针对蛋白S的特异性siRNA在治疗血友病中的用途的制作方法
本发明涉及使用针对蛋白s的sirna治疗血友病。
背景技术:
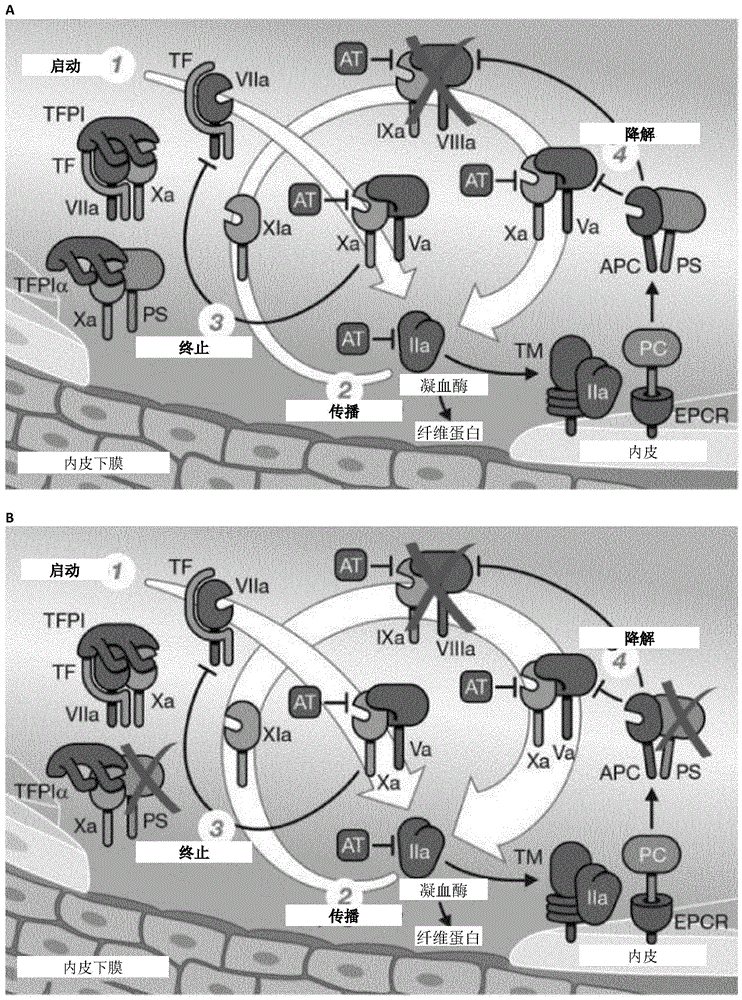
血友病a(ha)和b(hb)是遗传性x连锁疾病。它们分别是由因子viii(fviii)(f8)或因子ix(fix)基因(f9)的突变引起的,从而导致编码蛋白质的缺失,而该蛋白质是凝血内在途径的重要组成部分(图1a)。
患有严重血友病的患者经常会在肌肉骨骼系统内自发性出血,例如关节积血(hemarthrosis)。如果不及时治疗,可能导致年轻时的残疾。
目前的血友病治疗包括因子替代治疗。这种疗法可改善生活质量(qualityoflife)(qol),但仍存在一些缺点。因子是通过静脉内施用的,由于其半衰期短,因此必须反复输注,这种方式会使患者感到严重不适,并有感染和静脉损伤的风险。更重要的是,接受因子替代疗法的患者会发展成抑制性同种抗体(alloantibodies)。抑制剂会使得替代疗法无效,并限制了患者获得安全有效的标准护理,并增加了他们的易发病率和死亡风险。
新疗法专注于开发能够减少预防性(prophylactic)输注频率的产品,从而有可能同时提高对疗法和qol的依从性(compliance)。除了持久的fviii和fix,新方法还包括替换产生内源性凝血因子所需的基因,模仿缺失因子的凝血功能的双特异性抗体技术以及靶向凝血抑制剂(例如组织因子途径抑制剂)(tissuefactorpathwayinhibitor,tfpi)或抗凝血酶作为血友病患者凝血功能再平衡的策略。最近,研究表明,在血友病小鼠模型中,活化蛋白c(apc)特异的丝氨酸蛋白酶抑制蛋白可在体外(invitro)拯救凝血酶的产生并恢复止血作用。
基于上述现有技术,本发明的目的是提供新颖的血友病治疗方法和手段。该目的通过本说明书的权利要求书实现。
技术实现要素:
本发明的第一方面提供了针对蛋白s的sirna在治疗血友病的方法中的用途。
在本说明书的上下文中,术语血友病涉及身体的使血液凝结的能力受损的病症(condition)。血友病的基因型包括遗传性血友病a,血友病b和血友病c。
在本说明书的上下文中,蛋白s涉及由基因pros1(ncbi基因id:5627)编码的人“维生素k依赖性蛋白s”(uniprotidp07225)。
存在人蛋白s的两个转录本变体。与转录本变体1相比,转录本变体2在5′编码区缺少一个选择性框内外显子。编码的同种型(isoform)2比同种型1短。转录本变体2的特征在于seqidno001。转录本变体1的特征在于seqidno002。
在本说明书的上下文中,术语sirna(小/短干扰rna)涉及能够干扰包含与该序列互补的核酸序列的基因的表达(换句话说:阻止表达)的rna分子,该过程称为rna干扰。术语sirna旨在涵盖单链sirna和双链sirna。sirna通常以17-24bp的长度为特征。双链sirna衍生自1个较长的双链rna分子(dsrna)。长的dsrna被内切核糖核酸酶(称为dicer)切割,形成双链sirna。在核蛋白复合体(称为risc)中,双链sirna解螺旋(unwound)形成单链sirna。rna干扰通常通过将sirna分子与具有互补序列的mrna分子结合而起作用,从而导致mrna降解。rna干扰也可能通过将sirna分子与细胞核内的pre-mrna(未成熟的,未剪接的mrna)的内含子(intronic)序列结合而发生,从而导致pre-mrna降解。
发明人研究了靶向蛋白s(ps)是否可以通过重新平衡凝血来促进血友病中的止血(图1b)。由pros1基因编码的ps,在因子va(fva)和fviiia失活时,充当apc的辅助因子,在fxa抑制中,充当tfpi的辅助因子。这种双重作用使ps成为凝血酶生成的关键调节剂。
在某些实施方案中,sirna包含17-24个核苷酸。
在某些实施方案中,sirna包含18-22个核苷酸。
在某些实施方案中,sirna包含19个核苷酸。
在某些实施方案中,所述sirna其特征在于,其是与seqid001反向互补的序列。
在某些实施方案中,所述sirna其特征在于,其是与seqid002反向互补的序列。
在某些实施方案中,所述其特征在于,其是与选自由seqidno003、seqidno004、seqidno005、seqidno006、seqidno007、seqidno008、seqidno009、seqidno010、seqidno011、seqidno012、seqidno013、seqidno014、seqidno015、seqidno016、seqidno017和seqidno018组成的组的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno003反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno004反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno005反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno006反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno007反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno008反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno009反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno010反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno011反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno012反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno013反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno014反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno015反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno016反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno017反向互补的序列。
在某些实施方案中,所述sirna其特征在于与seqidno018反向互补的序列。
在某些实施方案中,sirna其特征在于与通过并列上文定义的序列的任何两个连续序列形成的序列反向互补的序列。作为非限制性实例,sirna其特征在于与通过并列seqidno014和seqidno015形成的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与选自seqidno014(外显子12)、seqidno011(外显子9)、seqidno006(外显子4)、seqidno012(外显子10)和seqidno013(外显子11)组成的组的序列反向互补。
在某些实施方案中,所述sirna其特征在于,其是与以seqidno001的核苷酸1-100、101-200、201-300、301-400、301-400、401-500、501-600、601-700、701-800、801-900、901-1000、1001-1100、1101-1200、1201-1300、1301-1400、1501-1600、1701-1800、1801-1900、1901-2000、2001-2100、2101-2200、2201-2300、2301-2400、2501-2600、2701-2800、2801-2900、2901-3000、3001-3100、3101-3200、3201-3300、3301-3400、3401-3500或3501-3580为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1-100为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸101-200为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸201-300为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸301-400为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸401-500为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸501-600为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸601-700为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸701-800为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸801-900为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸901-1000为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1001-1100为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1100-1200为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1201-1300为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1301-1400为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1401-1500为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1501-1600为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1601-1700为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1701-1800为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1801-1900为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸1901-2000为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2001-2100为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2100-2200为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2201-2300为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2301-2400为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2401-2500为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2501-2600为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2601-2700为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2701-2800为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2801-2900为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸2901-3000为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3001-3100为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3100-3200为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3201-3300为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3301-3400为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3401-3500为特征的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于与以seqidno001的核苷酸3501-3580为特征的序列反向互补的序列。
在某些实施方案中,sirna其特征在于与通过并列上文定义的序列的任何两个连续序列形成的序列反向互补的序列。作为非限制性实例,sirna其特征在于与通过并列seqidno001的1501-1600和seqidno001的1501-1600形成的序列反向互补的序列。
在某些实施方案中,sirna针对蛋白s的内含子序列。
在某些实施方案中,所述rna的特征在于与选自由seqidno014(外显子12),seqidno011(外显子9),seqidno006(外显子4),seqidno012(外显子10)和seqidno013(外显子11)组成的组的序列反向互补的序列。
在某些实施方案中,所述sirna其特征在于包含选自以下的序列或由其组成的序列组成:seqidno019(sirna_1)、seqidno020(sirna_2)、seqidno021(sirna_3)、seqidno022(sirna_4)、seqidno023(sirna_5)、seqidno024(sirna_6)、seqidno025(sirna_7)、seqidno026(sirna_8)、seqidno027(sirna_9)、seqidno028(sirna_10)、seqidno029(sirna_11)、seqidno030(sirna_12)、seqidno031(sirna_13)、seqidno032(sirna_14)、seqidno033(sirna_15)、seqidno034(sirna_16)、seqidno035(sirna_17)、seqidno036(sirna_18)、seqidno037(sirna_19)、seqidno038(sirna_20)、seqidno039(sirna_21)、seqidno040(sirna_22)、seqidno041(sirna_23)、seqidno042(sirna_24)、seqidno043(sirna_25)、seqidno044(sirna_26)、seqidno045(sirna_27)、seqidno046(sirna_28)、seqidno047(sirna_29)、seqidno048(sirna_30)、seqidno049(sirna_31)、seqidno050(sirna_32)、seqidno051(sirna_33)、seqidno052(sirna_34)、seqidno053(sirna_35)、seqidno054(sirna_36)、seqidno055(sirna_37)、seqidno056(sirna_38)、seqidno057(sirna_39)、seqidno058(sirna_40)、seqidno059(sirna_41)、seqidno060(sirna_42)、seqidno061(sirna_43)、seqidno062(sirna_44)、seqidno063(sirna_45)、seqidno064(sirna_46)、seqidno065(sirna_47)、seqidno066(sirna_48)、seqidno067(sirna_49)、andseqidno068(sirna_50)。
在某些实施方案中,提供了所述sirna在治疗血友病a的方法中的用途。
在某些实施方案中,提供了所述sirna在治疗血友病b的方法中的用途。
类似地,在本发明范围内的是一种在有需要的患者中治疗血友病的方法,其包括向所述患者施用包含根据本发明的sirna的分子。
类似地,提供了用于预防或治疗血友病的剂型(dosageform),其包括包含根据本发明的sirna的分子。
在本说明书的上下文中,表述“包含根据本发明的sirna的分子”意在涵盖根据本发明的sirna和分子,特别是dsrna分子,可以通过rna干扰途径在哺乳动物细胞内,从该分子产生本发明的sirna。
在本发明的上下文中,“核苷酸”是核酸或核酸类似物结构单元,其寡聚物能够基于碱基配对与rna寡聚物形成选择性杂交体。在本文中,术语核苷酸包括经典的核糖核苷酸结构单元、腺苷、鸟苷、尿苷(和核糖基胸苷(ribosylthymin))、胞苷、经典的脱氧核糖核苷酸、脱氧腺苷、脱氧鸟苷、胸苷、脱氧尿苷和脱氧胞苷。它进一步包括核酸的类似物,例如磷酸酯、2′-o-甲基硫代磷酸酯、肽核酸(pna;通过肽键连接的n-(2-氨基乙基)-甘氨酸单元,其核碱基附着在甘氨酸的α-碳上)或锁定的核酸(lna;2′o,4′c亚甲基桥接的rna结构单元)。杂交序列可以由任何上述核苷酸或其混合物组成。
在某些实施方案中,根据本发明的sirna的杂交序列包含17、18、19、20、21、22、23、24、25、26或27个核苷酸。
在某些实施方案中,杂交序列与seqid001或seqid002的反向互补序列具有至少95%同一性,更优选具有96%、97%、98%、99%或100%同一性。在某些实施方案中,杂交序列包括脱氧核苷酸、硫代磷酸酯(phosphothioate)脱氧核苷酸、lna和/或pna核苷酸或其混合物。
在本说明书的上下文中,术语序列同一性和序列同一性百分比是指通过比较两个比对的序列而确定的值。用于比较的序列比对方法是本领域众所周知的。用于比较的序列的比对可以通过smith和waterman,adv.appl.math.2:482(1981)的局部同源性算法来进行,通过needleman和wunsch,j.mol.biol.48:443(1970)的全局比对算法进行,通过pearson和lipman,proc.nat.acad.sci.85:2444(1988)寻找相似性的方法进行,或通过这些算法的计算机实现,包括但不限于:clustal、gap、bestfit、blast、fasta和tfasta。用于进行blast分析的软件是可公开获得的,例如通过国家生物技术信息中心(http://blast.ncbi.nlm.nih.gov/)。
比较氨基酸序列的一个例子是使用默认设置的blastp算法:expectthreshold:10;wordsize:3;maxmatchesinaqueryrange:0;matrix:blosum62;gapcosts:existence11,extension1;compositionaladjustments:conditionalcompositionalscorematrixadjustment。比较核酸序列的一个这样的例子是使用默认设置的blastn算法:expectthreshold:10;wordsize:28;maxmatchesinaqueryrange:0;match/mismatchscores:1.-2;gapcosts:linear。除非另有说明,否则本文提供的序列同一性值是指使用blast程序套件(altschuletal.,j.mol.biol.215:403-410(1990))使用上面确定的默认参数分别进行蛋白质和核酸比较。
在一些实施方案中,杂交序列包含核糖核苷酸,硫代磷酸酯和/或2′-o-甲基修饰的硫代磷酸酯核糖核苷酸。
在一些实施方案中,杂交序列包含脱氧核苷酸、硫代磷酸酯脱氧核苷酸、硫代磷酸酯核糖核苷酸和/或2′-o-甲基修饰的硫代磷酸酯核糖核苷酸。
在本文的任何地方出现的,将单个可分离特征的替代方案设置为“实施例”的情况,应当理解,这样的替代方案可以自由地组合以形成本文公开的本发明的离散实施例。
通过以下实施例和附图进一步说明本发明,从中可以得出其他实施方案和优点。这些实施例旨在说明本发明,而不是限制其范围。
附图说明
图1显示x酶活性的丧失拯救了pros1-/-小鼠。a,血友病情况下凝血酶产生的示意图。主要的凝血复合物之一是固有的十酶(tenase)(x-ase)复合物。x酶包含激活的fix(fixa)作为蛋白酶,激活的fviii(fviiia)作为辅因子和x因子(fx)作为底物。尽管在损伤部位组织因子(tf)的产生或暴露是通过外在途径引发凝血的主要事件,但内在途径x酶很重要,因为体内可用的活性tf数量有限,并且tfpi的存在(与活化的fx(fxa)复合时),可抑制tf/活化的vii因子(fviia)复合物(图1a)。因此,持续的凝血酶生成取决于fix和fviii的激活(图1a)。该过程由于以下原因被放大:由于fviii被fxa和凝血酶激活,fix被fviia和激活的因子xi(fxia)激活,后者之前已经被凝血酶激活。因此,随着fxa和凝血酶的形成,fviii和fix的激活逐渐增加。b,通过靶向pros1在严重血友病a和b中增强凝血酶生成的实验方法。c-d,鼠模型验证以及在有或没有pros1缺乏的血友病成年小鼠中的dic血液学参数评估:ps(抗原),fviii(凝血活性)或fix(凝血活性)在f8-/-pros1+/+、f8-/-pros1+/-和f8-/-pros1-/-(c)f9-/-pros1+/+、f9-/-pros1+/-和f9-/-pros1-/-成年小鼠(d)中的血浆水平(n=5/组);血友病a组的血小板(n=7/组)、纤维蛋白原(n=8/组)、pt(n=6/组)和tat(n=6/组)(c);血友病b组(d)的血小板(n=5/组)、纤维蛋白原(n=4/组)、pt(n=4/组)和tat(n=4/组)。e-f,单次静脉内注射2u/g重组fviii
图2显示了血栓形成的小鼠模型。a-c,在f8+/+pros1+/+、f8-/-pros1+/+、f8-/-pros1+/-和f8-/-pros1-/-小鼠中,tf诱导的静脉血栓栓塞(n=10/基因型)。麻醉的小鼠通过下腔静脉静脉注射不同剂量的重组tf(innovin):a中1/2稀释度(~4.3nmtf)和b-c中1/4稀释度(~2.1nmtf)。在(a)中,一组f8+/+pros1+/+小鼠注射了低分子量肝素(heparin)(依诺肝素(enoxaparin)60μg/g皮下注射(s.c.))。记录到了呼吸停止发作的时间至少持续2分钟。实验在20分钟时终止。kaplan-meier生存曲线(a-b)。c,在呼吸停止发作后2分钟或在20分钟观察期完成时,肺被切除,并检查纤维蛋白凝块(对不溶性纤维蛋白进行免疫染色,mab克隆102-10)。d,在f8+/+pros1+/+、f8-/-pros1+/+和f8-/-pros1-/-小鼠中通过活体内显微镜记录的fecl3损伤下的肠系膜动脉中的血栓形成,代表性实验(n=3/基因型)。d,在f8+/+pros1+/+、f8-/-pros1+/+和f8-/-pros1-/-小鼠中通过活体内显微镜记录的fecl3损伤下的肠系膜动脉中的血栓形成,代表性实验(n=3/基因型)。
图3显示了尾端出血模型。分别在2毫米(a)和4毫米(b)的尾部横切30分钟(a)和10分钟(b)后,将血液收集在新鲜的生理盐水试管中;然后测量总失血量(μl)。f8+/-pros1+/+和f8+/+pros1+/+小鼠(白色柱)用作对照(a组中的所有组n=5,a组中的所有组n=6)。c,抗人ps抗体改变了4毫米横切后的尾端出血。
图4显示了急性血栓形成模型。a,在f8-/-pros1+/+、f8-/-pros1+/-、f8-/-pros1-/-和f8+/+pros1+/+小鼠中,受伤后72h和受伤前膝盖直径的差别。b,在f8+/+pros1+/+、f8-/-pros1+/+和f8-/-pros1-/-中小鼠,未受伤和腿部受伤的代表的膝关节关节腔内的显微镜评估(masson三色染色和不溶性纤维蛋白的免疫染色)。c,使用特定sirna的体内(invivo)mps沉默:用单次腹膜内(i.p.)输注mpssirna或对照sirna处理的f8-/-pros1+/-和f8-/-pros1+/+小鼠,对其受伤后72小时的关节直径进行评估。d,在先前用mpssirna或ctrlsirna处理过的f8-/-pros1+/+小鼠中,对腿部受伤72h后的代表的膝关节腔内进行显微镜评估(masson三色染色)。测量值以平均值±标准误表示。*,p<0.05;**,p<0.005;***,p<0.0005;****,p<0.0001。
图5显示ps和tfpi均在鼠滑膜中表达。a,来自先前用ctrl-sirna或mps-sirna处理过的f8-/-pros1+/+小鼠的受伤膝盖的膝关节腔内ps和tfpi的免疫染色。箭头(arrowheads)指向滑膜组织,箭头(arrows)指向血管结构,ps和tfpi均为阳性。上图中的方框(比例尺:200μm)显示了下面面板中放大的区域(比例尺:50μm)。b,来自f8-/-pros1+/+和f8-/-pros1-/-小鼠的未受伤膝盖的膝关节腔内tfpi的免疫染色。c-e,使用抗ps(c)和抗tfpi(d)抗体对原代鼠成纤维样滑膜细胞(fibroblast-likesynoviocytes)(fls)培养的条件培养基进行蛋白质印迹(westernblot)分析。无血小板血浆(pfp),血小板蛋白裂解物(plt),鼠ps(mps)用作阳性对照(c)。通过比较去糖基化的tfpi和完全糖基化的tfpi的分子量来确定tfpi同种型的表达。鼠胎盘被用作tfpiα的阳性对照。e-f,使用抗ps(f)和抗tfpi(e)抗体在凝血酶(thr,+)或溶剂(vehicle)(-)存在下培养24小时后,从fls分离的总蛋白裂解物的蛋白质印迹分析。将人重组tfpi全长用作tfpiα(hrtfpi)的阳性对照。印迹代表三次独立的实验。
图6显示了人滑膜中的ps和tfpi。a,ps和tfpi在ha(按需和预防),按需或骨关节炎(oa)的hb患者的滑膜组织中表达。箭头指向滑膜衬里层(lininglayer),箭头指向衬里层中的血管结构,对ps和tfpi均呈阳性。比例尺:50μm。b,使用抗tfpi抗体在去糖基化之前和之后,对来自健康个体和oa患者的原代人fls(hfls)培养物的条件培养基的蛋白质印迹分析。人血小板裂解物(hplt)用作tfpiα的阳性对照。印迹代表三次独立的实验。
图7显示了血友病a中的凝血酶生成和血纤蛋白网络,tf-(1pm)诱导了来自f8-/-pros1+/+和f8-/-pros1-/-小鼠的prp中的凝血酶生成,描绘了tfpi依赖的ps活性。b,来自f8-/-pros1+/+和f8-/-pros1-/-小鼠的prp和pfp中apc依赖的ps活性。c,来自f8+/+pros1+/+、f8-/-pros1+/+和f8-/-pros1-/-以及来自f9+/+pros1+/+、f9-/-pros1+/+和f9-/-pros1-/-的代表性纤维蛋白结构的扫描电子显微镜图像。dg,严重的ha患者(pviii<1%)中,没有(d,f)和有高滴度抑制剂(e,g)下,pfp(d-e)和prp(f-g)中的凝血酶生成由低浓度tf(1pm)触发。测量值以平均值±标准误表示。**,p<0.005;***,p<0.0005。
图8显示了基因分型方法。通过杂交f8-/-pros1+/-(a-c)和f9-/-pros1+/-(d-f)小鼠获得的基因型。a,通过多重pcr扩增pros1等位基因。然后将pcr产物进行电泳;根据saller,2009,与无效带(nullband)(571bp)相比,wt带具有较低的分子量(234bp)。b,设置多重pcr,从基因组dna扩增f8等位基因的wt带(620bp)和无效带(420bp)。c,与(a)中相同的样品上的f8等位基因扩增的pcr产物(无效带:420bp)。d,通过多重pcr扩增pros1等位基因。然后将pcr产物进行电泳;根据saller,2009,与无效带(571bp)相比,wt带具有较低的分子量(234bp)。e,设置多重pcr,从基因组dna扩增f9等位基因的wt带(320bp)和无效带(550bp)。f,与(d)中相同的样品上的f9等位基因扩增的pcr产物(无效带:550bp)。
图9示出了生理状态下的组织学。在f8-/-pros1-/-和f8-/-pros1+/+以及f9-/-pros1+/+和f9-/-pros1-/-小鼠的肝、肺、肾、脑切片上对不溶性纤维蛋白进行免疫染色。比例尺:100μm。
图10显示pros1的遗传缺失可预防血友病b小鼠的关节积血形成。a,f9-/-pros1+/+、f9-/-pros1+/-、f9-/-pros1-/-和f9+/+pros1+/+小鼠受伤后72小时与受伤前膝盖直径之间的差异。b,在f9+/+pros1+/+、f9-/-pros1+/+和f9-/-pros1-/-小鼠中,未受伤和腿部受伤72小时后的代表的膝关节腔内的显微镜评估(masson三色染色和不溶性纤维蛋白染色,mab克隆102-10)。例尺:500μm。测量值以平均值±标准误表示。***,p<0.0005。
图11显示了纤维蛋白网络密度和纤维分支的定量。a-b,来自f8+/+pros1+/+,f8-/-pros1+/+和f8-/-pros1-/-小鼠的纤维蛋白网络。c-d,来自f9+/+pros1+/+,f9-/-pros1+/+和f9-/-pros1-/-的纤维蛋白网络。纤蛋白网络密度的定量(a和c)。分支的纤维的定量(b和d)。测量值以平均值±标准误表示。***,p<0.0005。
具体实施方式
实施例1:x酶活性的丧失拯救pros1-/-小鼠
pros1+/-雌性与f8-/-雄性杂交产生了25%的f8+/-pros1+/-后代。f8+/-pros1+/-雌性与f8-/-雄性一起繁殖产生了25%的f8-/-pros1+/-后代(图8a-c)。用f9-/-pros1+/-小鼠进行了类似的观察(图8d-f)。如预期的那样,f8-/-pros1-/-和f9-/-pros1-/-小鼠各自不显示fviii和fix血浆活性,并且在f8-/-pros1-/-和f9-/-pros1-/-小鼠血浆中未检测到ps(图1c-d)。据报道,f8-/-pros1+/-和f9-/-pros1+/-中的ps水平比f8-/-pros1+/+和f9-/-pros1+/+小鼠低约50-60%(图1c-d)。
在f8-/-pros1+/-育种对中的295只幼崽中,有72个(24%)是f8-/-pros1+/+,有164个(56%)是f8-/-pros1+/-,有59个(20%)是f8-/-pros1-/-(χ2=4.8,p=0.09)。因此,f8-/-pros1-/-小鼠以预期的孟德尔比率存在。相比之下,在f9-/-pros1+/-配对中的219只幼崽中,有56个(26%)是f9-/-pros1+/+,有132个(60%)f9-/-pros1+/-和有31个(14%)为f9-/-pros1-/-(χ2=14.95,p=0.001)。这与f9-/-pros1-/-小鼠的传递比失真兼容,与f9+/+pros1+/+小鼠的交配相比产仔数减少一致(5.2±0.7对9.8±1.8,n=4交配/超过3t代,p=0.046)。
f8-/-pros1-/-和f9-/-pros1-/-小鼠看起来完全正常。分别监测了它们的生存能力,分别长达20(n=4)和16个月(n=2),分别与f8-/-pros1+/+和f9-/-pros1+/+小鼠相比没有任何差异。
由于小鼠中完全pros1缺乏会导致消耗性凝血病(consumptivecoagulopathy)15,我们评估了f8-/-pros1-/-和f9-/-pros1-/-小鼠是否患有dic。dic参数在f8-/-pros1+/+、f8-/-pros1+/-和f8-/-pros1-/-小鼠(图1c),和在f9-/-pros1+/+、f9-/-pros1+/-和f9-/-pros1-/-小鼠(图1d)中具有可比性。在f8-/-pros1+/+(69±2秒),f8-/-pros1+/-(68±3秒)和f8-/-pros1-/-(63±6)小鼠(平均值±标准误,每组n=6,p=0.3)中,等同地延长了激活的部分促凝血酶原激酶(thromboplastin)时间(aptt),因为不存在fviii。在f9-/-pros1+/+、f9-/-pros1+/-和f9-/-pros1-/-小鼠中获得了可比较的数据。此外,在f8-/-pros1-/-和f9-/-pros1-/-小鼠的脑、肺、肝和肾中未发现血栓形成或纤维蛋白沉积(图9)。
因此,x酶活性的丧失挽救了完全pros1缺乏症的胚胎致死率。然而,挽救对于丧失了fix活性的作用只是部分的。可能的解释是,与重度ha相比,重度hb的病情较轻。因此,pros1-/-小鼠中的f9干扰与f8干扰相比,在重新平衡凝血方面效率较低。
为了探讨通过fviii注入恢复内在的x酶活性是否在f8-/-pros1-/-小鼠中诱导dic,血栓形成和暴发性紫癜(purpurafulminans),我们静脉注射重组fviii(rfviii)。rfviii注射后没有小鼠死亡。单次注射过量的rfviii后24小时,在f8-/-pros1-/-小鼠中发现大量血管中的血栓和肺部出血(图1e-f)。重复施用正常剂量的rfviii后24小时,凝血分析显示不凝固的凝血酶原时间(prothrombintime)(pt)(未显示),低纤维蛋白原和高凝血酶-抗凝血酶(thrombin-antithrombin)(tat)水平,与明显的dic兼容(图1g)。相反,在f8-/-pros1-/-小鼠中单次注射正常剂量的rfviii后,纤维蛋白原和tat的水平与未处理的f8-/-pros1-/-小鼠相当(图1g)。尽管在肺和肝中可见大量血栓(图1h-1),但这些小鼠均未出现暴发性紫癜。
实施例2:x酶活性的丧失不能阻止tf诱导的pros1-/-小鼠血栓栓塞(thromboembolism)致死
先前我们证明,尽管88%的pros1+/+小鼠在tf诱导的血栓栓塞模型中存活下来,但在注射低tf剂量(约1.1nm)后20分钟,仅25%的pros1+/-小鼠仍然存活。当使用更高的tf剂量(约4.3nm)时,pros1+/+和pros1+/-小鼠均在20分钟内死亡。但是,pros1+/-比pros1+/+更早死亡。ha和wt小鼠对这种高tf剂量同样敏感,其中超过85%的小鼠在15分钟内死亡(图2a)。相反,接受血栓预防的低分子量肝素(lowmolecularweightheparin)(lmwh)>75%wt小鼠存活(图2a)。因此,与lmwh相比,ha不能保护小鼠免于tf引起的血栓栓塞。然后,我们在同一模型中研究了f8-/-pros1+/+,f8-/-pros1+/-和f8-/-pros1-/-小鼠。输注tf(约2.1nm)后,有40-60%的小鼠死亡(p>0.05),而与它们的pros1基因型无关(图2b)。但是,存在f8-/-pros1-/-和f8-/-pros1+/-小鼠早于f8-/-pros1+/+小鼠死亡的趋势,和f8-/-pros1+/-早于f8-/-pros1+/+小鼠死亡的趋势(平均死亡时间:f8-/-pros1+/+为12±4分钟,f8-/-pros1+/-为7±2分钟,f8-/-pros1-/-为8±3分钟,n=4-6/组,p=0.43)。用f9-/-pros1+/+,f9-/-pros1+/-和f9-/-pros1-/-小鼠获得了相似的数据(数据未显示)。
在f8-/-pros1+/+和f8-/-pros1-/-小鼠的肺动脉中检测到了纤维蛋白凝块,它们在tf诱导的血栓栓塞攻击(challenge)期间死亡(图2c)。重要的是,来自f8-/-pros1-/-的小鼠的血栓比来自f8-/-pros1+/+的小鼠的多(分别为n=48和26)。此外,f8-/-pros1-/-肺中的大多数动脉被完全闭塞,而在f8-/-pros1+/+肺中仅部分被闭塞。
在tf诱导的血栓栓塞性攻击期间死亡的f8-/-pros1-/-小鼠均未出现暴发性紫癜。在f9-/-pros1+/+,f9-/-pros1+/-和f9-/-pros1-/-小鼠获得了相似的数据(未显示)。
实施例3:fviii的丢失部分保护pros1-/-小鼠免受肠系膜小动脉血栓形成的影响
然后,我们记录了肠系膜小动脉中的血栓形成,该模型对内在凝血途径的缺陷敏感。在f8+/+pros1+/+小鼠中,血栓在20分钟内增长到闭塞大小,所有受伤的小动脉均被闭塞(图2d)。不出所料,f8-/-pros1+/+的小动脉均未显示血栓形成,而f8-/-pros1-/-小鼠显示部分血栓形成(图2d)。
在f8+/+pros1+/+小鼠血栓形成过程中产生栓塞物(emboli),但在f8-/-pros1+/+小鼠中不会。在f8-/-pros1-/-小鼠中,多个微栓塞物在部分血栓生长过程中脱落,阻止了闭塞性血栓的形成。
实施例4:pros1靶向限制但不能消除ha小鼠的尾端出血
使用轻度或重度出血模型通过尾部横断评估出血表型。
在两个模型中,与f8-/-pros1+/+小鼠相比,f8-/-pros1-/-的失血量减少(图3a-b)。当受到轻度模型的攻击时,f8-/-pros1+/-小鼠的出血少于f8-/-pros1+/+小鼠(图3a)。相反,当暴露于严重模型时,f8-/-pros1-/-和f8-/-pros1+/-小鼠表现出相当的失血量(图3b)。但是,在两个模型中,f8-/-pros1-/-小鼠的出血均超过f8+/-pros1+/+和f8+/+pros1+/+小鼠(图3a-b),表明f8-/-小鼠中pros1的丢失仅能部分纠正出血表型。
然后,使用ps中和抗体来研究ps活性的抑制如何改变f8-/-pros1+/-小鼠的尾端出血。该抗体将f8-/-pros1+/-小鼠(图3c)的血液流失限制在与pros1完全基因丢失的程度相同(图3b)。
实施例5:pros1靶向或ps抑制完全保护ha或hb小鼠免于急性关节积血形成(ah)
尽管出血可能出现在血友病患者的任何地方,但是大多数出血发生在关节处。为了确定pros1缺失是否可以预防血友病小鼠的关节积血形成,我们将ah模型应用于f8-/-pros1+/+、f8-/-pros1+/-、f8-/-pros1-/-和f8+/+pros1+/+小鼠。与f8-/-pros1+/+和f8-/-pros1+/-小鼠相比,f8-/-pros1-/-和f8+/+pros1+/+小鼠受伤后的膝盖肿胀降低了(图4a)。f8-/-pros1-/-和f8+/+pros1+/+小鼠之间的膝盖肿胀也没有差异(图4a)。在f8-/-pros1+/+(ibs=2,n=5)的关节腔和滑膜中观察到出血,但在f8-/-pros1-/-(ibs=0,n=5)和f8+/+pros1+/+小鼠(ibs=0,n=5)的关节腔和滑膜中未观察到出血(图4b)。与f8-/-pros1-/-和f8+/+pros1+/+小鼠相比,f8-/-pros1+/+的关节腔和滑膜中的纤维蛋白更多(图4b)。用f9-/-pros1+/+和f9-/-pros1-/-小鼠获得了相似的数据(分别为ibs=0,n=3和ibs=2,n=3)(图9a-b)。
通过在f8-/-pros1+/-小鼠中(在ah诱导前1天开始)连续4天ps中和抗体或对照抗体皮下输注(ps中和抗体组的膝关节肿胀为0.43±0.07相对于对照组的0.69±0.09mm,n=9,p=0.04)证实了这些结果。ps中和抗体组的ps血浆水平为26±6%,而对照组为45±3%(n=5,p=0.017)。另外,在对f8-/-pros1+/-和f8-/-pros1+/+小鼠的ah攻击之前,通过静脉内注射鼠ps(mps)sirna替代性地实现了ps抑制(图4c-d)。ibs评估证实,与对照组sirna(ibs=2,n=3)相比,mpssirna(ibs=0.5,n=3)处理的f8-/-pros1+/+小鼠缺乏关节腔内出血,图4c)。重要的是,在血浆中(26±3%,对照组中为84±11%,n=3,p=0.006)和滑膜中的ps表达均被mpssirna降低(图5a)。
实施例6:ps和tfpi都在小鼠滑膜中表达
为了了解血友病小鼠中,pros1基因缺失和ps抑制在关节内止血的突出作用,对膝盖切片进行了ps和tfpi免疫染色。ps主要出现在用对照sirna处理的ah的f8-/-pros1+/+小鼠的滑膜组织的衬里层中,而ps的滑膜染色在接受mps的ah的f8-/-pros1+/+小鼠中明显减少sirna(图5a)。相反,在接受mpssirna的血友病小鼠的滑膜组织中,tfpi染色比对照sirna处理的小鼠滑膜组织更显著(图5a)。但是,tfpi表达在f8-/-pros1+/+和f8-/-pros1-/-小鼠的滑膜衬里层中具有可比性(图5b)。
为了进一步证明ps由成纤维样滑膜细胞(fls)表达,我们在从f8+/+pros1+/+、f8-/-pros1+/+和f8-/-pros1-/-fls收集的条件培养基上进行了蛋白质印迹。如图5c所示,f8+/+pros1+/+和f8-/-pros1+/+fls培养基在约75kda的分子量处具有与ps相当的条带,与血浆和血小板中观察到的相似。如预期的那样,在从f8+/+pros1-/-fls获得的培养基中未检测到染色(图5c)。
我们还研究了f8-/-pros1+/+和f8-/-pros1-/-fls条件培养基中的tfpi表达(图5d)。所有培养基均显示约50kda的条带,与胎盘裂解液观察到的相似。在蛋白质去糖基化之后研究了tfpi同种型的表达,因为完全糖基化的tfpiα和tfpiβ在相同的分子量下迁移30。来自fls培养基的去糖基化的tfpi以tfpiα的分子量单条带的形式迁移,其tfpiα的分子量类似于胎盘tfpi(tfpiα的阳性对照)(图5d)。这表明fls表达tfpiα,但不表达tfpiβ。此外,在凝血酶刺激后,f8-/-pros1+/+fls中的ps和tfpi表达增加(图5e-f)。
实施例7:ps和tfpi均在ha或hb患者的滑膜中表达
之后对人ha,hb和骨关节炎膝滑膜组织的ps和tfpi进行了分析(图6a)。在ha患者的滑膜衬里层和亚衬层中发现了tfpi和ps的按需(ondemand)强烈信号(n=7)。相比之下,在预防下的ha患者中ps和tfpi的免疫染色均降低了(n=5)。与按需ha患者相比,按需hb患者在滑膜衬里层和亚衬层(n=4)中所显示的ps和tfpi信号更少。骨关节炎患者(n=7)的切片与预防性血友病患者相似,未显示tfpi和ps强烈染色。为了评估人fls表达哪种tfpi的同种型,在从健康受试者和骨关节炎患者分离的人fls的条件培养基上进行了蛋白质印迹分析。与鼠fls相似,人fls表达tfpiα,但不表达tfpiβ(图6b)。
实施例8:pros1的丧失是造成ha小鼠缺乏tfpi依赖的ps活性和对apc的抵抗力的原因
在缺乏pros1或ps被抑制的ha或hb小鼠中,针对ah的全面保护至少可以部分解释为关节中缺乏针对apc和tfpi的ps辅因子活性。但是,在尾端出血攻击模型中,缺乏pros1或ps抑制的ha小鼠中部分止血作用的原因需要进一步研究。
体外tf引发的凝血酶生成测试表明,血浆生成凝血酶的能力与血友病的临床严重程度之间存在相关性。因此,我们研究了pros1丢失对ha小鼠血浆中凝血酶生成的影响。在无血小板血浆(platelet-freeplasma)(pfp)中未评估tfpi依赖性ps活性,但在富含血小板的血浆(platelet-richplasma)(prp)中评估了tfpi依赖性ps活性,因为无法借助凝血酶生成测试在小鼠血浆中证明ps的tfpi辅因子活性。这可以通过小鼠血浆中tfpiα的缺乏及其在小鼠血小板中的存在来解释。
响应1pmtf时,f8-/-pros1-/-的prp的凝血酶峰和内源性凝血酶电位(etp)均显著高于f8-/-pros1+/+(1072±160vs590±10nmol/l分钟,n=3/组,p=0.04),表明在f8-/-pros1-/-的prp中缺乏pstfpi-辅因子活性(图7a)。与以前的工作一致,在存在1、2.5或5pmtf的情况下,f8-/-pros1+/+和f8-/-pros1-/-小鼠的pfp中的凝血酶峰和etp均相当(数据未显示)。
为了评估f8-/-pros1-/-小鼠是否表现出有缺陷的功能性apc依赖性ps活性,我们在不存在apc、存在野生型(wt)重组apc或存在突变(l38d)重组小鼠apc(l38dapc,具有ps辅助因子活性降低(ablated)的变体)的情况下,在ca2+离子载体激活的prp中进行了凝血酶生成测试。在该测定中,apc滴定表明,添加8nmwtapc能够在wt小鼠的活化prp中将etp降低90%,而相同浓度的l38dapc仅使etp降低30%(数据未显示)。基于这些数据,记录了激活的prp的凝血酶生成曲线(3只小鼠/测定)。计算得出的apc比率(etp+apcwt/etp+apcl38d)表明f8-/-pros1-/-血浆中存在apc抗性,但f8-/-pros1+/+血浆中没有(分别为0.87±0.13、0.23±0.08,p=0.01)(图7b)。
在存在2nmwtapc和l38dapc的情况下,还在f8-/-pros1+/+和f8-/-pros1-/-小鼠(2只小鼠/试验)的pfp中测试了apc依赖性ps活性。计算的apc比率显示f8-/-pros1-/-小鼠中的apc抗性,但在f8-/-pros1+/+小鼠中没有(分别为1.08±0.04,0.25±0.09,p=0.0003)(图7b)。
实施例9:缺乏pros1小鼠的ha小鼠中纤维蛋白网络的改善
尾端出血的小鼠模型不仅对血小板功能异常敏感,而且对凝血和纤维蛋白溶解改变也很敏感。为了了解在已研究过基因型之间的尾端出血的差异,我们使用扫描电子显微镜成像研究纤维蛋白结构(图7c)。与f8-/-pros1+/+血浆凝块相比,f8+/+pros1+/+和f8-/-pros1-/-血浆的凝块显示出密集的高度分支的纤维蛋白纤维网络(图11a-b)。相比之下,来自f9+/+pros1+/+和f9-/-pros1-/-血浆的凝块没有显示出比f9-/-pros1+/+血浆凝块更密集的网络,而是纤维分支增加的趋势(图11c-d)。
与来自f8+/+pros1+/+小鼠或f9+/+pros1+/+小鼠的纤维相比,来自f8-/-pros1-/-和f8-/-pros1+/+小鼠以及f9-/-pros1-/-和f9-/-pros1+/+小鼠的纤维蛋白纤维显示出更大的直径。尽管如此,与f8-/-pros1+/+或f9-/-pros1+/+小鼠相比,f8-/-pros1-/-和f9-/-pros1-/-小鼠的纤维表面分别显示出较少的孔隙率,这表明:与源自f8-/-pros1+/+或f9-/-pros1+/+的纤维相比,源自f8-/-pros1-/-和f9-/-pros1-/-的纤维可能渗透性较低,因此对纤维蛋白溶解的抵抗力更高38。
这些数据补充了tfpi和apc辅助因子的活性结果(图7a-b),有助于解释为什么与f8-/-pros1+/+小鼠相比,f8-/-pros1-/-的尾端出血得以改善,但是没有像f8+/+pros1+/+小鼠中那样完全纠正。
实施例10:血浆中的ps抑制恢复了ha患者的凝血酶生成
然后,我们检查了ps抑制对人ha血浆中凝血酶生成的影响。在存在ps中和抗体的情况下,pfp中的etp增加2-4倍。使用针对c末端域的抗人tfpi抗体,即使在存在fviii抑制剂的情况下,也可以有效抑制fxa,获得相似的结果(图7d-e)。ps抑制在prp样品中具有显著效果,在prp样品中,其etp增加10倍以上(1912±37和1872±64nm*)(分别为图7f和g)。因此,ps抑制作用完全恢复了血友病血浆中的etp(相比之下,正常血浆中的etp:1495±2nm*min)。使用抗tfpi抗体获得了相似的结果(图7d-g)。这些数据在人类中证实了我们在小鼠中观察到的ps抑制驱动的hapfp和prp中凝血酶生成的改善。
实施例11:材料和方法
小鼠
具有c57bl/6j背景的f8-/-小鼠(b6;129s4-f8tm1kaz/j)和f9-/-小鼠(b6.129p2-f9tm1dws/j)从jackson实验室获得。pros1+/-小鼠是原始群落15的后代。瑞士联邦兽医局批准了实验。如所述对小鼠进行基因分型15-17。
tf诱导的肺栓塞
静脉血栓栓塞模型修改自weiss等18,并做微小调整15。6-9周龄的麻醉小鼠以4.25nm(1:2稀释度)或2.1nm(1:4稀释度)静脉内注射(2μl/g)人重组tf(hrtf,dadeinnovin,siemens)。呼吸停止发作后两分钟或20分钟观察期完成后,收集肺并将其固定在4%pfa中。肺切片用苏木精和曙红对纤维蛋白染色。肺中纤维蛋白凝块的程度以随机选择的10个非重叠区域(×10放大倍数)中血管内血栓的数量进行评估。
ha小鼠的断尾模型
如所描述的14,评估了两种不同的断尾模型以评估出血表型。简而言之,将8-10周龄小鼠的尾巴末端横断2mm(轻度损伤),出血为静脉或横断4mm(重度损伤),出血为动脉和静脉19。将出血定量为分别在30或10分钟后的失血量。在严重损伤模型中,一些f8-/-pros1+/-小鼠在尾横切前2分钟以2.1mg/kg的剂量静脉内接受了兔抗人ps-igg(dako)或兔同型(isotype)igg(r&dsystems)。
急性关节积血形成模型
根据
体内ps抑制
10周大的小鼠通过皮下渗透微型泵(型号2001,alzet),以1mg/kg/天的速度连续输注兔抗人ps-igg(dakobasel,瑞士)或兔同型igg(r&dsystems)。
或者,通过转染剂(invivofectamine3.0,invitrogen,lifetechnologies)将单剂量小鼠特异性sirna(s72206,lifetechnologies)或对照sirna(4459405,invivonegativecontrol#1ambion,lifetechnologies)以1mg/kg处理10周龄的小鼠,按照制造商的说明进行操作。ps抑制后2.5天,使用急性关节积血模型。
统计方法
值表示为平均值±标准误。非关联基因位点(non-linkedgeneticloci)的卡方(chi-square)用于评估孟德尔等位基因的分离。使用kaplan-meier方法绘制tf诱导的静脉血栓栓塞模型中的生存数据。使用对数秩(log-rank)检验对曲线进行统计比较(prism6.0d;graphpad)。其他数据通过graphpadprism6.0d的t检验,单向和双向anova试验进行分析。p值小于0.05认为具有统计学意义。
鼠血浆的制备
用戊巴比妥(40mg/kg)麻醉6-9周龄的小鼠,将全血从下腔静脉抽取到3.13%柠檬酸盐中(1体积抗凝剂/9体积血液)。将离心机预热至26℃,将血液以1031g离心10分钟,以获得富含血小板的血浆(prp)。其它血液在室温(rt)下以2400g离心10分钟,以获得乏血小板血浆(platelet-poorplasma)(ppp)。为了获得无血小板血浆(pfp),在10000g下另外离心10分钟。
血小板计数和凝血参数的测量
用自动细胞计数器(procytedxhematologyanalyzer,idexx)进行血小板计数。在自动sysmexca-7000凝血分析仪(sysmexdigitana)上测量纤维蛋白原,fviii和fix的活性。在凝血计(mc4plus,merlinmedical)上测量凝血酶原时间(pt)和活化的部分凝血活酶时间(aptt)。
elisa法测定鼠ps抗原和tat复合物
将来自96孔板(maxisorb,thermo)的孔用每孔50μl的10μg/ml兔多克隆抗人ps(dakocytomation)的包被,并在4℃下孵育过夜。用tbs缓冲液(0.05m3-(羟甲基)氨基甲烷,0.15mnacl,ph7.5、0.05%吐温20)洗涤3次后,用2%tbs-bsa封闭板。将稀释的血浆样品(稀释范围:1:300-1:600)添加至孔中,并在室温下孵育2小时。洗涤3次后,加入50μl的1μg/ml生物素化的鸡多克隆抗鼠蛋白s,在室温下孵育2小时。加入链霉亲和素-hrp偶联的辣根过氧化物酶(thermo)扩增信号,并将板孵育1h。将板洗涤3次,并添加100μl的tmb底物(kpl)。加入100μlhcl(1m)终止反应。在450nm处测量吸光度。使用连续稀释的合并(pooled)正常血浆稀释液建立标准曲线,该血浆来自14只健康小鼠(7-12周龄,雄性8只、雌性6只)。结果以相对于合并的正常血浆的百分比表示。
根据制造商的说明,使用市售的elisa(enzygnosttatmicro,siemens),对每个血浆样品的tat水平进行一式两份的测量。
小鼠组织加工和切片,免疫组织化学和显微镜检查
将未经预处理的组织切片(4μm)用苏木精/曙红或masson三色染色,或对不溶性纤维蛋白ps或tfpi,进行免疫染色。使用了以下抗体:纤维蛋白(mab克隆102-10)1终浓度15.6μg/ml,在室温下孵育30分钟,兔抗人二抗(ab7155abcam,剑桥,英国),1:200稀释,在室温下孵育30分钟;ps(mab4976,r&d,稀释度1:50)在室温下孵育30分钟,兔抗大鼠二抗(ab7155abcam)1:200稀释,在室温下孵育30分钟;tfpi(pahtfpi-s,hematologicaltechnologies)终浓度为18.6μg/ml,在室温下孵育30分钟,将兔抗绵羊igg二抗(ab7106,abcam)1:200稀释,在室温下孵育30分钟。按照制造商的说明,所有染色均使用免疫染色剂bondrx(leicabiosystems,muttenz,瑞士)进行。使用3dhistechpanoramic250flashii扫描整个玻片,使用20倍(na0.8),40倍(na0.95)的空气物镜。使用panoramicviewer软件进行图像处理。
将fviii体内给药至具有完全f8基因缺失的小鼠
将6至9周龄的小鼠用氯胺酮(ketamine)(80mg/kg)和甲苯噻嗪(xylazine)(16mg/kg)麻醉。我们静脉给予0.3u/kg的重组fviii(
肠系膜动脉中的fecl3损伤性血栓形成模型
根据参考文献2并进行了微小改动,使用活体显微镜对肠系膜动脉血栓形成模型进行了研究。通过腹膜内注射氯胺酮(80mg/kg)和甲苯噻嗪(16mg/kg)的混合物麻醉小鼠。通过注射100μl罗丹明6g(rhodamine6g)(1.0mm)在体内直接标记血小板。在选择了研究的区域后,通过用局部用10%fecl3饱和的滤纸(来自于1mwhatmann纸的直径为1毫米的补丁纸)局部使用1分钟来产生血管壁损伤。在配备有亲和力校正的水浸泡光学元件(zeiss,德国)的fitc滤光片组的荧光显微镜(iv-500,microninstruments,sandiego,ca)下实时监测血栓形成。亮荧光标记的血小板和白细胞可以通过视频触发的频闪落射式落射照明(stroboscopicepi-illumination)(chadwickhelmuth,elmonte,ca)观察到1355μmx965μm的视野。使用了带有na0.3的10倍物镜zeissplan-neofluar。所有场景均使用定制的低延迟硅增强目标相机(dagemti,michigancity,in),时基发生器和hi-8vcr(ev,c-100,索尼,日本)在录像带上录制。测量到血管壁阻塞的时间,这由血细胞流的停止确定。
成纤维样滑膜细胞(fls)分离,培养和流式细胞仪
分离并培养8-10周龄小鼠的鼠fls3。三代后,拍摄细胞的相衬图像,并将细胞与fitc缀合的大鼠抗小鼠cd11b抗体(m1/70,pharmingen,bdbiosciences),pe缀合的大鼠抗小鼠cd90.2抗体(30-h12,pharmingen,bdbiosciences),fitc缀合的大鼠抗小鼠cd106抗体(429mvcam.a,pharmingen,bdbiosciences),pe缀合的仓鼠抗小鼠cd54抗体(3e2,pharmingen,bdbiosciences)和荧光染料缀合的同型对照抗体在黑暗中于4℃孵育30分钟。在最后的洗涤和离心步骤之后,在lsrii流式细胞仪(bdbiosciences)和facsdiva7.0软件(bdbiosciences)上分析所有孵育的细胞。来自健康个体和oa患者的人fls购自bioscience,asterand,并根据制造说明进行培养。
蛋白质印迹
在还原条件下,通过十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(12%梯度sds-page,bio-rad)在人和小鼠样品中检测ps和tfpi。将蛋白质转移到硝酸纤维素膜(bio-rad)上,然后使用以下方式进行可视化:用于鼠ps的2μg/ml单克隆mab-4976(r&d系统),用于鼠tfpi(r&dsystem)的1μg/ml多克隆af2975。重组鼠ps4(30ng),全长重组人tfpi(由t.hamuro,kaketsuken,japan提供),洗涤过的血小板裂解物,来自f8-/-pros1+/+小鼠的pfp和来自f8+/+pros1+/+小鼠的胎盘裂解物用作ps,tfpiα作为对照。在无血清培养基(optimem)中孵育24小时后,从融合的鼠和人类fls条件培养基中收集样品,并使用amicon过滤器(millipore,截断值为10kda)浓缩40倍。对于tfpi蛋白质印迹,样品被加载到凝胶上之前,将该样品用五种蛋白质去糖苷酶的混合物(pngasef,o-糖苷酶,神经氨酸苷酶,β1-4半乳糖苷酶,β-n-乙酰氨基葡萄糖苷酶,去糖基化试剂盒,v4931,promega)在37℃下处理12小时。最终检测通过使用辣根过氧化物酶偶联的第二抗体(dako)和supersignalwestdura延长化学发光底物(pierce)进行,并由fujilas3000irccd相机进行监控。
人体膝关节滑膜的免疫组织化学
从佛罗伦萨大学实验与临床医学系解剖学和组织学科的档案中,收集了由于严重膝关节病而经历了关节置换术的12例ha患者和4例hb患者的石蜡包埋的滑膜组织样本,如在其它地方描述的那样5,6。7例ha患者进行按需治疗,另外5例进行二级预防。所有4名hb患者均按需治疗。来自7名骨关节炎(oa)患者的滑膜样品用作对照5,6。为了进行免疫组织化学分析,将滑膜组织切片(5μm厚)去石蜡,再水化,在柠檬酸钠缓冲液(10mm,ph6.0)中煮沸10分钟以回收抗原,然后在室温下用3%h2o2的甲醇溶液处理15分钟,阻止内源性过氧化物酶活性。然后将切片在pbs中洗涤,并根据制造商的规程,在室温下与ultrav阻滞剂(ultravision大容量检测系统抗多价抗体,hrp,目录号tp-125-hl,labvision)一起孵育10分钟。阻断非特异性位点结合后,将切片与在pbs中稀释的兔多克隆抗人蛋白s/pros1抗体(1:50稀释,目录号nbp1-87218,novusbiologics)或绵羊多克隆抗人组织因子途径抑制剂(tfpi)抗体(1:500稀释,目录号pahtfpi-s,haematologictechnologies)在4℃孵育过夜。对于ps免疫染色,然后根据制造商的规程,将组织切片依次与生物素化的第二抗体,链霉亲和素过氧化物酶(ultravision大容量检测系统抗多价抗体,hrp;labvision)进行孵育。为了进行tfpi免疫染色,将组织切片与结合了hrp的驴抗绵羊igg(1:1000稀释;目录号ab97125;abcam)孵育30分钟。使用3-氨基-9-乙基咔唑(aec试剂盒,目录号ta-125-sa;labvision)作为显色剂激发了免疫反应性。最后,将滑膜切片用mayer的苏木精(bio-optica)复染色,清洗,封固在水性封固剂中,并在leicadm4000b显微镜(leicamicrosystems)下观察。未暴露于一抗或未与同型匹配和浓度匹配的非免疫igg(sigma-aldrich)孵育的切片作为抗体特异性的阴性对照。用配备有leica软件应用程序套件lasv3.8(leicamicrosystems)的leicadfc310fx1.4百万像素数码彩色相机捕获光学显微镜图像。
纤维蛋白凝块超微结构研究
在37℃下,通过添加约5nmtf(dadeinnovin,siemens)从pfp制备纤维蛋白凝块。据zubairova等7,然后将它们固定在2%的戊二醛中,脱水,干燥并用金钯喷镀,以便使用扫描电子显微镜进行可视化。使用stepanizer软件(www.stepanizer.com)对网络密度和纤维分支进行了半定量评估。
鼠标本中的校准自动血栓(thrombography)分析
使用校准的自动血栓图(calibratedautomatedthrombogram)(cat)方法确定pfp和prp中的凝血酶生成。
在prp(150g/l)中评估tfpi依赖的ps活性,如下所示。简而言之,将10μl小鼠prp(150g/l),10μlprp试剂(diagnosticastago)和30μl缓冲液a(25mmhepes,175mmnacl,ph7.4,5mg/mlbsa)混合。凝血酶的产生是在37℃下用10μl荧光底物/cacl2混合物开始的。最终浓度如下:16.6%小鼠血浆,1pmhrtf,4μm磷脂,16mmcacl2和0.42mm荧光底物。
根据dargaudy等8,在小鼠的pfp和prp中,基于cat的apc抗性测试中评估了apc依赖的ps活性。预先使用40μmca2+离子载体(a23187)在37℃激活prp(150g/l)5分钟。终浓度如下:16.6%小鼠血浆,22μma23187、1pmhrtf,4μm磷脂,2nm(对于pfp)或8nm(对于prp)野生型重组小鼠apc(wt-rmapc)5或突变的重组小鼠apc(rmapcl38d),16mmcacl2和0.42mm荧光底物。根据参考文献9,10进行rmapcl38d的生成和表征,根据参考文献11,12进行纯化。
对于在pfp上进行tf滴定,使用了以下试剂:ppp试剂和mp试剂(diagnosticastago)。
使用配备有分配器的fluoroscanascent荧光计测量荧光。在390nm(激发滤光片)和460nm(发射滤光片)的波长处检测荧光强度。专用软件程序,
人体样品中的cat分析
从患者获得书面知情同意。通过在静脉穿刺抽出静脉血置于3.2%柠檬酸钠(体积/体积)中,并以2000g离心5分钟。然后将乏血小板血浆(ppp)以10000g离心10分钟,以获得pfp。将pfp等分,速冻,并保存在-80℃直至使用。对于prp,将血液以180gx10分钟的速度离心。所有受试者均获得知情同意参加。根据参考文献13,并有微小变化,在人类pfp和prp中评估了凝血酶的产生。简而言之,将68μlpfp或prp(150g/l)与12μl多克隆兔抗人ps-igg抗体(0.42mg/ml,dako)或针对tfpi的单克隆抗体或缓冲液a,在37℃孵育15分钟(0.66μm,mw1848,sanquin)。对于pfp样品,使用20μlppp低和ppp5pm试剂(diagnosticastago)的7:1混合物(对于pfp样品)或prp试剂(diagnosticastago)对prp样品进行混凝。加入20μlcacl2和荧光底物(i-1140;bachem)后,在fluoroskanascent阅读器(thermolabsystems)中跟踪凝血酶的产生。
讨论
ps是凝血酶生成的关键调节因子,我们认为靶向ps可能构成血友病的潜在疗法。
在小鼠中进行的广泛研究提供了概念数据证明,证明了ps和tfpi在导致血友病小鼠出血和严重关节损伤中起着核心作用。靶向pros1或抑制ps具有改善小鼠血友病的能力,这可以通过体内尾端出血试验中出血表型的改善,以及对关节积血形成的全面保护来判断(图3a-c和4)。由于关节表现出非常弱的tf表达,而滑膜细胞产生大量的tfpiα和ps(图5),因此关节内的外源性凝血途径的活性大大降低,易于诱发血友病关节出血。此外,血栓调节蛋白(thrombomodulin)(tm)和内皮蛋白c受体(epcr)均通过fls表达,这表明在ah的情况下,tm-凝血酶复合物激活epcr结合的pc以产生非常有效的抗凝剂apc。重要的是,tfpiα的表达被凝血酶上调(图5f)。因此,通常导致明显的局部炎症和关节症状的ah可持续数天至数周,还促进了多种抗凝剂,即apc,tfpiα及其相互辅因子ps的局部生成和分泌,这有助于解释血友病中关节损伤的病理生理学。
使用来自血友病患者的临床样本进行的观察与从鼠类研究中发现的内容是一致的。在人类中,有或没有抑制剂的ha患者的血浆中的ps阻断都能使etp正常化(图7d-g)。hb患者的tfpi和ps的关节内表达少于ha患者,这与目前hb患者的出血少于ha患者的出血的认知一致(图6)。此外,在考虑出血的情况下,接受预防治疗的ha患者显示的tfpi和ps滑膜表达少于接受fviii浓缩物的患者(即所谓的“按需治疗”)(图6a)。最后,与在小鼠中观察到的一致,人类fls分泌tfpiα和ps,从而加强了鼠类血友病数据对人类的推断。
本报告中的广泛发现使我们建议靶向ps可能会转化为对血友病有用的疗法。人和鼠关节中的ps是关节积血的新型病理生理贡献者,并且构成了诱人的潜在治疗靶标,特别是因为它对关节内的apc和tfpiα具有双重辅助因子活性。在ps的存在下,关节积血会增加滑膜中tfpiα的表达。在小鼠中靶向ps可以保护它们免于关节积血。因此,我们认为,由fls产生的tfpiα及其辅因子ps,连同tm-epcr-pc途径,都构成了一种有效的关节内抗凝系统,对关节积血具有重要的病理影响。已成功用于血友病小鼠的鼠ps沉默rna(图4h-1和图5a)是我们将为血友病患者开发的一种治疗方法。使rna沉默而不是现有的因子替代疗法的优势在于半衰期更长,从而减少了注射频率及其可能的皮下给药途径。
参考文献
1.hisaday,yasunagam,hanaokas,etal.discoveryofanuncoveredregioninfibrinclotsanditsclinicalsignificance.scirep.2013;3:2604.
2.angelillo-scherrera,fontanap,burnierl,etal.connexin37limitsthrombuspropensitybydownregulatingplateletreactivity.circulation.2011;124(8):930-939.
3.armakam,vassilikig,kontoyiannisd,kolliasg.astandardizedprotocolfortheisolationandcultureofnormalandarthritogenicmurinesynovialfibroblastsprotocolexchange.2009.
4.fernandezja,heebmj,xux,singhi,zlokovicbv,griffinjh.species-specificanticoagulantandmitogenicactivitiesofmurineproteins.haematologica.2009;94(12):1721-1731.
5.melchiorred,linaris,manettim,etal.clinical,instrumental,serologicalandhistologicalfindingssuggestthathemophiliabmaybelessseverethanhemophiliaa.haematologica.2016;101(2):219-225.
6.melchiorred,miliaaf,linaris,etal.rank-rankl-opginhemophilicarthropathy:fromclinicalandimagingdiagnosistohistopathology.jrheumatol.2012;39(8):1678-1686.
7.zubairovald,nabiullinarm,nagaswamic,etal.circulatingmicroparticlesalterformation,structure,andpropertiesoffibrinclots.scirep.2015;5:17611.
8.dargaudy,luddingtonr,graye,etal.standardisationofthrombingenerationtest--whichreferenceplasmafortgt?aninternationalmulticentrestudy.thrombres.2010;125(4):353-356.
9.harmons,prestonrj,niainlef,etal.dissociationofactivatedproteincfunctionsbyeliminationofproteinscofactorenhancement.jbiolchem.2008;283(45):30531-30539.
10.prestonrj,ajznere,razzaric,etal.multifunctionalspecificityoftheproteinc/activatedproteincgladomain.jbiolchem.2006;281(39):28850-28857.
11.burnierl,fernandezja,griffinjh.antibodyspc-54providesacuteinvivoblockageofthemurineproteincsystem.bloodcellsmoldis.2013;50(4):252-258.
12.mosnierlo,yangxv,griffinjh.activatedproteincmutantwithminimalanticoagulantactivity,normalcytoprotectiveactivity,andpreservationofthrombinactivablefibrinolysisinhibitor-dependentcytoprotectivefunctions.jbiolchem.2007;282(45):33022-33033.
13.maurissenlf,castoldie,simionip,rosingj,hackengtm.thrombingeneration-basedassaystomeasuretheactivityofthetfpi-proteinspathwayinplasmafromnormalandproteins-deficientindividuals.jthrombhaemost.2010;8(4):750-758.
14.polderdijksg,adamste,ivanciul,camirerm,baglintp,huntingtonja.designandcharacterizationofanapc-specificserpinforthetreatmentofhemophilia.blood.2016.
15.sallerf,brissetac,tchaikovskisn,etal.generationandphenotypicanalysisofproteins-deficientmice.blood.2009;114(11):2307-2314.
16.bil,lawleram,antonarakisse,highka,gearhartjd,kazazianhh,jr.targeteddisruptionofthemousefactorviiigeneproducesamodelofhaemophiliaa.natgenet.1995;10(1):119-121.
17.linhf,maedan,smithieso,straightdl,stafforddw.acoagulationfactorix-deficientmousemodelforhumanhemophiliab.blood.1997;90(10):3962-3966.
18.weissej,hamiltonjr,leaseke,coughlinsr.protectionagainstthrombosisinmicelackingpar3.blood.2002;100(9):3240-3244.
- 还没有人留言评论。精彩留言会获得点赞!