小麦抗纹枯病基因TaGST_B3及其应用
小麦抗纹枯病基因tagst_b3及其应用
技术领域
1.本发明属于植物基因工程技术领域,具体涉及一种小麦纹枯病抗性相关基因tagst_b3、其表达产物、其表达载体及应用。
背景技术:
2.由禾谷丝核菌(rhizoctonia cerealis,rc)引起的小麦纹枯病是一种极具破坏性的小麦茎基部真菌病害,对小麦的产量和品质均有很大影响。病害发生区平均减产10%~40%,严重发生时会引起枯孕穗和枯白穗,甚至造成颗粒无收(mcbeath jh,mcbeath j.plant diseases,pests and food security:environmental change and food security in china.2010.;hamada ms,yin yn,chen hg,ma zh.the escalating threat of rhizoctonia cerealis,the causal agent of sharp eyespot in wheat.pest manag sci.2011,67(11):1411-1419.)。据统计,2015至2022年我国小麦纹枯病年平均发生面积约为820万hm2,构成对小麦生产的重大威胁(https://www.natesc.org.cn/)。由于rc具有广泛的寄主范围,且病原菌在土壤或受感染的作物残基上可以长期存活,因此小麦-玉米轮作和秸秆还田的耕作方式导致我国小麦纹枯病愈发严重,已成为我国多个小麦主产区最重要的病害之一。
3.小麦纹枯病是由多基因控制的数量性状(quantitative trait locus,qtl)(chen j,li gh,du zy et al.mapping of qtl conferring resistance to sharp eyespot(rhizoctonia cerealis)in bread wheat at the adult plant growth stage.theor appl genet,2013,126:2865-2878),普通小麦材料中尚无对纹枯病免疫或高抗的种质(ren y,yu pb,wang y,hou wx,yang x,fan jl,wu xh,lv xl,zhang n,zhao l,dong zd,chen f.development of a rapid approach for detecting sharp eyespot resistance in seedling-stage wheat and its application in chinese wheat cultivars.plant dis.2020,104(6):1662-1667.)。当前,对小麦纹枯病的研究多集中在材料的纹枯病抗性鉴定和抗病基因的初步定位上,与其他植物病害相比,如小麦条锈病、白粉病、水稻纹枯病等,小麦纹枯病抗性遗传研究要少得多。自2002年起,利用不同的群体和不同类型的分子标记,约有27个纹枯病抗性qtl被定位,分布在小麦的14条染色体上(蔡士宾,任丽娟,颜伟等.小麦抗纹枯病种质创新及qtl定位的初步研究.中国农业科学,2006,39(5):928-934;chen j,li gh,du zy et al.mapping of qtl conferring resistance to sharp eyespot(rhizoctonia cerealis)in bread wheat at the adult plant growth stage.theor appl genet,2013,126:2865-2878;wu xj,cheng k,zhao rh.quantitative trait loci responsible for sharp eyespot resistance in common wheat ci12633.scientific reports,2017,7(1):11799)。这些qtl主要集中在小麦a和b基因组,d组的qtl相对较少。根据这些标记在中国春基因组中的物理位置(http://202.194.139.32/),可以看出普通小麦的3b染色体上有3个控制纹枯病抗性的qtl。此外,在小麦的1a、2b、5a、6b和7d染色体上也均存在2个以上控制小麦纹枯病抗性的qtl。但目前尚无纹枯病抗性qtl被图位克隆,抗纹枯病
基因的分子机制仍不十分清楚。因此,挖掘和克隆调控纹枯病抗性的重要基因,将丰富人类对作物纹枯病抗性分子机理的认识,为小麦纹枯病抗性基因利用和抗病育种奠定基础。
技术实现要素:
4.本发明提供一种小麦纹枯病抗性相关基因tagst_b3、其表达产物、其表达载体,并将其应用于制备转基因植物细胞和植物育种中,以解决小麦纹枯病抗性相关基因缺乏的技术问题,为小麦育种提供更多的选择。
5.本发明采用如下技术方案:
6.本发明的小麦纹枯病抗性相关基因tagst_b3,其核苷酸序列如seq id no:1所示,具体为:
7.5'-ggtcgacgcgggtgtgggcgagtagaccaggggcgatggcgacgcgctgaccttctcccctcaccggcgatctgctccttctgggtgggggtcgccggctgacgttctgttgcggggtgggggtcgccggctggcgttctgctgcggggtgggagtcgccgaccggcgtgctgctgctaggacaatcggtgaggccagttaggtgctagccgatcgattggcgaagagatccgagtcctggggagatcagtgaggccaggtgctatttggcctatcaattggccaggttctgggaacggggcgtggcgtgatcaacgaggtgctaggctgctagctagggaactggatcctggaacgtggaggaggcaagtccggtatgctaagtactttaactttccttcttcacatccacctgattcagattattttgatctaaattaacttgcaaaaaatatatgtgtgatatccatctactataattgcttacaatcaaaattatatgtgattttttttagtttagaagatttatatgcacagtaaatctgaatgttcttcacatgcatgatttagtttaactttaaagagttatactaactagtcttgataaagagatcttttggagccaacaccaaacctcgtgaggtgttttgcctacggaaaggttgtgttatgtaatgattattattaggatcaaagttgtaggataaacgtaaaaccttctcgatgtatcttttatacaacattgtagtttagttatatatggagagagtgatttaacactttgtgtttaagagtagaataagttattccacactctagccaaacgaactatttggcaaatatctggctagctggtgagagccagagccgtggaaagtctgtcttgctattaaggcacaagcatcaaacaggaacatttagagccatggaaaagtgatgtgtcgcctaccaatgggccaactgctagcgatgtaataatagcatccaagttgattttttatagaacatgcaaggcgttggcaagtgggaaaatgattgatcgctggcaagcttaactctcggaacttgtagcattcaactgaatcagaacaaagattttaaaaaaatacatttccatcgatagtgaaaaattattcaattgagtaacaacgaaaatcatattggaatgtacatttacttgttgattttaaattagaggcatttttctaccttttttagttaataagatatgcatatacccacccttagtgttttcgagacaacgagagggcacattgcttttggtgctaccatctctctcaagcctcaaataagttgtgcggacacgattatcttcccgcgttggaatatcgtggcctggtagagctagcgaaaaatcttccatgttggaatatgtcggcagccggatagccgccatgcatgtaaagtctcttttacctttacacttgctcaagtgacactgtatgtcgcctaccacttgctaaatcaatgggccaactgctagcgacgtaatagtagcaagttgatttacagtgttttgctacagttctctgactttgtttcttcattttagactagctgactactgtcgcttacctgccttcccttctccacgttagaggatccagttctgatgttgagacctcgacgatgggaggaagggcgcgatcgatgtggagtaatttgaatttcaaatctatctatctggggtatattggtccttcaccgatgttttgggggctgtcggaaattggttccgcgatctacaaaagtgaatggagggagtaattgtttctccaatccgtaccaacgcacgtgtttctaactagtacttccttcgcaccacaatatggaatagagggagtatcgataaactaacaaagatgattacttacccggtttaaatgattcaagagctcatttaatttggcactcatcatttcatatatcttttttggtagaaatgaaataaagcagatctagacactagctaaaaagtcgatgtagccttgttatttccttgggccacgcgggccgggtgtggtgctccctgctctgtgtataaatggagatcaacatccaaggcctcctcccacacacacacgctacagagcagagcagagtcttgctccagtatctgccctctcctgcctgcctgtagagcatccatcacgtgaagttcacggacaaaca
tgtctccggtgaaggtgttcgggcacccgatgttgacaaacgtcgcacgggtgctgctcttcctggaggaggtcggcgccgagtacgagctcgtgcccatggacttcgtcgccggcgagcacaagaggccccaacacgtccagctaaacgtaagcagacgacaattcgtcgatcggtcaccgtatcaggttcaggtgatgatttctgacagagatgctacgtgtttttcttccttgcagccgtttgcgaagatgcctgggttccaagatggcgatctcgtcctgttcggtgagccgtctaacacatcagtgcacacgacacgaatcattcctgtctaggagcatatatcatgttaaaaatccattggttggtttctctaatctaacaaatcgatgagtgagtgagtgcagagtcgcgcgccatcgccaagtatatcctccgcaagtacggggggacagccggcctggacctcctcggagaaaacagcggaatcgaagaattagcaatggtggacgtgtggacggaggtggaggcccagcagtactacccggccatctcgcccgtggtgttcgagtgcatcatcatccccttcatcatccctggcggtggcgcggcgccgaaccagaccgtcgtggacgagagcctggagcggctgaggggtgtgctggggatctacgaggcccggctggagaagagcaggtacttggccggggactccatcagcttcgccgatctgaaccacatcccgttcaccttctacttcatgaccaccccgtacgccaaggtgtttgatgactaccccaaggtgaaggcctggtgggagatgctcatggcgaggccggcggtgcagagggtctgcaaccatatgcctaccgagtttaagctaggtgcgcagtactaggtgtttatgcactccgcgcgggtgtgtgccgtgccgtgccgtgccgtgccggtgatgattccgttcaagagaagagacctccctcccgtgggtgggtgatcatctgcccgtctacttccagtagtatacgctttcagtttcagcagcacgaggagtccgcttgcgcttctgttcgtgcgctggattccttgctttaatttatgcactagtactagtaaataaacaacggactggtcaatgctgctaatccgtactgtacgtcctatatagtagtaccccactagagatcagtatatatatacatcttggatacgtgtgcatttgcatgcaaatatatatggacagggataactggcacatgtgtgcccgttaggaaaattaccatcgctctccttcggtcggtggcccgctcgctcgatcgagcgttccttcggtcggtgtcccgctcgcccttcggtccgtggcccgctcgctcgatcgagcgttccttcggtcggtgtcctgctcgctcgatcgagcgttgaggccagttggctcgatcgctccgctttgatcgagttgtgaatgctttcatcggttttcttttcttctgatttttattttcagaacggtctaccaaatagtcactacacccaataaccaactcggccatcctaccttgttatccacaatacgtgaacaacggaacttgaattcttcttctttttaccatgtacacaaaaaggttaattttagccttgataactttggcatgagcagcatgataactatgacatatgcatcatgattactttaatattaacacctggtaaccaaaaaattgacatgataaagcgggtaaatatatcagagaaggtc-3'。
8.基因tagst_b3的全长cdna核苷酸序列如seq id no:2所示,具体为:
9.5'-gtataaatggagatcaacatccaaggcctcctcccacacacacacgctacagagcagagcagagtcttgctccagtatctgccctctcctgcctgcctgtagagcatccatcacgtgaagttcacggacaaacatgtctccggtgaaggtgttcgggcacccgatgttgacaaacgtcgcacgggtgctgctcttcctggaggaggtcggcgccgagtacgagctcgtgcccatggacttcgtcgccggcgagcacaagaggccccaacacgtccagctaaacccgtttgcgaagatgcctgggttccaagatggcgatctcgtcctgttcgagtcgcgcgccatcgccaagtatatcctccgcaagtacggggggacagccggcctggacctcctcggagaaaacagcggaatcgaagaattagcaatggtggacgtgtggacggaggtggaggcccagcagtactacccggccatctcgcccgtggtgttcgagtgcatcatcatccccttcatcatccctggcggtggcgcggcgccgaaccagaccgtcgtggacgagagcctggagcggctgaggggtgtgctggggatctacgaggcccggctggagaagagcaggtacttggccggggactccatcagcttcgccgatctgaaccacatcccgttcaccttctacttcatgaccaccccgtacgccaaggtgtttgatgactaccccaaggtgaaggcctggtgggagatgctcatggcgaggccggcggtgcagagggtctgcaaccatatgcctaccgagtttaagctaggtgcgcagtactaggtgtttatgcactccgcgcgggtgtgtgccgtgccgtgccgtgccgtgccggtgatgattccgttcaagagaagagacctccctcccgtgggtgggtgatcatctgcccgtctacttccagtagtatacgctttcagtttcagcagcacgaggagtccgcttgcgcttctgttcgtgcgctggattccttgctttaatttatgcactagtactagtaaataaacaacgga
ctggtcaatgctgctaatccgtactgtacgtcctatatagtagtaccccactagagatcagtatatatatacatcttggatacgtgtgcatttgcatgcaaatatatatggacagggataactggcacatgtgtgcccgttaggaaaat-3'。
10.基因tagst_b3的cds序列如seq id no:3所示,具体为:
11.5'-atgtctccggtgaaggtgttcgggcacccgatgttgacaaacgtcgcacgggtgctgctcttcctggaggaggtcggcgccgagtacgagctcgtgcccatggacttcgtcgccggcgagcacaagaggccccaacacgtccagctaaacccgtttgcgaagatgcctgggttccaagatggcgatctcgtcctgttcgagtcgcgcgccatcgccaagtatatcctccgcaagtacggggggacagccggcctggacctcctcggagaaaacagcggaatcgaagaattagcaatggtggacgtgtggacggaggtggaggcccagcagtactacccggccatctcgcccgtggtgttcgagtgcatcatcatccccttcatcatccctggcggtggcgcggcgccgaaccagaccgtcgtggacgagagcctggagcggctgaggggtgtgctggggatctacgaggcccggctggagaagagcaggtacttggccggggactccatcagcttcgccgatctgaaccacatcccgttcaccttctacttcatgaccaccccgtacgccaaggtgtttgatgactaccccaaggtgaaggcctggtgggagatgctcatggcgaggccggcggtgcagagggtctgcaaccatatgcctaccgagtttaagctaggtgcgcagtactag-3'。
12.本发明的小麦纹枯病抗性相关基因编码的蛋白tagst_b3,其氨基酸序列如seq id no:4所示,具体为:
13.mspvkvfghpmltnvarvllfleevgaeyelvpmdfvagehkrpqhvqlnpfakmpgfqdgdlvlfesraiakyilrkyggtagldllgensgieelamvdvwteveaqqyypaispvvfeciiipfiipgggaapnqtvvdeslerlrgvlgiyearleksrylagdsisfadlnhipftfyfmttpyakvfddypkvkawwemlmarpavqrvcnhmptefklgaqy。
14.鉴定上述小麦纹枯病基因tagst_b3的特异性引物对,所述特异性引物对包括:
15.核苷酸序列如seq id no:5所示的上游引物和核苷酸序列如seq id no:6所示的下游引物。
16.利用上述特异性引物检测小麦纹枯病等位基因型的方法,包括如下步骤:
17.步骤1:提取待测小麦的基因组dna;
18.步骤2:以待测小麦的基因组dna为模板,用核苷酸序列如seq id no:5所示的上游引物和seq id no:6所示的下游引物进行pcr扩增,得到扩增产物,然后进行如下判断:
19.判断扩增产物自启动子区atg前第363位有4个碱基的idel以确定基因类型;其中,缺失类型(
‑‑‑‑
),其对应等位基因型命名为tagst_b3a;插入类型(tact),其对应等位基因型命名为tagst_b3b。
20.进一步表型鉴定结果显示,缺失类型(
‑‑‑‑
)植株对应病情指数较低,植株抗病性较强,插入类型(tact)植株对应病情指数较高,植株抗病性较弱;因此,tagst_b3基因对小麦纹枯病的抗性具有十分重要的作用,同时也有助于小麦抗病育种的遗传和分子生物学的更深入研究。
21.本发明还设计一种由上述基因构建的重组表达载体,以及由上述的重组表达载体构建的重组菌。
22.重组表达载体具体可为在植物表达载体plgy-oe3-gai的酶切位点bamh i和saci之间插入seq id no:3所示的核酸序列得到的重组质粒。
23.植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。所述植物表达载体还可包含外源基因的3
′
端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加
工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3
′
端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂合成酶nos基因)、植物基因(如大豆贮存蛋白基因)3
′
端转录的非翻译区均具有类似功能。使用目的基因构建重组植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型启动子或组成型启动子,如花椰菜花叶病毒(camv)35s启动子、玉米的泛素启动子(ubiquitin),它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
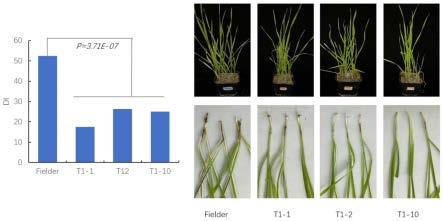
24.本发明还提供上述tagst_b3基因在制备转基因植物中的应用。即将所述基因导入目的植物中,得到可调节纹枯病抗性的转基因植物。携带有所述基因的表达载体可通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导、基因枪等常规生物学方法转化植物细胞或组织,并将转化的植物组织培育成植株。
25.所述目的植物既可以是单子叶植物也可以是双子叶植物。所述单子叶植物具体可为小麦,如小麦fielder。
26.同样的,还可将上述基因和/或上述蛋白在植物育种中应用。
27.本发明的有益效果为:
28.1.本发明首次公开、并确认了一种新的小麦纹枯病抗性相关基因,命名为tagst_b3,该基因位于小麦染色体3b上,其表达的蛋白可调控小麦纹枯病抗性。
29.2.本发明明确了小麦纹枯病抗性相关基因tagst_b3的dna序列,cds序列和编码蛋白序列,为小麦抗纹枯病育种的应用实践打下基础。
30.3.本发明中的特异性引物seq id no:5所示的上游引物和seq id no:6所示的下游引物,可直接用于小麦纹枯病抗病基因型的检测,有益于小麦抗病品种的培育和优异种质资源的创制,实现早期筛选,节省时间和资源。
31.4.本发明首次利用转基因技术将小麦纹枯病抗病基因tagst_b3转入普通小麦进行过表达,验证了tagst_b3基因在小麦纹枯病抗病育种中的作用。
32.5.本发明有助于揭示小麦纹枯病的分子遗传基础,同时对利用基因工程技术检测和改良小麦品种起到重要作用,为抗病、安全的小麦新品种培育提供了新途径。
附图说明
33.图1为pm群体小麦纹枯病病情指数(di)的频次分布图。图中,a:纹枯病田间di;b:纹枯病温室di;di:病情指数;i:重复1;ii:重复2。
34.图2为小麦纹枯病抗性的qtl定位图。
35.图3为tagst_b3、tafaoα和tapef1在抗、感池中的表达水平。图中,rb1为抗池1,rb2
为抗池2;sb1为感池1,sb2为感池2。
36.图4为接种5d后tagst_b3在抗病亲本平原50和感病亲本铭贤169中的相对表达量。bjj表示不接种纹枯菌,jj表示接种纹枯菌。
37.图5为tagst_b3在抗病亲本平原50和感病亲本铭贤169间的序列多态性。
38.图6为vigs沉默试验中tagst_b3基因的相对表达量柱图。
39.图7为vigs沉默试验中不同处理材料对小麦纹枯病的抗性对比照片。
40.图8为野生型kwt和tagst_b3突变体(k1027)的在苗期和成株期纹枯病抗性的表型比较图。图中tagst_b3表示tagst_b3基因突变体(k1027)。
41.图9为plgy-oe3-gai载体质粒图谱。
42.图10为过表达转基因株系(t1-1、t1-2和t1-10)与空载对照(fielder)的表型比较图。
43.图11为tagst_b3基因在过表达转基因系(t1-1,t1-2,t1-10)和野生型fielder中的表达水平。
具体实施方式
44.下面通过具体实施方式对本发明进行更加详细的说明,以便于对本发明技术方案的理解,但并不用于对本发明保护范围的限制。
45.在以下实施例中所涉及的仪器设备如无特别说明,均为常规仪器设备;所涉及的试剂如无特别说明,均为市售常规试剂;所涉及的试验方法,如无特别说明,均为常规方法。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。引物合成及测序工作均由生工生物工程(上海)股份有限公司完成。
46.以下所涉及的小麦材料为:
47.pm群体:由平原50和铭贤169杂交f1代经秋水仙素诱导加倍构成的包含117个家系的dh群体,此群体用于qtl定位和验证tagst_b3等位基因效应;
48.平原50:基因型与表型属抗小麦纹枯病材料,用于vigs试验验证tagst_b3基因效应;
49.kronos突变体:野生型kronos-wt和kronos1027(tagst_b3基因错义突变)的突变体系,用于比对tagst_b3基因效应;(kronos突变体材料来自美国加州大学戴维斯分校jorge dubcovsky院士课题组创建的四倍体小麦kronos的突变体库(krasileva等,pnas,2017,114:913-921))。
50.fielder:基因型与表型属感小麦纹枯病材料,为过表达转tagst_b3基因的受体材料。
51.实施例1:小麦纹枯病相关基因tagst_b3的定位和获得
52.利用温室病麦粒混土法及田间牙签接种法(ren yan et al.,development of a rapid approach for detecting sharp eyespot resistance in seedling-stage wheat and its application in chinese wheat cultivars.plant disease,2020,104(6):1662-1667)对pm群体进行苗期及乳熟期纹枯病抗性鉴定,表型鉴定结果显示,该dh群体的纹枯病di在乳熟期和苗期均呈现连续性分布并有超亲分析(图1),表明平原50所含有的纹枯病抗性基因属于典型的数量性状遗传。
53.根据pm群体田间和温室的表型数据,挑选极端抗、感家系构建抗感池,利用小麦660ksnp芯片(sun congwei et al.,the wheat 660k snp array demonstrates great potential for marker-assisted selection in polyploid wheat,plant biotechnology journal,2020(18):1354-1360)进行双亲及混池的bsa分析,获得在双亲及抗、感混池间一致性多态性的差异snp,主要位于3b和6b染色体上,进而,在上述定位区域均匀开发35个kasp标记,进行群体基因型分析,并结合ssr标记的检测信息,最终在小麦的3b染色体上检测到一个纹枯病抗性相关的qtl位点,命名为qse.hnau-3bs(图2),位于kasp标记a011839至a011841之间,对应物理区段约7.4mb。表型贡献率分析表明,无论乳熟期还是苗期该qtl均能显著降低pm群体的di值(表1)。该qtl前人未报道,是一个新的纹枯病抗性qtl位点。
54.表1存在和不存在qse.hnau-3bs对纹枯病抗性的影响
[0055][0056]
进一步结合pm群体中抗感池中的个体田间接种小麦纹枯病菌后的转录组结果,发现在上述qtl的定位区段内有3个差异表达基因(tagst_b3、tafaoα和tapef1),但只有一个注释为谷胱甘肽转移酶的基因(gst)tagst_b3在抗池中显著上调表达(图3),而另外2个基因在抗池中显著下调表达。荧光定量的结果也表明接种5天后,tagst_b3在抗病亲本平原50中的表达量显著高于感病亲本铭贤169(图4)。因此将该基因预测为小麦纹枯病抗性相关基因。
[0057]
参考primer5设计2对pcr扩增引物对扩增tagst_b3基因的启动子区和编码区序列。
[0058]
引物序列如下:
[0059]
引物对1:
[0060]
seq id no:5(tagst-f1):5'-gcgaggtgactggatgacg-3';
[0061]
seq id no:6(tagst-r1):5'-acggctcaccgaacaggac-3'。
[0062]
引物对2:
[0063]
seq id no:7(tagst-f2):5'-cttacttccttcgcaccaca-3';
[0064]
seq id no:8(tagst-r2):5'-acctatcgtgcttctgatgtt-3'。
[0065]
引物对1扩增的是tagst_b3基因启动子区序列和少部分基因编码区序列,引物对2扩增的是整个基因编码区序列。
[0066]
利用上述2对引物对小麦平原50和铭贤169等材料的基因组dna进行扩增,pcr扩增体系见表2,pcr扩增程序见表3。
[0067]
表2引物1和引物对2对tagst_b3基因的pcr扩增体系
[0068]
组分体积2
×
phanta buffer12.5μl
dntp0.5μlphanta enzyme0.5μlprimer f0.5μlprimer r0.5μldna2μlddh2oto 25μl
[0069]
表3引物1和引物对2对tagst_b3基因pcr扩增程序
[0070][0071]
将抗病亲本平原50和感病亲本铭贤169的tagst_b3基因的启动区及编码区的序列进行比较,发现抗、感亲本在编码区的多态性均未造成氨基酸的改变,但双亲启动子区存在丰富的多态性(图5)。平原50的tagst_b3基因的核苷酸序列如seq id no:1所示,全长cdna核苷酸序列如seq id no:2所示。
[0072]
实施例2:tagst_b3基因特异标记的检测
[0073]
以pm群体的5个极抗材料及5个极感材料的dna为模板,用核苷酸序列如seq id no:5所示的上游引物和seq id no:6所示的下游引物进行pcr扩增,得到扩增产物,然后进行如下判断:
[0074]
判断扩增产物自启动子区atg前第363位有4个碱基的idel以确定基因类型;其中,缺失类型(
‑‑‑‑
),其对应等位基因型命名为tagst_b3a;插入类型(tact),其对应等位基因型命名为tagst_b3b。
[0075]
结果显示5个极抗dh系的基因型均为tagst_b3a,5个极抗dh系的基因型均为tagst_b3b类型。
[0076]
实施例3:tagst_b3基因vigs沉默试验及其表型鉴定
[0077]
根据tagst_b3a基因的cdna序列设计引物,分别在正、反向引物的5’端加上酶切位点和保护碱基,形成上游引物:tagst-f3:5'-tttttttagctagctgattaattaacccaacacgtccagctaaac-3'和下游引物:tagst-r3:5'-cttccgttgctagctgagcggccgccggaggatgtacttggcgat-3'。利用此引物对扩增tagst_b3基因的vigs沉默片段并回收备用。
[0078]
进一步将tagst_b3基因的vigs沉默片段和γ-pds-as载体用pac1和not1双酶切,回收沉默片段和对应γ线性化载体片段,利用无缝克隆方法完成重组载体构建(γ-tagst_b3)。
[0079]
(2)tagst_b3基因的植物vigs沉默试验
[0080]
将病毒载体(α、β、γ、γ-pds)和tagst_b3基因沉默重组载体(γ-tagst_b3)质粒
分别酶切线性化,进一步用ribomaxtm large scale rna production systems-t7试剂盒将线性化的质粒体外转录,获得不同组分的病毒体外转录产物。取体外转录产物α、β、γ/γ-pds/γ-dir各2.5μl,按照1:1:1的比例混合并用depc水等体积稀释,取5μl稀释好的混合液加入到90μl fes缓冲液中充分吸打混匀。每次试验设置4组不同处理,分别为:完全空白对照组(wt),病毒空白对照组(α+β+γ),白化阳性对照组(α+β+γ-pds)和基因沉默组(α+β+γ-tagst_b3)。
[0081]
病毒侵染时,在要侵染的植株(平原50)表面先喷上少量depc水,取8~10μl fes混合液点于干净手套上,从叶片基部到叶尖处摩擦幼苗第二叶叶片3次,摩擦时控制好力度,摩擦完后从上往下喷少许depc水保持湿度,每个处理均需换干净手套操作。病毒接种完后置于23
±
2℃培养箱中保温保湿避光培养24h,后调为16h/8h光暗周期培养,定期观察并记录表型变化。
[0082]
(3)tagst_b3基因植物vigs沉默后表型鉴定
[0083]
vigs沉默2周后,用qrt-pcr技术检测tagst_b3基因在沉默植株(bsmv
tagst_b3
)、病毒空载植株(bsmv0)和未沉默植株(wt,平原50)中的相对表达情况,如图6所示,结果表明沉默植株中tagst_b3基因的相对表达量显著下调,tagst_b3基因被有效沉默。
[0084]
进一步对沉默植株进行纹枯病抗性调查,结果显示,tagst_b3基因沉默植株(di=80)比病毒空载植株(平原50,di=67)的病情指数显著升高,植株抗病能力显著下降(见图7)。
[0085]
综合来看,tagst_b3基因沉默后植株抗病能力显著降低,说明此基因是小麦纹枯病响应过程中的一个正向调控基因。
[0086]
实施例4:ems突变体的检测
[0087]
进一步利用四倍体kronos突变体来验证tagst_b3基因的功能,kronos1027在3b染色体上对应tagst_b3基因序列自5’端atg后10bp处核苷酸由g突变为a,从而引起基因错义突变。对突变体kronos1027和野生型kronos-wt进行小麦纹枯病苗期及成株期抗性检测,结果显示,在苗期与kronos-wt(di=60)相比,tagst_b3基因突变体植株kronos1027(di=90)的病情指数显著升高,植株抗病能力明显降低(见图8),推测tagst_b3基因在植物体内起正调控作用。
[0088]
实施例5:tagst_b3转基因小麦的获得及鉴定
[0089]
(1)tagst_b3植物过表达载体的构建
[0090]
根据小麦平原50中tagst_b3的全长cds序列(seq id no:3所示)设计引物,并在引物两端分别引入限制性内切酶bamh i和sac i识别位点及保护碱基,引物序列如下:
[0091]
tagst-f4:5'-gatgacgatgacaagggatcctctccggtgaaggtgttcg-3';
[0092]
(带下划线碱基为限制性内切酶bamh i识别位点及保护碱基);
[0093]
tagst-r4:5'-acgaacgaaagctctgagctcctagtactgcgcacctagctta-3';
[0094]
(带下划线碱基为限制性内切酶sac i识别位点及保护碱基)。
[0095]
利用trizol试剂提取平原50苗期总rna,以此rna为模板,使用superscript iii反转录酶(invitrogen,cat no.18080-044)进行反转录得到cdna,以此cdna为模板,利用引物tagstf-f4和tagstf-r4进行pcr扩增小麦tagst_b3基因的编码序列。
[0096]
表4引物tagst-f4和tagst-r4对tagst_b3基因的pcr扩增体系
[0097][0098][0099]
pcr反应程序为:
[0100]
95℃5min;94℃30s,60℃30s,72℃30s,35cycles;最后68℃延伸10min。
[0101]
反应结束后,对扩增产物进行1%琼脂糖凝胶电泳检测,回收并纯化690bp的dna片段进行测序,测序结果表明,扩增得到的dna片段与seq id no:3所示序列一致。
[0102]
表达载体选择plgy-oe3-gai,其载体图谱如图9所示。
[0103]
将上述690bp的dna片段克隆入植物表达载体plgy-oe3-gai的酶切位点bamh i和sac i之间,得到含有小麦tagst_b3基因的重组表达载体,命名为plgy-oe3-gai-tagst_b3。
[0104]
(2)tagst_b3转基因小麦的获得
[0105]
将plgy-oe3-gai-tagst_b3用农杆菌浸染法转化小麦fielder的幼胚愈伤组织,经过筛选、预分化、分化得到阳性植株。将阳性植株利用引物tagstf-f5和tagstf-r5进行pcr鉴定,得到t0代转基因植株。
[0106]
引物序列如下:
[0107]
tagst-f5:5'-ttagccctgccttcatacgc-3';
[0108]
tagst-r5:5'-tgcggaggatatacttggcg-3'。
[0109]
扩增体系和pcr程序:
[0110]
表5引物tagstf-f5和tagstf-r5对tagst_b3基因的pcr扩增体系
[0111]
组分体积2
×
phanta buffer12.5μldntp0.5μlphanta enzyme0.5μlprimer f0.5μlprimer r0.5μldna2μlddh2oto 25μl
[0112]
表6引物tagst-f5和tagst-r5对tagst_b3基因pcr扩增程序
[0113][0114]
(3)转基因小麦的pcr鉴定和表型鉴定
[0115]
利用tagst-f5和tagst-r5引物对进行pcr筛选阳性转基因植株,共获得10个阳性转基因植株,经过加代,获得t1代转基因株系(t1-1、t1-2和t1-10)。
[0116]
过表达转基因株系t1(t1-1、t1-2和t1-10)与野生型fielder在温室的纹枯病接种鉴定的表型如图10所示:
[0117]
同野生型fielder植株比较,过表达转基因株系t1(t1-1、t1-2和t1-10)的纹枯病病情指数显著降低,植株抗病能力显著提高。
[0118]
利用tagst-f6和tagst-r6引物,荧光实时定量pcr检测基因tagst_b3在t1代阳性转基因植株叶片中的表达量。
[0119]
引物序列如下:
[0120]
tagst-f6:5'-tcgtcctgttcggtgagc-3';
[0121]
tagst-r6:5'-taattcttcgattccgctg-3'。
[0122]
扩增体系和pcr程序:同表5和表6。
[0123]
荧光实时定量pcr结果如图11所示,目的基因tagst_b3在t1代阳性转基因植株叶片中的表达量显著高于野生型fielder,是空载叶中表达量的5~14倍左右。
[0124]
综上,本发明的小麦纹枯病抗性相关基因tagst_b3表达的蛋白可调控小麦纹枯病抗性,可用于研究控制小麦纹枯病相关的分子机理研究,在小麦抗病育种方面有着重要的应用价值。
[0125]
以上所述之实施例,只是本发明的较佳实施例而已,并非限制本发明的实施范围,故凡依本发明专利范围所述的构造、特征及原理所做的等效变化或修饰,均应包括于本发明申请专利范围内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1