一种柱花草苯丙氨酸解氨酶SgPAL2及其编码基因和应用
一种柱花草苯丙氨酸解氨酶sgpal2及其编码基因和应用
技术领域
1.本发明属于基因工程技术领域。更具体地,涉及一种柱花草苯丙氨酸解氨酶sgpal2及其编码基因和应用。
背景技术:
2.锰(manganese,mn)是植物生长发育必需的微量营养元素之一,参与植物体内光合作用、呼吸作用和蛋白质合成等一系列生理生化的过程。但是,锰也是重金属元素之一,因土壤污染等导致的锰过量积累会对植物产生毒害作用,锰毒害的典型特征是成熟叶片上出现褐色斑点、萎黄和坏死,最终会抑制植物生长。
3.苯丙氨酸解氨酶(phenylalanine ammonia-lyase,pal)是植物苯丙烷代谢途径的限速酶。苯丙氨酸经苯丙烷代谢途径转化形成的酚类、类黄酮和木质素等次级代谢物,在植物生长发育、抗病和抗逆中起关键作用。研究发现,金属锰毒胁迫会影响植物苯丙烷代谢途径,从而改变次级代谢物含量。例如,在过量锰处理条件下,豇豆叶片酚类物质和胼胝质含量显著增加;类似的,过量锰处理下水稻叶片中的木质素和黄酮类化合物增加,过量锰处理下杨树叶片中苯丙氨酸的含量增加,进而缓解了锰对植物体的毒害。目前对于苯丙氨酸解氨酶与植物锰毒胁迫相关的研究中,多数在生理水平,关于苯丙氨酸解氨酶基因是否参与缓解植物金属锰毒胁迫的研究尚无明确的报道。
4.柱花草(stylosanthes guianensis)是一种多年生豆科草本植物,原产于拉丁美洲,其可用作家畜饲草,用于果园间作和酸性土壤改良等,是热带和亚热带地区广泛种植的优良豆科牧草。在长期的自然进化和人工选育过程中,柱花草进化出具有较强的适应酸性土壤金属锰和铝毒害的能力,这可能与其体内的基因调控相关,但仍未挖掘到具体的基因。因此,挖掘柱花草适应金属锰毒胁迫的相关基因对培育耐锰毒害的作物新品种具有重要意义。
技术实现要素:
5.本发明要解决的技术问题是克服现有仍未有可提高植物对锰毒害耐受能力相关的蛋白或基因的不足,提供一种柱花草苯丙氨酸解氨酶sgpal2及其编码基因和应用。
6.本发明的第一个目的是提供一种柱花草苯丙氨酸解氨酶sgpal2。
7.本发明的第二个目的是提供一种柱花草苯丙氨酸解氨酶sgpal2的编码基因。
8.本发明的第三个目的是提供一种重组表达载体。
9.本发明的第四个目的是提供一种重组工程菌。
10.本发明的第五个目的是提供所述柱花草苯丙氨酸解氨酶sgpal2、所述编码基因、所述重组表达载体或所述重组工程菌在提高植物对锰毒害耐受能力中的应用。
11.本发明的第六个目的是提供所述柱花草苯丙氨酸解氨酶sgpal2、所述编码基因、所述重组表达载体或所述重组工程菌在制备耐锰毒害的转基因植物中的应用。
12.本发明的第七个目的是提供所述柱花草苯丙氨酸解氨酶sgpal2、所述编码基因、
所述重组表达载体或所述重组工程菌在降低植物锰积累中的应用。
13.本发明上述目的通过以下技术方案实现:
14.本发明提供了一种柱花草苯丙氨酸解氨酶sgpal2,其氨基酸序列如seq id no.2所示。
15.本发明还提供了所述苯丙氨酸解氨酶sgpal2的编码基因。
16.具体地,所述编码基因的核苷酸序列如seq id no.1所示,长2076bp。
17.本发明还提供了seq id no.1所示柱花草苯丙氨酸解氨酶基因sgpal2的克隆引物,其中,上游引物的核苷酸序列如seq id no.3所示,下游引物序列如seq id no.4所示。
18.本发明还提供了seq id no.1所示柱花草苯丙氨酸解氨酶基因sgpal2的克隆方法,即以柱花草的cdna为模板,用seq id no.3和4所示引物进行扩增。
19.本发明还提供了一种重组表达载体,其含有本发明所述苯丙氨酸解氨酶sgpal2的编码基因。
20.具体地,所述表达载体为超量表达载体。
21.更具体地,所述超量表达载体为ptf101s。
22.本发明还提供了一种重组工程菌,其含有本发明所述重组表达载体。
23.具体地,所述工程菌为大肠杆菌trans
1-t或农杆菌感受态细胞gv3101。
24.本发明通过转基因技术在植物中超量表达seq id no.1所示柱花草苯丙氨酸解氨酶基因sgpal2发现,超量表达sgpal2的转基因植株较野生型,其对锰毒害的耐受能力明显增强,并证实了该基因是通过减少植株中锰的积累而增强其对锰毒害的耐受性。
25.因此,本发明申请保所述柱花草苯丙氨酸解氨酶sgpal2、所述柱花草苯丙氨酸解氨酶sgpal2的编码基因、所述重组表达载体或所述重组工程菌在提高植物对锰毒害耐受能力中的应用。
26.具体地,所述提高植物对锰毒害耐受能力是通过在植物中表达柱花草苯丙氨酸解氨酶sgpal2实现的。
27.本发明还申请保护所述柱花草苯丙氨酸解氨酶sgpal2、所述柱花草苯丙氨酸解氨酶sgpal2的编码基因、所述重组表达载体或所述重组工程菌在制备耐锰毒害的转基因植物中的应用。
28.本发明还申请保护所述柱花草苯丙氨酸解氨酶sgpal2、所述柱花草苯丙氨酸解氨酶sgpal2的编码基因、所述重组表达载体或所述重组工程菌在降低植物锰积累中的应用。
29.具体地,所述植物为双子叶植物。
30.更具体地,所述双子叶植物为柱花草或拟南芥。
31.本发明具有以下有益效果:
32.本发明在柱花草中克隆到了一个表达水平与柱花草对锰毒害响应过程高度吻合的苯丙氨酸解氨酶基因sgpal2,并通过转基因拟南芥表达体系,证明了sgpal2的表达能够促进转基因拟南芥对锰毒害的耐受能力,同时证明了该基因的作用机制是通过减少植株中锰的积累,从而提高植物对锰毒害的耐受能力,因此可将其用于制备耐锰毒害的转基因植物。本发明不仅丰富了植物耐锰毒害的基因库,还有助于耐锰毒害的作物品种的培育。
附图说明
33.图1为不同金属和不同浓度的锰处理对柱花草地上部中sgpal2基因表达的影响结果;其中,图a为不同金属处理对sgpal2基因的表达的影响结果;图b为不同浓度的锰处理对sgpal2基因的表达的影响结果;图中数据为3个重复的平均值和标准误差,*号表示处理与对照之间差异显著,p《0.05。
34.图2为sgpal2基因的亚细胞定位结果;图中第一列为gfp荧光信号,第二列为叶绿体自发荧光信号,第三列为明场,第四列为gfp与叶绿体自发荧光信号的融合结果,图中标尺为20μm。
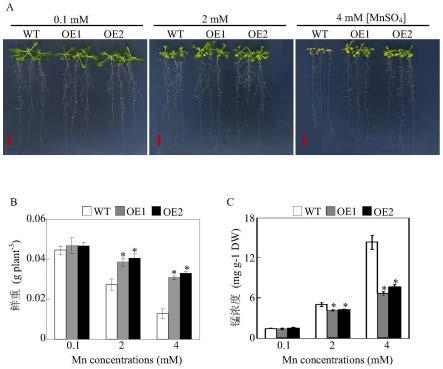
35.图3为超量表达sgpal2基因对拟南芥锰毒害耐受性的影响结果;其中,图a为超量表达拟南芥株系和野生型拟南芥在不同浓度的锰处理条件下的表型分析结果,0.1mm为对照处理,2mm和4mm mnso4为过量锰处理,wt为野生型株系,oe1/oe2为超量表达株系,图中标尺为2cm;图b为超量表达拟南芥株系和野生型拟南芥生物量的比较结果,试验中每个处理设4个生物学重复,图中柱子为4个生物学重复数据的平均值和标准误差;图c为不同浓度锰处理条件下超量表达拟南芥株系和野生型拟南芥中锰浓度的比较结果;图中*号表示超量表达拟南芥株系与野生型拟南芥之间差异显著,p《0.05。
具体实施方式
36.以下结合说明书附图和具体实施例来进一步说明本发明,但实施例并不对本发明做任何形式的限定。除非特别说明,本发明采用的试剂、方法和设备为本技术领域常规试剂、方法和设备。
37.除非特别说明,以下实施例所用试剂和材料均为市购。
38.实施例1柱花草苯丙氨酸解氨酶基因sgpal2的克隆
39.1、柱花草苯丙氨酸解氨酶基因sgpal2的克隆
40.克隆柱花草苯丙氨酸解氨酶基因sgpal2的引物的核苷酸序列如下所示:
41.上游引物:5
’‑
atggcagccattcatccacgtga-3’(seq id no.3)
42.下游引物5
’‑
taaactgagatgggaaccccgtt-3’(seq id no.4)
43.柱花草苯丙氨酸解氨酶基因sgpal2的克隆方法为:
44.参考trnzol universal总rna提取试剂(tiangen)说明书提取热研5号柱花草地上部总rna,然后根据revertaid first strand cdna synthesis试剂盒(thermo fisher)方法合成cdna第一链,以得到的cdna作为模板,利用上述克隆柱花草苯丙氨酸解氨酶基因sgpal2的引物进行pcr扩增。
45.pcr反应体系(20μl)为:2μl 10
×
ex taq buffer,1.6μl dntp mixture,1μl引物(10μm),0.12μl ex taq,1μl cdna模板,加ddh2o补足20μl;反应程序为:94℃1min,94℃30s,58℃30s,72℃120s,循环数35个,72℃延伸10分钟。
46.pcr扩增完成后,配制1%的琼脂糖凝胶(内含golden view核酸染料染色剂),取6μl pcr产物加1μl 6
×
loading buffer进行电泳检测,在凝胶成像系统成像。采用sanprep柱式dna胶回收试剂盒(生工)回收长度为2076bp的pcr产物。
47.将回收得到的pcr产物克隆到trans-t1(全式金公司)载体上进行测序鉴定,获得柱花草苯丙氨酸解氨酶基因sgpal2全长cdna,核苷酸序列如seq id no.1所示,其编码的氨
基酸序列如seq id no.2所示。
48.实施例2柱花草苯丙氨酸解氨酶基因sgpal2的亚细胞定位和表达分析
49.1、载体的构建
50.(1)超量表达载体的构建:
51.以实施例1中获得的柱花草地上部cdna为模板,用上游特异引物5
’‑
ccggggatcctctagaatggcagccattcatccacg-3’(seq id no.5)和下游特异引物5
’‑
gcaggtcgactctagattaaactgagatgggaaccccg-3’(seq id no.6)扩增sgpal2基因orf片段(2076bp);pcr片段经测序确认无误后,通过限制性内切酶xbal i对目的载体进行单酶切,将sgpal2基因连接到目的载体ptf101s并转化大肠杆菌trans1-t(全式金),进行测序分析,测序无误则成功获得ptf101s-sgpal2重组超量表达载体。
52.(2)亚细胞定位分析表达载体的构建:
53.以实施例1中获得的柱花草地上部cdna为模板,用上游特异引物5
’‑
tctagcgctaccggtatggcagccattcatccacg-3’(seq id no:7)和下游特异引物5
’‑
atggtggcgaccggtaactgagatgggaaccccg-3’(seq id no:8)扩增sgpal2基因orf片段,pcr片段测序无误后,将sgpal2基因pcr扩增片段连接到限制性内切酶ageⅰ单酶切线性化目的载体pegad,获得由35s启动子启动的sgpal2-gfp融合表达载体(35s::sgpal2-gfp)。
54.2、sgpal2基因的表达分析
55.本发明用不同的金属以及不同浓度的锰对柱花草种子进行处理后观察不同处理对sgpal2基因表达的影响。
56.具体地,选取热研5号柱花草种子,去种皮并于80℃加热2min,放置于黑暗条件下萌发2~3d后移至hoagland营养液(培养液含400μm nh4no3、1500μm kno3、500μm mgso4·
7h2o、250μm kh2po4、300μm k2so4、1.5μm mnso4·
4h2o、1μm znso4.7h2o、2.5μm na2moo4·
2h2o、0.5μm cuso4·
5h2o、25μm mgcl2、40μm fe-edta、1200μm ca(no3)2,ph 5.8)中预培养两周,分别用800μm fe、400μm mn、20μm zn和10μm cu处理,另用不同浓度的锰(0.5、50、400和800μm)处理;处理两周后收获样品;提取每个处理的柱花草地上部rna。
57.将上述rna反转录成cdna,进一步用定量pcr检测sgpal2的表达,以柱花草的看家基因sgefα作为内参。用于定量pcr检测基因表达量的引物分别如下所示:
58.柱花草sgefα基因的定量引物为:
59.sgefαf:5
’‑
cacttcaggacgtgtacaagatc-3’(seq id no.9)
60.sgefαr:5
’‑
cttggagagcttcatggtgca-3’(seq id no.10)
61.sgpal2基因的定量引物为:
62.sgpal2f:5
’‑
aactctgtcaacgacaaccc-3’(seq id no.11)
63.sgpal2 r:5
’‑
tggatggcaaaccgttgtta-3’(seq id no.12)
64.不同金属和不同浓度锰处理对柱花草地上部中sgpal2基因表达的影响结果如图1所示。其中,图1a为不同金属处理对sgpal2基因表达的影响结果,图中数据为3个重复的平均值和标准误差,*号表示处理与对照之间差异显著,p《0.05。由图1a所示结果可知,相比于对照ck,过量锰处理显著增加了sgpal2在柱花草地上部的表达。在过量锰处理下,柱花草地上部sgpal2表达量是对照的31.8倍。而过量fe、zn和cu处理对sgpal2表达影响不明显。上述结果说明sgpal2特异受过量锰胁迫诱导表达。
65.图1b为不同浓度的锰处理对sgpal2基因表达的影响结果,图中数据为3个重复的平均值和标准误差,*号表示不同锰处理与对照之间差异显著,p《0.05。由图1b所示结果可知,随着锰处理浓度的增加,柱花草sgpal2基因在地上部的表达呈先增加后下降的趋势,且在400μm锰处理下表达量最高,是正常锰处理(0.5μm)的1.6倍。
66.3、sgpal2的亚细胞定位分析
67.通过农杆菌介导的拟南芥原生质体表达体系,将构建得到的sgpal2-gfp融合表达载体和pegad空载体在拟南芥原生质体中进行行瞬时表达,然后用激光共聚焦显微镜(zeiss)观察gfp荧光,明确sgpal2的亚细胞定位。
68.sgpal2基因的亚细胞定位结果如图2所示,图中第一列为gfp荧光信号,第二列为叶绿体自发荧光信号,第三列为明场,第四列为gfp与叶绿体自发荧光信号的融合结果,图中标尺为20μm。由图2所示结果可知,柱花草sgpal2蛋白定位于细胞质中。
69.实施例3转基因实验
70.1、转基因拟南芥的获得
71.将实施例2中构建好的超量表达载体(ptf101s-sgpal2)转化至根癌农杆菌感受态细胞gv3101(唯地生物)中,用农杆菌介导的拟南芥花序侵染转化法进行拟南芥转化,获得转基因拟南芥。
72.2、转基因拟南芥的筛选
73.取合适数量的拟南芥种子于1.5ml离心管中,加入1ml无菌水浸泡1min后接着用75%的酒精灭菌1min,用无菌水清洗3次,最后加入10%的次氯酸钠灭菌10min后用灭菌水清洗3次,酒精和次氯酸钠灭菌过程中需不断颠倒离心管。种子灭菌后置于ms(含有5mg/l草铵膦)培养基培养7~10d,挑选耐除草剂并且正常生长的拟南芥栽培在基质土中,培养两周后提取叶片dna进行pcr验证。
74.20μl pcr反应体系:10μl 2
×
rapid master mix,0.6μl下游引物(10mol/l),0.6μl上游引物(10mol/l),2μl dna模板,加ddh2o至20μl;pcr扩增程序:95℃预变性1min,95℃变性15s,58℃退火30s,72℃120s进行35个循环,最后72℃延伸10min。
75.保留检测正确的拟南芥继续培养至种子成熟,此时种子为t1代种子。按照上诉方法连续筛选3次获取t3代纯合转基因拟南芥种子后保留。pcr确认得到sgpal2基因t3拟南芥种子用于后续实验。
76.3、转基因拟南芥的锰处理:
77.为了进行转基因拟南芥耐锰毒害能力分析,将灭菌后的野生型和转基因拟南芥种子播种在含有0.8%琼脂的ms培养基上培养7d,然后挑选大小一致的幼苗分别移至含0.1mm,2mm和4mm mnso4的固体ms培养基,处理7d后收获植株,测定植株鲜重及锰含量。
78.超量表达sgpal2基因对拟南芥的锰毒害耐受性的影响结果如图3所示。其中,图3a为超量表达拟南芥株系(即转基因拟南芥株系)和野生型拟南芥在不同浓度的锰处理条件下的表型分析结果,0.1mm为对照处理,2mm和4mm mnso4为过量锰处理,wt为野生型株系,oe1/oe2为超量表达株系,图中标尺为2cm。由图3a所示结果可知,在对照0.1mm锰条件下,wt、oe1和oe2转基因株系生长状况一致;而在2mm和4mm过量锰处理下,oe1和oe2转基因株系的植株生长状况明显比wt好,表明超量表达sgpal2可以缓解拟南芥金属锰毒害,提高其对锰毒害的耐受性。
79.图3b为超量表达拟南芥株系和野生型拟南芥生物量的比较,试验中每个处理设4个生物学重复,图中柱子为4个生物学重复数据的平均值和标准误差,*号表示超量表达株系与野生型对照株系之间差异显著,p《0.05。由图3b所示结果可知,在对照0.1mm处理条件下,超量表达sgpal2转基因株系(oe1和oe2)与wt野生型之间的鲜重差异不明显;而在2mm和4mm锰处理下,oe1和oe2植株鲜重显著高于野生型植株鲜重。在2mm过量锰处理下,oe1和oe2植株鲜重分别是wt的1.38和1.4倍;在4mm锰处理下,oe1和oe2植株鲜重分别是wt的2.1和1.8倍。
80.图3c为不同浓度锰处理条件下超量表达拟南芥株系和野生型拟南芥中锰浓度的比较结果,wt为野生型拟南芥,oe1/oe2为超量表达株系,图中*号表示超量表达拟南芥株系与野生型拟南芥之间差异显著,p《0.05。由图3c所示结果可知,在2mm和4mm过锰处理下,与野生型拟南芥wt相比,超量表达sgpal2可以显著降低拟南芥植株中的锰浓度。
81.综上结果表明,在拟南芥中超量表达sgpal2基因可以提高拟南芥对锰毒害的耐受性,其是通过减少植株中锰的积累而增强了植株对锰的耐受性。
82.上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1