光敏核隐性雄性不育基因Ghpsm5及其在棉花中的应用
本发明属于基因工程和农业作物遗传育种,具体涉及光敏核隐性雄性不育基因ghpsm5及其在棉花中的应用。
背景技术:
1、棉花是世界上最主要的农作物之一,目前有七十多个国家种植,中国、印度和美国是主要的棉花生产国。提高棉花产量和品质是棉花科学研究的主要任务。杂种优势是一种被人类在农业上应用最为成功的生物学现象,棉花也是一种杂种优势十分明显的经济作物。棉花杂交种具有营养生长优势、产量优势和耐高温、耐湿、耐旱、耐瘠、耐病等抗逆优势,因此杂交种应用成为棉花育种的重要手段。美国于20世纪40年代便开始了棉花杂种优势利用的相关研究,直到70年代培育出具有哈克尼西棉的胞质雄性不育系,并实现了三系配套。中国杂交棉育种发展迅速,利用胞质雄性不育系的三系法、隐性核雄性不育系的两系法及人工去雄辅助授粉方法均有应用。我国杂交棉利用在2007年达到高峰,其中长江流域棉区90%以上面积为杂交棉,但杂交种主要是通过人工去雄辅助授粉方法制种。2007年之后随着劳动力成本增高,杂交棉种子成本急剧攀升,杂交棉种植面积迅速减少。水稻的“三系法”“两系法”杂交水稻的相继成功已经产生了巨大的社会效益和经济效益,而棉花“胞质互作不育系”的恢复系筛选困难,没有很好的优势组合;棉花核不育系制种时要拔除50%的可育株,其制种产量又受到限制,因此棉花杂交种的生产仍以人工去雄制种为主。创制既能作为不育系应用,自身又能繁殖的“两用系”材料是解决棉花杂交种应用的关键技术,也是世界难题。
2、棉花的核不育系发现的种类较多,到目前为止,一共发现了17个不同类型的核不育系,包括9个隐性核不育系,分别是ms1、ms2、ms3、ms5ms6、ms8ms9、ms13、ms14、ms15和ms16;8个显性核不育系,分别是ms4、ms7、ms10、ms11、ms12、ms17、ms18以及ms19。这17种不育系中,12个是在陆地棉中发现的,5个发现于海岛棉中;ms2和ms4等为完全不育,ms1和ms3等仅是部分不育,ms8ms9则表现为花药不开裂。其中ms5、ms8和ms9等有分子标记定位在染色体上。核雄性不育基因的研究处于寻找标记的阶段,未见上述相关基因的专利和报道。
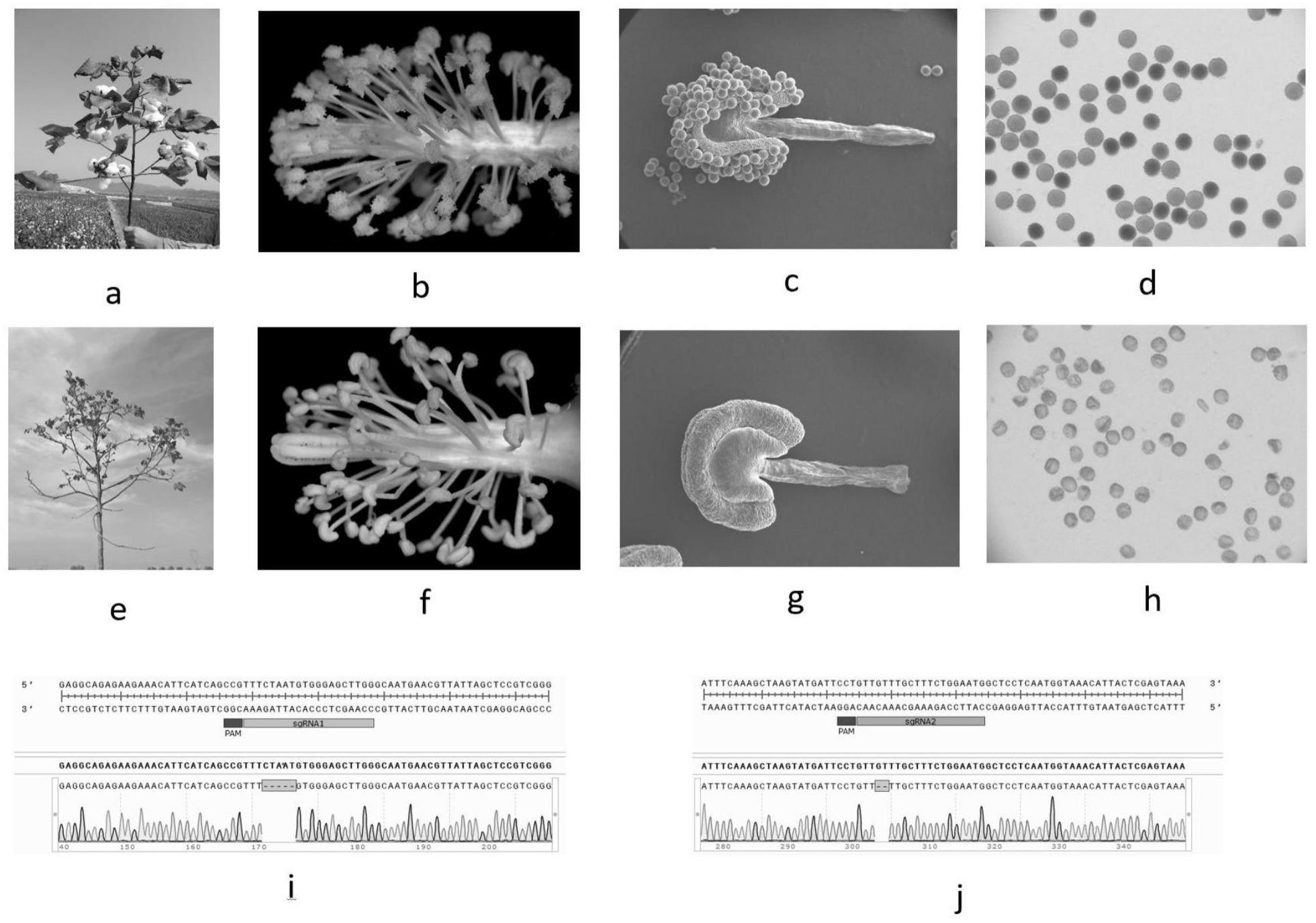
3、中国农业科学院棉花研究所利用航天诱变育种技术,培育棉花芽黄光敏雄性不育突变体中9106,在光照周期大于13.5h的长日照条件下表现为雄性不育不育,在光照周期小于13.5小时的短日照条件,且日平均温度大于等于21.5℃的条件下表现可育。张朝军应用棉花材料w10进行大量组织培养,获得了一个光敏核隐性雄性不育突变体psm4(zl201810132189.8);并获得了与光敏核隐性雄性不育性状相关的分子标记(zl202010869171.3)。通过对分子标记关联区间的基因功能验证,找到了控制棉花光敏核隐性雄性育性的基因ghpsm5;通过对ghpsm5基因的编辑,获得光敏核隐性雄性不育材料ps201。
技术实现思路
1、本发明的目的在于提供光敏核隐性雄性不育基因ghpsm5及其在棉花中的应用,基于所述基因和/或启动子,利用光周期的变化改变植物育性,从而创制棉花或其他植物光敏核隐性雄性不育材料。
2、本发明提供了一种棉花光敏核隐性雄性不育基因ghpsm5,所述光敏核隐性雄性不育基因ghpsm5编码的蛋白质的氨基酸序列包括seq id no.2所示的序列,或与seq id no.2所示的氨基酸序列具有75%以上同一性的蛋白质。
3、本发明还提供了一种棉花光敏核隐性雄性不育基因ghpsm5,所述光敏核隐性雄性不育基因ghpsm5的核苷酸序列包括seq id no.1所示,或对seq idno.2所示蛋白质经过取代和/或缺失和/或添加一个或几个氨基酸残基得到的具有调控花药开裂功能的衍生蛋白质的核苷酸序列。
4、本发明还提供了一种调控上述棉花光敏核隐性雄性不育基因ghpsm5表达的启动子,所述启动子的核苷酸序列如seq id no.3所示。
5、本发明还提供了上述棉花光敏核隐性雄性不育基因ghpsm5或上述启动子在创制植物光敏核雄性不育材料中的应用。
6、优选的,所述植物光敏核隐性雄性不育材料在日照时数大于等于12.5小时条件下花粉发育异常,花药不开裂,造成雄性不育;在日照时数小于等于12.0小时时,花粉发育恢复正常,花药正常开裂,恢复雄性育性;日照时数在高于12.0小时但低于12.5小时时,接近花朵基部的部分花药偶然开裂散粉,散出的花粉活力正常,但数量较少(一般低于5个花药),很难自交结铃,仍可用于杂交制种;在长日照条件下开花的光敏核隐性雄性不育棉花材料,转入短日照处理15-18天后,可转变为雄性可育,具体转化时间受蕾发育速度影响,ghpsm5基因编辑材料ps201在安阳8月份处理时转化时间为17天;在短日照条件下开花的棉花光敏核隐性雄性不育材料,表现为正常可育,转入长日照条件种植15-18天,可转变为雄性不育,具体转化时间受蕾发育速度影响,ps201在安阳8月份处理时转化时间为17天。
7、本发明还提供了一种植物光敏核隐性雄性不育材料的创制方法,包括调控和/或改变上述棉花光敏核隐性雄性不育基因ghpsm5的活性和\或表达规律。
8、优选的,所述调控和/或改变的方法,包括基因编辑、rnai、反义rna和dna甲基化中的一种或多种。
9、优选的,所述调控和/或改变的方法,包括通过基因工程方法,利用上述启动子作为启动子构建表达载体,表达能够对雄性器官发育发生作用的蛋白质、rna和/或dna序列。
10、优选的,所述雄性器官包括花药。
11、本发明还提供了依托上述创制方法得到的光敏核隐性雄性不育材料在植物育种中的应用。
12、有益效果:本发明提供了一种棉花光敏核隐性雄性不育基因ghpsm5,可通过改变正常棉花植株中ghpsm5基因的序列,使其不能正常表达,可以导致棉花在日照时数大于等于12.5小时的长日照条件下花粉无活性,花药不开裂导致雄性不育;日照时数小于等于12.0小时的短日照条件下花粉发育正常,花药开裂正常而恢复雄性育性;日照时数在高于12小时但低于12.5小时时,接近花朵基部的部分花药可以开裂散粉,散出的花粉活力正常,但数量较少(一般低于5个花药),很难自交结铃,仍可用于杂交制种。
13、改变该基因对雌性育性无影响,且ghpsm5与psm1的低温黄化无关,因此通过生物技术阻止该基因的表达,可以创制棉花光敏核隐性雄性不育系,用于棉花育种。
技术特征:
1.一种棉花光敏核隐性雄性不育基因ghpsm5,其特征在于,所述光敏核隐性雄性不育基因ghpsm5编码的蛋白质的氨基酸序列包括seq id no.2所示的序列,或与seq id no.2所示的氨基酸序列具有75%以上同一性的蛋白质。
2.一种棉花光敏核隐性雄性不育基因ghpsm5,其特征在于,所述光敏核隐性雄性不育基因ghpsm5的核苷酸序列包括seq id no.1所示,或对seq idno.2所示蛋白质经过取代和/或缺失和/或添加一个或几个氨基酸残基得到的具有调控花药开裂功能的衍生蛋白质的核苷酸序列。
3.一种调控权利要求1或2所述棉花光敏核隐性雄性不育基因ghpsm5表达的启动子,其特征在于,所述启动子的核苷酸序列如seq id no.3所示。
4.权利要求1或2所述棉花光敏核隐性雄性不育基因ghpsm5或权利要求3所述启动子在创制植物光敏核隐性雄性不育材料中的应用。
5.根据权利要求4所述应用,其特征在于,所述植物光敏核隐性雄性不育材料在日照时数大于等于12.5小时条件下花粉无活性,花药不开裂,造成雄性不育;日照时数在高于12小时但低于12.5小时时,接近花朵基部的少量花药能够开裂散粉,散出的花粉活力正常,但数量较少,很难自交结铃,仍可用于杂交制种;在日照时数小于等于12.0小时时,花粉发育恢复正常,花药正常开裂,恢复雄性育性。
6.一种植物光敏核隐性雄性不育材料的创制方法,其特征在于,包括调控和/或改变权利要求1或2所述棉花光敏核隐性雄性不育基因ghpsm5的活性和\或表达规律。
7.根据权利要求6所述创制方法,其特征在于,所述调控和/或改变的方法,包括基因编辑、rnai、反义rna和dna甲基化中的一种或多种。
8.根据权利要求6所述创制方法,其特征在于,所述调控和/或改变的方法,包括通过基因工程方法,利用权利要求3所述启动子作为启动子构建表达载体,表达能够对雄性器官发育发生作用的蛋白质、rna和/或dna序列。
9.根据权利要求8所述创制方法,其特征在于,所述雄性器官包括花药。
10.依托权利要求6~9任一项所述创制方法得到的光敏核隐性雄性不育材料在植物育种中的应用。
技术总结
本发明公开了光敏核隐性雄性不育基因Ghpsm5及其在棉花中的应用,涉及基因工程和农业作物遗传育种技术领域。本发明提供了一种棉花光敏核隐性雄性不育基因Ghpsm5,可通过改变正常棉花植株中Ghpsm5基因的序列,使其不能正常表达,可以导致棉花在日照时数大于等于12.5小时的长日照条件下雄性不育;日照时数小于等于12.0小时的短日照条件下恢复雄性育性;日照时数在大于12.0小时但小于12.5小时时不能自交结铃,仍可用于杂交制种。改变该基因对雌性育性无影响。通过生物技术阻止该基因的表达,可以创制棉花光敏核隐性雄性不育系,用于棉花育种。
技术研发人员:范术丽,张朝军,马启峰,乔凯凯
受保护的技术使用者:中国农业科学院棉花研究所
技术研发日:
技术公布日:2024/1/11
- 还没有人留言评论。精彩留言会获得点赞!