先兆子痫的制作方法
导言
可溶的fms-样酪氨酸激酶-1(sFlt-1或sVEGF-1)是一种VEGF受体1(Flt-1)的剪接变体,作为血管内皮生长因子(VEGF)的受体(Kendall et al.,1996)。然而,sFlt-1缺少受体的跨膜和胞浆结构域,及因此通过结合游离的循环VEGF而使VEGF信号途径变钝(Wu et al.,2010)。异常的sFlt-1表达在许多病理学状况中鉴别,包括不同类型的癌症、肝硬化、糖尿病和周围动脉疾病中(Bando et al.,2005;Ebos et al.,2004;Lamszus et al.,2003;Jaroszewicz et al.,2008;Blann et al.,2002;Findley et al.,2008)。
在这些疾病中,sFlt-1的作用在先兆子痫的发病机制中最佳鉴定。先兆子痫是在妊娠女性中以高血压、蛋白尿和肾损伤定义的一种孕母身体状况。目前,先兆子痫是孕母及围产期死亡率和发病率的主要原因(Steegers et al.,2010),影响世界范围内3%-8%的所有孕妇和女性(Sibai et al.,2005)。先兆子痫的发病机制是复杂的,但是已提出胎盘中内皮功能失调及导致的异常血管发生(新血管从现有血管中形成的过程)是这种医学状况的临床表现的基础(Hladunewich et al.,2007;Silasi et al.,2010;Maynard et al.,2008)。经鉴别sFlt-1在先兆子痫女性的血浆和胎盘中过量表达,sFlt-1的血浆水平可用作先兆子痫的潜在临床标记(Stepan et al.,2007)。此外,升高的sFlt-1被认为是导致先兆子痫的主要原因(Maynard et al.,2003;Powe et al.,2011)。我们先前的数据表明一种保护性酶-血红素加氧酶1(Hmoxl)负调节sFlt-1释放,而VEGF生长因子促进sFlt-1表达(Cudmore et al.,2007;Ahmad et al.,2011)。基于这些发现,我们及其它人使用强力的Hmoxl诱导剂statin开始进行预实验,以在高危女性中预防先兆子痫(Costantine et al.,2013)。然而,Hmoxl或VEGF怎样调节sFlt-1的分子学机制尚未知。
MicroRNA是一种21-23-nt的小非编码RNA,其以部分互补方式结合其靶基因的mRNA的3'-UTR,及因此导致靶基因的翻译抑制(Bartel 2009;Lewis et al.,2003;Williams 2008)。近年来,microRNA示出在调节许多发育、生理和病理过程中的重要作用(Williams 2008;He and Hannon,2004)。微阵列分析和进一步的定量PCR分析已经表明microRNAs在先兆子痫和正常女性的胎盘中不同表达(Pineles et al.,2007;Zhu et al.,2009;Hu et al.,2009;Mayor-Lynn et al.,2011;Enquobahrie et al.,2011;Noack et al.,2011)。进一步报道了microRNA与Hmoxl在干细胞分化、肺癌和氧化损伤中的相互作用(Kozakowska et al.,2014;Skrzypek et al.,2013;Hou et al.,2012)。因此,我们假设Hmoxl可以通过microRNA调节sFlt-1释放及在胎盘中microRNA的调节失常是导致先兆子痫的发病机制。使用基于qPCR的微阵列,本研究鉴别了两种microRNA,即miR-122和miR-374b,其可以由强力的Hmoxl诱导剂血红素(hemin)调节。进一步地,经证实这些microRNA由statin、Hmoxl和VEGF调节,其也调节sFlt-1表达。此外,sFlt-1证实是这些microRNA的直接靶位,及Hmoxl对sFlt-1的调节由这些microRNA进行。最重要地,这些microRNA在先兆子痫患者和RuPP先兆子痫模型中降低。也揭示了在动物模型中这些microRNAs与血压之间的负相关性。我们的研究提供了关于sFlt-1调节的新分子学机制及这些microRNA可作为sFlt-1相关疾病的新的治疗靶位。
MicroRNA-152是广泛保守的miR-148/152家族的成员。已经示出其通过靶向DNMT1调节表观形成(epigenesis)(Xiang et al.,2013;Ji et al.,2013;Huang et al.,2010;Braconi et al.,2010),抑制癌细胞增殖和粘附(Zhou et al.,2012;Mancini et al.,2012)及增加细胞溶解(Zhu et al.,2010。
进一步的研究证实其抗肿瘤作用,及因此在各种癌症类型中通过超甲基化负调节miR-152表达(Hiroki et al.,2010;Tsuruta et al.,2011;Braconi et al.,2010;Chen et al.,2010;Huang et al.,2010;Wang et al.,2010;Stumpel et al.,2011;Kitano et al.,2011;Zhou et al.,2012)。最重要地,已经示出在先兆子痫患者的胎盘中是正调节的(Zhu et al.,2009)。进一步地,miR-152降低肿瘤细胞血管发生(Zheng et al.,2013;Xu et al.,2013)及负调节重要的妊娠相关基因(HLA-G)的表达(Manaster et al.,2012)。
由于通过微阵列分析表明miR-152抑制癌细胞增殖和血管发生及还参与先兆子痫,我们假设miR-152在先兆子痫中调节内皮功能导致先兆子痫发病机制。在目前的研究中,我们证实在不同妊娠期胎盘、更准确定义的先兆子痫患者以及在动物先兆子痫模型的胎盘中miR-152的正调节,表明在低氧和炎症条件下miR-152表达增加。进一步的研究表明P1GF表达降低,内皮细胞粘附和血管发生能力基于miR-152的过表达。此外,腹膜内注射表达miR-152的病毒导致胎盘中去血管化及受限的胎儿生长。进一步地,我们在内皮细胞和小鼠模型中均鉴别了miR-152的新靶位ITGA5,证实miR-152和ITGA5在先兆子痫患者胎盘组织中的负相关性。miR-152及其靶位ITGA5在先兆子痫的发病机制中的鉴别将有助于理解这种医学状况及可以提供新的治疗靶位。
MicroRNA-195是广泛保守的miR-15/107超家族的成员。已经示出其是肿瘤阻抑物及通过靶向不同的下游因子而抑制癌细胞增殖、迁移、侵润及血管发生(Amer et al.,2014;Zhao et al.,2014;Jain et al.,2014;Guo et al.,2014;Wang et al.,2014a and b;Yang et al.,2014;Luo et al.,2014;Fu et al.,2013;Wang et al.,2013)。此外,miR-195调节胰岛素信号化途径并与2型糖尿病以及糖尿病相关的肾损伤和视网膜病变相关联(Chen et al.,2012;Mortuza et al.,2014;Yang et al.,2014;Ortega et al.,2014;Herrera et al.,2010;Guo et al.,2013)。此外,miR-195通过抑制心肌细胞的细胞周期还与急性心肌梗塞和心肌肥大相关(Long et al.,2012;You et al.,2014;van Rooij et al.,2006;Busk and Cirera,2010;Porrello et al.,2011)。令人感兴趣地,目前已经证实miR-195调节大动脉细胞外基质及可以用作深静脉血栓的生物标记(Zampetaki et al.,2014;Qin et al.,2014)。进一步研究表明了其在妊娠和相关并发症中的作用。miR-195的表达在妊娠晚期人胎盘中与妊娠早期相比是显著正调节的(Gu et al.,2013)。更重要地,miR-195在先兆子痫胎盘中的表达与在正常妊娠胎盘中的表达不同,提示mir-195在先兆子痫发病机制中的可能的作用(Xu et al.,2014;Zhu et al.,2009;Hu et al.,2009)。
内皮一氧化氮合成酶(eNOS或NOS3)是在内皮细胞中将L-精氨酸分解产生一氧化氮(NO)气并且在血管内皮中起关键作用的酶(Huang,2003)。eNOS产生低浓度NO,其为内皮功能和完整性提供保护作用,eNOS的丧失导致多种疾病,包括先兆子痫(Albrecht et al.,2003;Forstermann and Miinzel,2006;Fatini et al.,2006)。此外,已经鉴别eNOS/NO是新生血管的关键介导物(Duda et al.,2004),VEGF在eNOS-/-小鼠中不能血管发生(Lin and Sessa,2006)。然而,eNOS表达的调节机制还未知。迄今为止,仅一种microRNA,即miR-155,被报道在内皮细胞中直接靶向eNOS(Sun et al.,2012)。
由于通过微阵列分析表明miR-195抑制癌细胞增殖和血管发生及也参与先兆子痫,我们假设miR-195负调节内皮功能导致先兆子痫发病机制。在目前的研究中,我们证实miR-195在不同的妊娠胎盘、肥胖妊娠妇女、更准确而言的先兆子痫患者以及在动物先兆子痫模型中的正调节,表明the increase of miR-195表达在炎症(或者组合缺氧)条件下增加。进一步研究表明在miR-195过表达时,P1GF表达、内皮细胞增殖、存活力、粘附和血管发生能力降低。此外,我们鉴别了eNOS是miR-195的新靶位,及eNOSS1177D的过表达拯救miR-195介导的内皮细胞中血管发生的抑制。鉴于eNOS在先兆子痫发病机制中的重要性,miR-195的鉴别将有助于目前对于这种医学状况的了解及可以提供新的治疗靶位。
本发明提供了通过调节microRNA(miRNA)活性治疗先兆子痫的方法。所述活性典型是miRNA在参与先兆子痫、胎儿生长受限(FGR或IUGR)、妊娠肥胖、产后心肌病(心力衰竭)、癌症和糖尿病视网膜病变、心肌病、心肌梗塞、湿性黄斑变性以及其中血管发生是减少或加剧异常的其它病症的一或多个基因的基因表达的RNA沉默或者转录后调节中的功能,以在对象如具有这些疾病症状的人对象中减少症状或者诊断这些疾病。
MiRNA典型选自一或多个的miR-122、miR-374b、miR-152和miR-195或者其功能片段或同系物。例如,一或多个的miR-122、miR-374b加上miR-152,或者miR-122、miR-374b加上miR-195。
本领域通常已知miR-122、miR-374b、miR-152和miR-195及其抑制剂,可商购自例如英国曼彻斯特Qiagen公司。
它们可以是:
典型地,抑制剂是与miRNA基本互补的核酸分子以使得可以形成双链体。例如,一或两个碱基可以不是互补的,但是仍可以形成双链体。
术语“功能片段”是指保留与天然miRNA或其抑制剂相同生物活性的miRNA(或miRNA抑制剂)的片段。天然miRNA或其抑制剂也可以具有另外的核酸序列,如酌情在miRNA序列或其互补序列的5’或3’末端添加1、2、3、5、10或15个核酸。为了增加稳定性或者实现更好的效力,天然miRNA或其抑制剂可以经化学修饰,例如聚乙二醇化。所述活性典型是治疗先兆子痫一或多个症状的能力。
术语“同系物”是指具有所述miRNA相同活性的miRNA,如与所述miRNA调节相同基因。例如,已知动物miRNA通过使用在miRNA的5’末端的少如6-7个核苷酸(种子区)能识别靶mRNA。因此,典型地这种同系物是在5’末端具有与天然miRNA相同的至少6-8或者至少10-12个核苷酸的miRNA。所述miRNA可以是miRNA模拟物。这种模拟物是化学合成的dsRNA,其在转染进细胞中之后模拟内源miRNA。
所述治疗可包括如下一或多项措施:在先兆子痫、FGR或IUGR、妊娠肥胖、产后心肌病(心力衰竭)、癌症和糖尿病视网膜病变、心肌病、心肌梗塞、湿性黄斑变性以及其中血管发生是减少或加剧异常的其它病症的对象中增加miR-122、miR-374b、miR-152或者miR-195活性;或者降低miR-122、miR-374b、miR-152或者miR-195活性。
MiRNA活性可以通过正调节miRNA产生而增加。或者可以将miRNA或其功能片段或同系物注射进生物体中,例如注射进胎盘中。此外,miRNA或者其功能片段或同系物可以克隆进表达质粒中或者包装进腺病毒、AAV或者慢病毒中或者通过静脉内(IV)、肌内(IM)或者皮下(SC)注射进生物体中。
MiRNA可以被miRNA抑制剂抑制。例如,可以使用单链siRNA抑制剂,如miR-152或mirR-195抑制剂。此外,可以将miRNA、其功能片段或同系物的抑制剂克隆进表达质粒或者包装进腺病毒、AAV或慢病毒及通过静脉内(IV)、肌内(IM)或者皮下(SC)注射进生物体中。
本发明还提供了miR-122或miR-374b调节血管发生的应用。例如,可以将miRNA在细胞中低表达或者过表达以研究对于血管发生的作用。者可例如用于其中血管发生是减少或者加剧异常的疾病模型中或者例如妊娠妇女先兆子痫或者产后母亲心肌病中。其也可用于治疗癌症。
药物组合物包含两或多种的miR-122、miR-374b、miR-152抑制剂和miR-195抑制剂、或者其功能片段或同系物或者其药物可接受的盐。所述miRNA可以其天然形式或者修饰形式如聚乙二醇化形式使用。
miR-122和miR374b可用于通过将miRNAs导入其中血管发生是减少或加剧异常的对象中以抑制sFlt-1。其可例如用于体外细胞中或者动物模型体内,以调节sFlt-1及研究sFlt-1对于细胞或动物模型的作用。此外,其可例如用于先兆子痫妇女胎盘中以抑制sFlt-1水平。
通过参考仅为举例说明的下图描述本发明:
图1:可溶的fms相关的酪氨酸激酶1(sFlt-1)由各种化合物和生长因子调节。(A)将HUVEC细胞用VEGF-A(20ng/ml)和VEGF-E(20ng/ml)刺激24小时,分析条件培养基的sFlt-1情况。(B)将HUVEC细胞用2μM和20μM普伐他汀处理24小时,分析条件培养基的sFlt-1情况。(C)将HUVEC细胞用0.1μM、1μM和10μM Hemin载体或者VEGF-E刺激24小时。收集条件培养基并通过ELISA分析sFlt-1情况。
图2:MiR-122和miR-374由调节sFlt-1释放的各种制剂调节。将HUVEC细胞用10μM Hemin(A)、2μM或者20μM普伐他汀(B)20ng/ml VEGF-A(C)或者20ng/ml VEGF-E(D)处理24小时。分离处理细胞的总RNA,通过定量PCR测量miR-122和miR-374b的表达。将相对的miR-122和miR-374b表达归一化为RNU6。
图3:Hmoxl调节miR-122和miR-374b表达。将HUVEC细胞用过表达Hmoxl的腺病毒感染(A)或者用抗Hmoxl的siRNA转染(B)24小时,通过qPCR测量miR-122和miR-374b的相对表达。
图4:MiR-122和miR-374b在内皮细胞中直接调节sFlt-1释放。(A)图中示出miR-122和miR-374b的种子序列及sFlt-1 mRNA的3’-UTR中相应的靶位点。(B)将HUVEC细胞用模拟miR-122和miR-374b电穿孔并静置14小时。然后将转染的细胞在有或无VEGF-E(20ng/ml)的条件培养基中保温24-48小时,通过ELISA分析培养基中sFlt-1的情况。(C)将HUVEC细胞用miR-122和miR-374b的antagomir电穿孔并静置24小时。然后将转染的细胞在有或无普伐他汀(20μM)的条件培养基中保温24小时,通过ELISA分析培养基的sFlt-1情况。(D)将HUVEC细胞用siHO-1、miR-122或mi-374b单独或者组合电穿孔。然后将转染的细胞在条件培养基中用运载体或者VEGF-E(20ng/ml)刺激,通过ELISA分析sFlt-1的释放。
图5:MiR-122和miR-374b在人重度先兆子痫患者和先兆子痫RuPP小鼠模型中均降低,且在RuPP小鼠中与血压负相关。(A)收集重度先兆子痫患者(N=9-10)和对照妊娠妇女(N=9-10)的胎盘样品,通过qPCR分析miR-122和miR-374b的相对表达。(B)妊娠Sprague Dawley大鼠经历降低子宫灌注压力(RUPP)手术,收集足月胎盘组织。通过定量PCR分析假对照组(N=11-12)和RuPP大鼠(N=11-12)的胎盘中microRNA-122的表达。数据以平均值+SEM示出并通过Student t检验分析。将相对的miR-152表达标准化为RNU6。(C)线图示出平均动脉压(MAP)与RuPP小鼠(N=6)胎盘中miR-122或miR-374b表达之间的反相关性。相关性通过Pearson's相关系数计算(Pearson's相关性r=-0.8101,P=0.05,及r=-0.8451,P=0.034)。通过定量分析检测RuPP小鼠胎盘中miR-122和miR-374b的相对表达。
图6:将合成的microRNA-122和374b模拟物(B,mi-122和miR-374b)和antagomir(A,抗-122和抗-374b)电穿孔进HUVEC细胞中,miR-122和miR-374b的相对表达通过定量PCR测量。
图7:重度先兆子痫患者中miR-122或miR-374b表达与收缩压或舒张压(SBP和DBP)之间的反相关性。线图示出重度先兆子痫患者(N=8-9)胎盘中SBP或DBP与miR-122(A)或miR-374b(B)表达之间的反相关性。通过Pearson's相关系数计算相关。通过定量分析检测重度先兆子痫患者胎盘中miR-122和miR-374b的相对表达。
图9:miR-152的表达在重度先兆子痫妇女和动物先兆子痫模型的胎盘中增加。(A)收集重度先兆子痫患者(PE,N=14)和孕龄匹配的对照患者(N=17)的胎盘样品,使用qPCR分析miR-152表达。(B)妊娠Sprague Dawley大鼠经历降低子宫灌注压(RUPP)手术,收集胎盘组织。通过定量PCR分析正常孕妇(N.P.,N=12)和RuPP大鼠(n=17)的胎盘中microRNA-152的表达。(C)为妊娠C57BL/6小鼠在E 9.5通过尾静脉注射109PFU腺病毒,ad-CMV或者ad-sFlt-1。使用针对Flt-1的ELISA测量循环sFlt-1表达水平,在E 17.5在颈动脉中评估平均动脉压(MAP)(图17)。收集来自ad-CMV对照病毒(N=7)和ad-sFlt-1病毒(N=5)注射的小鼠的胎盘组织,使用定量PCR分析microRNA-152表达水平。此外,miR-152的表达在子宫内生长受限(IUGR)、肥胖孕妇和糖尿病小鼠模型中均增加。(D)IUGR(N=12)以及孕龄匹配的对照患者(N=17)的胎盘中miR-152的表达。(E,F)通过qPCR确定BMI超过30(内脏脂肪和皮下脂肪分别为N=25和24)或者正常BMI(18-25)(内脏脂肪N=22,皮下脂肪N=20)孕妇的内脏和皮下脂肪中miR-152的表达。(G,H)测量db/db糖尿病小鼠和对照小鼠品系的肝脏和肠系膜脂肪中miRNA-152的表达(N=5)。数据以平均值+SEM示出,并通过Student t检验分析。miR-152相对表达标准化为RNU6。***P<0.001vs第一孕期胎盘样品,##P<0.01vs第二孕期胎盘样品。
图10:缺氧、炎症细胞因子或者血管发生生长因子刺激增加在内皮细胞和胎盘外植体中microRNA-152的表达。将人胎盘外植体用(A)炎症细胞因子混合物(TNF-α,20ng/ml,IFN-γ,20ng/ml和IL-Ιβ,2ng/ml)、(B)缺氧(1%O2)条件或者(C)细胞因子混合物与缺氧条件组合进行处理,并使用qPCR分析miR-152表达。相似地,将HUEVC细胞用(D)缺氧(1%O2)条件、VEGF-E(20ng/ml,E)或者FGF-2(20ng/ml,F)处理24小时,并用于qPCR分析miR-152的表达。此外,在内皮细胞中miR-152的过表达抑制P1GF释放,但是不抑制sFlt-1和sEng。将HUVEC用模拟-152或者对照模拟-con电穿孔。在静置24小时后,将转染的细胞在条件培养基中用运载体或VEGF-E(20ng/ml)处理。在保温24小时后收集该培养基,使用ELISA分析sFlt-1(G)、sEng(H)和P1GF(I)浓度。数据以平均值+SEM示出,通过Student t检验分析。
图11:miR-152的阻断增加内皮细胞存活力,但是miR-152的抑制或者过表达不改变内皮细胞增殖或者凋亡。(A)将HUVEC细胞用抗-152(I)或者模拟-152(II)电穿孔。在恢复过夜后,将转染的细胞进行胰蛋白酶化并以1×104/孔铺板于96孔平板中,用运载体或者VEGF-A(20ng/ml)处理。48小时后,将这些细胞进行MTT分析。(B)对于细胞增殖,将抗-152(I)或模拟-152(II)转染的HUVEC细胞以4×104/孔铺板于24孔平板中,用VEGF-A(20ng/ml)或者VGEF-E(20ng/ml)刺激。处理48小时后,将细胞进行胰蛋白酶化,使用血细胞计数器在显微镜下计数每孔中细胞数目。数据以平均值+SEM示出,通过Student t检验分析。(C)将HUVEC细胞用抗-152(I)或模拟-152(II)转染并静置24小时。将细胞裂解物用裂解的胱天蛋白酶3的抗体印迹。
图12:miR-152负调节内皮细胞粘附及血管发生能力。(A)将HUVEC细胞用模拟-con或模拟-152电穿孔以过表达miR-152。48小时后,将2×104个模拟-152或模拟-con转染的细胞铺板于层黏蛋白(1mg/ml)、明胶(2%溶液)和胶原I型(0.1%solution)包被的96孔平板中,并用VEGF-A(20ng/ml)或VEGF-E(20ng/ml)处理30分钟。之后,将细胞用PBS洗涤3次,用Calcein AM荧光染料染色并进行荧光显微镜分析。在X4放大倍率下获取代表性图像,计数X4放大倍率下每个视野的粘附细胞数目。从至少5个单独视野中收集数据,以平均值+SEM示出。此外,将HUEVC细胞用模拟-152(B)或抗-152(C)电穿孔并在转染后静置48小时。随后,将1×104个转染的细胞铺板于基质胶包被的96孔平板上,用运载体、VEGF-A(20ng/ml)或者VEGF-E(20ng/ml)处理。在保温6-8小时后,在X4放大倍率下获取管状结构的相衬图像,量化每个X4视野的总管长度。从至少3-5个单独试验中收集数据,以平均值+SEM示出。
图13:在肢体缺血(HLI)小鼠模型中,miR-152的过表达减弱血管再生。为雄性C57B1/6小鼠(3-4月龄)动脉内注射ad-CMV或ad-152,然后通过手术切除左侧股动脉进行HLI。(A)在术后第3天(N=4,二者)和第14天(ad-CMV组N=11,ad-152组N=9)通过qPCR量化对照ad-CMV和ad-152腺病毒注射的腓肠肌中miR-152的表达。(B)对麻醉小鼠的足跖面通过LASER Doppler测量血流灌注。计算术后0/4和7天的血流恢复情况,以左侧缺血足与右侧非缺血足对比观测的血流比率表示。*P<0.05。(C)计算在第7天的血流恢复情况(ad-CMV组N=11,ad-152组N=9),在图右侧示出血流灌注的代表性图像。通过同工凝集素B4染色,通过组织学评估量化在术后14天的非缺血和缺血腓肠肌中毛细血管密度。图中示出在第14天在非缺血和缺血腓肠肌中毛细血管密度的代表性图像(D)和量化(E,每组N=3)。(F)通过qPCR测量ad-CMV注射的小鼠(每组N=9)的非缺血和缺血腓肠肌中miR-152的相对表达情况。
图14:miR-152的过表达降低胎儿体重及削弱妊娠小鼠胎盘中的血管。为雌性C57B1/6小鼠(3-4月龄)在胎龄11.5天(E11.5)经腹膜内注射5×109PFU ad-CMV或者ad-152,在胎龄17.5天终止妊娠并分析。(A)通过qPCR测量注射ad-CMV和ad-152的小鼠的血浆中miR-152的表达。(B)在胎龄17.5天(E17.5)的注射ad-CMV或ad-152的小鼠的胎儿的代表性图表。(C)在胎龄17.5天测量腹膜内注射ad-CMV(N=5)和ad-152(N=5)的C57B1/6小鼠中平均胎儿体重。(D)示出经腹膜内注射ad-CMV和ad-152的小鼠胎盘的代表图表。(E)在荧光显微镜下在10X和20X放大倍率观测的注射ad-CMV和ad-152的小鼠胎盘中血绒毛迷路区的同工凝集素B4和DAPI染色。
图15:MiR-152直接靶向ITGA5。(A)图中示出miR-152的种子序列及ITGA5mRNA的3'-UTR中相应靶位点。(B)将HUVEC细胞用抗-152或模拟-152电穿孔并静置48小时之后进行处理。然后将转染的细胞用VEGF-A(20ng/ml)刺激1小时,并用Neuropilin 1(NRP1)、小窝蛋白-2(CAV-2)、fms相关酪氨酸激酶1(Flt-1)和ITGA5的抗体进行Western印迹。(C)将HEK293细胞用pmiR-ITGA5或pmiR-ITGA5M(microRNA靶位点中突变)与模拟-con或模拟-152一起转染。含有海肾荧光素酶基因的质粒也进行转染作为转染对照物。在保温过夜后,测量相对荧光素酶活性并标准化为海肾荧光素酶活性。(D)ITGA5的过表达拯救模拟-152抑制的血管形成。将HUEVC细胞用ITGA5腺病毒感染过夜,然后用mi-con或mi-152电穿孔。24-48小时后,将细胞胰蛋白酶化以进行基质胶管形成分析。在相衬显微镜下在X4放大倍率获取荧光图像,计算每个X4视野的总管长度。ad-ITGA5的过表达通过western印迹使用抗-ITGA5抗体(插入)证实。
图16:在小鼠模型和重度先兆子痫患者中,ITGA5表达与miR-152水平反相关。(A)将2×1012GC的对照腺伴随病毒(AAV-con)或miR-152 AAV病毒注射进C57BL/6小鼠的内收肌中,在4周后收集肌肉样品并使用ITGA5和GFP抗体进行western印迹。计算ITGA5相对表达,以在注射AAV-152的肌肉中与注射AAV-con的肌肉中ITGA5表达的比率表示(每组N=4)。(B)将后肢缺血的腓肠肌样品用ITGA5抗体印迹。计算在注射ad-CMV(N=11)和ad-152(N=9)的小鼠的缺血和非缺血肌肉中ITGA5表达的比率。(C)通过western印迹检测经腹膜内注射ad-CMV(N=4)和ad-152(N=4)的小鼠胎盘中ITGA5蛋白质表达情况。(D)通过定量PCR检测8个重度先兆子痫患者的胎盘中miR-152的相对表达情况,相应ITGA5蛋白质表达也通过western印迹使用抗-ITGA5抗体确定。(E)线图示出重度先兆子痫患者的胎盘中miR-152表达与ITGA5水平之间的反相关性。从8个重度先兆子痫患者样品中通过Pearson's相关系数计算相关性(Pearson's相关性r=-0.823,P=0.0121)。
图17:注射ad-sFlt-1的小鼠中sFlt-1表达及平均动脉压(MAP)。为妊娠的C57BL/6小鼠在E 9.5通过尾静脉注射109PFU腺病毒ad-CMV或ad-sFlt-1。(A)在E 17.5在注射ad-CMV(N=6)和ad-sFlt-1(N=5)的小鼠血浆中使用ELISA针对Flt-1测量sFlt-1表达水平。(B)在E 17.5在注射ad-CMV(N=4)和ad-sFlt-1(N=5)的小鼠颈动脉中计算平均动脉压(MAP)。
图18:将合成的microRNA-152antagomir(A,抗-152)和模拟物(B,模拟-152)电穿孔进HUVEC细胞中,通过定量PCR测量miR-152相对表达水平。
图19:MiR-152正调节HUVEC细胞迁移。将HUEVC细胞用抗-microRNA-152(A)或者模拟-152(B)电穿孔。在恢复48小时后,在12或6孔平板中转染的融合细胞中心进行刮擦。在进行刮擦后0小时及16-20小时立即测量刮擦宽度。通过减去在0小时和16-24小时的刮擦宽度计算细胞迁移距离。从至少4-5个孔收集数据,在显微镜下测量每次刮擦的三个随机区域。
图20:(A)用于microRNA靶确认的pMirTarget质粒图。(B)在用pmir-ITGA5质粒转染的HUEVC细胞中红色荧光蛋白(RFP)的表达。
图21:AAV-152感染的C57BL/6小鼠中miR-152的表达。将每只动物2×1012GC的AAV-con或AAV-152局部注射进左侧内收肌(LAD),并将相同体积的盐水溶液注射进右侧(RAD)作为对照。4周后,收集左侧和右侧内收肌,将每个样品分为两部分进行qPCR分析、western印迹及冷冻切片。(A)通过定量PCR分析4周后注射AAVl-con和AAV1-152的内收肌中miR-152的表达。(B)将LAD和RAD的冷冻肌肉样品用低温器切成5μM切片,使用荧光显微镜在X20放大倍率获取GFP表达图像。(C)在LAD肌肉中重叠的GFP与DAPI代表性图像。
图22:在重度先兆子痫妇女和动物先兆子痫模型的胎盘中,miR-195表达增加。(A)收集来自第一孕期(N=6)、第二孕期(N=6)和第三孕期(N=4)的妊娠妇女的胎盘组织,通过qPCR测量miR-195相对表达水平。此外,通过qPCR确定内脏脂肪(B)和皮下脂肪(C)的BMI超过30(两组分别为N=25和24)或者正常BMI(18-25)(内脏组N=22,皮下组N=20)妊娠妇女的中miR-195表达水平。(D)测量重度先兆子痫(N=14)、IUGR(宫内生长受限,N=12)以及孕龄匹配的对照患者(N=17)的胎盘中miR-195表达水平。(E)在E9.5通过尾静脉为妊娠C57BL/6小鼠注射109PFU腺病毒ad-CMV或ad-sFlt-1。血液循环中sFlt-1表达水平使用ELISA针对Flt-1测量,在E 17.5在颈动脉中评估平均动脉压(MAP)(图26)。收集注射ad-CMV对照病毒(N=7)和ad-sFlt-1病毒(N=5)的小鼠胎盘组织,使用qPCR分析miR-195表达情况。(F)为妊娠Sprague Dawley大鼠进行降低子宫灌注压(RUPP)手术并收集胎盘组织。通过qPCR分析正常妊娠大鼠(N.P.,N=12)和RuPP大鼠(n=17)胎盘中miR-195表达水平。此外,单独的炎症细胞因子混合物或者组合缺氧条件增加内皮细胞和胎盘外植体中microRNA-195表达。(G,H)将HUEVCs用单独的炎症细胞因子混合物(TNF-α,20ng/ml,IFN-γ,20ng/ml及IL-Ιβ,2ng/ml)或者组合缺氧条件(1%O2)刺激12-24小时,通过qPCR测量miR-195表达。(I)将胎盘外植体用炎症细胞因子混合物组合缺氧条件(1%O2)处理,通过qPCR测量miR-195表达。
图23:microRNA-195负调节内皮细胞增殖、存活力、粘附能力和血管发生。(A)将HUVECs用mi-con或mi-195电穿孔并用运载体或者VEGF-A(20ng/ml)处理24小时。收集培养上清针对P1GF进行ELISA测定,从三个单独试验中产生数据。(B)将HUEVC用mi-195转染并铺板于24孔平板中,用VEGF-A(20ng/ml)或VGEF-E(20ng/ml)刺激。处理48小时后,将细胞胰蛋白酶化,并使用血细胞计数器在显微镜下计数每孔的细胞数目。(C)将HUEVC细胞用mi-195电穿孔并以1×104/孔密度铺板于96孔平板中,用运载体或者VEGF-A(20ng/ml)处理。保温48小时后,将细胞用于MTT测定。(D)将HUVEC细胞用mi-195或者对照mi-con电穿孔。48小时后,将2×104个mi-195或者mi-con转染的细胞铺板于2%明胶包被的96孔平板中,用VEGF-A(20ng/ml)或VEGF-E(20ng/ml)处理30分钟。随后,将细胞用PBS洗涤3次,用Calcein AM荧光染料染色及进行荧光显微镜分析。计数在10X放大倍率下每个视野的粘附细胞数目。从至少5个单独视野中收集数据。(E)将HUEVC细胞用mi-195电穿孔并静置48小时。之后,将1×104个转染的细胞铺板于基质胶包被的96孔平板中,用运载体、VEGF-A或VEGF-E处理。在保温6-8小时后,获取管样结构的图像进行定量分析。
图24:microRNA-195增加HUVEC细胞迁移。将HUEVC细胞用antagomir抗-195(A)或者模拟物mi-195(B)电穿孔并静置48小时以达到铺满融合。在12孔或6孔平板中在转染的融合细胞的孔中心位置进行刮擦。在进行刮擦后0小时和16-24小时立即测量刮擦宽度。通过减去在0小时和16-24小时的刮擦宽度计算细胞迁移距离。从至少4-5个孔中收集数据,在显微镜下测量每次刮擦的三个随机区域。
图25:miR-195直接靶向eNOS。(A)图表示出人miR-195的种子序列及不同物种eNOS mRNA的3'-UTR中相应靶位点。(B)将HEK293细胞用pmiR-eNOS或者pmiR-eNOSM(micro RNA靶位点中突变)以及mi-con或mi-195一起转染。共转染含有海肾荧光素酶基因的质粒作为对照。在保温过夜后,测量相对荧光素酶活性并标准化为海肾对照活性。(C)将HUVEC用mi-con或mi-195转染,分离总RNA以进行qPCR分析。使用针对eNOS编码序列的引物测量eNOS mRNA的表达。(D)将HUVEC用mi-con或mi-195电穿孔,在处理之前恢复48小时。48小时之后,将细胞用VEGF-A(20ng/ml)刺激1小时,及用抗-磷-Enos和总Enos抗体进行Western印迹。(E)使用Image J量化磷-eNOS和总NOS的相对表达。(F)将HUEVC用ad-195感染(MOI=100),使用Sievers NO化学发光分析仪分析一氧化氮释放情况。(G)eNOS的过表达拯救mi-195介导的管形成降低。将HUEVC用腺病毒ad-eNOS感染(MOI=50)过夜,随后用mi-con或mi-195电穿孔。24-48小时后,将细胞胰蛋白酶化以进行基质胶(matri-gel)管形成测定。在保温6-8小时后在X4放大倍数取管状结构的相位对比图像。(H)通过Image Pro Plus软件量化每个X4视野的总管长度。从至少3-5个单独试验中收集数据并以平均值+SEM示出。
图26:注射ad-sFlt-1的小鼠中sFlt-1表达及平均动脉压(MAP)。为妊娠C57BL/6小鼠在E 9.5通过尾静脉注射109PFU腺病毒,ad-CMV或ad-sFlt-1。(A)使用针对Flt-1的ELISA测量在E 17.5在注射ad-CMV(N=6)和ad-sFlt-1(N=5)的小鼠的血浆中sFlt-1表达水平。(B)在E 17.5在注射ad-CMV(N=4)和ad-sFlt-1(N=5)的小鼠颈动脉中评估平均动脉压(MAP)。数据以平均值+SEM示出并通过未配对t检验分析。
图27:用microRNA-195的合成的antagomir(抗-195)或者模拟物(mi-195)转染的或者用过表达miR-195的腺病毒(ad-195)感染的HUVEC中miR-195的表达。(A)通过定量PCR确定合成的抗-195和mi-195的效力。抗-195显著降低而mi-195增加在转染的HUVEC细胞中microRNA-195表达。(B)将HUVEC用ad-195感染,MOI为100。通过实时定量PCR测量miR-195的表达并与对照腺病毒感染的细胞对比。
图28:(A)用于microRNA靶确认的pMirTarget质粒的图表。(B)用pmir-eNOS质粒转染的HUEVC细胞中红色荧光蛋白(RFP)的表达。
miR-122和miR-374b
方法与材料:
试剂与化合物
重组生长因子、血管内皮生长因子A(VEGF-A)、VEGF-E购自RELIATech(Brauschweig,Germany)。血红素(Hemin)和普伐他汀(pravastatin)购自Sigma-Aldrich(USA)。M199培养基购自Invitrogen(Paisley,UK)。miR-122和miR-374b的定量PCR引物、qScriptTMmicroRNAcDNA和SynthesisGreen SuperMix试剂盒购自Quanta Biosciences。
人胎盘组织收集和制备
伦理委员会学会(Institutional Ethics Committee)许可胎盘组织收集并获得书面知情同意书。所有妇女从注册至分娩均被前瞻性跟踪。从妊娠并发先兆子痫妇女(N=14)及宫内生长受限妇女(IUGR,N=12)及从血压正常妊娠妇女(N=17)中收集人胎盘组织。将收集的胎盘组织进一步用于定量PCR和western印迹。先兆子痫定义为在至少两次连续测量血压>140/90mm Hg及母体蛋白尿为至少300mg/24小时,IUGR定义为胎儿的估计重量低于其胎龄体重的10%,腹围低于2.5%。
降低子宫灌注压(RUPP)的先兆子痫大鼠模型
RUPP先兆子痫模型的胎盘样品友情得自Dr.Fergus McCarthy(Cork University Maternity Hospital,Wilton,Cork,Ireland)。经历RUPP手术的大鼠的.实验程序和鉴定在先前描述(McCarthy et al.,2011)。
细胞培养
分离人脐静脉内皮细胞(HUVEC)并如先前所述在M199培养基中培养(Bussolati et al,2001).对第三代或第四代HUVEC进行试验。
基于PCR的microRNA微阵列
使用基于qPCR的阵列系统确定用血红素刺激的HUVEC细胞中MicroRNA表达模式。将HUVEC细胞用10μM Hemin处理24小时,分离总RNA进行Human miFinder RT2miRNA PCR阵列分析(Qiagen,Cat.No.331211)。根据厂商指导进行PCR程序及结果分析。
miR-122和miR-374b的miRNA模拟物和抑制剂
miR-122(mi-122和抗-122)及miR-374b(mi-374b和抗-374b)的化学合成的双链microRNA模拟物和单链抑制剂购自Qiagen,其效力通过Qpcr检测(图6A、B)。
将HUVECs进行胰蛋白酶化,并使用如先前所述电穿孔技术(Amaxa GmbH,Cologne,Germany)将1×106个细胞用~0.6μg模拟-122或模拟-374b、~3μg抗-122或抗-374或者对照分子电穿孔(Cudmore et al.,2007)。
腺病毒基因转移
如先前所述使用重组的复制缺陷的腺病毒编码大鼠HO-1(AdHO-1)(Cudmore et al.,2008)。
针对Hmoxl的siRNA
针对Hmoxl的siRNA如先前所述(Cudmore et al.,2008)。
实时聚合酶链反应
如先前所述进行样品制备与实时定量PCR(Cudmore et al.,2007)。
酶联免疫吸附测定
对于人sFlt-1的酶联免疫吸附测定(ELISA)试剂盒得自R&D Systems,根据厂商说明书进行。
MicroRNA靶测定
含有sFlt-1的3'UTR的质粒pmiR-Flt1得自GeneCopoeia(Cat No.HmiT054531-MT01,MD,USA)。将萤火虫荧光素酶cDNA与sFlt-1的3'UTR融合,对照海肾荧光素酶基因由在相同质粒中CMV启动子驱动。使用定向诱变技术产生含有miR-122和miR-374b结合位点的突变质粒pmiR-Flt1-M1和pmiR-Flt1-M2。miR-122的结合位点由TTTGTAGCATTGTCATCACTCCT突变为TTTGTCGACGGATAGAGAAT。miR-374b的两个结合位点从GTC AAAAT AGATTATT ATAA突变为GTCAAGAGCAAGGCGCA,及从TACAATATTTGTACTATTATAT突变为TACAATATTTAGACGCGCT。
对于microRNA靶测定,将HEK293细胞用pmiR-ITGA5或pmiR-ITGA5M及模拟-con或模拟-152一起转染。在保温过夜后,测量相对萤火虫荧光素酶活性并标准化为海肾荧光素酶活性,根据Dual-Reporter Assay System(E1910,Promega)的厂商方案进行。
microRNA靶预测
使用在线程序预测miR-122和miR-374b的直接靶位,microRNA.org(http://www.microrna.org/microrna/home.do)、PicTar(http://pictar.mdc-beriin.de/)及Target Scan(http://www.targetscaii.org/)。
统计学分析
所有数据均以平均值+S.E.M表示。使用Student's t检验或者Mann-Whitney U检验进行统计学对比。统计学意义设定为P值<0.05。
结果:
使用微阵列鉴别应答血红素刺激的microRNA
我们先前的研究示出内皮细胞中sFlt-1的释放可以由血管内皮生长因子(VEGF)和血红素加氧酶1(Hmox1)(Cudmore et al.,2007;Ahmad et al.,2011)调节。然而,还不清楚sFlt-1调节的分子学机制。因此,我们假设microRNAs在VEGF或Hmox1下游可被调节并直接靶向sFlt-1 mRNA翻译。初步研究证实我们先前的观测,即VEGF-E显著诱导sFlt-1从内皮细胞中释放(图1A),而作为强力Hmoxl诱导剂的他汀类制剂在2μM和20μM均降低sFlt-1水平(图IB)。此外,另一Hmoxl诱导剂血红素正如预期地在内皮细胞在10μM降低sFlt-1释放,这种降低不能通过VEGF-E刺激而恢复,提示VEGF-E和血红素是单独的sFlt-1调节物(图1C)。
为了鉴别应答血红素刺激的microRNA,使用分离自血红素处理的HUVEC细胞的RNA样品进行基于qPCR的微阵列。简而言之,将HUVEC细胞用10μM血红素处理24小时,从Human miFinder RT2 miRNA PCR阵列(Qiagen,Cat.No.331211)中分离总RNA。使用由厂商提供的程序分析结果并确定基于血红素处理不同表达的microRNA。概括而言,鉴别了24个正调节的microRNA及26个负调节的microRNA。为了进一步选择直接靶向sFlt-1的microRNA,使用在线生物信息工具包括microRNA.org、PicTar和Target Scan分析正调节的microRNA直接靶向sFlt-1 mRNA的可能性。最终,预测3个microRNA即miR-122、miR-144和miR-374b直接靶向sFlt-1并用于进一步研究。
miR-122和miR-374b表达由VEGF和Hmox1诱导剂调节
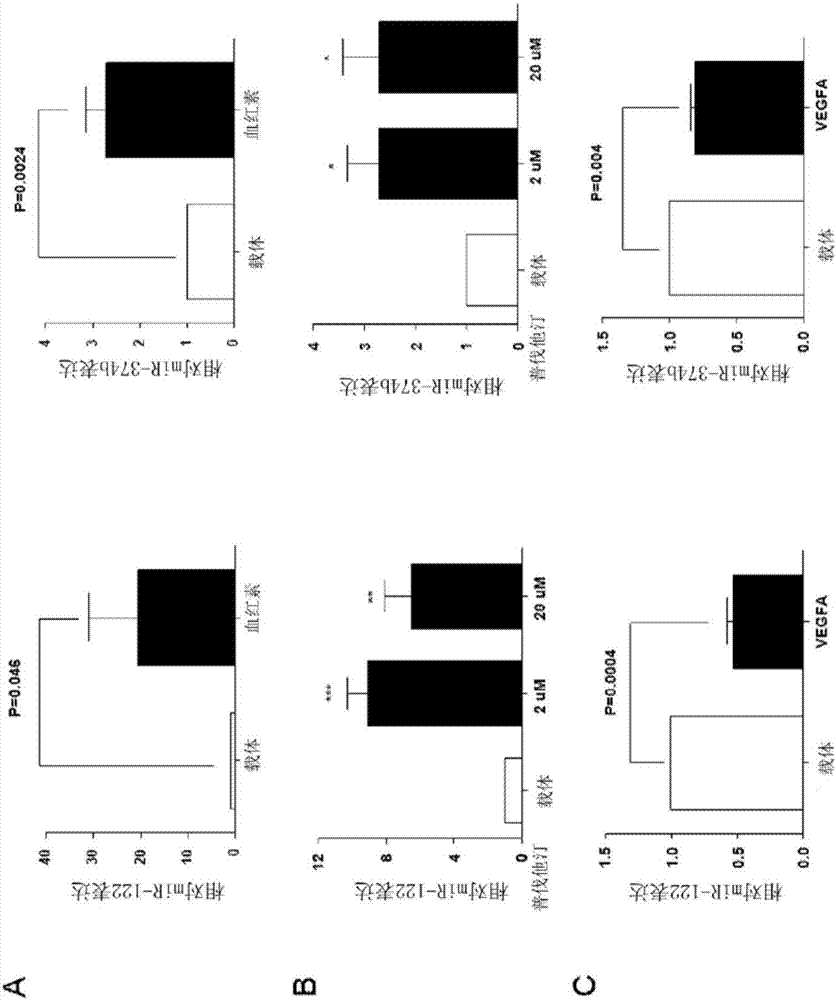
由于预测miR-122、miR-144和miR-374b是sFlt-1的直接调节物及VEGF或Hmox1诱导剂调节sFlt-1释放,我们检测这些microRNA是否可以由VEGF或Hmox1诱导剂调节。将HUVEC用血红素(10μM)、VEGF-A(20ng/ml)或者VEGF-E(20ng/ml)处理,通过qPCR确定这些microRNA的表达。用血红素处理使miR-122和miR-374b表达分别增加20倍和2.7倍(图2A),但是对于miR-144表达无显著影响(数据未示出)。此外,2μM和20μM普伐他汀刺激显著诱导miR-122和miR-374b表达分别为3-9倍和2.7倍(图2B)。相反,用VEGF-A和VEGF-E刺激使miR-122表达分别降低47%和75%(图2C,D)。用VEGF-A和VEGF-E处理使miR-374b的表达分别降低20%和34%(图2C,D)。
Hmox1调节miR-122和miR-374b表达
由于Hmox1诱导剂调节miR-122和miR-374b表达,我们研究了这些microRNA是否可以由Hmox1直接调节。将HUVEC细胞用过表达Hmoxl的腺病毒感染,这些细胞中miR-122和miR-374b的表达通过qPCR量化。正如预期的,Hmoxl的过表达使miR-122和miR-374表达分别增加7.8倍和70倍(图3A),但是miR-144表达未改变。相反,用针对Hmox1的siRNA转染的HUVEC细胞示出miR-122和miR-374b表达分别降低85%和25%(图3B)。
MiR-122和miR-374b直接靶向directly target sFlt-1mRNA
先前的生物信息分析推测miR-122和miR-374b可以直接结合sFlt-1 mRNA的3’非翻译区(UTR)(图4A)。为了进一步证实sFlt-1 mRNA由这些microRNA直接靶向,使用sFlt-1的3’-UTR及miR-122(mi-122)或miR-374b(mi-374b)模拟物进行荧光素酶分析。将含有sFlt-1的3’-UTR及其完整序列或者miR-122和miR-374b结合位点被修饰的突变序列的质粒与萤火虫荧光素酶融合并转染进HEK293细胞中。将这些细胞也用模拟对照物mi-122和mi-374b单独或者组合共转染。荧光素酶活性测定表明mi-122和mi-374b转染与模拟对照物转染的细胞相比显著抑制荧光素酶活性,而这种抑制作用在含有修饰的microRNA结合位点的突变质粒中未观测到(图4A)。令人感兴趣地,mi-122与mi-374b组合与单独的mi-122或mi-374b转染相比不进一步降低荧光素酶活性(图4A)。miR-122和miR-374b对sFlt-1的靶向另外通过ELISA证实,通过测量用mi-122和mi-374b单独或者组合转染的HUVEC培养基中sFlt-1水平而证实。在用运载体和VEGF-E刺激的两种条件下,用mi-122或mi-374b转染的HUVEC的培养基中的sFlt-1水平与模拟对照物相比降低大约30%(图4B)。相反,用miR-122(抗-122)和miR-374b(抗-374b)的antagomir转染HUVEC显著释放更多的sFlt-1进入培养基中,尤其是在用他汀类制剂刺激的条件下(图4C)。更重要地,这些microRNA在Hmox1下游起作用靶向sFlt-1表达。将HUVEC用siHO-1组合mi-122或mi-374b转染,通过ELISA测量sFlt-1释放。正如预期的,siHO-1转染显著增加sFlt-1表达,而用mi-122或者mi-374b共转染则完全消除这种由siHO-1介导的增加(图4D)。
miR-122和miR-374b在先兆子痫发病机制中的意义
由于血管发生失衡是导致先兆子痫的主要因素及sFlt-1拮抗VEGF信号途径(Levine et al.,2004;Venkatesha et al.,2006;Ramma and Ahmed,2011),我们通过qPCR测量了在先兆子痫患者和RuPP先兆子痫小鼠模型的胎盘中miR-122和miR-374b的表达。miR-122和miR-374b在先兆子痫胎盘中的表达与孕龄匹配的对照组相比分别降低大约60%和45%(图5A)。进一步地,其在RuPP小鼠胎盘中的表达与假对照组相比分别降低大约81%和64%(图5B)。更重要地,在RuPP小鼠(图5C,N=6,分别R=-0.81及R=-0.845)及注射ad-sFlt-1病毒的小鼠模型(图8,N=5,分别R=-0.9778及R=-0.9397)中,miR-122和miR-374b的表达与血压负相关。此外,这种相关性在重度先兆子痫患者中进一步证实。miR-122和miR-374b在重度先兆子痫患者胎盘中的表达与其收缩压和舒张压(SBP和DBP)负相关(图7,N=8-9)。
miR-152
方法和材料:
试剂和抗体
重组生长因子、血管内皮生长因子A(VEGF-A)、VEGF-E、肿瘤坏死因子α(TNF-α)、干扰素γ(IFN-γ)及白细胞介素1β(IL-1β)购自KELIATech(Brauschweig,Germany)。抗ITGA5兔抗体、Caspase 3、Flt-1、CAV2和NRP1得自Cell Signaling Technology(Danvers,MA 01923,United States)。小鼠抗-β-肌动蛋白抗体和兔抗-GAPDH分别得自Sigma-Aldrich(A5441,St.Louis,MO,United States)和Santa Cruze Biotech。miR-152的定量PCR引物、qScriptTM microRNA Cdna及SynthesisGreen SuperMix试剂盒购自Quanta Biosciences。低生长因子基质胶(Growth factor reduced Matrigel)购自Becton Dickinson(Oxford,UK)。M199培养基购自Invitrogen(Paisley,UK)。Calcein AM荧光染料购自BD Bioscience(P.O.Box 999 Sparks,MD,USA21152)。DyLight 594标记的Lycopersicon Esculentum(Tomato)Lectin(LEL,TL)购自Vector Laboratories(Burlingame,CA94010)。Alexa594 Conjugated Isolectin GS-IB4购自Life Technologies(Paisley,UK)。OCT化合物购自VWR International Ltd.(Leicestershire,LE17 4XN,England)。
人胎盘组织收集和制备
伦理委员会学会(Institutional Ethics Committee)许可胎盘组织收集并获得书面知情同意书。所有妇女从注册至分娩均被前瞻性跟踪。从妊娠并发先兆子痫妇女(N=14)及宫内生长受限妇女(IUGR,N=11)及从血压正常妊娠妇女(N=17)中收集人胎盘组织。将收集的胎盘组织进一步用于定量PCR和western印迹。先兆子痫定义为在至少两次连续测量血压>140/90mm Hg及母体蛋白尿为至少300mg/24小时,IUGR定义为胎儿的估计重量低于其胎龄体重的10%,腹围低于2.5%。
细胞和胎盘外植体培养
分离人脐静脉内皮细胞(HUVEC)并如先前所述在M199培养基中培养(Bussolati et al,2001)。对第三代或第四代HUVEC进行试验。从经历选择性终止妊娠的正常孕妇中获得第一孕期胎盘组织(6-9周孕龄)。如先前所述制备胎盘绒毛组织外植体(Ahmad and Ahmed,2004)。简而言之,将人胎盘绒毛组织外植体在检测物质或者缺氧条件刺激下保温,收集进行miR-152定量PCR。。为了产生缺氧条件,将70-80%铺满融合的HUEVCs或者胎盘外植体在保温仪中在具有1%O2和5%CO2及在37℃条件下培养。
针对miR-152的MiRNA模拟物和抑制剂
针对miR-152的化学合成的双链micro RNA模拟物和单链抑制剂(模拟-152和抗-152)购自Qiagen。
将HUVEC进行胰蛋白酶化,并使用如先前所述(Cudmore et al.,2007)电穿孔技术将l×106个细胞用~0.6μg模拟-152、~3μg抗-152或者对照分子电穿孔(Amaxa GmbH,Cologne,Germany)。
腺病毒及腺伴随病毒动物注射
腺病毒ad-sFlt-1友情得自Prof.Richard Mulligan(Harvard Medical School,Boston,USA)。将ad-sFlt-1和ad-CMV对照病毒扩增并滴定,将109PFU腺病毒在E 9.5通过尾静脉注射进C57BL/6小鼠中以过表达sFlt-1。
产生过表达miR-152的腺伴随病毒(血清型1)(AAV- 152)并由Vector Biolabs(Philadelphia,USA)滴定。将GFP与miR-152 cDNA融合作为报道基因。在后肢肌肉注射中,将2×1012GC的AAVl-con或者AAV1-152 AAVs局部注射进C57BL/6小鼠的左侧内收肌中,相同体积的盐水溶液注射进右侧肌肉中作为对照。通过共表达的GFP报道蛋白证实病毒感染,miR-152的过表达在注射4周后在所述肌肉中通过定量PCR确定(图21)。
降低子宫灌注压(RUPP)的大鼠先兆子痫模型
RUPP先兆子痫模型的胎盘样品友情得自Dr.Fergus McCarthy(Cork University Maternity Hospital,Wilton,Cork,Ireland)。试验程序及经历RUPP手术的大鼠的鉴别在先前描述(McCarthy et al.,2011)。
实时聚合酶链反应
如先前所述进行样品制备和实时定量PCR(Cudmore et al.,2007)。
Western印迹
如先前所述将来自HUVEC或动物组织的总蛋白质在RIPA缓冲液中裂解并进行分析(Ahmad and Ahmed,2004)。
酶联免疫吸附测定
对于人sFlt-1、sEng和PIGF的酶联免疫吸附测定(ELISA)的试剂盒得自R&D Systems,根据厂商说明书进行测定。
体外细胞粘附测定
将HUVECs用模拟-152或者对照模拟-con电穿孔。48小时后,将2X104模拟-152或模拟-con转染的细胞铺板于2%明胶包被的96孔平板中,用VEGF-A(20ng/ml)或VEGF-E(20ng/ml)处理30分钟。之后,将细胞用PBS洗涤3次,用Calcein AM荧光染料染色并进行荧光显微镜分析。计数在4×放大倍率下每个视野粘附细胞的数目。
体外管形成测定
如先前所述确定低生长因子基质胶上模拟-152、抗-152或模拟-152及ad-ITGA5处理的HUEVC的毛细血管样结构的形成(Bussolati et al,2001)。
体外细胞存活力和增殖测定
将HUVEC细胞用抗-152或模拟-152电穿孔。在过夜恢复后,将转染的细胞进行胰蛋白酶化并以1×104/孔密度铺板于96孔平板中,用运载体或VEGF-A(20ng/ml)处理。48小时后,将这些细胞使用细胞生长确定试剂盒(CDG1,Sigma-Aldrich)进行MTT测定。对于细胞增殖测定,将转染的细胞以4×104/孔密度铺板于24孔平板中,用VEGF-A(20ng/ml)或者VGEF-E(20ng/ml)刺激。48小时后,将细胞进行胰蛋白酶化,并在显微镜下使用血细胞计数器计数每孔中细胞数目。
体外细胞迁移测定
将HUEVC细胞用抗-152或模拟-152电穿孔。在恢复过夜后,在12或6孔平板中转染的铺满融合细胞的中心进行刮擦。在刮擦后0小时和16-24小时立即测量刮擦宽度。通过减去在0小时和16-24小时的刮擦宽度计算细胞迁移距离。
后肢缺血小鼠模型
动物研究方案由Aston University Ethical Review Committee许可,并根据United Kingdom Animals(Scientific Procedures)Act,1986所述进行。为雄性C57B1/6小鼠(3-4月龄)经动脉内注射ad-152或ad-CMV,然后如先前所述(Murdoch et al.,2014)通过手术切除左侧股动脉进行HLI。分离左侧股动脉。在插入套管之前对静脉和动脉进行暂时近端结扎,并将ad-152或ad-CMV注射进左侧股动脉中。5分钟后,在切除之前大约1.5mm的近端和远端位置对动脉进行两个进一步结扎。除去与静脉的暂时结扎。如先前所述(Murdoch et al.,2014),在麻醉小鼠(氯胺酮(100mg/kg)、甲苯噻嗪(10mg/kg);i.p.)的足跖面通过激光多普勒LASER Doppler(Moor Instruments UK)测量血流灌注情况。血流量恢复通过在左侧缺血足与右侧非缺血足相比观测的血流量比率计算。通过同工凝集素B4染色,通过组织学评估量化非缺血和缺血腓肠肌中毛细血管密度。
小鼠妊娠研究
交配3-4月龄的C57BL/6小鼠。妊娠第一天(E0.5)定义为第二天早上存在阴道栓。在第E11.5天将妊娠小鼠随机分为两组,经腹膜内注射ad-152或ad-CMV,剂量为5×109PFU/小鼠。6天后,将妊娠小鼠使用氯胺酮/甲苯噻嗪混合物麻醉,随后处死取样。技术活的胎儿和胎盘并称重。取血样并收集肾、肝、脾和胎盘。通过qPCR测量在血液和胎盘中miR-152表达。随后将胎盘组织用同工凝集素B4染色以分析胎盘血管结构。
micro RNA靶测定
含有ITGA5非翻译区(UTR)的人cDNA购自Origene(Rockville,MD20850,USA),将该UTR区亚克隆进pMirTarget中产生ITGA5-UTR-荧光素酶(萤火虫)表达质粒pmiR-ITGA5。pmiR-ITGA5在内皮细胞中的成功转染通过红色荧光蛋白(RFP)的表达确认(图21)。使用定向诱变试剂盒(Stratagene)与含有突变位点的PCR引物产生在miR-152和ITGA5-UTR结合位点有突变的质粒,所得质粒称作pmiR-ITGA5M。用于PCR扩增的引物是mITGA5-F,TCCCTCCCCCCCATGCTGTGG,及mITGA5-R,TGTAAACAAGGGTCCACAGCA。对于microRNA靶测定,将HEK293细胞用pmiR-ITGA5或pmiR-ITGA5M与模拟-con或模拟-152一起转染。对含有海肾荧光素酶基因的质粒也进行转染作为内部对照。在保温过夜后,根据Dual-Reporter Assay System(E1910,Promega)的厂商方案测量相对萤火虫荧光素酶活性并标准化为海肾活性。
microRNA靶预测
使用在线程序预测miR-152的直接靶位:microRNA.org(http://www.microrna.org/microrna/home.do)、PicTar(http://pictar.mdc-berlin.de/)和Target Scan(litip://www.targetscan.org/)。
微阵列分析
将HUVEC细胞用模拟-152转染并收集总RNA样品,使用Affymetrix genechip Human Gene 1.0 ST进行全基因组阵列分析。使用+1.25倍截止值选择基于模拟-152转染的正调节的和负调节的基因。
统计学分析
所有数据均以平均值+S.E.M表示。使用Student's t检验或者Mann-Whitney U检验进行统计学对比。统计学意义设定为p<0.05。
结果:
在先兆子痫患者及动物模型、IUGR、妊娠肥胖妇女和糖尿病小鼠模型的胎盘中miR-152表达增加
尽管Zhu及其同事证实在先兆子痫中miR-152表达水平升高(Zhu et al.,2009),但是其研究中使用的胎盘样品是从平均孕龄35.9周的先兆子痫妇女中收集的。为了在更准确定义的重度先兆子痫患者中研究miR-152表达,从平均孕龄29.7周的先兆子痫妇女中收集胎盘样品并通过定量PCR检验miR-152表达。定量PCR表明在先兆子痫胎盘(N=14)中与孕龄匹配的对照血压正常孕妇胎盘(N=17)中相比miR-152显著增加,但是在IUGR胎盘(N=11)中无变化,提示miR-152在先兆子痫中是特别正调节的(图9A)。
为了进一步证实先兆子痫中miR-152的正调节,在两个先兆子痫动物模型中检验miR-152的表达。接受ad-sFlt-1腺病毒注射的妊娠小鼠与注射ad-CMV病毒的对照小鼠相比呈现出显著增加的胎盘miR-152水平(图9B)以及增加的sFlt-1表达和平均动脉压(图17)。此外,经历降低子宫灌注压手术的妊娠Sprague Dawley大鼠(N=17)的胎盘中的miR-152表达与正常妊娠大鼠(N=12,图9C)相比也显著增加,提示miR-152增加与先兆子痫发病机制相关。重度先兆子痫患者与胎儿生长受限(IUGR)通常相关联,因此我们检验了在IUGR患者胎盘中miR-152水平。IUGR患者(N=11)中的miR-152表达与孕龄匹配的对照组(N=17,图9D)相比增加88%。
由于妊娠肥胖妇女具有先兆子痫高危险性,我们检验了在妊娠肥胖妇女的脂肪组织中miR-152的表达。正如预期的,妊娠肥胖妇女的内脏脂肪(N=25)和皮下脂肪(N=24)中miR-152表达与瘦妊娠妇女(N=20-22)相比显著增加(图9E和F)。此外,我们检测了糖尿病db/db小鼠的肝脏和肠系膜脂肪中miR-152水平。结果证实在db/db小鼠中与对照组相比miR-152表达显著增加(N=5,图9G和H)。
缺氧、炎症和生长因子使miR-152表达升高
先兆子痫胎盘与缺氧、炎症状况以及VEGF和FGF-2生长因子增多相关联(Hunter et al.,2000;Ozkan et al.,2008),因此通过qPCR确定在缺氧、炎症和VEGF刺激条件下胎盘外植体和内皮细胞中miR-152的表达。在胎盘外植体中,混合的炎症细胞因子刺激使miR-152增加大约55%(图10A)。此外,在胎盘外植体(图10B)和内皮细胞(图10D)中的缺氧处理使miR-152表达分别增加75%和84%。在胎盘外植体中,缺氧和炎症细胞因子混合物的组合刺激使miR-152表达增加87%(图10C)。此外,VEGF和FGF-2刺激使miR-152表达分别增加大约100%和65%(图10E和F)。这些数据证实miR-152在先兆子痫胎盘中受病理学状况控制。
MiR-152调节内皮细胞中P1GF表达
抗血管发生因子sFlt-1和sEng的释放及胎盘生长因子表达的抑制是导致先兆子痫的主要因素(Powe et al.,2011;Ahmad and Ahmed,2004)。此外,VEGF和炎症细胞因子刺激增加sFlt-1和sEng释放(Cudmore et al.,2008),因此,我们检验在内皮细胞中miR-152是否调节sFlt-1、sEng或者P1GF表达。将HUVECs用模拟-152转染以过表达miR-152(图18),通过ELISA检验sFlt-1、sEng和PlGF水平。HUVECs中miR-152的过表达在运载体和VEGF-E刺激下均降低PlGF表达(图10I)。尽管sFlt-1(图10G)和sEng(图10H)的表达不变,但是PlGF水平显著降低,因此sFlt-1/P1GF比率增高。这些数据进一步支持了miR-152正调节可导致先兆子痫发病机制的观点。
MiR-152对细胞增殖无作用
由于内皮功能失调是导致先兆子痫的主要因素,我们检测miR-152是否调节内皮细胞功能。将HUVEC细胞用合成的抑制剂(抗-152)(I)或者miR-152模拟物(模拟-152)(II)转染,进行MTT(图11A)或者细胞增殖测定(图11B)。尽管抗-152在HUVECs中显著增加细胞存活力,但是模拟-15在运载体或VEGF-A刺激下不改变细胞存活力(图11A).此外,细胞增殖测定证实抗—152或者模拟-152对于HUEVC细胞增殖均无影响(图11B).此外,western印迹分析表明在用抗-152或者模拟-152转染的HUVECs细胞中裂解的Caspase-3的表达无变化(图11C)。
MiR-152抑制内皮细胞粘附及体外内皮细胞管形成,但是促进细胞迁移
进一步检测miR-152在内皮细胞粘附、迁移和血管发生中的作用。HUVEC细胞粘附能力在模拟-152转染的细胞中显著降低,与VEGF-A或是VEGF-E刺激无关。细胞计数证实与模拟-con转染的HUVECs对比,在模拟-152转染的HUVECs中在运载体刺激条件下粘附细胞的数目降低84%,而在VEGF-A和VEGF-E刺激条件下降低大约89%(图12A)。
由于miR-152在肿瘤细胞中抑制血管发生,我们推测miR-152在胎盘内皮细胞中也抑制血管发生,导致先兆子痫的发病机制。用模拟-152转染的HUVEC细胞在基质胶上在运载体、VEGF-A或VEGF-E刺激下呈现出降低的管形成能力(图12B)。定量分析表明在模拟-152转染的细胞中在运载体刺激条件下管形成降低37%,在VEGF-A和VEGF-E刺激条件下管形成降低34%-66%。相反,在抗-152转染的HUVEC细胞中在VEGF-A刺激条件下使管形成增加大约33%(图12C)。这些数据证实miR-152负调节胎盘内皮细胞体外血管发生能力。
此外,刮擦测定表明在运载体或生长因子刺激条件下miR-152的过表达增加HUVEC细胞迁移能力(图19A),而miR-152的抑制显著降低HUVEC细胞迁移能力(图19B)。
miR-152的过表达在后肢缺血模型中削弱血液恢复及血管再生
为了进一步检测miR-152在体内的抗血管发生性质,利用小鼠后肢缺血模型。将过表达miR-152的腺病毒注射进随后除去的股动脉中。与非缺血肌肉相比,在第3天所述腓肠肌中miR-152的表达增加大约18倍,在第14天为55%(图13A)。在注射ad-152的小鼠与注射的ad-CMV对照小鼠中对比后肢缺血的血流恢复情况。在HLI手术后(第0、3、7和14天)在足跖面通过激光多普勒连续评定血流量(图13C)。在注射的ad-CMV小鼠中在第3天证实血流恢复,注射的小鼠呈现出在第3天和第7天一致降低的血流量(图13B)。在第7天,在注射ad-152的小鼠中与对照组相比示出大约30%的最大降低(图13C)。由于血流量恢复与增加的组织毛细管密度相关,我们在腓肠肌中检验毛细管密度(图13D)。正如预期的,在注射ad-152和ad-CMV的小鼠的缺血腓肠肌中毛细血管密度均增加;然而在注射ad-152的缺血肌肉中,毛细血管增加明显降低(图13E)。此外,与在体外观测结果一致,在对照小鼠的缺血肌肉中的miR-152表达与对照的非缺血肌肉相比显著增加(图13F)。
miR-152的过表达削弱妊娠小鼠中胎盘血管发生及抑制胎儿生长
为了研究miR-152过表达在妊娠中的作用,将过表达miR-152的腺病毒在E11.5天经腹膜内注射进妊娠小鼠体内。在注射6天后,获取来自注射ad-CMV和ad-152的小鼠的血样,通过qPCR测量miR-152的表达。与注射ad-CMV对照组相比,注射ad-152显著增加miR-152表达(图14A)。与这些数据一致,在注射ad-152的妊娠小鼠中胎儿生长显著受限(图14B)。这个观测结果通过测量这些小鼠中平均胎儿体重可进一步证实。
注射ad-152的小鼠的胎儿与注射ad-CMV的小鼠相比呈现出体重降低大约25%(图14C)。
更重要地,胎儿生长受限可以是由于胎盘中削弱的新血管发生所致,这可以由注射的小鼠中无血管发生(avasculated)的胎盘而证实(图14D)。对胎盘迷路区的组织学分析进一步证实这个观测结果。迷路区由滋养层细胞和胎儿内皮细胞组成,形成大表面积以供母体与胎儿之间的营养和气体交换。使用同工凝集素B4标示胎儿内皮细胞,分析注射ad-152和ad-CMV的小鼠中迷路区的解剖学特点。对照小鼠迷路区中的血管具有高密度胎儿血管分支的良好组构。然而,观测到在注射ad-152的小鼠迷路区中的血管结构是较低密度的不规则分支(图14E)。
MiR-152在内皮细胞中直接靶向ITGA5
为了鉴别miR-152的直接靶位,使用来自模拟-152和模拟-con转染的HUVEC的RNA样品进行基因表达微阵列分析。以±1.25倍截止值选择基于miR-152过表达的14个正调节的和34个负调节的基因。这些表达改变的基因对于血管重建、细胞粘附、血管发生、细胞迁移性和存活力具有广泛功能。此外,使用在线预测工具microRNA.org、PicTar和Target Scan进一步分析负调节的基因以通过序列比对证实可能的miR-152直接靶位。最终,选择34个负调节基因中的18个基因作为miR-152直接靶位的候选者,这些候选靶位的表达在HUVEC细胞中通过western印迹进一步检测(图15B)。
在检测的靶中,ITGA5是最受miR-152调节的一个靶。通过在线预测工具预测ITGA5是miR-152直接靶位(图15A),且通过western印迹也证实,即在HUVECs细胞中miR-152过表达降低ITGA5表达而miR-152抑制增加ITGA5表达(图15B)。MicroRNA靶测定进一步证实模拟-152转染在pmiR-ITGA5转染的HEK293细胞中显著抑制荧光素酶活性,但是在突变pmiR-ITGA5M转染的细胞中则否(图15C,图20)。为了确认miR-152靶向ITGA5以履行使内皮细胞功能失调的作用,我们检测了ITGA5过表达是否可以拯救模拟-152介导的血管发生的抑制。正如预期的,ITGA5的过表达在HUVEC细胞中自发性增加管形成及显著恢复模拟-152抑制的管样结构(图15D)。
MiR-152在体内靶向ITGA5及在重度先兆子痫患者中与ITGA5相关
更重要地,miR-152对ITGA5的调节在本研究使用的动物模型中也经证实。与注射对照的AAV病毒的小鼠肢体肌肉(图16A)相比,注射AAV-152病毒介导的小鼠肢体肌肉中的miR-152过表达(图21)显著降低ITGA5表达。此外,在后肢缺血条件下,注射ad-152的小鼠缺血肌肉中的ITGA5表达与在注射ad-CMV的小鼠缺血肌肉中的表达相比降低(图16B)。此外,注射ad-152的妊娠小鼠胎盘中的ITGA5表达与在注射ad-CMV的胎盘中的表达相比显著降低(图16C)。
由于miR-152在先兆子痫患者中是正调节的以及鉴别ITGA5是miR-152的直接靶位,我们推测在先兆子痫患者中miR-152和ITGA5的表达是负相关的。分别使用Qpcr和western印迹,从8个先兆子痫患者和8个孕龄匹配的对照患者中检验miR-152和ITGA5的表达(图16D)。miR-152和ITGA5表达的负相关性在先兆子痫胎盘中发现(R=-0.823,图16E),但是在对照的血压正常的胎盘中未发现,提示在先兆子痫病理情况下miR-152特异性靶向ITGA5。
讨论:
先兆子痫中MiR-152
尽管微阵列分析及下一代测序技术表明microRNA在先兆子痫妇女胎盘中不同表达(Yang et al.,2011;Wu et al.,2012;Hromadnikova et al.,2012;Pan et al.,2012),但是仅鉴定了极有限数目的microRNAs在先兆子痫发病机制中的作用,特别是在内皮功能失调中的作用。通过微阵列分析示出mir-152表达在先兆子痫患者中是正调节的(Zhu et al.,2009)。
鉴于此,我们的数据证实在更精确限定的平均孕龄为29.7周的重度先兆子痫患者群中miR-152表达升高。此外,我们证实在小鼠和大鼠先兆子痫模型中正调节的miR-152表达,进一步强化了miR-152与先兆子痫发病机制相关的可能性。
先兆子痫胎盘与缺氧和炎症状况相关(Lockwood et al.,2008;Soleymanlou et al.,2005)。我们的数据表明缺氧和炎症细胞因子增加胎盘外植体中miR-152表达,提示miR-152可以是理解先兆子痫发病机制的分子学线索。此外,已经表明血管发生的失衡是导致先兆子痫的主要因素(Ramma and Ahmed,2011;Ramma et al.,2012)。在我们的研究中,鉴别miR-152在内皮细胞中是对于胎盘功能重要的生长因子PlGF的负调节物。尽管sFlt-1和sEng水平随着miR-152过表达未改变,但是sFlt-l/PlGF比率由于P1GF水平降低而增加。由于sFlt-1/PlGF比率升高是血管发生失衡的重要指征且是评估先兆子痫的可靠生物标记(De Vivo et al.,2008;Verlohren et al.,2012),因此在先兆子痫胎盘中miR-152表达升高在先兆子痫发病机制中可导致血管发生功能紊乱。
MiR-152通过ITGA5调节内皮功能
已经示出mir-152抑制细胞增殖和粘附(Zhou et al.,2012;Mancini et al.,2012)以及肿瘤细胞血管发生(Zheng et al.,2013;Xu et al.,2013),但是在内皮细胞中则否。我们的数据表明miR-152降低内皮细胞粘附和血管发生能力,及增强细胞迁移,但是对于细胞增殖无作用。内皮功能紊乱在先兆子痫发病机制中起主要作用且导致先兆子痫的大多数临床症状(Poston 2006;Baumwell and Karumanchi,2007),因此,miR-152可通过调节内皮功能导致先兆子痫的发病机制。
ITGA5属于整联蛋白α链家族,其促进在癌细胞中的细胞粘附、侵润及迁移(Hood and Cheresh,2002;Wang et al.,2008;Qin et al.,2011)。此外,已经示出其促进内皮细胞和小鼠胚胎中血管发生和血管生成(Francis et al.,2002;Bonauer et al.,2009)。在我们的研究中,发现ITGA5是miR-152的直接靶位及也是miR-152诱导的内皮功能紊乱的介导物。通过直接靶向ITGA5,miR-152抑制内皮细胞粘附,降低的细胞粘附随后导致促进的细胞迁移(Moh and Shen,2009;Grzesiak et al.,2005)。更重要地,ITGA5的过表达拯救内皮细胞中模拟-152抑制的血管发生,提示ITGA5是miR-152介导的内皮功能紊乱的至少破坏血管发生的下游效应子。显然,我们的微阵列数据揭示了更可能的miR-152靶位,其调节血管重建、细胞粘附、血管发生、细胞能动性及存活过程。这些潜在的miR-152靶位可进一步导致先兆子痫的不同的临床表现。
MiR-152对于先兆子痫的治疗
由于目前尚无对于先兆子痫的有效治疗方法,因此高危妇女的早期预测对于预防和初级护理这种病症是重要的(Leslie et al.,2011)。此外,最近的Meta分析提示在高危妇女妊娠16周开始给予低剂量阿司匹林可以预防直至50%的PE、重度PE及胎儿宫内生长受限(IUGR)(Bujold et al.,2009)。因此,精确预测先兆子痫的生物标记具有重大临床价值。近年来,miR-152已经涉及妊娠妇女血清,提示其作为生物标记的潜在用途(Williams et al.,2013)。基于我们目前的研究,在重度先兆子痫中miR-152升高,因此应检验先兆子痫高危妇女的早期妊娠血清中的miR-152表达,以验证其作为生物标记的潜力。
先兆子痫是一种多因素疾病,导致世界范围的母亲和胎儿发病率和死亡率(Pennington et al.,2012),因此迫切需要新方法以处理这个复杂的病症。我们的数据表明miR-152在先兆子痫中升高,随后通过靶向ITGA5导致内皮功能紊乱,提示miR-152和ITGA5在先兆子痫中可作为潜在治疗靶位。更重要地,先兆子痫妇女胎盘中的miR-152和ITGA5表达是负相关的,进一步证实miR-152和ITGA5在先兆子痫中的治疗潜力。
miR-195
方法和材料:
试剂和抗体
重组生长因子、血管内皮生长因子A(VEGF-A)、VEGF-E、肿瘤坏死因子α(TNF-α)、干扰素γ(IFN-γ)和白细胞介素1β(IL-1β)购自KELIATech(Brauschweig,Germany)。抗总eNOS和phosphor-eNOS的兔抗体得自Cell Signaling Technology(Danvers,MA 01923,United States)。小鼠抗-β-肌动蛋白抗体得自Sigma-Aldrich(A5441,St.Louis,MO,United States)。针对miR-195的定量PCR引物qScriptTMmicroRNA cDNA及Synthesis Green SuperMix试剂盒购自Quanta Biosciences。低生长因子基质胶购自Becton Dickinson(Oxford,UK)。M199培养基购自Invitrogen(Paisley,UK)。Calcein AM荧光染料购自BD Bioscience(P.O.Box999Sparks,MD,USA21152)。
人胎盘组织收集和制备
伦理委员会学会(Institutional Ethics Committee)许可胎盘组织收集并获得书面知情同意书。所有妇女从注册至分娩均被前瞻性跟踪。从第一孕期(N=6)、第二孕期r(N=6)和第三孕期(N=4)的妊娠妇女中收集不同孕龄的胎盘组织并通过qPCR测量相对miR-195表达水平。从BMI超过30(内脏脂肪N=25,皮下脂肪N=24)或者具有正常BMI(18-25)(内脏脂肪N=22,皮下脂肪N=20)的妊娠妇女中收集人脂肪组织,通过qPCR确定miR-195表达。此外,从并发先兆子痫(N=14)和胎儿子宫内生长受限(IUGR,N=12)的妊娠妇女及从血压正常的妊娠妇女(N=17)中收集人胎盘组织。将收集的胎盘组织进一步用于定量PCR。先兆子痫定义为在至少两次连续测量血压>140/90mm Hg及母体蛋白尿为至少300mg/24小时,IUGR定义为胎儿的估计重量低于其胎龄体重的10%,腹围低于2.5%。
细胞和胎盘外植体培养
如先前所述分离人脐静脉内皮细胞(HUVEC)并在M199培养基中培养(Bussolati et al,2001)。对第三代或第四代HUVEC进行试验。从经历选择性终止妊娠的正常妊娠妇女中获得第一孕期胎盘组织(6-9周孕龄)。如先前所述制备胎盘绒毛组织外植体(Ahmad and Ahmed,2004)。简而言之,将人胎盘绒毛外植体在检测物质刺激下或者取样条件下保温,收集进行miR-195的定量PCR。为了产生缺氧条件,将70-80%铺满融合的HUEVCs或者胎盘外植体在保温仪中1%O2和5%CO2条件下在37℃培养。为了模拟炎症状况,将HUVECs或者胎盘外植体在含有TNF-α(20ng/ml)、IFN-γ(20ng/ml)和IL-1β(2ng/ml)的cyto-mix混合物中刺激。
针对miR-195的miRNA模拟物和抑制剂
针对miR-195的化学合成的双链micro RNA模拟物和单链抑制剂(mi-195和抗-195)购自Qiagen。这些分子的效力使用miR-195的引物通过qPCR检测(图27A)。
如先前所述(Cudmore et al.,2007),将HUVECs胰蛋白酶化,并使用电穿孔技术将1×106个细胞用~0.6μg mi-195、~3μg抗-195或者等价对照分子电穿孔(Amaxa GmbH,Cologne,Germany)。
针对miR-195的腺病毒
过表达eNOS组成型激活形式EnosS1177D的腺病毒ad-eNOSS1177D友情得自Prof.Ingrid Fleming(Johann Wolfgang Goethe University,Germany)。过表达miR-195的腺病毒ad-195购自Vector Biolabs(Philadelphia,PA,USA),其效力使用qPCR检测(图27B)。
Ad-sFlt-1诱导的小鼠先兆子痫模型
腺病毒ad-sFlt-1友情得自Prof.Richard Mulligan(Harvard Medical School,Boston,USA)。为妊娠的C57BL/6小鼠在E9.5天经尾静脉注射109PFU腺病毒ad-CMV或者ad-sFlt-1。在E17.5天使用针对Flt-1的ELISA测量循环血液中sFlt-1表达水平并在颈动脉评估平均动脉压(MAP)(图26)。收集来自注射ad-CMV对照病毒(N=7)和ad-sFlt-1病毒(N=5)的小鼠的胎盘组织并使用qPCR分析miR-195表达水平。
降低子宫灌注压(RUPP)的先兆子痫大鼠模型
RUPP先兆子痫模型的胎盘样品友情得自Dr.Fergus McCarthy(Cork University Maternity Hospital,Wilton,Cork,Ireland)。经历RUPP手术的大鼠的实验程序和鉴定如先前所述(McCarthy et al.,2011)。
实时聚合酶链反应
如先前所述进行样品制备和实时PCR(Cudmore et al.,2007)。miR-195的逆转录试剂盒和引物购自Quanta Biosciences。
酶联免疫吸附测定
人PlGF的酶联免疫吸附测定(ELISA)得自R&D Systems并根据厂商说明书进行测定。
Western印迹
如先前所述,将来自HUVEC或动物组织的总蛋白质在RIPA缓冲液中裂解并分析(Ahmad and Ahmed,2004)。
体外细胞存活力和增殖分析
将HUVEC细胞用抗-195或mi-195电穿孔。在过夜恢复后,将转染的细胞胰蛋白酶化并以1×104/孔密度铺板于96孔平板中,用运载体或VEGF-A(20ng/ml)处理。48小时后,使用细胞生长确定试剂盒(Cat No.CDGl,Sigma-Aldrich)将这些细胞进行MTT测定。对于细胞增殖测定,将转染的细胞以4×104/孔密度铺板于24孔平板中,用VEGF-A(20ng/ml)或VGEF-E(20ng/ml)刺激。48小时后,将细胞胰蛋白酶化,使用血细胞计数器在显微镜下计数每孔细胞数。
体外细胞粘附测定
将HUVEC用mi-195或者对照mi-con电穿孔。48小时后,将2×104mi-195或者mi-con转染的细胞铺板于2%明胶包被的96孔平板中,用VEGF-A(20ng/ml)或者VEGF-E(20ng/ml)处理30分钟。之后,将细胞用PBS洗涤3次,用Calcein AM荧光染料染色并进行荧光显微镜检测。计数在4×放大倍率下每个视野的粘附细胞数目。
体外管形成测定
如先前所述在低生长因子基质胶上确定用mi-195或抗-195转染的或者或者mi-195与ad-EnosS1177D共转染的HUVEC的毛细血管样结构的形成(Bussolati et al,2001)。
体外细胞迁移测定
将HUEVC细胞用抗-195或mi-195电穿孔。在过夜恢复后,在12或6孔平板中转染的铺满融合细胞中心进行刮擦。在进行刮擦后0小时和16-24小时立即测量刮擦的宽度。通过减去在0小时和16-24小时的刮擦宽度计算细胞迁移距离。
NO释放测定
如先前所述(Ahmad et al.,2006),使用Sievers NO化学发光分析仪(Analytix,Sunderland,UK)测定条件培养基中以NO的稳定分解产物亚硝酸盐形式的总NO。简而言之,将HUVECs用ad-con或ad-195感染,MOI=100,收集条件培养基进行NO测量。
MicroRNA靶荧光素酶分析
含有eNOS未翻译区(UTR)的人cDNA购自Origene(Rockville,MD20850,USA),将UTR区亚克隆进pMirTarget产生eNOS-UTR-荧光素酶(萤火虫)表达质粒pmiR-eNOS。内皮细胞中pmiR-eNOS的成功转染通过红色荧光蛋白(RFP)的表达证实(图28)。使用定向诱变试剂盒(Stratagene)及含有突变位点的PCR引物产生携带miR-195结合位点的eNOS UTR突变形式的质粒,所得质粒称作pmiR-Enosm。用于PCR扩增的引物是:mNOS3-F(CTCTCAGGAGTAGAGTACCTGTAAAGGAGAATCTCTAAATCAAGT)和mNOS3-R(ACTTGATTTAGAGATTCTCCTTT AC AGGT ACTCTACTCCTGAGAG)。
对于microRNA靶测定,将HEK293细胞用pmiR-eNOS或pmiR-eNOSM与mi-con或mi-195一起转染。将含有海肾荧光素酶的质粒也进行转染作为内部对照。在保温过夜后,测量相对的萤火虫荧光素酶活性并标准化为海肾荧光素酶活性,根据Dual-Reporter Assay System(E1910,Promega)的厂商指导方案进行。
MicroRNA靶预测
使用在线程序预测miR-195的直接靶位:microRNA.org(http://www.microrna.org/microrna/home.do)、PicTar(http://pictar.mdc-beriin.de/)及Target Scan(http://www.targetscaii.org/)、RNAhybrid 2.2和RNA22 microRNA靶预测工具
微阵列分析
将HUVEC细胞用mi-195转染,收集总RNA样品,使用Affymetrix genechip Human Gene 1.0ST进行全基因组基因阵列分析。使用±1.25倍截止值选择基于mi-195转染的正调节和负调节的基因。
统计学分析
所有数据均以平均值+S.E.M表示。使用Student's t检验或者Mann-Whitney U检验进行统计学对比。统计学意义设定为p<0.05。
结果:
miR-195表达在重度先兆子痫患者和动物模型、IUGR、妊娠肥胖妇女中升高
各种微阵列分析表明miR-195表达在先兆子痫妇女中是异常的Xu et al.,2014;Zhu et al.,2009;Hu et al.,2009)。因此,我们检验了miR-195在与先兆子痫发病机制相关的不同人体组织中的表达模式。首先,通过qPCR确定不同孕龄的妊娠妇女胎盘中的miR-195表达。在第三孕期胎盘中miR-195表达与在第一孕期(N=6)和第二孕期d(N=6)胎盘的表达中相比分别正调节12.5倍和8.9倍(图22A)。此外,由于肥胖妊娠妇女具有先兆子痫高危性,我们检验了肥胖妊娠妇女的脂肪组织中的miR-195表达。与偏瘦的妊娠妇女(N=20-22)相比,在肥胖妊娠妇女的内脏脂肪(N=25)和皮下脂肪(N=24)中的miR-195表达分别增加2.28倍和7.8倍(图22B和C)。尽管各种微阵列研究表明先兆子痫中miR-195异常表达,但是这些研究的结果是相矛盾的。更重要地,这些研究中使用的胎盘样品是从34.7-35.9周的晚期孕龄先兆子痫妇女中收集的。为了研究在更精确定义的早期重度先兆子痫患者中的miR-195表达,收集平均孕龄为29.7周的先兆子痫妇女的胎盘样品,通过qPCR检验miR-195表达水平。定量PCR表明与孕龄匹配的对照的血压正常的胎盘(N=17)相比在先兆子痫胎盘(N=14)和IUGR胎盘(N=12)中的miR-195表达分别增加10.9倍和2.6倍(图22D)。
为了进一步证实在先兆子痫中miR-195的正调节,在两个先兆子痫动物模型中检验miR-195的表达。接受ad-sFlt-1腺病毒注射的妊娠小鼠(N=5)与ad-CMV对照病毒注射的小鼠(N=7)相比呈现出显著增加的胎盘miR-195水平(图22E)以及升高的sFlt-1表达及平均动脉压(图26)。此外,经历降低子宫灌注压手术的妊娠Sprague Dawley大鼠(N=17)胎盘中的miR-195表达与正常妊娠大鼠(N=12,图22F)相比也增加7.7倍。总之,这些数据提示miR-195升高与先兆子痫的发病机制相关。
缺氧和炎症细胞因子增加miR-195表达
先兆子痫胎盘与缺氧、炎症以及VEGF和FGF-2生长因子升高相关(Hunter et al.,2000;Ozkan et al.,2008),因此通过qPCR确定在缺氧和炎症刺激下在内皮细胞和胎盘外植体中的miR-195表达。在HUVEC细胞中混合的炎症细胞因子的刺激使miR-195表达增加大约43%(图22G)。此外,缺氧与炎症细胞因子混合物的组合刺激在HUVEC细胞和胎盘外植体中使miR-195表达分别增加138%和48%(图22H和I)。这些数据证实miR-195表达受与先兆子痫胎盘相关的病理状况的调节。
MiR-195在内皮细胞中调节PlGF表达
抗血管发生因子sFlt-1和sEng的释放及胎盘生长因子(PlGF)表达的抑制是导致先兆子痫的主要因素(Powe et al.,2011;Ahmad and Ahmed,2004),因此我们检验了miR-195在内皮细胞中是否调节sFlt-1、sEng或者PlGF表达。将HUVEC用mi-195转染以过表达miR-195(图27),sFlt-1、sEng和PlGF释放通过ELISA检验。尽管在运载体和VEGF刺激条件下,HUVEC中miR-195的过表达不改变sFlt-1和sEng表达,但是PlGF水平显著降低(图23A)。结果,sFlt-l/PlGF的比率增加。这些数据进一步支持了miR-195正调节可导致先兆子痫发病机制的观点。
MiR-195抑制内皮细胞增殖、存活力、粘附和体外管形成,但是促进细胞迁移
由于削弱的内皮是导致先兆子痫的主要因素,因此我们检测了miR-195是否调节内皮细胞功能。将HUVEC用合成的模拟物(mi-195)转染,并进行细胞增殖测定。与对照组相比在运载体、VEGF-A或VEGF-E刺激下,miR-195过表达使细胞数目减少35%-62%(图23B)。此外,在用mi-195转染的HUVEC中进行的MTT测定进一步证实miR-195过表达在运载体或者VEGF刺激下使细胞存活力分别降低35%或31%(图23C)。
也检测了miR-195过表达在内皮细胞粘附中的作用。粘附能力在mi-195转染的细胞中与VEGF-A或VEGF-E刺激无关地显著降低。细胞计数证实与在转染的细胞中相比,在mi-195转染的HUVEC中在运载体、VEGF-A或VEGF-E刺激下粘附细胞减少大约70%(图23D)。
由于miR-195在肿瘤细胞中抑制血管发生,我们推测miR-195还抑制导致先兆子痫发病机制的胎盘内皮细胞中血管发生。
用mi-195转染的HUVEC呈现出在运载体、VEGF-A或VEGF-E刺激下在基质胶上降低的管形成(图23E)。定量分析表明在mi-195转染的细胞中在运载体刺激条件下管形成降低40%,在VEGF-A或VEGF-E刺激条件下管形成降低40%-84%。
此外,通过刮擦测定确定miR-195在内皮细胞迁移中的作用。用抗-195转染的HUVEC与抗-con转染的对照细胞相比示出在运载体或VGEF刺激条件下迁移能力降低11%-26%(图24A)。相反,HUVEC中miR-195的过表达在运载体或VEGF-A刺激条件下使细胞迁移能力显著增加大约1.25倍(图24B)。
MiR-195在内皮细胞中直接靶向eNOS
为了鉴别miR-195的直接靶位,使用来自mi-195和mi-con转染的HUVEC细胞的RNA样品进行基因表达微阵列分析。以±1.25倍截止值基于miR-152过表达选择10个正调节的基因和12个负调节的基因。这些表达改变的基因对于血管重建、细胞粘附、迁移性和血管发生具有广泛的功能。此外,使用在线预测工具进一步分析负调节的基因以通过序列比对证实miR-195的可能的直接靶位。最终,选择4个负调节的基因作为miR-195直接靶位的候选者。
在这些直接靶位中,eNOS是最重要的且在文献中经充分鉴定。首先,通过在线预测工具在各种动物物种中预测eNOS是miR-195直接靶位(图25A)。进一步地,microRNA靶测定证实mi-195的pmiR-eNOS转染在转染的HEK293细胞中显著荧光素酶活性,但是在eNOS UTR突变质粒pmiR-eNOSM转染的细胞中则否(图25B;图28)。荧光素酶测定数据通过eNOS mRNA和蛋白质水平在mi-195转染的HUVEC中降低这一证据而得以进一步支持。定量PCR表明在mi-195转染的HUVEC中降低22%(图25C),使用总eNOS和phosphor-eNOS的抗体的western印迹分析表明在mi-195转染的细胞中总eNOS降低大约55%及phosphor-eNOS降低24%-56%(图25D和E)。更重要地,用mi-195转染的HUVEC中NO的释放与在mi-con转染的细胞中相比降低27%(图25F)。最终,为了证实eNOS是miR-195介导的内皮功能失调的下游效应子,我们检测了eNOS过表达是否可拯救mi-195介导的血管发生抑制作用。正如预期的,在HUVEC细胞中,miR-195过表达降低自发性管形成,而eNOSS1177D过表达完全恢复mi-195介导的管样结构的抑制作用(图25G和H)。
参考文献:
Ahmad S,Ahmed A.Elevated placental soluble vascular endothelial growth factor receptor- 1 inhibits angiogenesis in preeclampsia.Circ Res.2004;95:884-891.
Bussolati B,Dunk C,Grohman M,Kontos CD,Mason J,Ahmed A.Vascular endothelial growth factor receptor- 1 modulates vascular endothelial growth factor-mediated angiogenesis via nitric oxide.Am J Pathol 2001;159:993-1008.
Sibai B,Dekker G,Kupferminc M.Pre-eclampsia.Lancet.2005;365:785-799.
Steegers EA,von Dadelszen P,Duvekot JJ,Pijnenborg R.Pre-eclampsia.Lancet.2010;376:631-644.
Hladunewich M,Karumanchi SA,Lafayette R.Pathophysiology of the clinical manifestations of preeclampsia.Clin J Am Soc Nephrol.2007;2:543-549.
Silasi M,Cohen B,Karumanchi SA,Rana S.Abnormal placentation,angiogenic factors,and the pathogenesis of preeclampsia.Obstet Gynecol Clin North Am.2010;37:239-253.
Maynard S,Epstein FH,Karumanchi SA.Preeclampsia and angiogenic imbalance.Annu Rev Med.2008;59:61-78.
Wang A,Rana S,Karumanchi SA.Preeclampsia:the role of angiogenic factors in its pathogenesis.Physiology(Bethesda).2009;24:147-158.
Ramma W,Ahmed A.Is inflammation the cause of pre-eclampsia?Biochem Soc Trans.2011;39:1619-1627.
Ramma W,Buhimschi IA,Zhao G,Dulay AT,Nayeri UA,Buhimschi CS,Ahmed A.The elevation in circulating抗-angiogenic factors is independent of markers of neutrophil activation in preeclampsia.Angiogenesis.2012;15:341-348.
Lockwood CJ,Yen CF,Basar M,Kayisli UA,Martel M,Buhimschi I,Buhimschi C,Huang SJ,Krikun G,Schatz F.Preeclampsia -related inflammatory cytokines regulate interleukin-6 expression in human decidual cells.Am J Pathol.2008;172:1571-1579.
Powe CE,Levine RJ,Karumanchi SA.Preeclampsia,a disease of the maternal endothelium:the role of antiangiogenic factors and implications for later cardiovascular disease.Circulation.2011;123:2856-2869.
Poston L.Endothelial dysfunction in pre-eclampsia.Pharmacol Rep.2006;58Suppl:69-74.
Soleymanlou N,Jurisica I,Nevo O,Ietta F,Zhang X,Zamudio S,Post M,Caniggia I.Molecular evidence of placental hypoxia in preeclampsia./Clin Endocrinol Metab.2005;90:4299-4308.
Grill S,Rusterholz C,Zanetti-Dallenbach R,Tercanli S,Holzgreve W,Hahn S,Lapaire O.Potential markers of preeclampsia-a review.Reprod Biol Endocrinol.2009;7:70-83.
Bartel DP.MicroRNAs:target recognition and regulatory functions.Cell.2009;136:215-233.
Lewis BP,Shih IH,Jones-Rhoades MW,Bartel DP,Burge CB.Prediction of mammalian microRNA targets.Cell.2003;115:787-798.
Williams AE.Functional aspects of animal microRNAs.Cell Mol Life Sci.2008;65:545-562.
He L,Hannon GJ.MicroRNAs:small RNAs with a big role in gene regulation.Nat Rev Genet.2004;5:522-531.
Zhu XM,Han T,Sargent IL,Yin GW,Yao YQ.Differential expression profile of microRNAs in human placentas from preeclamptic pregnancies vs normal pregnancies.Am J Obstet Gynecol.2009;200:661.el-7.
Hu Y,Li P,Hao S,Liu L,Zhao J,Hou Y.Differential expression of microRNAs in the placentae of Chinese patients with severe pre-eclampsia.Clin Chem Lab Med.2009;47:923-929.
Pineles BL,Romero R,Montenegro D,Tarca AL,Han YM,Kim YM,Draghici S,Espinoza J,Kusanovic JP,Mittal P,Hassan SS,Kim CJ.Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia.Am J Obstet Gynecol.2007;196:261.el-6.
Mayor-Lynn K,Toloubeydokhti T,Cruz AC,Chegini N.Expression profile of microRNAs and mRNAs in human placentas from pregnancies complicated by preeclampsia and preterm labor.Reprod Sci.2011;18:46-56.
Enquobahrie DA,Abetew DF,Sorensen TK,Willoughby D,Chidambaram K,Williams MA.Placental microRNA expression in pregnancies complicated by preeclampsia.Am J Obstet Gynecol.2011;204:178.el2-21.
Noack F,Ribbat-Idel J,Thorns C,Chiriac A,Axt-Fliedner R,Diedrich K,Feller AC.miRNA expression profiling in formalin-fixed and paraffin-embedded placental tissue samples from pregnancies with severe preeclampsia.J Perinat Med.2011;39:267-271.
Zhang Y,Diao Z,Su L,Sun H,Li R,Cui H,Hu Y.MicroRNA- 155 contributes to preeclampsia by down-regulating CYR61.Am J Obstet Gynecol.2010;202:466.el-7.
Cheng W,Liu T,Jiang F,Liu C,Zhao X,Gao Y,Wang H,Liu Z.microRNA- 155 regulates angiotensin II type 1 receptor expression in umbilical vein endothelial cells from severely pre-eclamptic pregnant women.Int J Mol Med.2011;27:393-399.
Zhang Y,Fei M,Xue G,Zhou Q,Jia Y,Li L,Xin H,Sun S.Elevated levels of hypoxia-inducible microRNA-210 in pre-eclampsia:new insights into molecular mechanisms for the disease./Cell Mol Med.2012;16:249-259.
Dai Y,Diao Z,Sun H,Li R,Qiu Z,Hu Y.MicroRNA-155 is involved in the remodelling of human-trophoblast-derived HTR-8/SVneo cells induced by lipopoly saccharides.Hum Reprod.2011;26:1882-1891.
Lee DC,Romero R,Kim JS,Tarca AL,Montenegro D,Pineles BL,Kim E,Lee J,Kim SY,Draghici S,Mittal P,Kusanovic JP,Chaiworapongsa T,Hassan SS,Kim CJ.miR-210 targets iron-sulfur cluster scaffold homologue in human trophoblast cell lines:siderosis of interstitial trophoblasts as a novel pathology of preterm preeclampsia and small-for-gestational-age pregnancies.Am J Pathol.2011;179:590-602.
Dai Y,Qiu Z,Diao Z,Shen L,Xue P,Sun H,Hu Y.MicroRNA-155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin Dl.Placenta.2012;33:824-829.
Wang Y,Fan H,Zhao G,Liu D,Du L,Wang Z,Hu Y,Hou Y.miR-16 inhibits the proliferation and angiogenesis-regulating potential of mesenchymal stem cells in severe preeclampsia.FEBS J.2012;279:4510-4524.
Muralimanoharan S,Maloyan A,Mele J,Guo C,Myatt LG,Myatt L.MIR-210modulates mitochondrial respiration in placenta with preeclampsia.Placenta.2012;33:816-823.
Li P,Guo W,Du L,Zhao J,Wang Y,Liu L,Hu Y,Hou Y.microRNA-29b contributes to pre-eclampsia through its effects on apoptosis,invasion and angiogenesis of trophoblast cells.Clin Sci(Lond).2013;124:27-40.
Liu L,Wang Y,Fan H,Zhao X,Liu D,Hu Y,Kidd AR 3rd,Bao J,Hou Y.MicroRNA-181a regulates local immune balance by inhibiting proliferation and immunosuppressive properties of mesenchymal stem cells.Stem Cells.2012;30:1756-1770.
Luo L,Ye G,Nadeem L,Fu G,Yang BB,Honarparvar E,Dunk C,Lye S,Peng C.MicroRNA-378a-5p promotes trophoblast cell survival,migration and invasion by targeting Nodal.J Cell Sci.2012;125:3124-3132.
Xiang Y,Ma N,Wang D,Zhang Y,Zhou J,Wu G,Zhao R,Huang H,Wang X,Qiao Y,Li F,Han D,Wang L,Zhang G,Gao X.MiR-152和miR-185 co-contribute to ovarian cancer cells cisplatin sensitivity by targeting DNMT1directly:a novel epigenetic therapy independent of decitabine.Oncogene.2013;doi:10.1038/onc.2012.575.
Ji W,Yang L,Yuan J,Yang L,Zhang M,Qi D,Duan X,Xuan A,Zhang W,Lu J,Zhuang Z,Zeng G.MicroRNA-152 targets DNA methyltransferase 1 in NiS -transformed cells via a feedback mechanism.Carcinogenesis.2013;34:446-453.
Huang J,Wang Y,Guo Y,Sun S.Down-regulated microRNA-152 induces aberrant DNA methylation in hepatitis B virus-related hepatocellular carcinoma by targeting DNA methyltransferase 1.Hepatology.2010;52:60-70.
Braconi C,Huang N,Patel T.MicroRNA-dependent regulation of DNA methyltransferase- 1 and tumor suppressor gene expression by interleukin-6 in human malignant cholangiocytes.Hepatology.2010;51:881-890.
Zhou X,Zhao F,Wang ZN,Song YX,Chang H,Chiang Y,Xu HM.Altered expression of miR-152和miR-148a in ovarian cancer is related to cell proliferation.Oncol Rep.2012;27:447-454.
Mancini M,Saintigny G,Mahe C,Annicchiarico-Petruzzelli M,Melino G,Candi E.MicroRNA-152 and-181a participate in human dermal fibroblasts senescence acting on cell adhesion and remodeling of the extra-cellular matrix.Aging(Albany NY).2012;4:843-853.
Zhu XM,Han T,Wang XH,Li YH,Yang HG,Luo YN,Yin GW,Yao YQ.Overexpression of miR-152leads to reduced expression of human leukocyte antigen-G and increased natural killer cell mediated cytolysis in JEG-3 cells.Am J Obstet Gynecol.2010;202:592.el-7.
Hiroki E,Akahira J,Suzuki F,Nagase S,Ito K,Suzuki T,Sasano H,Yaegashi N.Changes in microRNA expression levels correlate with clinicopathological features and prognoses in endometrial serous adenocarcinomas.Cancer Sci.2010;101:241-249.
Tsuruta T,Kozaki K,Uesugi A,Furuta M,Hirasawa A,Imoto I,Susumu N,Aoki D,Inazawa J.miR-152 is a tumor suppressor microRNA that is silenced by DNA hypermethylation in endometrial cancer.Cancer Res.2011;71:6450-6462.
Tsuruta T,Kozaki K,Uesugi A,Furuta M,Hirasawa A,Imoto I,Susumu N,Aoki D,Inazawa J.miR-152 is a tumor suppressor microRNA that is silenced by DNA hypermethylation in endometrial cancer.Cancer Res.2011;71:6450-6462.
Chen Y,Song Y,Wang Z,Yue Z,Xu H,Xing C,Liu Z.Altered expression of MiR-148a和miR-152 in gastrointestinal cancers and its clinical significance./Gastrointest Surg.2010;14:1170-1179.
Wang XY,Wu MH,Liu F,Li Y,Li N,Li GY,Shen SR.Differential miRNA expression and their target genes between NGX6-positive and negative colon cancer cells.Mol Cell Biochem.2010;345:283-290.
Stumpel DJ,Schotte D,Lange-Turenhout EA,Schneider P,Seslija L,de Menezes RX,Marquez VE,Pieters R,den Boer ML,Stam RW.Hypermethylation of specific microRNA genes in MLL-rearranged infant acute lymphoblastic leukemia:major matters at a micro scale.Leukemia.2011;25:429-439.
Kitano K,Watanabe K,Emoto N,Kage H,Hamano E,Nagase T,Sano A,Murakawa T,Nakajima J,Goto A,Fukayama M,Yatomi Y,Ohishi N,Takai D.CpG island methylation of microRNAs is associated with tumor size and recurrence of non-small-cell lung cancer.Cancer Sci.2011;102:2126-2131.
Zheng X,Chopp M,Lu Y,Buller B,Jiang F.MiR-15b和miR-152 reduce glioma cell invasion and angiogenesis via NRP-2 and MMP-3.Cancer Lett.2013;329:146-154.
Xu Q,Jiang Y,Yin Y,Li Q,He J,Jing Y,Qi YT,Xu Q,Li W,Lu B,Peiper SS,Jiang BH,Liu LZ.A regulatory circuit of miR-148a/152 and DNMTl in modulating cell transformation and tumor angiogenesis through IGF-IR and IRS 1.J Mol Cell Biol.2013;5:3-13.
Manaster I,Goldman-Wohl D,Greenfield C,Nachmani D,Tsukerman P,Hamani Y,Yagel S,Mandelboim O.MiRNA-mediated control of HLA-G expression and function.PLoS One.2012;7:e33395.
Cudmore M,Ahmad S,Al-Ani B,Fujisawa T,Coxall H,Chudasama K,Devey LR,Wigmore SJ,Abbas A,Hewett PW,Ahmed A.Negative regulation of soluble Flt-1 and soluble endoglin release by heme oxygenase-1.Circulation.2007;115:1789-1797.
McCarthy FP,Drewlo S,Kingdom J,Johns EJ,Walsh SK,Kenny LC.Peroxisome proliferator-activated receptor-γas a potential therapeutic target in the treatment of preeclampsia.Hypertension.2011;58:280-286.
Yang Q,Lu J,Wang S,Li H,Ge Q,Lu Z.Application of next-generation sequencing technology to profile the circulating microRNAs in the serum of preeclampsia versus normal pregnant women.Clin Chim Acta.2011;412:2167-2173.
Wu L,Zhou H,Lin H,Qi J,Zhu C,Gao Z,Wang H.Circulating microRNAs are elevated in plasma from severe preeclamptic pregnancies.Reproduction.2012;143:389-397.
Hromadnikova I,Kotlabova K,Doucha J,Dlouha K,Krofta L.Absolute and relative quantification of placenta- specific micrornas in maternal circulation with placental insufficiency-related complications.J Mol Diagn.2012;14:160-167.
Pan M,Ge Q,Li H,Yang Q,Lu J,Zhang D,Lu Z.Sequencing the miRNAs in maternal plasma from women before and after parturition./Nanosci Nanotechnol.2012;12:4035-4043.
De Vivo A,Baviera G,Giordano D,Todarello G,Corrado F,D'anna R.Endoglin,P1GF and sFlt-1 as markers for predicting pre-eclampsia.Acta Obstet Gynecol Scand.2008;87:837-842.
Verlohren S,Herraiz I,Lapaire O,Schlembach D,Moertl M,Zeisler H,Calda P,Holzgreve W,Galindo A,Engels T,Denk B,Stepan H.The sFlt-l/PlGF ratio in different types of hypertensive pregnancy disorders and its prognostic potential in preeclamptic patients.Am J Obstet Gynecol.2012;206:58.el-8.
Baumwell S,Karumanchi SA.Pre-eclampsia:clinical manifestations and molecular mechanisms.Nephron Clin Pract.2007;106x72-81.
Hood JD,Cheresh DA.Role of integrins in cell invasion and migration.Nat Rev Cancer.2002;2:91-100.
Wang QY,Zhang Y,Shen ZH,Chen HL.alpha 1,3 fucosyltransferase-VII up-regulates the mPvNA of alpha5 integrin and its biological function./Cell Biochem.2008;104:2078-2090.
Qin L,Chen X,Wu Y,Feng Z,He T,Wang L,Liao L,Xu J.Steroid receptor coactivator-1 upregulates integrin as expression to promote breast cancer cell adhesion and migration.Cancer Res.2011;71:1742-1751.
Francis SE,Goh KL,Hodivala-Dilke K,Bader BL,Stark M,Davidson D,Hynes RO.Central roles of alpha5betal integrin and fibronectin in vascular development in mouse embryos and embryoid bodies.Arterioscler Thromb Vase Biol.2002;22:927-933.
Bonauer A,Carmona G,Iwasaki M,Mione M,Koyanagi M,Fischer A,Burchfield J,Fox H,Doebele C,Ohtani K,Chavakis E,Potente M,Tjwa M,Urbich C,Zeiher AM,Dimmeler S.MicroRNA-92a controls angiogenesis and functional recovery of ischemic tissues in mice.Science.2009;324:1710-1713.
Moh MC,Shen S.The roles of cell adhesion molecules in tumor suppression and cell migration:a new paradox.Cell Adh Migr.2009;3:334-336.
Grzesiak JJ,Smith KC,Chalberg C,Burton DW,Deftos LJ,Bouvet M.Type I collagen and divalent cation shifts disrupt cell-cell adhesion,increase migration,and decrease PTHrP,IL-6,and IL-8 expression in pancreatic cancer cells.Int J Gastrointest Cancer.2005;36:131-146.
Leslie K,Thilaganathan B,Papageorghiou A.Early prediction and prevention of preeclampsia.Best Pract Res Clin Obstet Gynaecol.2011;25:343-354.
Bujold E,Morency AM,Roberge S,Lacasse Y,Forest JC,Giguere Y.Acetylsalicylic acid for the prevention of preeclampsia and intra-uterine growth restriction in women with abnormal uterine artery Doppler:a systematic review and meta-analysis./Obstet Gynaecol Can.2009;31:818-826.
Williams Z,Ben-Dov IZ,Elias R,Mihailovic A,Brown M,Rosenwaks Z,Tuschl T.Comprehensive profiling of circulating microRNA via small RNA sequencing of cDNA libraries reveals biomarker potential and limitations.Proc Natl Acad Sci U S A.2013;110:4255-4260.
Pennington KA,Schlitt JM,Jackson DL,Schulz LC,Schust DJ.Preeclampsia:multiple approaches for a multifactorial disease.Dis Model Mech.2012;5:9-18.
Murdoch CE,Shuler M,Haeussler DJ,Kikuchi R,Bearelly P,Han J,Watanabe Y,Fuster JJ,Walsh K,Ho YS,Bachschmid MM,Cohen RA,Matsui R.Glutaredoxin-1 up-regulation induces soluble vascular endothelial growth factor receptor 1,attenuating post-ischemia limb revascularization./Biol Chem.2014;289:8633-44.
Hunter Al,Aitkenhead M,Caldwell C,McCracken G,Wilson D,McClure N.
Serum levels of vascular endothelial growth factor in preeclamptic and normotensive pregnancy.Hypertension.2000;36:965-969.
Ozkan S I,Vural B,Filiz S,Co§tur P,Dalcik H.Placental expression of insulin-like growth factor-I,fibroblast growth factor-basic,and neural cell adhesion molecule in preeclampsia./Matern Fetal Neonatal Med.2008;21:831-838.
参考文献:
Sibai B,Dekker G,Kupferminc M.Pre-eclampsia.Lancet.2005;365:785-799.
Steegers EA,von Dadelszen P,Duvekot JJ,Pijnenborg R.Pre-eclampsia.Lancet.2010;376:631-644.
Hladunewich M,Karumanchi SA,Lafayette R.Pathophysiology of the clinical manifestations of preeclampsia.Clin J Am Soc Nephrol.2007;2:543-549.
Silasi M,Cohen B,Karumanchi SA,Rana S.Abnormal placentation,angiogenic factors,and the pathogenesis of preeclampsia.Obstet Gynecol Clin North Am.2010;37:239-253.
Maynard S,Epstein FH,Karumanchi SA.Preeclampsia and angiogenic imbalance.Annu Rev Med.2008;59:61-78.
Wang A,Rana S,Karumanchi SA.Preeclampsia:the role of angiogenic factors in its pathogenesis.Physiology(Bethesda).2009;24:147-158.
Ramma W,Ahmed A.Is inflammation the cause of pre-eclampsia?Biochem Soc Trans.2011;39:1619-1627.
Ramma W,Buhimschi IA,Zhao G,Dulay AT,Nayeri UA,Buhimschi CS,Ahmed A.The elevation in circulating抗-angiogenic factors is independent of markers of neutrophil activation in preeclampsia.Angiogenesis.2012;15:341-348.
Lockwood CJ,Yen CF,Basar M,Kayisli UA,Martel M,Buhimschi I,Buhimschi C,Huang SJ,Krikun G,Schatz F.Preeclampsia-related inflammatory cytokines regulate interleukin-6 expression in human decidual cells.Am J Pathol.2008;172:1571-1579.
Powe CE,Levine RJ,Karumanchi SA.Preeclampsia,a disease of the maternal endothelium:the role of antiangiogenic factors and implications for later cardiovascular disease.Circulation.2011;123:2856-2869.
Poston L.Endothelial dysfunction in pre-eclampsia.Pharmacol Rep.2006;58 Suppl:69-74.
Soleymanlou N,Jurisica I,Nevo O,Ietta F,Zhang X,Zamudio S,Post M,Caniggia I.Molecular evidence of placental hypoxia in preeclampsia./Clin Endocrinol Metab.2005;90:4299-4308.
Grill S,Rusterholz C,Zanetti-Dallenbach R,Tercanli S,Holzgreve W,Hahn S,Lapaire O.Potential markers of preeclampsia-a review.Reprod Biol Endocrinol.2009;7:70-83.
Bartel DP.MicroRNAs:target recognition and regulatory functions.Cell.2009;136:215-233.
Lewis BP,Shih IH,Jones-Rhoades MW,Bartel DP,Burge CB.Prediction of mammalian microRNA targets.Cell.2003;115:787-798.
Williams AE.Functional aspects of animal microRNAs.Cell Mol Life Sci.2008;65:545-562.
He L,Hannon GJ.MicroRNAs:small RNAs with a big role in gene regulation.Nat Rev Genet.2004;5:522-531.
Hu Y,Li P,Hao S,Liu L,Zhao J,Hou Y.Differential expression of microRNAs in the placentae of Chinese patients with severe pre-eclampsia.Clin Chem Lab Med.2009;47:923-929.
Pineles BL,Romero R,Montenegro D,Tarca AL,Han YM,Kim YM,Draghici S,Espinoza J,Kusanovic JP,Mittal P,Hassan SS,Kim CJ.Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia.Am J Obstet Gynecol.2007;196:261.el-6.
Mayor-Lynn K,Toloubeydokhti T,Cruz AC,Chegini N.Expression profile of microRNAs and mRNAs in human placentas from pregnancies complicated by preeclampsia and preterm labor.Reprod Sci.2011;18:46-56.
Enquobahrie DA,Abetew DF,Sorensen TK,Willoughby D,Chidambaram K,Williams MA.Placental microRNA expression in pregnancies complicated by preeclampsia.Am J Obstet Gynecol.2011;204:178.el2-21.
Noack F,Ribbat-Idel J,Thorns C,Chiriac A,Axt-Fliedner R,Diedrich K,Feller AC.miRNA expression profiling in formalin-fixed and paraffin-embedded placental tissue samples from pregnancies with severe preeclampsia.J Perinat Med.2011;39:267-271.
Zhang Y,Diao Z,Su L,Sun H,Li R,Cui H,Hu Y.MicroRNA- 155 contributes to preeclampsia by down-regulating CYR61.Am J Obstet Gynecol.2010;202:466.el-7.
Cheng W,Liu T,Jiang F,Liu C,Zhao X,Gao Y,Wang H,Liu Z.microRNA- 155 regulates angiotensin II type 1 receptor expression in umbilical vein endothelial cells from severely pre-eclamptic pregnant women.Int J Mol Med.2011;27:393-399.
Zhang Y,Fei M,Xue G,Zhou Q,Jia Y,Li L,Xin H,Sun S.Elevated levels of hypoxia-inducible microRNA-210 in pre-eclampsia:new insights into molecular mechanisms for the disease.Cell Mol Med.2012;16:249-259.
Dai Y,Diao Z,Sun H,Li R,Qiu Z,Hu Y.MicroRNA- 155 is involved in the remodelling of human-trophoblast-derived HTR-8/SVneo cells induced by lipopoly saccharides.Hum Reprod.2011;26:1882-1891.
Lee DC,Romero R,Kim JS,Tarca AL,Montenegro D,Pineles BL,Kim E,Lee J,Kim SY,Draghici S,Mittal P,Kusanovic JP,Chaiworapongsa T,Hassan SS,Kim CJ.miR-210 targets iron-sulfur cluster scaffold homologue in human trophoblast cell lines:siderosis of interstitial trophoblasts as a novel pathology of preterm preeclampsia and small-for-gestational-age pregnancies.Am J Pathol.2011;179:590-602.
Dai Y,Qiu Z,Diao Z,Shen L,Xue P,Sun H,Hu Y.MicroRNA- 155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin Dl.Placenta.2012;33:824-829.
Wang Y,Fan H,Zhao G,Liu D,Du L,Wang Z,Hu Y,Hou Y.miR-16 inhibits the proliferation and angiogenesis-regulating potential of mesenchymal stem cells in severe preeclampsia.FEBS J.2012;279:4510-4524.
Muralimanoharan S,Maloyan A,Mele J,Guo C,Myatt LG,Myatt L.MIR-210 modulates mitochondrial respiration in placenta with preeclampsia.Placenta.2012;33:816-823.
Li P,Guo W,Du L,Zhao J,Wang Y,Liu L,Hu Y,Hou Y.microRNA-29b contributes to pre-eclampsia through its effects on apoptosis,invasion and angiogenesis of trophoblast cells.Clin Sci(Lond).2013;124:27-40.
Liu L,Wang Y,Fan H,Zhao X,Liu D,Hu Y,Kidd AR 3rd,Bao J,Hou Y.MicroRNA-181a regulates local immune balance by inhibiting proliferation and immunosuppressive properties of mesenchymal stem cells.Stem Cells.2012;30:1756-1770.
Luo L,Ye G,Nadeem L,Fu G,Yang BB,Honarparvar E,Dunk C,Lye S,Peng C.MicroRNA-378a-5p promotes trophoblast cell survival,migration and invasion by targeting Nodal.J Cell Sci.2012;125:3124-3132.
Amer M,Elhefnawi M,El-Ahwany E,Awad AF,Gawad NA,Zada S,Tawab FM.
Hsa-miR-195 targets PCMT1 in hepatocellular carcinoma that increases tumor life span.Tumour Biol.2014.[Epub ahead of print]
Zhao FL,Dou YC,Wang XF,Han DC,Lv ZG,Ge SL,Zhang YK.Serum microRNA-195 is down-regulated in breast cancer:a potential marker for the diagnosis of breast cancer.Mol Biol Rep.2014;41:5913-5922.
Jain M,Zhang L,Boufraqech M,Liu-Chittenden Y,Bussey K,Demeure MJ,Wu X,Su L,Pacak K,Stratakis CA,Kebebew E.ZNF367 inhibits cancer progression and is targeted by miR-195.PLoS One.2014;9:el01423.
Guo H,Li W,Zheng T,Liu Z.miR-195 Targets HDGF to inhibit proliferation and invasion of NSCLC cells.Tumour Biol.2014;35:8861-8866.
Wang X,Wang Y,Lan H,Li J.miR-195 inhibits the growth and metastasis of NSCLC cells by targeting IGF1R.Tumour Biol.2014;35:8765-8770.
Wang L,Qian L,Li X,Yan J.MicroRNA-195 inhibits colorectal cancer cell proliferation,colony-formation and invasion through targeting CARMA3.Mol Med Rep.2014;10:473-478.
Yang Y,Li M,Chang S,Wang L,Song T,Gao L,Hu L,Li Z,Liu L,Yao J,Huang C.MicroRNA-195acts as a tumor suppressor by directly targeting Wnt3a in HepG2hepatocellular carcinoma cells.Mol Med Rep.2014;10:2643-2648.
Luo Q,Wei C,Li X,Li J,Chen L,Huang Y,Song H,Li D,Fang L.MicroRNA-195-5p is a potential diagnostic and therapeutic target for breast cancer.Oncol Rep.2014;31:1096-1102.
Fu MG,Li S,Yu TT,Qian LJ,Cao RS,Zhu H,Xiao B,Jiao CH,Tang NN,Ma JJ,Hua J,Zhang WF,Zhang HJ,Shi RH.Differential expression of miR-195 in esophageal squamous cell carcinoma和miR-195expression inhibits tumor cell proliferation and invasion by targeting of Cdc42.FEBS Lett.2013;587:3471-3479.
Chen YQ,Wang XX,Yao XM,Zhang DL,Yang XF,Tian SF,Wang NS.Abated microRNA-195expression protected mesangial cells from apoptosis in early diabetic renal injury in mice.J Nephrol.2012;25:566-576.
Mortuza R,Feng B,Chakrabarti S.miR-195 regulates SIRT1 -mediated changes in diabetic retinopathy.Diabetologia.2014;57:1037-1046.
Yang WM,Jeong HJ,Park SY,Lee W.Saturated fatty acid-induced miR-195 impairs insulin signaling and glycogen metabolism in HepG2 cells.FEBS Lett.2014;5793:00681-00684.
Ortega FJ,Mercader JM,Moreno-Navarrete JM,Rovira O,Guerra E,Esteve E,Xifra G,Martinez C,Ricart W,Rieusset J,Rome S,Karczewska-Kupczewska M,Straczkowski M,Fernandez-Real JM.Profiling of circulating microRNAs reveals common microRNAs linked to type 2 diabetes that change with insulin sensitization.Diabetes Care.2014;37:1375-1383.
Herrera BM,Lockstone HE,Taylor JM,Ria M,Barrett A,Collins S,Kaisaki P,Argoud K,Fernandez C,Travers ME,Grew JP,Randall JC,Gloyn AL,Gauguier D,McCarthy MI,Lindgren CM.Global microRNAexpression profiles in insulin target tissues in a spontaneous rat model of type 2 diabetes.Diabetologia.2010;53:1099-1109.
Guo ST,Jiang CC,Wang GP,Li YP,Wang CY,Guo XY,Yang RH,Feng Y,Wang FH,Tseng HY,Thorne RF,Jin L,Zhang XD.MicroRNA-497 targets insulin-like growth factor 1 receptor and has a tumour suppressive role in human colorectal cancer.Oncogene.2013;32:1910-1920.
Long G,Wang F,Duan Q,Yang S,Chen F,Gong W,Yang X,Wang Y,Chen C,Wang DW.Circulating miR-30a,miR-195 and let- 7b associated with acute myocardial infarction.PLoS One.2012;7:e50926.
You XY,Huang JH,Liu B,Liu SJ,Zhong Y,Liu SM.HMGA1 is a new target of miR-195 involving isoprenaline-induced cardiomyocyte hypertrophy.Biochemistry(Mosc).2014;79:538-544.
van Rooij E,Sutherland LB,Liu N,Williams AH,Mc Anally J,Gerard RD,Richardson JA,Olson EN.A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure.Proc Natl Acad Sci U S A.2006;103:18255-18260.
Busk PK,Cirera S.MicroRNA profiling in early hypertrophic growth of the left ventricle in rats.Biochem Biophys Res Commun.2010;396:989-993.
Porrello ER,Johnson BA,Aurora AB,Simpson E,Nam YJ,Matkovich SJ,Dorn GW 2nd,van Rooij E,Olson EN.MiR-15 family regulates postnatal mitotic arrest of cardiomyocytes.Circ Res.2011;109:670-679.
Zampetaki A,Attia R,Mayr U,Gomes RS,Phinikaridou A,Yin X,Langley SR,Willeit P,Lu R,Fanshawe B,Fava M,Barallobre-Barreiro J,Molenaar C,So PW,Abbas A,Jahangiri M,Waltham M,Botnar R,Smith A,Mayr M.Role of miR-195 in Aortic Aneurysmal Disease.Circ Res.2014;115:857-866.
Qin J,Liang H,Shi D,Dai J,Xu Z,Chen D,Chen X,Jiang Q.A panel of microRNAs as a new biomarkers for the detection of deep vein thrombosis./Thromb Thrombolysis.2014;Epub ahead of print.
Gu Y,Sun J,Groome LJ,Wang Y.Differential miRNAexpression profiles between the first and third trimester human placentas.Am J Physiol Endocrinol Metab.2013;304:E836-843.
Wang R,Zhao N,Li S,Fang JH,Chen MX,Yang J,Jia WH,Yuan Y,Zhuang SM.MicroRNA-195 suppresses angiogenesis and metastasis of hepatocellular carcinoma by inhibiting the expression of VEGF,VAV2,and CDC42.Hepatology.2013;58:642-653.
Xu P,Zhao Y,Liu M,Wang Y,Wang H,Li YX,Zhu X,Yao Y,Wang H,Qiao J,Ji L,Wang YL.Variations of microRNAs in human placentas and plasma from preeclamptic pregnancy.Hypertension.2014;63:1276-1284.
Zhu XM,Han T,Sargent IL,Yin GW,Yao YQ.Differential expression profile of microRNAs in human placentas from preeclamptic pregnancies vs normal pregnancies.Am J Obstet Gynecol.2009;200:661.el-7.
Hu Y,Li P,Hao S,Liu L,Zhao J,Hou Y.Differential expression of microRNAs in the placentae of Chinese patients with severe pre-eclampsia.Clin Chem Lab Med.2009;47:923-929.
Huang PL.Endothelial nitric oxide synthase and endothelial dysfunction.Curr Hypertens Rep.2003;5:473-480.
Albrecht EW,Stegeman CA,Heeringa P,Henning RH,van Goor H.Protective role of endothelial nitric oxide synthase.J Pathol.2003;199:8-17.
Forstermann U,Miinzel T.Endothelial nitric oxide synthase in vascular disease:from marvel to menace.Circulation.2006;113:1708-1714.
Duda DG,Fukumura D,Jain RK.Role of eNOS in neovascularization:NO for endothelial progenitor cells.Trends Mol Med.2004;10:143-145.
Lin MI,Sessa WC.Vascular endothelial growth factor signaling to endothelial nitric oxide synthase:more than a FLeeTing moment.Circ Res.2006,99:666-668.
Fatini C,Sticchi E,Gensini F,Genuardi M,Tondi F,Gensini GF,Riviello C,Parretti E,Mello G,Abbate R.Endothelial nitric oxide synthase gene influences the risk of preeclampsia,the recurrence of negative pregnancy events,and the maternal-fetal flow./Hypertens.2006;24:1823-1929.
Sun HX,Zeng DY,Li RT,Pang RP,Yang H,Hu YL,Zhang Q,Jiang Y,Huang LY,Tang YB,Yan GJ,Zhou JG.Essential role of microRNA-155 in regulating endothelium-dependent vasorelaxation by targeting endothelial nitric oxide synthase.Hypertension.2012;60:1407-1414.
Ahmad S,Ahmed A.Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia.Circ Res.2004;95:884-891.
Cudmore M,Ahmad S,Al-Ani B,Fujisawa T,Coxall H,Chudasama K,Devey LR,Wigmore SJ,Abbas A,Hewett PW,Ahmed A.Negative regulation of soluble Flt-1 and soluble endoglin release by heme oxygenase-1.Circulation.2007;115:1789-1797.
McCarthy FP,Drewlo S,Kingdom J,Johns EJ,Walsh SK,Kenny LC.Peroxisome proliferator-activated receptor-γas a potential therapeutic target in the treatment of preeclampsia.Hypertension.2011;58:280-286.
Bussolati B,Dunk C,Grohman M,Kontos CD,Mason J,Ahmed A.Vascular endothelial growth factor receptor- 1 modulates vascular endothelial growth factor-mediated angiogenesis via nitric oxide.Am J Pathol 2001;159:993-1008.
Ahmad S,Hewett PW,Wang P,Al-Ani B,Cudmore M,Fujisawa T,Haigh JJ,le Noble F,Wang L,Mukhopadhyay D,Ahmed A.Direct evidence for endothelial vascular endothelial growth factor receptor- 1 function in nitric oxide-mediated angiogenesis.Circ Res.2006;99:715-722.
Hunter Al,Aitkenhead M,Caldwell C,McCracken G,Wilson D,McClure N.Serum levels of vascular endothelial growth factor in preeclamptic and normotensive pregnancy.Hypertension.2000;36:965-969.
Ozkan S I,Vural B,Filiz S,Co§tur P,Dalcik H.Placental expression of insulin-like growth factor-I,fibroblast growth factor-basic,and neural cell adhesion molecule in preeclampsia./Matern Fetal Neonatal Med.2008;21:831-838.
参考文献:
Cudmore M,Ahmad S,Al-Ani B,Fujisawa T,Coxall H,Chudasama K,Devey LR,Wigmore SJ,Abbas A,Hewett PW,Ahmed A.Negative regulation of soluble Flt-1 and soluble endoglin release by heme oxygenase-1.Circulation.2007;115:1789-1797.
Ahmad S,Hewett PW,Al-Ani B,Sissaoui S,Fujisawa T,Cudmore MJ,Ahmed A.Autocrine activity of soluble Flt-1 controls endothelial cell function and angiogenesis.Vase Cell.2011;3:15.
Levine RJ,Maynard SE,Qian C,Lim KH,England LJ,Yu KF,Schisterman EF,Thadhani R,Sachs BP,Epstein FH,Sibai BM,Sukhatme VP,Karumanchi SA.Circulating angiogenic factors and the risk of preeclampsia.N Engl J Med.2004;350:672-683.
Venkatesha S,Toporsian M,Lam C,Hanai J,Mammoto T,Kim YM,Bdolah Y,Lim KH,Yuan HT,Libermann TA,Stillman IE,Roberts D,D'Amore PA,Epstein FH,Sellke FW,Romero R,Sukhatme VP,Letarte M,Karumanchi SA.Soluble endoglin contributes to the pathogenesis of preeclampsia.Nat Med.2006;12:642-649.
Ramma W,Ahmed A.Is inflammation the cause of pre-eclampsia?Biochem Soc Trans.2011;39:1619-1627.
Kendall RL,Wang G,Thomas KA.Identification of a natural soluble form of the vascular endothelial growth factor receptor,FLT-1,and its heterodimerization with KDR.Biochem Biophys Res Commun.1996;226:324-328.
Wu FT,Stefanini MO,Mac Gabhann F,Kontos CD,Annex BH,Popel AS.A systems biology perspective on sVEGFRl:its biological function,pathogenic role and therapeutic use./Cell Mol Med.2010;14:528-552.
Stepan H,Unversucht A,Wessel N,Faber R.Predictive value of maternal angiogenic factors in second trimester pregnancies with abnormal uterine perfusion.Hypertension.2007;49:818-824.
Bando H,Weich HA,Brokelmann M,Horiguchi S,Funata N,Ogawa T,Toi M.Association between intratumoral free and total VEGF,soluble VEGFR-1,VEGFR-2 and prognosis in breast cancer.Br J Cancer.2005;92:553-561.
Ebos JM,Bocci G,Man S,Thorpe PE,Hicklin DJ,Zhou D,Jia X,Kerbel RS.A naturally occurring soluble form of vascular endothelial growth factor receptor 2 detected in mouse and human plasma.Mol Cancer Res.2004;2:315-326.
Lamszus K,Ulbricht U,Matschke J,Brockmann MA,Fillbrandt R,Westphal M.Levels of soluble vascular endothelial growth factor(VEGF)receptor 1 in astrocytic tumors and its relation to malignancy,vascularity,and VEGF-A.Clin Cancer Res.2003;9:1399-1405.
Jaroszewicz J,Januszkiewicz M,Flisiak R,Rogalska M,Kalinowska A,Wierzbicka I.Circulating vascular endothelial growth factor and its soluble receptors in patients with liver cirrhosis:possible association with hepatic function impairment.Cytokine.2008;44:14-17.
Blann AD,Belgore FM,McCollum CN,Silverman S,Lip PL,Lip GY.Vascular endothelial growth factor and its receptor,Fit-1,in the plasma of patients with coronary or peripheral atherosclerosis,or Type II diabetes.Clin Sci(Lond).2002;102:187-194.
Findley CM,Mitchell RG,Duscha BD,Annex BH,Kontos CD.Plasma levels of soluble Tie2 and vascular endothelial growth factor distinguish critical limb ischemia from intermittent claudication in patients with peripheral arterial disease.J Am Coll Cardiol.2008;52:387-393.
Sibai B,Dekker G,Kupferminc M.Pre-eclampsia.Lancet.2005;365:785-799.
Steegers EA,von Dadelszen P,Duvekot JJ,Pijnenborg R.Pre-eclampsia.Lancet.2010;376:631-644.
Silasi M,Cohen B,Karumanchi SA,Rana S.Abnormal placentation,angiogenic factors,and the pathogenesis of preeclampsia.Obstet Gynecol Clin North Am.2010;37:239-253.
Maynard S,Epstein FH,Karumanchi SA.Preeclampsia and angiogenic imbalance.Annu Rev Med.2008;59:61-78.
Maynard SE,Min JY,Merchan J,Lim KH,Li J,Mondal S,Libermann TA,Morgan JP,Sellke FW,Stillman IE,Epstein FH,Sukhatme VP,Karumanchi SA.Excess placental soluble fms-like tyrosine kinase 1(sFltl)may contribute to endothelial dysfunction,hypertension,and proteinuria in preeclampsia./Clin Invest.2003;111:649-658.
Powe CE,Levine RJ,Karumanchi SA.Preeclampsia,a disease of the maternal endothelium:the role of antiangiogenic factors and implications for later cardiovascular disease.Circulation.2011;123:2856-2869.
Costantine MM,Cleary K;Eunice Kennedy Shriver National Institute of Child Health and Human Development Obstetric—Fetal Pharmacology Research Units Network.Pravastatin for the prevention of preeclampsia in high-risk pregnant women.Obstet Gynecol.2013;121:349-353.
Bartel DP.MicroRNAs:target recognition and regulatory functions.Cell.2009;136:215-233.
Lewis BP,Shih IH,Jones-Rhoades MW,Bartel DP,Burge CB.Prediction of mammalian microRNA targets.Cell.2003;115:787-798.
Williams AE.Functional aspects of animal microRNAs.Cell Mol Life Sci.2008;65:545-562.
He L,Hannon GJ.MicroRNAs:small RNAs with a big role in gene regulation.Nat Rev Genet.2004;5:522-531.
Zhu XM,Han T,Sargent IL,Yin GW,Yao YQ.Differential expression profile of microRNAs in human placentas from preeclamptic pregnancies vs normal pregnancies.Am J Obstet Gynecol.2009;200:661.el-7.
Hu Y,Li P,Hao S,Liu L,Zhao J,Hou Y.Differential expression of microRNAs in the placentae of Chinese patients with severe pre-eclampsia.Clin Chem Lab Med.2009;47:923-929.
Pineles BL,Romero R,Montenegro D,Tarca AL,Han YM,Kim YM,Draghici S,Espinoza J,Kusanovic JP,Mittal P,Hassan SS,Kim CJ.Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia.Am J Obstet Gynecol.2007;196:261.el-6.
Mayor-Lynn K,Toloubeydokhti T,Cruz AC,Chegini N.Expression profile of microRNAs and mRNAs in human placentas from pregnancies complicated by preeclampsia and preterm labor.Reprod Sci.2011;18:46-56.
Enquobahrie DA,Abetew DF,Sorensen TK,Willoughby D,Chidambaram K,Williams MA.Placental microRNA expression in pregnancies complicated by preeclampsia.Am J Obstet Gynecol.2011;204:178.el2-21.
Noack F,Ribbat-Idel J,Thorns C,Chiriac A,Axt-Fliedner R,Diedrich K,Feller AC.miRNA expression profiling in formalin-fixed and paraffin-embedded placental tissue samples from pregnancies with severe preeclampsia.J Perinat Med.2011;39:267-271.
Kozakowska M,Szade K,Dulak J,Jozkowicz A.Role of heme oxygenase- 1 in postnatal differentiation of stem cells:a possible cross-talk with microRNAs.Antioxid Redox Signal.2014;20:1827-1850.
Skrzypek K,Tertil M,Golda S,Ciesla M,Weglarczyk K,Collet G,Guichard A,Kozakowska M,Boczkowski J,Was H,Gil T,Kuzdzal J,Muchova L,Vitek L,Loboda A,Jozkowicz A,Kieda C,Dulak J.Interplay between heme oxygenase- 1 and miR-378 affects non-small cell lung carcinoma growth,vascularization,and metastasis.Antioxid Redox Signal.2013;19:644-660.
Hou W,Tian Q,Steuerwald NM,Schrum LW,Bonkovsky HL.The let-7 microRNA enhances heme oxygenase-1 by suppressing Bachl and attenuates oxidant injury in human hepatocytes.Biochim Biophys Acta.2012;1819:1113-1122.
序列表
<110> 阿斯顿大学
<120> 先兆子痫
<130> P344117PCT
<160> 29
<170> PatentIn version 3.5
<210> 1
<211> 22
<212> RNA
<213> Homo sapiens
<400> 1
uggaguguga caaugguguu ug 22
<210> 2
<211> 22
<212> RNA
<213> Homo sapiens
<400> 2
auauaauaca accugcuaag ug 22
<210> 3
<211> 21
<212> RNA
<213> Homo sapiens
<400> 3
ucagugcaug acagaacuug g 21
<210> 4
<211> 21
<212> RNA
<213> Homo sapiens
<400> 4
uagcagcaca gaaauauugg c 21
<210> 5
<211> 22
<212> RNA
<213> Homo sapiens
<400> 5
caaacaccau ugucacacuc ca 22
<210> 6
<211> 22
<212> RNA
<213> Homo sapiens
<400> 6
cacuuagcag guuguauuau au 22
<210> 7
<211> 21
<212> RNA
<213> Homo sapiens
<400> 7
ccaaguucug ucaugcacug a 21
<210> 8
<211> 21
<212> RNA
<213> Homo sapiens
<400> 8
gccaauauuu cugugcugcu a 21
<210> 9
<211> 23
<212> DNA
<213> Homo sapiens
<400> 9
tttgtagcat tgtcatcact cct 23
<210> 10
<211> 20
<212> DNA
<213> Homo sapiens
<400> 10
tttgtcgacg gatagagaat 20
<210> 11
<211> 20
<212> DNA
<213> Homo sapiens
<400> 11
gtcaaaatag attattataa 20
<210> 12
<211> 17
<212> DNA
<213> Homo sapiens
<400> 12
gtcaagagca aggcgca 17
<210> 13
<211> 22
<212> DNA
<213> Homo sapiens
<400> 13
tacaatattt gtactattat at 22
<210> 14
<211> 19
<212> DNA
<213> Homo sapiens
<400> 14
tacaatattt agacgcgct 19
<210> 15
<211> 21
<212> DNA
<213> Homo sapiens
<400> 15
tccctccccc ccatgctgtg g 21
<210> 16
<211> 21
<212> DNA
<213> Homo sapiens
<400> 16
tgtaaacaag ggtccacagc a 21
<210> 17
<211> 45
<212> DNA
<213> Homo sapiens
<400> 17
ctctcaggag tagagtacct gtaaaggaga atctctaaat caagt 45
<210> 18
<211> 45
<212> DNA
<213> Homo sapiens
<400> 18
acttgattta gagattctcc tttacaggta ctctactcct gagag 45
<210> 19
<211> 22
<212> RNA
<213> Homo sapiens
<400> 19
guuuguggua acagugugag gu 22
<210> 20
<211> 23
<212> RNA
<213> Homo sapiens
<400> 20
uuuguagcau ugucaucacu ccu 23
<210> 21
<211> 22
<212> RNA
<213> Homo sapiens
<400> 21
gugaaucguc caacauaaua ua 22
<210> 22
<211> 20
<212> RNA
<213> Homo sapiens
<400> 22
gucaaaauag auuauuauaa 20
<210> 23
<211> 21
<212> RNA
<213> Homo sapiens
<400> 23
gguucaagac aguacgugac u 21
<210> 24
<211> 21
<212> RNA
<213> Homo sapiens
<400> 24
ucccuccccc ccaugcacug u 21
<210> 25
<211> 16
<212> DNA
<213> Mus musculus
<400> 25
taaagtctaa tctcta 16
<210> 26
<211> 21
<212> DNA
<213> Macaca mulatta
<400> 26
acctgtaaag tctaatctct a 21
<210> 27
<211> 21
<212> DNA
<213> Gorilla gorilla
<400> 27
acctgtaaag tctaatctct a 21
<210> 28
<211> 21
<212> DNA
<213> Pan troglodytes
<400> 28
acctgtaaag tctaatctct a 21
<210> 29
<211> 21
<212> DNA
<213> Homo sapiens
<400> 29
acctgtaaag tctaatctct a 21
- 还没有人留言评论。精彩留言会获得点赞!