将核酸递送至耳蜗和前庭细胞的材料和方法与流程
相关申请的交叉引用
本申请主张2017年2月6日提交的美国临时申请no.62/455,197的优先权和权益,该临时申请的公开内容通过引用而整体并入本文。
背景技术:
遗传性听力损失是一个显著的问题,且鲜有除人工耳蜗以外的治疗选择。先天性听力问题往往由单基因缺陷造成。每500个婴儿中诊断出1例语前聋,其中大约50%具有遗传病因。乌谢尔综合征(ushersyndrome)与大量不同的临床亚型相关,每一种亚型均可由大量不同基因中任一个的突变造成,乌谢尔综合征是3%至6%的儿童早期耳聋的原因。被评估为所有遗传性耳聋的1%至2%的一种更普遍的基因缺陷,出现在tmc1基因中。最严重的乌谢尔综合征形式,ush1,与下述六种基因的缺陷相关:ush1、myo7a(肌球蛋白(myosin)7a)、ush1c(harmonin)、cdh23(钙粘附蛋白(cadherin23)、pcdh15(原钙粘蛋白(protocadherin)15)、sans(sans;也称为ush1g)和cib2(钙整合素结合蛋白2)。
内耳,例如耳蜗,尤其是耳蜗中的内毛细胞和外毛细胞(ihc和ohc),对于用以干预多种病因的听力损失和耳聋、最直接的单基因形式的先天性耳聋的多核苷酸治疗途径来说,是有吸引力的目标。但是,有效地以ihc和ohc及其它可能与基因治疗途径相关的内耳细胞为靶点并转到这些细胞是个挑战。
技术实现要素:
本发明提供以受试者的内耳细胞(例如,内毛细胞或外毛细胞)为靶点并表达编码所关注的多肽(例如,tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c))的转基因的组合物和方法。在一个具体实施例中,将内耳毛细胞靶向的aav给药至在听觉和/或前庭机械感受中具有基因缺陷的受试者的内耳。
如本文所示,含有被称为anc80或特异性anc80衣壳蛋白(例如,anc80-0065)的祖先骨架衣壳蛋白(ancestralscaffoldcapsidprotein)的腺相关病毒(aav),有效地以内耳中的多种细胞诸如体内的ihc和ohc为靶点。
由于一种或多种类型的细胞(例如外毛细胞)的限制性转导,将基因转移至动物模型的内耳细胞的效力有限。但是,本文所述的新颖基因递送方式包括基于含有anc80的衣壳蛋白aav的新组合物和方法,其提供以包括ihc和ohc两者在内的内耳细胞为目标的高效基因转移。
一方面,本发明提供内耳毛细胞靶向的合成腺相关病毒(aav)载体,其中该载体含有编码人ush1多肽的多核苷酸,其中该ush1多肽是肌球蛋白7a、harmonin(例如,harmonin-a、harmonin-b或harmonin-c)、钙粘蛋白23、原钙粘蛋白15、sans和钙整合素结合蛋白2、或本文所述的任何其它多肽。
另一方面,本发明提供内耳毛细胞靶向的合成腺相关病毒(aav)载体,其中该载体编码与anc80l65的序列一致性为至少约85%的衣壳,且含有引导人tmc1或tmc2多核苷酸表达的启动子。
另一方面,本发明提供内耳毛细胞靶向的合成腺相关病毒(aav)载体,其中该载体含有引导下游多核苷酸表达的启动子,该启动子是espin启动子、pcdh15启动子、ptprq启动子或tmhs(lhfpl5)启动子。
另一方面,本发明提供一种细胞,该细胞含有前述方面所述的内耳毛细胞靶向的合成腺相关病毒(aav)载体。
另一方面,本发明提供表达受试者的内耳中的多肽的方法,该方法涉及令内耳细胞与编码所关注多肽且内耳毛细胞靶向的合成腺相关病毒(aav)载体接触,其中该aav载体转染该内耳细胞的至少约70%且含有祖先aav序列。
另一方面,本发明提供表达受试者的内耳中的多肽的方法,该方法涉及令内耳细胞与编码所关注的人多肽的合成腺相关病毒(aav)载体接触,其中该aav载体编码与anc80l65的序列一致性为至少约85%的衣壳。
另一方面,本发明提供治疗受试者的感觉传导缺陷的方法,该方法涉及令该受试者的细胞与内耳毛细胞靶向的合成腺相关病毒(aav)载体接触,其中该载体含有编码人ush1多肽的或和感受,其中该ush1多肽是肌球蛋白7a、harmonin、钙粘蛋白23、原钙粘蛋白15、sans和钙整合素结合蛋白2中的任意一者或多者。
另一方面,本发明提供治疗受试者的感觉传导缺陷的方法,该方法涉及令该受试者的细胞与内耳毛细胞靶向的合成腺相关病毒(aav)载体接触,其中该载体含有启动子,该启动子是espin启动子、pcdh15启动子、ptprq启动子或tmhs(lhfpl5)启动子中的任一种。
另一方面,本发明提供治疗受试者的感觉传导缺陷的方法,该方法涉及令该受试者的细胞与内耳毛细胞靶向的合成腺相关病毒(aav)载体接触,其中该载体编码与anc80l65的序列一致性为至少约85%的衣壳,且含有可操作地链接至编码ush1多肽的多核苷酸的启动子,该ush1多肽是肌球蛋白7a、harmonin、钙粘蛋白23、原钙粘蛋白15、sans和钙整合素结合蛋白2。
在本文所述的本发明的上述方面或任何其它方面的多种具体实施例中,该感觉传导缺陷是与在内耳中被表达的多肽中的基因改变相关的遗传性障碍。在上述方面的一种具体实施例中,该启动子是espin启动子、pcdh15启动子、ptprq启动子或tmhs(lhfpl5)启动子中的任意一者或多者。在上述方面的其它具体实施例中,该载体以至少约70%或更高的效率转导内毛细胞和外毛细胞。在上述方面的其它具体实施例中,该harmonin多肽是harmonin-a、harmonin-b或harmonin-c。在上述方面的其它具体实施例中,该细胞是外毛细胞或内毛细胞。在上述方面的一种具体实施例中,该载体含有引导下游多核苷酸表达的启动子,且该启动子是espin启动子、pcdh15启动子、ptprq启动子或tmhs(lhfpl5)启动子。在上述方面的其它具体实施例中,该下游多核苷酸是tmc1、tmc2或ush1多肽,且该ush1多肽是肌球蛋白7a、harmonin、钙粘蛋白23、原钙粘蛋白15、sans和钙整合素结合蛋白2。在上述方面的特定具体实施例中,该harmonin多肽是harmonin-a、harmonin-b或harmonin-c。在上述方面的其它具体实施例中,该内耳毛细胞靶向的合成腺相关病毒(aav)载体以内毛细胞和外毛细胞为靶点,靶向效率为至少约70%、80%、90%、95%或更高,甚至高达100%。在上述方面的其它具体实施例中,该人类多肽是tmc1、tmc2、harmonin-a、harmonin-b或harmonin-c。在上述方面的其它具体实施例中,该感觉传导缺陷是听力障碍或前庭障碍。在上述方面的其它具体实施例中,该感觉传导缺陷是乌谢尔综合征。
定义
除非另做定义,否则本文中使用的所有科技术语均具有本发明所属领域技术人员所一般理解的意义。下述参考文献对技术人员提供本发明中使用的多个术语的一般性定义:singleton等人所著《微生物学和分子生物学词典(第二版)》(singletonetal.,dictionaryofmicrobiologyandmolecularbiology(2nded.1994));《剑桥科技词典》(thecambridgedictionaryofscienceandtechnology(walkered.,1988));rieger等人编撰的《遗传性术语表(第五版)》(theglossaryofgenetics,5thed.,r.riegeretal.(eds.),springerverlag(1991));以及hale和marham所著《哈珀·柯林斯生物学词典》(hale&marham,theharpercollinsdictionaryofbiology(1991))。如本文中所用,除非明确排除,否则下述术语具有归于其下方的意义。
“祖先aav序列”意为设计的序列,其是由于对天然存在的aav的分析和对进化祖先的预测而产生的基因工程产物。
“anc800多肽”意为衣壳多肽,其与下述多肽序列的氨基酸一致性为至少约85%:
maadgylpdwlednlsegirewwdlkpgapkpkanqqkqddgrglvlpgykylgpfngldkgepvnaada
aalehdkaydqqlkagdnpylrynhadaefqerlqedtsfggnlgravfqakkrvleplglveegaktap
gkkrpveqspqepdsssgigkkgqqparkrlnfgqtgdsesvpdpqplgeppaapsgvgsntmaagggapmadnnegadgvgnasgnwhcdstwlgdrvittstrt
alptynnhlykqissqsggstndntyfgystpw
gyfdfnrfhchfsprdwqrlinnnwgfrpkklnfklfniqvkevttndgtttiannltstvqvftdseyqlpyvlgsahqgclppfpadvfmi
pqygyltlnngsqavgrssfycleyfpsqmlrtgnnfqfsytfedvp
fhssyahsqsldrlmnplidqylyylsrtqttsgtagnrtlqfsqagpssmanqaknwlpgpcyrqqrvs
kttnqnnnsnfawtgatkyhlngrdslvnpgpamathkddedkffpmsgvlifgkqgagnsnvdldnvmi
tneeeikttnpvateeygtvatnlqsantapatgtvnsqgalpgmvwqdrdvylqgpiwakiphtdghfhpsplmggfglkhpppqilikntpvpanppttfspakfasfitqystgqvsveieelqkenskrwnpeiqytsnynkstnvdfavdtngvyseprpigtryltrnl
“anc80多核苷酸”意为编码anc80多肽的核酸分子。
“血清阳性率”意为,基于血清学(血清)试样,所测试的群体中对于具体疾病呈阳性的人数。在一种具体实施例中,血清阳性率表征为占总测试试样的百分比或表征为每100,000个被测试人中所占的比例。
“内耳毛细胞靶向的aav”意为一种腺相关病毒,其在给药至受试者的内耳或与源自内耳的细胞进行体外接触后,转染至少70%的内毛细胞和70%的外毛细胞。优选地,内耳毛细胞靶向的aav是在注射入活体耳蜗后转染至少90%的内毛细胞和90%的外毛细胞的aav。可使用编码gfp的基因在小鼠模型内评估转染效率。
“机械感受(mechanosensation)”意为对机械刺激的反应。将机械刺激转化为神经元信号的触觉、听觉和示例的平衡。机械感觉传导输入被转化为通过名为“机械感觉转导(mechanotransduction)”的过程对机械刺激的反应。
“tmc1多肽”意为具有机械转导通道活性的多肽或其片段,该多肽与ncbi参考序列:np_619636.2的氨基酸序列一致性为至少约85%或更大。示例性的tmc1氨基酸序列提供于下:
“tmc1多核苷酸”意为编码tmc1多肽的多核苷酸。例示性的tmc1多核苷酸的序列提供为ncbi参考序列:nm_138691.2,其转载于下:
“tmc2多肽”意为在机械转导中发挥作用的多肽或其片段,该多肽与ncbi参考序列:np_542789的氨基酸序列一致性为至少约85%或更大。例示性的tmc2氨基酸序列提供于下:
“harmonin”多肽意为在机械感受中发挥作用或与ush1c、ush1g、cdh23和myo7a中的任一者或多者相互作用的多肽或其片段,该多肽与q9y6n9-1(亚型(isoform)1)、q9y6n9-2、q9y6n9-3、q9y6n9-4、q9y6n9-5的氨基酸序列一致性为至少约85%。例示性的harmonin-a多肽(亚型1)的序列提供于下:
>sp|q9y6n9|ush1c_humanharmoninos=智人gn=ush1cpe=1sv=3
mdrkvarefrhkvdfliendaekdylydvlrmyhqtmdvavlvgdlklvinepsrlplfdairpliplkhqveydqltprrsrklkevrldrlhpeglglsvrgglefgcglfishlikggqadsvglqvgdeivringysissctheevinlirtkktvsikvrhiglipvksspdepltwqyvdqfvsesggvrgslgspgnrenkekkvfislvgsrglgcsissgpiqkpgifishvkpgslsaevgleigdqivevngvdfsnldhkeavnvlkssrsltisivaaagrelfmtdrerlaearqrelqrqellmqkrlamesnkilqeqqemerqrrkeiaqkaaeeneryrkemeqiveeeekfkkqweedwgskeqlllpktitaevhpvplrkpkydqgvepelepaddldggteeqgeqdfrkyeegfdpysmftpeqimgkdvrllrikkegsldlaleggvdspigkvvvsavyergaaerhggivkgdeimaingkivtdytlaeaeaalqkawnqggdwidlvvavcppkeyddeltff
“ush1c多核苷酸”意为编码harmonin多肽的核酸分子。例示性的ush1c多核苷酸nm_005709的序列提供于下:
其它例示性的harmonin序列提供于下:
harmonin-b
>xm_011519832.2predicted:智人ush1蛋白质网络组件(proteinnetworkcomponent)harmonin(ush1c),转录变体(transcriptvariant)x3,mrna
agctccgagggcggctggcccggtcgcggtcgcggctctttccagctcctggcagccgggcacccgaagg
aacgggtcgtgcaacgacgcagctggacctggcccagccatggaccgaaaagtggcccgagaattccggc
ataaggtggattttctgattgaaaatgatgcagagaaggactatctctatgatgtgctgcgaatgtacca
ccagaccatggacgtggccgtgctcgtgggagacctgaagctggtcatcaatgaacccagccgtctgcct
ctgtttgatgccattcggccgctgatcccactgaagcaccaggtggaatatgatcagctgaccccccggc
gctccaggaagctgaaggaggtgcgtctggaccgtctgcaccccgaaggcctcggcctgagtgtgcgtgg
tggcctggagtttggctgtgggctcttcatctcccacctcatcaaaggcggtcaggcagacagcgtcggg
ctccaggtaggggacgagatcgtccggatcaatggatattccatctcctcctgtacccatgaggaggtca
tcaacctcattcgaaccaagaaaactgtgtccatcaaagtgagacacatcggcctgatccccgtgaaaag
ctctcctgatgagcccctcacttggcagtatgtggatcagtttgtgtcggaatctgggggcgtgcgaggc
agcctgggctcccctggaaatcgggaaaacaaggagaagaaggtcttcatcagcctggtaggctcccgag
gccttggctgcagcatttccagcggccccatccagaagcctggcatctttatcagccatgtgaaacctgg
ctccctgtctgctgaggtgggattggagataggggaccagattgtcgaagtcaatggcgtcgacttctct
aacctggatcacaaggaggctgtaaatgtgctgaagagtagccgcagcctgaccatctccattgtagctg
cagctggccgggagctgttcatgacagaccgggagcggctggcagaggcgcggcagcgtgagctgcagcg
gcaggagcttctcatgcagaagcggctggcgatggagtccaacaagatcctccaggagcagcaggagatg
gagcggcaaaggagaaaagaaattgcccagaaggcagcagaggaaaatgagagataccggaaggagatgg
aacagattgtagaggaggaagagaagtttaagaagcaatgggaagaagactggggctcaaaggaacagct
actcttgcctaaaaccatcactgctgaggtacacccagtaccccttcgcaagccaaagtatgatcaggga
gtggaacctgagctcgagcccgcagatgacctggatggaggcacggaggagcagggagagcagaaaggaa
aagataagaagaaagccaagtatggcagcctgcaggacttgagaaagaataagaaagaactggagtttga
gcaaaagctttacaaagagaaagaggaaatgctggagaaggaaaagcagctaaagatcaaccggctggcc
caggaggatttccggaaatatgaggaaggctttgacccctactctatgttcaccccagagcagatcatgg
ggaaggatgtccggctcctacgcatcaagaaggagggatccttagacctggccctggaaggcggtgtgga
ctcccccattgggaaggtggtcgtttctgctgtgtatgagcggggagctgctgagcggcatggtggcatt
gtgaaaggggacgagatcatggcaatcaacggcaagattgtgacagactacaccctggctgaggctgagg
ctgccctgcagaaggcctggaatcagggcggggactggatcgaccttgtggttgccgtctgccccccaaa
ggagtatgacgatgagctgaccttcttctgaagtccaaaaggggaaaccaaattcaccgttaggaaacag
tgagctccggccccacctcgtgaacacaaagcctcggatcagccttgagagaggccacactacacacacc
agatggcatccttgggacctgaatctatcacccaggaatctcaaactccctttggccctgaaccagggcc
agataaggaacagctcgggccactcttctgaaggccaacgtggaggaaagggagcagccagccatttggg
agaagatctcaaggatccagactctcattcctttcctctggcccagtgaatttggtctctcccagctctg
ggggactccttccttgaaccctaataagaccccactggagtctctctctctccatccctctcctctgccc
tctgctctaattgctgccaggattgtcactccaaaccttactctgagctcattaataaaatagatttatt
ttccaharmonin-b多肽
mdrkvarefrhkvdfliendaekdylydvlrmyhqtmdvavlvgdlklvinepsrlplfdairpliplkhqveydqltprrsrklkevrldrlhpeglglsvrgglefgcglfishlikggqadsvglqvgdeivringysissctheevinlirtkktvsikvrhiglipvksspdepltwqyvdqfvsesggvrgslgspgnrenkekkvfislvgsrglgcsissgpiqkpgifishvkpgslsaevgleigdqivevngvdfsnldhkeavnvlkssrsltisivaaagrelfmtdrerlaearqrelqrqellmqkrlamesnkilqeqqemerqrrkeiaqkaaeeneryrkemeqiveeeekfkkqweedwgskeqlllpktitaevhpvplrkpksfgwfyrydgkfptirkkgkdkkkakygslqdlrknkkelefeqklykekeemlekekqlkinrlaqevseteredleesekiqywverlcqtrleqissadneisemttgppppppsvsplapplrrfagglhlhttdlddipldmfyyppktpsalpvmphpppsnpphkvpappvlplsghvsassspwvqrtpppipippppsvptqdltptrplpsaleealsnhpfrtgdtgnpvedweaknhsgkptnspvpeqsfpptpktfcpspqpprgpgvstiskpvmvhqepnfiyrpavksevlpqemlkrmvvyqtafrqdfrkyeegfdpysmftpeqimgkdvrllrikkegsldlaleggvdspigkvvvsavyergaaerhggivkgdeimaingkivtdytlaeaeaalqkawnqggdwidlvvavcppkeyddelaslpssvaespqpvrklledraavhrhgfllqleptdlllkskrgnqihr"
harmonin-c
>nm_001297764.1智人ush1蛋白质网络组件harmonin(ush1c),转录变体3,mrna
agctccgagggcggctggcccggtcgcggtcgcggctctttccagctcctggcagccgggcacccgaagg
aacgggtcgtgcaacgacgcagctggacctggcccagccatggaccgaaaagtggcccgagaattccggc
ataaggtggattttctgattgaaaatgatgcagagaaggactatctctatgatgtgctgcgaatgtacca
ccagaccatggacgtggccgtgctcgtgggagacctgaagctggtcatcaatgaacccagccgtctgcct
ctgtttgatgccattcggccgctgatcccactgaagcaccaggtggaatatgatcagctgaccccccggc
gctccaggaagctgaaggaggtgcgtctggaccgtctgcaccccgaaggcctcggcctgagtgtgcgtgg
tggcctggagtttggctgtgggctcttcatctcccacctcatcaaaggcggtcaggcagacagcgtcggg
ctccaggtaggggacgagatcgtccggatcaatggatattccatctcctcctgtacccatgaggaggtca
tcaacctcattcgaaccaagaaaactgtgtccatcaaagtgagacacatcggcctgatccccgtgaaaag
ctctcctgatgagcccctcacttggcagtatgtggatcagtttgtgtcggaatctgggggcgtgcgaggc
agcctgggctcccctggaaatcgggaaaacaaggagaagaaggtcttcatcagcctggtaggctcccgag
gccttggctgcagcatttccagcggccccatccagaagcctggcatctttatcagccatgtgaaacctgg
ctccctgtctgctgaggtgggattggagataggggaccagattgtcgaagtcaatggcgtcgacttctct
aacctggatcacaaggagggccgggagctgttcatgacagaccgggagcggctggcagaggcgcggcagc
gtgagctgcagcggcaggagcttctcatgcagaagcggctggcgatggagtccaacaagatcctccagga
gcagcaggagatggagcggcaaaggagaaaagaaattgcccagaaggcagcagaggaaaatgagagatac
cggaaggagatggaacagattgtagaggaggaagagaagtttaagaagcaatgggaagaagactggggct
caaaggaacagctactcttgcctaaaaccatcactgctgaggtacacccagtaccccttcgcaagccaaa
gtatgatcagggagtggaacctgagctcgagcccgcagatgacctggatggaggcacggaggagcaggga
gagcaggatttccggaaatatgaggaaggctttgacccctactctatgttcaccccagagcagatcatgg
ggaaggatgtccggctcctacgcatcaagaaggagggatccttagacctggccctggaaggcggtgtgga
ctcccccattgggaaggtggtcgtttctgctgtgtatgagcggggagctgctgagcggcatggtggcatt
gtgaaaggggacgagatcatggcaatcaacggcaagattgtgacagactacaccctggctgaggctgagg
ctgccctgcagaaggcctggaatcagggcggggactggatcgaccttgtggttgccgtctgccccccaaa
ggagtatgacgatgagctgaccttcttctgaagtccaaaaggggaaaccaaattcaccgttaggaaacag
tgagctccggccccacctcgtgaacacaaagcctcggatcagccttgagagaggccacactacacacacc
agatggcatccttgggacctgaatctatcacccaggaatctcaaactccctttggccctgaaccagggcc
agataaggaacagctcgggccactcttctgaaggccaacgtggaggaaagggagcagccagccatttggg
agaagatctcaaggatccagactctcattcctttcctctggcccagtgaatttggtctctcccagctctg
ggggactccttccttgaaccctaataagaccccactggagtctctctctctccatccctctcctctgccc
tctgctctaattgctgccaggattgtcactccaaaccttactctgagctcattaataaaatagatttatt
ttccagctta
harmonin-c多肽
mdrkvarefrhkvdfliendaekdylydvlrmyhqtmdvavlvgdlklvinepsrlplfdairpliplkhqveydqltprrsrklkevrldrlhpeglglsvrgglefgcglfishlikggqadsvglqvgdeivringysissctheevinlirtkktvsikvrhiglipvksspdepltwqyvdqfvsesggvrgslgspgnrenkekkvfislvgsrglgcsissgpiqkpgifishvkpgslsaevgleigdqivevngvdfsnldhkegrelfmtdrerlaearqrelqrqellmqkrlamesnkilqeqqemerqrrkeiaqkaaeeneryrkemeqiveeeekfkkqweedwgskeqlllpktitaevhpvplrkpkydqgvepelepaddldggteeqgeqdfrkyeegfdpysmftpeqimgkdvrllrikkegsldlaleggvdspigkvvvsavyergaaerhggivkgdeimaingkivtdytlaeaeaalqkawnqggdwidlvvavcppkeyddeltff"
“espin启动子”意为源自ncbi参考序列:ng_015866.1的调节性多核苷酸序列,其足以引导下游多核苷酸在耳蜗细胞中的表达。在一种具体实施例中,espin启动子包含位于espin编码序列上游的至少约350、500、1000、2000、3000、000、5000或更多个碱基对。
“原钙粘蛋白相关15(pcdh15)启动子”意为源自ncbi参考序列:ng_009191的调节性多核苷酸序列,其足以引导下游多核苷酸在耳蜗细胞中的表达。在一种具体实施例中,pcdh15启动子包含位于pcdh15编码序列上游的至少约350、500、1000、2000、3000、000、5000或更多个碱基对。
“蛋白酪氨酸磷酸酶q型受体(ptprq)启动子”意为源自geneid:374462的调节性多核苷酸序列,其足以引导下游多核苷酸在耳蜗细胞中的表达。在一种具体实施例中,ptprq启动子包含位于ptprq编码序列上游的至少约350、500、1000、2000、3000、000、5000或更多个碱基对。
“脂肪瘤hmgic融合伴侣样(partner-like)5(lhfpl5)启动子”也称“tmhs”启动子,意为源自ncbi参考序列:geneid:222662的调节性多核苷酸序列,其足以引导下游多核苷酸在耳蜗细胞中的表达。在一种具体实施例中,tmhs启动子包含位于pcdh15编码序列上游的至少约350、500、1000、2000、3000、000、5000或更多个碱基对。
“剂”意为多肽、多核苷酸或小化合物。
“缓解”意为减少、阻抑、衰减、缩减、迟滞或稳定化疾病或病变的发展或进展。
“改变”意为基因或多肽的表达水平或活性的变化(增加或减少),如通过该领域已知的方法如本文中揭示的那些方法所检测。如本文中所用,改变包括表达水平变化至少10%,优选变化25%,更优选变化40%,且最优选表达水平变化50%或更高。
本公开中,“包含(comprise)”、“包含(comprising)”、“含有(containing)”和“具有(having)”等可具有美国专利法中规定的属于它们的意义,且可意为“包括”等;“主要由...组成”等具有美国专利法中规定的属于它们的意义,且该术语是开放性的,允许超过其所引述者的存在,只要其所引述的基本或新颖特征没有被超过其所引述者的存在改变即可,但不包括先前技术具体实施例。
“检测”指的是证实待检测的被分析物的存在、不存在或量。
“疾病”意为损害或干扰细胞、组织或器官的正常功能的任何病症或病变。疾病的示例包括遗传障碍,其以在例如受试者的内耳中被表达且在机械感觉传导转导中发挥作用的蛋白质功能的损失为特征。在另一具体实施例中,该疾病是乌谢尔综合征(例如,ush1)或老年性听力损失。在一种具体实施例中,疾病是与基因缺陷诸如tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7、ush1c(例如,harmonin-a、harmonin-b或harmonin-c)中的缺陷相关的听觉障碍。
“有效量”意为相对于未治疗患者缓解疾病症状所需的剂的量。用来实践本发明的用于治疗性处理疾病的活性化合物的有效量可变,取决于给药模式以及受试者的年龄、体重和一般健康情况。最终,主治医生或兽医将决定适宜的量和给药方案。这一量被称为“有效”量。
“片段”意为多肽或核酸分子的一部分。这一部分优选地含有参考核酸分子或多肽总长度的至少10%、20%、30%、40%、50%、60%、70%、80%或90%。片段可包含10、20、30、40、50、60、70、80、90、或100、200、300、400、500、600、700、800、900、或1000核苷酸或氨基酸。
“杂交”意为互补核酸碱基之间的氢键键合,其可以是沃森-克里克(watson-crick)、胡斯坦(hoogsteen)和反向胡斯坦氢键键合。举例而言,腺嘌呤和胸腺嘧啶是通过形成氢键而配对的互补核酸碱基。
术语“单离的”、“纯化的”或“生物学上纯的”指的是材料没有不同程度的在其天然状态下发现的正常伴生组分。“单离”表示与原始来源或周围物质分隔的程度。“纯化”表示高于“单离”的分隔程度。“纯化的”或“生物学上纯的”蛋白质充分地不含其它材料,使得任何杂质均不在材料上影响该蛋白质的生物学性质或不造成其它负面后果。换言之,如果本发明的核酸或肽基本上在通过重组dna技术生产时不含细胞材料、病毒材料或培养基,或者当化学合成时不含化学前体或其它化学品,则该核酸或肽是纯化的。典型使用分析化学技术例如聚丙烯酰胺凝胶电泳或高校液相色谱来测定纯度和均质性。术语“纯化的”可表示核酸或蛋白质在电泳凝胶中给出实质上的一条谱带。对于可进行修饰如磷酸化或糖基化的蛋白质,不同的修饰可给出不同的单离的蛋白,它们可独立纯化。
“单离的多核苷酸”意为一种核酸(例如,dna),其不含在本发明的核酸分子所源自的有机体的天然出现的基因组中位于该基因侧翼的基因。该术语因此包括,举例而言,重组dna,其被并入载体内、并入自主复制质粒或病毒内、或并入原核生物或真核生物的基因组dna内,或作为独立于其它序列的单独分子而存在(举例而言,通过pcr或限制性内切酶消化制备的cdna或基因组或cdna片段)。此外,该术语包括从dna分子转录的rna分子,以及作为编码附加多肽序列的杂交基因的一部分的重组dna。
“单离的多肽”意为已经与其天然伴随组分单离的本发明的多肽。典型地,以重量计,当多肽的至少60%不含其天然关联的蛋白质和天然出现的有机分子时,则该多肽是单离的。优选该制剂含有至少75重量%、更优选至少90重量%、最优选99重量%的本发明的多肽。可通过例如从天然来源提取、编码多肽的重组核酸的表达、或化学合成蛋白质来获得本发明的单离的多肽。可通过任何适宜的方法例如柱色谱、聚丙烯酰胺凝胶电泳或hplc分析来测量纯度。
“标记物”意为任意蛋白质或多核苷酸,其在表达水平或活性上有与疾病或病变相关的改变。
如本文中所用,如“获得一剂”中的“获得”包括合成、购买或以其它方法取得该剂。
“启动子”意为足以引导下游多核苷酸的转录的多核苷酸。
“降低”意为至少10%、25%、50%、75%或100%的负向改变。
“参考”意为标准或对照条件。
“参考序列”是用作序列比对基础的界定序列。参考序列可以是特定序列的一部分或整体;例如,全长度cdna或基因序列的连段(segment)、或完整的cdna或基因序列。对于多肽,参考多肽序列的长度将通常为至少约16个氨基酸,优选至少约20个氨基酸,更优选至少约25个氨基酸,且甚至更优选约35个氨基酸、约50个氨基酸或约100个氨基酸。对于核酸,参考核酸序列的长度将通常为至少约50个核苷酸,优选至少约60个核苷酸,更优选至少约75个核苷酸,且甚至更优选约100个核苷酸或约300个核苷酸或与之接近或之间的任何整数。
可用于本发明方法的核酸分子包括编码本发明的多肽或其片段的任何核酸分子。此类核酸分子无须与内源性核酸序列100%一致,但典型将会展现大致的一致性。具有与内源性序列“基本一致性”的多核苷酸典型能与双链核酸分子的至少一条链杂交。可用于本发明方法的核酸分子包括编码本发明的多肽或其片段的任何核酸分子。此类核酸分子无须与内源性核酸序列100%一致,但典型将会展现大致的一致性。具有与内源性序列“基本一致性”的多核苷酸典型能与双链核酸分子的至少一条链杂交。
“杂交”意为在互补多核苷酸序列(如,本文中揭示的基因)或其部分在多种严格条件下配对以形成双链分子。(见,例如,wahl,g.m.ands.l.berger(1987)methodsenzymol.152:399;kimmel,a.r.(1987)methodsenzymol.152:507)。
举例而言,严格的盐浓度一般为少于约750mmnacl和75mm柠檬酸三钠,优选少于约500mmnacl和50mm柠檬酸三钠,且更优选少于约250mmnacl和25mm柠檬酸三钠。低严格度杂交可在有机溶剂如甲酰胺不存在下获得,而高严格度杂交可在至少约35%甲酰胺且更优选至少约50%甲酰胺的存在下获得。严格的温度条件一般将包括至少约30℃,更优选至少约37℃,且最优选至少约42℃的温度。可变的附加因素,如杂交时间、洗涤剂如十二烷基硫酸钠(sds)的浓度、以及包含或不饱和载体dna,是该领域技术人员所周知的。通过按需要组合这些多种条件来实施各种水平的严格度。在优选的实施方式中,杂交将出现在30℃的750mmnacl、75mm柠檬酸三钠和1%sds中。在更优选的实施方式中,杂交将出现在37℃的500mmnacl、50mm柠檬酸三钠、1%sds、35%甲酰胺和100μg/ml变性鲑鱼精dna(ssdna)中。在最优选的实施方式中,杂交将出现在42℃的250mmnacl、25mm柠檬酸三钠、1%sds、50%甲酰胺和200μg/mlssdna中。对这些条件的有用改变对于该领域技术人员是显而易见的。
对于大多数应用,杂交后的洗涤步骤的严格度也将改变。可通过盐浓度和温度来界定洗涤严格度条件。如上,可通过降低盐浓度或增加温度来增加洗涤严格度。举例而言,洗涤步骤的严格的盐浓度优选为少于约30mmnacl和3mm柠檬酸三钠,且最优选少于约15mmnacl和1.5mm柠檬酸三钠。用于洗涤步骤的严格的温度条件一般将包括至少约25℃,更优选至少约42℃,且甚至更优选至少约68℃的温度。在优选的实施方式中,洗涤步骤将出现在25℃的30mmnacl、3mm柠檬酸三钠和0.1%sds中。在更优选的实施方式中,洗涤步骤将出现在42℃的15mmnacl、1.5mm柠檬酸三钠和0.1%sds中。在更优选的实施方式中,洗涤步骤将出现在68℃的15mmnacl、1.5mm柠檬酸三钠和0.1%sds中。对这些条件的额外改变对于该领域技术人员是显而易见的。杂交技术是该领域技术人员所周知的,且揭示在例如bentonanddavis(science196:180,1977);grunsteinandhogness(proc.natl.acad.sci.,usa72:3961,1975);ausubeletal.(currentprotocolsinmolecularbiology,wileyinterscience,newyork,2001);bergerandkimmel(guidetomolecularcloningtechniques,1987,academicpress,newyork);以及sambrooketal.,molecularcloning:alaboratorymanual,coldspringharborlaboratorypress,newyork中。
“基本相同”意为多肽或核酸分子展现与参考氨基酸序列(例如,任何一种本文所述的氨基酸序列)或核酸序列(例如,任何一种本文所述的核酸序列)的至少50%一致性。优选地,此序列在氨基酸水平或核酸水平上与用于比较的序列的一致性为至少60%、更优选80%或85%、且更优选90%、95%或99%。
通常使用序列分析软件(例如,威斯康星大学生物技术中心的遗传学计算机公司(geneticscomputergroup,universityofwisconsinbiotechnologycenter,1710universityavenue,madison,wis.53705)的序列分析软件包blast、bestfit、gap、或pileup/prettybox程序)测量序列一致性。该软件通过设定多种替换、删除、及/或其它修饰的同源性程度来匹配一致或相似的序列。保守替换典型包括下述各组的组内替换:甘氨酸、丙氨酸;缬氨酸、异亮氨酸、亮氨酸;天冬氨酸、谷氨酸、天冬酰胺、谷氨酰胺;丝氨酸、苏氨酸;赖氨酸、精氨酸;以及苯丙氨酸、酪氨酸。在例示性的测定一致性程度的途径中,可使用blast程序,其中介于e-3与e-100之间的可能性得分指示密切相关的序列。
“受试者”意为哺乳动物,包括但不限于人类和非人哺乳动物,诸如牛、马、犬、羊或猫科动物。
“转基因”意为被人工插入细胞内并变为从该细胞发展而来的有机体基因组的一部分,或者在线虫转基因的情况下,变为遗传染色体外阵列的一部分的任何dna碎片。此类转基因可包括与转基因有机体部分上或整体上异源(即,异质)的基因,或可表示与该有机体的内源基因同源的基因。
本文中提供的范围理解为该范围内所有值的略写。例如,1至50的范围理解为包括来自由1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49或50所组成的组的任意数字、数字组合或子范围。
如本文中所用,“治疗(treat,treating,treatment)”等指的是减轻或缓解与其相关的病变和/或症状。应知晓,尽管未排除,但治疗病症或症状并不需要完全消除与该病症或症状相关的病症、症状或症候。
除非明确指出或从语境中明显可见,否则如本文中所用,术语“或”理解为包含的。除非明确指出或从语境中明显可见,否则如本文中所用,术语“一(a,an)”和“该(the)”理解为单数或复数。
除非明确指出或从语境中明显可见,否则本文中使用的术语“约”理解为处于该领域正常公差范围内,例如,处于均值的2标准偏差内。“约”可理解为处于所指出值的10%、9%、8%、7%、6%、5%、4%、3%、2%、1%、0.5%、0.1%、0.05%或0.01%内。除非从语境中明确排除,否则本文中提供的所有数值均以术语“约”修饰。
本文中,对任何变量定义中一系列化学基团的描述包括该变量作为任何单一基团或作为所列基团的组合的定义。对本文中变量或方面的具体实施例的描述包括该具体实施例作为任何单一输送法南或作为与任何其它具体实施例或其部分的组合。
本文中提供的任何组合物或方法可与本文中提供的一种或多种任何其它组合物和方法组合。
除非另做定义,否则本文中使用的所有科技术语均具有与该方法和物质组合物所属领域技术人员所一般理解的相同的意义。尽管与本文中揭示者类似或等效的方法和材料可用于实践或测试该方法和物质组合物,但适当的方法和材料揭示如下。此外,材料、方法和实施例仅做例示性说明之用,而非试图限制。本文中提及的所有出版物、专利申请、专利和其它参考文献通过引用而以其整体并入本文。
附图说明
第一部分:高效率的耳蜗基因转移
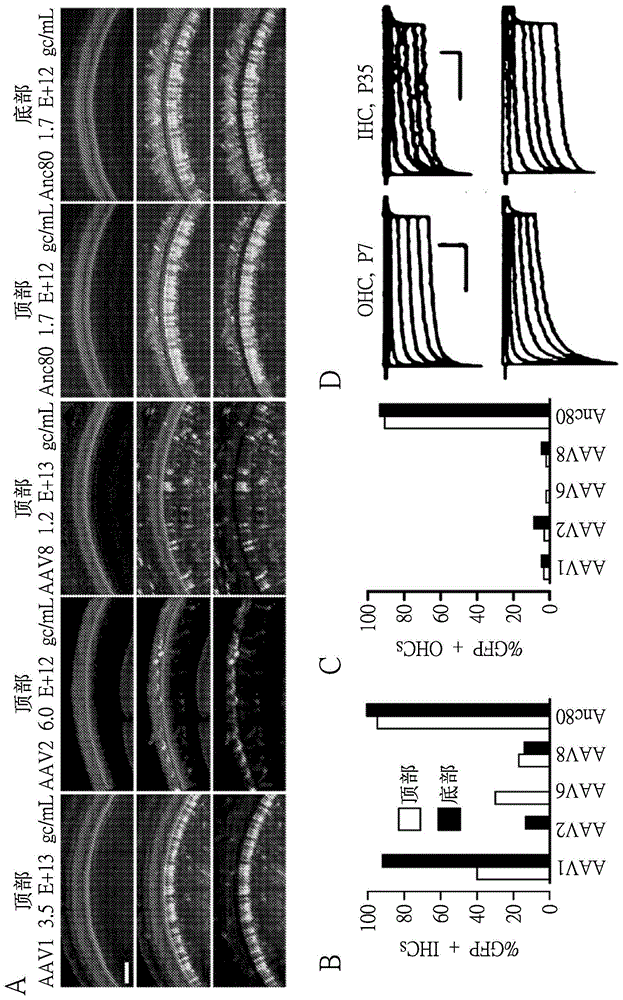
图1a至图1g提供了一系列显微照片(a)和六组图表(b至g)。天然aav血清型和anc80的体内耳蜗转导。(a)小鼠的柯蒂(corti)氏器官的共聚焦图像,用alexa-546-鬼笔环肽(红色)复染并对egfp成像(绿色,本文档中显示为各组中图和下图中的明亮染色)。对小鼠注射1μl的各组图上方所标注效价的aav原液。比例尺为50μm。(b)对注射aav-egfp的耳蜗的底部和定点处egfp阳性ihc的定量。(c)对注射aav-egfp的耳蜗的底部和定点处egfp阳性ohc的定量。对c57bl/6小鼠的一只耳朵注射,数量为5只动物(aav1)、4只动物(aav2)、2只动物(aav8)、1只动物(aav6)、3只动物(anc80)。(d)在p7(左侧)从egfp阴性ohc(黑色)和egfp阳性ohc(绿色)记录的多组感觉转导电流。纤毛束以0.1μm的增量在-0.1μm与1μm之间偏转。纵向比例尺表示200pa;水平则表示20msec。来自egfp阴性(黑色)和egfp阳性(绿色)p35ihc的电流显示在右侧。纵向比例尺表示100pa;水平则表示20msec。(e)对底部所标注年龄的103个ihc和ohc绘制感觉转导电流幅度。显示来自egfp阴性(黑色)和egfp阳性(绿色)的数据。每组中的细胞数显示在该图上。所有小鼠均在p1注射。(f)均值±标准偏差(sd)。对四只anc80注射耳(绿色)和四只未注射耳(黑色)与由于注射相关的损伤而没有egfp荧光的一只注射耳(红色)的数据一起绘制abr阈值。(g)均值±sd。对四只anc80注射耳(绿色)和四只未注射耳(黑色)以及一只具有注射损伤而没有egfp荧光的阴性对照耳(红色)绘制dpoae阈值。b至g中数据点的注射效价同a。
图2a至图2d是显示前庭感觉上皮细胞中anc80-egfp转导的图像。(a)来自注射1μlanc80-egfp(1.7x1012gc/ml)的p1小鼠的小鼠椭圆囊。在p10收获该组织,固定,以alexa546-鬼笔环肽(红色)染色,并对egfp成像(绿色)。横跨egfp阳性细胞的多个聚焦平面的形态学评估表明,在所检查的每个样品(未显示)中,i型细胞的常规圆锥状形态以及ii型细胞的圆柱状形态。比例尺为100μm。(b)来自与组图a中所述相同的小鼠的后半规管壶腹嵴。比例尺为50μm。(c)人椭圆囊的感觉上皮。将该组织曝露于1010gcanc80.cmv.egfp.wpre中24小时,培养10天,固定,以alexa546-鬼笔环肽(红色)染色,并对egfp荧光(绿色)成像。比例尺为100μm。(d)以alexa546-鬼笔环肽(红色)和myo7a(蓝色)染色并对egfp(绿色)成像的椭圆囊中的人上皮细胞的高倍率视图,转导条件同c。重叠图中的白色箭头表示所选择的egfp阳性/myo7a阳性细胞。比例尺为20μm。
图3a至图3e是显示以anc80转导的小鼠耳蜗中大量内毛细胞和外毛细胞的图像。(a)在p1注射1μl的1.7x1012gc/ml的anc80-egfp的小鼠耳蜗的整个顶部的低倍率图像。在p10收获该耳蜗,以alexa546-鬼笔环肽(红色)染色,并对egfp成像(绿色)。比例尺为100μm。(b)在p1注射1μl的1.7x1012gc/ml的anc80-493的不同小鼠耳蜗的基底部的高倍率图像。在p10收获该耳蜗,以alexa546-鬼笔环肽(红色)染色,并对egfp成像(绿色)。比例尺为20μm。(c、d)对p1至p2的c57bl/6小鼠进行圆窗注射后,对在相同剂量下的所有血清型的内毛细胞和外毛细胞转导效率的定量比较。对c、d中的小鼠耳蜗注射1.36x1012的aav1、aav2、aav8和anc80,在7至9天后收获耳蜗进行活体细胞成像,并通过落射荧光显微镜定量(n=8只每组)。(e)anc80毛细胞转导的剂量依赖性。将曝露于两个不同的anc80-egfp效价(1.8x1012对比1.36x1012gc)下的耳蜗固定,用alexa546-鬼笔环肽(红色)染色,并对egfp(绿色)成像。比例尺为20μm
图4a和图4b分别是一张图像和图表,显示在anc80耳蜗转导后的前庭功能。在p1经由rwm对小鼠注射anc80.cmv.egfp,并在转棒装置上评估表达和平衡功能。经由使用对myo7a(红色)进行免疫荧光染色的共聚焦显微镜评估前庭组织中egfp(绿色)的表达。转棒数据显示,在注射对照组与未注射对照组之间没有差异。绘制直到小鼠从该装置上摔落为止的时间+/-sem。n=3只动物,各自进行5次实验(注射组);以及,n=2只动物,各自进行5次试验(对照组)(b)。比例尺为50μm
第二部分:基因疗法复原了1c型乌谢尔综合征小鼠模型的听觉和前庭功能
图5a至图5l是显示p8的ush1cc.216g>a突变小鼠的柯蒂氏器官的扫描电子显微镜检查的图像。(a至f)对p8的c.216ga(n=3只小鼠)和c.216aa(n=4只小鼠)突变鼠的柯蒂氏器官的基底部、中部和顶部成像。在杂合小鼠体内,ohc和ihc纤毛束得以保存,但在纯合216aa小鼠体内,一些纤毛束表现为沿着柯蒂氏器官而紊乱。(g至l)高倍率图像显示,在多数但非全部ohc(g至h)和ihc(i至j)中的具有阶梯阵列性扰乱的片段化且紊乱的纤毛束。在p8对器官中部的ohc纤毛束成像的示例示出了存在于相同处理器官中的被保存的纤毛束(k)和紊乱的纤毛束(l)。星形表示被保存的纤毛束;单个三角形(arrowhead)表示紊乱的纤毛束;而多个箭头表示波状的ihc纤毛束。比例尺:低倍率:5μm(a至f);高倍率:2μm(g)、3μm(h)、2μm(i-863、j)和1μm(k、l)。
图6a至图6h是显示ush1cc.216g>a新生突变小鼠的毛细胞内的机械感觉转导的图像。(a至d)使用渗透性的苯乙烯基染料fm1-43评估c.216ga和c.216aa小鼠的毛细胞内开放的转导通道的存在。在柯蒂氏器官中,p4的c.216aa小鼠感觉毛细胞中的fm吸收被降低(a至b,中下部)。注意,由于ihc处于不同的聚焦平面内,ihcfm1-43荧光出现减光。左图:dic,右图:fm1-43;比例尺为10μm。在椭圆囊中,fm1-43吸收被限制在p6的c.216aa突变体的横纹外区域(extra-striolaregion)(c;比例尺为50μm),而椭圆囊毛细胞保持总体正规束形貌,如通过dic所评估(d;比例尺为10μm)。图d中的白线勾勒出横纹(无吸收)区域和横纹外区域(吸收)。实验重复三次。(e至h)在新生c.216ga和c.216aa小鼠的ohc、ihc和vhc中评估机械感觉转导(自其记录的小鼠数分别为:对于ohc,n=7、6;对于ihc,n=2、4,且对于vhc,n=2、6,细胞数标注在柱状图上方)。绘制了代表性的转导电流(e)、使用二阶玻尔兹曼函数拟合的它们的相关电流/位移图(f)和平均峰值转导电流(g至h)。在耳蜗中,在p3至p6的中回和中顶回中获得记录值。在椭圆囊中,从p5与p7之间的横纹外区域和横纹区域的vhc记录转导电流(e-f)。尽管纤毛束似乎在dic下得以良好保存,但在c.216aa突变体中诱发了较小的平均转导电流(h)。两种基因型之间,ohc、ihc和vhc中的平均峰值转导显著不同(***p<0.01,单因素方差分析)。
图7a至图7e是显示,在体外和体内曝露于腺相关病毒载体的组织中,荧光标记的harmonin的表达和定位的图像。(a至c)将细致剖开的p0至p1内耳组织曝露于aav2/1载体中24h,在培养物中保持7至8天之后,固定,复染(alexafluor鬼笔环肽,invitrogen),并使用zeisslsm共聚焦显微镜成像。野生型椭圆囊中大量感觉毛细胞被感染,且在大多数特异性地位于感觉纤毛束顶部的毛细胞中证实了融合至egfp的harmonin-b1表达(a,比例尺:10μm,上图;5μm,下图)。类似地,在c.216aa小鼠和野生型小鼠的ohc和ihc内的静纤毛尖处证实了egfp::harmonin-b1的表达(b,比例尺:10μm;c,比例尺:3μm)。当在p1注射aav2/1.cmv.egfp::harmonin-b1载体时,在一些处于p60的左侧注射耳内的ihc和ohc中检测到了egfp信号(d,比例尺:30μm)。在p0曝露于aav2/1.cmv.tdtomato::harmonin-a1中24h的p7器官型培养物中的细胞体中检测到了外源
tdtomato::harmonin-a1(e,比例尺:5μm)。一些harmonin-a1色斑与ctbp2共同定位(蓝色;鼠抗ctbp21/200,bdbioscience),尤其在大概接近带状突触的感觉细胞的基底部处。在静纤毛束中未观察到表达。
图8a至图8c是显示注射anc80harmonin载体的小鼠的毛细胞内的机械感觉转导恢复的图像。(a至c)记录未注射的c.216aa对照小鼠(n=8细胞,一只小鼠)和在p1注射aav2/anc80.cmv.harmonin-b1的c.216aa小鼠(0.8μl,n=15细胞,一只小鼠)、或联合注射aav2/anc80.cmv.harmonin-b1和aav2/anc80.cmv.harmonin-a1的小鼠(0.5μl+0.5μl,n=7细胞,一只小鼠)的ihc中的机械感觉转导电流。在p6制备器官型培养物,并在p15与p16(9至10div)之间进行记录。尽管可通过c.216aa小鼠的纤毛束刺激诱导小的机械感觉转导电流,但更大的电流在注射驱动harmonin-b1表达或harmonin-a1和harmonin-b1双重表达的载体的c.216aa小鼠体内被诱发(a)。对每个数据集绘制响应的i/x曲线,并使用二阶玻尔兹曼拟合方程进行拟合。最大机械感觉转导电流imax分别为102.1pa(c.216aa)、424.3pa(c.216aa+harmonin-b1)和341.1pa(c.216aa+harmonin-a1&-b1)(b)。平均反应(均值±s.d.)显示,注射harmonin-b1的小鼠和注射harmonin-a1+harmonin-b1的小鼠相对于未注射小鼠的显著的转导恢复(***p<0.001)。在注射harmonin-b1的小鼠与c.216ga对照小鼠之间没有平均转导电流的显著差异(n.s.p>0.5)。当将harmonin-a与harmonin-b组合时,机械感觉转导的恢复也未得以显著改善。(c),单因素方差分析。
图9a至图9e是显示在p1注射aav2/anc80.cmv.harmonin-b1的小鼠体内的abr和dpoae阈值恢复的图像。(a)6周龄c.216aa对照小鼠和和在p1经由rwm注射编码harmonin-a1、harmonin-b1或两者组合的载体的c.216aa小鼠体内,对于16khz音调的代表性abr反应。在单独注射harmonin-b1的小鼠或同时注射harmonin-a1和harmonin-b1的小鼠体内,测量到接近30dbspl的恢复的abr阈值。(b)对于下列获得的均值abr反应:c.216aa(n=13);c.216ga(n=12);c.216aa+harmonin-a1(n=12);c.216aa+harmonin-b1(n=19只修复/25只测试鼠);c.216aa+harmonin-a1&-b1(n=6只修复/11只测试鼠)。均值±s.e,实线。虚线:其16khz记录显示在图a中的小鼠体内,对于整个频率范围的abr阈值。(c)对于下列获得的均值dpoae反应:c.216aa(n=13);c.216ga(n=12);c.216aa+harmonin-a1(n=12);
c.216aa+harmonin-b1(n=15只修复鼠-dpoae<70dbspl/25只测试鼠);c.216aa+harmonin-a1&-b1(n=4只修复鼠dpoae<70dbspl/11只测试鼠)。均值±s.e,实线。虚线:其记录在图a中示出的四只小鼠的dpoae阈值。箭头表示阈值高于所测试的最大刺激水平。(d至e)在6周和3个月,在八只小鼠体内获得的abr和dpoae反应,显示低于或等于45db的初始abr阈值。将八只小鼠中的六只保持6个月,并进行abr和dpoae评估(虚线)。均值±s.e.。尽管在最开始的三个月内证实了abr和dpoae阈值偏移,但在6个月时间内的较低频率范围内听力修复仍然突出。
图10a至图10e是显示在p1注射aav2/anc80.cmv.harmonin-a1和aav2/anc80.cmv.harmonin-b1的小鼠体内的惊吓反应、转棒能力和开放场域行为恢复的图像。(a)记录6周龄c.216ga对照小鼠、c.216aa对照小鼠和c.216aa注射小鼠对于白噪音刺激的惊吓反应。在注射harmonin-b1的小鼠体内证实了部分的惊吓反应修复,但注射harmonin-a1的小鼠无此反应(数据与c.216aa对照小鼠重叠)。平均值显示为±s.e.。(b)记录4至6周之间的c.216ga对照小鼠、c.216aa对照小鼠和c.216aa注射小鼠的转棒能力。在注射harmonin-b1的小鼠和注射harmonin-a1/b1的小鼠体内观察到了完全恢复。在单独注射harmonin-a1的小鼠体内未观察到恢复。平均值显示为±s.e.。(c至e)对6周龄的c.216ga对照小鼠、c.216aa对照小鼠、c.216aa注射小鼠和c.216ga注射小鼠实施5分钟的开放场域观察。显示了2.5min内的代表性痕迹(b)。尽管c.216aa突变小鼠探索了整个场域并实施了重复性的全身旋转,但在p1注射harmonin-a1、harmonin-b1或两种载体的组合的c.216aa小鼠演示了与其杂合c.216ga配对物或注射截短的载体的c.216ga小鼠相类似的正常行为(c)。图表示出了每分钟转动圈数和覆盖距离的均值±s.d.。在未注射的小鼠与注射的小鼠之间观察到显著的恢复差异***p<0.001。通过单因素方差分析进行统计学分析。
图11是注射aav2/anc80.cmv.harmonin-b1的小鼠的柯蒂氏器官的扫描电子显微镜图像。对六周龄的c.216ga小鼠、c.216aa小鼠和在p1注射(rmw注射0.8μlaav2/anc80.cmv.harmonin-b1)的c.216aa小鼠的柯蒂氏器官的基底部、中部和顶部成像。在c.216ga小鼠体内,ohc和ihc纤毛束得以保存,但在c.216aa小鼠体内,表现为沿着柯蒂氏器官而紊乱。在c.216aa小鼠体内观察到了明显的毛细胞损失(星号)和纤毛束紊乱,且在该器官的基底端具有更明显的退化。c.216aa小鼠的纤毛束缺乏正常的静纤毛行。较短的行看起来似乎是可伸缩的,而最高的行在c.216aa小鼠中定义维持(箭头)。尽管在修复的c.216aa小鼠体内仍然证实了毛细胞损失和纤毛束紊乱,但毛细胞生存率显著高于该器官的基底部和中部。毛细胞计数汇总在柱状图中。在c.216aa小鼠(4只耳朵)中计数到总计1824个细胞,而在修复的c.216aa小鼠(2只耳朵)中计数到总计792个细胞。均值±s.e.。高倍率成像显示了对经注射的c.216aa小鼠(箭头)体内多个但非全部细胞(三角形)的阶梯阵列状修复。比例尺:低倍率:5μm;高倍率:1μm。
图12a至图12i是显示通过sem在p18对ush1cc.216g>a小鼠进行的纤毛束形貌分析的图像。(a至c)杂合c.216ga小鼠在p18展示正常的纤毛束形貌。(d至i)沿着p18纯合c.216aa突变小鼠的器官观察到了紊乱的纤毛束。(j至l)c.216aa小鼠体内的ihc纤毛束被轻微扰乱。所测量的与顶尖的距离:底部:3.5至4mm;中部:1.8至2.2mm;顶部:0.6至0.8mm。比例尺:低倍率:5μm;高倍率:1μm。
图13a至13j是显示c.216aa突变小鼠的机械感觉转导特性的图像。(a至e)对来自p3至p6的耳蜗中回和中顶回的新生ohc内的机械感觉转导分析。使用双指数衰减函数拟合来自~po=0.5的代表性电流痕迹,以评估c.216ga和c.216aa突变体的适应性(a)。使用拟合来生成快时间常数(c)和慢时间尝试(d)以及适应程度(e)。10%至90%的操作范围没有显著改变(b)。c.216aa小鼠的适应程度显著低于这一散点图中所示的杂合ohc(e)。(f至j)新生ihc中的机械感觉转导分析。与相比,c.216gaihc中的10%至90%操作范围值比c.216aaihc中的小(g)。尽管略低,但适应性一直存在,而在c.216aaihc中的程度显著更低(h至j)。统计学分析标注在各图中:*p<0.05,**p<0.01且***p<0.001,单因素方差分析。
图14a至图14c是一组图像,其显示在p1进行双载体注射后6周的c.216aa的柯蒂氏器官内荧光标记的harmonin-a和harmonin-banc80载体的表达。(a至c)在p1进行aav2/anc80.cmv.tdtomato::harmonin-a1(0.5μl;4.11e^12gc/ml)和aav2/anc80.cmv.egfp::harmonin-b1(0.5μl;2.99e^12gc/ml)联合注射后的6周龄c.216aa小鼠的基底回的共聚焦图像。总细胞数的69%和74%分别表达egfp(a)和tdtomato(c),且65%表达两种标记物,表明了成功的共转导。比例尺:20μm。
图15a至图15f是显示对6周龄c.216ga对照小鼠和注射修复的c.216aa小鼠的abr反应分析的数据。(a、d)c.216ga对照小鼠和修复的c.216aa小鼠在8和16khz的abr反应的实施例。(b至c、e至f)6周龄小鼠在8至11.3khz和16khz的平均峰1幅度(b至d)和延迟(c至d),且阈值相当(n=8c.216ga,n=5c.216aa+harmonin-b1,rwmp1)。均值±s.e.:单因素方差分析。
图16a至图16d显示,在ush1cc.216g>a小鼠体内表达的harmonin突变形式并不改变毛细胞或听觉功能。(a)野生型harmonin-b1蛋白质与作为隐秘剪接结果的截短的harmonin之间的序列对齐,和与ush1c基因的外显子3中的acadiang>a突变相关的框移。(b)来自p2至p3野生型小鼠、c.216ga小鼠和c.216aa突变小鼠的听觉器官的半定量rt-pcr确认了野生型(450bp)和截短的(-35bp)harmonin在c.216ga小鼠和c.216aa小鼠体内的表达。(c至d)测量6周龄c.216ga注射小鼠和c.216ga对照小鼠以及c.216aa对照小鼠的听脑干反应(abr,c)和畸变产物(dpoae,d)。图示显示为平均±s.e.。与216ga对照小鼠相比,注射的小鼠体内没有观察到阈值偏移。
图17a至图17c是显示注射aav2/anc80.cmv.harmonin-b1的6周龄小鼠内耳中的准确ush1c剪接的恢复的图像。(a)对来自ush1c.216a基因座的准确剪接的(450bp)和异常的(415bp)mrna的半定量rt-pcr定量显示了#1和#2c.216aa修复小鼠的注射耳(i)和对侧耳(c)中准确ush1c剪接的修复(在11.3khz,来自注射耳的35dbspl反应)。具有极差abr反应(在11.3khz,90dbspl)的#3小鼠显示了准确mrna表达的中度恢复,而#4小鼠(在11.3khz,100dbspl)没有显示修复。尽管在未注射的c.216aa小鼠(#5和#6小鼠)体内没有检测到准确剪接形式,但在c.216ga小鼠(#7、#8、#9小鼠)体内检测到了准确和截短的两种剪接形式。将底图中显示的相应小鼠gapdh放大,以确认材料的相对量。(b)半定量放射标记pcr分析确认了aav-mush1c在ush1c.216aa小鼠的注射耳和对侧耳中的存在。在#3和#4小鼠体内存在相对水平的aav-mush1cdna,但含量减少。(c)相对量的aav-mush1c与abr阈值相关联。示出了在11.3khz和16khz进行的分析。线性回归显示,两者之间高度相关。
图18是显示长期abr阈值恢复与听觉器官中至顶部中ohc存活率相关联的图。在三只未注射的6月龄c.216aa小鼠和五只注射(p1rwm注射,0.8μlaav2/anc80.cmv.harmonin-b1)的6月龄c.216aa小鼠死后,对其左耳实施横跨整个柯蒂氏器官的毛细胞计数。插入:尽管小鼠中有两只(#1和#2)显示极差的横跨整个测试范围的abr反应阈值(≥95dbspl),但有三只(#3至#5)以35至55dbspl范围内的阈值对5.6至16khz之间的声音刺激产生反应。在注射的小鼠体内,ihc和ohc毛细胞的总数有所增加。将注射修复的小鼠与那些具有极差修复的注射小鼠比较,显示ihc的数目没有差异,但在修复的小鼠体内注意到大量的ohc。横跨整个器官长度的分析显示,该差异可以解释为从该器官中部至顶部中毛细胞存活率的增加。
第三部分:听力损失所牵涉的附加突变的基因疗法
图19a至图19d是来自ush1c突变小鼠的耳蜗的代表性共聚焦图像,其中在p2通过rwm向小鼠注射1ml的anc80-harmonin::gfp(即,gfp被融合至harmonin多肽)(6×1012gc/ml),在p9收获耳蜗,使用肌动蛋白(红色;19a),myo7a(蓝色;19b)染色,并进行gfp成像(绿色;19c)。(19a)、(19b)和(19c)的合并图像显示在(19d)中。
图20是显示将ush1c突变小鼠(方块)和注射anc80-harmonin::gfp载体的ush1c突变小鼠(圆圈)的abr阈值作为声音频率的函数而绘制的图。
图21a至图21c显示在p2通过rwm注射1μl的anc80-kcnq4(6×1012gc/ml)、在p9收获、使用alexa546–鬼笔环肽(红色)和抗kcnq4的抗体(绿色)染色的kcnq4-/-耳蜗的代表性共聚焦图像,分别为低倍率(21a)或高倍率(21b),相对于高倍率下的未注射耳蜗(21c)。
图22a至22c是一系列图表,其显示p10野生型小鼠(22a)、p10kcnq4-/-小鼠(22b)、以及经由rwm(2.4×1013gc/ml)注射anc80-kcnq4的p10kcnq4-/-小鼠(22c)体内的kcnq4电流。在注射后8天,收获耳蜗。
图23是注射anc80-cmv-tmc1载体的tmc1-/-组织中的fm1-43吸收(fm1-43仅穿透功能性tmc1通道)的一系列三张图像。经由rwm对p2tmc1-/-小鼠注射anc80-cmv-tmc1载体(2.4×1013gc/ml),在注射后6至7天收获耳蜗。
图24a显示从p10野生型小鼠(左)、p10tmc1-/-小鼠(中)、和在p2经由rwm(2.4×1013gc/ml)注射anc80-cmv-tmc1的p10tmc1-/-小鼠(右)的ihc记录的代表性的几组感觉转导电流。在注射后8天,收获耳蜗。
图24b是图24a中所示小鼠的恢复速率的图解。图24中的图表明了野生型小鼠(左)、tmc1-/-小鼠(中)、和注射anc80-cmv-tmc1的tmc1-/-小鼠(右)体内功能性细胞的百分比。
图25是显示野生型小鼠、tmc1-/-小鼠、和注射anc80-cmv-tmc1的tmc1-/-小鼠的作为刺激频率的函数的畸变产物耳声发射(dpoae)阈值的图。
图26a至图26c显示各种启动子对于表达的效果。图26a显示膜细胞中的anc80-pcdh15-gfp表达,支持细胞gfp和myosin7a如所标注者显示,且毛细胞被myosin7a染为红色,myosin7a是一种毛细胞标记物。图26b显示anc80-myo6-gfp与显示gfp表达的内毛细胞和外毛细胞。在这一实验中没有进行计数染色。图26c显示anc80-kcnq4-gfp-gfp与显示gfp表达的外毛细胞。使用鬼笔环肽对该组织进行计数染色,以照亮毛细胞。
具体实施方式
本发明提供将包括听力和/或前庭功能在内的机械感觉传导所需的蛋白质(例如,tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c))递送至受试者的内耳细胞注入耳蜗细胞(例如,内毛细胞或外毛细胞)并令其在该细胞内表达的组合物和方法,其中该受试者体内的该蛋白质损失或水平下降。
本发明至少部分地基于下述发现:合理设计的编码anc80衣壳的合成载体(“anc80载体”)可用于将转基因有效递送至耳蜗;以及,这一载体将会被用来将野生型ush1c递送至乌谢尔综合征小鼠模型的内耳中。圆窗膜注射导致小鼠体内的内毛细胞和外毛细胞的高效转导,相对于传统的腺相关病毒(aav)载体具有实质性改善。anc80圆窗注射耐受良好,如由感觉细胞功能、听力和前庭功能以及免疫学参数所指示。恢复负载的听觉功能所需的anc80高效率地以外毛细胞为靶点的能力,可以令用于听力和平衡障碍的未来的基因疗法成为可能。注射编码ush1c的anc80载体的小鼠演示了基因和蛋白质表达的恢复、感觉细胞功能的复原、复杂听觉功能的修复、以及听力和平衡行为恢复至接近野生型水平。数据表示了前所未有的内耳功能的恢复,且表明,用以治疗耳聋的生物疗法可能适合转用于患有遗传性内耳障碍的人类。
乌谢尔综合征(乌谢尔syndrome)
人类乌谢尔综合征(ush)是导致又聋又盲的罕见基因状况。作为常染色体隐性性状遗传,其在美国影响16,000至20,000人,且是3%至6%的儿童早期耳聋的原因。根据症状的严重性,乌谢尔综合征分为三个临床亚型(ush-1、ush-2和ush-3)。ush1是最严重的形式。受ush1影响的患者苦于先天性双侧深感神经性听力损失、前庭反射消失和青春期前色素性视网膜炎(视网膜视杆视锥细胞功能的进行性的、双侧的、对称性的退化)。除非配备人工耳蜗,否则个体通常无法发展出说话的能力。尽管目前尚无用于乌谢尔综合征患者的生物学治疗手段,但有缺陷的基因的野生型形式的早期再引入可能考虑到疾病的逆转。
6个乌谢尔基因与ush1有关:myo7a(myosin7a)、ush1c(harmonin)、cdh23(钙粘蛋白23)、pcdh15(原钙粘蛋白15)、sans(sans)和cib2(钙整合素结合蛋白2)。这些基因编码内耳中纤毛束形态发生所牵涉且作为相互作用组的一部分的蛋白质(见,例如,mathur&yang,2015,biochim.biophys.acta,1852:406-20)。harmonin停留在ush1相互作用组的中心,在该处,其结合至其它乌谢尔1蛋白。因为它的pdz(psd-5995/dlg/zo-1)相互作用域,harmonin已经被提出发挥支架蛋白的功能。体外结合研究已经显示,和两种ush2蛋白(即usherin和vlgr1)一样,所有其它已知的ush1蛋白均结合至harmonin的pdz域。ush1c基因由28个外显子组成,其编码10个交替剪接形式的harmonin,并依据蛋白质的域组成而分为三个不同的子类(a、b和c)。三种亚型的区别在于pdz蛋白质-蛋白质相互作用域、卷曲螺旋(cc)域、和富含脯氨酸-丝氨酸-苏氨酸(pst)的域的数目。
ush1蛋白质定位在毛细胞顶部的机械感觉纤毛束中,该纤毛束由数百个通过大量细胞外链接而互相连接的静纤毛构成。钙粘蛋白23和原钙粘蛋白15分别为乌谢尔基因为ush1d和ush1e的产物,形成位于静纤毛远端的尖端链接。harmonin-b结合至cdh23、pcdh15、f-肌动蛋白和它本身。其可见于接近毛细胞内尖端链接插入点的静纤毛的尖端,且被认为在毛细胞的转导和适应中扮演功能性角色。
harmonin-b在产后早期阶段被表达,但其在耳蜗和前庭两者中的表达在产后30天(p30)左右减少。harmonin-a也结合至钙粘蛋白23,且见于静纤毛中。近期的报导透露了harmonin-a在突触处的额外角色,在该处,harmonin-a与cav1.3ca2+通道关联以通过泛素依赖途径限制通道的可用性。
在过去10年间,已经认定并工程化了若干种用于乌谢尔综合征的小鼠模型,其中的7种影响harmonin。这些模型中,仅一种即ush1cc.216g>a模型再次产生了作为人类乌谢尔综合征特征的听觉缺陷和视网膜缺陷。ush1cc.216g>a是敲入小鼠模型,由于点突变类似于在french-acadianush1c患者队列中所见者,该模型影响所有传统harmonin亚型的表达。该突变引入了位于ush1c基因的外显子3末端的隐蔽剪接位点。使用这一隐蔽剪接位点产生了具有35bp删除的框移转录物,并导致缺乏pdz域、pst域和cc域的严重截短的蛋白质的转化。纯合c.216aa敲入小鼠在1月龄时苦于严重的听力损失,而杂合c.216ga小鼠不存在任何异常的表型。c.216aa小鼠的耳蜗组织学显示,在p30时中回和基底回中的纤毛束紊乱、异常细胞行、以及内毛细胞和外毛细胞两者的损失。
本文中证明,含有祖先aav衣壳蛋白的aav成功地转导毛细胞,并驱动harmonin剪接形式的表达和准确定位。此外,本文中证明,在产后早期进行如本文所述的含有祖先aav衣壳蛋白的aav的圆窗注射,成功地复原了纯合c.216aa小鼠的听觉和前庭功能。注射小鼠的听觉功能的恢复与编码野生型harmonin的mrna表达的恢复以及纤毛束形貌和机械感觉转导的保存相关。本文中提供的结果表明,使用如本文所述的含有祖先aav衣壳蛋白的aav进行野生型harmonin的早期再引入可能有用于治疗ush1c。
tmc1/tmc2
在造成耳聋的tmc1中已经鉴别出超过40种截然不同的突变。它们被分为35种隐性突变和5种显性突变。大多数隐性突变造成严重的先天性听力损失(例如,dfnb7/11),但一些也造成迟发性的中度至重度听力损失。所有显性突变均造成进行性听力损失(例如,dfna36),并且在青少年时期的中段发作。特别地,可使用如本文所述的包括anc80衣壳的aav载体递送非突变(例如,野生型)tmc1序列或tmc2序列,从而预防听力损失(例如,进一步的听力损失)和/或恢复听力功能。
用于治疗听力损失的治疗性策略
由于成年哺乳动物耳蜗的感觉细胞缺乏自我修复的能力,当前的治疗性策略(取决于损伤的水平和确切位置)依靠放大(助听器)、更好的声音传播(中耳假体/活性移植物)、或直接的神经刺激(人工耳蜗)来补偿原代感觉毛细胞或螺旋神经节神经元的永久性损害,这些细胞形成听觉神经且将声学信息传达至脑部。尽管这些途径已经是变革性的,但它们在复原对于现代生活非常重要的复杂的人类听力功能来说仍远非最优。具体而言,主要问题仍包括频率选择度有限、声音听起来不正常、以及在嘈杂环境中语音识别受限。
转移至耳蜗的治疗性基因已经被视为相对于当前从老年性和环境诱发的听力损失到遗传形式的耳聋范围内的护理标准而进一步改善。超过300个基因座已经被关联至遗传性听力损失,且对超过70个致病基因有所描述(parker&bitner-glindzicz,2015,arch.dis.childhood,100:271-8)。这些途径在治疗上的成功显著依赖于将外源基因构造安全有效地递送至耳蜗中柯蒂氏器官(oc)中的治疗性细胞靶点。
oc包括两类感觉毛细胞:ihc,其将声音所携带的机械信息转变为传播至神经元结构的电信号;以及,ohc,其用来放大并调谐耳蜗反应,这是复杂听力功能所需的过程。内耳中的其它潜在靶点包括螺旋神经节神经元、螺旋缘的柱状细胞,这些细胞对于维持相邻的盖膜或支持细胞来说是重要的,这些细胞具有保护功能且可被触发以转分化为毛细胞直到新生儿早期阶段为止。
对充满高钾内淋巴液的耳蜗管进行注射,可提供对毛细胞的直接抵达。但是,这一微妙液体环境的改变可扰乱耳蜗内电位,提高了注射相关毒性的风险。环绕耳蜗管、鼓阶和前庭阶的充满外淋巴的空间,可从中耳抵达,或通过卵圆窗膜圆窗膜(rwm)抵达。rwm是进入内耳的唯一非骨性开口,在多种动物模型中可相对早的抵达,且使用这一路径进行的病毒载体给药耐受良好。在人体内,人工耳蜗的安放通常依赖于通过rwm插入手术电极。
先前在器官型耳蜗外植体以及体内的内耳注射中评估aav表型的研究已经导致仅在先天性耳聋小鼠模型中导致部分修复。出乎意料的是,含有祖先aav衣壳蛋白的腺相关病毒(aav)以高效率转导ohc。这一发现克服了低转导速率,而该低转导速率限制了使用传统aav血清型的耳蜗基因疗法的成功发展。如本文所述的含有祖先aav衣壳蛋白的aav,提供了一种用于将内耳基因递送至ihc和ohc以及其它被遗传性听力和平衡障碍连累的内耳细胞类型阵列的有价值的平台。如本文所述的含有祖先aav衣壳蛋白的aav除了提供高转导速率之外,在进行全身性注射时还显示在小鼠和非人灵长动物体内具有相似的安全性,且在抗原性上与循环aav截然不同,就限制传统aav载体效力的事先存在的免疫性而言,提供潜在的益处。
但是,本文中描述组合物和方法,其允许将引导编码tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c)中的一者或多者的多核苷酸表达的核酸(例如,aav载体,诸如包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5))的anc80载体)高效递送至细胞,尤其是内耳内例如耳蜗内的细胞(或耳蜗的细胞或耳蜗细胞)。如本文中所用,内耳细胞是指而非限于,内毛细胞(ihc)、外毛细胞(ohc)、螺旋神经节神经元、血管纹、前庭毛细胞、前庭神经节神经元、和支持细胞。支持细胞是指耳内的不兴奋的细胞,例如,除毛细胞或神经元以外的细胞。支持细胞的示例为许旺细胞(schwanncell)。
将本文所述的一种或多种核酸递送至内耳细胞可用来治疗任何先天性或后天性听力障碍,这些听力障碍典型由部分听力损失或完全耳聋定义。本文所述的方法可用来治疗听力障碍,诸如而不限于,隐性耳聋、显性耳聋、乌谢尔综合征、和其它有症状的耳聋,以及由于创伤或老龄化造成的听力损失。
制作携带特异性转基因的病毒的方法
如本文所述,含有祖先aav衣壳蛋白的腺相关病毒(aav)在将核酸(例如,转基因,包括但不限于,编码下列一者或多者的多核苷酸tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c))递送至内耳细胞中尤其有效,且一类尤其有效的祖先aav衣壳蛋白指定为祖先支架衣壳蛋白anc80,其显示在seqidno:1中。anc80载体是内耳毛细胞靶向的aav的示例,其有利地转导超过约60%、70%、80%、90%、95%、或甚至100%的内毛细胞或外毛细胞。但是,一种落入anc80祖先衣壳蛋白类别内的特定祖先衣壳蛋白是anc80-0065(seqidno:2),以其整体并入本文的wo2015/054653描述了大量落入anc80祖先衣壳蛋白类别内的附加祖先衣壳蛋白。
在特定具体实施例中,腺相关病毒(aav)含有具有天然或工程化的毛细胞趋性的祖先aav衣壳蛋白。在一些具体实施例中,该病毒是内耳毛细胞靶向的aav,其将转基因(例如,编码下列一者或多者的多核苷酸:tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c))递送至受试者的内耳。在一些具体实施例中,该病毒是包含纯化的粘粒多肽的aav。在一些具体实施例中,该病毒是人工的。在一些具体实施例中,该病毒含有祖先aav序列。在一些具体实施例中,该病毒是血清阳性率低于aav2的aav。在一些具体实施例中,该病毒是外显子相关的aav。在一些具体实施例中,该病毒是外显子相关的aav1。在一些具体实施例中,该病毒包含衣壳蛋白,该衣壳蛋白与anc80衣壳蛋白的氨基酸序列一致性为至少95%或与anc80衣壳蛋白同源。
本文所述的含有anc80衣壳蛋白的病毒可用来将多种核酸递送至内耳细胞。在一种具体实施例中,内耳细胞靶向的aav(例如,anc80载体)包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5)),该启动子引导编码下列一者或多者的多核苷酸的表达:tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7ush1c(例如,harmonin-a、harmonin-b或harmonin-c)。用于表达目的而被递送至细胞的核酸序列时常被称为转基因。可被递送至内耳细胞并在该细胞内表达的代表性转基因包括而不限于,编码在听觉和/或前庭机械感觉传导中发挥功能的多肽(例如,tmc1、tmc2、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7(例如,harmonin-a、harmonin-b或harmonin-c))的转基因,编码神经营养因子(例如,gdnv、bdnf或hsp70)、免疫调节蛋白或抗致癌性转录物的转基因。此外,可被递送至内耳细胞并在该细胞内表达的代表性转基因还包括而不限于,编码抗体或其片段、反义、静默或非编码长rna碎片、或基因组编辑系统(例如,基因修饰的锌指核酸酶、转录激活因子样效应子核酸酶(talen)、或成簇规则间隔短回文重复序列(crispr))的转基因。再者,可被递送至内耳细胞并在内耳细胞内表达的代表性转基因包括核酸指定的actg1、adcy1、atohi、atp6v1b1、bdnf、bdp1、bsnd、datsper2、cabp2、cd164、cdc14a、cdh23、ceacam16、chd7、ccdc50、cib2、cldn14、clic5、clpp、clrn1、coch、col2a1、col4a3、col4a4、col4a5、col9a1、col9a2、col11a1、col11a2、crym、dcdc2、dfna5、dfnb31、dfnb59、diaph1、edn3、ednrb、elmod3、emod3、eps8、eps8l2、espn、esrrb、eya1、eya4、fam65b、foxi1、gipc3、gjb2、gjb3、gjb6、gpr98、grhl2、gpsm2、grxcr1、grxcr2、hars2、hgf、homer2、hsd17b4、ildr1、kars、kcne1、kcnj10、kcnq1、kcnq4、kitlg、lars2、lhfpl5、loxhd1、lrtomt、marveld2、mcm2、met、mir183、mirn96、mitf、msrb3、mt-rnr1、mt-ts1、myh14、myh9、myo15a、myo1a、myo3a、myo6、myo7a、nars2、ndp,nf2、nt3、osbpl2、otoa、otof、otog、otogl、p2rx2、pax3、pcdh15、pdzd7、pjvk、pnpt1、polr1d、polr1c、pou3f4、pou4f3、prps1、ptprq、rdx、s1pr2、sans、sema3e、serpinb6、slc17a8、slc22a4、slc26a4、slc26a5、six1、six5、smac/diablo、snai2、sox10、strc、syne4、tbc1d24、tcof1、tecta、timm8a、tjp2、tnc、tmc1、tmc2、tmie、tmem132e、tmprss3、trpn、triobp、tspear、ush1c、ush1g、ush2a、ush2d、vlgr1,wfs1、whrn和xia。在特定的具体实施例中,该转基因是myo7a、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7、ush1c(例如,harmonin-a、harmonin-b或harmonin-c)中的一种或多种。
转基因的表达可由该转基因的天然启动子(即,天然见于该转基因编码序列中的启动子)引导,或者转基因的表达可由异种启动子(例如,cmv启动子、espin启动子、pcdh15启动子、ptprq启动子和tmhs(lhfpl5)启动子)引导。例如,本文所述的任何转基因可与其天然启动子合用。或者,本文所述的任何转基因可与异种启动子合用。如本文中所用,异种启动子是指并非天然地引导该序列表达(即,未天然见于该序列中)的启动子。可用以引导本文中指明的任何转基因的表达的代表性异种启动子包括,例如,cmv启动子、cba启动子、casi启动子、p启动子、ef-1启动子、alpha9烟碱受体启动子、prestin启动子、gfi1启动子、和vglut3启动子。此外,天然地引导上述转基因之一的表达的启动子(例如,kcnq4启动子、myo7a启动子、myo6启动子、或atoh1启动子)可用作异种启动子以引导转基因的表达。在其它具体实施例中,该启动子是espin启动子、pcdh15启动子、ptprq启动子或tmhs(lhfpl5)启动子。
制作用于封装在含有anc80衣壳蛋白的病毒内的转基因(例如,tmc1、tmc2、ush1c(例如,harmonin-a、harmonin-b或harmonin-c)、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1,pdzd7)的方法是该领域中已知的,并使用传统分子生物学技术和重组核酸技术。在一种具体实施例中,提供了包括编码anc80衣壳蛋白的核酸序列的构造和携带两翼带有合适的末端反向重复序列(itr)的转基因的构造,这使得转基因可封装在该anc80衣壳蛋白内。
可使用例如封装宿主细胞将转基因封装在含有anc780衣壳蛋白的aav内。可使用一种或多种如本文所述的构造将病毒颗粒的成分(例如,rep序列、cap序列、反向末端重复(itr)序列)短暂地或稳定地引入封装宿主细胞内。本文所述的病毒至少含有anc80衣壳蛋白;病毒颗粒的其它成分(例如,rep序列、itr序列)可以是基于祖先序列或当代序列的。在一些情况下,例如,整个病毒颗粒可以是基于祖先序列的。此类病毒可使用常规方法纯化。
通常,如本文中所使用,“核酸”可包括dna和rna,且也可包括含有一种或多种核酸类似物或骨架修饰的核酸。核酸可以是单链的或双链的,一般取决于其用途。可用于本文所述方法的核酸可与已知核酸序列一致,或可用于本文所述方法的核酸可在序列上不同于已知序列。举例来说,核酸(或所编码的多肽)与已知序列的序列一致性可为至少75%(例如,至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的序列一致性)。
在计算序列一致性百分比时,将两个序列对齐,并确定两个序列的核苷酸或氨基酸残基之间一致匹配的数目。一致匹配的数目除以对齐区域的长度(即,对齐的核苷酸或氨基酸残基的数目)再乘以100即为序列一致性百分比的值。应知悉,对齐区域的长度可以是一个序列或两个序列的一部分至最短序列的全长度大小。也应知悉,单个序列可与超过一个其它序列对齐,并因此可具有在不同对齐区域内的不同序列一致性百分比的值。
使用计算机程序clustalw和缺省参数实施用以确定序列一致性百分比的两个或多个序列的对齐,该程序允许在待实施核酸或多肽序列的整个长度上进行对齐(全局对齐)。chennaetal.,2003,nucleicacidsres.,31(13):3497-500。clustalw计算查询序列与一个或多个受试序列之间的最佳匹配,并将它们对齐,从而确定一致性、相似性和差异性。可将一个或多个残基的空位插入查询序列、受试序列或两者中,以最大化序列对齐。对于核苷酸序列的成对地对齐,使用缺省参数(即,字长:2;窗口长:4;评分方法:百分比;顶对角线数:4;空位罚分:5);对于多个核酸序列的对齐,使用下列参数:空位开启罚分:10.0;空位延伸罚分:5.0;权重转移:是。对于多肽序列的成对地对齐,使用下列参数:字长:1;窗口大小:5;评分方法:百分比;顶对角线数:5;空位罚分:3。对于多肽序列的多次对齐,使用下列参数:权重矩阵:blosum(模块替换矩阵);空位开启罚分:10.0;空位延伸罚分:0.05;亲水性空位:开;亲水性残基:gly、pro、ser、asn、asp、gln、glu、arg和lys;以及,残基特异性空位罚分:开。例如,clustalw可在互联网的贝勒医学院搜索载入器(baylorcollegeofmedicinesearchlauncher)网站上或在欧洲生物信息研究中心(europeanbioinformaticsinstitute)网站上运行。
可将改变引入核酸序列中,如果该核酸是编码序列,则可导致所编码的多肽的氨基酸序列的改变。例如,可使用诱变(例如,定点诱变、pcr介导的诱变)或通过化学合成具有此类改变的核酸分子而将改变引入核酸编码序列中。此类核酸改变可导致在一个或多个氨基酸残基处的保守的和/或非保守的氨基酸替换。“保守氨基酸替换”是将一个氨基酸残基替代为具有相似侧链的不同氨基酸残基的氨基酸替换(见,例如,dayhoffetal.(1978,《蛋白质序列和结构图谱》(atlasofproteinsequenceandstructure,5(suppl.3):345-352)),其提供氨基酸替换的密码表),而非保守替换是将一个氨基酸残基替代为不具有相似侧链的氨基酸残基。
核酸可包含在亦可称为载体或质粒的构造中。构造可商购,或可通过该领域的常规重组技术生产。含有核酸的构造可具有引导和/或调节此核酸表达的表达元件,也可包括序列诸如用于维持该构造的那些序列(例如,复制起点、可选择的标记物)。表达元件是该领域中已知的且包括,例如,启动子序列、内含子序列、增强子序列、反应元件、或诱导元件。
药物组合物
包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs
(lhfpl5))以及ush1、myo7a、ush1c(harmonin-a、harmonin-b、harmonin-c)、cdh23、pcdh15、sans和cib2中的一种或多种多核苷酸的内耳毛细胞靶向aav(例如,anc805载体),通常悬浮在生理上相容的赋形剂中,可通过经由圆窗注射入受试者内耳中而给药至受试者(例如,人类或非人哺乳动物)。合适的载剂包括盐水,其可使用多种缓冲溶液(例如,磷酸盐缓冲盐水)、乳糖、蔗糖、磷酸钙、明胶、葡聚糖、琼脂、果胶和水配制。以足够的量给药内耳毛细胞靶向的aav,以转导或感染该细胞并提供足够的基因转移水平和表达水平,从而提供治疗性有益效果而没有过度的副作用。
给药至受试者的内耳毛细胞靶向的aav的剂量将主要取决于一下因素:诸如待治疗的病症以及该受试者的年龄、体重和健康状况。例如,待给药至人类受试者的内耳毛细胞靶向的aav的治疗有效剂量通常在约0.1ml至10ml溶液的范围内,其中该溶液含有约1x10'至1x1012基因组拷贝(gc)的aav浓度(例如,约1x103至1x109gc)。
将核酸递送至内耳细胞的方法
将核酸递送至细胞的方法通常是该领域中已知的,而将含有转基因的病毒(其也被称为病毒颗粒)在体内递送至内耳细胞的方法在本文中描述。如本文所述,可将约108至1012个病毒颗粒给药至受试者,且该病毒可悬浮在合适体积(例如,10μl、50μl、100μl、500μl或1000μl)的例如人工外淋巴液中。
可使用任何数量的手段将如本文所述的含有启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5))和转基因(例如,tmc1、tmc2、ush1c(例如,harmonin-a、harmonin-b或harmonin-c)、myo7a、usch1c、cdh23、pcdh15、sans、cib2、ush2a、vlgr1、whrn、clrn1、pdzd7)的病毒递送至内耳细胞(例如,耳蜗中的细胞)。例如,可通过圆窗或卵圆窗典型以相对简单(例如,门诊)的过程注射治疗有效量的包括含有一种或多种不同类型的本文所述转基因的病毒的组合物。在一些具体实施例中,可在外科手术(例如,耳蜗造口术或耳道造口术)过程中,将如本文所述的包含治疗有效量的含有转基因的病毒颗粒或含有一组或多组不同病毒颗粒的组合物递送至耳部的适当位置,其中,一组病毒颗粒中的每个颗粒可含有相同类型的转基因,但各组颗粒含有与其它组不同类型的转基因。
在一种具体实施例中,通过有此需要的受试者的圆窗注射包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5))以及ush1、myo7a、ush1c(harmonin-a、harmonin-b、harmonin-c)、cdh23、pcdh15、sans和cib2中的一种或多种多核苷酸的内耳毛细胞靶向的aav(例如anc80载体)。
此外,促进将剂转移跨过鼓膜和/或通过圆窗的递送媒介(例如,聚合物)是可获得的,且任何此类递送介质可用来递送本文所述的病毒。见,例如,arnoldetal.,2005,audiol.neurootol.,10:53-63。
本文所述的组合物和方法令将核酸高效递送至内耳细胞如耳蜗细胞成为可能。例如,本文所述的组合物和方法能够将转基因递送至并表达在至少80%(例如,至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%)的内毛细胞中,或递送至并表达在至少80%(例如,至少85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%)的外毛细胞中。
如本文所证明,使用含有anc80衣壳蛋白的aav递送的转基因的表达可导致内毛细胞(ihc)、外毛细胞(ohc)、螺旋神经节神经元、血管为、前庭毛细胞、和/或前庭神经节神经元(例如,atoh1、nf2)的再生,使得听力或前庭功能得以长时间(例如,几个月、几年、几十年或余生)复原。
如wo2015/054653中所讨论,含有anc80衣壳蛋白的aav的特征可能在于其血清阳性率和/或其相对于传统aav(即,不含anc80衣壳蛋白的aav)的中性化程度。在该领域中,血清阳性率理解为是指群体中血清阳性受试者(即,已经暴露于特定的病原体或免疫原)的比例,且计算为所检查的群体中产生对抗特定病原体或免疫原的抗体的受试者数除以该群体中的个体总数。确定病毒的血清阳性率在该领域中常规地实施,且典型包括使用免疫检验来确定来自特定个体群体的样品(例如,血液样品)中一种或多种抗体的发生率。此外,可获得几种用以确定血清样品中抗体中和程度的方法。例如,中和抗体试验测量了实验样品含有的抗体浓度相较于没有抗体的对照样品将感染中和50%或更高时的滴度。又见,fisheretal.(1997,naturemed.,3:306-12)和manningetal.(1998,humangenether.,9:477-85)。代表性的传统aav包括而不限于,aav8(或包含aav8衣壳蛋白的病毒)和/或aav2(或包含aav2衣壳蛋白的病毒)。
试剂盒
本发明也提供包含一个或多个填充有本发明药物组合物的一种或多种成分(例如,包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5))以及ush1、myo7a、ush1c(harmonin-a、harmonin-b、harmonin-c)、cdh23、pcdh15、sans和cib2中的一种或多种多核苷酸的内耳毛细胞靶向的aav(例如,anc80载体))的容器的药物包装或试剂盒。与此类容器关联的可以是由政府机构规定的调节药物或生物产品的制造、使用或规格形式的注意事项,该注意事项反映了该机构批准的用于人类给药的制造、使用或规格。
本发明也提供用于治疗或预防与听觉和/或前庭机械感觉传导中的缺陷相关的疾病或其病变(或症状)。一个具体实施例中,试剂盒包括单位剂型的有效量的内耳毛细胞靶向的aav(例如,anc80载体),该aav包含启动子(例如,cmv、espin、pcdh15、ptprq、tmhs(lhfpl5))和作为下列一者或多者的多核苷酸:ush1、myo7a、ush1c(harmonin-a、harmonin-b、harmonin-c)、cdh23、pcdh15、sans和cib2;同时还包括将抑制血管生成的化合物给药至苦于或易患与血管生成相关的疾病或病变或症状的受试者的使用说明书,其中该有效量的抑制血管生成的化合物是少于500mg的化合物。在优选的具体实施例中,该试剂盒包含含有抑制血管生成的化合物的无菌容器;此类容器可为盒、安瓿、瓶、小瓶、管、袋、包、或其它该领域中已知的适合的容器形式。此类容器可由塑料、玻璃、层压纸、金属箔或其它适用于容纳药物的材料制作。使用说明书通常将包括关于该抑制血管生成的化合物用于治疗血管形成相关疾病或病变或症状的用途的信息;在优选的具体实施例中,使用说明书包括下列至少一者:对该抑制血管生成的化合物的说明;用于治疗与血管生成相关的疾病或病变或其症状的剂量方案和给药途径;预防措施;警告;适应症;禁忌症;剂量过量信息;副作用;动物药理学;临床研究;和/或参考文献。该说明书可直接印在容器(当存在时)上,或作为用于该容器的标签,或作为独立的页、册、卡或折叠式印刷品提供在给容器内或与该容器一起提供。
可根据本公开使用该领域中熟悉的传统分子生物学、微生物学、生物化学和重组dna技术。此类技术在文献中完整地诠释。在下述实施例中将进一步描述本发明,这些实施例并不限制权利要求书中描述的方法和物质组合物的范畴。
[实施例]
实施例1:含有祖先aav衣壳蛋白的腺相关病毒(aav)导致安全有效的耳蜗基因转移
下述方法和材料用于实施例1中。
病毒载体
如先前所述,在马萨诸塞州眼耳医院(massachusettseyeandear)的基因转移载体中心(genetransfervectorcore)(vector.meei.harvard.edu)制备了具有cmv驱动的egfp转基因和woodchuck肝炎病毒转录后调节元件(wpre)盒的aav2/1、2/2、2/6、2/8、2/9和aav2/anc80。aav2/anc80质粒试剂通过addgene.com获得。
动物模型和通用方法
所有实验均得到波士顿儿童医院(bostonchildren’shospital)(条款#12-02-2146)和生物安全委员会机构(institutionalbiosafetycommittee)(条款#ibc-p00000447)的批准。野生型c57bl/6j小鼠和cba/caj小鼠从杰克逊实验室(jacksonlaboratory(barharbor,me))获得,且用于实验的每种性别的动物预计为50/50比率。通过所获取的标本和技术可行性确定用于体外和体内转导试验和后续端点的每一实验的组内动物数。在后续实验中,使用大量载体定性地验证所报告的对于anc80转导的观察(但由于获取标本的独特性和天然限制性,人类前庭组织转导除外)。由于获取标本受限和所报告的发现的定量特性受限,没有实施血清型转导效率之间的统计学分析。
实施例1a:体内注射
使用坡口玻璃显微注射穿刺针经由圆窗膜对幼鼠(p0至p2)注射。在p-2000穿刺针拉制仪(sutterinstruments,novato,ca)上从毛细玻璃管(wpi)拉出穿刺针,并使用微穿刺针坡口机(sutterinstrument,novato,ca)切出坡口(~20μm尖端直径,28°角)。使用无菌拭子在外部施加用于镇痛的emla乳霜(2.5%利多卡因和2.5%丙胺卡因),以覆盖手术部位(左侧乳突状突出)。手术前,在38℃取暖垫上维持体温。通过将幼鼠在冰/水中快速降低体温2至3分钟而麻醉直至失去意识为止,并在手术过程中在冷却平台上将这一状态维持5至10分钟。使用碘伏(betadine)刷洗并使用70%乙醇擦拭,重复三次,将手术部位消毒。作出耳后切口以暴露透明的耳泡,手动操纵微穿刺针前行通过耳泡和上覆筋膜,并借由微穿刺针的尖端穿透rwm。在1分钟内,手动将大约1μl的病毒单侧注射入5只(aav1)、4只(aav2)、2只(aav8)、1只(aav6)、3只(anc80)c57bl/6小鼠的左耳内。为了控制与具体载体制剂相关的因素诸如品质和纯度,在后续研究中使用不同的来自独立制剂的大量载体证实anc80的结果,证实了此处呈递的我们的定性发现(数据未显示)。以非盲模式实施各组的注射。注射针偶尔插入过深、过浅或角度错误。如果存在肉眼可见的对中耳或内耳结构的损伤,则将样本从进一步的分析中剔除。注射成功率介于~50%至~80%之间,取决于注射人员的经验水平。注射后,使用6-0黑色单丝缝线(surgicalspecialties,wyomissing,pa)闭合皮肤切口。然后将幼鼠送回38℃取暖垫上,放置5至10分钟,随后放回其母亲处饲养。
与先前的报告一致,aav1以中至高效率转导ihc(图1a、图1b)。这些研究表明,aav2、aav6和aav8以少量的ihc为靶点,仅aav8演示了在顶部和基底部的大致等效的转导(图1b)。而且,与先前的报告一致,对于所测试的全部传统aav血清型,这些都是最小ohc转导(<5%)。但是,anc80在低20倍(对于aav1)至3倍(对于aav2)的剂量下转导了接近100%的ihc和~90%的ohc(图2a至图2c)。在1.36x1012gc的相等剂量下对所有血清型进行转导,结果,anc80导致了实质性的ihc和ohc转导,但aav1、aav2和aav8的ihc靶向最低且无一在ohc中出现值得注意的转导,如通过由落射荧光显微镜进行的活体细胞成像所观察的(图8c、图8d)。
随后将anc80转导的样本固定,染色,并通过共聚焦显微镜成像,表明了毛细胞转导的剂量依赖性(图1e)。空前的ohc靶向(图1c、图3)示出了anc80相较于其它aav的定性上截然不同的转导生物学。在总计三只注射anc80的小鼠体内,发现了遍及从基底部到顶部的耳蜗的相似水平的anc80转导(图1a、图1b、图1c)。耳蜗顶部的低倍率图(图3a)显示了远离注射部位的强egfp表达。基底部的高倍率图像显示了100%ihc和95%ohc转导(图3b)。
由于一些形式的遗传性耳聋也造成前庭功能障碍,anc80可能是可用于将基因递送至人类前庭器官的载体。为了评估之一可能性,从四位进行前庭神经鞘瘤切除术的成年患者采集人前庭上皮;将感觉上皮置于培养中,如先前所述。对于aav转导的样本,图3c表明了遍及人前庭上皮的毛细胞和支持细胞两者中的强egfp荧光。图3d中,使用myo7a复染的上皮的高倍率图显示,83%(19/23)的myo7a阳性毛细胞也是egfp阳性的,表明anc80可有效地转导鼠和人的毛细胞。
实施例1b:毛细胞电生理学
切除耳蜗,将其封固在盖玻片上,并使用配备63x水浸物镜和微分干涉相称光学系统的axioexaminer.a1直立式显微镜(carlzeiss,oberkochen,germany)观察。在室温(22℃至24℃)下,在含有下列(以mm计)的标准溶液中实施电生理记录:137nacl、5.8kcl、10hepes、0.7nah2po4、1.3cacl2、0.9mgcl2、以及mem中的5.6d-葡萄糖、维生素(1:100)和氨基酸(1:50)(lifetechnologies,carlsbad,ca)(ph7.4;~310mosm/kg)。从r-6玻璃(kingprecisionglass,claremont,ca)拉出记录电极(3至4mω),并充满含有下列(以mm计)的细胞内溶液:140cscl、5egta-koh、5hepes、2.5na2atp、3.5mgcl2和0.1cacl2(ph7.4;~280mosm/kg)。使用高阻抗密封膜片箝全细胞技术,使用axopatch200b(moleculardevices,sunnyvale,ca)记录机械感觉转导电流。将毛细胞保持在-84mv。使用低通量bessel滤波器在5khz将电流滤波,使用12位采集板(digidata1440a,oleculardevices,sunnyvale,ca)在≥20khz数字化,并使用pclamp10软件(moleculardevices,sunnyvale,ca)记录。使用封固在picma芯片压电执行机构(physikinstrumente,karlsruhe,germany)上、由lvpzt放大器(e-500.00,physikinstrumente,karlsruhe,germany)驱动并使用8极bessel滤波器(3384型滤波器,krohn-hitecorporation,brockton,ma)在40khz滤波以最小化残留吸管共振的硬玻璃探针,令ihc和ohc的纤毛束偏转。硬玻璃探针设计为恰好嵌入用于全束记录的毛细胞静纤毛阵列的凹面内(用于ohc时,直径为3至4μm;用于ihc时,直径为4至5μm)。对于在>p10时的全细胞电生理学记录,在p5至p7切除耳蜗组织并将其在具有1%fbs的mem(1x)+glutamaxtm-i培养基中在37℃和5%co2下温育长达30天。
来自p7ohc和p35ihc的由纤毛束偏转诱发的代表性电流显示,在egfp阳性和egfp阴性对照细胞的波幅、灵敏度或动力学之间没有差异(图1d)。在暴露于anc80中之后,从耳蜗全部区域以及从1至5周龄之间记录到51个egfp阳性和52个egfp阴性毛细胞。在所有情况下,野生型的反应是不能区分的(图1e),这证实了anc80转导对于感觉细胞功能没有损害效应。
实施例1e:听力测试
如先前所述,收集听性脑干反应(abr)和畸变产物耳声发射(dpoae)数据。dpoae是进行适宜耳蜗放大和调谐的试验,且是外毛细胞生存能力的灵敏量度。在被麻醉小鼠体内测试的刺激以5.6、8、11.3、16、22.6和32khz的频率在10至90db声压级之间变动。在p28至p30,分析了4只注射anc80的耳朵、4只未注射的耳朵、和1只具有注射损伤而没有egfp荧光的阴性对照耳朵。
绘制诱发abr所需的最小声音阈值(图1f),显示在注射耳与未注射耳的阈值之间没有差异。组织学分析表明,在4只注射耳中均有强烈的egf荧光(数据未显示)。在一种情况下,没有egfp阳性细胞且没有评估abr阈值(图1f),表示注射失败且注射针可能已经穿透耳蜗管并造成永久性损害。尽管由anc80-egfp造成的外毛细胞转导强健,但未见相对于未注射的对照耳的dpoae差异(图1g)。因此,来自abr和dpoae的数据表明,对于听觉安全来说,rwm注射、anc80转导以及ihc和ohc中的转基因表达都是安全的。
实施例1f:转棒测试
在转棒装置上测试五只c57bl/6小鼠的平衡行为。已知前庭功能受损的小鼠极难完成转棒测试。先前的研究突出了这一转棒测试检测当一只耳朵受影响时的平衡功能障碍的能力。三只在p1注射的小鼠在p36测试,而两只未注射的对照小鼠在p79测试。使用下述转棒方案测试所有小鼠。第一天,在以4rpm转动的转棒上对小鼠进行五分钟的平衡训练。第二天,对小鼠进行五次试验,每两次试验之间间隔五分钟。对于每次试验,棒的转速均从最初的2rpm速率增加1rpm。记录到小鼠从装置上摔落为止的时间(秒)。
由于耳蜗的外淋巴液与前庭迷路的外淋巴液是连续的,评估经由耳蜗rwm注射的anc80-egfp是否会转导前庭感觉器官。事实上,前庭上皮的整装包埋(wholemount)显示了在椭圆囊、对重力和头部线性移动敏感的前庭器官、以及对头部转动敏感的半规管中的i型和ii型两种毛细胞中的强健的egfp表达(图2a、图2b)。因此,为了解决anc80转导可能影响平衡性的安全问题,对前庭表达已经得以证实的经注射小鼠实施与未注射的对照小鼠相似的前庭功能转棒测试(图4)。
第二部分:基因疗法复原了1c型乌谢尔综合征小鼠模型的听觉和前庭功能
实施例2:乌谢尔综合征的小鼠模型
下述方法和材料用于实施例2中。
组织制备
在出生后0至8天(p0至p8),从ush1cc.216g>a杂合或纯合突变小鼠采集椭圆囊和柯蒂氏器官进行电生理学研究。通过快速斩首杀死出生后的幼鼠。切除颞骨并将其浸泡在以10mmhepes(ph7.4)补充的mem(invitrogen,carlsbad,ca)中。如先前所述,不使用酶而解剖柯蒂氏器官(53)。在以0.1mg/ml的蛋白酶(proteasexxiv,sigma)处理10分钟后,移除椭圆囊。将切除的器官封固在圆形盖玻片上。将事先用胶水粘在该盖玻片上的一对薄玻璃纤维置于组织边缘上以将其稳定在平坦位置。组织很快被使用,或保存在具有1%胎牛血清的培养物中。将培养物维持7至8天,每2至3天更换培养基一次,以便进行牵涉体外病毒载体感染的实验。
动物
从路易斯安那州立大学健康科学中心获得ush1cc.216g>a敲入小鼠。事先将在c57bl6背景上引入的种系培育出造成老年性听力损失的cdh23(ahl)突变(48、49)。用于这一工作的所有过程均符合nih实验动物护理和使用指南,并且得到波士顿儿童医院的动物护理和使用委员会机构的批准(条款#12-02-2146、#14-03-2659r和#15-01-2878r)。如先前所述,使用趾夹(p8之前)或耳标(p8之后)将小鼠基因分型,并实施pcr(32)。对于全部研究,均大约以相等的比例使用雄性和雌性小鼠。没有另外应用的随机范式。
病毒载体生成
从c.216aa突变小鼠的耳蜗中单离出总rna(rnaqueous微量试剂盒,ambion),并使用quantitect逆转录试剂盒(qiagen)逆转录。使用高保真度platinumtaqdna聚合酶(invitrogen)和下列引物通过pcr扩增trunc-harmonin的cdna:trunc-harmonin.f(kpni)gaggtaccatggaccggaaggtggcccgag;trunc-harmomin.rv(bamhi)caggatccggacaatttcatcccctac。使用ta克隆试剂盒(invitrogen)克隆387bppcr产物,并测序证实。为了生成gfp融合构造,使用kpni和bamhi将截短的harmonin片段亚克隆入pegfp-c1中。将nhei-xbaiegfp::trunc-harmonincdna转移至aav穿梭载体内。使用aav2末端反向重复序列(itr)将定制载体封装在aav1衣壳内,其中转基因盒由cmv启动子(aav2/1.cmv.egfp::trunc-harmomin.hgh,1.92e14gc/m,bch)驱动。
在我们的实验室中,从lilyzheng和jamesbartles(52)(西北大学范伯格医学院细胞与分子生物学系(departmentofcellandmolecularbiology,northwesternuniversity,feinbergschoolofmedicine,chicago,il))慷慨提供的egfp标签标记的构造制备harmonin-a1和harmonin-b1质粒。harmonin-a1的原始来源为小鼠肾脏,而harmonin-b1则从小时耳蜗感觉上皮单离。我们进一步修饰该harmonin-a1构造以使用tdtomato替换其n端的egfp标签。将经荧光标记的构造和未标记的构造封装在aav载体中。通过波士顿儿童医院的病毒核设施和马萨诸塞州眼耳医院的基因转移载体核生成病毒载体。生成下述载体:aav2/1.cmv.tdtomato::harmonin-a14.3310^13gc/ml(bch);aav2/1.cmv.egfp::harmonin-b12.7356410^14gc/ml(bch);aav2/1.cmv.egfp-harmonin-a1:2.8110^12gc/ml(meei);aav2/1.cmv.egfp-trunc-harmonin:1.9210^14gc/ml(bch);aav2/anc80.cmv.harmonin-a1:1.9310^12gc/ml(meei);aav2/anc80.cmv.harmonin-b1:1.7410^12gc/ml(meei);aav2/anc80.cmv.trunc-harm.wpre:9.0256710^12gc/ml(meei);对于体外实验,将10μl的浓缩载体施加至1mlmem补充培养基,将其在1%胎牛血清的存在下在紧急切除的组织上放置24h。然后将培养物维持长达10天。
圆窗膜(rwm)注射
如波士顿儿童医院的动物护理和使用委员会机构所批准(条款#15-01-2878r),实施rwm注射。将0.8μl至1ul的aav载体注射到p0至p1和p10至p12的新生小鼠体内。首先使用低温暴露麻醉p0至p1小鼠,而p10至p12小鼠使用异氟烷麻醉。麻醉后,作出耳廓后切口以暴露鼓泡并令耳蜗可视化。使用由显微操控器控制的玻璃微穿刺针通过rwm进行注射(askewetal.2015)。将所注射材料的体积控制为大约0.02μl/min,注射10min。施用标准术后护理。在持续的基础上确定用于体内研究的样本尺寸,以优化该样本尺寸并降低方差。
电生理学记录
在含有下列(以mm计)的标准人工外淋巴液中实施记录:144nacl、0.7nah2po4、5.8kcl、1.3cacl2、0.9mgcl2、5.6d-葡萄糖和10hepes-naoh,调节为ph7.4和320mosmol/kg。加入来自浓缩物(invitrogen,carlsbad,ca)的维生素(1:50)和氨基酸(1:100)。使用配备具有微分干涉相称光学系统的63x水浸物镜的直立式axioskopfs显微镜(zeiss,oberkochen,germany)从顶面观察毛细胞。从硼硅酸盐玻璃毛细管(garnerglass,claremont,ca)拉出记录穿刺针(3至5mω),并充满含有下列(以mm计)的细胞内溶液:135kcl、5egta-koh、10hepes、2.5k2atp、3.5mgcl2、0.1cacl2,ph7.4。在室温下,在-64mv的保持电位的全细胞电压钳下记录电流。使用axopatchmulticlamp700a或axopatch200a(moleculardevices,paloalto,ca)撷取数据,使用低通量bessel滤波器在10khz滤波,使用12位采集板(digidata1322)和pclamp8.2及10.5(moleculardevices,paloalto,ca)在≥20khz数字化。使用originlab软件在线下分析数据,且除非明确标注,否则表示为均值±标准偏差。
统计学分析
在每个时间点,以每组至少三只小鼠评估测试载体和对照载体,以确保可重现性。样本尺寸标注在图注说明中。所有成功进行rwm注射的动物均包括在研究分析中。那些未成功注射的动物在计算均值时排除,但包括在完整公开的图注说明中。根据阈值>90dbspl的abr恢复来确定注射成功。使用origin2016(originlabcorporation)实施统计学分析。如文本和图注说明中所标注的,数据表示为均值±标准偏差(s.d)或均值的标准误差(s.e.m)。使用单向方差分析(anova)确定均值之间的显著差异。
实施例2a:扫描电子显微镜检查法(sem)
在p7、p18和~p42(6周)实施对照小鼠和突变小鼠的柯蒂氏器官的sem。与华盛顿大学的edwinrubel博士合作实施p18sem。将内耳在4℃的0.1m磷酸钠中以4%戊二醛中固定过夜。第二天,将标本在0.1m磷酸钠缓冲液(pb)中冲洗三次,并在冰浴中在0.1mpb中的1%四氧化锇中再固定30分钟。然后,将标本在0.1mpb中冲洗,并通过下列浓度梯度的乙醇脱水:35%、70%、95%、和100%(x2)。将样本进行临界点干燥,封固在sem载物台上,并溅射涂覆au/pd。使用jeoljsm-840a扫描电子显微镜实施sem。由géléoc博士和indzhykulian博士在p8和6周阶段实施类似的处理。在室温下,将柯蒂氏器官外植体在以2mmcacl2补充的0.1m二甲砷酸盐缓冲液(electronmicroscopysciences)中以2.5%戊二醛固定1小时。将标本在梯度浓度的丙酮中脱水,从液体co2临界点干燥,溅射涂覆4至5nm的铂(q150t,quorumtechnologies,unitedkingdom),并使用场发射扫描电子显微镜(s-4800,hitachi,japan)观察。
纯合c.216aa突变小鼠是聋的,并显示前庭功能障碍的转圈和摇头行为特征。来自lentz等人的先前工作(34)描述了在p30时耳蜗基底部的显著的内毛细胞和外毛细胞退化。在中回内也观察到了退化和毛细胞死亡,而该器官顶部在1月龄时保持良好。假设毛细胞退化在内耳器官发育过程中进行性地出现,且为了评估毛细胞在较早阶段的生存率,在p8和p18对柯蒂氏器官实施sem分析。杂合c.216ga小鼠的外毛细胞(ohc)和内毛细胞(inc)得以保持,且它们的束在这些阶段适宜地取向(图5a至图5c、图5g、图5i、图12a至图12c、和图12k)。但是,在所分析的两个阶段,在纯合c.216aa小鼠的柯蒂氏器官的整个长度上均证明有紊乱的纤毛束(图12d至图12f、图12h、图12j至图12l、图19d至图19i、和图19l)。在p8,基底部区域、中部区域和顶部区域的ihc束轻度紊乱(图12d至图12f、图12j)。大量ihc束展现了静纤毛行的波纹状图案和轻度紊乱(图12j)。尽管c.216aa突变小鼠的很多ohc具备得以良好保持的纤毛束(图12h、图12k),该器官中仍广泛存在片段化的和紊乱的纤毛束(图12d至图12f、图12l)。在p18时扰乱更为明显,但大多数毛细胞仍如先前报导而存在(35)(图12d至图12f)。
为了评估已经使用harmonin-b1进行基因疗法的小鼠体内的纤毛束形貌,制备6周龄未治疗(或未注射)小鼠和经治疗(或经注射)小鼠的颞骨进行sem分析。在未治疗的c.216aa小鼠的该器官基底部区域和中部区域展现了严重的毛细胞损失(图11)。在基底部区域,ohc在第一行中大量缺席但广泛存在于第二行和第三行中。在该器官的中部区域,第一行ohc也大量缺席。在顶端观察到较轻的表型。高倍率sem也显示了沿着c.216aa突变小鼠的该器官整个长度的严重紊乱的纤毛束。明显地,在6周龄c.216aa小鼠体内,没有观察到保留具有三行静纤毛的典型阶梯状结构的纤毛束。相反,来自c.216aa小鼠的毛细胞展现了具有沿着第一行具有可伸缩的静纤毛的紊乱纤维素、异常的第二行和还算得以保持的最高行。相比之下,在经harmonin-b1治疗后的c.216aa小鼠体内观察到了降低的毛细胞损失和正常的纤毛束。从代表性的视野中纤毛束的存在或不存在预计毛细胞计数。数据表明,在经注射的小鼠体内,从该器官的基底部至顶部,毛细胞数目得以显著保持,在基底部为40%至79%,中部为68%至95%,且在顶部为93至99%(n=1824个细胞,来自n=4只c.216aa小鼠耳朵;n=792,来自n=2修复的c.216aa耳朵)。尽管在注射harmonin-b1的小鼠体内仍存在不正常的纤毛束,但大多数纤毛束具备三行静纤毛且具有几乎与其杂合对照体无差别的形貌(图11)。
实施例2b:fm1-43成像
将5毫摩尔的fm1-43(invitrogen)稀释在细胞外记录溶液中,施加至组织10秒,随后在细胞外记录溶液中洗涤3次以移除过量染料并防止经由内吞作用吸收。5分钟后,使用落射荧光光源、微分干涉相称光学系统、和fm1-43滤光套件(chromatechnologies)在具有20x、40x和63x水浸物镜的zeissaxioscopefsplus上对细胞内fm1-43成像。使用ccd相机和argus-20图像处理器(hamamatsu)使用背景荧光减法获取16位的图像。对全部图像的获取均维持相同的增益和对比度设定,并使用adobephotoshop或image-j软件在线下分析。
为了评估较早阶段的毛细胞功能,在p4分析紧急切除的内耳器官中的fm1-43吸收。简短应用(<10s)时,fm1-43渗入具备功能性机械感觉通道的毛细胞(36、37、38)。在c.216ga小鼠的毛细胞中观察到均匀的fm1-43吸收(图6a),但c.216aa小鼠的ohc吸收水平之间有所变动,表明一些但非全部细胞保留了功能性转导通道(图6b)。沿着耳蜗整个长度做出类似的观察。没有注意到音频差异。在产后第一周的c.216aa小鼠的ihc中的fm1-43吸收也有所降低(数据未显示)。也评估了突变小鼠的椭圆囊毛细胞中的fm1-43吸收。有趣的是,在c.216aa突变小鼠体内,在p6时吸收被限制在横纹外区域,表明横纹区域的毛细胞缺少静息时开放的机械感觉通道(图6c、图6d)。
实施例2c:机械刺激
ohc和ihc:经由封固在一维524picma芯片压电致动器(physikinstruments,waldbronn,gernamy)上且由400maenv400放大器(piezosystemjenagermany,54)驱动的硬玻璃探针传输机械刺激。将探针尖端烧制抛光(火抛光器,h602,worldprecisioninstrumentsinc.,sarasota,fl)以嵌入静纤毛束(51)。通过施加电压阶跃诱发偏转,使用8极bessel滤波器(khron-hite,528brockton,ma)在50khz滤波以消除残余的吸管共振。使用c2400ccd相机(hamamatsu,japan)监控纤毛束偏转。使用电压阶跃来校准刺激探针在其静息位置的±2μm左右的运动。记录探针的视频图像,以证实偏轴运动的不存在并校准探针运动(空间分辨率为~4nm)。探针的10%至90%上升时间为~20μsec。
vhc:经由封固在双压电晶片元件上的硬玻璃探针传输机械刺激。通过将动纤毛轻柔地吸入该刺激穿刺针内而实施偶合。通过将电压阶跃施加至压电装置而诱发偏转,其中该压电装置由依次封固在刺激探针上并与该探针直接偶合的两个双压电晶片组成。通过pclamp8.0软件控制电压阶跃,并使用8极bessel滤波器在1khz(khron-hite,brockton,ma)滤波。使用c2400ccd相机(hamamatsu,japan)监控纤毛束偏转。在实验之前,将刺激探针的运动校准为大约(±2μm)在其静息位置。
在产后第一周期间,听觉和前庭上皮保留机械敏感毛细胞,包括一些具有相对正常形貌的细胞(图5)。在柯蒂氏器官中,从来自p3至p6的c.216aa小鼠耳蜗的中回和顶回的具有看起来正常的束的毛细胞和那些具有更严重紊乱的纤毛束的毛细胞获得记录。在c.216aa突变体中,ohc保留机械敏感性,但反应幅度显著降低~63%,达到170±80pa(n=24;p<0.001,图6e、图6f、图6g)。在c.216aa小鼠的ohc中观察到了宽范围的反应幅度,介于31和292pa之间。当根据纤毛束形貌将数据分组时,观察到了显著差异(p<0.01):在具备严重紊乱的束的突变体毛细胞内诱发的电流比在具有更多被保持的纤毛束的突变体细胞内诱发的电流小,分别为120±65pa(n=9)和201±74pa(n=15)。尽管电流幅度有所下降,仍保留与杂合c.216ga类似的对于机械位移的毛细胞反应。使用二阶玻尔兹曼方程拟合刺激反应[i(x)]曲线(图6f),该拟合用来确定10%至90%操作范围(图13b)。在从c.216ga和c.216aa记录的ohc之间没有观察到操作范围的显著差异(p=0.054)。类似地,尽管来自c.216aa突变小鼠的ihc的纤毛束在dic显微镜下看起来轻度紊乱,但转导电流在p6时显著降低(图13e、图13f、图13g)。在-64mv的保持电位下,杂合c.216gaihc(p6至p7)的最大转导电流平均为587±96pa(n=21),但c.216aaihc中的最大转导电流降低46%,达到316±127pa(n=19;p<0.001)。在c.216aa突变小鼠的ihc中测量到操作范围的显著(p<0.01)减小(图13g)。
定义为转导电流在恒定束偏转存在时衰退的适应(adaptation)也存在于c.216aa突变小鼠体内。使用双指数拟合以确定快成分和慢成分来分析适应动力学。虽然两个成分在来自c.216aa突变小鼠的ihc和ohc中均较慢,但仅仅慢成分的差异是显著的(在ohc中,p<0.05,且在ihc中,p<0.001;图13c、图13d、图13h、图13i)。另一方面,在c.216aa的ohc和ihc中在popen=0.5时测量的适应程度显著低于在c.216ga毛细胞中所测(图20e、图20j;p<0.001)。综上,这些结果表明,在c.216aa小鼠的内毛细胞和外毛细胞中,机械敏感性轻度受损,且重要的是,两种类型的细胞在整个产后第一周期间存活,这是进行基因疗法和细胞功能修复的先决条件。
在c.216aa小鼠的前庭毛细胞中,也观察到了机械感觉转导电流的下降。在横纹外区域,与其数值为231±53pa(n=8,p6至p7)的c.216ga的电流相比,c.216aa的电流显著(p<0.001)下降至109±30pa(n=9,p5至p7)(图6e、图6f、图6h)。从横纹区域的毛细胞记录到非常小的电流或没有电流(6±13pa,n=6,p5至p7),与该区域中fm1-43吸收的缺失相符(见下文;图6c、图6d)。尽管通过dic显微镜观察,椭圆囊纤毛束看起来很好地得以保持,但来自横纹外区域和横纹区域毛细胞的转导电流分别显著降低或不存在。因此,除了横纹区域之外,这些结果说明,突变小鼠体内的转导装置得以正确地组装并作为靶点,但新生小鼠体内的功能配合物数目减少。
之后评估了暴露于aav载体的c.216aa毛细胞中驱动harmonin表达的功能。为了提升使用外源性harmonin进行功能复原的可能性,将由cmv启动子驱动的无标签harmonin-a1或harmonin-b1编码序列封装在被称为anc80的aav衣壳中(39)。最近,已经显示anc80衣壳在体内转导100%的ihc和80至90%的ohc(40)。我们假设,harmonin-b为ihc和ohc两者中的机械感觉转导所需,且对于两种类型细胞的听觉功能是必要的。分别实施aav2/anc80.cmv.harmonin-b1(0.8μl,1.9×10^12gc/ml)和aav2/anc80.cmv.harmonin-a1(1.7×10^12gc/ml)+aav2/anc80.cmv.harmonin-b1(0.5μl+0.5μl)混合物的rwm注射,并在治疗后2周评估机械感觉转导反应。在耳蜗尚未骨化的p5至p6时提取组织,并将其在培养物中维持10天。虽然成熟的ohc(>p10)在间接体内记录模式下不存活,但在等效的p14至p16时从ihc获得了稳健的电生理学记录。结果呈现在图8中。尽管来自未注射小鼠的ihc展示了在p16时严重降低的转导电流(79±43pa,n=8),但在接受aav治疗的小鼠体内发现了感觉转导恢复的证据。在p1时注射harmonin-b1或注射harmonin-b1和harmonin-a1的组合的小鼠体内观察到显著的恢复(***p<0.001),平均最大转导电流分别为388±66pa(n=15)和352±28pa(n=7;图8c)。在使用harmonin-b1治疗后的ihc中的转导电流幅度与对照c.216ga小鼠没有显著差异。恢复水平未通过共同注射harmonin-b1和harmonin-a1而得以显著改变。这些结果表明,在早期阶段经由rwm注射而递送外源性harmonin-b1可修复inc中的机械感觉转导。
实施例2d:共聚焦成像
为了从p0至p8的产后小鼠制备用于共聚焦成像的组织,使用4%多聚甲醛(pfa)进行15分钟的固定。使用0.01%triton透化,并使用alexafluor鬼笔环肽(invitrogen,1/200)复染以标记肌动蛋白长丝。在lsm700zeiss共聚焦显微镜上获得图像。在较年长的小鼠(4至8周)中,在安乐死后移除颞骨并在4%pfa中放置1小时,之后使用120mmedta进行24至36小时的脱钙作用。随后切除感觉上皮,并如上述者注射进行免疫染色。施加鼠抗ctbp2(bdbioscience#612044,1/200)48小时,使用alexafluor山羊抗鼠(1/200)在4℃复染过夜以标记带状突触。在zeisslsm710激光共聚焦显微镜(iddrcimagingcoregrantp30hd18655)上撷取图像,并使用zeisslsm图像浏览器4.2处理。
先前的工作显露了两种选择性剪接形式的harmonin在感觉毛细胞中的表达。为了评估aav载体驱动外源性harmonin剪接形式的表达的能力,将来自新生c.216aa小鼠和野生型(c57bl/6j)小鼠的椭圆囊和柯蒂氏器官暴露于aav2/1载体,该载体编码融合至harmonin-b1的n端的egfp(egfp::harmonin-b1)或融合至harmonin-a1的181n端的tdtomato(tdtomato::harmonin-a1)。在p1,或在体外或在体内通过rwm注射(1μl)施加该载体。当在体外施加时,将p0至p1的组织在该载体的存在下温育24小鼠,并在培养物中维持一周。共聚焦图像显示,野生型小鼠、c.216ga小鼠和c.216aa小鼠的毛细胞被成功转导(图7a至图7c、图7e)。egfp::harmonin-b1信号在vhc(图7a)、ihc和ohc(图7b、7c)中的静纤毛的尖端被证实。在p1时注射的小鼠耳蜗的基底部的ohc和ihc中,也在p60时检测到了egfp信号(图14d)。在听觉毛细胞的基底部检测到tdtomato::harmonin-a1(图14e)。使用带状突触标记物ctbp2进行的共染色常常显露在p7ihc中的共定位(图14e),但在p7椭圆囊中不显露共定位(数据未显示)。外源性融合构造的定位与先前工作一致,harmonin-b被定位在静纤毛的远端且接近尖端细丝插入(tip-linkinsertion)(26、27、28),而harmonin-a被定位在突触(30、31)。
实施例2e:听脑干反应(abr)和畸变产物(dpoae)
从使用甲苯噻嗪(5至10mg/kgi.p.)和氯胺酮(60至100mg/kgi.p.)麻醉的小鼠记录abr和dpoae。将皮下针电极插入以下部位的皮肤内:a)两耳之间的背侧(参考电极);b)左耳廓后(记录电极);和c)动物尾部背侧(接地电极)。切掉耳廓基底部的管道以暴露耳道。对于abr记录,耳道和听力仪器(epl声学系统,meei,boston)给出了5毫秒的短音。使用基于pc的数据采集系统(epl,耳蜗功能测试套件,meei,boston)的模数转换板将反应放大(10,000倍),滤波(0.1至3khz),并平均。以5至10db的阶跃,将声音水平从0升高至110db声压级(分贝spl)。在每一水平,将512至1024个反应在“去伪”后平均(使用交替刺激极性)。通过目测确定阈值。使用origin-2015(originlabcorporation,ma)分析数据并绘图。除非明确排除,否则阈值表示为平均值±标准偏差。对于dpoae,f1和f2主音(f2/f1=1.2)呈现为,f2以半音级阶跃在5.6至45.2khz之间变动,且l1–l2=10dbspl。在每个f2,l2均以10dbspl增量在10至80dbspl之间变动。从平均谱将dpoae阈值定义为诱发超过本底噪声5dbspl量级的dpoae的l2水平。在所有频率下,平均本底噪声均低于0dbspl。使用24位i–o数位板(nationalinstrumentspxi-4461)在pxi-1042q底盘中生成刺激,通过sa-1扬声器(tucker–davistechnologies,inc.)放大,并从两个静电驱动器(cuicdmg15008-03a)在我们的定制声学系统中递送。使用位于小电极管末端的驻极体传声器(knowlesfg-23329-p07)监控耳道声压。这些实验大多数并非在盲测条件下实施。
为了确定截短的harmonin是否干扰正常的听觉功能,生成anc80.cmv.trunc-harm载体以过表达该截短蛋白质。经由rwm将载体注射到c.216ga小鼠的内耳中。在4周、6周和12周时测量abr和dpoaes,并且没有发现经注射的和未注射的c.216ga小鼠阈值之间的差别(来自6周龄小鼠的记录显示在图16c至图16d中)。该数据用作注射技术的对照,但重要的是,该载体不符合外源性截短harmonin不与内源性全长度harmonin竞争的说法,而该说法意味着c.216aa毛细胞中的内源性截短形式不可能干扰经由基因疗法载体表达的外源性全长度harmonin。
为了确定harmonin基因增补能否复原ush1c小鼠的听觉和平衡功能,在p0至p1实施aav2/anc80.cmv.harmonin-a1(0.8μl,1.7×10^12gc/ml)或aav2/anc80.cmv.harmonin-b1(0.8μl,1.9×10^12gc/ml)的rwm注射,并评估听脑干反应(abr)、畸变产物耳声发射(dpoae)、声学惊吓反射、开放场域和转棒行为。在6周时评估小鼠,6周是c.216aa小鼠苦于深刻听力损失和前庭功能障碍的时期。一些小鼠在3个月和6个月时进一步测试。12只注射aav2/anc80.cmv.harmonin-a1的小鼠无一在6周时恢复听觉功能(图9a至9c),表明harmonin-a1的外源性表达对于听觉复原是不够的。但是,25只注射aav2/anc80.cmv.harmonin-b1的小鼠中的19只的听觉功能在6周时显著恢复。在低频(5.6至16khz)下,注射aav2/anc80.cmv.harmonin-b1的耳朵的最佳abr阈值为25至30dbspl,与野生型小鼠的阈值惊人地相似(图16a至16b)。在22.6khz观察到部分复原,而在32khz观察到很低程度的复原或未观察到复原。dpoae阈值的复原也得到证实,与ohc中功能的复原相一致(图16c)。在较晚阶段,测试了8只对于8至11.3khz刺激的听觉阈值<45dbspl的小鼠,以评估该复原的持续时长。从6周到3个月,在低频范围内观察到了~10dbspl的abr阈值偏移,在高频范围内观察到了~30dbspl的abr阈值偏移(图16d)。在dpoae阈值中也观察到了类似的偏移(图16e)。在这一时间点之后,abr阈值和dpoae在6月龄之前保持稳定(图16d至16e),而6月龄是所测试的最晚时间点。
为了评估harmonin-a1和harmonin-b1两者是否为更完全的听觉复原尤其是高频端听觉复原所需,共同注射了aav2/anc80.cmv.tdtomato::harmonin-a1(0.5μl;2384.1e^12gc/ml)和aav2/anc80.cmv.egfp::harmonin-b1(0.5μl;3.0e^12gc/ml)。65%的毛细胞表达harmonin-a1和harmonin-b1两者,如从对于两种荧光标签为阳性的细胞中证实的(图14)。在暴露于aav2/anc80.cmv.tdtomato::harmonin-a1的小鼠的静纤毛中偶尔观察到荧光标记的harmonin-a1,可能是因为过度表达。共同注射未标记的harmonin-a1载体和harmonin-b1载体的小鼠的abr和dpoae阈值(图9)与单独注射harmonin-b1的小鼠类似,且并不提供进一步的改善,表明harmonin-a1对于听觉功能来说可能是可有可无的。重要的是,数据表明单独使用harmonin-b1就足以造成低频听觉阈值的显著复原(图9)。
为了进一步评估复原的程度,分析来自阈值≤45dbspl的小鼠的abr波形,并在8只c.216ga对照小鼠和5只注射aav2/anc80.cmv.harmonin-b1的c.216aa小鼠之间进行比较。对在8至11.3khz和16khz的反应的分析显露了正常的波1波幅(非显著性差异,p>0.2,学生t测试)和更长的峰值1延迟(p>0.001)(图15),表明在突触处可能存在神经传递滞后。在多数动物中,也在对侧耳中观察到了听觉复原,且abr阈值在11.3khz低至20dbspl(harmonin-b1:平均59.7±5.3dbspl,n=15/25;harmonin-a1+harmonin-b1:255平均76.2±10.3dbspl,n=4至6)。先前已经观察到了aav载体扩散至对侧耳(37),该扩散可能经由在新生小鼠体内与蛛网膜下腔保持连续的外淋巴管而出现。
我们也想知道,在较晚发育阶段进行注射是否可导致部分听觉复原。在p10至p12实施aav2/anc80.cmv.harmonin-b1(0.8μl)的rwm注射,并在6周评估听觉阈值。p10至p12的注射小鼠无一具有可检测的dpoae,且它们的abr阈值与未注射的c.216aa对照小鼠没有差别(n=10;数据未显示),表明干预机会窗口可能被限制在产后早期阶段,这可能是由于在较年长组织中的低病毒转导效率或者柯蒂氏器官在较晚发育阶段的退化。
实施例2f:乌谢尔小鼠模型中的rt-pcr
使用quantitect逆转录试剂盒(qiagen)从6只p2至p3的野生型、杂合和纯合ush1cc.216g>a小鼠的听觉器官制备cdna。使用下列引物扩增编码全长度harmonin(450bp)或截短harmonin(-35bp)的cdna:正向引物mush1c_ex2f:5’ctcattgaaaatgacgcagagaagg3’,反向引物mush1c_ex5r:5’tctcactttgatggacacggtctt3’。由于靶点序列位于ush1cc.216a基因座的人类敲入部分区域之外,这些引物对于小鼠ush1c序列是特异性的,并将扩增内源性ush1c和源自aav2的ush1c。也评估了在治疗后6周收集的小鼠组织的dna和rna水平。根据制造商的建议,使用trizol试剂(lifetechnologies,carlsbad,ca)从耳蜗中单离dna和rna。使用goscript逆转录系统来逆转录rna(promega,madison,wi)。使用gotaqgreen预混物(promega,madison,wi)进行放射标记的pcr。对于病毒dna扩增,使用对于小鼠ush1c:mush1c_ex3f(5’-gaacccaaccgcctgccg)和mush1c_ex4wtr(5’-tgcagacggtccaagcgt-3’)为特异性的引物。这些引物将仅扩增病毒ush1cdna,这是因为纯合ush1c.216aa小鼠具有敲入外显子3和4中并替代小鼠序列(32)的人类ush1cc.216a基因。对于全长度(450bp)和异常剪接/截短的harmonin(415bp)的cdna扩增,使用与上述相同的引物(mush1c_ex2f和mush1c_ex5r)。gapdh引物为:mgapdh_ex3f(5’-611gtgaggccggtgctgagtatg-3’)和mgapdh_ex4r(5’-gccaaagttgtcatggatgac-3’)。在6%非变性聚丙烯酰胺凝胶上单离产物,并使用typhoon9400磷光成像仪(gehealthcare)对其定性。
由于先前研究中浮现了截短harmonin可能通过与全长度harmonin竞争内源性结合伴侣而扰乱功能的可能性(34、35),我们想知道截短蛋白质的持续表达是否可能限制注射表达外源性全长度harmonin的载体的c.216aa小鼠的恢复(图16a)。为了解决这一顾虑,使用rt-pcr分析检查了c.216ga小鼠和c.216aa小鼠体内的ush1c转录物。与先前的报导一致,在c.216ga耳蜗中检测到了编码全长度harmonin和截短harmonin的ush1c转录物,而在c.216aa耳蜗中仅检测到了编码截短harmonin的转录物(图16b)。
为了证实aav2/anc80.cmv.harmonin-b1的表达并探究病毒表达水平与abr阈值之间的关系,从注射耳蜗和对侧耳蜗单离出dna和rna,并分别通过pcr和rt-pcr定量。在6周龄c.216ga小鼠以及注射和未注射aav2/anc80.cmv.harmonin-b1(0.8μl;1.9310^12gc/ml)的c.216aa小鼠体内评估表达。样本包括两只具有良好abr复原(在11.3khz,阈值≤35dbspl)的注射小鼠和两只具有极差abr复原(在11.3khz,阈值≥90dbspl)的小鼠。在所有被注射的耳蜗中,在较小程度上,在所有测试动物的对侧耳蜗中,均检测到了编码正确剪接形式的harmonin的rna(图18a)和aav2/anc80.cmv.harmonin-b1dna(图18b)。动物的abr阈值以及所表达的dna和rna的量之间存在可变性(图18c)。但是,在aav2/anc80.cmv.harmonin-b1dna水平、编码正确剪接形式的harmonin的rna的量和abr阈值水平之间发现强相关,这表示abr数据的可变性可能是aav表达的直接结果。为了评估具有成功恢复的abr阈值的小鼠体内的毛细胞长期存活,制备组织,并计数5只6月龄小鼠体内的ihc和ohc数目(图17)。尽管两个队列中的ihc数并未改变,但三只小鼠体内保留50%或更多的显示长期abr复原的ohc。观察除基底回之外整个器官的ohc存活(图17)。
实施例2g:听觉惊吓反应
使用惊吓监控器(kinderscientific)测量听觉惊吓反应(asr)。将小鼠放置在固定在压电/有机玻璃感应组件上的小尺寸的、非限制性的、立方有机玻璃记录室(27cm×10cm×65212.5cm)内,并令其在60dbspl背景白噪声下适应5分钟。每个环节由35次试验组成,在此期间,递送强度为60至120dbspl的10dbspl范围内的单个噪声脉冲,每次试验间隔平均为30秒(25至35秒范围)。脉冲以伪随机顺序布置,背景噪声为恒定的60dbspl,以限制外部噪声干扰。惊吓监控系统将对每一脉冲的反应还原为第一n、maxn和反应的最大时间(ms),以便计算峰值惊吓反应(asr幅度)以及从刺激到峰值惊吓反应的时间(asr延迟)。asr全部为盲测。
为了评估abr/dpoae恢复是否导致行为上相关的听觉功能恢复,测量了注射aav2/anc80.cmv.harmonin-a1的小鼠、注射aav2/anc80.cmv.harmonin-b1的小鼠和注射两种载体的小鼠的声学惊吓反应。对于白噪声惊吓反应的分析显示,注射aav2/anc80.cmv.harmonin-b1的6周龄小鼠和共同注射两种载体的小鼠的该反应部分复原(图10a)。仅接受harmonin-a1的小鼠与未注射的c.216aa小鼠类似,并未恢复惊吓反应。
实施例2h:前庭评估
使用开放场域测试和转棒平衡测试评估前庭功能。使用直径测量为42cm的圆形框进行开放场域测试,将其置于昏暗房间内的具有顶部led照明的声室内,led设定为在其中心为30lux。每次一只地将小鼠置于该圆形开发场域内,并令其暴露5分钟。记录行为,并使用ethovisionxt跟踪,令行动距离和速度的测量成为可能。开放场域平均全部为盲测。转棒表现牵涉小鼠在密闭笼内的棒上的安置,其中该棒以4rpm开始旋转并以0.1rpms-1的速率加速。第一天,将小鼠在棒上放置5分钟,以熟悉该设备。第二天,将小鼠放置在棒上,总计进行5次试验。每两次试验之间设置5分钟的休息时间。将动物在坠落至笼的仪器底板上之前在该装置上停留的时间长度显示在计时器上,并在每次测试运行结束后记录该时间长度。
由于耳蜗的外淋巴空间与前庭迷路的外淋巴空间是连续的,经由rwm注射的aav载体也可转导前庭感觉器官。为了评估前庭行为,测试小时在转棒上的表现。尽管在c.216aa小鼠和注射aav2/anc80.cmv.harmonin-a1的c.216aa小鼠中观察到了极差的转棒表现(坠落前平均停留时间<22sec),但注射aav2/anc80.cmv.harmonin-b1的c.216aa小鼠和那些同时注射harmonin-a1和harmonin-b1载体的小鼠在转棒上维持平衡功能60至120秒,与c.216ga对照小鼠一致(图10b)。
在注射harmonin-b1的c.216aa小鼠以及进行harmonin-a1和harmonin-b1双重注射的c.216aa小鼠中也观察到了开放场域行为的恢复。代表性的开放场域探索示踪绘制在图17c中。c.216ga小鼠探索了更广阔的场域且展示了最小程度的全身转动,而c.216aa小鼠在整个室内均展示了更大的活跃性且定性为旋转/分钟的全身转动增加(图10d至图10e)。令人惊奇的是,尽管在注射aav2/anc80.cmv.harmonin-a1的小鼠体内没有观察到abr复原,但开放场域数据表明了前庭功能恢复至对照小鼠的水平。注射aav2/anc80.cmv.trunc-harmonin的c.216ga小鼠的行为与c.216ga对照小鼠没有差别,再次表明截短的harmonin与野生型harmonin之间缺乏干扰(图10c至图10e)。
实施例3:听力损失所牵涉的附加突变的多核苷酸疗法
实施例3a:体内实验
随着转圈行为消失但注射harmonin-a1的小鼠未能成功通过转棒测试,行为实验表明使用harmonin-a1实现了部分的前庭复原。另一方面,注射harmonin-b1的小鼠在两个测试中均表现出功能恢复(图10)。横纹区域内不存在转导和fm1-43吸收,表明横纹区域的毛细胞和可能的i型细胞功能取决于适宜的harmonin表达(图6)。
尽管听觉的低频复原卓越而高频并非如此(图9),但在6周时在整个器官上均观察到了纤毛束形貌的保持(图11)。高频复原的缺失可能是由于注射造成的损害。在任何注射aav载体的c.216ga体内均未观察到高频听力损失(图16c至图16d)。作为一种解释,aav在整个耳蜗长度上的靶向性反对基底部缺乏转导效率的说法。一种可能性为,其它harmonin亚型诸如短harmonin-c可能是耳蜗的基底部高频端功能复原所必需的。或者,由于耳蜗的发育在基底端开始,来自基底高频端的毛细胞在p0时就已经成熟而超过修复点是可能的。如果是这种情况,则胚胎干预可能导致在高频区域的更好复原。
使用辅助无病毒系统和双重转染方法,如先前所述,生成携带由经修饰的cmv启动子驱动的小鼠tmc1编码序列的anc80载体(grimmetal.,2003,mol.ther.,7:839:50)。将三重旗标(tripleflag-tag)(flag)序列融合至tmc编码序列的c端,以令被表达的蛋白质能够可视化。使用不连续梯度的碘克沙醇以及其后进行的离子交换色谱来纯化anc80-cmv-tmc载体。使用对于人类β-球蛋白固有元件为特异性的引物进行定量pcr,测量的滴度范围是1×1012至1×1013gc/ml。将病毒等量小样在-80℃储存,并在融化后立即使用。
如下文所述,根据波士顿儿童医院动物护理和使用委员会机构皮疹的条款(条款#2659、#2146),使用p0至p2年龄的小鼠进行病毒载体的体内递送。将c57bl/6j(杰克逊实验室)或swisswebster小鼠种系(taconic)用作野生型对照小鼠,而携带tmc1突变基因座的小鼠(tmc1δ/δ或tmc1-/-)是如先前所述基于c57bl/6j而获得(kawashimaetal.,2011,j.clin.invest.,121:4796-809)。
为了制备用于评价的组织,从p0至p10的幼鼠采集颞骨。通过快速斩首杀死幼鼠,切除颞骨,将其置于以10mmhepes、0.05mg/ml氨苄青霉素和0.01mg/ml环丙沙星补充的ph7.40的mem(invitrogen)中。在解剖镜下单离膜迷路,剥落reissner氏膜,并用机械方法移除盖膜和血管纹。柯蒂氏器官培养物被扁平地固定在其一端使用sylgard粘附在18mm圆形盖玻片的一对薄玻璃纤维下。该组织很快被用于电生理学研究。对于年龄超过p10的小鼠,在吸入co2对动物实施安乐死后采集颞骨,并生成耳蜗的整装包埋。
图中的所有均值和误差棒均表示均值±sd。使用双尾成对t测试实施注射耳与未注射耳之间的统计学显著性的比较。将p<0.05视为显著。
实施例3b:病毒载体的体内注射
使用坡口玻璃显微注射穿刺针经由圆窗膜对幼鼠(p0至p2)注射。在p-2000穿刺针拉制仪(sutterinstruments)上从毛细玻璃管拉出穿刺针,并使用微穿刺针坡口机(sutterinstruments)切出坡口(~20μm尖端直径,28°角)。使用无菌拭子在外部施加用于镇痛的emla乳霜(2.5%利多卡因和2.5%丙胺卡因),以覆盖手术部位(左侧乳突状突出)。手术前,在37℃取暖垫上维持体温30至60分钟。
通过在2至3分钟内快速降低体温将幼鼠麻醉直至失去意识为止,并在手术过程中在冷却平台上将这一状态维持10至15分钟。使用碘伏(betadine)刷洗并使用70%乙醇擦拭,重复三次,将手术部位消毒。作出耳后切口以暴露透明的耳泡,通过显微操作器(mp-30,sutterinstrumentcompany)操纵微穿刺针前行通过耳泡和上覆筋膜,并借由微穿刺针的尖端穿透rwm。
使用气动显微注射器(wpinanoliter2010)将大约1μl的滴度为1012与1014gc/ml(109和1011总病毒颗粒)之间的病毒以0.1μl/min单侧注射入左耳。使用6-0单丝缝线(ethicon)闭合皮肤切口。随后将幼鼠送回取暖垫上进行恢复。
实施例3c:免疫荧光
实施免疫染色来确定通过病毒载体递送的转基因的表达分布。为了做到这一点,对新鲜切除并在室温使用稀释于pbs中的4%多聚甲醛浸没固定1h的柯蒂氏器官实施免疫染色。随后将该组织在pbs中冲洗,在0.01%至0.1%tritonx-100中透化处理30分钟,使用alexafluor546-鬼笔环肽(molecularprobes,1:200稀释)复染1h以标记丝状肌动蛋白。
对于外源性表达的tmc::flag融合蛋白的定位,使用2%bsa和5%正常山羊血清将该组织封闭1小时,使用flag基序抗体(bdbiosciences,1:200稀释)在4℃温育过夜。对于毛细胞计数,将组织在正常山羊血清中封闭1小时,使用兔抗-myosinviia一级抗体(proteusbiosciences,1:1000稀释)在4℃染色过夜,并使用偶联到alexafluor488的山羊抗兔抗体(lifetechnologies,1:200稀释)标记1h。使用vectashield封固剂(vectorlaboratories)将样品封固在盖玻片上,并使用zeisslsm700共聚焦显微镜以10x至63x放大成像。
图13显示证明harmonin至ush1c突变小鼠的均匀的anc80递送的免疫荧光,而图28显示证明kcnq4至kcnq4突变小鼠的anc80递送。
实施例3d:毛细胞电生理学
将器官型耳蜗培养物沐浴在含有137mmnacl、0.7mmnah2po4、5.8mmkcl、1.3mmcacl2、0.9mmmgcl2、10mmhepes和5.6mmd-葡萄糖的标准人工外淋巴液中。将维生素(1:50)和氨基酸(1:100)加入来自浓缩物(invitrogen)的溶液中,并使用naoh将最终ph调节为7.40(310mosmol/kg)。从r6毛细玻璃管(kingprecisionglass)拉出记录穿刺针(3至5megohm)并充满含有135mmcscl、5mmhepes、5mmegta、2.5mmmgcl2、2.5mm三磷酸腺苷二钠、和0.1mmcacl2的胞内液,其中,csoh用来将最终ph调节为7.40(285mosmol/kg)。在室温(22℃至24℃)下,使用axopatch200b放大器(moleculardevices)在-84mv进行全细胞密封电压钳记录。使用低通量bessel滤波器在10khz将感觉转导电流滤波,使用16位采集板(digidata1440a)和pclamp10软件(moleculardevices)在≥20khz数字化。存储数据,使用originpro8(originlab)进行线下分析。
图23显示,以anc80-kcnq4转染的kcnq4-/-小鼠细胞中钾电流相对于突变小鼠(图23b)的接近野生型水平(图23a)的恢复。
实施例3e:听觉脑干反应(abr)
如先前所述,进行abr记录(maisonetal.,2010,j.neurosci.,30:6751-62)。简单地说,经由ip注射(0.1ml/10g体重)稀释在5ml的0.9%盐水中的50mg氯胺酮和5mg甲苯噻嗪麻醉p25至p30的小鼠。在隔音室内在32℃实施abr实验。为了测试听力功能,将小鼠呈递至以5db阶跃的10db与115db之间的声压级的5.6khz、8khz、11.3khz、16khz、22.6khz、或32khz的纯音刺激,直到检测到诱发可再现的abr波形的abr阈值强度(峰i至iv)为止。使用交替极性刺激,对每个声压级收集512至1024个反应并取平均。通过“去伪”功能丢弃波幅(峰至谷)大于15μv的波形。在着手进行abr测试之前,使用解剖剪切掉典型遮盖外耳道入口的皮瓣和软骨,验证每一个体受试者在全部刺激频率下的耳道入口处的声压。通过由两个静电耳机(cuiminiaturedynamics)组成的用以生成主音的自定义探针管扬声器/扩音器组件(eplpxisystems)和用以记录耳道声压的knowles微型麦克风,将声学刺激直接递送至所研究的耳部。声音刺激由5毫秒猝发音(0.5ms升降,cos2始发,以40/s递送)组成。使用插入耳廓(活性电极)、头顶(参考电极)和尾部(接地电极)的皮下针电极收集abr信号。使用自定义数据采集软件(labview)将abr电位放大(10,000x)、滤波(0.3至10khz)并数字化。使用数字化i-o板(nationalinstruments)以40μs的间隔对声音刺激和电极电压采样并存储以备线下分析。阈值形象化地定义为最低分贝水平,在该水平下,任何波(i-iv)均可被检测并使用增加的声强再次产生。对每一实验组内的abr阈值取平均,并用于统计学分析。
图20图解性地阐明,递送编码并表达harmonin的anc80病毒载体可提供对于听觉功能的近乎完全的恢复,尤其是在较低频率(例如,约5至约22khz)的功能。
实施例3f:定量rt-pcr分析
进行实验以评估在体内给药后耳蜗中存在的病毒的量。在p1对两只tmc1-/-小鼠的左耳进行注射。将耳蜗从左耳和右耳切除,并在培养物中保持3天,等效于p10。提取rna,使用安捷伦生物分析仪(agilentbioanalyzer(agilenttechnologies))证实品质,再逆转录至cdna中,使用sybrgreenerqpcr试剂(invitrogen)以对tmc1为特异性的有效引物组进行定量rt-pcr分析,如先前所述(kawashimaetal.,2011,j.clin.invest.,121:4796-809)。
使用下列引物扩增tmc1的片段:5’-catctgcagccaactttggtgtgt-3’(seqidno:9)和5’-agaggtagccggaaattcagccat-3’(seqidno:10)。将表达水平归一化至使用5’-tgagcgcaagtactctgtgtggat-3’(seqidno:11)和5’-actcatcgtactcctgcttgctga-3’(seqidno:12)扩增的actb(编码β-肌动蛋白)的表达水平。所有引物均设计为跨内含子,并使用解链曲线分析和阴性对照验证。使用δδct方法相对于actb和注射区域与未注射区域间的差异分析数据。tmc1mrna在注射区域内的表达比未注射区域内高12倍。
实施例3g:fm1-43标记
如先前所述,实施fm1-43染料负载实验(galeetal.,2001,j.neurosci.,21:7013-25;meyersetal.,2003,j.neurosci.,23:4054-65;以及géléoc&holt,2003,nat.neurosci.,10:1019-20)。将粘附有耳蜗培养物的盖玻片放置在直立式显微镜(zeissaxioscopefsplus)下的玻璃底腔室上。将在人工外淋巴液中稀释的5μmfm1-43fx(invitrogen)施加10秒钟,将组织在人工外淋巴液中洗涤三次以从细胞膜的外叶移除染料。5分钟后,使用fm1-43滤波器组合具有63x水浸物镜的落射荧光光源对细胞内fm1-43成像。固定并处理组织,以进行如上所述的免疫荧光分析。
图23是免疫染色图像,显示了暴露于本文所述anc80病毒载体的细胞对fm1-43染料的吸收;而图28图解性地阐明,通过本文所述anc80病毒载体递送的tmc1在体内复原了tmc1缺陷性毛细胞的感觉转导。
实施例3h:畸变产物耳声发射(dpoae)
在于abr数据相对的条件下并在相同的记录环节内,收集dpoae数据。产生频率比为1.2(f2/f1)的用于在2f1-f2下生成dpoae的主音,其中,对于每个f2/f1对,f2水平比f1水平低10db声压级。f2水平以5db的阶跃从20db至80db扫频。在每一水平使用波形和频谱平均来增加所记录的耳道声压的信噪比。从平均频谱中提取2f1-f2下的dpoae幅度,以及频谱中附近点的本底噪声。从dpoae幅度与声级的对比图对iso反应曲线插值。阈值定义为在0db产生dpoae所需的f2水平。
图25图解性地阐明,使用本文所述的anc80病毒载体递送的tmc1复原了tmc1-/-小鼠的外耳毛细胞功能,尤其是在较低频率(例如,约5至16khz)下的功能。
图26表明启动子pcdh15、myo6和kcnq4对于毛细胞中gfp表达的效果。
其它具体实施例
应理解,尽管本位中已经联合大量不同方面描述了方法和物质的组合物,但前述对多个方面的说明旨在示出而非限制该方法和物质的组合物的范畴。其它方面、优点和修饰处于后附权利要求书的范畴内。
本文中,对任何变量定义中一系列元件的描述包括该变量作为任何单一元件或作为所列元件的组合(或亚组合)的定义。对本文中具体实施例的描述包括该具体实施例作为任何单一具体实施例或作为与任何其它具体实施例或其部分的组合。
本说明书中提及的全部专利和出版物通过引用并入本文,其并入程度与各独立的专利和出版物具体且独立地指明以待通过引用并入的程度相同。
公开了可用于、可用于制备所公开的方法和组合物或可与之合用或作为其产物的方法和组合物。本文中公开了这些和其它材料,应理解,也公开了这些方法和组合物的组合、子集、相互作用、组等。换句话说,尽管这些组合物和方法的每一个体以及集合性的组合和排列可不被明确地公开,但其各自在本文中被特异性地关注和描述。例如,如果特定的物质组合物或特定的方法被公开并谈论且大量组合物或方法被讨论,则除非另有特别指示,否则该组合物和方法的每一种组合和排列均被特异性地关注。同样,这些的任何子集或组合也被特别性地关注和公开。
- 还没有人留言评论。精彩留言会获得点赞!