvalprogly660665670aspglnthrserthrileileglngluleugluproglyvalglutyr675680685pheileargvalphealaileleugluasnlyslysserileproval690695700seralaargvalalathrtyrleuproalaprogluglyleulysphe705710715720lysserilelysgluthrservalgluvalglutrpaspproleuasp725730735ilealaphegluthrtrpgluileilepheargasnmetasnlysglu740745750aspgluglygluilethrlysserleuargargprogluthrsertyr755760765argglnthrglyleualaproglyglnglutyrgluileserleuhis770775780ilevallysasnasnthrargglyproglyleulysargvalthrthr785790795800thrargleuaspalaproserglnilegluvallysaspvalthrasp805810815thrthralaleuilethrtrpphelysproleualagluileaspgly820825830ilegluleuthrtyrglyilelysaspvalproglyaspargthrthr835840845ileaspleuthrgluaspgluasnglntyrserileglyasnleulys850855860proaspthrglutyrgluvalserleuileserargargglyaspmet865870875880serserasnproalalysgluthrphethrthrglyleuaspalapro885890895argasnleuargargvalserglnthraspasnserilethrleuglu900905910trpargasnglylysalaalaileaspsertyrargilelystyrala915920925proileserglyglyasphisalagluvalaspvalprolyssergln930935940glnalathrthrlysthrthrleuthrglyleuargproglythrglu945950955960tyrglyileglyvalseralavallysgluasplysgluserasnpro965970975alathrileasnalaalathrgluleuaspthrprolysaspleugln980985990valsergluthralagluthrserleuthrleuleutrplysthrpro99510001005leualalyspheaspargtyrargleuasntyrserleuprothr101010151020glyglntrpvalglyvalglnleuproargasnthrthrsertyr102510301035valleuargglyleugluproglyglnglutyrasnvalleuleu104010451050thralaglulysglyarghislysserlysproalaargvallys105510601065alaserthrgluglnalaprogluleugluasnleuthrvalthr107010751080gluvalglytrpaspglyleuargleuasntrpthralaalaasp108510901095glnalatyrgluhispheileileglnvalglnglualaasnlys110011051110valglualaalaargasnleuthrvalproglyserleuargala111511201125valaspileproglyleulysalaalathrprotyrthrvalser113011351140iletyrglyvalileglnglytyrargthrprovalleuserala114511501155glualaserthrglygluthrproasnleuglygluvalvalval116011651170alagluvalglytrpaspalaleulysleuasntrpthralapro117511801185gluglyalatyrglutyrphepheileglnvalglnglualaasp119011951200thrvalglualaalaglnasnleuthrvalproglyglyleuarg120512101215serthraspleuproglyleulysalaalathrhistyrthrile122012251230thrileargglyvalthrglnasppheserthrthrproleuser123512401245valgluvalleuthrglugluvalproaspmetglyasnleuthr125012551260valthrgluvalsertrpaspalaleuargleuasntrpthrthr126512701275proaspglythrtyraspglnphethrileglnvalglngluala128012851290aspglnvalgluglualahisasnleuthrvalproglyserleu129513001305argsermetgluileproglyleuargalaglythrprotyrthr131013151320valthrleuhisglygluvalargglyhisserthrargproleu132513301335alavalgluvalvalthrgluaspleuproglnleuglyaspleu134013451350alavalsergluvalglytrpaspglyleuargleuasntrpthr135513601365alaalaaspasnalatyrgluhisphevalileglnvalglnglu137013751380valasnlysvalglualaalaglnasnleuthrleuproglyser138513901395leuargalavalaspileproglyleuglualaalathrprotyr140014051410argvalseriletyrglyvalileargglytyrargthrproval141514201425leuseralaglualaserthralalysgluprogluileglyasn143014351440leuasnvalseraspilethrprogluserpheasnleusertrp144514501455metalathraspglyilephegluthrphethrilegluileile146014651470aspserasnargleuleugluthrvalglutyrasnilesergly147514801485alagluargthralahisileserglyleuproproserthrasp149014951500pheilevaltyrleuserglyleualaproserileargthrlys150515101515thrileseralathralathrthrglualaleuproleuleuglu152015251530asnleuthrileseraspileasnprotyrglyphethrvalser153515401545trpmetalasergluasnalapheaspserpheleuvalthrval155015551560valaspserglylysleuleuaspproglngluphethrleuser156515701575glythrglnarglysleugluleuargglyleuilethrglyile158015851590glytyrgluvalmetvalserglyphethrglnglyhisglnthr159516001605lysproleuargalagluilevalthrglualagluprogluval161016151620aspasnleuleuvalseraspalathrproaspglypheargleu162516301635sertrpthralaaspgluglyvalpheaspasnphevalleulys164016451650ileargaspthrlyslysglnsergluproleugluilethrleu165516601665leualaprogluargthrargaspilethrglyleuarggluala167016751680thrglutyrgluilegluleutyrglyileserlysglyargarg168516901695serglnthrvalseralailealathrthralametglyserpro170017051710lysgluvalilepheseraspilethrgluasnseralathrval171517201725sertrpargalaprothralaglnvalgluserpheargilethr173017351740tyrvalproilethrglyglythrprosermetvalthrvalasp174517501755glythrlysthrglnthrargleuvallysleuileproglyval176017651770glutyrleuvalserileilealametlysglypheglugluser177517801785gluprovalserglyserphethrthralaleuaspglyproser179017951800glyleuvalthralaasnilethraspserglualaleualaarg180518101815trpglnproalailealathrvalaspsertyrvalilesertyr182018251830thrglyglulysvalprogluilethrargthrvalserglyasn183518401845thrvalglutyralaleuthraspleugluproalathrglutyr185018551860thrleuargilephealaglulysglyproglnlysserserthr186518701875ilethralalysphethrthraspleuaspserproargaspleu188018851890thralathrgluvalglnsergluthralaleuleuthrtrparg189519001905proproargalaservalthrglytyrleuleuvaltyrgluser191019151920valaspglythrvallysgluvalilevalglyproaspthrthr192519301935sertyrserleualaaspleuserproserthrhistyrthrala194019451950lysileglnalaleuasnglyproleuargserasnmetilegln195519601965thrilephethrthrileglyleuleutyrpropheprolysasp197019751980cysserglnalametleuasnglyaspthrthrserglyleutyr198519901995thriletyrleuasnglyasplysalaglualaleugluvalphe200020052010cysaspmetthrseraspglyglyglytrpilevalpheleuarg201520202025arglysasnglyarggluasnphetyrglnasntrplysalatyr203020352040alaalaglypheglyaspargargglugluphetrpleuglyleu204520502055aspasnleuasnlysilethralaglnglyglntyrgluleuarg206020652070valaspleuargasphisglygluthralaphealavaltyrasp207520802085lyspheservalglyaspalalysthrargtyrlysleulysval209020952100gluglytyrserglythralaglyaspsermetalatyrhisasn210521102115glyargserpheserthrpheasplysaspthraspseralaile212021252130thrasncysalaleusertyrlysglyalaphetrptyrargasn213521402145cyshisargvalasnleumetglyargtyrglyaspasnasnhis215021552160serglnglyvalasntrpphehistrplysglyhisgluhisser216521702175ileglnphealaglumetlysleuargproserasnpheargasn218021852190leugluglyargarglysargala21952200<210>76<211>89<212>prt<213>人工序列<220><223>fibconfn3域<400>76leuaspalaprothraspleuglnvalthrasnvalthraspthrser151015ilethrvalsertrpthrproproseralathrilethrglytyrarg202530ilethrtyrthrproserasnglyproglygluprolysgluleuthr354045valproproserserthrservalthrilethrglyleuthrprogly505560valglutyrvalvalserleutyralaleulysaspasnglngluser65707580proproleuvalglythrglnthrthr85<210>77<211>94<212>prt<213>智人<400>77valseraspvalproargaspleugluvalvalalaalathrprothr151015serleuleuilesertrpaspalaproalavalthrvalargtyrtyr202530argilethrtyrglygluthrglyglyasnserprovalglngluphe354045thrvalproglyserlysserthralathrileserglyleulyspro505560glyvalasptyrthrilethrvaltyralavalthrglyargglyasp65707580serproalaserserlysproileserileasntyrargthr8590<210>78<211>4<212>prt<213>人工序列<220><223>接头<400>78glyserglyser1<210>79<211>25<212>prt<213>人工序列<220><223>接头<400>79glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyserglyglyglyglyser2025<210>80<211>4<212>prt<213>人工序列<220><223>接头<400>80alaproalapro1<210>81<211>10<212>prt<213>人工序列<220><223>接头<400>81alaproalaproalaproalaproalapro1510<210>82<211>20<212>prt<213>人工序列<220><223>接头<400>82alaproalaproalaproalaproalaproalaproalaproalapro151015alaproalapro20<210>83<211>40<212>prt<213>人工序列<220><223>接头<400>83alaproalaproalaproalaproalaproalaproalaproalapro151015alaproalaproalaproalaproalaproalaproalaproalapro202530alaproalaproalaproalapro3540<210>84<211>29<212>prt<213>人工序列<220><223>接头<400>84alaglualaalaalalysglualaalaalalysglualaalaalalys151015glualaalaalalysglualaalaalalysalaalaala2025<210>85<211>8<212>prt<213>人工序列<220><223>tenconbc环<400>85thralaproaspalaalapheasp15<210>86<211>7<212>prt<213>人工序列<220><223>tenconfg环<400>86lysglyglyhisargserasn15<210>87<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>87alaaspprohisglyphetyrasp15<210>88<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>88thrtyraspargaspglytyrasp15<210>89<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>89trpasppropheserphetyrasp15<210>90<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>90aspaspproargglyphetyrglu15<210>91<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>91thrtrpprotyralaaspleuasp15<210>92<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>92glytyrasnglyasphispheasp15<210>93<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>93asptyraspleuglyvaltyrasp15<210>94<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域bc环<400>94aspaspprotrpaspphetyrglu15<210>95<211>12<212>prt<213>人工序列<220><223>结合egfr的fn3域fg环<400>95hisasnvaltyrlysaspthrasnmetargglyleu1510<210>96<211>11<212>prt<213>人工序列<220><223>结合egfr的fn3域fg环<400>96leuglysertyrvalphegluhisaspvalmet1510<210>97<211>285<212>dna<213>人工序列<220><223>来自ecb97;p54ar4-83v22的结合egfr的fn3域<400>97atgttgccagcgccgaagaacctggtagttagcgaggttactgaggacagcgcgcgtctg60agctgggacgatccgtgggcgttctacgagagctttctgatccagtatcaagagagcgag120aaagtcggtgaagcgattgtgctgaccgtcccgggctccgagcgttcctacgacctgacc180ggtttgaagccgggtaccgagtatacggtgagcatctacggtgttcacaatgtctataag240gacactaatatccgcggtctgcctctgagcgccattttcaccacc285<210>98<211>285<212>dna<213>人工序列<220><223>来自ecb15;p54ar4-83v2的结合egfr的fn3域<400>98atgctgccagcccctaagaatctggtcgtgagcgaagtaaccgaggacagcgcccgcctg60agctgggacgacccgtgggcgttctatgagtctttcctgattcagtatcaagaaagcgaa120aaagttggcgaagcgatcgtcctgaccgtcccgggtagcgagcgctcctacgatctgacc180ggcctgaaaccgggtacggagtacacggtgtccatttacggtgttcacaatgtgtataaa240gacaccaacatgcgtggcctgccgctgtcggcgattttcaccacc285<210>99<211>89<212>prt<213>人工序列<220><223>tencon27fn3域<400>99leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvallysglyglyhisargser65707580asnproleuseralailephethrthr85<210>100<211>89<212>prt<213>人工序列<220><223>tcl14文库<220><221>尚未归类的特性<222>(32)..(32)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(34)..(34)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(36)..(36)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(38)..(38)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(39)..(39)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(40)..(40)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(41)..(41)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(68)..(68)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(70)..(70)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(72)..(72)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(78)..(78)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(79)..(79)<223>xaa为任何氨基酸<220><221>尚未归类的特性<222>(81)..(81)<223>xaa为任何氨基酸<400>100leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphexaa202530ilexaatyrxaagluxaaxaaxaaxaaglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrxaavalxaailexaaglyvallysglyglyxaaxaaser65707580xaaproleuseralailephethrthr85<210>101<211>1408<212>prt<213>智人<400>101metlysalaproalavalleualaproglyileleuvalleuleuphe151015thrleuvalglnargserasnglyglucyslysglualaleualalys202530serglumetasnvalasnmetlystyrglnleuproasnphethrala354045gluthrproileglnasnvalileleuhisgluhishisilepheleu505560glyalathrasntyriletyrvalleuasnglugluaspleuglnlys65707580valalaglutyrlysthrglyprovalleugluhisproaspcysphe859095procysglnaspcysserserlysalaasnleuserglyglyvaltrp100105110lysaspasnileasnmetalaleuvalvalaspthrtyrtyraspasp115120125glnleuilesercysglyservalasnargglythrcysglnarghis130135140valpheprohisasnhisthralaaspileglnsergluvalhiscys145150155160ilepheserproglnileglugluproserglncysproaspcysval165170175valseralaleuglyalalysvalleuserservallysaspargphe180185190ileasnphephevalglyasnthrileasnsersertyrpheproasp195200205hisproleuhisserileservalargargleulysgluthrlysasp210215220glyphemetpheleuthraspglnsertyrileaspvalleuproglu225230235240pheargaspsertyrproilelystyrvalhisalaphegluserasn245250255asnpheiletyrpheleuthrvalglnarggluthrleuaspalagln260265270thrphehisthrargileileargphecysserileasnserglyleu275280285hissertyrmetglumetproleuglucysileleuthrglulysarg290295300lyslysargserthrlyslysgluvalpheasnileleuglnalaala305310315320tyrvalserlysproglyalaglnleualaargglnileglyalaser325330335leuasnaspaspileleupheglyvalphealaglnserlysproasp340345350seralagluprometaspargseralametcysalapheproilelys355360365tyrvalasnaspphepheasnlysilevalasnlysasnasnvalarg370375380cysleuglnhisphetyrglyproasnhisgluhiscyspheasnarg385390395400thrleuleuargasnserserglycysglualaargargaspglutyr405410415argthrgluphethrthralaleuglnargvalaspleuphemetgly420425430glnphesergluvalleuleuthrserileserthrpheilelysgly435440445aspleuthrilealaasnleuglythrsergluglyargphemetgln450455460valvalvalserargserglyproserthrprohisvalasnpheleu465470475480leuaspserhisprovalserprogluvalilevalgluhisthrleu485490495asnglnasnglytyrthrleuvalilethrglylyslysilethrlys500505510ileproleuasnglyleuglycysarghispheglnsercyssergln515520525cysleuseralaproprophevalglncysglytrpcyshisasplys530535540cysvalargsergluglucysleuserglythrtrpthrglnglnile545550555560cysleuproalailetyrlysvalpheproasnseralaproleuglu565570575glyglythrargleuthrilecysglytrpasppheglypheargarg580585590asnasnlyspheaspleulyslysthrargvalleuleuglyasnglu595600605sercysthrleuthrleusergluserthrmetasnthrleulyscys610615620thrvalglyproalametasnlyshispheasnmetserileileile625630635640serasnglyhisglythrthrglntyrserthrphesertyrvalasp645650655provalilethrserileserprolystyrglyprometalaglygly660665670thrleuleuthrleuthrglyasntyrleuasnserglyasnserarg675680685hisileserileglyglylysthrcysthrleulysservalserasn690695700serileleuglucystyrthrproalaglnthrileserthrgluphe705710715720alavallysleulysileaspleualaasnarggluthrserilephe725730735sertyrarggluaspproilevaltyrgluilehisprothrlysser740745750pheileserthrtrptrplysgluproleuasnilevalserpheleu755760765phecysphealaserglyglyserthrilethrglyvalglylysasn770775780leuasnservalservalproargmetvalileasnvalhisgluala785790795800glyargasnphethrvalalacysglnhisargserasnsergluile805810815ilecyscysthrthrproserleuglnglnleuasnleuglnleupro820825830leulysthrlysalaphephemetleuaspglyileleuserlystyr835840845pheaspleuiletyrvalhisasnprovalphelyspropheglulys850855860provalmetilesermetglyasngluasnvalleugluilelysgly865870875880asnaspileaspproglualavallysglygluvalleulysvalgly885890895asnlyssercysgluasnilehisleuhisserglualavalleucys900905910thrvalproasnaspleuleulysleuasnsergluleuasnileglu915920925trplysglnalaileserserthrvalleuglylysvalilevalgln930935940proaspglnasnphethrglyleuilealaglyvalvalserileser945950955960thralaleuleuleuleuleuglyphepheleutrpleulyslysarg965970975lysglnilelysaspleuglysergluleuvalargtyraspalaarg980985990valhisthrprohisleuaspargleuvalseralaargservalser99510001005prothrthrglumetvalserasngluservalasptyrargala101010151020thrpheprogluaspglnpheproasnserserglnasnglyser102510301035cysargglnvalglntyrproleuthraspmetserproileleu104010451050thrserglyaspseraspileserserproleuleuglnasnthr105510601065valhisileaspleuseralaleuasnprogluleuvalglnala107010751080valglnhisvalvalileglyproserserleuilevalhisphe108510901095asngluvalileglyargglyhispheglycysvaltyrhisgly110011051110thrleuleuaspasnaspglylyslysilehiscysalavallys111511201125serleuasnargilethraspileglygluvalserglnpheleu113011351140thrgluglyileilemetlysasppheserhisproasnvalleu114511501155serleuleuglyilecysleuargsergluglyserproleuval116011651170valleuprotyrmetlyshisglyaspleuargasnpheilearg117511801185asngluthrhisasnprothrvallysaspleuileglyphegly119011951200leuglnvalalalysglymetlystyrleualaserlyslysphe120512101215valhisargaspleualaalaargasncysmetleuaspglulys122012251230phethrvallysvalalaasppheglyleualaargaspmettyr123512401245asplysglutyrtyrservalhisasnlysthrglyalalysleu125012551260provallystrpmetalaleugluserleuglnthrglnlysphe126512701275thrthrlysseraspvaltrpserpheglyvalleuleutrpglu128012851290leumetthrargglyalaproprotyrproaspvalasnthrphe129513001305aspilethrvaltyrleuleuglnglyargargleuleuglnpro131013151320glutyrcysproaspproleutyrgluvalmetleulyscystrp132513301335hisprolysalaglumetargproserphesergluleuvalser134013451350argileseralailepheserthrpheileglygluhistyrval135513601365hisvalasnalathrtyrvalasnvallyscysvalalaprotyr137013751380proserleuleusersergluaspasnalaaspaspgluvalasp138513901395thrargproalaserphetrpgluthrser14001405<210>102<211>697<212>prt<213>智人<400>102glnarglysargargasnthrilehisgluphelyslysseralalys151015thrthrleuilelysileaspproalaleulysilelysthrlyslys202530valasnthralaaspglncysalaasnargcysthrargasnlysgly354045leuprophethrcyslysalaphevalpheasplysalaarglysgln505560cysleutrpphepropheasnsermetserserglyvallyslysglu65707580pheglyhisglupheaspleutyrgluasnlysasptyrileargasn859095cysileileglylysglyargsertyrlysglythrvalserilethr100105110lysserglyilelyscysglnprotrpsersermetileprohisglu115120125hisserpheleuprosersertyrargglylysaspleuglngluasn130135140tyrcysargasnproargglyglugluglyglyprotrpcysphethr145150155160serasnprogluvalargtyrgluvalcysaspileproglncysser165170175gluvalglucysmetthrcysasnglyglusertyrargglyleumet180185190asphisthrgluserglylysilecysglnargtrpasphisglnthr195200205prohisarghislyspheleuprogluargtyrproasplysglyphe210215220aspaspasntyrcysargasnproaspglyglnproargprotrpcys225230235240tyrthrleuaspprohisthrargtrpglutyrcysalailelysthr245250255cysalaaspasnthrmetasnaspthraspvalproleugluthrthr260265270glucysileglnglyglnglygluglytyrargglythrvalasnthr275280285iletrpasnglyileprocysglnargtrpaspserglntyrprohis290295300gluhisaspmetthrprogluasnphelyscyslysaspleuargglu305310315320asntyrcysargasnproaspglysergluserprotrpcysphethr325330335thraspproasnileargvalglytyrcysserglnileproasncys340345350aspmetserhisglyglnaspcystyrargglyasnglylysasntyr355360365metglyasnleuserglnthrargserglyleuthrcyssermettrp370375380asplysasnmetgluaspleuhisarghisilephetrpgluproasp385390395400alaserlysleuasngluasntyrcysargasnproaspaspaspala405410415hisglyprotrpcystyrthrglyasnproleuileprotrpasptyr420425430cysproileserargcysgluglyaspthrthrprothrilevalasn435440445leuasphisprovalilesercysalalysthrlysglnleuargval450455460valasnglyileprothrargthrasnileglytrpmetvalserleu465470475480argtyrargasnlyshisilecysglyglyserleuilelysgluser485490495trpvalleuthralaargglncyspheproserargaspleulysasp500505510tyrglualatrpleuglyilehisaspvalhisglyargglyaspglu515520525lyscyslysglnvalleuasnvalserglnleuvaltyrglyproglu530535540glyseraspleuvalleumetlysleualaargproalavalleuasp545550555560aspphevalserthrileaspleuproasntyrglycysthrilepro565570575glulysthrsercysservaltyrglytrpglytyrthrglyleuile580585590asntyraspglyleuleuargvalalahisleutyrilemetglyasn595600605glulyscysserglnhishisargglylysvalthrleuasngluser610615620gluilecysalaglyalaglulysileglyserglyprocysglugly625630635640asptyrglyglyproleuvalcysgluglnhislysmetargmetval645650655leuglyvalilevalproglyargglycysalaileproasnargpro660665670glyilephevalargvalalatyrtyralalystrpilehislysile675680685ileleuthrtyrlysvalproglnser690695<210>103<211>276<212>dna<213>人工序列<220><223>结合c-met的fn3域<400>103ctgccggctccgaagaacttggtggtgagccgtgttaccgaagatagcgcacgcctgagc60tggacggcaccggatgcggcgttcgatagcttctggattcgctattttgagtttctgggt120agcggtgaggcaattgttctgacggtgccgggctctgaacgctcctacgatttgaccggt180ctgaaaccgggcaccgagtatgtggtgaacattctgagcgttaagggcggtagcatcagc240ccaccgctgagcgcgatcttcacgactggtggttgc276<210>104<211>267<212>dna<213>人工序列<220><223>结合c-met的fn3域<400>104ctgccggcaccgaagaacctggttgtcagccgtgtgaccgaggatagcgcacgtttgagc60tggaccgctccggatgcagcctttgacagcttctggattcgttactttgaatttctgggt120agcggtgaggcgatcgttctgacggtgccgggctctgaacgcagctatgatttgacgggc180ctgaagccgggtactgagtacgtggttaacatcatgggcgttaagggtggtaaaatcagc240ccgccattgtccgcgatctttaccacg267<210>105<211>5<212>prt<213>人工序列<220><223>接头<400>105glyglyglyglyser15<210>106<211>194<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>106metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthr<210>107<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>107leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthr8590<210>108<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>108leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthr8590<210>109<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>109leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprohisalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthr8590<210>110<211>94<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>110leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthr8590<210>111<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>111leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnileleuglyvallysglyglylysileser65707580proproleuseralailephethrthr85<210>112<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>112leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphegluphevalglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnileleuglyvallysglyglyserileser65707580proproleuseralailephethrthr85<210>113<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>113leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphegluphevalserlysglyaspalailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnileleuglyvallysglyglyserileser65707580proproleuseralailephethrthr85<210>114<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域<400>114leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrpheglupheleuglyserglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnileleuservallysglyglyserileser65707580proproleuseralailephethrthr85<210>115<211>591<212>dna<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>115atgttgccagcgccgaagaacctggtagttagcgaggttactgaggacagcgcgcgtctg60agctgggacgatccgtgggcgttctacgagagctttctgatccagtatcaagagagcgag120aaagtcggtgaagcgattgtgctgaccgtcccgggctccgagcgttcctacgacctgacc180ggtttgaagccgggtaccgagtatacggtgagcatctacggtgttcacaatgtctataag240gacactaatatccgcggtctgcctctgagcgccattttcaccaccgcaccggcaccggct300ccggctcctgccccgctgccggctccgaagaacttggtggtgagccgtgttaccgaagat360agcgcacgcctgagctggacggcaccggatgcggcgttcgatagcttctggattcgctat420tttgagtttctgggtagcggtgaggcaattgttctgacggtgccgggctctgaacgctcc480tacgatttgaccggtctgaaaccgggcaccgagtatgtggtgaacattctgagcgttaag540ggcggtagcatcagcccaccgctgagcgcgatcttcacgactggtggttgc591<210>116<211>582<212>dna<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>116atgctgccagcccctaagaatctggtcgtgagcgaagtaaccgaggacagcgcccgcctg60agctgggacgacccgtgggcgttctatgagtctttcctgattcagtatcaagaaagcgaa120aaagttggcgaagcgatcgtcctgaccgtcccgggtagcgagcgctcctacgatctgacc180ggcctgaaaccgggtacggagtacacggtgtccatttacggtgttcacaatgtgtataaa240gacaccaacatgcgtggcctgccgctgtcggcgattttcaccaccgcgcctgcgccagcg300cctgcaccggctccgctgccggcaccgaagaacctggttgtcagccgtgtgaccgaggat360agcgcacgtttgagctggaccgctccggatgcagcctttgacagcttctggattcgttac420tttgaatttctgggtagcggtgaggcgatcgttctgacggtgccgggctctgaacgcagc480tatgatttgacgggcctgaagccgggtactgagtacgtggttaacatcatgggcgttaag540ggtggtaaaatcagcccgccattgtccgcgatctttaccacg582<210>117<211>51<212>prt<213>人工序列<220><223>白蛋白结合域<400>117thrileaspglutrpleuleulysglualalysglulysalaileglu151015gluleulyslysalaglyilethrserasptyrtyrpheaspleuile202530asnlysalalysthrvalgluglyvalasnalaleulysaspgluile354045leulysala50<210>118<211>250<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>118metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyraspgluvalval130135140valglyglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrtyrvalasnile165170175leuglyvallysglyglyserileservalproleuseralailephe180185190thrthralaproalaproalaproalaproalaproleualagluala195200205lysvalleualaasnarggluleuasplystyrglyvalserasptyr210215220tyrlysasnleuileasnasnalalysthrvalgluglyvallysala225230235240leuleuaspgluileleualaalaleupro245250<210>119<211>250<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>119metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyraspgluvalval130135140valglyglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrtyrvalasnile165170175leuglyvallysglyglyserileservalproleuseralailephe180185190thrthralaproalaproalaproalaproalaproleualagluala195200205lysvalleualaasnarggluleuasplystyrglyvalserasptyr210215220tyrlysasnleuileasnasnalalysthrvalgluglyvallysala225230235240leuleuaspgluileleualaalaleupro245250<210>120<211>250<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>120metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175metglyvallysglyglylysileserproproleuseralailephe180185190thrthralaproalaproalaproalaproalaproleualagluala195200205lysvalleualaasnarggluleuasplystyrglyvalserasptyr210215220tyrlysasnleuileasnasnalalysthrvalgluglyvallysala225230235240leuleuaspgluileleualaalaleupro245250<210>121<211>250<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>121metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175metglyvallysglyglylysileserproproleuseralailephe180185190thrthralaproalaproalaproalaproalaproleualagluala195200205lysvalleualaasnarggluleuasplystyrglyvalserasptyr210215220tyrlysasnleuileasnasnalalysthrvalgluglyvallysala225230235240leuleuaspgluileleualaalaleupro245250<210>122<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>122metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>123<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>123metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpthrtyraspargaspglytyraspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>124<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>124metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpglytyrasnglyasphispheaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>125<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>125metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpaspaspproargglyphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>126<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>126metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpthrtrpprotyralaaspleuaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>127<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>127metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpglytyrasnglyasphispheaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>128<211>96<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>128metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpasptyraspleuglyvaltyrpheaspser202530pheleuileglntyrglngluserglulysvalglyglualaileasn354045leuthrvalproglysergluargsertyraspleuthrglyleulys505560proglythrglutyrthrvalseriletyrglyvalhisasnvaltyr65707580lysaspthrasnmetargglyleuproleuseralagluphethrthr859095<210>129<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>129metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>130<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>130metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpthralaproaspalaalapheaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalleuglysertyrval65707580phegluhisaspvalmetleuproleuseralagluphethrthr859095<210>131<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>131metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthr859095<210>132<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>132metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpthralaproaspalaalapheaspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalleuglysertyrval65707580phegluhisaspvalmetleuproleuseralailephethrthr859095<210>133<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>133metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015serleuargleusertrpthrtrpprotyralaaspleuaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralagluphethrthr859095<210>134<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>134metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnmetargglyleuproleuseralailephethrthr859095<210>135<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>135metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthr859095<210>136<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>136metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthr859095<210>137<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域<400>137metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpalaaspprohisglyphetyraspserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthr859095<210>138<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>138leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyraspgluvalvalvalglyglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrtyrvalasnileleuglyvallysglygly180185190serileservalproleuseralailephethrthr195200<210>139<211>203<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>139leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glyserleuproalaprolysasnleuvalvalserargvalthrglu115120125aspseralaargleusertrpthralaproaspalaalapheaspser130135140phetrpileargtyrpheglupheleuglyserglyglualaileval145150155160leuthrvalproglysergluargsertyraspleuthrglyleulys165170175proglythrglutyrvalvalasnilemetglyvallysglyglylys180185190ileserproproleuseralailephethrthr195200<210>140<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>140leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyrpheglupheleuglyserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrvalvalglnileileglyvallysglygly180185190hisileserleuproleuseralailephethrthr195200<210>141<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>141leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphepheileargtyraspglupheleuargserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrtrpvalthrileleuglyvallysglygly180185190leuvalserthrproleuseralailephethrthr195200<210>142<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>142leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyrpheglupheleuglyserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrvalvalasnilemetglyvallysglygly180185190lysileserproproleuseralailephethrthr195200<210>143<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>143leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyrpheglupheleuglyserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrvalvalglnileileglyvallysglygly180185190hisileserleuproleuseralailephethrthr195200<210>144<211>204<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>144leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthrglygly859095glyglyserglyglyglyglyserglyglyglyglyserglyglygly100105110glysermetleuproalaprolysasnleuvalvalserargvalthr115120125gluaspseralaargleusertrpthralaproaspalaalapheasp130135140serphetrpileargtyrpheglupheleuglyserglyglualaile145150155160valleuthrvalproglysergluargsertyraspleuthrglyleu165170175lysproglythrglutyrvalvalglnileileglyvallysglygly180185190hisileserleuproleuseralailephethrthr195200<210>145<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>145leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnilemet165170175glyvallysglyglylysileserproproleuseralailephethr180185190thr<210>146<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>146leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyraspgluvalvalval130135140glyglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrtyrvalasnileleu165170175glyvallysglyglyserileservalproleuseralailephethr180185190thr<210>147<211>194<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>147leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaprometleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175metglyvallysglyglylysileserproproleuseralailephe180185190thrthr<210>148<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>148leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyraspgluvalvalval130135140glyglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrtyrvalasnileleu165170175glyvallysglyglyserileservalproleuseralailephethr180185190thr<210>149<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>149leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglylysileserproproleuseralailephethr180185190thr<210>150<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>150leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalgly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>151<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>151leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalser130135140lysglyaspalailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>152<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>152leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175servallysglyglyserileserproproleuseralailephethr180185190thr<210>153<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>153leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprohisalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglylysileserproproleuseralailephethr180185190thr<210>154<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>154leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprohisalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalgly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>155<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>155leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprohisalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalser130135140lysglyaspalailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>156<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>156leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprohisalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175servallysglyglyserileserproproleuseralailephethr180185190thr<210>157<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>157leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglylysileserproproleuseralailephethr180185190thr<210>158<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>158leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalgly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>159<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>159leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrphegluphevalser130135140lysglyaspalailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175glyvallysglyglyserileserproproleuseralailephethr180185190thr<210>160<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>160leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175servallysglyglyserileserproproleuseralailephethr180185190thr<210>161<211>193<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>161leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnileargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnileleu165170175servallysglyglyserileserproproleuseralailephethr180185190thr<210>162<211>249<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>162leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyraspgluvalvalval130135140glyglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrtyrvalasnileleu165170175glyvallysglyglyserileservalproleuseralailephethr180185190thralaproalaproalaproalaproalaproleualaglualalys195200205valleualaasnarggluleuasplystyrglyvalserasptyrtyr210215220lysasnleuileasnasnalalysthrvalgluglyvallysalaleu225230235240leuaspgluileleualaalaleupro245<210>163<211>249<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>163leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyraspgluvalvalval130135140glyglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrtyrvalasnileleu165170175glyvallysglyglyserileservalproleuseralailephethr180185190thralaproalaproalaproalaproalaproleualaglualalys195200205valleualaasnarggluleuasplystyrglyvalserasptyrtyr210215220lysasnleuileasnasnalalysthrvalgluglyvallysalaleu225230235240leuaspgluileleualaalaleupro245<210>164<211>249<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>164leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpaspaspprotrpalaphetyrgluserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnilemet165170175glyvallysglyglylysileserproproleuseralailephethr180185190thralaproalaproalaproalaproalaproleualaglualalys195200205valleualaasnarggluleuasplystyrglyvalserasptyrtyr210215220lysasnleuileasnasnalalysthrvalgluglyvallysalaleu225230235240leuaspgluileleualaalaleupro245<210>165<211>249<212>prt<213>人工序列<220><223>双特异性egfr/c-met结合域<400>165leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015alaargleusertrpalaaspprohisglyphetyraspserpheleu202530ileglntyrglngluserglulysvalglyglualailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrglyvalhisasnvaltyrlysasp65707580thrasnmetargglyleuproleuseralailephethrthralapro859095alaproalaproalaproalaproleuproalaprolysasnleuval100105110valserargvalthrgluaspseralaargleusertrpthralapro115120125aspalaalapheaspserphetrpileargtyrpheglupheleugly130135140serglyglualailevalleuthrvalproglysergluargsertyr145150155160aspleuthrglyleulysproglythrglutyrvalvalasnilemet165170175glyvallysglyglylysileserproproleuseralailephethr180185190thralaproalaproalaproalaproalaproleualaglualalys195200205valleualaasnarggluleuasplystyrglyvalserasptyrtyr210215220lysasnleuileasnasnalalysthrvalgluglyvallysalaleu225230235240leuaspgluileleualaalaleupro245<210>166<211>588<212>dna<213>人工序列<220><223>双特异性egfr/c-met结合域<400>166ttgccagcgccgaagaacctggtagttagcgaggttactgaggacagcgcgcgtctgagc60tgggacgatccgtgggcgttctacgagagctttctgatccagtatcaagagagcgagaaa120gtcggtgaagcgattgtgctgaccgtcccgggctccgagcgttcctacgacctgaccggt180ttgaagccgggtaccgagtatacggtgagcatctacggtgttcacaatgtctataaggac240actaatatccgcggtctgcctctgagcgccattttcaccaccgcaccggcaccggctccg300gctcctgccccgctgccggctccgaagaacttggtggtgagccgtgttaccgaagatagc360gcacgcctgagctggacggcaccggatgcggcgttcgatagcttctggattcgctatttt420gagtttctgggtagcggtgaggcaattgttctgacggtgccgggctctgaacgctcctac480gatttgaccggtctgaaaccgggcaccgagtatgtggtgaacattctgagcgttaagggc540ggtagcatcagcccaccgctgagcgcgatcttcacgactggtggttgc588<210>167<211>579<212>dna<213>人工序列<220><223>双特异性egfr/c-met结合域<400>167ctgccagcccctaagaatctggtcgtgagcgaagtaaccgaggacagcgcccgcctgagc60tgggacgacccgtgggcgttctatgagtctttcctgattcagtatcaagaaagcgaaaaa120gttggcgaagcgatcgtcctgaccgtcccgggtagcgagcgctcctacgatctgaccggc180ctgaaaccgggtacggagtacacggtgtccatttacggtgttcacaatgtgtataaagac240accaacatgcgtggcctgccgctgtcggcgattttcaccaccgcgcctgcgccagcgcct300gcaccggctccgctgccggcaccgaagaacctggttgtcagccgtgtgaccgaggatagc360gcacgtttgagctggaccgctccggatgcagcctttgacagcttctggattcgttacttt420gaatttctgggtagcggtgaggcgatcgttctgacggtgccgggctctgaacgcagctat480gatttgacgggcctgaagccgggtactgagtacgtggttaacatcatgggcgttaagggt540ggtaaaatcagcccgccattgtccgcgatctttaccacg579<210>168<211>282<212>dna<213>人工序列<220><223>来自ecb97的结合egfr的fn3域<400>168ttgccagcgccgaagaacctggtagttagcgaggttactgaggacagcgcgcgtctgagc60tgggacgatccgtgggcgttctacgagagctttctgatccagtatcaagagagcgagaaa120gtcggtgaagcgattgtgctgaccgtcccgggctccgagcgttcctacgacctgaccggt180ttgaagccgggtaccgagtatacggtgagcatctacggtgttcacaatgtctataaggac240actaatatccgcggtctgcctctgagcgccattttcaccacc282<210>169<211>282<212>dna<213>人工序列<220><223>来自ecb15的结合egfr的fn3域<400>169ctgccagcccctaagaatctggtcgtgagcgaagtaaccgaggacagcgcccgcctgagc60tgggacgacccgtgggcgttctatgagtctttcctgattcagtatcaagaaagcgaaaaa120gttggcgaagcgatcgtcctgaccgtcccgggtagcgagcgctcctacgatctgaccggc180ctgaaaccgggtacggagtacacggtgtccatttacggtgttcacaatgtgtataaagac240accaacatgcgtggcctgccgctgtcggcgattttcaccacc282<210>170<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>170metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglylysileserproproleuseralailephe180185190thrthrcys195<210>171<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>171metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>172<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>172metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140serlysglyaspalailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>173<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>173metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>174<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>174metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglylysileserproproleuseralailephe180185190thrthrcys195<210>175<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>175metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>176<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>176metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheval130135140serlysglyaspalailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuglyvallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>177<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>177metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprohisalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>178<211>195<212>prt<213>人工序列<220><223>结合egfr/c-met的双特异性分子<400>178metleuproalaprolysasnleuvalvalsergluvalthrgluasp151015seralaargleusertrpaspaspprotrpalaphetyrgluserphe202530leuileglntyrglngluserglulysvalglyglualailevalleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnileargglyleuproleuseralailephethrthrala859095proalaproalaproalaproalaproleuproalaprolysasnleu100105110valvalserargvalthrgluaspseralaargleusertrpthrala115120125proaspalaalapheaspserphetrpileargtyrpheglupheleu130135140glyserglyglualailevalleuthrvalproglysergluargser145150155160tyraspleuthrglyleulysproglythrglutyrvalvalasnile165170175leuservallysglyglyserileserproproleuseralailephe180185190thrthrcys195<210>179<211>12<212>prt<213>人工序列<220><223>结合egfr的fn3域的共有fg环<220><221>变体<222>(9)..(9)<223>xaa可为met或ile<400>179hisasnvaltyrlysaspthrasnxaaargglyleu1510<210>180<211>12<212>prt<213>人工序列<220><223>结合egfr的fn3域的gf环<400>180leuglysertyrvalphegluhisaspvalmetleu1510<210>181<211>8<212>prt<213>人工序列<220><223>结合egfr的fn3域的共有bc环<220><221>变体<222>(1)..(1)<223>xaa可为ala、thr、gly或asp<220><221>变体<222>(2)..(2)<223>xaa可为ala、asp、tyr或trp<220><221>变体<222>(3)..(3)<223>xaa可为pro、asp或asn<220><221>变体<222>(4)..(4)<223>xaa可为leu或不存在<220><221>变体<222>(5)..(5)<223>xaa可为asp、his、arg、gly、tyr或trp<220><221>变体<222>(6)..(6)<223>xaa可为glu、asp或ala<220><221>变体<222>(7)..(7)<223>xaa可为ala、phe、gly、his或asp<220><221>变体<222>(8)..(8)<223>xaa可为tyr、phe或leu<400>181xaaxaaxaaxaaxaaxaaxaaxaa15<210>182<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域共有序列<220><221>变体<222>(22)..(22)<223>xaa可为ala、thr、gly或asp<220><221>变体<222>(23)..(23)<223>xaa可为ala、asp、tyr或trp<220><221>变体<222>(24)..(24)<223>xaa可为pro、asp或asn<220><221>变体<222>(25)..(25)<223>xaa可为leu或不存在<220><221>变体<222>(26)..(26)<223>xaa可为asp、his、arg、gly、tyr或trp<220><221>变体<222>(27)..(27)<223>xaa可为gly、asp或ala<220><221>变体<222>(28)..(28)<223>xaa可为ala、phe、gly、his或asp<220><221>变体<222>(29)..(29)<223>xaa可为tyr、phe或leu<220><221>变体<222>(84)..(84)<223>xaa可为met或ile<400>182leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpxaaxaaxaaxaaxaaxaaxaaxaaaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalhisasnvaltyrlys65707580aspthrasnxaaargglyleuproleuseralagluphethrthr859095<210>183<211>95<212>prt<213>人工序列<220><223>结合egfr的fn3域共有序列<220><221>变体<222>(22)..(22)<223>xaa可为ala、thr、gly或asp<220><221>变体<222>(23)..(23)<223>xaa可为ala、asp、tyr或trp<220><221>变体<222>(24)..(24)<223>xaa可为pro、asp或asn<220><221>变体<222>(25)..(25)<223>xaa可为leu或不存在<220><221>变体<222>(26)..(26)<223>xaa可为asp、his、arg、gly、tyr或trp<220><221>变体<222>(27)..(27)<223>xaa可为gly、asp或ala<220><221>变体<222>(28)..(28)<223>xaa可为ala、phe、gly、his或asp<220><221>变体<222>(29)..(29)<223>xaa可为tyr、phe或leu<400>183leuproalaprolysasnleuvalvalsergluvalthrgluaspser151015leuargleusertrpxaaxaaxaaxaaxaaxaaxaaxaaaspserphe202530leuileglntyrglngluserglulysvalglyglualaileasnleu354045thrvalproglysergluargsertyraspleuthrglyleulyspro505560glythrglutyrthrvalseriletyrglyvalleuglysertyrval65707580phegluhisaspvalmetleuproleuseralagluphethrthr859095<210>184<211>15<212>prt<213>人工序列<220><223>结合c-met的fn3域c链和cd环共有序列<220><221>变体<222>(4)..(4)<223>xaa为trp、phe或val<220><221>变体<222>(8)..(8)<223>xaa为asp、phe或leu<220><221>变体<222>(10)..(10)<223>xaa为val、phe或leu<220><221>变体<222>(11)..(11)<223>xaa为val、leu或thr<220><221>变体<222>(12)..(12)<223>xaa为val、arg、gly、leu、thr或ser<220><221>变体<222>(13)..(13)<223>xaa为gly、ser、ala、thr或lys<220><221>变体<222>(15)..(15)<223>xaa为glu或asp<400>184aspserphexaaileargtyrxaagluxaaxaaxaaxaaglyxaa151015<210>185<211>17<212>prt<213>人工序列<220><223>结合c-met的fn3域f链和fg环共有序列<220><221>变体<222>(4)..(4)<223>xaa为tyr、trp、ile、val、gly或ala<220><221>变体<222>(6)..(6)<223>xaa为asn、thr、qln或gly<220><221>变体<222>(8)..(8)<223>xaa为leu、met、asn或ile<220><221>变体<222>(9)..(9)<223>xaa为gly或ser<220><221>变体<222>(14)..(14)<223>xaa为ser、leu、gly、tyr、thr、arg、his或lys<220><221>变体<222>(15)..(15)<223>xaa为ile、val或leu<220><221>变体<222>(17)..(17)<223>xaa为val、thr、his、ile、pro、tyr或leu<400>185thrglutyrxaavalxaailexaaxaavallysglyglyxaaxaaser151015xaa<210>186<211>89<212>prt<213>人工序列<220><223>结合c-met的fn3域共有序列<220><221>变体<222>(32)..(32)<223>xaa为trp、phe或val<220><221>变体<222>(36)..(36)<223>xaa为asp、phe或leu<220><221>变体<222>(38)..(38)<223>xaa为val、phe或leu<220><221>变体<222>(39)..(39)<223>xaa为val、leu或thr<220><221>变体<222>(40)..(40)<223>xaa为val、arg、gly、leu、thr或ser<220><221>变体<222>(41)..(41)<223>xaa为gly、ser、ala、thr或lys<220><221>变体<222>(43)..(43)<223>xaa为glu或asp<220><221>变体<222>(68)..(68)<223>xaa为tyr、trp、ile、val、gly或ala<220><221>变体<222>(70)..(70)<223>xaa为asn、thr、qln或gly<220><221>变体<222>(72)..(72)<223>xaa为leu、met、asn或ile<220><221>变体<222>(73)..(73)<223>xaa为gly或ser<220><221>变体<222>(78)..(78)<223>xaa为ser、leu、gly、tyr、thr、arg、his或lys<220><221>变体<222>(79)..(79)<223>xaa为ile、val或leu<220><221>变体<222>(81)..(81)<223>xaa为val、thr、his、ile、pro、tyr或leu<400>186leuproalaprolysasnleuvalvalserargvalthrgluaspser151015alaargleusertrpthralaproaspalaalapheaspserphexaa202530ileargtyrxaagluxaaxaaxaaxaaglyxaaalailevalleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrxaavalxaailexaaxaavallysglyglyxaaxaaser65707580xaaproleuseralagluphethrthr85<210>187<211>94<212>prt<213>人工序列<220><223>双特异性egfr/c-met分子的egfr共有fn3域<220><221>变体<222>(11)..(11)<223>xaa为glu、asn或arg<220><221>变体<222>(14)..(14)<223>xaa为glu或pro<220><221>变体<222>(17)..(17)<223>xaa为leu或ala<220><221>变体<222>(25)..(25)<223>xaa为his或trp<220><221>变体<222>(29)..(29)<223>xaa为glu或asp<220><221>变体<222>(37)..(37)<223>xaa为glu或pro<220><221>变体<222>(46)..(46)<223>xaa为asn或val<220><221>变体<222>(73)..(73)<223>xaa为gly或tyr<220><221>变体<222>(83)..(83)<223>xaa为met或ile<220><221>变体<222>(91)..(91)<223>xaa为glu或ile<400>187leuproalaprolysasnleuvalvalserxaavalthrxaaaspser151015xaaargleusertrpaspaspproxaaalaphetyrxaaserpheleu202530ileglntyrglnxaaserglulysvalglyglualailexaaleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrthrvalseriletyrxaavalhisasnvaltyrlysasp65707580thrasnxaaargglyleuproleuseralaxaaphethrthr8590<210>188<211>89<212>prt<213>人工序列<220><223>双特异性egfr/c-met分子的c-met共有fn3域<220><221>变体<222>(11)..(11)<223>xaa为glu、asn或arg<220><221>变体<222>(14)..(14)<223>xaa为glu或pro<220><221>变体<222>(17)..(17)<223>xaa为leu或ala<220><221>变体<222>(37)..(37)<223>xaa为glu或pro<220><221>变体<222>(39)..(39)<223>xaa为val或leu<220><221>变体<222>(40)..(40)<223>xaa为gly或ser<220><221>变体<222>(41)..(41)<223>xaa为ser或lys<220><221>变体<222>(43)..(43)<223>xaa为glu或asp<220><221>变体<222>(46)..(46)<223>xaa为asn或val<220><221>变体<222>(72)..(72)<223>xaa为leu或met<220><221>变体<222>(73)..(73)<223>xaa为gly或ser<220><221>变体<222>(78)..(78)<223>xaa为ser或lys<220><221>变体<222>(86)..(86)<223>xaa为glu或ile<400>188leuproalaprolysasnleuvalvalserxaavalthrxaaaspser151015xaaargleusertrpthralaproaspalaalapheaspserphetrp202530ileargtyrphexaaphexaaxaaxaaglyxaaalailexaaleuthr354045valproglysergluargsertyraspleuthrglyleulysprogly505560thrglutyrvalvalasnilexaaxaavallysglyglyxaaileser65707580proproleuseralaxaaphethrthr85<210>189<211>125<212>prt<213>智人<400>189glnvalglnleuvalgluserglyglyglyvalvalglnproglyarg151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530glymethistrpvalargglnalaproglylysglyleuglutrpval354045alavaliletrpaspaspglysertyrlystyrtyrglyaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargaspglyilethrmetvalargglyvalmetlysasptyrphe100105110asptyrtrpglyglnglythrleuvalthrvalserser115120125<210>190<211>107<212>prt<213>智人<400>190alaileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnaspileserserala202530leuvaltrptyrglnglnlysproglylysalaprolysleuleuile354045tyraspalaserserleugluserglyvalproserargphesergly505560sergluserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnpheasnsertyrproleu859095thrpheglyglyglythrlysvalgluilelys100105<210>191<211>121<212>prt<213>智人<400>191gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530trpmetasntrpvalargglnalaproglylysglyleuglutrpval354045alaasnilelyslysaspglyserglulystyrtyrvalaspserval505560lysglyargphethrileserargaspasnalalysasnserleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargaspleuglytrpglytrpglytrptyrpheaspleutrpgly100105110argglythrleuvalthrvalserser115120<210>192<211>107<212>prt<213>智人<400>192gluilevalleuthrglnserproalathrleuserleuserprogly151015gluargalathrleusercysargalaserglnservalsersertyr202530leualatrptyrglnglnlysproglyglnalaproargleuleuile354045tyraspalaserasnargalathrglyileproalaargphesergly505560serglyserglythraspphethrleuthrileserserleuglupro65707580gluaspphealavaltyrtyrcysglnglnargserasntrppropro859095thrpheglyglnglythrlysvalgluilelys100105<210>193<211>119<212>prt<213>智人<400>193glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercysgluthrserglytyrthrphethrsertyr202530glyilesertrpvalargglnalaproglyhisglyleuglutrpmet354045glytrpileseralatyrasnglytyrthrasntyralaglnlysleu505560glnglyargvalthrmetthrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargaspleuargglythrasntyrpheasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>194<211>107<212>prt<213>智人<400>194aspileglnmetthrglnserproserservalseralaservalgly151015aspargvalthrilethrcysargalaserglnglyileserasntrp202530leualatrppheglnhislysproglylysalaprolysleuleuile354045tyralaalaserserleuleuserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnalaasnserpheproile859095thrpheglyglnglythrargleugluilelys100105<210>195<211>125<212>prt<213>智人<400>195gluvalglnleuvalgluserglyglyglyleuvallysproglygly151015serleulysleusercysalaalaserglyphethrpheserasptyr202530tyrmettyrtrpvalargglnthrproglulysargleuglutrpval354045alathrileseraspaspglysertyrthrtyrtyrproaspserval505560lysglyargphethrileserargaspasnalalysasnasnleutyr65707580leuglnmetserserleulyssergluaspthralamettyrtyrcys859095alaarggluglyleutyrtyrtyrglyserglysertyrtyrasngln100105110asptyrtrpglyglnglythrleuvalthrvalserser115120125<210>196<211>106<212>prt<213>智人<400>196glnleuthrglnserproserserleuseralaservalglyasparg151015valthrilethrcysargalaserglnglyleuserseralaleuala202530trptyrargglnlysproglylysalaprolysleuleuiletyrasp354045alaserserleugluserglyvalproserargpheserglysergly505560serglythraspphethrleuthrileserserleuglnprogluasp65707580phealathrtyrtyrcysglnglnphethrsertyrproglnilethr859095pheglyglnglythrargleugluilelys100105<210>197<211>457<212>prt<213>智人<400>197glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercysglnalaserglytyrargpheserasnphe202530valilehistrpvalargglnalaproglyglnargpheglutrpmet354045glytrpileasnprotyrasnglyasnlysglupheseralalysphe505560glnaspargvalthrphethralaaspthrseralaasnthralatyr65707580metgluleuargserleuargseralaaspthralavaltyrtyrcys859095alaargvalglyprotyrsertrpaspaspserproglnaspasntyr100105110tyrmetaspvaltrpglylysglythrthrvalilevalserserala115120125serthrlysglyproservalpheproleualaproserserlysser130135140thrserglyglythralaalaleuglycysleuvallysasptyrphe145150155160progluprovalthrvalsertrpasnserglyalaleuthrsergly165170175valhisthrpheproalavalleuglnserserglyleutyrserleu180185190serservalvalthrvalproserserserleuglythrglnthrtyr195200205ilecysasnvalasnhislysproserasnthrlysvalasplysarg210215220valgluprolyssercysasplysthrhisthrcysproprocyspro225230235240alaprogluleuleuglyglyproservalpheleupheproprolys245250255prolysaspthrleumetileserargthrprogluvalthrcysval260265270valvalaspvalserhisgluaspprogluvallyspheasntrptyr275280285valaspglyvalgluvalhisasnalalysthrlysproarggluglu290295300glntyrasnserthrtyrargvalvalservalleuthrvalleuhis305310315320glnasptrpleuasnglylysglutyrlyscyslysvalserasnlys325330335alaleuproalaproileglulysthrileserlysalalysglygln340345350proarggluproglnvaltyrthrleuproproserarggluglumet355360365thrlysasnglnvalserleuthrcysleuvallysglyphetyrpro370375380seraspilealavalglutrpgluserasnglyglnprogluasnasn385390395400tyrlysthrthrproprovalleuaspseraspglyserpheleuleu405410415tyrserlysleuthrvalasplysserargtrpglnglnglyasnval420425430phesercysservalmethisglualaleuhisasnhistyrthrgln435440445lysserleuserleuserproglylys450455<210>198<211>457<212>prt<213>智人<400>198glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercysglnalaserglytyrargpheserasnphe202530valilehistrpvalargglnalaproglyglnargpheglutrpmet354045glytrpileasnprotyrasnglyasnlysglupheseralalysphe505560glnaspargvalthrphethralaaspthrseralaasnthralatyr65707580metgluleuargserleuargseralaaspthralavaltyrtyrcys859095alaargvalglyprotyrsertrpaspaspserproglnaspasntyr100105110tyrmetaspvaltrpglylysglythrthrvalilevalserserala115120125serthrlysglyproservalpheproleualaproserserlysser130135140thrserglyglythralaalaleuglycysleuvallysasptyrphe145150155160progluprovalthrvalsertrpasnserglyalaleuthrsergly165170175valhisthrpheproalavalleuglnserserglyleutyrserleu180185190serservalvalthrvalproserserserleuglythrglnthrtyr195200205ilecysasnvalasnhislysproserasnthrlysvalasplysarg210215220valgluprolyssercysasplysthrhisthrcysproprocyspro225230235240alaprogluleuleuglyglyproservalpheleupheproprolys245250255prolysaspthrleumetileserargthrprogluvalthrcysval260265270valvalaspvalserhisgluaspprogluvallyspheasntrptyr275280285valaspglyvalgluvalhisasnalalysthrlysproarggluglu290295300glntyrasnserthrtyrargvalvalservalleuthrvalleuhis305310315320glnasptrpleuasnglylysglutyrlyscyslysvalserasnlys325330335alaleuproalaproileglulysthrileserlysalalysglygln340345350proarggluproglnvaltyrthrleuproproserarggluglumet355360365thrlysasnglnvalserleuthrcysleuvallysglyphetyrpro370375380seraspilealavalglutrpgluserasnglyglnprogluasnasn385390395400tyrlysthrthrproprovalleuaspseraspglyserphepheleu405410415tyrserargleuthrvalasplysserargtrpglnglnglyasnval420425430phesercysservalmethisglualaleuhisasnhistyrthrgln435440445lysserleuserleuserproglylys450455<210>199<211>455<212>prt<213>智人<400>199glnvalglnleuvalgluserglyglyglyvalvalglnproglyarg151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530glymethistrpvalargglnalaproglylysglyleuglutrpval354045alavaliletrpaspaspglysertyrlystyrtyrglyaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargaspglyilethrmetvalargglyvalmetlysasptyrphe100105110asptyrtrpglyglnglythrleuvalthrvalserseralaserthr115120125lysglyproservalpheproleualaproserserlysserthrser130135140glyglythralaalaleuglycysleuvallysasptyrpheproglu145150155160provalthrvalsertrpasnserglyalaleuthrserglyvalhis165170175thrpheproalavalleuglnserserglyleutyrserleuserser180185190valvalthrvalproserserserleuglythrglnthrtyrilecys195200205asnvalasnhislysproserasnthrlysvalasplysargvalglu210215220prolyssercysasplysthrhisthrcysproprocysproalapro225230235240gluleuleuglyglyproservalpheleupheproprolysprolys245250255aspthrleumetileserargthrprogluvalthrcysvalvalval260265270aspvalserhisgluaspprogluvallyspheasntrptyrvalasp275280285glyvalgluvalhisasnalalysthrlysproargglugluglntyr290295300asnserthrtyrargvalvalservalleuthrvalleuhisglnasp305310315320trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu325330335proalaproileglulysthrileserlysalalysglyglnproarg340345350gluproglnvaltyrthrleuproproserarggluglumetthrlys355360365asnglnvalserleuthrcysleuvallysglyphetyrproserasp370375380ilealavalglutrpgluserasnglyglnprogluasnasntyrlys385390395400thrthrproprovalleuaspseraspglyserpheleuleutyrser405410415lysleuthrvalasplysserargtrpglnglnglyasnvalpheser420425430cysservalmethisglualaleuhisasnhistyrthrglnlysser435440445leuserleuserproglylys450455<210>200<211>214<212>prt<213>智人<400>200alaileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnaspileserserala202530leuvaltrptyrglnglnlysproglylysalaprolysleuleuile354045tyraspalaserserleugluserglyvalproserargphesergly505560sergluserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnpheasnsertyrproleu859095thrpheglyglyglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>201<211>449<212>prt<213>智人<400>201glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercysgluthrserglytyrthrphethrsertyr202530glyilesertrpvalargglnalaproglyhisglyleuglutrpmet354045glytrpileseralatyrasnglytyrthrasntyralaglnlysleu505560glnglyargvalthrmetthrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargaspleuargglythrasntyrpheasptyrtrpglyglngly100105110thrleuvalthrvalserseralaserthrlysglyproservalphe115120125proleualaproserserlysserthrserglyglythralaalaleu130135140glycysleuvallysasptyrpheprogluprovalthrvalsertrp145150155160asnserglyalaleuthrserglyvalhisthrpheproalavalleu165170175glnserserglyleutyrserleuserservalvalthrvalproser180185190serserleuglythrglnthrtyrilecysasnvalasnhislyspro195200205serasnthrlysvalasplysargvalgluprolyssercysasplys210215220thrhisthrcysproprocysproalaprogluleuleuglyglypro225230235240servalpheleupheproprolysprolysaspthrleumetileser245250255argthrprogluvalthrcysvalvalvalaspvalserhisgluasp260265270progluvallyspheasntrptyrvalaspglyvalgluvalhisasn275280285alalysthrlysproargglugluglntyrasnserthrtyrargval290295300valservalleuthrvalleuhisglnasptrpleuasnglylysglu305310315320tyrlyscyslysvalserasnlysalaleuproalaproileglulys325330335thrileserlysalalysglyglnproarggluproglnvaltyrthr340345350leuproproserarggluglumetthrlysasnglnvalserleuthr355360365cysleuvallysglyphetyrproseraspilealavalglutrpglu370375380serasnglyglnprogluasnasntyrlysthrthrproprovalleu385390395400aspseraspglyserphepheleutyrserargleuthrvalasplys405410415serargtrpglnglnglyasnvalphesercysservalmethisglu420425430alaleuhisasnhistyrthrglnlysserleuserleuserprogly435440445lys<210>202<211>214<212>prt<213>智人<400>202aspileglnmetthrglnserproserservalseralaservalgly151015aspargvalthrilethrcysargalaserglnglyileserasntrp202530leualatrppheglnhislysproglylysalaprolysleuleuile354045tyralaalaserserleuleuserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnalaasnserpheproile859095thrpheglyglnglythrargleugluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>203<211>330<212>prt<213>智人<400>203alaserthrlysglyproservalpheproleualaproserserlys151015serthrserglyglythralaalaleuglycysleuvallysasptyr202530pheprogluprovalthrvalsertrpasnserglyalaleuthrser354045glyvalhisthrpheproalavalleuglnserserglyleutyrser505560leuserservalvalthrvalproserserserleuglythrglnthr65707580tyrilecysasnvalasnhislysproserasnthrlysvalasplys859095argvalgluprolyssercysasplysthrhisthrcysproprocys100105110proalaprogluleuleuglyglyproservalpheleuphepropro115120125lysprolysaspthrleumetileserargthrprogluvalthrcys130135140valvalvalaspvalserhisgluaspprogluvallyspheasntrp145150155160tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu165170175gluglntyrasnserthrtyrargvalvalservalleuthrvalleu180185190hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn195200205lysalaleuproalaproileglulysthrileserlysalalysgly210215220glnproarggluproglnvaltyrthrleuproproserarggluglu225230235240metthrlysasnglnvalserleuthrcysleuvallysglyphetyr245250255proseraspilealavalglutrpgluserasnglyglnprogluasn260265270asntyrlysthrthrproprovalleuaspseraspglyserphephe275280285leutyrserargleuthrvalasplysserargtrpglnglnglyasn290295300valphesercysservalmethisglualaleuhisasnhistyrthr305310315320glnlysserleuserleuserproglylys325330<210>204<211>330<212>prt<213>智人<400>204alaserthrlysglyproservalpheproleualaproserserlys151015serthrserglyglythralaalaleuglycysleuvallysasptyr202530pheprogluprovalthrvalsertrpasnserglyalaleuthrser354045glyvalhisthrpheproalavalleuglnserserglyleutyrser505560leuserservalvalthrvalproserserserleuglythrglnthr65707580tyrilecysasnvalasnhislysproserasnthrlysvalasplys859095argvalgluprolyssercysasplysthrhisthrcysproprocys100105110proalaprogluleuleuglyglyproservalpheleuphepropro115120125lysprolysaspthrleumetileserargthrprogluvalthrcys130135140valvalvalaspvalserhisgluaspprogluvallyspheasntrp145150155160tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu165170175gluglntyrasnserthrtyrargvalvalservalleuthrvalleu180185190hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn195200205lysalaleuproalaproileglulysthrileserlysalalysgly210215220glnproarggluproglnvaltyrthrleuproproserarggluglu225230235240metthrlysasnglnvalserleuthrcysleuvallysglyphetyr245250255proseraspilealavalglutrpgluserasnglyglnprogluasn260265270asntyrlysthrthrproprovalleuaspseraspglyserpheleu275280285leutyrserlysleuthrvalasplysserargtrpglnglnglyasn290295300valphesercysservalmethisglualaleuhisasnhistyrthr305310315320glnlysserleuserleuserproglylys325330<210>205<211>1368<212>dna<213>智人<400>205caggtgcagctggtcgagagcggcggaggggtggtgcagcccggcagaagcctgaggctg60tcctgcgccgccagcggcttcaccttcagcacctacggcatgcactgggtgcggcaggcc120ccaggcaagggcctggagtgggtggccgtgatctgggacgacggcagctacaagtactac180ggcgacagcgtgaagggcaggttcaccatcagcagggacaacagcaagaacaccctgtac240ctgcagatgaacagcctgagggccgaggacaccgccgtgtactactgcgccagggacggc300atcaccatggtgcggggcgtgatgaaggactacttcgactactggggccagggcaccctg360gtgaccgtgagcagcgccagcaccaagggcccaagcgtgttccccctggcccccagcagc420aagagcaccagcggcggcacagccgccctgggctgcctggtgaaggactacttccccgag480ccagtgaccgtgtcctggaactctggcgccctgacctccggcgtgcacaccttccccgcc540gtgctgcagagcagcggcctgtacagcctgagcagcgtggtgaccgtgcccagcagcagc600ctgggcacccagacctacatctgcaacgtgaaccacaagcccagcaacaccaaggtggac660aagagagtggagcccaagagctgcgacaagacccacacctgccccccctgcccagcccca720gagctgctgggcggacccagcgtgttcctgttcccccccaagcccaaggacaccctgatg780atcagcaggacccccgaggtgacctgcgtggtggtggacgtgagccacgaggacccagag840gtgaagttcaactggtacgtggacggcgtggaggtgcacaacgccaagaccaagcccaga900gaggagcagtacaacagcacctacagggtggtgtccgtgctgaccgtgctgcaccaggac960tggctgaacggcaaggaatacaagtgcaaggtctccaacaaggccctgccagcccccatc1020gaaaagaccatcagcaaggccaagggccagccacgggagccccaggtgtacaccctgccc1080cccagccgggaggagatgaccaagaaccaggtgtccctgacctgtctggtgaagggcttc1140taccccagcgacatcgccgtggagtgggagagcaacggccagcccgagaacaactacaag1200accacccccccagtgctggacagcgacggcagcttcctcctgtacagcaagctgaccgtg1260gacaagtccaggtggcagcagggcaacgtgttcagctgcagcgtgatgcacgaggccctg1320cacaaccactacacccagaagtccctgagcctgagccccggcaaatga1368<210>206<211>642<212>dna<213>智人<400>206atccagctgacccagagccccagcagcctgagcgccagcgtgggcgaccgggtgaccatc60acctgccgggccagccaggacatcagcagcgccctggtctggtatcagcagaagcccggc120aaggcccccaagctgctgatctacgacgccagctccctggaaagcggcgtgcccagccgg180ttcagcggcagcgagagcggcaccgacttcaccctgaccatcagcagcctgcagcccgag240gacttcgccacctactactgccagcagttcaacagctaccccctgacctttggcggcgga300acaaaggtggagatcaagcgtacggtggccgctcccagcgtgttcatcttcccccccagc360gacgagcagctgaagagcggcaccgccagcgtggtgtgcctgctgaacaacttctacccc420cgggaggccaaggtgcagtggaaggtggacaacgccctgcagagcggcaacagccaggag480agcgtcaccgagcaggacagcaaggactccacctacagcctgagcagcaccctgaccctg540tccaaggccgactacgagaagcacaaggtgtacgcctgcgaggtgacccaccagggcctg600tccagccccgtgaccaagagcttcaacaggggcgagtgctga642<210>207<211>1350<212>dna<213>智人<400>207caggttcagctggtgcagtctggagctgaggtgaagaagcctggggcctcagtgaaggtc60tcctgcgagacttctggttacacctttaccagctatggtatcagctgggtgcgacaggcc120cctggacacgggcttgagtggatgggatggatcagcgcttacaatggttacacaaactat180gcacagaagctccagggcagggtcaccatgaccacagacacatccacgagcacagcctac240atggagctgaggagcctgagatctgacgacacggccgtgtattactgtgcgagagatctg300agaggaactaactactttgactactggggccagggaaccctggtcaccgtctcctcagcc360tccaccaagggcccaagcgtgttccctctggcccccagcagcaagagcacatctggcgga420acagccgccctgggctgcctggtgaaagactacttccccgagcccgtgaccgtgtcctgg480aactctggcgccctgaccagcggcgtgcacacctttccagccgtgctgcagagcagcggc540ctgtacagcctgtccagcgtggtgaccgtgcccagcagctccctgggcacccagacctac600atctgcaacgtgaaccacaagcccagcaacaccaaggtggacaagcgggtggaacccaag660agctgcgacaagacccacacctgtcccccctgccctgcccctgaactgctgggcggaccc720tccgtgttcctgttccccccaaagcccaaggacaccctgatgatcagccggacccccgaa780gtgacctgcgtggtggtggacgtgtcccacgaggaccctgaagtgaagttcaattggtac840gtggacggcgtggaagtgcacaacgccaagaccaagcccagagaggaacagtacaacagc900acctaccgggtggtgtccgtgctgacagtgctgcaccaggactggctgaacggcaaagag960tacaagtgcaaggtctccaacaaggccctgcctgctcccatcgagaaaaccatcagcaag1020gccaagggccagccccgcgagcctcaggtgtacacactgcctcccagccgggaagagatg1080accaagaaccaggtgtccctgacctgtctggtgaaaggcttctaccccagcgatatcgcc1140gtggaatgggagagcaacggacagcccgagaacaactacaagaccaccccccctgtgctg1200gacagcgacggctccttcttcctgtactctcggctgaccgtggacaagagccggtggcag1260cagggaaacgtgttcagctgcagcgtgatgcacgaggccctgcacaaccactacacccag1320aagtccctgagcctgagccccgggaagtga1350<210>208<211>645<212>dna<213>智人<400>208gacatccagatgacccagtccccctcctccgtgtccgcctctgtgggcgacagagtgacc60atcacctgtcgggcctcccagggcatctccaactggctggcctggttccagcacaagccc120ggcaaggcccccaagctgctgatctacgccgcctcctccctgctgtccggcgtgccctcc180agattctccggctctggctccggcaccgacttcaccctgaccatctccagcctgcagccc240gaggacttcgccacctactactgccagcaggccaactccttccccatcaccttcggccag300ggcacccggctggaaatcaagcgtacggtggccgctcccagcgtgttcatcttccccccc360agcgacgagcagctgaagagcggcaccgccagcgtggtgtgcctgctgaacaacttctac420ccccgggaggccaaggtgcagtggaaggtggacaacgccctgcagagcggcaacagccag480gagagcgtcaccgagcaggacagcaaggactccacctacagcctgagcagcaccctgacc540ctgtccaaggccgactacgagaagcacaaggtgtacgcctgcgaggtgacccaccagggc600ctgtccagccccgtgaccaagagcttcaacaggggcgagtgctga645<210>209<211>215<212>prt<213>智人<400>209gluilevalleuthrglnserproglythrleuserleuserprogly151015gluargalathrphesercysargserserhisserileargserarg202530argvalalatrptyrglnhislysproglyglnalaproargleuval354045ilehisglyvalserasnargalaserglyileseraspargpheser505560glyserglyserglythraspphethrleuthrilethrargvalglu65707580progluaspphealaleutyrtyrcysglnvaltyrglyalaserser859095tyrthrpheglyglnglythrlysleugluarglysargthrvalala100105110alaproservalpheilepheproproseraspgluglnleulysser115120125glythralaservalvalcysleuleuasnasnphetyrproargglu130135140alalysvalglntrplysvalaspasnalaleuglnserglyasnser145150155160glngluservalthrgluglnaspserlysaspserthrtyrserleu165170175serserthrleuthrleuserlysalaasptyrglulyshislysval180185190tyralacysgluvalthrhisglnglyleuserserprovalthrlys195200205serpheasnargglyglucys210215<210>210<211>5<212>prt<213>智人<400>210thrtyrglymethis15<210>211<211>17<212>prt<213>智人<400>211valiletrpaspaspglysertyrlystyrtyrglyaspservallys151015gly<210>212<211>16<212>prt<213>智人<400>212aspglyilethrmetvalargglyvalmetlysasptyrpheasptyr151015<210>213<211>11<212>prt<213>智人<400>213argalaserglnaspileserseralaleuval1510<210>214<211>7<212>prt<213>智人<400>214aspalaserserleugluser15<210>215<211>9<212>prt<213>智人<400>215glnglnpheasnsertyrproleuthr15<210>216<211>5<212>prt<213>智人<400>216sertyrglyileser15<210>217<211>17<212>prt<213>智人<400>217trpileseralatyrasnglytyrthrasntyralaglnlysleugln151015gly<210>218<211>10<212>prt<213>智人<400>218aspleuargglythrasntyrpheasptyr1510<210>219<211>11<212>prt<213>智人<400>219argalaserglnglyileserasntrpleuala1510<210>220<211>7<212>prt<213>智人<400>220alaalaserserleuleuser15<210>221<211>9<212>prt<213>智人<400>221glnglnalaasnserpheproilethr15<210>222<211>5<212>prt<213>智人<400>222sertyrtrpmetasn15<210>223<211>17<212>prt<213>智人<400>223asnilelyslysaspglyserglulystyrtyrvalaspservallys151015gly<210>224<211>12<212>prt<213>智人<400>224aspleuglytrpglytrpglytrptyrpheaspleu1510<210>225<211>11<212>prt<213>智人<400>225argalaserglnservalsersertyrleuala1510<210>226<211>7<212>prt<213>智人<400>226aspalaserasnargalathr15<210>227<211>9<212>prt<213>智人<400>227glnglnargserasntrpproprothr15<210>228<211>5<212>prt<213>智人<400>228asptyrtyrmettyr15<210>229<211>17<212>prt<213>智人<400>229thrileseraspaspglysertyrthrtyrtyrproaspservallys151015gly<210>230<211>16<212>prt<213>智人<400>230gluglyleutyrtyrtyrglyserglysertyrtyrasnglnasptyr151015<210>231<211>11<212>prt<213>智人<400>231argalaserglnglyleuserseralaleuala1510<210>232<211>7<212>prt<213>智人<400>232aspalaserserleugluser15<210>233<211>10<212>prt<213>智人<400>233glnglnphethrsertyrproglnilethr1510<210>234<211>451<212>prt<213>智人<400>234gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530trpmetasntrpvalargglnalaproglylysglyleuglutrpval354045alaasnilelyslysaspglyserglulystyrtyrvalaspserval505560lysglyargphethrileserargaspasnalalysasnserleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargaspleuglytrpglytrpglytrptyrpheaspleutrpgly100105110argglythrleuvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplysargvalgluprolyssercys210215220asplysthrhisthrcysproprocysproalaprogluleuleugly225230235240glyproservalpheleupheproprolysprolysaspthrleumet245250255ileserargthrprogluvalthrcysvalvalvalaspvalserhis260265270gluaspprogluvallyspheasntrptyrvalaspglyvalgluval275280285hisasnalalysthrlysproargglugluglntyrasnserthrtyr290295300argvalvalservalleuthrvalleuhisglnasptrpleuasngly305310315320lysglutyrlyscyslysvalserasnlysalaleuproalaproile325330335glulysthrileserlysalalysglyglnproarggluproglnval340345350tyrthrleuproproserarggluglumetthrlysasnglnvalser355360365leuthrcysleuvallysglyphetyrproseraspilealavalglu370375380trpgluserasnglyglnprogluasnasntyrlysthrthrpropro385390395400valleuaspseraspglyserpheleuleutyrserlysleuthrval405410415asplysserargtrpglnglnglyasnvalphesercysservalmet420425430hisglualaleuhisasnhistyrthrglnlysserleuserleuser435440445proglylys450<210>235<211>214<212>prt<213>智人<400>235gluilevalleuthrglnserproalathrleuserleuserprogly151015gluargalathrleusercysargalaserglnservalsersertyr202530leualatrptyrglnglnlysproglyglnalaproargleuleuile354045tyraspalaserasnargalathrglyileproalaargphesergly505560serglyserglythraspphethrleuthrileserserleuglupro65707580gluaspphealavaltyrtyrcysglnglnargserasntrppropro859095thrpheglyglnglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>236<211>455<212>prt<213>智人<400>236gluvalglnleuvalgluserglyglyglyleuvallysproglygly151015serleulysleusercysalaalaserglyphethrpheserasptyr202530tyrmettyrtrpvalargglnthrproglulysargleuglutrpval354045alathrileseraspaspglysertyrthrtyrtyrproaspserval505560lysglyargphethrileserargaspasnalalysasnasnleutyr65707580leuglnmetserserleulyssergluaspthralamettyrtyrcys859095alaarggluglyleutyrtyrtyrglyserglysertyrtyrasngln100105110asptyrtrpglyglnglythrleuvalthrvalserseralaserthr115120125lysglyproservalpheproleualaproserserlysserthrser130135140glyglythralaalaleuglycysleuvallysasptyrpheproglu145150155160provalthrvalsertrpasnserglyalaleuthrserglyvalhis165170175thrpheproalavalleuglnserserglyleutyrserleuserser180185190valvalthrvalproserserserleuglythrglnthrtyrilecys195200205asnvalasnhislysproserasnthrlysvalasplysargvalglu210215220prolyssercysasplysthrhisthrcysproprocysproalapro225230235240gluleuleuglyglyproservalpheleupheproprolysprolys245250255aspthrleumetileserargthrprogluvalthrcysvalvalval260265270aspvalserhisgluaspprogluvallyspheasntrptyrvalasp275280285glyvalgluvalhisasnalalysthrlysproargglugluglntyr290295300asnserthrtyrargvalvalservalleuthrvalleuhisglnasp305310315320trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu325330335proalaproileglulysthrileserlysalalysglyglnproarg340345350gluproglnvaltyrthrleuproproserarggluglumetthrlys355360365asnglnvalserleuthrcysleuvallysglyphetyrproserasp370375380ilealavalglutrpgluserasnglyglnprogluasnasntyrlys385390395400thrthrproprovalleuaspseraspglyserphepheleutyrser405410415argleuthrvalasplysserargtrpglnglnglyasnvalpheser420425430cysservalmethisglualaleuhisasnhistyrthrglnlysser435440445leuserleuserproglylys450455<210>237<211>213<212>prt<213>智人<400>237glnleuthrglnserproserserleuseralaservalglyasparg151015valthrilethrcysargalaserglnglyleuserseralaleuala202530trptyrargglnlysproglylysalaprolysleuleuiletyrasp354045alaserserleugluserglyvalproserargpheserglysergly505560serglythraspphethrleuthrileserserleuglnprogluasp65707580phealathrtyrtyrcysglnglnphethrsertyrproglnilethr859095pheglyglnglythrargleugluilelysargthrvalalaalapro100105110servalpheilepheproproseraspgluglnleulysserglythr115120125alaservalvalcysleuleuasnasnphetyrproargglualalys130135140valglntrplysvalaspasnalaleuglnserglyasnserglnglu145150155160servalthrgluglnaspserlysaspserthrtyrserleuserser165170175thrleuthrleuserlysalaasptyrglulyshislysvaltyrala180185190cysgluvalthrhisglnglyleuserserprovalthrlysserphe195200205asnargglyglucys210<210>238<211>15<212>prt<213>智人<400>238proglupheargaspsertyrproilelystyrvalhisalaphe151015<210>239<211>16<212>prt<213>智人<400>239phealaglnserlysproaspseralagluprometaspargserala151015<210>240<211>16<212>prt<213>智人<400>240proglyalaglnleualaargglnileglyalaserleuasnaspasp151015当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1