脑机接口行为学模型的构建方法与流程
本发明涉及脑机接口技术领域,特别是涉及脑机接口行为学模型的构建方法。
背景技术:
脑机接口(brain-machineinterfaces,bmi)在大脑与外部设备之间建立一种新型的信息交流与控制通道,实现大脑与外界的直接交互,该技术不依赖于常规的脊髓/外周神经肌肉系统。1969年fetz,e.e检测皮层神经元的活性,通过操作性条件强化导致皮层神经元放电增加构建volitionalcontrol。1999年通过检测并解码大鼠皮层m1神经元利用m1皮层神经元的电信号控制外部机械臂获得奖赏,建立真正的大脑与外界的直接交互。过去的十几年,利用大鼠、小鼠、非人灵长类动物和瘫痪的病人进行脑机接口的研究蓬勃发展,利用实时控制外部的设备,在小鼠、类人猿、瘫痪病人成功控制神经义肢,而且脑机接口的临床预实验的研究正在进行之中。虽然目前研究已经在信号检测、信号解码和bmi学习的反馈方面取得很大的进步,极大地提高bmi的学习,但是机器和脑的交互不仅仅是恢复断开的连接,而且涉及到神经义肢的学习和神经适应。控制和使用神经义肢控制并不能接近正常的运动。神经义肢学习是通过强迫性学习理论,以目标导向性的方式,通过意念控制神经义肢。因此,脑机接口的学习是一个漫长的过程,目前除了加强训练,通过工程技术的方法改进解码和记录神经信号外,没有更好的方法促进脑机接口的学习。
技术实现要素:
基于此,本发明提供一种新的脑机接口行为学模型的构建方法,所述构建方法成功率高,构建的模型稳定。
具体技术方案为:
一种脑机接口行为学模型的构建方法,包括以下步骤:
操作性行为阶段:训练小鼠在提示出现后的t1内通过操作性行为获得奖赏,成功获得奖赏的操作性行为的次数为n次,同时,分别记录所述n次操作性行为发生时,小鼠m1皮层神经元的钙信号f1值,将所述提示出现时小鼠m1皮层神经元的钙信号记为f0,计算所述n次操作性行为中,所述钙信号的变化率(f1-f0)/f0的平均值,将所述平均值设定为域值;
意念控制阶段:训练小鼠在所述提示出现后的t2内通过意念控制小鼠m1皮层神经元的钙信号,记录所述t2内的小鼠m1皮层神经元的钙信号f2值,待钙信号的变化率(f2-f0)/f0超过所述域值,小鼠获得奖赏,记为一次成功的试验,统计小鼠在每天试验中的成功率,并记录每天完成m次成功的试验所需总时间;
以每天试验中的所述成功率和所述完成m次成功的试验所需总时间作为评价指标,完成所述脑机接口行为学模型的构建。
在其中一个实施例中,所述t1内为所述提示出现后的0-30s或0-20s内。
在其中一个实施例中,所述t2内为所述提示出现后的0-30s或0-20s内。
在其中一个实施例中,所述意念控制阶段还包括:
训练小鼠在所述提示出现前的t3内通过意念控制小鼠m1皮层神经元的钙信号,记录所述t3内的小鼠m1皮层神经元的钙信号f3值,待所述f3<f1,且所述钙信号的变化率(f2-f0)/f0超过所述域值,小鼠获得奖赏,记为一次成功的试验。
在其中一个实施例中,所述t3内为所述提示出现前的15s内。
在其中一个实施例中,通过比较每天试验中的所述成功率的情况和所述完成m次成功的实验所需总时间的长短,判断所述脑机接口行为学模型是否成功构建。
在其中一个实施例中,所述操作性行为阶段的训练时间为5-8天。
在其中一个实施例中,所述意念控制阶段的训练时间为8-15天。
在其中一个实施例中,所述n次为40-60次。
在其中一个实施例中,所述m次为40-60次。
在其中一个实施例中,所述操作性行为为压杆行为。
在其中一个实施例中,所述提示为声音提示。
与现有技术相比,本发明具有以下有益效果:
本发明提供一种新的脑机接口行为学模型的构建方法,该模型的成功构建为脑机接口学习提供了新的研究载体。通过测试,本发明所述的构建方法成功率高,尤其是意念控制阶段保持较高的成功率,模型稳定。
附图说明
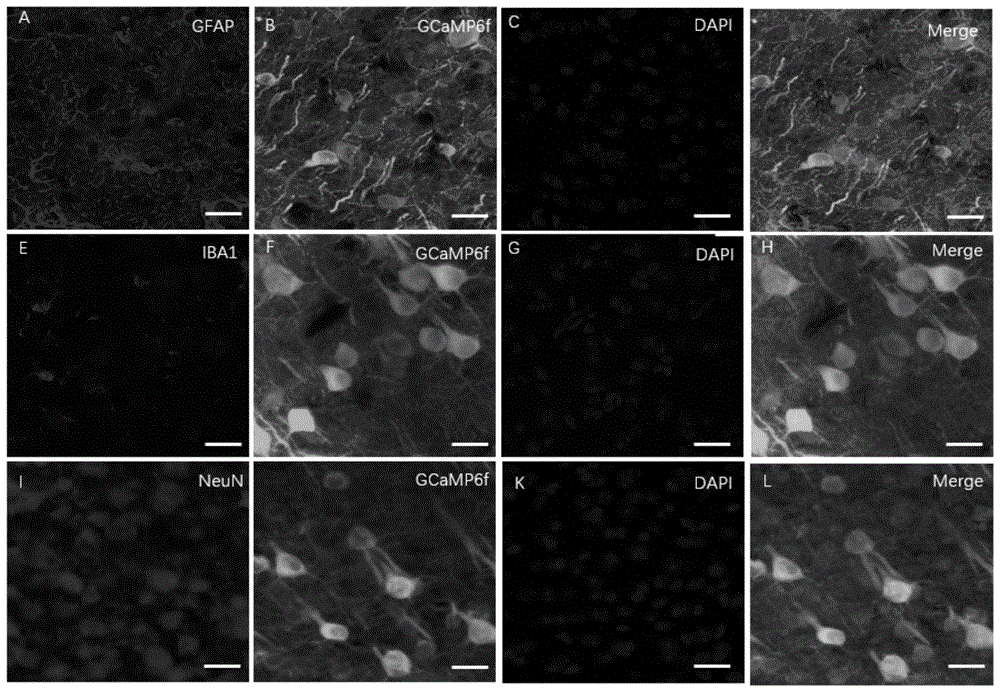
图1为gcamp6f表达细胞的鉴定图;
图2为脑机接口行为学模型构建方法示意图;
图3为实施例1的测试结果示意图;
图4为实施例2、实施例3和实施例4的测试结果示意图;
图5为实施例5的试验方法和测试结果示意图。
具体实施方式
以下结合具体实施例对本发明的腺苷a2a受体拮抗剂类药物的新应用作进一步详细的说明。本发明可以以许多不同的形式来实现,并不限于本文所描述的实施方式。相反地,提供这些实施方式的目的是使对本发明公开内容理解更加透彻全面。
除非另有定义,本文所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。本文中在本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不是旨在于限制本发明。本文所使用的术语“和/或”包括一个或多个相关的所列项目的任意的和所有的组合。
实施例1利用小鼠m1皮层神经元群体细胞的钙信号构建脑机接口行为学模型
脑机接口能够帮助瘫痪患者恢复运动功能,脑机接口(神经义肢)行为学模型的构建是适应脑机接口的核心。在本实施例中,利用小鼠m1皮层群体神经元的钙信号作为“控制器”,在自由运动小鼠中构建脑机接口行为学模型(图3a-d),具体方法为:
首先,利用aav9-syn-gcamp6f-wpre-sv40(aav病毒)在小鼠m1皮层神经元表达遗传编码的ca2+指示剂gcamp6f,并在病毒的注射的位点插入光纤。经证明:gcamp6f是选择性的表达在皮层的l4-6层(l4/l5/l6)神经元并和神经元的标记物neun共表达,而不是和小胶细胞标记(gfpa+)和小胶质细胞标记(iba1)共表达(图1a-l)。采用光纤记录系统实时记录m1皮层神经元l5层的钙信号的变化(图2a),利用该钙信号作为“控制器”构建脑机接口行为学模型(图2b)。
其次,由于小鼠学习本来不存在的工作结构是很困难的,所以应用两步法转换的学习方式(操作性行为阶段模型构建和意念控制阶段模型构建)帮助小鼠学会脑机接口学习(图2c-d)。
(1)操作性行为阶段:训练小鼠在特定的时间内通过操作性行为(压杆)获得奖励,并伴随着在m1皮层神经元特定的钙信号变化的出现。
具体方法为:
取m1皮层神经元表达gcamp6f的小鼠,在固定间隔(半小时)无条件给予糖水,接着,强迫学习三天,每天50次通过压杆获得糖水(若未完成50次,但是总的训练时间达到1小时,也视为完成每天试验)。接着,进入为期6天的操作性行为训练,增加声音提示,在声音提示出现后的30s内,小鼠压杆才能获得糖水(图2c),统计小鼠在这一阶段每天试验中的成功率和每天获得50次糖水所需的总时间(若未完成50次,但是总的训练时间达到1小时,也视为完成每天试验,总时间记为1小时),可知,小鼠压杆获得糖水的成功率不断增加(图3a,n=8:onewayanovap<0.0001),每天训练的时间(获得50次糖水的总时间)减少(图3b,n=8;onewayanovp<0.0001)。研究发现,训练第二天,成功率达到平台期,在随后的四天,成功率维持在95%以上。
将声音提示出现时的小鼠m1皮层神经元的钙信号记为钙信号的基线(f0);δf定义为钙信号f1在f0基础之上的变化值(f1-f0)。通过分析δf/f[(f1-f0)/f0]压杆前2s和后5s的变化,发现,钙信号在压杆之前开始升高,压杆之后开始降低(图2c),因此得出钙信号的变化和压杆行为密切相关的结论,说明神经元活性的变化是压杆行为触发的。为了进一步证明钙信号变化和压杆行为之间的相关性,还对小鼠前肢的肌电变化进行记录(图2c)。证实在皮层m1钙信号的上升或下降瞬时精确的和小鼠前肢肌电上升或下降一致(图2c;p<0.05),进一步证明,m1神经元钙信号的变化和小鼠的前肢的压杆行为是相关的。
分别记录每天的操作性行为阶段中,50次成功压杆时小鼠m1皮层神经元的钙信号f1值。计算所述钙信号的变化率(f1-f0)/f0的平均值,将6天操作性行为阶段中钙信号的变化率(f1-f0)/f0的平均值设定为域值;
(2)意念控制阶段:训练小鼠在特定的时间通过意念控制m1皮层神经元的出现和操作性行为相似的钙信号来获得奖赏。
具体方法为:操作性行为的6天训练结束后,第7天,撤回杆,小鼠转换到为期10天的脑机接口的意念控制阶段的训练。训练声音提示出现后的30s内,小鼠通过意念控制m1皮层神经元的钙信号,将声音提示出现后0-30s内小鼠m1皮层神经元的钙信号记为f2。小鼠需要在声音提示出现后的30s内将δf2[(f2-f0)/f]升高到设定的域值,才能获得糖水,将此记为一次成功的实验,统计小鼠在每天试验中的成功率,并记录每天完成50次成功试验所需的总时间(若未完成50次,但是总的训练时间达到1小时,也视为完成每天试验,总时间记为1小时)。
基线f0对小鼠每次成功通过增加钙信号超过设定的域值获得奖励、完成每次实验非常重要,因为每次声音提示出现,表明实验开始,而我们定义的基线(f0)为:声音提示出现的10ms时,小鼠m1皮层神经元的钙信号的值,所以f0在每次实验都是不确定。
小鼠只有在实验开始前(声音提示出现前)减少钙信号,在声音提示出现后增加钙信号,满足这两个条件,才会成功完成实验任务,我们将小鼠在声音提示出现前的15s内小鼠m1皮层神经元的钙信号记为f3,训练小鼠在声音提示出现前减少钙信号f3。
在意念训练的开始阶段,钙信号有很大的变异,随着训练的增加,钙信号的变化慢慢的趋同。表现为:钙信号在声音出现前10s开始下降,接着在声音提示出现后开始增加(图3c-d;n=7)。最终,结果显示,在声音提示出现后,小鼠不仅条件性的意念增加钙信号超过设定域值,而且在声音提示出现之前,意念性的降低钙信号。
经过训练,判断脑机接口行为学模型构建是否成功有两个指标:1、在经过10天训练后,成功率是否有明显的差异;2、在经过10天训练后,完成时间是否明显减少。
结果显示,成功率随着训练的天数的增加而增加(图3e;one-wayanova,p<0.05)。在10天的意念训练期间,每次的训练的时间也减少(图3f;one-wayanova,p<0.05)。在训练的第一天,小鼠在声音提示出现后,并没有明显升高钙信号(图3c),但是在第十天,已经出现明显的趋势(图3d),而且,第十天(图3g)的每个训练的完成时间明显低于第一天(图3h)。说明小鼠慢慢学会调节钙信号的变化完成脑机接口行为学模型的构建,并且,模型较稳定。
最后,为了证实上述脑机接口行为学模型是由小鼠意念控制的,进行了如下验证:
(1)在意念控制阶段,同时记录小鼠左前肢的肌电和小鼠m1钙信号变化。观察到肌电信号和钙信号产生分离(图2d)。
(2)成功获得糖水的小鼠的f0附近的钙信号在比失败的更低,说明小鼠意念控制成功率的增加是小鼠有意识的在f0降低钙信号。
(3)在声音提示出现后,成功获得糖水的小鼠δf/f增加,失败的没有增加δf/f。
(4)录像结果显示,小鼠自由运动和抬头并不会达到设定的域值。在录像中我们可以看到,在小鼠自由运动和抬头的过程中,钙信号并没有明显的变化,说明钙信号的变化和这些运动没有关系,只有在小鼠想喝水的时候,才会出现钙信号增加的情况,说明钙信号是由意念产生的。
通过以上试验验证,证明上述脑机接口行为学模型是由小鼠意念控制的。
实施例2
本实施例在构建脑机接口行为学模型的操作性行为阶段和意念控制阶段,向参加训练的小鼠腹腔注射腺苷a2a受体拮抗剂kw6002(5mg/kg),在训练开始的前半小时开始注射,每日一次。
结果显示,在操作性行为阶段,小鼠获得糖水的成功率除在第一天明显上升外,其与五天的训练成功率并无显著差别(图4a;two-wayanova,p<0.050,),每天训练的时间除在前两天明显下降外,其与四天训练时间并无显著差别(图4b)。在意念控制阶段,小鼠获得糖水的成功率均上升(图4c;two-wayanova,p<0.051,p=0.05)。但是,每次的完成训练的时间并没有显著的变化(图4d)。
实施例3
本实施例仅在构建脑机接口行为学模型的意念控制阶段,向参加训练的小鼠腹腔注射腺苷a2a受体拮抗剂kw6002(5mg/kg),在训练开始的前半小时开始注射,每日一次。
结果显示:在意念控制阶段,小鼠获得糖水的成功率上升,与实施例2的结果进行比较,发现,在操作性行为阶段是否注射kw6002,不影响意念控制阶段的训练成功率,意念控制阶段的训练成功率仅与该阶段是否在意念控制阶段注射kw6002有关(图4e,two-way-anova,p<0.05),同时,注射kw6002对于每次训练的完成时间没有影响(图4f)。
实施例4
本实施例分两组训练小鼠构建脑机接口行为学模型。
第一组的训练方法与实施例1基本相同,区别在于,在操作性行为阶段,将训练条件改为:增加声音提示后,在声音提示出现后的20s内,小鼠压杆才能获得糖水。在意念控制阶段,将训练条件改为:在声音提示出现后的20s内,小鼠需要在声音提示出现后的20s内将δf2升高到设定的域值,才能获得糖水。本组小鼠在操作性行为阶段和意念控制阶段均未注射腺苷a2a受体拮抗剂kw6002。
第二组的训练方法与第一组的训练方法基本相同,区别在于:本组小鼠在操作性行为阶段和意念控制阶段均注射腺苷a2a受体拮抗剂kw6002。
结果显示,第一组的小鼠不能成功的完成脑机接口行为学模型的构建,在意念控制阶段,第一组的小鼠训练成功率与第二组小鼠的训练成功率有明显的差异(图4g,two-way-anova,p<0.05),在相同的训练条件下,注射腺苷a2a受体拮抗剂kw6002的小鼠训练成功率较高。训练完成的时间也有明显的差异(图4h,two-way-anova,p<0.05),相同的训练条件下,注射腺苷a2a受体拮抗剂kw6002的小鼠训练完成的时间较少。第二组的小鼠成功的完成了脑机接口行为学模型的构建,而第一组并不能成功完成该模型的构建。
实施例5
本实施例利用cre酶介导的floxeda2a受体基因敲除的方法验证阻断a2a受体对脑机接口行为学模型构建的影响。具体方法为:
取a2a-flox的转基因小鼠,在背侧纹状体注射aav-cre敲低a2a受体的表达(图5a),接着,采用与实施例1相同的方法,在上述小鼠中构建脑机接口行为学模型。
结果表明,在背侧纹状体敲除a2a受体后,同样能够促进小鼠在操作性行为(图5b,two-way-anova,p<0.05)和意念控制阶段(图5d,two-way-anova,p<0.05)的成功率,同样对训练完成时间都没有明显的影响(图5c和图5e;two-way-anova,p>0.05)。
以上研究表明,腺苷a2a受体拮抗剂类药物能够作为促进脑机接口学习的药物,提高脑机接口行为学模型的构建的成功率,尤其是,腺苷a2a受体拮抗剂能够作为促进脑机接口学习中意念控制阶段模型的构建的药物,提高完成意念控制阶段任务的成功率。
进一步地,腺苷a2a受体拮抗剂类药物除了可选自腺苷a2a受体拮抗剂kw6002之外,还可选自腺苷a2a受体拮抗剂cpi-444等其他腺苷a2a受体拮抗剂中的一种或几种。
腺苷a2a受体拮抗剂kw6002还可以与药学上可接受的辅料结合,制备成各种剂型的药物,应用于脑机接口的学习中。
进一步地,通过研究发现,纹状体-苍白球通路(简介通路)参与到脑机接口学习中,通过调控高表达与间接通路的a2a受体,可影响脑机接口学习。
以上所述实施例的各技术特征可以进行任意的组合,为使描述简洁,未对上述实施例中的各个技术特征所有可能的组合都进行描述,然而,只要这些技术特征的组合不存在矛盾,都应当认为是本说明书记载的范围。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
- 还没有人留言评论。精彩留言会获得点赞!