提高利用nic1位点选育低尼古丁烟草的分子标记及其应用的制作方法
1.本发明属于生物技术领域,具体涉及一种利用烟草尼古丁合成主效调控突变位点nic1后提高烟草品系遗传背景回复率的共显性和显性分子标记的应用。
背景技术:
2.栽培烟草(nicotianatabacuml.)是茄科一年生草本植物,也是重要的经济作物之一。尼古丁(nicotine)是栽培烟草中一类重要的特征性生物碱。根据烟草遗传背景的不同,烟叶中尼古丁含量占烟叶总生物碱含量的90~95%。烟叶中尼古丁含量的高低也直接与烟草品质和商业用途密切相关。与传统卷烟相比,即将推出的vln烟草制品(如vln
®
king、vln
®
menthol king)中尼古丁含量下降约95%,同时具有低危害、低焦油,不造成代偿性吸食的特点。国内外烟草公司和相关研究机构正致力于利用尼古丁合成的主效调控基因位点nic1来调控烟叶尼古丁含量,实现减少烟草制品中的尼古丁含量,从而达到降低烟草对人体的危害性的目的。此外,国内外大量研究机构正在利用改变烟草栽培措施、与不同茄科作物嫁接、基因编辑为主的生物技术手段开展降低烟草尼古丁的生物合成或尼古丁运输过程来达到调控烟叶尼古丁含量的目的。由于美国烟草制品为混合型烟草,22世纪烟草公司也正与北卡罗莱纳州立大学合作开发白肋烟、香料烟和晾晒烟的低尼古丁烟草品种。
3.烟草尼古丁不同含量的突变体最早是在雪茄烟中发现的,后来由烟草育种家们通过杂交回交选育将这些突变位点分别转育到烤烟和白肋烟的不同品系当中。在白肋烟中,烟草育种家们通过花药培养和染色体加倍技术选育出了四份白肋烟尼古丁合成的近等基因系(near isogenic lines,nils)材料,分别命名为ha burley 21、hiburley 21、liburley 21和laburley21。对这四份材料的经典遗传学研究发现,烟草尼古丁含量受到两个独立遗传的办显性位点调控(命名为nicotine 1和nicotine 2,简称nic1和nic2)(legg,p.d.,collins,g.b.,1971.inheritance of per cent total alkaloids in nicotiana tabacum l.:ii.genetic effects of two loci in burley 21x la burley 21populations.can.j.genet.cytol.13,287-291.legg,p.d.,chaplin,j.f.,collins,g.b.,1969.inheritance of percent total alkaloids in nicotiana tabacum l.:populations derived from crosses of low alkaloid lines with burley and flue-cured varieties.j.hered.60,213-217)。其中,nic1位点对尼古丁合成的调控能力是nic2位点的2.4倍。根据相关报道,烟草中nic2位点至少由7个乙烯响应因子erf(ethylene response factor)转录因子基因所组成,包括nterf189,nterf115、nterf179等。这些转录因子基因能够结合尼古丁合成途径上的代谢酶基因(如ntpmt2和ntqpt2)的启动子区域来转录激活下游基因表达(shoji,t.,kajikawa,m.,hashimoto,t.,2010.clustered transcription factor genes regulate nicotine biosynthesis in tobacco.plant cell 22,3390-3409;shoji,t.,hashimoto,t.,2012.dna-binding and transcriptional activation properties of tobacco nic2-locus erf189 and related transcription factors.plant biotechnology 29,35-42.)。其中,la burley 21品系中的突变型nic2位
点是由于其所在的染色体片段丢失所造成的(kajikawa m.,sierro n.,kawaguchi h.,et al.,2017.genomic insights into the evolutionof the nicotine biosynthesis pathway in tobacco,plant physiology 174,999-1011.)。另一方面,近期研究还发现其他类的转录因子(transcription factor)家族基因也参与到烟草尼古丁生物合成的转录调控中。例如,basic helix-loop-helix(bhlh)家族转录因子ntmyc2,能够与nic2位点转录因子(如nterf189)协同调控,二者能够分别结合到尼古丁合成途径中关键代谢限速酶基因(如ntpmt、ntqpt)启动子中的g-box和gcc-box转录元件上来来最大限度激活这些基因的表达(shoji t.and hashimoto t.2011.tobacco myc2 regulates jasmonate-inducible nicotine biosynthesis genes directly and by way of the nic2-locus erf genes,plant cell physiol.52(6):1117
–
1130.)。另一方面,烟草生长素响应因子ntarf6转录因子过表达所引起的烟草生物碱含量降低是由于茉莉酸途径和其他植物激素信号转导途径(如乙烯、水杨酸、脱落酸)之间的拮抗作用所引起的。茉莉酸生物合成途径受到抑制的同时也伴随着乙烯、水杨酸、脱落酸以及病菌侵染防御反应的激活,从而拮抗茉莉酸所诱导的次生代谢调整并降低烟草体内尼古丁合成基因的表达(hu et al.,2021.transcriptomic analysis provides insights into the auxin response factor 6-mediated repression of nicotine biosynthesis in tobacco(nicotiana tabacum l.),plant mol biol.107(1-2):21-36.)。此外,近期发现myb转录因子家族基因ntmyb305a可通过结合烟草ntpmt1a启动子上的gag区域中的at-rich元件。同时ntmyb305a也以类似的调控方式来调控其他尼古丁合成基因表达,从而正向调控烟草中尼古丁的生物合成(bian et al.,2022.ntmyb305a binds to the jasmonate-responsive gag region of ntpmt1a promoter to regulate nicotine biosynthesis.plant physiol.188(1):151-166.)。国内外学者分别在烤烟(云烟87)和白肋烟(haburely 21)基因组通过生物信息学手段中发现了一个新的erf(nic2-like)基因簇,其组成与nic2基因簇高度类似(kajikawa m.,sierro n.,kawaguchi h.,et al.,2017.genomic insights into the evolutionof the nicotine biosynthesis pathway in tobacco,plant physiology 174,999-1011;sui x.et al.,2019.ethylene response factor nterf91 positively regulates alkaloidaccumulations in tobacco(nicotiana tabacum l.),biochembiophys res commun.517(1):164-171;sui x.et al.,2020.unravel the mystery of nic1-locus on nicotine biosynthesis regulation in tobacco,biorxiv)。最近,通过遗传群体定位和基因功能验证的手段已经证明上述nic2-like基因簇就是nic1位点,并对其簇内多个基因的功能进行了验证(sui x.et al.,2020.unravel the mystery of nic1-locus on nicotine biosynthesis regulation in tobacco,biorxiv)。同时开发出多个用于检测白肋烟nic1基因的snp(single nucleotide polymorphism)分子标记,且将检测的标记及方法申请了专利(pct/us2018/038679)。另一方面,对nic1突变体中尼古丁合成基因表达量下降的原因也已基本明确。突变型nic1基因簇所在染色体区域的侧翼发生了两个染色体片段(nic1-s和nic1-b)缺失,最终导致nic1基因簇所处染色体发生结构变异从而影响了基因簇内基因的正常表达(sui x.et al.,2020.unravel the mystery of nic1-locus on nicotine biosynthesis regulation in tobacco,biorxiv)。
4.然而,利用突变型nic1基因簇开展低尼古丁含量烟草品种作为常用的育种手段仍
面临一些问题。根据相关文献报道,突变型nic1烟草品系(la burley 21)与正常烟草相比存在许多农艺性状上的缺点,如落黄慢、难烘烤、产量低、烤后烟叶质量差(chaplin,j.f.,&burk,l.g.(1983).agronomic,chemical,and smoke characteristics of flue-cured tobacco lines with different levels of total alkaloids.crop science,75,133
–
136.chaplin,j.f.,&weeks,w.w.(1976).association between percent total alkaloids and other traits in flue-cured tobacco.crop science,16,416
–
418.;legg,p.d.,collins,g.b.,&littion,c.c.(1970).registration of la burley 21tobacco germplasm.crop science,10,212.)。因此,利用lafc53作为种质材料通过回交选育低尼古丁烟草品种的过程中,需要在最大程度保留nic1突变型位点的同时,最好能够将系统选育或回交世代中的轮回亲本的染色体片段得以保留。因此,利用全基因组重测序技术对比轮回亲本基因组(如nc95 ha)和非轮回亲本(lafc53)在nic1所在染色体区域周围区域寻找差异,开发能够区分两亲本基因组差异的特异(共显性或显性)分子标记,对采用分子标记辅助选育低尼古丁含量的烟草品种具有重要意义。
5.截止目前,针对烟草尼古丁合成nic1位点的分子标记开发研究虽有报道,但现有报道的研究成果均是基于nic1突变型位点遗传前景选择开展的,即仅仅选择造成nic1基因簇内基因表达水平下调的缺失片段(nic1-s和nic1-b)进行标记选择(sui x.et al.,2020.unravel the mystery of nic1-locus on nicotine biosynthesis regulation in tobacco,biorxiv)。例如,与nic1基因簇连锁的标记报道是基于snp变异位点开发的dcaps标记和ssr标记。在烟草育种实践及烟叶生产中存在着一些不足:1)存在严重的使用范围局限性,即已报道并申请专利的检测鉴定nic1基因的dcaps标记的使用范围较小,其仅适用于白肋烟类型;而栽培烟草中除了白肋烟类型外,还有烤烟、香料烟、雪茄烟、晾烟、晒烟等多种不同类型及黄花烟草种,上述众多烟草类型或种是dcaps标记所不能检测鉴定的。2)目前所开发的分子标记仅仅基于对nic1突变型基因簇本身的标记辅助选择(即前景选择),而对无法对轮回亲本或非轮回亲本(如lafc53)遗传背景进行选择。在利用lafc53作为非轮回亲本通过系统选育或回交转育低尼古丁烟草品种的过程中,无法在最大程度保留基因nic1突变型位点的同时,同时将回交世代中的轮回亲本的染色体片段得以最大程度地保留,从而有效地改良由突变型nic1烟草非轮回亲本(lafc53)上存在的农艺性状上的缺点(落黄慢、难烘烤、产量低、烟叶质量差)。
6.本发明利用尼古丁含量不同的两份近等基因系材料nc95 ha(nic1nic1nic2nic2)和低尼古丁含量的烟草材料lafc53(nic1nic1nic2nic2)为实验材料,通过烟草基因组重测序技术对比两个近等基因系在nic1位点所在染色体区域存在的结构变异,开发出可区分两亲本基因组差异的特异(共显性和显性)分子标记。通过构建自交二代(f2)分离群体,对筛选出选择群体中基因型为(nic1nic1nic2nic2)的个体后,单株留种。对留种的单株株系在f3代利用特异标记进行遗传背景选择和株系农艺性状比较试验,筛选出f3代中比非轮回亲本(lafc53)遗传背景回复高和农艺性状有显著改良的低尼古丁烟草品系,加速分子标记辅助选择(marker assistant selection,mas)在低尼古丁含量烟草品种选育中的利用,从而实现方便、快捷、稳定、可靠的培育出具有低尼古丁含量且高品质的烟草品种。
技术实现要素:
7.本发明的第一目的在于提供一种利用烟草尼古丁合成主效调控nic1突变型位点后,有效提高系统选育或回交选育后代遗传背景选择效率的特异性(显性和共显性)分子标记;第二目的在于提供所述的与烟草尼古丁合成主效调控nic1突变体位点后提高回交后代遗传背景选择效率来而改良非轮回亲本(携带突变型nic1位点的亲本)所具有的不良农艺性状,加速突变型nic1位点在选育低尼古丁烟草品种中的应用。
8.利用烟草尼古丁合成主效调控突变位点nic1后能提高烟草品系遗传背景回复率的共显性和显性分子标记的应用,检测烟草分离群体中烟草单株基因组dna在保留突变型nic1位点的同时,通过系统选育或回交育种手段提高nic1位点所在染色体区域回复为轮回亲遗传背景的选择效率,最终有效改良由突变型nic1烟草非轮回亲本所引入的农艺性状上的缺点。
9.一种利用烟草尼古丁合成主效调控突变位点nic1的共显性和显性分子标记,本发明所述分子标记可检测到在烟草尼古丁合成主效调控突变型位点nic1所在染色体区域周围区域所产生的染色体结构变异,特异共显性分子标记的编号为target1~target3和显性分子标记的编号为target4~target6;其pcr扩增产物核苷酸序列分别为seq id no.1和seq id no.2、seq id no.3和seq id no.4、seq id no.5和seq id no.6、seq id no.7、seq id no.8和seq id no.9所示;所述的特异分子标记所对应的6个区域的引物序列分别为:
10.target1序列为:
11.target1f:5
’‑
gacttatggcaattcaaagataaga
ꢀ‑3’
;
12.target1r:5
’‑
cagtttctggaaatgtttgttaagt-3’;
13.target2序列为:
14.target2f:5
’‑
agttcaactattgttttctcgacat-3’;
15.target2r:5
’‑
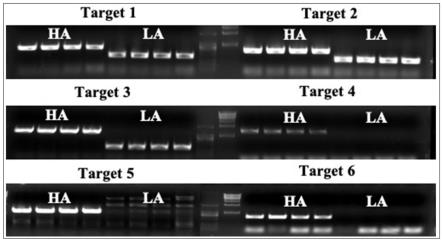
atttaggcactgttattacttgtgg-3’;
16.target3序列为:
17.target3f:5
’‑
gcaccatccaaacacaaggttaaac-3’;
18.target3r:5
’‑
cctaatcctcttcgaatcttaaatc-3’;
19.target4序列为:
20.target4f:5
’‑
tactactgtgcagcagatgatttag-3’;
21.target4r:5
’‑
taccttgcatatgttcctatatggt-3’;
22.target5序列为:
23.target5f:5
’‑
tttaagttcttgtttttctccttga
ꢀ‑3’
;
24.target5r:5
’‑
aatcagttccttcctcacactaac-3’;
25.target6序列为:
26.target6f:5
’‑
aaattcagagagatttttggaaagt-3’;
27.target6r:5
’‑
ataaagaagcagaaatagggaaaat-3’。
28.本发明所述的共显性和显性分子标记的应用,所述应用为:提高烟草品系遗传背景回复率。
29.本发明上述应用具体为:检测烟草分离群体中烟草单株基因组dna在保留nic1位点的同时,通过系统选育或回交育种手段提高nic1位点所在染色体区域回复为轮回亲遗传
背景的选择效率,从而改良非轮回亲本(携带突变型nic1位点亲本)所具有的不良农艺性状(如难落黄、产量低、及植株矮小等),提高后代烟株遗传背景选择效率,加速突变型nic1位点在选育低尼古丁烟草品种中的应用。
30.本发明上述应用为:是分别以target1序列的引物、target2序列的引物、target3序列的引物或target4序列的引物、target5序列的引物和target6序列的引物,分别扩增待检测烟草基因组dna,检测pcr扩增产物,依照扩增产物结果进行分析。
31.本发明如果pcr扩增产物结果中同时含有如seq id no.1、seq id no.3和seq id no.5所示的三条核苷酸序列,即为具有纯合轮回亲本的烟草遗传背景。
32.本发明如果pcr扩增产物结果中同时含有如seq id no.2、seq id no.4和seq id no.6所示的三条核苷酸序列,即为具有纯合突变型nic1的非轮回亲本遗传背景。
33.本发明如果pcr扩增产物结果中同时含有如seq id no.1、seq id no.2、seq id no.3、seq id no.4、seq id no.5和seq id no.6所示的6条核苷酸序列,即其烟草的遗传背景为杂合型,即具有轮回亲本和非轮回亲本遗传背景的烟草植株。
34.本发明如果pcr扩增产物结果中同时含有如seq id no.7、seq id no.8和seq id no.9所示的三条核苷酸序列,即具有轮回亲本或兼有轮回亲本和非轮回亲本的杂合型烟草遗传背景。
35.本发明如果pcr扩增产物结果为无pcr扩增产物,即为具有具有纯合突变型nic1的非轮回亲本遗传背景的烟草植株。
36.为了简便、高效选择具有低尼古丁含量且农艺性状良好的烟草品种,有针对性、特异性的选择含纯合基因型nic1nic1的后代材料,同时最大程度替换来源于低尼古丁的亲本(携带nic1突变型位点,如lafc53)中nic1位点所在染色体区域所携带的染色体片段。本发明提供一种利用烟草尼古丁合成主效调控突变位点nic1后提高烟草品系遗传背景回复率的共显性和显性分子标记target1、target2、target3、target4、target5和target6,该分子标记采用烟草基因组重测序分析(tobacco wholegenome resequencing)的方法,通过比较两份尼古丁含量不同的近等基因系材料nc95 ha(nic1nic1nic2nic2)和lafc53(nic1nic1nic2nic2)在突变型nic1位点所在染色体区域存在的结构变异,开发出可区分两亲本基因组差异的特异性分子标记。此分子标记可以用于低尼古丁含量烟草品种选育的辅助手段,以利用分子标记辅助选择提高回交选育后代遗传背景选择效率并改良农艺性状,从而加速低尼古丁含量烟草品种的选育进程。
37.本发明利用两份具有不同尼古丁含量的近等基因系作为材料nc95 ha(高尼古丁含量:nic1nic1 nic2nic2)和lafc53(低尼古丁含量:nic1nic1 nic2nic2)为亲本,采用烟草基因组重测序分析法,利用在突变体nic1所在染色体侧翼区域存在的染色体结构变异(structural variation,sv)开发设计能够有效区分来源于轮回亲本和非轮回亲本(如nc95 ha和lafc53)染色体片段的特异性分子标记。同时通过杂交、自交,筛选出自交二代(f2)群体中基因型nic1nic1nic2nic2的个体单株24株,套袋留种。提取f2代各单株基因组dna后利用所开发的特异性标记对各株系遗传背景进行筛选。另一方面,将24个单株自交后代(即f3)进行田间种植调查各株系的农艺性状和统计分析,获得烟草株系的各农艺性状(一个或多个)与非轮回亲本相比有明显改善的株系。结果表明利用这些标记可提高分子标记辅助选择(marker assistant selection,mas)在低尼古丁含量烟草品种选育中的选择
效率。
38.本发明所述的利用烟草尼古丁合成主效调控突变位点nic1后能提高烟草品系遗传背景回复率的共显性和显性分子标记具有稳定、可靠、简便、快捷和低成本的特点,因此该分子标记可以作为低尼古丁含量烟草品种培育中利用突变型nic1位点的同时提高遗传背景选择效率的分子标记的应用。
附图说明
39.图1是与烟草尼古丁合成主效调控位点突变体nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记在2份近等基因系材料中的pcr扩增产物凝胶电泳图;其中,a,共显性标记target1;b,共显性标记target2;c,共显性标记target3;d,显性标记target4;e,显性标记target5;f,显性标记target6;la,低尼古丁含量亲本lafc53;ha,高尼古丁含量亲本nc95 ha;m,1000bp dna ladder,长度片段分别为:100bp,200bp,300bp,400bp,500bp;
40.图2是nc95 ha
ꢀ×
lafc53的f2群体中四种基因型a_b_、a_bb、aab_、和aabb个单株的生物碱含量图(其中,a:nic1;b:nic2;a:nic1;b:nic2)。
41.图3是共显性标记target1、target2和target3和显性标记target4、target5和target6在8份f2代(基因型为nic1nic1 nic2nic2)各单株中的pcr扩增产物条带统计图表,其中,a表示来源于轮回亲本nc95 ha的染色体片段,b表示来源于具有nic1突变型位点的非轮回亲本lafc53的染色体片段;
42.图4是所筛选到f2代单株所得到的f3代烟草株系与对照品种lafc53的农艺性状的统计分析图,包括自然株高、自然叶片、打顶株高、茎围、腰叶长、腰叶宽、有效叶数、节距。
具体实施方式
43.本领域技术人员将会理解,下列实施例仅用于说明本发明,而不应视为限定本发明的范围。实施例中未注明具体技术或条件者,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。所用试剂或仪器未注明生产厂商者,均为可以通过购买获得的常规产品。
44.本发明所述的利用烟草尼古丁合成主效调控突变位点nic1后能提高烟草品系遗传背景回复率的共显性和显性分子标记的编号为target1、target2、target3 target4、target5和target6,其pcr扩增产物核苷酸序列分别为seq id no.1和seq id no.2、seq id no.3和seq id no.4、seq id no.5和seq id no.6、seq id no.7、seq id no.8和seq id no.9所示。
45.所述的分子标记所对应的6个位点的引物序列分别为:
46.target1序列为:
47.target1f:5
’‑
gacttatggcaattcaaagataaga
ꢀ‑3’
(seq id no.1),
48.target1r:5
’‑
cagtttctggaaatgtttgttaagt-3’(seq id no.2);
49.target2序列为:
50.target2f:5
’‑
agttcaactattgttttctcgacat-3’(seq id no.3),
51.target2r:5
’‑
atttaggcactgttattacttgtgg-3’(seq id no.4);
52.target3序列为:
53.target3f:5
’‑
gcaccatccaaacacaaggttaaac-3’(seq id no.5),
54.target3r:5
’‑
cctaatcctcttcgaatcttaaatc-3’(seq id no.6);
55.target4序列为:
56.target4f:5
’‑
tactactgtgcagcagatgatttag-3’(seq id no.7),
57.target4r:5
’‑
taccttgcatatgttcctatatggt-3’;
58.target5序列为:
59.target5f:5
’‑
tttaagttcttgtttttctccttga
ꢀ‑3’
(seq id no.8),
60.target5r:5
’‑
aatcagttccttcctcacactaac-3’;
61.target6序列为:
62.target6f:5
’‑
aaattcagagagatttttggaaagt-3’(seq id no.9),target6r:5
’‑
ataaagaagcagaaatagggaaaat-3’;
63.本发明所述的与烟草尼古丁合成主效调控位点突变体nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记的应用,其特征在于检测烟草基因组dna中是否存在突变型基因型nic1位点的同时而通过利用系统选育和回交选育手段来提高育种后代中提高烟草遗传背景选择效率,改善所选育烟草植株田间农艺性状,从而加速选育低尼古丁烟草品种中的应用。
64.所述的与烟草尼古丁合成主效调控位点突变体nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记的应用是分别以target1序列的引物、target2序列的引物和target3序列的引物分别扩增待检测烟草基因组dna,检测pcr扩增产物,如果pcr扩增产物中同时含有如seq id no.1、seq id no.3和seq id no.5所示的三条核苷酸序列,即为具有轮回亲本nc95 ha的烟草遗传背景;如果pcr扩增产物中同时含有如seq id no.2、seq id no.4和seq id no.6所示的三条核苷酸序列,即为具有非轮回亲本lafc53烟草的遗传背景;如果pcr扩增产物中同时含有如seq id no.1、seq id no.2、seq id no.3、seq id no.4、seq id no.5和seq id no.6所示的6条核苷酸序列,即其烟草的遗传背景为具有轮回亲本和非轮回亲本杂合型遗传背景的烟草植株。如果以target4序列的引物、target5序列的引物和target6序列的引物分别扩增待检测烟草基因组dna,检测pcr扩增产物,如果pcr扩增产物中同时含有如seq id no.7、seq id no.8和seq id no.9所示的三条核苷酸序列,即为具有轮回亲本nc95 ha的纯合型或具有轮回亲本和非轮回亲本的杂合型的烟草遗传背景;如果pcr无扩增产物,即为具有纯合突变型nic1的非轮回亲本lafc53遗传背景的烟草植株。
65.下面以具体实施案例对本发明做进一步说明:
66.实施例1
67.利用烟草全基因组重测序分析(tobacco wholegenome resequencing)法,对两份近等基因系材料高尼古丁含量烟草品系nc95 ha(基因型nic1nic1nic2nic2)和低尼古丁含量烟草lafc53品系(基因型nic1nic1nic2nic2)进行深度重测序。通过序列读段比对来检测和对比nic1所在染色体侧翼区域是否在两亲本之间存在的染色体结构变异,并根据变异类型获取相关染色体片段序列信息。烟草全基因组重测序分析结果可视化在igv(integrative genomics viewer)软件上进行,相关特异性标记(引物)的设计利用primer3
在线工具完成(https://bioinfo.ut.ee/primer3-0.4.0/)。
68.一、实验材料
69.以两份近等基因系材料高尼古丁含量烟草品系nc95 ha(nic1nic1nic2nic2)为母本,以低尼古丁含量烟草lafc53品系(nic1nic1nic2nic2)为父本。2017年种植高、低尼古丁含量亲本材料,经杂交,获得f1代。2018年冬季种植杂交一代(f1),于2019年初获得nic1、nic2基因分离群体(f2代)。2019年种植两亲本、f1和f2世代材料,并在f2世代材料中获得基因型为(nic1nic1nic2nic2)的单株,套袋收种获得f3代株系。2021年种植高、低尼古丁含量亲本材料,以及f3代株系,获得亲本及f3代田间农艺性状调查数据。
70.二、亲本及f2分离群体尼古丁含量测定
71.试验材料成苗后移栽至大田,行株距为100cm
×
50cm;采用常规栽培和田间管理,f2群体整体现蕾后打顶,打顶2周后,对各单株鲜烟叶进行采烤。采烤后分别将每株叶片研磨并测定尼古丁含量。对得到的亲本和f2群体各单株叶片的尼古丁含量数据进行分析,基因型数据分析的依据为:根据各个单株的带型进行数据统计,即,两个位点与nc95 ha亲本带型一致的单株记作“a_”或“b_”;两个位点与lafc53亲本带型一致的单株条带标记作“aa”或“bb”,条带不清晰或者无扩增条带的记做“u”。
72.三、特异性标记设计
73.打顶前,在田间采集双亲、f2群体的各单株新鲜叶片,参试材料全基因组dna的提取,采用常规ctab法或植物组织dna提取试剂盒均可,方法可参考已有的文献或试剂盒中的说明书。pcr反应体系配制、产物扩增及扩增产物的2%琼脂糖凝胶电泳检测,参照sui et al.(sui x.,huang y.,et al.2011,molecular authentication of an ethnomedicinal plant sabia parviflora and its adulterants by dna barcoding technique.planta medica 2011,77(5):492-496.)提供的方法进行。
74.所述pcr反应体系为:30-50ng/μl dna,正向和反向引物各1.0μmol/l,1.5mmol/l dntps,2μl 10
×
pcr buffer(mg
2+
plus),0.75~1.0u q5高保真dna聚合酶(high-fidelity dna polymerase,neb),加双蒸水至20μl。
75.所述pcr扩增程序为:95℃预变性5min,95℃变性30sec,55℃复性30sec,72℃延伸30sec,30个循环后,72℃再延伸5min。
76.所述凝胶电泳检测指采用2%的琼脂糖凝胶,1
×
tbe电泳缓冲液,于100v恒压电泳45分钟分离,最后用溴化乙锭法显影。
77.四、基于nic1位点染色体结构变异开发特异性标记的亲本验证
78.在苗期按实施例1所述方法,利用重测序数据分析结果所设计的引物对中高尼古丁含量亲本nc95 ha(nic1nic1nic2nic2)和低尼古丁亲本lafc53(nic1nic1nic2nic2)2份材料进行pcr扩增,筛选验证能够区分两亲本在突变型nic1位点所在染色体区域染色体片段的特异性标记(显性或共显性)。筛选的结果如图1所示:共显性特异性标记target1、target2和target3能够区分两亲本在nic1位点所在染色体区域周围的染色体片段,即,标记target1、target2和target3在高尼古丁含量亲本nc95 ha中的带型完全一致,仅出现一条776bp(序列如seq id no.1)、716bp(序列如seq id no.3)和1458bp(序列如seq id no.5)的特异性条带;在低尼古丁含量亲本lafc53中的带型完全一致,仅出现一条457bp(序列如seq id no.2)、396bp(序列如seq id no.4)和420bp(序列如seq id no.6)的特异性条
ha亲本带型一致的单株记作“a”;与lafc53亲本带型一致的单株条带标记作“b”。
86.f2群体单株尼古丁含量的结果为:377个单株中,基因型为a_b_的单株有224个,基因型为a_bb的单株有36个,基因型为aab_的单株有94个,基因型为aabb的单株有23个。对f2代377个单株进行基因型分析如图2所示。
87.实施例3
88.利用与烟草尼古丁合成主效调控位点突变体nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记提高亲本nc95 ha和亲本lafc53杂交后代中f3代低尼古丁含量株系的田间农艺性状。
89.一、实验材料
90.所用的标记为target1、target2、target3、target4、target5和target6六个nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记。植物材料为ha nc95(基因型nic1nic1nic2nic2)、lafc53(基因型nic1nic1nic2nic2)、f2分离群体(nc95 ha
×
lafc53)中基因型为nic1nic1nic2nic2的8个单株、及其所对应的f3代8个株系。
91.二、数据处理
92.首先,按实施例1所述方法对上述烟草材料进行烟草基因组dna提取、纯化。其次,利用与烟草尼古丁合成主效调控位点突变体nic1所在染色体侧翼区域存在的染色体结构变异检测的特异性分子标记对上述烟草单株进行基因型分析。最后,对各个单株的带型进行数据分析,即,pcr扩增产物中同时出现如seq id no.1、seq id no.3和seq id no.5所示的核苷酸序列即为具有高尼古丁含量烟草轮回亲本nc95 ha遗传背景的特异性pcr扩增核苷酸序列;若pcr扩增产物中同时出现如seq id no.1和seq id no.2、seq id no.3和seq id no.4及seq id no.5和seq id no.6所示的核苷酸序列即为具有轮回亲本和非轮回亲本(携带nic1位点突变型)的杂合型染色体片段;若pcr扩增产物中同时出现如seq id no.2、seq id no.2和seq id no.4所示的核苷酸序列即为具有含有nic1突变型位点的非轮回亲本lafc53遗传背景的特异性pcr扩增核苷酸序列。
93.由图3可知:在8份候选待测的f3代烟草株系中,株系nc95 ha
×
lafc53 f
3-72是在保证具有具有低尼古丁性状(nic1nic1nic2nic2)前景的同时,具有最多的nc95 ha亲本遗传背景的株系,其农艺性状与亲本nc95 ha无明显差异,主要农艺性状得到了较大改善;而株系nc95 ha
ꢀ×
lafc53 f
3-112是在保证具有具有低尼古丁性状(nic1nic1nic2nic2)前景下,同时与亲本lafc53的遗传背景最接近的株系,其有除茎围略高于亲本lafc53外和有效叶数低于亲本lafc53,其他农艺性状(如自然株高、自然叶数、打顶株高)与亲本lafc53无明显差异,总体农艺性状较差;而其他f3株系(如nc95 ha
ꢀ×
lafc53 f
3-112、nc95 ha
ꢀ×
lafc53f
3-12、nc95 ha
×
lafc53 f
3-151和nc95 ha
×
lafc53 f
3-308)其nic1位点周围染色体区域不同程度地被亲本(nc95 ha)的染色体片段替换(纯合或杂合),其总体农艺性状与亲本lafc53相比也不同程度地改善,如图3所示。这些候选株系可以进一步通过系统选育加速品系纯合或通过与亲本nc95 ha进行回交来加快f3候选株系中不同单株的遗传背景选择效率,最终使其农艺性状得到不断改善,最终分子辅助选择选育低尼古丁含量的烟草品种。
94.结论:利用上述六个特异性(显性或共显性)标记可简便、快捷、稳定地检测烟草nic1位点所在染色体周围染色体片段替换的情况,又可清晰地鉴定出来源于不同亲本染色体片段的纯合或杂合情况,同时有针对性、特异性的选择遗传背景回复率高且各农艺性状
有较大改善的低尼古丁含量的烟草中间材料,可极大提高具有低尼古丁含量烟草品种的选育效率。
95.以上所述的仅是本发明的部分具体实施例,方案中公知的具体内容或常识在此未作过多描述(包括但不仅限于简写、缩写、本领域惯用的单位)。应当指出,上述实施例不以任何方式限制本发明,对于本领域的技术人员来说,凡是采用等同替换或等效变换的方式获得的技术方案均落在本发明的保护范围内。本技术要求的保护范围应当以其权利要求的内容为准,说明书中的具体实施方式等记载可以用于解释权利要求的内容。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1