基于藻类细胞能量代谢的磷营养限制判别方法与流程
本发明涉及藻类生理学、生物热力学、生态化学计量学等研究领域,特别是涉及一种基于藻类细胞能量代谢的磷营养限制判别方法。
背景技术:
在水生生态系统中,藻类生物量时常受到水体磷含量的限制。在藻类营养盐的绝对限制法则中,当磷限制发生时,环境中磷浓度会下降至较低水平,而0.1μmol/l被认为是藻类磷营养吸收的最低阈值。这时藻细胞无法输出足够的能量来驱动磷的转运过程,即不足以克服跨膜形成的电化学势梯度。这种情况下,藻类一般通过特殊机制开发周围环境磷浓度,使之瞬时升高以维持正常代谢活动,例如利用水生动物的排泄物等。因此,当环境磷浓度超过某一特征值时,藻细胞可输出足够的能量以启动吸收系统,使磷快速地积聚在细胞膜外侧并完成胞外至胞内的转运过程,进而以聚合磷酸盐的形式储存起来。
技术实现要素:
针对上述问题,本发明提供一种基于细胞能量代谢定量判断藻类磷营养限制的判别方法。该判别方法通过条件设定建立藻类细胞磷营养通量基础的模型,得到定态能量下环境磷浓度与营养通量之间的数学关系式,结合藻类磷吸收的实验数据,作图计算磷营养通量为零时的磷浓度阈值,以获得用于判别的参数,最后判别出一定胞内营养状态的藻细胞是否处于磷营养限制水平。
该判别方法的目的在于解决以下问题:通过对一种基于藻类细胞能量代谢的磷营养通量模型的条件设定,将磷的绝对限制与藻类胞内营养状态相联系,以得到藻细胞生理特异的磷浓度阈值,该阈值可用于判别藻类细胞是否处于磷的绝对限制状态,以及可反映出藻类对环境磷浓度变化的生长响应。
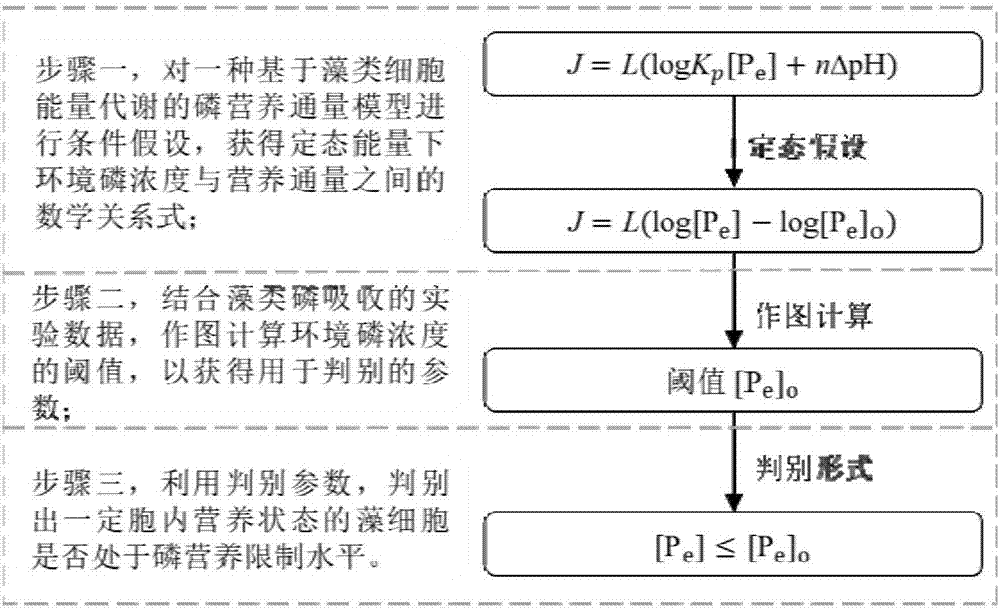
为实现上述目的及其他相关目的,本发明提供一种基于藻类细胞能量代谢的磷营养限制判别方法,包括如下步骤:
1)对基于藻类细胞能量代谢的磷营养通量模型进行条件假设,获得定态能量下的磷营养通量模型;
2)结合藻类磷吸收的实验数据,通过磷营养通量模型计算得到阈值,通过比较环境中磷浓度与阈值的大小,判别藻细胞是否处于磷营养限制水平。
在本发明的一些实施例中,步骤2)中,作图计算磷营养通量为零时的磷浓度阈值[pe]o,通过该浓度阈值[pe]o,利用判别式[pe]≤[pe]o判别一定胞内营养状态的藻细胞是否处于磷营养限制水平,如果上述判别式成立,则为绝对限制;若[pe]>[pe]o,藻细胞将吸收磷营养盐直至下一次绝对限制发生;pe表示环境中磷浓度,从环境中检测得到,单位nmol/l,当然,根据换算,其单位也可以为μmol/l等。
在本发明的一些实施例中,步骤1)中的藻类细胞能量代谢的磷营养通量模型表达式为:
j=l(logkp[pe]+nδph)(1)
式中,j为磷营养通量,l为模型系数,kp为环境中磷经跨膜转运、胞内转化生成聚合磷酸盐的平衡常数,δph为h+迁移形成的ph梯度,n为迁移的h+数目。
在本发明的一些实施例中,步骤1)中,所述条件假设如下:对所得的线性磷营养通量模型进行能量的定态假设,该假设下,一定营养状态的藻类细胞处于磷绝对限制状态,此时细胞膜对磷的主动运输能力仅受吉布斯自由能约束,若要激发吸收行为则需通过提高环境磷浓度来实现。
在本发明的一些实施例中,在上述假设条件下,藻类的磷营养通量为零时,(logkp+nδph)=-log[pe]o,得到线性表达式为:
j=l(log[pe]-log[pe]o)(2)
在本发明的一些实施例中,步骤2)中的作图方法为:以j为y轴,log[pe]为x轴进行作图,结合藻类磷吸收的实验数据,得到与藻类细胞生理特异的直线,x轴的截距即为磷营养通量为零时的磷浓度阈值[pe]o,直线斜率即为模型系数l,通过控制实验条件可得到不同生理含义的模型系数的值。
本发明采用的藻类磷营养通量模型是一种从细胞能量代谢角度,定量描述环境磷浓度与营养通量之间的线性非平衡热力学关系,可定量分析藻细胞在不同营养状态下的受环境磷浓度约束的能量水平。
附图说明
为了使本发明专利的目的、构建步骤和有益效果更加清楚,本发明专利提供如下附图进行说明:
图1为本发明提供的一种基于藻类细胞能量代谢的磷营养限制判别方法流程图。
图2为本发明提供的图1中一种基于藻类细胞能量代谢的磷营养限制判别方法详细流程图。
图3为本发明实施例1中磷营养通量与log[pe]的线性关系。
图4为本发明实施例1中不同生长速率下磷营养通量与log[pe]的线性关系图。
图5为本发明实施例2中不同生长策略藻种磷营养通量与log[pe]的线性关系图。
具体实施方式
以下通过特定的具体实例说明本发明的实施方式,本领域技术人员可由本说明书所阐释的内容清晰地了解本发明的其他优点与功能。本发明还可以通过另外不同的具体实施方式加以实施或应用,本说明书中的各项细节也可以基于不同观点与应用,在没有背离本发明的理念下进行相关修饰或改变。
本发明限定藻细胞磷吸收过程的能量代谢仅包括磷的转运、磷的同化和磷的储存三个环节,图1为基于藻类细胞能量代谢的磷营养通量模型构建流程图,图2为本发明提供的图1中一种基于藻类细胞能量代谢的磷营养通量模型构建流程说明图。首先,环境中磷通过细胞膜转运至细胞内部,然后胞内磷经同化作用与二磷酸腺苷(adp)转化为atp,最后atp合成聚合磷(polyp)完成磷的储存,具体为:1)环境中磷(pe)经细胞膜运输至胞内的转运过程[式(3)],2)胞内磷(pi)转化为三磷酸腺苷(atp)的光合磷酸化过程[式(4)],3)atp合成聚合磷酸盐(polyp)分子的磷酸根转移过程[式(5)],得到的过程总反应式[式(6)]。
过程①,相应的跨膜转运过程的化学反应方程式为:
过程②,相应的光合磷酸化过程的化学反应方程式为:
过程③,相应的磷酸根转移过程的化学反应方程式为:
统一组织上述三个化学反应式后,得到的藻类磷吸收过程的总反应式为:
本发明涉及的磷营养通量(j)的改变,是由上述总反应的吉布斯自由能变化(δg)推动,因此该能变是引起磷营养物质迁移的推动力(x),本发明未涉及的其他影响因素由线性系数(l)表征。结合吉布斯自由能变化的热力学函数δg=-rtlnk[s]/[p],r为气体常数,8.314j·mol–1·k–1;t为热力学温度,单位k;k为平衡常数,推导需要,不进行测定;s为反应物浓度,单位mol/l;p为生成物浓度,单位mol/l。
得到推动力x的表达式[式(7)],再结合线性非平衡热力学函数j=l·x,得到模型表达式[式(1)]:
将上式ln替换为log,lnn=2.3logn:
j=l(logkp[pe]+nδph)(1)
式中,j为磷营养通量,l为模型系数,kp为环境中磷经跨膜转运、胞内转化生成polyp的平衡常数,δph为h+迁移形成的ph梯度,n为迁移的h+数目。从物理意义上说,磷营养通量表示藻细胞在单位时间所吸收的磷营养物质的量。单位不唯一,因为藻细胞生物量可用叶绿素a浓度(chla,μg/l)、细胞密度(celld,个/l)、细胞干重(dw,mg/l)等指标进行表征,本发明的实施例采用叶绿素a浓度来表征藻细胞生物量,故j单位为μmol·(μgchla·h)–1,根据单位换算,作图时也采用μmol·(mgchla·min)–1或μmol·(mgchla·h)–1表示。模型中kp、nδph参数是热力学方法推导的要素,具有其本身的化学意义,但本发明不对其进行细胞分子层面的测定,关键是把握宏观磷营养盐与藻类吸收的关系,故测定指标仅包括胞外磷浓度和藻细胞生物量。j通过具体实验,作图计算得到。
实施例1—绝对限制
实验实施:本实验以蓝藻为研究对象,蓝藻是一藻属,本实验具体采用微囊藻microcystisflos-aquae(fachb-1028),来源于中国科学院武汉水生生物研究所淡水藻种库,详见http://algae.ihb.ac.cn/,用bg-11液体培养基对微囊藻进行间歇培养,以磷酸盐作为培养液中藻类吸收实验的唯一磷营养物来源。藻种的培养在光照培养箱中进行,培养温度为25±1℃,光照强度为25μmol·m–2·s–1,光暗比为12h:12h。配置磷浓度梯度为0.02、0.2、1.0、5.0mg/l(k2hpo4)的培养液,加入500μl藻液(0.1μgchla/ml)进行培养,一段时间后测量磷营养通量。
具体培养方法如下:在实验开始前,对藻细胞进行饥饿处理,将藻细胞离心收集,用无磷的bg-11培养基(k2hpo4由等摩尔的kcl代替)洗涤以去除吸附的磷,重复洗涤,离心3次后转入无磷bg-11培养基中,饥饿培养7d以消耗细胞内储存的磷,将饥饿培养的藻种接入实验设置的不同磷浓度培养液中进行间歇培养,接种浓度为1×107cell/l,3组平行。分别在接种1、2、3h后取样,用gf/c滤膜过滤后测定样品中磷酸盐的含量。根据已知的磷酸盐投入量和培养基中磷酸盐的剩余量求得细胞对磷的吸收量。藻类的生物量由叶绿素a(chla)计算,具体采用热乙醇萃取分光光度法。
将实验结果按判别形式所述的作图法进行绘制,log[pe]与磷营养通量j的线性关系如图3所示,可得到x轴截距为–1.71=log0.0195,即阈值为0.0195μmol/l,换算单位后,得到磷营养限制阈值为19.5nmol/l。而实验直接测得培养液中磷浓度为19±2nmol/l,与上述数学方法结果近似。
进一步地,将同一批次的藻种进行连续培养,控制稀释速率在0.055h–1、0.085h–1、0.125h–1,分别记作a、b和c组,用相同方法对实验数据进行处理,如图4所示为不同生长速率下磷营养通量与log[pe]的线性关系,纵坐标表示磷营养通量j,横坐标表示log[pe],得到x轴截距为–2.5=log0.00316,即阈值为0.00316μmol/l,换算单位后,得到磷营养限制阈值为3.16nmol/l。实验直接测得a、b和c的阈值分别为3.2、3.3和4.0nmol/l,与本方法结果近似。在本实验中,同一批次实验藻种的控制条件为生长速率,故其胞内营养状态相同,所体现的磷浓度阈值相同;同时,直线斜率所代表的生理含义即为藻类细胞的生长速率。
上述实验中,直接检测培养液中磷浓度的方法为磷酸盐测定方法,具体参照《水和废水监测分析方法》(第四版)进行,参考文献:国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4版.北京:中国环境科学出版社,2002.
叶绿素a的浓度测定参照:章宗涉,黄祥飞.淡水浮游生物研究方法.北京:科学出版社,1991.
实施例2—种间竞争
在获得不同生长策略藻种磷绝对限制阈值的基础上,对培养藻种进行横向比较,培养条件同实施例1,并从以下四个方面进行判别:
阈值较小,且培养期间阈值变化有递减趋势,表明在同期生境条件下该生长策略藻种具有更低资源需求,进而具有生长优势将在竞争中排斥其他藻而成为优势藻。
阈值较小,但培养期间阈值变化有递增趋势或基本不变,表明该生长策略藻种对资源需求变高,进而不具备竞争优势。
斜率较大,且培养期间生长速率变化有递增的趋势,表明该生长策略藻种对磷的亲和性增加,而后储存的能量用于生长增速,进而具有竞争优势。
斜率较小,但培养期间生长速率变化有递减趋势或基本不变,表明该生长策略藻种对磷的亲和性低,且储存的能量更多地用于合成细胞膜上的转运蛋白,相较而言不具备竞争优势。
以不同生长策略藻种为实验对象,选择小环藻(r型生长策略)、鱼腥藻(cs型生长策略)和微囊藻(s型生长策略)进行间歇培养,实验藻种水华鱼腥藻(anabaenaflos-aquae)、铜绿微囊藻(microcystisaeruginosa)、小环藻(cyclotellahubeiana)购买于中国科学院水生生物研究所(编号分别为fachb-245、fachb-1092、fachb-1030),实验条件同前,测得不同生长策略藻种的磷营养通量。实验结果如图5所示,小环藻(r型生长策略)的阈值明显地小于其他藻种,表明r型藻种相对处于胞内贫营养状态,对磷的亲和性较高,在物质限制条件下较其他藻种更具资源竞争优势;鱼腥藻(cs型生长策略)的磷浓度阈值较大且斜率较小,表明cs型藻种相对处于胞内富营养状态,对磷的亲和性较低,但对能量的储存能力较强;微囊藻(s型生长策略)的磷浓度阈值较大且斜率较大,表明s型藻种虽然对磷亲和性较低,但能够在能量限制条件下具有生长优势。
综上所述,本发明旨在提供一种在藻类磷营养吸收过程中绝对限制的热力学判别方法。其技术方案要点是:通过将藻类磷吸收的线性非平衡热力学模型转换成定态能量下的磷营养通量模型,结合藻类磷吸收的实验数据作图,计算磷营养通量为零时的磷浓度阈值,以获得用于判别的参数,最后判别出处于一定胞内营养状态的藻细胞是否处于磷营养限制水平。
最后说明的是,以上优选实施例仅用以说明本发明的验证技术方案而非限制,尽管通过上述优选实施例已经对本发明进行了详细的描述,但本领域技术人员应当理解,可以在形式上和细节上对其作出各种各样的改变,而不偏离本发明的权利要求书所限定的范围。
- 还没有人留言评论。精彩留言会获得点赞!