用于预防、减轻和/或治疗痴呆的系统和方法与流程
相关申请的交叉引用
本申请要求以下每个美国临时申请的优先权权益:于2017年10月10日提交且名称为“neuroprotectiveeffectsofcombinedsensorystimulation”的序列号62/570,250(代理人案卷号mitx-9699/00us);以及于2017年10月11日提交且名称为“gammaentrainmentbindshigherorderbrainregionsandoffersneuroprotection”的序列号为62/570,929(代理人案卷号mitx-0070/00us)。这些临时申请中的每一个以引用方式全文并入本文。
政府支持声明
本发明是在由美国国立卫生研究院授予的资助号rf1ag047661下在美国政府的支持下进行的。美国政府享有本发明的某些权利。
背景技术:
阿尔茨海默氏病(ad)是一种以记忆力、定向以及推理能力的下降为特征的进行性神经退化性疾病。它是世界上最常见的痴呆形式,大约影响八分之一的65岁以上的人,并且是美国第六大主要死因。在接下来的十年里,该进行性神经退化性疾病的患病率估计将增加40%。
在组织病理学上,ad可以以淀粉样斑的积累为特征,所述淀粉样斑包含淀粉样-β(aβ)肽和由tau蛋白制成的神经元纤维缠结(nft)。aβ肽是一种36-43个氨基酸的蛋白质,所述蛋白质的正常生理功能尚未确定。aβ肽由β-分泌酶1(bace1)和γ-分泌酶对淀粉样前体蛋白(app)进行顺序蛋白水解切割而形成。c端片段β(β-ctf)是在由bace1进行的app的促淀粉样(amyloidogenic)切割期间产生的app衍生物并由此是aβ肽产生的另一个指示。在正常情况下,可溶性aβ肽由神经元产生和分泌,随后经由脑脊髓液(csf)通路从大脑清除。然而,在患有ad的受试者中,aβ肽似乎以浓度相关方式聚集成更高级种属,以形成可溶性低聚物和不溶性斑块。该聚集可能引发许多神经毒性事件,包括脑代谢紊乱、神经炎症、功能连接降低、突触和神经元丢失和/或nft形成。
已经证明了aβ浓度与神经元活性之间的基本关系。首先,用河豚毒素处理由过表达app的转基因(tg)小鼠制备的海马体器官型脑片降低神经元活性,随后降低aβ水平。然后,在用印防己毒素处理时观察到相反的作用-神经元活性增加。也已经使用神经元活性证明了aβ肽浓度的动态调节和体内的最终斑块沉积。在人类ad患者中,神经影像学显示最严重的斑块沉积可能与被称为“默认-模式网络”的始终最活跃的脑区对齐。
目前ad尚不能治愈,并且治疗选项不抑制ad的病理进展,主要是姑息性的,和/或可能具有多种令人不安的副作用。例如,在临床试验中,针对aβ肽和/或其前体的预防和/或治疗策略(例如,aβ免疫疗法和对β-和γ-分泌酶的抑制)在降低ad病变时是有毒和/或无效的。由于缺乏认知益处,涉及淀粉样β疫苗(例如,巴品珠单抗)的临床试验失败。γ-分泌酶抑制剂(例如,司马西特)因加重受试者的认知缺陷而在临床试验中失败。即使是现有的药物,像乙酰胆碱酯酶抑制剂(例如,多奈哌齐和利凡斯的明)和n-甲基-d-天冬氨酸(nmda)-受体拮抗剂(例如,美金刚)也仅表现出轻微的认知益处。
技术实现要素:
如在2016年11月23日提交且名称为“systemsandmethodsforpreventing,mitigating,and/ortreatingdementia”的美国专利申请序列号15/360,637中公开的(在此以引用方式全文并入本文),经由视觉或听觉刺激在大脑中引起同步的γ振荡导致一些脑区中的淀粉样负荷降低和形态变化。然而,发明人已经认识到并理解,仍然需要治疗痴呆和阿尔茨海默氏病的系统和方法,所述系统和方法解决影响多个大脑中枢的全回路疾病,所述大脑中枢对学习和记忆以及其他高级脑功能负有重要责任。
根据前述内容,本公开至少部分涉及组合的听觉和视觉刺激,所述刺激根据在本文中通常称为“使用感官刺激的γ夹带(genus,gammaentrainmentusingsensorystimuli)”的各种技术,来在受试者的大脑中引起γ振荡。如本文所公开的组合的听觉和视觉刺激(例如,组合的视觉和听觉genus)出乎意料地生成了对于单独视觉或听觉genus未观察到的积极的生理和行为变化。由组合的听觉和视觉genus产生的对大脑的积极效果不限于听觉皮层(ac)和海马体(hpc),而是显著地,它们扩展到在内侧前额叶皮层(mpfc)中引起小神经胶质簇聚反应并减少整个新皮层中的淀粉样负荷。此外,组合的听觉和视觉genus的效果在较短的治疗/暴露时间范围(大约数周)内观察到。
在一方面,本公开提供了用于治疗有需要的受试者中的痴呆或阿尔茨海默氏病的装置、方法以及系统,方法包括以下步骤:向受试者非侵入性地递送具有约20hz至约60hz的频率的组合的听觉和视觉刺激,以在受试者的至少一个脑区中引起同步的γ振荡。在一些实施方案中,痴呆与ad、血管性痴呆、额颞痴呆、路易体痴呆和/或年龄相关认知衰退关联。受试者可以是人或动物。
在一些实施方案中,组合的听觉和视觉刺激具有约35hz至约45hz的频率或约40hz的频率。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激在选自听觉皮层(ac)、视觉皮层(vc)、海马体(hpc)和内侧前额叶皮层(mpfc)的至少一个皮层区域中的5%或更多的记录位点中引起周期性的尖峰反应。在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激在mpfc中以约40hz引起局部场电位(lfp)。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激提高了至少一个皮层区域中的小神经胶质反应。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在一些实施方案中,提高小神经胶质反应包含选自增加25微米的淀粉样斑内的小神经胶质数量;增大小神经胶质细胞体直径;减小小神经胶质投影长度;以及增加小神经胶质细胞计数的至少一个效果。在实施方案中,提高小神经胶质反应包含将小神经胶质细胞体直径增大至少10%、20%、30%、40%或50%。在实施方案中,提高小神经胶质反应包含将小神经胶质投影长度减小至少10%、20%、30%、40%或50%。在实施方案中,提高小神经胶质反应包含将小神经胶质细胞计数增加至少10%、20%、30%、40%或50%。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激包含减少选自新皮层、ac、vc、hpc和mpfc的至少一个皮层区域中的淀粉样斑。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在实施方案中,减少淀粉样斑包含将斑块尺寸减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。在实施方案中,减少淀粉样斑包含将斑块数量减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激包含减少选自新皮层、ac、vc、hpc和mpfc的至少一个皮层区域中的淀粉样-β(aβ)肽的数量。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在实施方案中,减少aβ肽的数量包含将数量减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。在一些实施方案中,aβ肽包括同种型aβ140肽和同种型aβ1-42肽中的至少一种。在一些实施方案中,aβ肽包括可溶性aβ肽和不溶性aβ肽中的至少一种。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。
在第二方面,本公开提供了一种用于治疗有需要的受试者中的痴呆或阿尔茨海默氏病的方法,所述方法包括以下步骤:控制至少一个视觉刺激器发射频率为约35hz至约45hz的视觉刺激;控制至少一个电声换能器将电音频信号转换为频率为约35hz至约45hz的对应听觉刺激;以及向受试者非侵入性地递送组合刺激,所述组合刺激包括同步对齐的视觉刺激和声音刺激,组合刺激在受试者的至少一个脑区中引起同步的γ振荡。同步的γ振荡导致受试者认知功能的改善。
在一些实施方案中,视觉刺激包含重复地将光打开12.5ms然后关闭12.5ms。在一些实施方案中,光遗传刺激器是具有40-80w功率的发光二极管。在一些实施方案中,听觉刺激包含以约4%至约80%的占空比以40hz播放的10khz音调。在一些实施方案中,视觉刺激包含以约10%至约80%的占空比以40hz闪烁10秒时段的光。
在一些实施方案中,视觉刺激和听觉刺激是同步的。在一些实施方案中,视觉刺激和听觉刺激的相差为从-180度到0度或从0度到180度。
应当理解,前述概念和下面更详细讨论的附加概念的所有组合(假设此类概念并不相互矛盾)被认为是本文所公开的发明主题的一部分。特别地,出现在本公开的结尾处的要求保护的主题的所有组合被认为是本文所公开的发明主题的一部分。还应当理解,本文明确采用的也可能出现在通过引用并入的任何公开中的术语应被赋予与本文所公开的特定概念最一致的含义。
通过查看以下附图和详细描述,其他系统、过程和特征对于所属领域的技术人员将变得显而易见。预期的是将所有此类附加系统、过程和特征包括在本说明书内,在本发明的范围内,并受所附权利要求书保护。
附图说明
本专利或申请文件包含至少一张彩色附图。
本领域技术人员将理解,附图主要是出于说明性目的,并且无意限制本文所述的发明主题的范围。附图不一定按比例绘制;在一些情况下,本文所公开的发明主题的各方面可以在附图中被扩大或放大示出,以促进对不同特征的理解。在附图中,相同的参考特性通常参见例如相同的特征(例如,功能上相似和/或结构上相似的元件)。
图1a至图1l示出40hz听觉刺激调节ac、ca1和mpfc中的尖峰活性。
图2a至图2j示出听觉genus改善5xfad小鼠的识别和空间记忆任务。
图3a至图3i示出听觉genus降低5xfad小鼠的ac和hpc中的淀粉样负荷。
图4a至图4k示出听觉genus在5xfad小鼠的ac和ca1中引起神经胶质反应。
图5a至图5f示出听觉genus增加淀粉样-脉管系统的关联。
图6a至图6i示出组合的听觉和视觉genus通过小神经胶质引起簇聚显型反应。
图7a至图7j示出组合的听觉和视觉genus减少mpfc和新皮层中的淀粉样负荷。
图8a至图8r示出20hz和80hz的听觉刺激调节ac、ca1和mpfc中的活性。
图9a至图9l示出听觉genus不影响小鼠行为。
图10a至图10h示出听觉genus改善app/ps1小鼠中的斑块负荷。
图11a至图11h示出听觉genus在app/ps1小鼠中引起小神经胶质反应。
图12a至图12l示出听觉genus减少p301s小鼠中的磷酸化tau。
图13a至图13u示出40hz的组合的听觉和视觉刺激调节ac、ca1和mpfc中的尖峰活性。
图14a至图14v示出1周的听觉或视觉genus仅不影响mpfc病变。
具体实施方式
使用感官刺激的组合的听觉和视觉γ夹带(genus)出乎意料地生成了对于单独视觉或听觉genus未观察到的积极的生理和行为变化。对大脑的积极效果不限于听觉皮层(ac)和海马体(hpc),而是扩展到在内侧前额叶皮层(mpfc)中引起小神经胶质簇聚反应并减少整个新皮层中的淀粉样负荷。此外,组合的听觉和视觉genus的效果在较短的时间范围(大约数周)内观察到。大约一周的组合的听觉和视觉genus改善跨越大型回路网络的脑区中的阿尔茨海默氏病(ad)病变。特别地,组合的听觉和视觉genus显著降低ac、视觉皮层(vc)、海马体亚区ca1和mpfc中的淀粉样负荷。组合的听觉和视觉genus在内侧前额叶皮层中产生小神经胶质簇聚反应和减少的淀粉样。组合的听觉和视觉genus广泛减少整个新皮层的淀粉样斑。
在一方面,本公开提供了用于治疗有需要的受试者中的痴呆或阿尔茨海默氏病的装置、方法以及系统,方法包括以下步骤:向受试者非侵入性地递送具有约20hz至约60hz的频率的组合的听觉和视觉刺激,以在受试者的至少一个脑区中引起同步的γ振荡。在一些实施方案中,痴呆与ad、血管性痴呆、额颞痴呆、路易体痴呆和/或年龄相关认知衰退关联。受试者可以是人或动物。
在一些实施方案中,组合的听觉和视觉刺激具有约35hz至约45hz的频率或约40hz的频率。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激在选自听觉皮层(ac)、视觉皮层(vc)、海马体(hpc)和内侧前额叶皮层(mpfc)的至少一个皮层区域中的5%或更多的记录位点中引起周期性的尖峰反应。在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激在mpfc中以约40hz引起局部场电位(lfp)。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激提高了至少一个皮层区域中的小神经胶质反应。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在一些实施方案中,提高小神经胶质反应包含选自增加25微米的淀粉样斑内的小神经胶质数量;增大小神经胶质细胞体直径;减小小神经胶质投影长度;以及增加小神经胶质细胞计数的至少一个效果。在实施方案中,提高小神经胶质反应包含将小神经胶质细胞体直径增大至少10%、20%、30%、40%或50%。在实施方案中,提高小神经胶质反应包含将小神经胶质投影长度减小至少10%、20%、30%、40%或50%。在实施方案中,提高小神经胶质反应包含将小神经胶质细胞计数增加至少10%、20%、30%、40%或50%。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10周后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10个月后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10年后产生。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激包含减少选自新皮层、ac、vc、hpc和mpfc的至少一个皮层区域中的淀粉样斑。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在实施方案中,减少淀粉样斑包含将斑块尺寸减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。在实施方案中,减少淀粉样斑包含将斑块数量减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10周后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10个月后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10年后产生。
在一些实施方案中,非侵入性地递送组合的听觉和视觉刺激包含减少选自新皮层、ac、vc、hpc和mpfc的至少一个皮层区域中的淀粉样-β(aβ)肽的数量。皮层区域可以包括新皮层、ac、vc、hpc和mpfc。在一些实施方案中,在mpfc中引起小神经胶质反应。在一些实施方案中,在多个皮层区域中引起小神经胶质反应。在一些实施方案中,跨整个新皮层引起小神经胶质反应。
在实施方案中,减少aβ肽的数量包含将数量减少至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%或60%。在一些实施方案中,aβ肽包括同种型aβ140肽和同种型aβ1-42肽中的至少一种。在一些实施方案中,aβ肽包括可溶性aβ肽和不溶性aβ肽中的至少一种。
在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10天后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10周后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10个月后产生。在一些实施方案中,提高小神经胶质反应在非侵入性地递送组合的听觉和视觉刺激的1、2、3、4、5、6、7、8、9或10年后产生。
在第二方面,本公开提供了一种用于治疗有需要的受试者中的痴呆或阿尔茨海默氏病的方法,所述方法包括以下步骤:控制至少一个视觉刺激器发射频率为约35hz至约45hz的视觉刺激;控制至少一个电声换能器将电音频信号转换为频率为约35hz至约45hz的对应听觉刺激;以及向受试者非侵入性地递送组合刺激,所述组合刺激包括同步对齐的视觉刺激和声音刺激,组合刺激在受试者的至少一个脑区中引起同步的γ振荡。同步的γ振荡导致受试者认知功能的改善。
在一些实施方案中,视觉刺激包含重复地将光打开12.5ms然后关闭12.5ms。在一些实施方案中,光遗传刺激器是具有40-80w功率的发光二极管。在一些实施方案中,听觉刺激包含以约4%至约80%的占空比以40hz播放的10khz音调。在一些实施方案中,视觉刺激包含以约10%至约80%的占空比以40hz闪烁10秒时段的光。
在一些实施方案中,视觉刺激和听觉刺激是同步的。在一些实施方案中,视觉刺激和听觉刺激的相差为从-180度到0度或从0度到180度。如本文所用的,“相位”是指以度数表达的听觉刺激与视觉刺激之间的滞后,其中0度意指同时的听觉和视觉刺激,而-180或+180是指交替的视觉和听觉刺激。
如本文所用的,术语“治疗”是指治疗处理和预防性或预防措施这两者。在一些实施方案中,需要治疗的受试者包括已经患有疾病或病症的那些受试者以及可能发展疾病或病症的那些受试者,并且在后者的受试者中,目的是预防、延迟或减少疾病或病症。例如,在一些实施方案中,本文所公开的装置、方法和系统可以被采用来预防、延迟或减少受试者从遗传学角度易感的疾病或病症,诸如ad。在一些实施方案中,本文所公开的装置、方法和系统可以被采用来治疗、减轻、减少受试者已经被诊断患有的疾病或病症(诸如ad)的症状和/或延迟其进展。
如本文所用的,术语“受试者”代表哺乳动物,诸如啮齿类、猫科、犬科或灵长类。优选地,根据本发明的受试者是人。
如本文所用的术语“约”是指“约”修饰的对象的正负百分之十。
痴呆是以智力能力丧失和/或记忆损害为特征的疾病。痴呆包括例如ad、血管性痴呆、路易体痴呆、皮克氏病、额颞痴呆(ftd)、aids痴呆、与年龄有关的认知损害和与年龄有关的记忆损害。痴呆也可能与神经和/或精神疾病关联,诸如例如,脑瘤、脑损伤、癫痫、多发性硬化症、唐氏综合征、雷特综合征、进行性核上性麻痹、额叶综合症、精神分裂症和外伤性脑损伤。
ad是发达国家中最常见的神经退化性疾病。ad在组织病理学上以淀粉样斑的积累为特征,所述淀粉样斑由aβ肽和由tau蛋白制成的nft组成。临床上,ad与进行性认知损害关联,该认知损害以记忆、功能、语言能力、判断力和执行功能的丧失为特征。ad在其后期常常导致严重的行为症状。
血管性痴呆也可以称为脑血管性痴呆,是指脑血管疾病(例如,脑半球梗塞),这些疾病通常具有波动过程,该过程具有改善和逐步恶化的时段。血管性痴呆可包括定向障碍、记忆损害和/或判断力损害的一个或多个症状。血管性痴呆可能由离散的多发性梗塞或其他血管原因引起,这些原因包括例如自身免疫性血管炎,诸如在系统性红斑狼疮中发现的;感染性血管炎,诸如莱姆病;复发性脑出血;以及/或中风。
额颞痴呆(ftd)是一种进行性神经退化性疾病。患有ftd的受试者通常表现出明显的行为和性格改变,通常伴有语言损害。
路易体痴呆以以下内容的一个或多个症状为特征:特征与ad的特征重叠的痴呆发展;帕金森氏病特征的发展;以及/或幻觉的早期发展。路易体痴呆通常以症状严重程度的每日波动为特征。
在一些方面,本公开提供了用于预防、减轻和/或治疗受试者中的痴呆的方法,所述方法包括在受试者的大脑中引起同步的γ振荡。在一些实施方案中,在患有神经疾病或障碍或与年龄有关的下降的受试者中引起γ振荡用于恢复由于疾病或障碍或与年龄有关的下降或与其关联而在受试者中破坏的γ振荡节律。
在一些实施方案中,γ振荡的引起减少了同种型aβ1-40和aβ1-42的生成。在一些实施方案中,γ振荡的引起增强了从受试者的大脑清除aβ(例如,同种型aβ1-40和aβ1-42)。在一些实施方案中,γ振荡的引起防止了aβ在受试者的大脑中积累。在一些实施方案中,相对于治疗前的受试者的大脑中的aβ水平,本文所提供的方法将受试者大脑中的aβ水平降低约10%、约20%、约30%、约40%、约50%、约60%、约70%或更多。在一些实施方案中,相对于治疗前的受试者的大脑中的aβ水平,受试者的大脑中的aβ水平降低了至少约50%。
在一些实施方案中,通过减少受试者的大脑中的app的切割来降低受试者的大脑中的aβ水平。在一些实施方案中,相对于治疗前的受试者的大脑中的app切割水平,本文所提供的方法将受试者大脑中的app切割降低约10%、约20%、约30%、约40%、约50%、约60%、约70%或更多。在一些实施方案中,相对于治疗前的受试者的大脑中的app切割水平,受试者的大脑中的app切割水平降低了至少约50%。在一些实施方案中,app切割的水平通过受试者大脑中的c端片段β(β-ctf)的水平来测量。在一些实施方案中,大脑中app切割的水平通过抑制β-和/或γ-分泌酶,诸如通过增加对β-和/或γ-分泌酶活性的抑制水平来降低。在一些实施方案中,本文所提供的方法减少了受试者大脑中aβ斑块的聚集。
在一些实施方案中,方法改善受试者的认知能力和/或记忆。
在另一方面,本公开提供了用于在受试者的大脑中引起神经保护姿态或神经保护环境的方法,所述方法包括在受试者的大脑中引起同步的γ振荡。例如,在一些实施方案中,神经保护姿态与神经保护小神经胶质细胞轮廓关联。在另外的实施方案中,神经保护性姿态由m-csf通路的活性增加引起或与其关联。在一些实施方案中,神经保护环境与抗炎信号传导通路关联。例如,在一些实施方案中,抗炎信号传导通路是抗炎小神经胶质信号传导通路。
在一些实施方案中,神经保护姿态与促炎神经胶质细胞活性的降低或缺乏关联。促炎神经胶质细胞活性与小神经胶质中的m1显型关联,并且包括产生活性氧(ros)、神经分泌蛋白嗜铬粒蛋白a、分泌性辅因子胱抑素c、nadph氧化酶、一氧化氮合酶(诸如inos)、nf-κb依赖性炎症反应蛋白以及促炎细胞因子和趋化因子(例如tnf、il-1β、il-6和ifnγ)。
相反,小神经胶质的m2显型与炎症的下调和炎症引起的损伤的修复关联。抗炎细胞因子和趋化因子(il-4、il-13、il-10和/或tgfβ)以及吞噬活性的增加与m2显型关联。由此,在一些实施方案中,本文所提供的方法在小神经胶质中引起神经保护性m2显型。在一些实施方案中,本文所提供的方法增加了受试者大脑中的吞噬活性。例如,在一些实施方案中,本文所提供的方法增加了小神经胶质的吞噬活性,使得增加了aβ的清除率。
γ振荡可以包括大约20hz至大约100hz。由此,在一些实施方案中,本公开提供了用于预防、减轻或治疗受试者中的痴呆的方法,所述方法包括在受试者的大脑中引起约20hz至约100hz、或约20hz至约80hz、或约20hz至约50hz、或约30hz至约60hz、或约35hz至约45hz、或约40hz的γ振荡。优选地,γ振荡约为40hz。
刺激可包括受试者的内部或外部环境的任何可检测到的变化,该变化直接或最终在至少一个脑区中引起γ振荡。例如,刺激可以被设计为刺激电磁辐射感受器(例如,光感受器、红外线感受器和/或紫外线感受器)、机械感受器(例如,机械应力和/或应变)、伤害感受器(即,疼痛)、声音感受器、电感受器(例如,电场)、磁感受器(例如,磁场)、水感受器、化学感受器、热感受器、渗透压感受器和/或本体感受器(即,位置觉)。引起来自受体的反应所需的绝对阈值或最小感觉量可基于刺激类型和受试者而变化。在一些实施方案中,基于个体敏感性来调整刺激。
在一些实施方案中,以脑区特异性方式引起γ振荡。例如,在一些实施方案中,在海马体、视觉皮层、桶状皮层、听觉皮层或其任何组合中引起γ振荡。举例来说,在一些实施方案中,使用闪光在视觉皮层中引起γ振荡;而在其他实施方案中,使用特定频率的听觉刺激在听觉皮层中引起γ振荡。在一些实施方案中,使用视觉、听觉和/或其他刺激的组合同时在多个脑区中引起γ振荡。在一些实施方案中,在虚拟现实系统中引起γ振荡。
在一些实施方案中,受试者经由被配置成引起γ振荡的环境(诸如被动或主动地阻挡无关的刺激(例如,光阻挡或噪声消除)的室)接收刺激。另选地或另外,受试者可以经由包括例如光阻挡或噪声消除方面的系统来接收刺激。在一些实施方案中,受试者经由刺激发射装置(诸如被设计为递送刺激的眼镜)接收视觉刺激。所述装置可以挡住其他光线。在一些实施方案中,受试者经由刺激发射装置(诸如被设计为递送刺激的耳机)接收听觉刺激。所述装置可以消除其他噪声。
除了用于发射刺激的至少一个接口之外,一些实施方案还可以包括至少一个处理器(例如,以生成刺激,控制刺激的发射,监控刺激/结果的发射,和/或处理关于刺激/结果的反馈)、至少一个存储器(存储例如处理器可执行指令、至少一个刺激、刺激生成策略、反馈和/或结果)、至少一个通信接口(与例如受试者、保健提供者、看护人、临床研究调查员、数据库、监控应用程序等通信)和/或检测装置(检测和提供有关例如刺激和/或受试者的反馈,包括是否引起了γ振荡、受试者的敏感性、认知功能、物理或化学变化、应力、安全性等)。
在一些实施方案中,γ振荡是由视觉刺激(诸如约20hz至约100hz的闪光)引起的。在特定实施方案中,γ振荡是由约20hz至约50hz的闪光引起的。在另外的实施方案中,γ振荡是由约35hz至约45hz的闪光引起的。在另一些实施方案中,γ振荡是由约40hz的闪光引起的。在一些实施方案中,受试者接收(例如,被置于具有挡光装置的室中或佩戴挡光装置,该装置发射)约20hz至约100hz的闪光、或约20hz至约50hz的闪光或约35hz至约45hz的闪光、或约40hz的闪光。
在一些实施方案中,γ振荡由诸如声音的听觉刺激引起,该声音的频率为约20hz至约100hz、或约20hz至约80hz、或约20hz至约50hz、或约35hz至约45hz、或约40hz。在一些实施方案中,受试者接收(例如,被置于具有噪声消除装置的室中或佩戴噪声消除装置,该装置发射)约20hz至约100hz、约20hz至约80hz、约20hz至约50hz、约35hz至约45hz或约40hz的听觉刺激。
在一些实施方案中,受试者接收(例如,被置于具有挡光装置的室中或佩戴挡光装置,该装置发射)视觉刺激和/或听觉刺激约一小时、约2小时、约3小时、约4小时、约5小时或更长时间。在一些实施方案中,受试者接收(例如,被置于具有挡光装置的室中或佩戴挡光装置,该装置发射)刺激不超过约6小时、不超过约5小时、不超过约4小时、不超过约3小时、不超过约2小时或不超过约一小时。在一些实施方案中,受试者接收(例如,被置于具有挡光装置的室中或佩戴挡光装置,该装置发射)刺激少于一个小时。
在一些实施方案中,受试者经历本文所提供的方法。在其他实施方案中,受试者在多个单独的场合用本文所提供的方法进行治疗。受试者可以定期或者随着症状出现或恶化进行治疗。在一些实施方案中,慢性治疗可有效减少可溶性aβ肽和/或不溶性aβ肽(即,斑块)。
在一些实施方案中,以细胞类型特异性的方式引起γ振荡。在一些实施方案中,在fs-pv-中间神经元中引起γ振荡。当用于描述一类神经元时,术语“快速放电”(fs)是指神经元在几乎没有峰值频率适应或峰值高度的衰减的情况下长时间以高速率放电的能力。由此,这些神经元能够持续的高频(例如,等于或大于约100hz或约150hz)放电而没有明显的适应。fs神经元的该特性在很大程度上可归因于其快速延迟的整流通道(换句话讲,非常快速地激活和失活的通道)的表达。
在一方面,刺激可以是非侵入性的。如本文所用的术语“非侵入性”是指不需要外科手术干预或对身体的操纵(诸如组合物或装置的注射或植入)的装置、方法和系统。例如,刺激可以是视觉的(例如,闪烁的光)、音频的(例如,声音振动)和/或触觉的(用力、振动或运动引起的机械刺激)。
在另一方面,刺激可以是侵入性的或至少部分侵入性的。例如,视觉、音频和/或触觉刺激可以与组合物(例如,光-敏蛋白)或装置(例如,集成的光纤和固态光源)的注射或植入结合。
实验数据
下面结合各种附图列出与本文所述的发明构思有关的实验数据,这些附图首先被概述,然后进行详细说明。
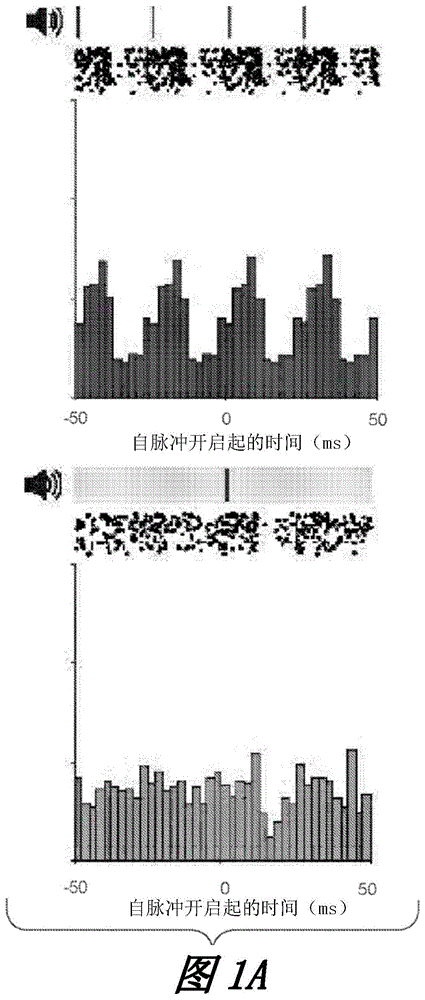
图1a示出了在ac中的40hz听觉刺激(每对的左图)和随机刺激(每对的右图)期间两个推定的单个单位的激发速率调节。刻点指示听觉脉冲;光条指示随机分布的脉冲。栅格图示出了两个实施例推定的单个单位对10秒的40hz听觉或随机刺激的尖峰反应。
图1b示出了对于所有单个单位在无刺激(标记为无刺激)、随机(标记为随机)和40hz听觉刺激(标记为40hz刺激)条件下,ac中的激发速率的峰之间的间隔分布(在5只小鼠中的9个记录阶段内n=292单位。刺激间间隔周围的间隔比例:p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)。
图1c示出了在40hz听觉刺激期间相对于刺激开始的激发速率调节(左边,刺激在0处开始)的示例性极坐标图、单个单位激发速率调节在40hz听觉刺激、随机和无刺激期间的矢量强度分布(中心,****p<0.0001,p=4x10-5440hz相对于无刺激,p=2x10-1340hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)、以及单个单位激发速率调节的瑞利统计值的分布(右边,****p<0.0001,p=5x10-6840hz相对于无刺激,p=6x10-7240hz相对于随机,柯尔莫哥洛夫-斯米尔诺夫检验;26个单位40hz的刺激rs值大于30;1个单位的随机刺激rs值大于30)。
图1d示出了ac中刺激条件之间的平均激发速率值的分布。
图1e对于ca1示出了与图1a相同的内容。
图1f对于ca1示出了与图1b相同的内容(在5只小鼠中的10个记录阶段内n=338单位。p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)。
图1g对于ca1示出了与图1c相同的内容(中心,****p<0.0001,p=4x10-4040hz相对于无刺激,p=9x10-1140hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=1x10-7440hz相对于无刺激,p=2x10-7340hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)。
图1h对于ca1示出了与图1d相同的内容。
图1i对于mpfc示出了与图1a相同的内容。
图1j对于mpfc示出了与图1b相同的内容(在4只小鼠中的7个记录阶段内n=115单位。p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)。
图1k对于mpfc示出了与图1c相同的内容(中心,****p<0.0001,p=2x10-2740hz相对于无刺激,p=4x10-540hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=1x10-2840hz相对于无刺激,p=5x10-3040hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)。
图1l对于mpfc示出了与图1d相同的内容。
图2a示出了5xfad听觉genus小鼠的行为实验的时间线。
图2b示出了5xfad听觉genus小鼠的新物体识别(nor)测试的识别指标(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,圆圈指示'n',条形图中的平均值s.e.m.,****p<0.0001,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2c示出了新物体识别测试期间的平均速度(cm/s)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2d示出了新物体识别测试期间新近的总距离(cm)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2e示出了5xfad听觉genus小鼠的新物体定位(nol)测试的识别指标(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,****p<0.0001,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2f示出了新物体定位测试期间的平均速度(cm/s)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2g示出了新物体定位测试期间新近的总距离(cm)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2h示出了5xfad非刺激、随机频率以及40hz听觉刺激小鼠在morris水迷宫中的逃避潜伏期(s)(无刺激组中n=25只小鼠,40hz组中n=28只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,****p<0.0001,n.s.=不显著,双因素方差分析与图基多重比较检验)。
图2i示出了在探测试验期间在目标象限内游泳所花费的时间(s)(无刺激组中n=25只小鼠,40hz组中n=28只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图2j示出了在探测试验期间平台穿越的次数(无刺激组中n=25只小鼠,40hz组中n=28只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,**p<0.01,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图3a示出了归一化至非刺激对照的、在40hz、8hz、80hz或随机频率听觉刺激(每天1小时,持续7天)后,6月龄5xfad小鼠的听觉皮层(ac)和海马体(hpc)中的相对可溶性aβ1-42水平(无刺激组中n=19只小鼠,40hz组中n=19只小鼠,8hz组中n=4只小鼠,80hz组中n=7只小鼠,随机频率组中n=6,条形图中的平均值s.e.m.,****p<0.0001,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图3b对于不溶性aβ1-42示出了与图3a相同的内容。
图3c示出了在听觉genus或无刺激(每天1小时,持续7天)后,5xfad小鼠的6月龄ac中用抗aβ(d54d2,绿色)抗体产生的免疫组织化学(每组n=7只小鼠,比例尺,50μm)。
图3d对于ca1示出了与图3c中相同的内容。
图3e示出了ac和ca1中的aβ阳性斑块的平均数量(每组n=7只小鼠,条形图中的平均值s.e.m.,*p<0.05,****p<0.0001;未配对的曼-惠特尼检验)。
图3f示出了ac和ca1中的aβ阳性斑块的平均面积(每组n=7只小鼠,条形图中的平均值s.e.m.,**p<0.01,***p<0.001;未配对的曼-惠特尼检验)。
图3g示出了在听觉genus或无刺激(每天1小时,持续7天)后,5xfad小鼠的6月龄ac中用抗aβ(12f4,红色)抗体产生的免疫组织化学(插图,20倍,比例尺,50μm)。
图3h对于ca1示出了与图3g中相同的内容。
图3i示出了归一化至非刺激对照的aβ(12f4)平均强度值(12f4抗体)(每组n=7只小鼠,条形图中的平均值s.e.m.,****p<0.0001,未配对的曼-惠特尼检验)。
图4a示出了每天1小时无刺激或听觉genus7天后,在5xfad小鼠的ac中用抗iba1(019-19741,绿色)和抗aβ(12f4,红色)抗体产生的免疫组织化学(每组n=8只小鼠,比例尺,50μm)。
图4b对于ca1示出了与图4a中相同的内容。
图4c示出了ac和ca1中的iba1阳性小神经胶质的数量(每组n=8只小鼠,条形图中的平均值s.e.m.,*p<0.05;未配对的曼-惠特尼检验)。
图4d示出了归一化至非刺激对照的ac和ca1中的iba1阳性小神经胶质细胞体的直径(每组n=8只小鼠,条形图中的平均值s.e.m.,****p<0.0001;未配对的曼-惠特尼检验)。
图4e示出了归一化至非刺激对照的ac和ca1中的iba1阳性小神经胶质原齿突的平均长度(每组n=8只小鼠,条形图中的平均值s.e.m.,****p<0.0001;未配对的曼-惠特尼检验)。
图4f示出了归一化至非刺激对照的ac和ca1中的iba1阳性小神经胶质的平均齿突分枝(每组n=8只小鼠,条形图中的平均值s.e.m.,*p<0.05,**p<0.01;未配对的曼-惠特尼检验)。
图4g示出了在ac和ca1中也为aβ阳性的iba1阳性小神经胶质细胞体的百分比(每组n=8只小鼠,条形图中的平均值s.e.m.,**p<0.01,***p<0.001;未配对的曼-惠特尼检验)。
图4h示出了在每天1小时无刺激或听觉genus7天后,在5xfad小鼠的ac中用抗s100b(ab868,紫色)和抗gfap(ab4674,灰色)抗体产生的免疫组织化学(每组n=8,比例尺,50μm)。
图4i对于ca1示出了与图4h中相同的内容。
图4j示出了ac和ca1中的s100b阳性星形细胞的数量(每组n=8只小鼠,*p<0.05;未配对的曼-惠特尼检验)。
图4k对于gfap阳性星形细胞示出了与图4i中相同的内容。
图5a示出了在每天1小时无刺激或听觉genus7天后,在6月龄的5xfad小鼠的ac中用凝集素染色剂(dl-1174,绿色)产生的免疫组织化学(比例尺,50μm)。
图5b对于ca1示出了与图5a中相同的内容。
图5c示出了归一化至无刺激对照的、在每天1小时无刺激或听觉genus7天后,在6月龄的5xfad小鼠的ac和ca1中的血管直径的百分比倍数变化(每组n=7只小鼠,条形图中的平均值s.e.m.,****p<0.0001;未配对的曼-惠特尼检验)。
图5d示出了在每天1小时无刺激或听觉genus7天后,在6月龄的5xfad小鼠的ac中用抗lrp1(28320,红色)、抗aβ(ab9234,绿色)以及凝集素染色剂(dl-1174,灰色)抗体产生的免疫组织化学(每组n=8只小鼠,比例尺,50μm)。
图5e对于ca1示出了与图5d中相同的内容。
图5f示出了在每天1小时无刺激或听觉genus7天后,在5xfad小鼠的ac和ca1中的aβ-lrp1共定位的百分比(每组n=8只小鼠,*p<0.05;未配对的曼-惠特尼检验)。
图6a示出了单个单位在40hz视听刺激期间的激发速率调节(左,下)。栅格图示出了两个实施例推定的单个单位对10秒的40hz听觉或随机刺激的尖峰反应(左,上)。40hz视听刺激、随机视听刺激和无刺激时段的矢量强度分布(右边,****p<0.0001,p=9x10-5940hz相对于无刺激,p=1x10-1340hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)。
图6b对于ca1示出了与图6a相同的内容(右边,****p<0.0001,p=6x10-4140hz相对于无刺激,p=2x10-1140hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)。
图6c对于mpfc示出了与图6a相同的内容(右边,****p<0.0001,p=2x10-2340hz相对于无刺激,p=9x10-540hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验)。
图6d示出了在每天1小时无刺激7天后,在6月龄的5xfad小鼠的ac、vc、ca1和mpfc中使用抗iba1(019-19741)和抗aβ(12f4)抗体的imaris(方法)产生的免疫组织化学和3d重建(每组n=6只小鼠,顶部插图:使用imaris来量化围绕淀粉样斑周围25μm半径的小神经胶质的数量的实施例。斑块被展示为红点,小神经胶质被展示为绿点,白色箭头指向簇。底部插图:来自ac的放大合并图像。比例尺,20μm)。
图6e对于组合的genus示出了与图6d中相同的内容。
图6f示出了归一化至无刺激对照的、在每天1小时无刺激或组合genus(a+v刺激)7天后,在6月龄的5xfad小鼠的ac、vc、ca1和mpfc中的平均小神经胶质细胞体直径(无对照组中n=6只小鼠,组合genus组中n=7只小鼠,条形图中的平均值s.e.m.,****p<0.0001;未配对的曼-惠特尼检验)。
图6g示出了归一化至无刺激对照的、在每天1小时无刺激或组合genus7天后,在6月龄的5xfad小鼠的ac、vc、ca1和mpfc中的平均小神经胶质齿突长度(无对照组中n=6只小鼠,组合genus组中n=7只小鼠,条形图中的平均值s.e.m.,**p<0.01,****p<0.0001;未配对的曼-惠特尼检验)。
图6h示出了在每天1小时无刺激或组合genus7天后在6月龄的5xfad小鼠的ac、vc、ca1和mpfc中的每个感兴趣区域的小神经胶质计数(无对照组中n=6只小鼠,组合genus组中n=7只小鼠,条形图中的平均值s.e.m.,*p<0.05,**p<0.01,未配对的曼-惠特尼检验)。
图6i示出了在无刺激或组合genus之后的ac、vc、ca1和mpfc中的斑块的25μm半径周围的小神经胶质的平均数量(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05;未配对的曼-惠特尼检验)。
图7a示出了在每天1小时无刺激7天后,在6月龄的5xfad小鼠的ac、vc、ca1和mpfc中的抗aβ斑块(d54d2,绿色)抗体的免疫组织化学(用40倍物镜拍摄的图像,比例尺,50μm)。
图7b对于组合的genus示出了与图7a中相同的内容。
图7c示出了归一化至无刺激对照的、在每天1小时的无刺激、40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz以及组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1、mpfc和vc中的平均斑块核心面积(每组n=12只小鼠,条形图中的平均值sem,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图7d示出了归一化至无刺激对照的、在每天1小时的无刺激、40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz以及组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1、mpfc和vc中的平均斑块数量(每组n=12只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图7e示出了归一化至非刺激对照的、在每天1小时的40hz听觉刺激、组合(a+v)genus、组合(a+v)8hz或组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的mpfc中的相对可溶性aβ1-42水平(每组n=4-5只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图7f对于不溶性aβ1-42示出了与图7e中相同的内容(n.s.=不显著,*p<0.05)。
图7g示出了在每天1小时无刺激7天后,6月龄5xfad小鼠的抗aβ斑块(d54d2,白色)的shield处理的全脑(大脑的25μm切片的矢状面)的免疫组织化学(光片照明显微镜,比例尺,700μm)。
图7h对于组合的(a+v)genus示出了与图7g中相同的内容。
图7i示出了在无刺激或组合的(a+v)genus之后的平均皮层斑块数量(每组n=6只小鼠,条形图中的平均值s.e.m.,*p<0.05;未配对的曼-惠特尼检验)。
图7j示出了在组合的(a+v)genus之后的平均皮层斑块体积(μm3)(每组n=6只小鼠,条形图中的平均值s.e.m.,*p<0.05;未配对的曼-惠特尼检验)。
图8a示出了对用于检测听觉皮层的听觉映射音调的平均lfp反应(左边)。蓝色区域指示何时播放50ms映射音调。簇聚推定的单个单位的实施例(右边)。
图8b示出了具有跨记录日的平均值和标准偏差的对40hz听觉闪烁刺激和无刺激时段的功率谱密度(psd)反应(左边)、ac中对所有记录日的听觉闪烁的功率谱lfp反应(示出了每个记录深度在40hz听觉闪烁期间具有最大40hz峰值的记录位点,参见方法)(右边)。
图8c示出了单个单位在40hz听觉刺激相对于无刺激时段期间在ac中的平均激发速率(fr)(左边)、单个单位在0hz周围在ac中心中在多个刺激条件之间的平均激发速率差(右边,****p<0.000140hz–无刺激,其他所有n.s.;对零中值进行威氏符号秩次检验)。
图8d示出了响应于20hz音频闪烁刺激的推定的单个单位的激发速率调节(左,下),栅格图示出了响应于10s刺激的尖峰(左,上)。对20hz音频刺激的激发速率反应中的峰值之间的间隔分布(右边,刺激间间隔周围的间隔比例:p=020hz相对于无刺激;对于两个比例进行z检验)。
图8e示出了响应于80hz音频闪烁刺激的d所示的同一单位的激发速率调节(左,下),栅格图示出了响应于10s刺激的尖峰(左,上)。对80hz音频刺激的激发速率反应中的峰值之间的间隔分布(右边,刺激间间隔周围的间隔比例:p=080hz相对于无刺激;对于两个比例进行z检验)。
图8f示出了20hz和80hz听觉刺激相对于无刺激条件的矢量强度分布(左边,****p<0.0001,p=3x10-6120hz相对于无刺激,p=3x10-6180hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)以及20hz和80hz听觉刺激相对于无刺激的瑞利统计分布(右边,****p<0.0001,p=3x10-7320hz相对于无刺激,p=1x10-6880hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;54个单位的20hz刺激rs值大于30;28个单位的80hz刺激rs值大于30)。
图8g示出了用于检测ca1的θ节律(海马体的标志)的实施例。
图8h对于ca1示出了与b相同的内容。
图8i对于ca1示出了与c相同的内容(右边,n.s.;对于零中值进行威氏符号秩次检验)。
图8j对于ca1示出了与d相同的内容(右边,p=020hz相对于无刺激;对于两个比例进行z检验)。
图8k对于ca1示出了与e相同的内容(右边,p=080hz相对于无刺激;对于两个比例进行z检验)。
图8l对于ca1示出了与f相同的内容(左边,****p<0.0001,p=1x10-4020hz相对于无刺激,p=9x10-4580hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=1x10-7120hz相对于无刺激,p=8x10-7380hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)。
图8m示出了组织学图像,该图像示出了mpfc中的探测轨迹和记录位置。红色箭头指示记录位置。
图8n对于mpfc示出了与b相同的内容。
图8o对于mpfc示出了与c相同的内容(右边,n.s.;对于零中值进行威氏符号秩次检验)。
图8p对于mpfc示出了与d相同的内容(右边,p=020hz相对于无刺激;对于两个比例进行z检验)。
图8q对于mpfc示出了与e相同的内容(右边,p=080hz相对于无刺激;对于两个比例进行z检验)。
图8r对于mpfc示出了与f相同的内容(左边,****p<0.0001,p=1x10-2320hz相对于无刺激,p=6x10-2480hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=2x10-1720hz相对于无刺激,p=4x10-2680hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)。
图9a示出了在5xfad非刺激、40hz和随机频率刺激的小鼠的nor测试期间探索熟悉和新物体花费的时间(秒)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,**p<0.01,****p<0.0001,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9b示出了小鼠在nor测试期间达到20s的总物体探索要求所需的时间(分钟)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9c示出了在5xfad非刺激、40hz和随机频率刺激的小鼠的nol测试期间探索熟悉和新位置中的物体花费的时间(秒)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,***p<0.001,****p<0.0001,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9d示出了小鼠在nol测试期间达到20s的总物体探索要求所需的时间(分钟)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9e示出了习惯化期间的平均速度(cm/s)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9f示出了在习惯化期间行进的总距离(cm)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)
图9g示出了在习惯化期间在行为室的中心花费的时间(秒)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9h示出了在习惯化期间在行为室的周边花费的时间(秒)(无刺激组中n=20只小鼠,40hz组中n=20只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9i示出了在morris水迷宫期间的平均游泳速度(cm/s)(无刺激组中n=25只小鼠,40hz组中n=28只小鼠,随机频率组中n=9,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9j示出了在1小时无刺激、听觉genus或随机频率刺激期间的平均速度(cm/s)(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,随机频率组中n=6,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9k示出了在1小时无刺激、听觉genus或随机频率刺激期间行进的总距离(cm)(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,随机频率组中n=6,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图9l示出了在1小时无刺激、听觉genus或随机频率刺激期间在2cm/s下花费的时间(秒)(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,随机频率组中n=6,条形图中的平均值s.e.m.,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图10a示出了归一化至非刺激对照的、在40hz、8hz、80hz或随机频率听觉刺激(每天1小时,持续7天)后,6月龄5xfad小鼠的听觉皮层(ac)和海马体(hpc)中的相对可溶性aβ1-40水平(注意:用于80hz和随机频率hpc样品的elisa未成功且未能报道,无刺激组中n=19只小鼠,40hz组中n=19只小鼠,8hz组中n=4只小鼠,80hz组中n=7只小鼠,随机频率组中n=6,条形图中的平均值s.e.m.,**p<0.01,n.s.=不显著,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图10b示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的6月龄app/ps1小鼠的听觉皮层(ac)和海马体(hpc)中的相对可溶性aβ1-42水平(无刺激组中n=4只小鼠,40hz组中n=4只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图10c示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1(“感兴趣区域”,roi)中的平均斑块数量(无刺激组中n=5只小鼠,40hz组中n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图10d示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1中的平均斑块核心面积(无刺激组中n=5只小鼠,40hz组中n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图10e示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1中的aβ(12f4)平均强度值(12f4抗体)(无刺激组中n=5只小鼠,40hz组中n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图10f示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的平均斑块数量(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图10g示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的平均斑块核心面积(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图10h示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的aβ(12f4)平均强度值(12f4抗体)(无刺激组中n=6只小鼠,40hz组中n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图11a示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1中的iba1阳性小神经胶质细胞体的直径(每组n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图11b示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1中的iba1阳性小神经胶质原齿突的平均长度(每组n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图11c示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后的9月龄app/ps1小鼠的ac和ca1中的iba1阳性小神经胶质的数量(每组n=5只小鼠,条形图中的平均值s.e.m.,*p<0.05,未配对的曼-惠特尼检验)。
图11d示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的iba1阳性小神经胶质细胞体的直径(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图11e示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的iba1阳性小神经胶质原齿突的平均长度(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图11f示出了归一化至非刺激对照的、在听觉genus(每天1小时,持续7天)后7天无刺激后的6月龄5xfad小鼠的ac和ca1中的iba1阳性小神经胶质的数量(每组n=6只小鼠,条形图中的平均值s.e.m.,*p<0.05,n.s.=不显著,未配对的曼-惠特尼检验)。
图11g示出了在6月龄wt和5xfad小鼠的ca1中用抗gfap(ab4674,红色)和凝集素染色剂(dl-1174,绿色)抗体产生的clarity处理的脑片的免疫组织化学(每组n=5只小鼠,比例尺,50μm)。
图11h示出了6月龄wt和5xfad小鼠的ca1中的gfap阳性细胞的数量(每个感兴趣图像,使用imaris)(每组n=5只小鼠,条形图中的平均值s.e.m.,**p<0.01,未配对的曼-惠特尼检验)。
图12a示出了在每天1小时无刺激或听觉genus7天后,在6月龄的p301s小鼠的ac中用抗ptau(t181,红色)抗体产生的免疫组织化学(用40倍物镜拍摄的图像,比例尺,50μm)。
图12b对于ca1示出了与图12a中相同的内容。
图12c示出了归一化至非刺激对照的、在每天1小时无刺激或听觉genus7天后,在p301s小鼠的ac和ca1中的相对ptau(t181)强度水平(每组n=10只小鼠,***p<0.001,****p<0.0001;未配对的曼-惠特尼检验)。
图12d示出了在每天1小时无刺激或听觉genus7天后,在6月龄的p301s小鼠的ac中用抗ptau(s396,绿色)抗体产生的免疫组织化学(比例尺,50μm)。
图12e对于ca1示出了与图12d中相同的内容。
图12f示出了归一化至非刺激对照的、在每天1小时无刺激或听觉genus7天后,p301s小鼠中在ac和ca1中的相对ptau(s396)强度水平(每组n=10只小鼠,****p<0.0001;未配对的曼-惠特尼检验)。
图12g示出了代表性蛋白质印记,该蛋白质印记示出了在每天1小时无刺激或听觉genus7天后,在6月龄的p301s小鼠的ac中的ptau(s396)和总tau的水平。
图12h对于海马体示出了与图12g中相同的内容。
图12i示出了代表性蛋白质印记,该蛋白质印记示出了在每天1小时无刺激或听觉genus7天后,在6月龄的p301s小鼠的海马体中的ptau(t181)和总tau的水平。
图12j示出了在每天1小时无刺激或听觉genus7天后,归一化至p301s小鼠的ac中的总tau(来自g中的蛋白质印迹)的ptau(s396)的相对免疫反应性(每组n=3只小鼠,条形图中的平均值s.e.m.,*p<0.05;未配对的曼-惠特尼检验)。
图12k示出了在每天1小时无刺激或听觉genus7天后,归一化至p301s小鼠的hpc中的总tau(来自h中的蛋白质印迹)的ptau(s396)的相对免疫反应性(40hz组中=2只小鼠且非刺激组中n=3,条形图中的平均值s.e.m.,***p<0.001;未配对的曼-惠特尼检验)。
图12l示出了在每天1小时无刺激或听觉genus7天后,归一化至p301s小鼠的海马体中的总tau(来自i中的蛋白质印迹)的ptau(t181)的相对免疫反应性(40hz组中=2只小鼠且非刺激组中n=3,条形图中的平均值s.e.m.,**p<0.01;未配对的曼-惠特尼检验)。
图13a示出了具有跨记录日的平均值和标准偏差的对40hz视听闪烁刺激和无刺激时段的功率谱密度(psd)反应(左边)、ac中对所有记录日的视听闪烁刺激的功率谱lfp反应(示出了每个记录深度在40hz视听闪烁期间具有最大40hz峰值的记录位点,参见方法)(右边)。
图13b示出了图13a所示的推定的单个单位到视听随机刺激的激发速率调节(下);栅格图(上)示出了单个单位到10s随机刺激的尖峰(左边)。ac中对视听刺激的激发速率反应峰值之间的间隔分布(中心,刺激间间隔周围的间隔比例:p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)、单个单位对40hz视听刺激的瑞利统计分布(右边,****p<0.0001,p=2x10-7940hz相对于无刺激,p=1x10-7240hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验;20个单位的40hz刺激rs值大于30;2个单位的随机刺激rs值大于30。
图13c示出了在所有视听刺激条件期间的单个单位平均激发速率。
图13d示出了响应于20hz视听闪烁刺激的推定的单个单位的激发速率调节(左,下),栅格图示出了响应于10s刺激的尖峰(左,上)。对20hz视听刺激的激发速率反应中的峰值之间的间隔分布(右边,刺激间间隔周围的间隔比例:p=020hz相对于无刺激;对于两个比例进行z检验)。
图13e示出了响应于80hz视听闪烁刺激的d所示的同一单位的激发速率调节(左,下),栅格图示出了响应于10s刺激的尖峰(左,上)。对80hz视听刺激的激发速率反应中的峰值之间的间隔分布(右边,刺激间间隔周围的间隔比例:p=080hz相对于无刺激;对于两个比例进行z检验)。
图13f示出了20hz和80hz听觉刺激高于无刺激条件的矢量强度分布(左边,****p<0.0001,p=2x10-6320hz相对于无刺激,p=1x10-5680hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)以及20hz和80hz听觉刺激高于无刺激的瑞利统计分布(右边,****p<0.0001,p=1x10-8820hz相对于无刺激,p=5x10-7280hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;50个单位的20hz刺激rs值大于30;19个单位的80hz刺激rs值大于30)。
图13g示出了单个单位在0hz周围在ac中心中在多个刺激条件之间的平均激发速率差(*p<0.0520hz–80hz,*p<0.0520hz–40hz,其他所有n.s.;对零中值进行威氏符号秩次检验)。
图13h对于ca1示出了与a相同的内容。
图13i对于ca1示出了与b相同的内容(右边,****p<0.0001,p=1x10-7140hz相对于无刺激,p=1x10-7140hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验;中心,刺激间间隔周围的间隔比例:p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)。
图13j对于ca1示出了与c相同的内容。
图13k对于ca1示出了与d相同的内容(右边,p=020hz相对于无刺激;对于两个比例进行z检验)。
图13l对于ca1示出了与e相同的内容(右边,p=080hz相对于无刺激;对于两个比例进行z检验)。
图13m对于ca1示出了与f相同的内容(左边,****p<0.0001,p=8x10-4320hz相对于无刺激,p=8x10-4080hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=2x10-7020hz相对于无刺激,p=1x10-5780hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)。
图13n对于ca1示出了与g相同的内容(所有n.s.;对于零中值进行威氏符号秩次检验)。
图13o对于mpfc示出了与a相同的内容。
图13p对于mpfc示出了与b相同的内容(右边,****p<0.0001,p=5x10-2340hz相对于无刺激,p=3x10-2140hz相对于随机;柯尔莫哥洛夫-斯米尔诺夫检验;中心,刺激间间隔周围的间隔比例:p=040hz相对于无刺激,p=040hz相对于随机;对于两个比例进行z检验)。
图13q对于mpfc示出了与c相同的内容。
图13r对于mpfc示出了与d相同的内容(右边,p=020hz相对于无刺激;对于两个比例进行z检验)。
图13s对于mpfc示出了与e相同的内容(右边,p=080hz相对于无刺激;对于两个比例进行z检验)。
图13t对于mpfc示出了与f相同的内容(左边,****p<0.0001,p=1x10-2320hz相对于无刺激,p=2x10-2580hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验;右边,****p<0.0001,p=1x10-2320hz相对于无刺激,p=8x10-2580hz相对于无刺激;柯尔莫哥洛夫-斯米尔诺夫检验)。
图13u对于mpfc示出了与g相同的内容(*p<0.0540hz-无刺激,所有其他n.s.;对于零中值进行威氏符号秩次检验)。
图14a示出了每天1小时听觉genus7天后,在6月龄5xfad小鼠的ac、ca1和mpfc中的抗iba1(019-19741,绿色)和抗aβ(12f4,红色)抗体的免疫组织化学(插图放大,100倍;比例尺,50μm)。
图14b示出了每天1小时视觉genus7天后,在6月龄5xfad小鼠的vc、ca1和mpfc中的抗iba1(019-19741,绿色)和抗aβ(12f4,红色)抗体的免疫组织化学(插图放大,100倍;比例尺,50μm)。
图14c示出了归一化至无刺激对照的、在每天1小时听觉genus7天后,在6月龄5xfad小鼠的ac、ca1和mpfc中的平均小神经胶质细胞体直径(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,**p<0.01;未配对的曼-惠特尼检验)。
图14d示出了归一化至无刺激对照的、在每天1小时听觉genus7天后,在6月龄5xfad小鼠的ac、ca1和mpfc中的平均小神经胶质齿突长度(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,**p<0.01;未配对的曼-惠特尼检验)。
图14e示出了在每天1小时听觉genus7天后,在6月龄5xfad小鼠的ac、ca1和mpfc中的每个感兴趣区域的小神经胶质计数(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,**p<0.01;未配对的曼-惠特尼检验)。
图14f示出了在无刺激或听觉genus之后的ac、ca1和mpfc中的斑块的25μm半径周围的小神经胶质的平均数量(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图14g示出了归一化至无刺激对照的、在每天1小时视觉genus7天后,在6月龄5xfad小鼠的vc、ca1和mpfc中的平均小神经胶质细胞体直径(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05;未配对的曼-惠特尼检验)。
图14h示出了归一化至无刺激对照的、在每天1小时视觉genus7天后,在6月龄5xfad小鼠的vc、ca1和mpfc中的平均小神经胶质齿突长度(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05;未配对的曼-惠特尼检验)。
图14i示出了在每天1小时视觉genus7天后,在6月龄5xfad小鼠的vc、ca1和mpfc中的每个感兴趣区域的小神经胶质计数(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图14j示出了在无刺激或视觉genus之后的vc、ca1和mpfc中的斑块的25μm半径周围的小神经胶质的平均数量(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,未配对的曼-惠特尼检验)。
图14k示出了归一化至非刺激对照的、在每天1小时的40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz或组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1和mpfc中的平均小神经胶质细胞体直径(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,**p<0.01;未配对的曼-惠特尼检验)。
图14l示出了归一化至非刺激对照的、在每天1小时的40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz或组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1和mpfc中的平均小神经胶质齿突长度(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,未配对的曼-惠特尼检验)。
图14m示出了归一化至非刺激对照的、在每天1小时的40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz或组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1和mpfc中的每个感兴趣区域的小神经胶质计数(每组n=6只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,未配对的曼-惠特尼检验)。
图14n示出了在每天1小时听觉genus7天后,在6月龄5xfad小鼠的ac、ca1和mpfc中的抗aβ斑块(d54d2,绿色)抗体的免疫组织化学(每组n=6只小鼠,比例尺,50μm)。
图14o示出了在每天1小时视觉genus7天后,在6月龄5xfad小鼠的vc、ca1和mpfc中的抗aβ斑块(d54d2,绿色)抗体的免疫组织化学(每组n=6只小鼠,比例尺,50μm)。
图14p示出了归一化至无刺激对照的、每个感兴趣区域的平均斑块核心面积(每组n=6,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,未配对的曼-惠特尼检验)。
图14q示出了归一化至无刺激对照的、听觉genus之后的ac、ca1和mpfc中的平均斑块数量(每组n=6,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,***p<0.001,未配对的曼-惠特尼检验)。
图14r示出了归一化至无刺激对照的、每个感兴趣区域的平均斑块核心面积(每组n=6,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,未配对的曼-惠特尼检验)。
图14s示出了归一化至无刺激对照的、视觉genus之后的vc、ca1和mpfc中的平均斑块数量(每组n=6,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,未配对的曼-惠特尼检验)。
图14t示出了归一化至无刺激对照的、在每天1小时的无刺激、40hz听觉刺激、组合(a+v)genus、组合(a+v)8hz以及组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac和hpc中的相对可溶性aβ1-42水平(每组n=4-5,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图14u示出了归一化至无刺激对照的、在每天1小时的无刺激、40hz听觉刺激、组合(a+v)genus、组合(a+v)8hz以及组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac和hpc中的相对不溶性aβ1-42水平(每组n=4-5,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
图14v示出了归一化至无刺激对照的、在每天1小时的无刺激、40hz听觉刺激、组合(a+v)genus、组合(a+v)80hz以及组合(a+v)随机频率刺激7天后的6月龄5xfad小鼠的ac、ca1和mpfc中的aβ(12f4)平均强度值(12f4抗体)(每组n=4-5只小鼠,条形图中的平均值s.e.m.,n.s.=不显著,*p<0.05,克鲁斯卡尔-沃利斯检验与邓恩多重比较检验)。
40hz的听觉刺激调节ac、ca1和mpfc中的尖峰活性
我们首先确定听觉音调刺激是否可以在听觉皮层(ac)、海马体亚区ca1和内侧前额叶皮层(mpfc)中产生genus。我们向动物呈现20hz、40hz、80hz的音调序列、或随机间隔的音调序列(每12.5ms、25ms、50ms或以平均25ms的随机化音调间间隔播放1ms长、10khz音调,此后称为"听觉闪烁刺激")。在音调呈现期间,我们在球形跑台上奔跑或休息的3-8月龄雄性野生型(c57bl6j)小鼠中的ac、ca1和mpfc中使用32通道硅探头执行电生理记录。为了定位ac,在不同的深度播放一系列的50ms听觉映射音调(此后称为“映射刺激”),直到在音调开始后约20ms检测到瞬时lfp反应为止(图8a)。基于电生理学标志定位ca1,并且在每只动物中进行最终记录后利用组织学确认mpfc记录位置(图8g和图8m)。
在我们到达目标区域后,向动物呈现安静和听觉闪烁刺激的交错时段,同时记录神经活动。刺激块在20hz、40hz、80hz以及随机的听觉闪烁刺激之间轮换。推定的单个单位的激发速率随着每个音调而周期性地增加和降低,从而夹带到40hz的听觉闪烁刺激(图1a、图1e和图1i,蓝色)。还通过随机刺激来调节单位。当所有随机脉冲对齐时,激发速率调节在刺激后发生变化,这指示单个单位对随机刺激脉冲作出反应。然而,听觉音调的随机序列不会引起周期性的激发调制(图1a、图1e和图1i,橙色)。夹带在相位分布和振幅这两者上在单个单位之间变化。在闪烁刺激期间,神经元根据刺激而激发,但它们并非在每个循环都激发,并且通常在很宽的相位范围内激发(图1a、图1e和图1i)。在大多数单个单位中,听觉闪烁刺激期间的激发速率峰值之间的间隔约为25ms(相当于40hz);ac中为75%,ca1中为79%,mpfc中为74%。(图1b、图1f和图1j)。
相反,在没有音调的基准时段和具有随机音调的时段期间,峰值之间的间隔具有较宽分布,ac中少于11%的细胞、ca1中12%的细胞以及mpfc中16%的细胞具有约25ms的峰值间隔(即,激发速率未在40hz下调节;图1b、图1f和图1j)。通过把单个单位激发速率看作刺激相位的函数并计算其矢量强度(vs)来量化调节强度(图1c、图1g和图1k,左边)。矢量强度值的范围是0到1,其中0表示未被刺激调节的均匀激发分布(vs=0),而1表示仅将神经元激发到特定刺激相位的分布(vs=1)。对40hz听觉刺激的单个单位反应的矢量强度的分布在ac中在0.002至1的范围内,在hpc中在0.0005至1的范围内,在pfc中在0.1至0.6的范围内,并且显著高于无刺激以及随机刺激的分布(图1c、图1g和图1k,中心)。随机刺激矢量强度也显著高于无刺激条件,因为矢量强度测量了由刺激进行的调节。然而,矢量强度未量化调节的周期性,并且虽然随机刺激确实引起单个单位反应,但它不引起周期性的激发调节。
相似地,单个单位在40hz听觉刺激期间的瑞利统计分布显著高于无刺激和随机刺激对照的分布(图1c、图1g和图1k,右边)。在40hz听觉闪烁刺激以及无刺激、随机刺激、20hz和80hz听觉闪烁刺激的对照之间,单个神经元的平均激发速率相似(图1d、图1h和图1l;图8c,图1i和图1o,右边)。ac中的局部场电位在听觉闪烁刺激期间在40hz处显示出提高的功率,但效果在记录位置、记录阶段和对映射音调的反应潜伏期之间变化(图8b、图1h和图1n)。这些发现表明40hz听觉闪烁刺激在ac、ca1和mpfc中强劲地引起genus。
听觉genus改善5xfad小鼠的记忆效能
假定听觉genus能够影响海马体神经活性,我们接下来评估其对6月龄5xfad小鼠的海马体相关学习和记忆的作用(图2a)。我们使用6月龄5xfad动物,因为这是行为损害首次变得明显的时候。对于所有后续实验,我们执行了1周的听觉genus;具体地,将小鼠置于安静的室中,并将其暴露于1ms长的10khz听觉音调的序列,每天1小时,持续7天,频率为40hz(由此,40(10khz)音调/秒)。我们首先在测试新物体识别(nor)和新物体定位(nol)记忆效能前24小时使小鼠习惯于行为室,这些效能评估了记住物体在特定环境中的身份或放置(即,已知在人类ad受试者中受影响的行为)的能力。这些测试使用识别指标来测量行为效能,所述识别指标是探索新物体或新位置物体分别花费的时间在探索的整个持续时间中的百分比。
在习惯化期间,听觉genus、随机频率或未刺激组均未示出平均速度、总距离、在中心花费的时间或在周边花费的时间的显著变化,这指示三组未示出一般活动或焦虑样行为的差异(图9e至图9h)。在听觉genus之后,5xfad小鼠表现出对物体记忆任务的显著更高的识别指标(65.50±1.40%)和对位置记忆任务的显著更高的识别指标(61.41±2.0%),而非刺激和随机频率对照组在两个测试中均未显示出对新物体或新位移物体的偏好(图2b和图2e)。在三组之间的任务时段期间,行进的距离或平均速度没有显著的差异,这指示这些影响并非归于活动的一般差异(图2c、图2d、图2f和图2g)。
检查了在nor期间探索新物体和熟悉物体所花费的时间量;我们观察到听觉genus之后的小鼠关于新物体花费显著更高的时间量,而非刺激和随机频率对照组未表现出探索偏好(图9a)。相似地,我们观察到听觉genus之后的小鼠关于新位置中的物体(nol)花费显著更高的时间量,而非刺激和随机频率对照组未表现出探索偏好(图9c)。作为检查探索活动差异的另外对照措施,我们测量了小鼠在物体任务期间达到20s的目标探索要求所花费的时间量(分钟)。我们观察到在三组之间达到物体探索要求所花费的时间没有显著差异(图9b和图9d)。
为了进一步表征听觉genus对海马体相关行为的影响,我们执行morris水迷宫测试,该测试测量了记住隐藏平台相对于周围环境线索的位置的能力。小鼠在连续的试验期间逐渐学习隐藏平台的位置,并且它们对平台位置的空间记忆通过逃避潜伏期来测量,逃避潜伏期是指单个小鼠找到隐藏平台所花费的时间量。在训练阶段期间,所有三组都能够成功地学习隐藏平台的位置,然而,接收听觉genus的组的逃避潜伏期始终且显著短于非刺激和随机频率对照组这两者(图2h)。三组之间的游泳速度没有显著差异(图9i)。在探测试验期间,或在从箱去除隐藏平台时,与非刺激和随机频率对照组这两者比较,接收听觉genus的小鼠花费探索包含缺失平台的象限的显著更长的时间段,并且在之前的平台位置上显示出更多的交叉(图2i和图2j)。
作为最终的行为测量,我们检查了5xfad小鼠在1小时听觉genus、无刺激或随机频率刺激期间的活动,并且观察到平均速度(cm/s)和行进距离(cm)没有显著差异(图9j和图9k)。为了探索在“睡眠”状态或静止期是否存在差异,我们测量了小鼠在1小时刺激组期间在2cm/s下花费的时间量。我们观察到三组之间没有显著差异(图9l)。这些结果一起示出,听觉genus可以改善6月龄5xfad小鼠的识别和空间记忆。
听觉genus减少5xfad小鼠的ac和海马体中的淀粉样负荷
听觉genus对认知功能的有益作用使我们研究了是否可以在5xfad小鼠模型中修改潜在的淀粉样病变。之前,我们示出了视觉genus对更年轻的3月龄小鼠的淀粉样负荷的改善作用。在这里,我们的目的是进一步研究听觉genus对6月龄小鼠的影响,这些小鼠处于更加进展的ad状态并表现出更高的淀粉样斑负荷。将小鼠置于安静的室中,并将其暴露于一系列不同的听觉音调序列频率(包括40hz、8hz、80hz、随机频率刺激)或无刺激。7天刺激完成后24小时,我们通过aβ酶联免疫吸附测定(elisa)分析了ac和整个海马体(hpc)中的淀粉样负荷。在40hz听觉刺激后,我们观察到,与没有音调或另外频率对照相比,可溶性aβ1-42水平在ac中降低了51.84±4.98%并且在hpc中降低了46.89±3.89%,而ac和hpc中的可溶性aβ1-40水平分别降低了20.65±3.21%和34.15±4.83%。(图3a和图10a)。相似地,不溶性aβ1-42水平在ac中降低了36.68±3.21%,并且在hpc中降低了43.84±2.42%(图3b)。在听觉genus或无刺激对照这两者中均无法经由elisa检测到不溶性aβ1-40。
我们的结果指示,与非刺激对照相比,观察到的淀粉样减少对40hz刺激是特定的,因为8hz、80hz和随机频率刺激均不显著改变aβ水平。为了确定这些作用是否适用于其他ad小鼠模型以及我们的结果是否特定于我们的5xfad模型,我们在7天的听觉genus后检查了6月龄app/ps1转基因小鼠(即,一种经过充分验证的ad模型)的aβ水平。我们发现,与无刺激对照相比,可溶性aβ1-42在ac中显著降低了48.39±3.50%,并且在hpc中显著降低了35.54±4.27%(图10b)。
接下来,我们使用β-淀粉样特异性抗体(cellsignalingtechnology;d54d2)以免疫组织化学分析检查了5xfad小鼠模型中的斑块负荷(图3c和图3d)。与无刺激对照相比,在7天的听觉genus后,斑块数量在ac和ca1中分别显著减少了45.73±2.818%和59.30±2.083%(图3e)。斑块尺寸在ac和ca1中也分别显著减小了54.37±5.603%和40.70±5.321%(图3f)。对aβ1-42特异性免疫染色的分析指示,aβ1-42沉积物在ac和ca1中分别显著减少了45.35±0.011%和43.21±0.0285%(图3g至图3i)。为了检查40hz刺激后的斑块负荷的动态,我们在5xfad小鼠中使用了β-淀粉样特异性抗体(cellsignalingtechnology;d54d2)执行了免疫组织化学分析,这些小鼠首先经受一周的听觉genus,然后在接下来的7天内不进行刺激。我们观察到平均斑块数量、面积和淀粉样强度略微降低,然而,这些差异并不显著(参见图10f至图10h)。为了检查另一个ad模型中的听觉genus之后的斑块负荷,我们使用了9月龄的app/ps1小鼠,因为6月龄小鼠没有表现出显著的斑块发展。
我们观察到,斑块数量在ac中显著降低了52.65±7.53%,在ca1中显著降低了62.90±15.5%。斑块尺寸在ac中显著降低了67.90±6.18%,在ca1中显著降低了64.06±15.2%。使用aβ1-42抗体(biolegend;12f4)进行的aβ1-42特异性免疫染色的分析指示,与无刺激对照相比,aβ1-42沉积物在ac和ca1中分别显著减少了38.77±4.21%和47.63±6.08%(图10c至图10e)。共同地,这些结果证明,听觉genus可以在ac和ca1中夹带γ活性,并减少ad小鼠模型中的淀粉样负荷。
听觉genus在5xfad小鼠中引起神经胶质和血管反应
越来越多的证据表明,小神经胶质响应于神经元活性的变化,并在ad病变中起作用(allen和barres,2005年;mosher和wyss-coray,2014年;walker和lue,2015年)。我们降低ac和hpc中淀粉样负荷的能力使我们检查了听觉genus是否可以刺激6月龄5xfad小鼠的小神经胶质反应的变化。已经示出小神经胶质在涉及吞噬的激活状态期间改变其细胞形态(davies等人,2016年),并且实际上,我们的早期研究证明,1小时的视觉genus足以引起符合vc中的激活和增加的吞噬细胞活动的小神经胶质的形态学变化(iaccarino等人,2016年)。使用抗小神经胶质标记物iba1的抗体(图4a和图4b),与无刺激对照相比,我们在听觉genus组中观察到ac和ca1这两者中的小神经胶质多大约60%(图4c)。与无刺激对照相比,在听觉genus后,小神经胶质细胞体面积在ac中增加了70.60±4.78%,并且在ca1中增加了117.17±10.4%(图4d)。我们还发现,与无刺激对照相比,小神经胶质齿突长度降低了46.44±3.2%(ac)和50.875.±4.8%(ca1),并且齿突分枝增加了36.00±9.5%(ac)和143.813±29.9%(ca1)(图4e和图4f)。为了评估小神经胶质对aβ的摄取,我们通过用iba1和对aβ1-42特异的aβ抗体(12f4,参见方法)对组织切片进行共免疫染色来测量小神经胶质内aβ的共定位。我们观察到,与无刺激对照相比,在听觉genus后,细胞体与aβ共定位的小神经胶质的百分比在ac中增加了58.75±1.25%,并且在ca1中增加了61.33±3.71%(图4g)。
为了检查在其他ad小鼠模型中在听觉genus后是否发生小神经胶质反应,我们测量了来自7天的听觉genus后的9月龄app/ps1小鼠的小神经胶质形态。与我们在7天的听觉genus之后的5xfad小神经胶质中看到的结果相似(参见图4a至图4g),我们观察到,与无刺激对照相比,小神经胶质细胞体直径和计数显著增加,并且ac和ca1中的平均齿突长度显著减小(图11a至图11c)。
为了理解听觉genus后的5xfad小鼠中的小神经胶质反应的纵向影响,我们在一周听觉genus后7天无刺激后检查小神经胶质形态。我们观察到与来自淀粉样的趋势相似的趋势(图10f至图10h),具体是小神经胶质细胞体直径的无显著增加、平均齿突长度的减小以及听觉皮层中的小神经胶质计数的增加(参见图11d至图11f)。然而,与非刺激对照相比,我们确实看到了ca1中的小神经胶质计数的显著增加(增加41.70±6.75%)。
星形细胞是中枢神经系统中的另一种原发神经胶质细胞,并且对于稳态维持、突触修剪、废物清除和其他重要的生物过程(诸如调节脑血流量)是关键的(chung等人,2015年;kisler等人,2017年)。反应性星形细胞表达神经胶质纤维酸性蛋白(gfap)(eng等人,1971年)。为了研究反应性星形细胞数量在6月龄5xfad和wt同窝对照小鼠之间是否存在基线变化,我们对针对gfap抗体染色的100μm的ca1脑片执行了clarity(图11g)。我们观察到,与wt对照相比,5xfad小鼠的gfap阳性星形细胞数量显著减少(图11h)。该观察符合示出具有相似神经胶质萎缩的其他ad转基因小鼠模型的报道(rodríguez等人,2009年)。为了确定听觉genus是否可能影响星形细胞的反应性,我们使6月龄的5xfad小鼠经受了7天的每天1小时的听觉genus或无刺激对照,然后使用针对gfap和s100钙结合蛋白b(s100b)的抗体对它们的脑片进行了免疫染色,s100b经证实是在反应性星形细胞中表达的另一种蛋白质(图4h和图4i)。对于gfap阳性的星形细胞在ac和ca1中分别增加27.66±0.954%和18.14±0.799%,并且s100b阳性星形细胞在ac中增加21.83±1.07%,在ca1中增加15.57±0.869%(图4j和图4k)。该观察到的听觉genus后的星形细胞数量的变化可能指示星形细胞存活潜在增加。
已知星形细胞在调节大脑的血管网络中起着重要的作用,并且越来越多的证据表明该网络在ad中的功能障碍可能会加剧病变。从大脑清除淀粉样是多层面的,并且已经提出了通过脉管系统进行的各种过程,诸如通过淋巴系统和经由内吞受体脂蛋白受体相关蛋白1(lrp1)的转运。
为了研究脉管系统的潜在变化,我们首先使用了番茄凝集素(番茄(lycopersiconesculentum))(该凝集素是血管内皮的有效标记物)来在听觉genus之后对5xfad脑片进行染色(图5a和图5b)。有趣的是,我们观察到,与无刺激对照相比,在听觉genus后,血管直径增加了49.70±7.80%(ac)和104.70±10.96%(ca1)(图5c)。我们进一步探索了淀粉样-血管相互作用在听觉genus后是否发生变化。我们检查了听觉genus是否可能影响aβ与lrp1的共定位,这已通过在来自5xfad小鼠的脑片中对lrp1和aβ染色借助脉管系统来示出在aβ转运和系统消除中起重要作用(storck等人,2016年),这些小鼠暴露于1小时/天的听觉genus或无刺激7天(图5d和图5e)。在无刺激对照中,我们观察到aβ与lrp1的共定位为8.17±2.70%(ac)和6.97±1.73%(ca1),而在听觉genus组中,aβ与lrp1的共定位在ac和ca1中分别显著增加至17.71±2.78%和16.50±3.90%(图5f)。这些结果一起表明,听觉genus后ac和ca1中aβ水平降低的一种解释可能是通过小神经胶质对aβ的清除增加以及脉管系统的改变。
听觉genus减少ac和海马体中的tau磷酸化
ad的另一个经典病理标志是磷酸化的tau聚集体的积累。已经证明与ad关联的特定氨基酸残基处的tau磷酸化改变其细胞骨架支持功能并降低其溶解度,由此被认为是神经元的主要损害。为了研究听觉genus是否可能影响另一种与ad关联的小鼠模型的病变,我们使用了taup301s小鼠。由于taup301s小鼠在6月龄时开始表现出空间和情境学习障碍,座椅我们检查了听觉genus是否可以导致6月龄taup301s小鼠的ac和hpc中的磷酸化tau减少。对来自这些小鼠的脑片的免疫组织化学分析(图12a、图12b、图12d和图12e)指示,听觉genus使苏氨酸-181(t181)处的tau磷酸化降低了36.20±2.828%(ac)和38.70±2.737%(ca1),并且使丝氨酸-396(s396)处的tau磷酸化降低了37.90±3.469%(ac)和40.80±4.528%(ca1)(图12c和图12f)。蛋白质印记(wb)实验证实了s396处的tau磷酸化的免疫组织化学结果,该结果示出,与总tau相比,ac和整个海马体组织中的磷酸化分别降低了33.83±0.20%和43.20±1.50%(图12g、图12h、图12j和图12k)。wb分析指示,海马体中磷酸化的t181tau降低了34.50±1.61%,但在ac中差异不显著(图12i和图12l)。我们的结果一起示出,听觉genus可以降低ad相关的过度磷酸化tau表位的水平,并且听觉genus可以影响tau蛋白病小鼠模型的病变。
组合的听觉和视觉genus引起小神经胶质的簇聚显型反应
由此,我们的发现证明,听觉genus可以降低淀粉样水平并引起皮层感觉区和海马体的神经胶质和脉管系统改变。这促使我们研究组合听觉与视觉genus是否可以引起更深刻的细胞效应。我们首先确定40hz听觉音调刺激与40hz光闪烁的组合是否可以在ac、ca1和mpfc中夹带神经反应。我们以40hz频率向3-8月龄的雄性野生型(c57bl6j)小鼠呈现了1ms长听觉音调和12.5mm长光脉冲的组合,同时随着动物在球形跑台上奔跑或休息而使用32通道硅探头记录ac、ca1和mpfc中的神经活动。尖峰关于每个音调和点亮时段周期性地增大和减小,由此在组合的听觉-视觉刺激期间夹带至40hz(图6a至图6c,左边)。矢量强度分布在40hz听觉-视觉刺激期间显著高于随机刺激或无刺激条件下(图6a至图6c,右边)。因此,ac、ca1和mpfc中的单个神经元的尖峰在听觉-视觉刺激时段期间比基线时段期间显著更多地夹带到40hz。ac、hpc和mpfc中的局部场电位在视听闪烁刺激期间在40hz处显示出提高的功率,然而,作用在mpfc中很小(图13a、图13h和图13o)。由此,40hz音调加光刺激在ac、ca1和mpfc中引起genus。
虽然小,但我们观察到mpfc的听觉和组合听觉-视觉刺激在局部场电位(lfp)反应和单个单位平均激发速率方面的差异。在组合刺激期间,lfp在40hz下的功率有小幅增加,但在仅听觉刺激期间没有该增加(图8n和图6o)。另外,在组合刺激与基线之间的平均激发速率差异的分布具有显著不同于零的中值,而关于仅听觉刺激,该中值与零没有显著不同(图8o和图6u)。
在每天1小时持续7天的组合genus之后,我们检查了ac、vc和ca1中的小神经胶质的形态特征及其与aβ的相互作用(图6d和图6e)。因为已知更高阶的认知区域来处理多模式感觉刺激,所以我们检查了组合的genus是否也可以在内侧前额叶皮层(mpfc)中引起小神经胶质效应。我们发现,与非刺激对照相比,小神经胶质表现出体细胞面积的显著增加,而投影长度显著减小(图6f和图6g)。小神经胶质数量在ac、vc、ca1和mpfc中在组合的genus后也显著增加(图6h)。单独的听觉或视觉刺激组中的小神经胶质(图14a至图14d、图14g和图14h)显示出了在ac、vc和ca1中的减小的投影长度和增大的体细胞面积,但在mpfc中则没有。
与视觉genus相反,听觉genus示出ca1中的小神经胶质计数显著增加;但是单独的听觉或视觉genus都不会引起mpfc中的小神经胶质计数的显著变化(图14e和图14i)。这些发现示出,在genus进行1周后,仅组合的听觉和视觉刺激(而不是单独的听觉或视觉刺激)促进了mpfc中的小神经胶质反应。
有趣的是,组合的genus组中的小神经胶质似乎通过显示围绕淀粉样沉积物的封装作用而示出活性变化。为了更佳地解决簇聚小神经胶质aβ的显型,我们由ac、vc、ca1和mpfc图像创建了三维(3d)渲染,这些图像从组合genus和无刺激对照之后的5xfad脑片获取(图6d和图6e中的“3d重构柱”中示出)。使用imaris成像软件(参见方法),我们创建了淀粉样沉积物(红点)和小神经胶质(绿点)的3d表面,并量化了淀粉样沉积物的25μm半径内的小神经胶质的接近度和数量(最右侧插图,图6d和图6e,在补充视频1和2中提供示例性视频,这些视频示出了组合genus和无刺激对照后的簇聚的小神经胶质aβ显型)。我们观察到,与无刺激对照相比,在组合genus之后,淀粉样斑周围的25μm半径内的小神经胶质数量在ac中显著增加48.88±0.651%,在vc中显著增加31.56±1.11%,在mpfc中显著增加38.64±0.959%(图6i)。我们还观察到ca1中非显著地增加33.05±2.65%。为了检查簇聚小神经胶质aβ显型在组合genus后是否为特定显型,我们分析了仅听觉或视觉genus后在淀粉样沉积物的25μm半径内的小神经胶质数量。我们观察到每个斑块的小神经胶质的数量在genus与无刺激小鼠之间没有显著差异(图14f和图14j)。
接下来,我们解决了在ac、ca1和mpfc中的40hz听觉genus、组合genus、80hz或随机频率刺激7天后,在6月龄5xfad小鼠中的小神经胶质反应的频率特异性。我们观察到,与另外频率和非刺激对照相比,在40hz听觉刺激和组合genus后,在ac和ca1中,小神经胶质细胞体直径和计数显著增加,而且平均齿突长度显著减小。
与40hz听觉刺激、另外频率和非刺激对照相比,仅组合的genus在mpfc中导致小神经胶质反应(图14k至图14m)。这些结果表明,组合的genus通过神经元活性的改变增强了小神经胶质反应。因此,我们得出结论,组合的genus在ac、vc和mpfc中引起延长的小神经胶质簇聚反应。
同时的听觉和视觉genus,而不是单独的听觉或视觉,减少mpfc中的淀粉样负荷
我们在ac、vc、ca1和mpfc中观察到的小神经胶质反应,促使我们研究在1小时的刺激7天后,组合的genus是否也可以改变这些区域的淀粉样水平。使用抗aβ抗体(d54d2)进行的免疫组织化学分析证明,与无刺激对照相比,在组合的genus之后,斑块面积减小(ac中减小56.34±6.35%,vc中减小71.50±6.51%,ca1中减小69.73±6.48%),并且数量减少(ac中减少50.02±3.74%,vc中减少50.60±10.9%,ca1中减少48.80±11.1%)。令人惊讶地,我们的结果证明,与无刺激对照相比,在组合的genus组中,mpfc中的斑块大小减小59.64±8.71%,斑块数量减少了2倍(图7a至图7d)。单独的听觉和视觉genus都无法引起mpfc中淀粉样斑染色的减少,这表明了对组合的genus具有特异性的反应(图14n至图14s)。视觉genus也未示出ca1中斑块尺寸或数量的变化。仅听觉和组合genus治疗均示出,通过ac和hpc中的aβ-elisa测量的可溶性aβ1-42和不溶性aβ1-42水平降低,而组合的随机闪烁、8hz或80hz刺激对ac或hpc中的淀粉样水平没有显著影响(图14t和图14u)。
接下来,我们以各种频率的感官刺激对6月龄的5xfad小鼠进行了处理,以便确定mpfc中的aβ的降低是否特定于刺激的类型(仅听觉相对于组合)或频率。使用aβ-elisa来测量mpfc中淀粉样水平的变化,我们观察到可溶性或不溶性aβ1-42在组合的8hz、40hz听觉刺激、组合随机频率刺激或无刺激组之间无显著差异。相反,与无刺激组相比,组合的genus组示出mpfc中的可溶性aβ1-42减少了59.58±7.26%,并且不溶性aβ1-42减少了34.17±8.20%(图7e和图7f)。
此外,我们经由免疫组织化学分析测量了斑块负荷,该分析在40hz听觉genus、组合genus、80hz或随机频率7天后的6月龄的5xfad小鼠中使用β-淀粉样特异性抗体(cellsignalingtechnology;d54d2)进行。我们观察到,在40hz听觉刺激和组合genus后,ac和ca1中的平均斑块数量显著减少,然而,与另外频率对照和非刺激对照相比,仅组合的genus导致mpfc中的斑块数量显著减少(图7c和图7d)。使用aβ1-42抗体对aβ1-42特异性免疫染色的分析指示,在40hz听觉刺激和组合genus后,ac和ca1中的aβ显著降低,然而,仅组合的genus导致mpfc中的免疫染色强度显著降低(图14v)。
mpfc中淀粉样负荷的降低表明组合的genus影响更广泛的皮层区域。为了确定组合genus对整个皮层中的淀粉样斑丰度的总体影响,我们在组合genus1周后的6月龄5xfad小鼠中执行了全脑shield处理(方法),并对淀粉样斑进行了免疫染色(使用d54d2抗体)(图7g和图7h)。使用光片显微镜来3d地分析斑块,我们发现与非刺激对照相比,新皮层中的总斑块体积和数量分别减少了37%和34%(图7i和图7j,在补充视频3和4中提供示例性视频,这些视频示出了组合genus和无刺激对照后对于斑块免疫染色的3d全脑shield样品)。这些结果一起指示,组合的genus显著降低跨5xfad小鼠模型新皮层的淀粉样斑负荷。
方法
动物
所有动物工作均由麻省理工学院比较医学系动物护理委员会和乔治亚理工学院机构动物护理与使用委员会批准。在标准的12小时明亮/12小时黑暗循环中,将小鼠分成不超过五个的组来安置;所有实验均在明亮循环期间执行。在乔治亚理工学院执行电生理学实验,从杰克逊实验室获得雄性(1-3月龄)wt小鼠(c57bl/6)。将小鼠安置在相反的12小时明亮/12小时黑暗循环中,所有实验均在黑暗循环期间执行。无限制地提供食物和水。
外科手术
所有外科手术均如iaccarino和singer等人(2016年)所述地执行。简而言之,在头板放置手术前,用异氟烷麻醉成年(2-3月龄)小鼠。使用牙粘固粉(c&bmetabond,parkell)固定定制的不锈钢头板,并在颅骨上标记用于lfp记录的目标开颅术部位(以mm为单位,从前卤:-2.0前/后,+/-1.8中/侧,用于靶向ca1,-2.0至-3.0前/后,+/-1.8中/侧,用于靶向听觉皮层,以及+1.3至+1.4前/后,+/-1.0中/侧,用于靶向前额叶皮层)。随后在3-8月龄小鼠中执行开颅术。在第一记录阶段之前,通过用牙钻减薄颅骨然后用27号针形成孔来进行开颅术(200-500μm直径)。不记录时,用无菌硅弹性体(kwik-silwpi)密封开颅术。
电生理记录
在记录期间,头部固定的动物在气浮8英寸球形跑台上奔跑。所有动物之前都已经学习在跑台上操纵,直到它们舒适为止,同时偶尔接收加糖炼乳(1:2水稀释)。动物在球上最多5小时,并且在此期间具有多个奔跑和休息的时段。将单柄32通道探头(neuronexus)推进到目标位置。记录位点跨度为250m。对于听觉皮层记录,将探头从平行于冠状平面的垂直方向以45°角推进到3-4.15mm的深度。呈现一系列5、10、15和20khz的50ms音调,以检测平均lfp中的听觉响应。对于ca1记录,探头通过开颅术垂直前进至1.14–2.05mm的深度,直到观察到海马体锥体层电生理学特性(大θ波和锐波波纹,多个通道上的150+μv尖峰)为止。对于前额叶皮层记录,将探头从垂直方向以20°角、从冠状平面以49°角推进到1.48-2.15mm的深度。如果在相同的记录阶段期间在多个深度收集数据;标出新的深度,以便确保记录位点的位置保持在目标位置中(对于ac,n=9个记录深度,来自5只小鼠中的9个阶段,对于ca1,12个记录深度,来自5只小鼠中的10个阶段,对于mpcf,n=7个记录深度,来自4只小鼠中的7个阶段)。使用intanrhd2000评估系统以20khz的采样速率获取数据,该系统使用磨碎的丸作为参考。
电生理记录的听觉和视觉刺激
向动物呈现与10s基线时段交错的10s刺激块。刺激块在20hz、40hz、80hz的仅听觉或听觉和视觉刺激或随机刺激之间轮换(以随机化脉冲间间隔递送脉冲,这些间隔由平均间隔为25ms的均匀分布确定)。交错刺激块,以确保观察到的结果不是由于神经元反应随时间变化。所有听觉脉冲均为1ms长的10khz音调。所有视觉脉冲均为50%占空比的刺激频率(长度为25ms、12.5ms或6.25ms)。对于组合刺激,将听觉和视觉脉冲与每个脉冲的开始对齐。
数据获取
使用intanrhd2000评估系统以20khz的采样速率获取数据。
峰值检测
原始迹线在300-6,000hz之间进行带通滤波。然后通过滤波信号的中值阈值加上估计的标准偏差的五倍(中值/0.675)来检测尖峰。
尖峰排序和单个单位的稳定性
使用mountainsort自动化尖峰排序进行尖峰检测和排序,然后基于波形和互相关图的视觉检查进行手动管理。在手动管理之前,应用质量阈值,以仅包括峰值snr大于或等于1、与噪声重叠小于10%、针对其他单位的隔离大于95%的单位,这产生良好分离的单个单位。为了考虑期间丢失单个单位的记录的不稳定时段,应用稳定准则,使得在分析中将仅考虑稳定时段(不会突然丢失单个单位的激发速率)。在记录期的过程期间,计算每个单位的激活速率(fr)。使用k均值聚类将激发速率聚类为两个分布,即,低fr和高fr。对于fr降低至高fr平均值的10%以下的单位,进一步的分析识别了稳定的记录时段,该时段被定义为fr比低fr平均值高2个标准偏差的最长时间长度。
局部场电位
lfp通过将原始迹线下采样至2khz并在1至300hz之间进行带通滤波获得。
用于分析的记录位点
在多个通道上分析ac和ca1中的数据。在ac中,利用跨越375m的32个通道中的下16个通道,因为探头上的最低通道用于确定ac的位置,并且确定最高的16个通道不在主感兴趣区域中。对于ca1,分析探头上跨越250m的所有起作用的通道(27/32或31/32)。ac和ca1两者中的探头上的最高通道用作功率谱分析的探头参考。使用地面作为参考获得了类似的结果。
前额叶皮层组织学
在每只动物的最终mpfc记录期间,用dii涂布探头并将其插入目标深度。在麻醉(异氟醚)下用磷酸缓冲盐水(pbs)中的4%多聚甲醛对小鼠进行心内灌注,然后将大脑在1xpbs中的4%多聚甲醛中进行固定后过夜。用leicavt1000s振动切片机(leica)将大脑切成100μm厚的片。将切片用1xpbs中的0.2%1mmoldapi染色,并用vectashield安装介质将切片安装到显微镜载玻片上。使用伴随的zenblue2软件在zeissaxioobserverz1倒置落射荧光显微镜上获取图像。
功率谱
使用来自chronux工具箱的多锥度法执行功率谱密度分析(时间带宽乘积=3,锥度=5)。lfp迹线被分为每个刺激条件的10s试验。参照头骨上方盐水中的研磨丸,在这些试验中对于每只动物计算平均功率谱密度(在相同的记录日和记录深度内)。最初针对ac、ca1和mpfc中的所有记录位点计算了功率谱密度分析。从每个记录深度,分析中都包括响应于40hz闪烁刺激而具有最大40hz峰值的迹线。在呈现的数据中显示的每深度迹线响应听觉闪烁刺激具有最大的40hz峰值。
闪烁刺激期间的激发
每个刺激频率的单个单位周围刺激时间直方图(psth)包含四个刺激循环
平均激发速率
对于每种刺激条件对于每个单个单位平均激发速率。仅每个单位的稳定时段有助于平均fr计算(参见尖峰排序和单个单位稳定性,上方)。通过取得每个单位在每种条件下的平均fr差异,每个单位内计算出刺激条件之间的平均发动速率差异。
40hz视觉闪烁刺激方案
为了进行生物化学和免疫组织化学分析,将5xfad小鼠置于在由发光二极管(led)灯泡照亮的暗室中,并将其暴露于以下四种刺激条件之一:暗,8hz,40hz(点亮12.5ms,关灯12.5ms,60w)或随机(以平均值为25ms的均匀分布确定的随机间隔递送光脉冲)刺激1小时持续七天。
40hz听觉音调刺激方案
为了进行生化、免疫组织化学或行为分析,将5xfad、app/ps1或p301s小鼠置于昏暗的室内,房间内装有隔音泡沫(mcmaster-carr,5692t49)。将扬声器(ayl,ac-48073)在室上方放置在小鼠够不着的位置。将小鼠暴露于五种刺激条件之一:没有音调、8hz音调、40hz音调、80hz音调或随机递送的音调(以通过平均25ms的均匀分布确定的随机间隔递送听觉音调)刺激。刺激条件的音调由持续时间为1ms并且以60db递送的10khz音调构成。对于电生理学记录,在放置探头后,关闭房间中的灯,并且向动物呈现与10s无光或音调时段交错的交替的10s仅音频和视听刺激时段。对于纯音频刺激,以4%的占空比以40hz播放10khz的音调。对于视听刺激,音频刺激伴随着以50hz占空比以40hz闪烁10s时段的周围光。以这种方式呈现刺激,持续20分钟,在阶段之间停顿1-10分钟,以检查动物的行为。
同时40hz听觉和视觉刺激方案
对于生物化学、免疫组织化学或行为分析,将5xfad小鼠置于由led灯泡照射的暗室中,并同时暴露于听觉音调序列。小鼠暴露于四种刺激之一:黑暗/安静、40hz光闪烁、40hz听觉音调序列、同时的40hz光闪烁和听觉音调、或随机光闪烁/音调刺激。
免疫组织化学
在麻醉(2:1的开他敏/安耐宁)下用磷酸缓冲盐水(pbs)中的4%多聚甲醛对小鼠进行心内灌注,然后将大脑在pbs中的4%多聚甲醛中进行固定后过夜。用leicavt1000s振动切片机(leica)将大脑切成40μm厚的片。切片在含有0.3%tritonx-100和10%驴血清的pbs中在室温下渗透和封闭2小时。切片在含有0.3%tritonx-100和10%驴血清的pbs的第一抗体中于4℃温育过夜。第一抗体是:抗β-淀粉样(cellsignalingtechnology;d54d2)、抗iba1(wakochemicals;019-19741)、抗神经胶质纤维酸性蛋白(gfap)(abcam;ab4674)、抗s100b(abcam;ab868)、抗lrp1(abcam;28320)、dylight488标记的番茄(番茄)凝集素(vector实验室;dl-1174)、抗淀粉样低聚物(milliporesigma;ab9234)、抗磷酸化tau(ser396)(cellsignalingtechnology;9632)、抗磷酸-tau(thr181)(cellsignalingtechnology,12885)、hoechst33342(thermofisherscientific;h3570)。使用抗aβ抗体12f4是因为它不与app反应,这允许我们确定我们的标记是否对aβ特异,而且允许与iba1共标记。抗淀粉样低聚物抗体ab9234用于与lrp1共标记。第二天,在室温下将脑片与荧光偶联的二抗(jacksonimmunoresearch)孵育2小时,并用hoechst33342(invitrogen)对细胞核染色。使用具有40倍物镜的共焦显微镜(lsm710;zeiss)在所有条件下以相同设置获取图像。由对治疗组不知情的实验者使用imagej1.42q对图像进行量化。对于每个实验条件,来自每个动物的两个冠状切片用于量化。比例尺为50μm,除非在图例中另有说明。imagej用于测量iba1+细胞体的直径并为了长度测量而跟踪齿突。另外,coloc2插件用于测量iba1和aβ的共定位。使用imarisx648.1.2(bitplane,瑞士苏黎世)量化小神经胶质齿突分枝。imagej中的“分析颗粒”功能用于计数斑块数量和面积、包括至少10μm的沉积物,并且将设定的阈值用于对照组和实验组这两者。
脉管系统-aβ共定位分析
imariscoloc模块用于量化3d中两个独立的源通道(即,凝集素和ab、凝集素和lrp1)之间的信号共定位。这些源通道被阈值化以掩蔽来自噪声或背景信号的任何强度。然后imariscoloc生成一个新的通道,该通道仅包含共定位在源通道的阈值组内的体素,并呈现关联的统计分析。
小神经胶质aβ簇聚分析
使用imaris来分析40um切片中淀粉样斑周围的小神经胶质簇聚模式。使用表面模块来基于12f4信号检测并3d渲染斑块(红色)。然后使用点模块对iba1阳性小神经胶质计数,这在每个细胞的体细胞(绿色)上放置一个球体。最后,运行接近表面xtension的点以寻找比定义的25μm阈值更接近表面物体的点的子集,并且排除落在该范围之外的点。该算法测量3d空间中从点中心到表面物体最近点的距离,这允许量化斑块附近的小神经胶质聚集。
脑片中的clarity免疫染色
小鼠用冰冷的pbs(1x)灌注,然后用冰冷的1xpbs中的4%pfa、1%戊二醛灌注。解剖出脑,在4℃下在4%pfa/1%戊二醛溶液中后固定72小时。通过在灭活溶液(1×pbs中的4%丙烯酰胺、1m甘氨酸、0.1%triton-x100)中在rt下孵育大脑48小时来终止固定。用1xpbs洗涤后,在1xpbs中在振动切片机(leicavt100s)上将大脑切成100um冠状切片。参照allen小鼠脑图谱,选择含有感兴趣区域(即,听觉皮层和海马体)的切片,并在透明缓冲液(ph8.5-9.0,200mm十二烷基硫酸钠,20mm氢氧化锂一水合物,4mm硼酸的ddh2o溶液)中孵育2-4小时,在55℃摇动,将清洁切片在1xpbst(0.1%triton-x100/1xpbs)中洗涤3×15分钟,并在rt下放入封闭溶液(2%牛血清白蛋白/1xpbst)中过夜。随后,在1xpbst中执行三次1小时洗涤,即,在rt下摇动。切片在弱结合缓冲液(ph8.5-9.0,37.75mmna2hpo4,3.53mmkh2po4,0.02%叠氮化钠的pbst溶液)中在rt下孵育1小时,然后转移至第一抗体,在37℃下在12小时内在1x弱结合缓冲液中稀释至1:100,然后在6小时内以每小时等份加入反向缓冲液(ph7.4、37.75mmna2hpo4、3.53mmkh2po4的0.02%叠氮化钠的pbst溶液),以等于第一抗体溶液的体积加上组织的体积。在将切片于hoechst33258(1:250)(sigma-aldrich,94403)和第二抗体(1:100)的1xpbs混合物中rt孵育12小时之前,进行另一组在1xpbst中3×1小时的洗涤。然后在1xpbs中洗涤切片过夜,并在rims(折射率匹配溶液:75ghistodenz、20ml0.1m磷酸盐缓冲液、60mlddh2o)中rt孵育1小时,然后安装。将脑切片用rims中的盖玻片(vwrvistavision、vwrinternational、llc、radnor、pa)安装到显微镜载玻片上。
在zeisslsm880显微镜上获得图像,该显微镜附有zenblack2.1软件(carlzeiss显微镜,德国耶拿)。以0.4-0.5μm的步长、4.1ms的像素停留、2的平均、适于3d重建的1024×1024的分辨率获取z叠堆图像。imarisx648.3.1(bitplane,瑞士苏黎世)用于3-d渲染和分析。
整个小鼠大脑的处理和清除
根据shield方案处理5xfad小鼠大脑。简而言之,先对5xfad小鼠进行心内灌注冰冷的pbs,然后再灌注20ml含4%pfa的shield-off溶液。解剖大脑并在4c下在相同溶液中后固定24小时。然后将大脑在4c下在没有pfa的shield-off溶液中于孵育过夜。然后将大脑在37c下在shield-on溶液中孵育24小时。固定后,将大脑在含有200mm十二烷基硫酸钠(sds)、20mm一水氢氧化锂、40mm硼酸、ph8.5-9.0的透明水溶液中孵育。然后使用smartclearpro(lifecanvastechnologies,马萨诸塞州剑桥)基于随机电传输(kim等人,pnas,2015年)清理大脑几天,直到透明为止。
对清除的整个半球进行免疫染色
使用etango(即,一种修改的随机电转移方法)在2天内用15ul与alexafluor-488(cst,#51374)偶联的β-淀粉样抗体对清除的半球进行染色(kim等人,pnas,2015年)。
光片显微镜
用hprotos(3g泛影酸、5gn-甲基-d-谷氨酸胺、125g碘海醇的105ml去离子水溶液)孵育免疫染色样品,以进行光学澄清,然后在hprotos中使用2%低温熔化琼脂糖将其固定在丙烯酸保持器上。使用装配有10倍clarity优化的物镜的定制光片显微镜用于将488通道用于β-淀粉样可视化和将647通道用于自发荧光来对整个半球体成像。
清晰的全脑图像处理,斑块检测和地图集对齐
使用cidre(即,在matlab中实施的开源软件包)对获取的图像数据进行照明校正,然后在imaristm
蛋白质印记
解剖海马体和听觉皮层,并从6月龄雄性5xfad中备裂解物。用手动匀浆器(sigma)在1mlripa(50mmtrishclph8.0、150mmnacl,1%np-40、0.5%脱氧胆酸钠,0.1%sds)缓冲液中匀浆组织,在冰上孵育15分钟,并在4℃下旋转30分钟。分离细胞碎片并以在14,000r.p.m下离心10分钟来丢弃。使用nanodrop量化分析裂解物,然后将25μg的蛋白质加载在10%的丙烯酰胺凝胶上。在100v下将蛋白质从丙烯酰胺凝胶转移至pvdf膜(invitrogen)120分钟。使用在tbs:tween中稀释的牛血清白蛋白(5%w/v)堵塞膜。将膜在第一抗体中于4℃孵育过夜,在第二抗体于室温下孵育90分钟。第一抗体是抗磷酸化tau(ser396)和抗磷酸化tau(thr181)。第二抗体是li-corirdye第二抗体。使用imagej1.46a量化信号强度,并将信号强度归一化为总tautau5的值(thermofisherscientific;ahb0042)。
elisa
从6月龄的5xfad雄性分离出初生听觉皮层、内侧前额叶皮层和海马体,并根据制造商的说明使用aβ42或aβ40elisa试剂盒(invitrogen)进行aβ测量。elisa测量前,用5m胍/50mmtrishcl(ph8.0)缓冲液处理不溶性aβ。
行为实验
新物体识别
如先前所述(leger等人,2013年),新物体识别(nor)任务包括习惯化阶段,然后在第二天执行训练和测试。训练前24小时,使小鼠习惯于开放的测试活动场所(40cm长x40cm宽x35cm高),持续5分钟,其间计算总距离(cm)、在中心中的时间(s)和速度(cm/s)(tse系统)。在训练期间,将小鼠放置于在同一个盒子中,两个相同的物体放在相对的角落。允许小鼠总共进行20秒的物体互动时间(最大时间范围为10分钟),然后立即将其从活动场所移开。1小时后,在训练期间使用相同的程序测试物体记忆,除了一个物体被其位置的一个新物体代替。当口鼻部接触到任一物体时,会记录物体探索,并通过识别指标ri=t新/(t新+t熟悉)计算,其中t新和t熟悉分别指示关于新物体和熟悉物体花费的时间。
新物体定位
使用与物体识别任务相同的过程执行新位置识别(nol)任务,除了将两个相同的物体用于训练和测试这两者,并且在测试期间将一个物体位移到新位置。
morris水迷宫测试
在填充有约22℃的白色不透明水的圆形箱(直径,1.2m)中执行空间参考记忆测试。沿着水箱周围的墙壁放置了由不同颜色和形状构成的参考线索。在水箱内是位于目标象限的固定平台(直径,10cm)。在测试期间,平台被淹没,小鼠在随机面向箱的墙壁的七个点中的一个处被放置到箱中。向小鼠提供60秒来搜索平台,如果没有找到平台,则将小鼠轻轻地引导到平台。将动物保持在平台上15s。每天进行两次试验,试验间间隔1小时。在试验之间,将小鼠轻轻地垫干并在加热垫上加温。使用tse系统对小鼠行为进行视频记录。每次试验对逃避潜伏期或小鼠到达平台所花费的时间进行评分,并在每个测试日进行平均。在第6天,去除平台并执行了记忆测试(探头测试)。记录4个象限中每个象限所花费的时间以及平台被使用过的区域的穿越次数。自动记录游泳速度。
结论
本公开的发明方面针对本文所述的每个单独的特征、系统、制品、材料、药盒和/或方法。另外,两个或更多个此类特征、系统、物品、材料、套件和/或方法(如果此类特征、系统、物品、材料、套件和/或方法并非互不一致)的任何组合包含在本公开的发明性范围内。
而且,各种发明概念可以体现为一种或多种方法,已经提供了其实例。作为该方法的一部分执行的动作可以以任何合适的方式排序。因此,可以构造按不同于所说明的次序执行动作的实施例,其可以包含同时执行一些动作,即使在说明性实施例中展示为依序的动作也是如此。
本文提及的所有出版物、专利申请、专利和其他参考文献通过引用整体并入本文。
如本文所定义和使用的所有定义应被理解为控制词典定义,通过引用并入的文档中的定义和/或所定义术语的普通含义。
除非明确指出相反的含义,否则本文中在说明书和权利要求书中使用的不定冠词“一个”和“一种”应理解为表示“至少一个”。
在说明书和权利要求书中使用的短语“和/或”应当理解为表示这样结合的元件中的“任一者或两者”,即在一些情况下共同存在而在其他情况下不连续存在的元件。用“和/或”列出的多个元件应以相同的方式解释,即,这样结合的元件中的“一者或多者”。除了由“和/或”子句具体指出的元件之外,还可以任选地存在其他元件,无论与具体指出的那些元件相关还是无关。因此,作为非限制性实例,当结合开放式语言(例如“包括”)使用时,提及“a和/或b”在一个实施例中可以仅指a(任选地包含除b之外的元素);在另一个实施例中仅指b(任选地包含除a之外的元素);在又一个实施例中,兼指a和b(任选地包含其他元素);等等。
如本文在说明书和权利要求书中所使用的,“或”应被理解为具有与如上文所定义的“和/或”相同的含义。例如,当将列表中的项目分开时,“或”或“和/或”应解释为包括的,即包括多个元件或元件列表中的至少一个(还包括一个以上)以及任选地其他未列出的项目。仅明确指出相反的术语,诸如“仅一个”或“恰好一个”,或当在权利要求书中使用时,“由…组成”将参见例如包括多个元件或元件列表中的恰好一个元件。一般地,本文所使用的术语“或”仅应解释为在排他性术语诸如“任一个”、“一个”、“仅一个”或“恰好一个”之前指示排他的替代方式(即“一者或另一者,但并非两者”)。当在权利要求书中使用时,“基本上由…组成”应具有其在专利法领域中所使用的普通含义。
如本文在说明书和权利要求书中所使用的,在提及一个或多个元件的列表时,短语“至少一个”应理解为指选自该元件列表中的任何一个或多个元件的至少一个元件,但不一定包括元件列表中具体列出的每个元件中的至少一个,并且不排除元件列表中的元件的任何组合。除了短语“至少一个”所指的元件列表中具体指出的元件之外,该定义还允许任选地存在其他元件,无论与具体指出的那些元件相关还是无关。因此,作为非限制性实例,在一个实施例中,“a和b中的至少一个”(或,等效地,“a或b中的至少一个”或,等效地“a和/或b中的至少一个”)可以指代至少一个,任选地包含多于一个a,不存在b(并且任选地包含除了b之外的元素);在另一个实施例中,可以指代至少一个,任选地包含多于一个b,不存在a(并且任选地包含除了a之外的元素);在又一实施例中,可以指代至少一个,任选地包含多于一个a,和至少一个,任选地包含多于一个b(并且任选地包含其他元素);等等。
在权利要求书以及以上说明书中,所有过渡短语,诸如“包含”、“包括”、“带有”、“具有”、“涵盖”、“涉及”、“容纳”、“由…组成”等都应理解为开放式的,即指包括但不限于。如美国专利局专利审查程序手册第2111.03节所述,只有过渡短语“由…组成”和“基本上由…组成”才应分别是封闭的或半封闭的过渡短语。
- 还没有人留言评论。精彩留言会获得点赞!