CH3结构域表位标签的制作方法
本发明的领域涉及异源抗体表位在促进生物样品中基于抗体的生物制品的检测中的用途。
背景技术:
基于抗体的生物制品,例如治疗性抗体和fc融合蛋白,通常在人免疫球蛋白g(igg)支架上开发,以最小化在其施用后对生物制品的不期望的受体介导的免疫应答。然而,因为人天然产生全身循环的igg,所以由于背景存在内源性人igg,施用的生物制品的igg支架环境使得难以检测患者样品内的生物制品。具有检测患者样品中的生物制品的能力是重要的,因为用于追踪生物制品的血清水平和药代动力学(“pk”)行为的测定常规可用于优化生物制品的给药。
执业医师通常依赖于抗独特型单克隆抗体来检测基于抗体的生物制品的独一无二的fab表位独特型,以针对内源性igg背景检测它们。然而,每种抗独特型单克隆抗体的开发是时间和资源密集型的,因为每一个基于抗体的生物制品需要其自身的检测抗体。
或者,可以通过将一个或多个非天然存在的表位掺入衍生自人iggfc区的ch3支架的ab、ef或cd环,继而将其掺入基于抗体的生物制品中消除产生抗独特型抗体的需要。
技术实现要素:
本发明涉及将异源抗体表位包含到基于抗体的生物制品中,以促进针对内源性抗体的背景(通常在患者样品的环境下)的生物制品检测。更具体地,将异源抗体表位掺入igg1衍生的ch3支架的ab、ef或cd结构环中的一个或多个,继而将其掺入基于抗体的生物制品中。本质上,根据本发明的ch3支架的一个或多个异源表位用作“表位标签”,以使得能够快速识别包括经表位标记的ch3结构域的蛋白或蛋白复合物。
此外,相同的表位可以被掺入到实际上无限数量的不同的基于抗体的生物制品中。因此,根据本发明的快速生物制品检测系统可以推广用于不同的生物制品,而用于检测样品中生物制品的常规方法依赖于对每个不同的生物制品单独使用不同的抗独特型抗体。
附图说明
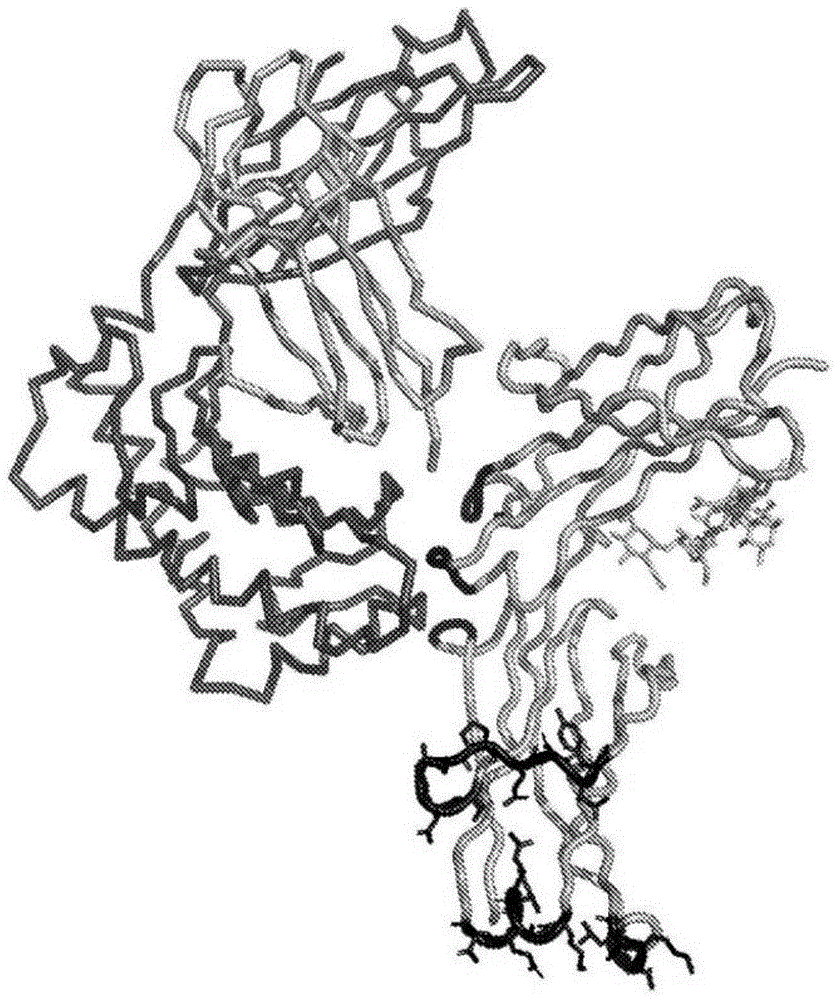
图1描绘了新生fc受体(“fcrn”)、人血清白蛋白(hsa)和fc区之间的复合物的晶体结构,如pdb4n0u所示,并使用pymol分子建模软件可视化。晶体结构的链a是igg受体fcrn大亚基p51(以深灰色带描绘)。晶体结构的链b是β2微球蛋白亚基(以浅灰色带描绘)。未描绘链d(hsa)。链e是igg1fc区同二聚体的一个亚基(浅灰色卡通,其中强调的区域为黑色)。以黑色棒强调的残基代表cd和ab/ef环。以黑色卡通描绘没有棒状物的fc残基在fc:fcrn界面的
图2a显示了各种人ig-折叠结构域蛋白的氨基酸序列的多序列比对,其晶体结构在结构生物信息学蛋白数据库(“pdb”)研究合作实验室中是可获得的,pdb4wi2:a的氨基酸104-108对应于人igg1ch3结构域序列的区域。使用序列比对软件程序mafft进行比对。
图2b显示了各种人ig-折叠结构域蛋白的氨基酸序列的多序列比对,其晶体结构在结构生物信息学蛋白数据库(“pdb”)研究合作实验室中是可获得的,pdb4wi2:a的氨基酸109-202对应于人igg1ch3结构域序列的区域。使用序列比对软件程序mafft进行比对。
图2c显示了各种人ig-折叠结构域蛋白的氨基酸序列的多序列比对,其晶体结构在结构生物信息学蛋白数据库(“pdb”)研究合作实验室中是可获得的,pdb4wi2:a的氨基酸203-208对应于人igg1ch3结构域序列的区域。使用序列比对软件程序mafft进行比对。
图3显示了代表来自图2中描绘的那些的ig-折叠结构域蛋白的选择组的氨基酸序列与衍生自pdb4wi2的人igg1ch3结构域序列的比对。
图4描绘了使用pymol分子建模软件以灰色卡通可视化的衍生自pdb4wi2序列的野生型人fc区的晶体结构。以灰色棒描绘碳水化合物。ab、cd和ef环以黑色描绘。ab和ef环内的表面暴露的侧链用棒表示。侧链的表面暴露预示着它们与抗体接触的潜力。
图5描绘了使用pymol分子建模软件以灰色卡通可视化的衍生自pdb4wi2序列的野生型人fc区的晶体结构。以灰色棒描绘碳水化合物。ab、cd和ef环以黑色描绘。cd环内的表面暴露的侧链用棒表示。侧链的表面暴露预示着它们与抗体接触的潜力。
图6描绘了人fc区的ab-ef环区的表面。ab-ef环中的表面残基被描绘为球。pdb结构4wi2使用pymol软件包可视化。
图7显示了ab和ef环中的模型化表面残基。将突变后掺入seqidno.38和seqidno.67的pdb#4wi2的pymol分别显示在ab和ef环中,以产生表位标签。
图8是免疫印迹(westernblot)的扫描图像,其评估抗glu抗体检测具有野生型或标记的(cd-glu或cd-4i2x)fc结构域的人抗体的能力。
图9描绘了在存在或不存在各种比例(微克(mcg)/ml:微克/ml)的cd-wt抗体的情况下基于elisa的cd-glu检测。
图10描绘了来自竞争性结合fret实验的数据,其评价了含有一系列不同标签的抗体竞争性抑制具有野生型(m1,17)fc的抗体与fcrn结合的能力。
图11描绘了来自竞争性fret测定的数据,其评价了含有一系列不同标签的抗体竞争性抑制具有野生型(m1,17)fc的抗体与cd16a结合的能力。
图12是m1,17和cd-glu对一系列fc受体的基于spr的亲和力测量。
图13绘制了与fcγri结合的cd-wt(深灰色)和cd-glu(浅灰色)的结合率(on-rates)(ka,1/ms)与解离率(off-rates)(kd,1/s)。数据表明,相对于cd-wt,cd-glu的解离率小幅(1.2倍)增长。
具体实施方式
本发明涉及与一种或多种异源抗体表位掺入免疫球蛋白fc结构的恒定重链结构域3(“ch3结构域”)或其含ch3的片段有关的组合物和方法。例如,本发明可以将异源表位掺入人igg抗体的ch3结构域。根据本发明的这种掺入ch3结构域的表位可以用作“表位标签”,以允许快速识别包括表位标记的ch3结构域的蛋白或蛋白复合物。通常,根据本发明的ch3结构域表位标签的氨基酸序列也衍生自或修饰自ch3结构域,并保留了ch3结构域的基本三级结构,并且因此在本文中也称为“ch3支架”。换言之,ch3结构域表位标签存在于“ch3支架”的环境中。尽管有衍生自任何具有ch3结构域的免疫球蛋白分子诸如人igg2、igg3或igg4的ch3支架,但是衍生自人igg1分子的ch3支架是根据本发明的ch3支架的优选结构背景。同样地,根据本发明的ch3支架也可以衍生自或修饰自非免疫球蛋白的结构域,其具有相对于iggch3结构域至少部分保守的三级结构。
实际上,根据本发明的ch3支架基本上保留了天然存在的ch3结构域的结构特征,称为免疫球蛋白折叠(ig-折叠),包含两个天然存在的ch3结构域的β片层(sheet)的堆积,即,含有反平行β链c、f和g的3链β片层,与以反平行方向排列的含有β链a、b、d和e的4链β片层相对堆积。涉及维持β片层的堆积的氨基酸残基在本领域中是已知的,包括形成氢键、疏水作用和二硫键的残基。在特定的实施方式中,保持ig-折叠至关重要的残基未被修饰。在某些实施方式中,基本上不修饰框架残基;例如,与野生型ch3结构域相比,在工程化的ch3支架中修饰不超过15%或10%或5%的框架残基。与对其他区域的修饰相比,在连接天然ch3的β片层(例如ab环)的两个β链或两个不同片层(例如cd或ef环)的链的环处的修饰或环附近的修饰可能更具耐受性(即不太可能破坏天然ch3的结构或构象)。在免疫球蛋白重链(“ihc”)的情况下,ch3支架保留野生型ch3分子的fcrn结合结构。例如,据信对于fcrn结合功能至关重要的残基。
已工程化以含有根据本发明的表位标记的ch3支架的蛋白通常是治疗性抗体(适用于向受试者施用以治疗或预防疾病或病症的抗体)和fc连接的治疗性产品,例如fc-融合蛋白和fc-连接的药物或治疗剂,它们可以统称为fc-生物制品。
根据本发明的抗体优选是单克隆抗体。单克隆抗体通常理解为是由单个克隆的b淋巴细胞群体、克隆的杂交瘤细胞群体或已转染单个抗体的基因或其部分的细胞的克隆群体产生。单克隆抗体通过本领域技术人员已知的方法产生,例如通过由骨髓瘤细胞与免疫淋巴细胞的融合制备杂交抗体形成细胞。
根据本发明的单克隆抗体还包含人源化单克隆抗体。更具体地,根据本发明的“人”抗体,也称为“全人”抗体,是包含人框架区和来自人免疫球蛋白的cdr的抗体。例如,抗体的框架和cdr来自相同的起源人重链,或人轻链氨基酸序列,或两者。或者,框架区可来源于一种人抗体,并被工程化以包含来自不同人抗体的cdr。
根据本发明的表位标记的抗体可以是单特异性的、双特异性的、三特异性的或具有更大的多特异性。多特异性抗体可以是对多肽的不同表位特异的,或者可以是对异源表位特异的,例如异源多肽或固体支持物材料。参见例如pct公开wo93/17715;wo92/08802;wo91/00360;wo92/05793;美国专利号4,474,893;4,714,681;4,925,648;5,573,920;5,601,819。
可以被工程化以包含根据本发明的ch3支架的治疗性抗体的实例包含但不限于:靶向cd20的嵌合小鼠/人igg1(例如,利妥昔单抗);靶向her2的人源化igg1(例如曲妥珠单抗(trastuzumab));在b淋巴细胞和t淋巴细胞上靶向cd52的人源化igg1(例如阿仑单抗);靶向rankl的人igg2(例如地诺单抗(denosumab));靶向α-4整联蛋白的人源化igg4(例如那他珠单抗);靶向egfr、erbb-1和her1的人igg2(例如帕尼单抗);靶向补体蛋白c5的人源化igg2/4κ(例如依库丽单抗);靶向egfr、erbb-1和her1的嵌合小鼠/人igg1(例如西妥昔单抗);靶向vegf的人源化igg1(例如贝伐单抗);靶向tnf-α的人igg1(例如阿达木单抗);和靶向tnf-α的嵌合小鼠/人igg1(例如英利昔单抗)。
与本发明的表位标记的抗体一样,根据本发明的fc融合蛋白的fc区也优选衍生自人免疫球蛋白g(“igg”)类框架,并且更特别是igg1亚类。fc融合蛋白可以是单体蛋白或多聚体蛋白,诸如二聚体蛋白或四聚体蛋白,其可以经由其fc区通过多聚化形成。fc区提供fc融合蛋白的pk行为。
通常,fc融合蛋白是生物工程化的多肽,将抗体的可结晶片段(fc)区与另一具有生物活性的蛋白结构域或肽连接在一起,以产生具有独特结构功能特性和治疗潜力的分子。fc融合蛋白(其中fc区与受体的天然形式的胞外结构域融合)可以充当配体的捕集器(trap)。可以被工程化以包含根据本发明的ch3支架的fc融合蛋白的实例包含但不限于:与人igg1的fc区融合的ctla-4(例如贝拉西普);与人igg1的fc区融合的vegfr1/vegfr2(例如阿柏西普);与人igg1的fc区融合的il-1r(例如利纳西普);与人igg1的fc区融合的促血小板生成素结合肽(例如罗米司亭(romiplostim));与人igg1的fc区融合的突变ctla-4(例如阿巴西普);与人igg1的fc区融合的lfa-3(例如阿法西普(alefacept));和与人igg1的fc区融合的tnfr(例如依那西普)。
根据本发明的用于检测表位标记的ch3支架的抗体和包括表位标记的ch3支架的蛋白在本文中通常称为“检测抗体”。如本文所用,术语“特异性结合(specificallybinds)”或“特异性结合(specificbinding)”是指在异源分子群体中决定目标同源配体的结合反应。因此,在指定条件(例如免疫测定条件)下,根据本发明的检测抗体结合其特定靶标,例如根据本发明的ch3表位标签,且不以显著量结合样品中存在的其它分子,诸如患者样品中的内源性抗体。特异性结合是指结合就对其靶标的亲和力而言是选择性的,并且通常在结合常数或结合动力学相差至少10倍,优选相差至少100倍,并且更优选相差至少1000倍时实现。
术语“表位”是指通常由氨基酸序列形成的结构,其能够被抗体结构特异性结合,包括天然存在的和单克隆抗体,以及此类分子的片段。换言之,表位可以是分子结构,其可以完全构成抗体或其片段的结合结构域的特异性结合配偶体或为特异性结合配偶体的一部分。根据本发明的ch3支架的表位标签通常将包含至少3个氨基酸,优选5至15个氨基酸,或10至20个氨基酸,并且可以包含一个或多个以表位标签的修饰的ab、ef或cd环序列为边界(border)的氨基酸。此外,尽管表位通常是线性的,但根据本发明的表位也可以是构象的;例如,通过折叠多肽将不连续的序列结合在一起形成表位的三级结构而形成的表位。
根据本发明的表位标签的特征在于ch3支架的ab、ef或cd环或其任何组合的野生型氨基酸序列的至少一种修饰,所述修饰导致形成不被内源性抗体识别的表位,所述内源性抗体由个体响应治疗性抗体或包括表位标记的ch3支架的fc融合蛋白产生。在表位标签的设计中可以使用各种策略。例如,ab、cd和ef环内的序列修饰可以基于用衍生自其它结构相关ig-折叠蛋白的相应结构区的序列取代ab、cd或ef环野生型序列。因此,可以通过进行各种ig-折叠蛋白的一级氨基酸序列与ihc的ch3结构域的序列的多序列比对来识别候选表位序列。例如,可以将登记在结构生物信息学蛋白数据库(“pdb”)研究合作实验室中的ig折叠蛋白晶体形式的一级氨基酸序列与ihcch3结构域的一级序列比对。
比对分析可以考虑序列位置保守性的一般类别,包含igg折叠内的间隔、氨基酸电荷、等电点(pl)、极性和三维(3d)结构的保守性,如晶体结构比较所获知的。序列比对还可强调保守序列的绝对同一性,从而预测氨基酸对于维持ch3结构域的三级结构(通常,或者是ab、ef或cd环结构)至关重要。这样的氨基酸也可以称为“锚定(anchor)残基”。例如,val-ser二肽序列、ab环的c末端和位于cd环的n末端的两个残基的trp是锚定残基。如果序列比对揭示了在其他方面保守的候选ig折叠衍生的表位中不存在锚定残基,则供体ig折叠蛋白的野生型序列可以通过用对应于其在野生型ch3序列中的位置的锚定残基取代供体序列中该位置的氨基酸来修饰。
可以衍生根据本发明的表位标签氨基酸序列的具有ig折叠结构域的ig折叠蛋白的实例是信号调节蛋白α(sirpα)和sirpγ(sirpγ)。sirpα和sirpγ的晶体形式在pdb中标识为2wng和4i2x:e。例如,根据本发明,ihc的ab环中与野生型序列诸如ltkn(seqidno.32)相对应的位置的氨基酸可以分别被sirpα和sirpγ衍生的序列tpqh(seqidno.43)和tpeh(seqidno.47)取代。其也可以分别被轻链恒定结构域(“cl”)衍生的序列ltsg(seqidno.45)取代。
类似地,ihc的ef环中与野生型序列诸如ksrwqq(seqidno.59)相对应的位置的氨基酸可以分别被sirpα-、sirpγ-和cl-衍生的序列诸如ltrwdv(seqidno.61)、ldrwdv(seqid65)和kdrwer(seqidno.63)取代。更特别地,seqidno.61、65和63分别对应于以下位置:seqidno.70、71和72描述的sirpα、sirpγ和cl序列的第186-191位;第187-192位;和第183-188位。seqidno.61、65和63中的序列“rw”分别在sirpα、sirpγ和cl中的相应序列位置上替换野生型序列“re”、“pw”和“ey”。更具体地,“rw”序列概括了野生型ch3结构域的ef环中相应序列位置上rw的存在。因此,“rw”用作锚定序列以保留ch3支架的整体结构。
sirpγ和sirpα在ihc的cd环的与野生型序列诸如sngqpenny(seqidno.2)相对应的位置也含有与氨基酸相对应的区域。实际上,ch3野生型序列可以被sirpγ和sirpα衍生的序列ngnelsdf(seqidno.4)取代。ch3cd环的野生型序列可以被cl衍生的序列idgserqng(seqidno.6)取代。seqidno.6和4分别对应于由seqidno.72和70描述的cl和sirpα序列的第150-158位和第156-163位。
设计表位标签的另一种策略涉及选择改变野生型序列的ab、ef和cd环的序列修饰,同时保留环的整体3d结构,包括避免会产生不期望空间效应的修饰。因此,根据本发明的ch3支架可以含有对ab、ef或cd环序列的氨基酸取代、缺失或插入,其中野生型环序列氨基酸的性质,诸如电荷、pi和极性可以保留以维持表位标签的天然框架,但其中改变暴露于溶剂的氨基酸的绝对序列同一性(即,表位特异性抗体可及的表面)以利于检测抗体特异性结合。
当设计用于ch3支架的新颖的表位标签时,考虑3d结构和空间关系的各种策略是本领域已知的。例如,分子可视化系统软件诸如pymol可用于在抗体(例如但不限于衍生自pdb4wi2或4n0u的晶体结构的人igg1fc区域)的背景下对候选ab、ef和cd环表位标签序列进行建模。
根据本发明,ab环表位标签的非限制性实例含有用isrq(seqidno.41)取代野生型序列诸如ltkn(seqidno.32)。而根据本发明,ef环表位标签的非限制性实例是用ndrwqq(seqidno.67)取代野生型序列诸如ksrwqq(seqidno.59)。根据本发明的cd表位标签序列的非限制性实例包括以下序列,其通常用:dnpvy(seqidno.8);sniaqprny(seqidno.10);sngqpekrnenny(seqidno.12);sngqpelanenny(seqidno.14);sngqpdrry(seqidno.16);sngqpdnf(seqidno.18);或sngqpdqqy(seqidno.20)替换igg1野生型序列,例如sngqpenny(seqidno.2)。
如上所述,根据本发明的表位标签的特征在于ch3结构域的ab、ef或cd环或其任何组合的野生型(wt)氨基酸序列的至少一种修饰。因此,根据本发明的具有ch3结构域的ihc或其部分可以具有仅位于其ab环、其ef环或其cd环内的单个表位标签。或者,根据本发明的具有ch3结构域的ihc或其部分可以仅具有两个表位标签,一个表位标签位于其ab环内,另一个位于其ef环内。同样,根据本发明的具有ch3结构域的ihc或其部分可以仅具有两个表位标签,一个表位标签位于其ab环内,另一个位于其cd环内。其还遵循,根据本发明的具有ch3结构域的ihc或其部分可以仅具有两个表位标签,一个表位标签位于其ef环内,另一个位于其cd环内。对于根据本发明的需要具有ch3结构域的ihc或其部分以具有三个表位标签的用途,ihc或其部分将含有分别位于其ab、ef和cd环的三个表位标签。
如上所述,根据本发明优选基于人igg1亚类或在其背景下的基于抗体的生物制品。更特别地,基于表位标记的抗体的生物制品的fc区可以衍生自各种igg1同种异型(jefferis&lefranc)。例如,fc区可以衍生自具有g1m1或ng1m1同种异型的一级氨基酸序列的igg1抗体。由于g1m1和ng1m1同种异型自然通过ab环内的它们的氨基酸序列差异来区分,因此ab环表位标签的设计可以通过在那些位置保持序列同一性来保留那些序列差异。更具体地,野生型g1m1同种异型包含氨基酸序列rdeltknqvs,并且相应的ng1m1序列是reemtknqvs。在前述序列中以粗体突出显示的氨基酸是同种异型的特定决定因素。因此,在ng1m1衍生的fc区的ab环中突出的e和m残基的存在会阻止g1m1特异性抗体与fc区结合。类似地,在g1m1衍生的fc区的ab环中强调的d和l残基的存在会阻止ng1m1特异性抗体与fc区结合。
同种异型也可能存在于igg1抗体的ihc的ch1结构域内。表位标记的基于抗体的生物制品的ihc可以衍生自各种igg1同种异型。例如,ihc可以衍生自具有同种异型g1m3(imgtr120;www.imgt.org)或g1m17(imgtk120)的一级氨基酸序列的igg1抗体。更特别地,表位标记的抗体,基于抗体的生物制品的ihc可以衍生自同种异型的组合。更具体地,ihc可以是g1m17,1(“m1,17”)同种异型,其掺入了g1m1和g1m17同种异型的组合。
除了前述在表位序列设计中对同种异型序列的考虑之外,根据本发明的表位标签可以但不必须包含以表位标签的部分为边界的野生型氨基酸序列,所述表位标签特别设计为使用例如上面讨论的策略充当表位。例如,ab环表位标签序列边界为和igg1的野生型g1m1同种异型相关的(氨基/羧基)边界序列。类似地,ab环表位标签序列边界为和igg1的野生型ng1m1同种异型相关的(氨基/羧基)边界序列。
ef环表位标签序列在其氨基末端侧和羧基末端侧分别以igg1野生型序列(d)和(gqv)为边界,也可以包含前述野生型序列的一部分或全部。最后,cd环表位标签序列在其氨基末端侧和羧基末端侧分别以igg1野生型序列(we)和(ktt)为边界,也可以包括前述野生型序列的一部分或全部。
本发明还包括编码根据本发明的ch3支架的多核苷酸。例如,根据本发明的多核苷酸可以编码单个ch3支架多肽,或含有根据本发明的ch3支架的多肽或其部分,诸如ihc、抗体片段或fc融合蛋白的组分。
可以通过本领域已知的任何方法获得编码本发明的分子的多核苷酸。实际上,可以采用众所周知的分子生物学方法来设计和生产编码具有ab、ef和cd结构环区的ch3支架的多核苷酸,其中至少一个结构环区包括抗体表位氨基酸序列。如上所述,根据本发明的抗体表位氨基酸序列含有ch3支架的ab、ef或cd结构环区中的至少一个的至少一种序列修饰。因此,根据本发明的编码ch3支架的多核苷酸的核苷酸序列可以包含导致ch3支架表达的序列修饰,所述ch3支架相对于野生型ch3结构域序列在其ab、ef或cd环中的至少一个内具有至少一种氨基酸取代、缺失或插入。在这方面,根据本发明的多核苷酸可含有修饰的核苷酸序列,其中核苷酸序列被修饰以表达ch3支架,其中一个或多个ab、ef或cd环含有衍生自结构上相关的ig折叠蛋白的氨基酸序列。例如,根据本发明的多核苷酸可含有编码ch3支架的核苷酸序列,其中其ab、ef或cd环中的一个或多个含有衍生自ig折叠蛋白sirpα、sirpγ或另一免疫球蛋白链诸如恒定轻链的氨基酸序列。
可以将根据本发明的多核苷酸掺入蛋白表达载体中,继而可以将其转染到蛋白表达系统宿主细胞中,以驱动ch3支架或含ch3支架的蛋白诸如ihc、抗体片段或fc融合蛋白的组分的表达。
本发明还提供了用于筛选和识别与根据本发明的ch3支架的工程化表位特异性结合的分子(包括抗体和抗原结合片段)的方法。结合表位的分子可以是例如单克隆抗体或其抗原结合片段。还提供了用于产生与根据本发明的表位标记的ch3支架反应的抗体及其抗原结合片段的方法或过程。
尽管本发明不限制用于识别表位结合分子的方法的可用性,但此类方法的实例包括:(i)使用根据本发明的表位标记的ch3支架作为探针筛选生物样品或肽库;(ii)分离与探针特异性结合的分子;和(iii)识别分子。因此,特异性结合根据本发明的ch3支架的工程化表位的抗体或抗体片段可用于检测含有过量未标记抗体或抗体片段的样品中具有表位标记的ch3支架的抗体。例如,特异性结合根据本发明的ch3支架的工程化表位的抗体可以在溶液中特异性地区别和检测工程化表位标记的(“标记的”)抗体,该溶液也含有未标记的抗体,标记的:未标记的比例至少为1:250000、1:100000、1:10000、1:1000、1:100或其中的任何比例。
实施例
以下实施例描述了人igg的ch3结构域的ab、cd和ef环内的氨基酸序列的设计和分析过程,以创建表位标签,从而允许使用标签的抗体同源物容易检测样品中的专门的抗体。使用四种一般策略设计ab、ef和cd环的表位标签序列:i)用衍生自其他ig折叠蛋白区域的序列取代野生型序列,所述ig折叠蛋白与ch3环结构共享序列或结构相似性,或两者;ii)使用分子建模软件来识别ab、ef和cd环中空间上有利的氨基酸取代;iii)基于就野生型序列而言的结构假设,引入对ab、ef和cd环序列的氨基酸序列、长度或两者的序列修饰;和iv)掺入可商购抗体的同源表位以替换cd环的氨基酸序列。
实施例1。ig-折叠蛋白衍生的表位标签。简而言之,ab、cd和ef环内的序列修饰是基于用衍生自其他结构相关ig折叠蛋白的相应结构区的序列取代ab、cd或ef环野生型序列。该方法导致识别出独特的表位以掺入人ch3结构域。ab环的序列修饰是在g1m1以及ng1m1同种异型的背景下产生的(分别是del与(vs)eem)。
通过进行各种ig折叠蛋白的一级氨基酸序列与衍生自和pdb4wi2相关的晶体结构的人igg1fc区的ch2和ch3结构域序列的多序列比对,识别对应于ab、ef和cd环序列的候选ig折叠ch3环序列,总结于图2a-2c中。截至2018年4月11日,比对包含八个ig-折叠蛋白,其晶体结构可在结构生物信息学蛋白数据库(“pdb”)研究合作实验室中获得。参见http://www.rcsb.org/。候选ig折叠蛋白的pdbid为:2wng;4i2x:a;4i2x:e;4grl;1exu;1t7w;3bvn;和4gup。
比对分析显示,ig折叠蛋白中ab、cd和ef环的间距大体保守。在每个环之间存在序列基序或“锚定残基”。蛋白之间的绝对序列同一性低。那些蛋白具体是4i2x:a、4i2x:e和2wng的子集的多序列比对,识别了与4wi2的ch3结构域具有较高序列同一性的区域(图3)。
比对分析考虑了几个一般的序列位置保守性类别,包括ig折叠区域内的间隔、电荷、等电点(pl)和极性。尽管重点放在潜在保守的锚定残基的绝对同一性上,诸如ab环c末端的缬氨酸-丝氨酸,和cd环n末端的两个残基处的色氨酸。与其他六个ig折叠蛋白相比,在两个晶体2wng和4i2x中发现的蛋白的氨基酸序列在ab、cd和ef环上与igg1ch3序列更保守。晶体2wng对应于调节膜糖蛋白,信号调节蛋白α(sirpα)。晶体4i2x含有两个ig折叠蛋白4i2x:a和4i2x:e,分别对应于抗体fab片段的轻链和sirpγ(sirpγ)。
基于衍生自pdb4wi2igg1晶体的氨基酸序列的ch3支架被设计为掺入sirpα、sirpγ和4i2x:a的cl结构域的区域,所述区域与人ch3的ab、cd和ef环相对应。表1-3总结了sirpα、sirpγ和cl衍生的表位标签氨基酸序列,分别用于替换cd、ab和ef环的野生型(wt)序列。然而,由于其潜在的结构显著性以及由于那些残基不会暴露并且因此不会被检测抗体接近,因此具有指向抗体结构内部的侧链的野生型序列中的氨基酸未被取代。
表1
表2
表3
*具有指向ab内部的侧链的氨基酸以小写字母表示。
实施例2。ab和ef环中的空间上有利的氨基酸取代。使用分子可视化系统软件pymol,使用衍生自pdb4wi2或4n0u晶体结构的人igg1fc区对环的位置进行建模,并识别ab和ef环内溶剂暴露的氨基酸侧链。使用pymol的诱变特征对氨基酸取代进行建模,可以分析ab和ef环内表面暴露残基的各种氨基酸取代的空间效应。未引起空间位阻的取代被考虑用于进一步分析。表4和表5分别含有候选ab和ef表位氨基酸序列,其通过识别ab和ef环的表面暴露环中的空间有利取代形成。
表4
表5
*具有指向ab内部的侧链的氨基酸以小写字母表示。
实施例3。cd环序列修饰。使用分子可视化系统软件pymol,使用衍生自pdb4wi2或4n0u的晶体结构的人igg1fc区对环的位置进行建模,并识别cd环序列内溶剂暴露的氨基酸侧链。使用该方法,可以分析cd环背景下序列修饰的空间效应。基于每个氨基酸位置的电荷、等电点(pl)和极性的相似性选择氨基酸取代。设计cd表位的另一个一般考虑是避免在抗体外侧的疏水区块。表6含有通过使用前述策略选择的候选cd表位标签序列的列表。
表6
实施例4。掺入商业上可获得的抗体的同源表位以替换cd环的氨基酸序列。表7含有通过用商业上可获得的抗体识别的已知表位标签序列替换野生型氨基酸序列而修饰的cd环的描述。
表7
将cd-glu表位掺入m1,17骨架允许用商业上可获得的抗glu抗体(西格玛奥德里奇,cat#ab3788)检测该抗体。如图8所描绘的,与含有野生型m1,17cd环(cd-wt)的抗体或包括cd-4i2x:e表位的抗体相比,通过免疫印迹选择性检测cd-glu。cd-glu可通过免疫印迹在至少15-400ng的范围内检测。在高达400ng的水平下,cd-wt或cd-4i2x:e均未检测到信号。
可以在cd-wt抗体存在下检测cd-glu。将cd-glu和cd-wt以0:100、0:1000、1:0、10:0、100:0、1:100、10:100、100:100(μg/mlcd:-glu:μg/mlcd-wt)的比例混合在磷酸盐缓冲盐水中,并涂覆在elisa板的表面,一式三份。不含cd-glu或cd-wt(0:0)的磷酸盐缓冲盐水用作背景对照。用1:1000稀释的抗cdgluglu多克隆抗血清(西格玛奥德里奇,cat#ab3788)作为一抗和1:5000稀释的小鼠抗兔igg-hrp偶联二抗(南方生物技术公司(southernbiotech),cat#4090-05)检测结合cd-glu。用opd底物显影信号,并在450nm处读取吸光度。如图9所描绘的,当以高达1000μg/ml的浓度涂板时,在背景水平以上未检测到cd-wt。相反,在没有cd-wt的情况下涂板时,可检测到低至10μg/ml的浓度的cd-glu。在存在100μg/mlcd-wt的情况下,还检测到浓度为10μg/ml和100μg/ml的cd-glu。
实施例5。表位的掺入不会显著改变与新生fc受体(fcrn)的结合。fcrn的结合亲和力与抗体的药代动力学(pk)行为相关并且因此与血清半衰期相关。为了解决设计的表位是否会改变igg与fcrn结合的能力并且因此对体内pk产生影响,一组含有cd-wt、cd-glu、cd-4i2x、abef-isnd和abef-4i2x:e表位标签的抗体在相同的可变结构域的背景下表达,并使用基于竞争性fret的测定(cisbio)评估与fcrn的结合。如图10所证明的,cd-wt能够以剂量依赖性方式有效竞争供体标记的人igg的结合。所测试的四个表位中任何一个的掺入并没有显著改变抗体与供体标记的人igg竞争的能力。如设计过程所预测的,这些数据表明,将表位掺入cd或abef环(不同于与fcrn的已知相互作用位点的位点)没有显著改变抗体结合fcrn的能力。因此,预测将表位掺入这些环中不会改变抗体的pk。
实施例6。与fcγ受体的结合在表位之间有所不同。不同类别的免疫效应细胞在其细胞表面表达fcγr的独特组合。抗体通过其fc结构域与那些fcγr的结合调节了免疫效应细胞的活性。例如,cd16a(fcγriiia)在自然杀伤(nk)细胞表面上的结合对于诱导抗体依赖性细胞毒性(adcc)是重要的。cd16a中自然发生的多态性,例如cd16av158f和cd16av176f,改变了受体对igg分子fc结构域的结合亲和力。这继而改变了igg在体外诱导adcc的能力,并与对某些基于抗体的疗法的临床反应相关。使用竞争性fret测定(cisbio),在m1,17同种异型的背景下,评估cd-wtfc,以及cd-glu、cd-4i2x:e、abef-isnd和abef-4i2x:e与cd16a158v结合的能力。如图11所示,cd-glu、abef-isnd和abef-4i2x:e能够以剂量依赖性方式与人igg竞争与cd16a158v的结合,这模拟了用含有野生型(m1,17同种异型)fc结构域的抗体所观察到的。相反,cd-4i2x:e抗体需要高于大约10倍的浓度来达到相同的竞争水平,这与对cd16a158v受体的亲和力降低是一致的。
fcγr家族的其他成员存在于免疫效应细胞的各种亚群上。其中有cd16b(fcγriiib)、cd32a(fcγriia)、cd32b(fcγriib)和cd64(fcγri)。cd32和cd16b是低亲和力igg受体,cd16a以中等亲和力结合,并且cd64以高亲和力与单体igg结合。与cd16a一样,其他改变结合亲和力的fcγr中也存在多态性,诸如cd32ah131r。在biacore8k上使用表面等离子体共振进一步表征野生型(cd-wt)和cd-glu,以测量并直接比较对cd16a176v、cd16a176f、cd16b、cd32a167h、cd32a167r和cd32b的亲和力。在固定有抗fab的系列scm5传感器芯片上捕获约150ru的cd-wt和cd-glu,目的是在与fcγr结合后产生50-100ru的rmax。使用最少10个重复测量每个fcγr的亲和力,并使用对各个fcγr特异的亲和力模型评估。利用二价分析物模型的单循环动力学用于cd64和cd16家族受体,并且稳态结合用于评估与受体cd32家族的结合。那些研究的结果在表8和9以及图12和13中描绘。总体而言,数据反映了cd-wt和cd-glu的相似结合,而cd-glu可能有适度下降的趋势,范围为所有fcγr家族的1.1-1.4倍。在fcγr1结合的情况下,相对于造成差异的cd-wt,数据指向小且一致增加的cd-glu解离率。已知与fcγr1的结合受fc结构域的糖基化状态影响。相对于cd-wt,cd-glu糖基化的细微差异可能造成观察到的差异。
表8.二价分析物模型
表9.研究状态亲和力拟合
实施例7。cd-glu的免疫原性预测。抗体序列的修饰,甚至使用不同的天然存在的同种异型骨架时,都与免疫原性的速率改变有关。为了评价将cd-glu表位标签掺入m1,17骨架中的潜在免疫原性风险,分别使用netmhccons和netmhciipan(3.2版)服务器,通过计算分析引入mhci类和ii类结合肽的氨基酸序列。该分析提供了预测值,以nm结合亲和力给出并作为%排名,与一组针对强结合肽和弱结合肽的200,000个随机天然肽相比。netmhccons评价了321个九氨基酸肽,它们通过一个氨基酸在m1,17同种异型fc氨基酸序列上对齐移位,其含有掺入的cd-glu表位标签(seqidno:73),跨越了人igg1的ch1、ch2和ch3结构域。netmhciipan在含有重链序列的相同cd-glu上评价了315个15氨基酸肽。强mhci结合肽是具有50nm亲和力且相对于对照肽在前0.5%内的肽。弱mhci结合肽是具有500nm亲和力且在前2%内的肽。强和弱mhcii结合肽分别是相对于天然存在的对照组在前2%和10%内的肽。该算法预测了三个强和三个弱mhci结合肽,以及零个强和11个弱mhcii结合肽。强和弱mhci结合肽的实例分别是肽133和116。同样,预测与mhcii弱结合的肽是肽178。所有预测的肽都位于野生型fc结构域的天然存在区域。cd-glu表位的插入既不引入mhci也不引入mhcii结合肽。表10和表11列出了含有cd-glu表位的任何部分的重叠肽的结果。
表10.跨ch3结构域中cd-glu插入物的肽的mhci结合
表11.跨ch3结构域中cd-glu插入物的肽的mhcii结合
实施例8。含有fc结构域的cd-glu保留了热稳定性。对仅在存在或不存在cd-glu表位方面不同的两种抗体进行差示扫描荧光法。两种抗体均以1.55mg/ml(cd_glu)或1.97mg/ml(cd-wt)的浓度在100mmhepes,100mmnacl,50mmnaoac,ph6.0中配制。通过差示扫描荧光法在25℃至95℃在1℃/min的温度斜升下分析热稳定性。绘制了荧光变化对温度变化(d荧光(dfluor)/d温度,nm/℃)的一阶导数图,以定义熔融温度(tms)。如表12所定义的,cd-wt具有三个熔点转变,且cd-glu具有两个熔点转变,这与igg分子的已知熔化相一致。75.2℃的熔点可能对应于cd-glu的fc结构域,比cd-wt测得的熔点低约六度。尽管所观察到的温度低于cd-wt的温度,但与其他临床相关抗体所观察到的温度一致(andersen等人)。
表12.cd-glu对抗体热稳定性的影响
参考文献
andersencb,mannom,rischelcetal(2010)aggregationofamultidomainprotein:acoagulationmechanismgovernsaggregationofamodeligg1antibodyunderweakthermalstress.proteinsci.19:279-290.
jefferisandlefranc(2009)humanimmunoglobulinallotypes:possibleimplicationsforimmunogenicity.mabs.1(4):332-338.
序列表
<110>伊米若梅有限公司
<120>ch3结构域表位标签
<130>172.0001-wo00
<140>62/672,738
<141>2018-05-17
<160>72
<170>patentin版本3.5
<210>1
<211>14
<212>prt
<213>智人
<400>1
trpgluserasnglyglnprogluasnasntyrlysthrthr
1510
<210>2
<211>9
<212>prt
<213>智人
<400>2
serasnglyglnprogluasnasntyr
15
<210>3
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-2wng环表位
<400>3
trpgluserasnglyasngluleuseraspphelysthrthr
1510
<210>4
<211>8
<212>prt
<213>人工
<220>
<223>cd-2wng表位
<400>4
asnglyasngluleuseraspphe
15
<210>5
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-4i2x:a表位
<400>5
trpgluileaspglysergluargglnasnglylysthrthr
1510
<210>6
<211>9
<212>prt
<213>人工
<220>
<223>cd-4i2x:a表位
<400>6
ileaspglysergluargglnasngly
15
<210>7
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-tlk表位
<400>7
trpgluaspasnprovaltyrlysthrthr
1510
<210>8
<211>5
<212>prt
<213>人工
<220>
<223>cd-tlk表位
<400>8
aspasnprovaltyr
15
<210>9
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-cd2hv表位
<400>9
trpgluserasnilealaglnproargasntyrlysthrthr
1510
<210>10
<211>9
<212>prt
<213>人工
<220>
<223>cd-cd2hv表位
<400>10
serasnilealaglnproargasntyr
15
<210>11
<211>18
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-krne表位
<400>11
trpgluserasnglyglnproglulysargasngluasnasntyrlys
151015
thrthr
<210>12
<211>13
<212>prt
<213>人工
<220>
<223>cd-krne表位
<400>12
serasnglyglnproglulysargasngluasnasntyr
1510
<210>13
<211>18
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-lane表位
<400>13
trpgluserasnglyglnprogluleualaasngluasnasntyrlys
151015
thrthr
<210>14
<211>13
<212>prt
<213>人工
<220>
<223>cd-lane表位
<400>14
serasnglyglnprogluleualaasngluasnasntyr
1510
<210>15
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-drr表位
<400>15
trpgluserasnglyglnproaspargargtyrlysthrthr
1510
<210>16
<211>9
<212>prt
<213>人工
<220>
<223>cd-drr表位
<400>16
serasnglyglnproaspargargtyr
15
<210>17
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-dnf-衍生的表位
<400>17
trpgluserasnglyglnprogluaspasnphelysthrthr
1510
<210>18
<211>9
<212>prt
<213>人工
<220>
<223>cd-dnf-衍生的表位
<400>18
serasnglyglnprogluaspasnphe
15
<210>19
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-dqq表位
<400>19
trpgluserasnglyglnproaspglnglntyrlysthrthr
1510
<210>20
<211>9
<212>prt
<213>人工
<220>
<223>cd-dqq表位
<400>20
serasnglyglnproaspglnglntyr
15
<210>21
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-opn表位
<400>21
trpgluthrtrpleuasnproaspproserglnlysthrthr
1510
<210>22
<211>9
<212>prt
<213>人工
<220>
<223>cd-opn表位
<400>22
thrtrpleuasnproaspprosergln
15
<210>23
<211>13
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-glu表位
<400>23
trpglutyrmetprometgluasnasntyrlysthrthr
1510
<210>24
<211>8
<212>prt
<213>人工
<220>
<223>cd-glu表位
<400>24
tyrmetprometgluasnasntyr
15
<210>25
<211>14
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-myc表位
<400>25
trpgluglnlysleuileserglugluaspleulysthrthr
1510
<210>26
<211>9
<212>prt
<213>人工
<220>
<223>cd-myc表位
<400>26
glnlysleuileserglugluaspleu
15
<210>27
<211>12
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-flag表位
<400>27
trpgluasptyrlysaspaspaspasplysthrthr
1510
<210>28
<211>7
<212>prt
<213>人工
<220>
<223>cd-flag表位
<400>28
asptyrlysaspaspaspasp
15
<210>29
<211>15
<212>prt
<213>人工
<220>
<223>具有边界序列的cd-his表位
<400>29
trpgluserasnglyhishishishishishistyrlysthrthr
151015
<210>30
<211>10
<212>prt
<213>人工
<220>
<223>cd-his表位
<400>30
serasnglyhishishishishishistyr
1510
<210>31
<211>10
<212>prt
<213>智人
<400>31
argaspgluleuthrlysasnglnvalser
1510
<210>32
<211>4
<212>prt
<213>智人
<400>32
leuthrlysasn
1
<210>33
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>33
lysaspgluleuserlysasnglnvalser
1510
<210>34
<211>4
<212>prt
<213>人工
<220>
<223>ab环序列
<400>34
leuserlysasn
1
<210>35
<211>10
<212>prt
<213>人工
<220>
<223>ab环表位和边界序列
<400>35
glnaspgluleuserlysasnglnvalser
1510
<210>36
<211>4
<212>prt
<213>人工
<220>
<223>ab环序列
<400>36
leuserlysasn
1
<210>37
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>37
argaspgluileserlysasnglnvalser
1510
<210>38
<211>4
<212>prt
<213>人工
<220>
<223>ab环序列
<400>38
ileserlysasn
1
<210>39
<211>10
<212>prt
<213>人工
<220>
<223>ab环表位
<400>39
glnaspgluileserlysasnglnvalser
1510
<210>40
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab-isrq表位
<400>40
argaspgluileserargglnglnvalser
1510
<210>41
<211>4
<212>prt
<213>人工
<220>
<223>ab-isrq表位
<400>41
ileserarggln
1
<210>42
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab-2wng表位
<400>42
argaspgluthrproglnhisglnvalser
1510
<210>43
<211>4
<212>prt
<213>人工
<220>
<223>ab-2wng表位
<400>43
thrproglnhis
1
<210>44
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab-4i2x:a表位
<400>44
argaspgluleuthrserglyglnvalser
1510
<210>45
<211>4
<212>prt
<213>人工
<220>
<223>ab-4i2x:a表位
<400>45
leuthrsergly
1
<210>46
<211>9
<212>prt
<213>人工
<220>
<223>具有边界序列的ab-4i2x:e表位
<400>46
trpgluthrprogluhislysthrthr
15
<210>47
<211>4
<212>prt
<213>人工
<220>
<223>ab-4i2x:e表位
<400>47
thrprogluhis
1
<210>48
<211>10
<212>prt
<213>智人
<400>48
arggluglumetthrlysasnglnvalser
1510
<210>49
<211>4
<212>prt
<213>智人
<400>49
metthrlysasn
1
<210>50
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>50
lysgluglumetserlysasnglnvalser
1510
<210>51
<211>4
<212>prt
<213>人工
<220>
<223>ab环表位
<400>51
metserlysasn
1
<210>52
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>52
glnglugluileserlysasnglnvalser
1510
<210>53
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab表位环
<400>53
lysglugluileserlysasnglnvalser
1510
<210>54
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>54
glnglugluileserlysasnglnvalser
1510
<210>55
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>55
argglugluileserargglnglnvalser
1510
<210>56
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>56
argglugluthrproglnhisglnvalser
1510
<210>57
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ab环表位
<400>57
argglugluleuthrserglnglnvalser
1510
<210>58
<211>10
<212>prt
<213>智人
<400>58
asplysserargtrpglnglnglyasnval
1510
<210>59
<211>6
<212>prt
<213>智人
<400>59
lysserargtrpglngln
15
<210>60
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ef-2wng表位
<400>60
aspleuthrargtrpaspvalglyasnval
1510
<210>61
<211>6
<212>prt
<213>人工
<220>
<223>ef-2wng表位
<400>61
leuthrargtrpaspval
15
<210>62
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ef-4i2x:a表位
<400>62
asplysaspargtrpgluargglyasnval
1510
<210>63
<211>6
<212>prt
<213>人工
<220>
<223>ef-4i2x:e表位
<400>63
lysaspargtrpgluarg
15
<210>64
<211>11
<212>prt
<213>人工
<220>
<223>具有边界序列的ef-4i2x:e表位
<400>64
trpgluleuaspargtrpaspvallysthrthr
1510
<210>65
<211>6
<212>prt
<213>人工
<220>
<223>ef-4i2x:e表位
<400>65
leuaspargtrpaspval
15
<210>66
<211>10
<212>prt
<213>人工
<220>
<223>具有边界序列的ef-nd表位
<400>66
aspasnaspargtrpglnglnglyasnval
1510
<210>67
<211>6
<212>prt
<213>人工
<220>
<223>ef-nd表位
<400>67
asnaspargtrpglngln
15
<210>68
<211>106
<212>prt
<213>智人
<400>68
glyglnproarggluproglnvaltyrthrleuproproserargasp
151015
gluleuthrlysasnglnvalserleuthrcysleuvallysglyphe
202530
tyrproseraspilealavalglutrpgluserasnglyglnproglu
354045
asnasntyrlysthrthrproprovalleuaspseraspglyserphe
505560
pheleutyrserlysleuthrvalasplysserargtrpglnglngly
65707580
asnvalphesercysservalmethisglualaleuhisasnhistyr
859095
thrglnlysserleuserleuserprogly
100105
<210>69
<211>106
<212>prt
<213>智人
<400>69
glyglnproarggluproglnvaltyrthrleuproproserargglu
151015
glumetthrlysasnglnvalserleuthrcysleuvallysglyphe
202530
tyrproseraspilealavalglutrpgluserasnglyglnproglu
354045
asnasntyrlysthrthrproprovalleuaspseraspglyserphe
505560
pheleutyrserlysleuthrvalasplysserargtrpglnglngly
65707580
asnvalphesercysservalmethisglualaleuhisasnhistyr
859095
thrglnlysserleuserleuserprogly
100105
<210>70
<211>327
<212>prt
<213>智人
<400>70
gluglugluleuglnvalileglnproasplysservalservalala
151015
alaglygluseralaileleuhiscysthrvalthrserleuilepro
202530
valglyproileglntrppheargglyalaglyproalaarggluleu
354045
iletyrasnglnlysgluglyhispheproargvalthrthrvalser
505560
gluserthrlysarggluasnmetasppheserileserileserasn
65707580
ilethrproalaaspalaglythrtyrtyrcysvallysphearglys
859095
glyserproaspthrgluphelysserglyalaglythrgluleuser
100105110
valargalalysproseralaprovalvalserglyproalaalaarg
115120125
alathrproglnhisthrvalserphethrcysgluserhisglyphe
130135140
serproargaspilethrleulystrpphelysasnglyasngluleu
145150155160
serasppheglnthrasnvalaspprovalglygluservalsertyr
165170175
serilehisserthralalysvalvalleuthrarggluaspvalhis
180185190
serglnvalilecysgluvalalahisvalthrleuglnglyasppro
195200205
leuargglythralaasnleusergluthrileargvalproprothr
210215220
leugluvalthrglnglnprovalargalagluasnglnvalasnval
225230235240
thrcysglnvalarglysphetyrproglnargleuglnleuthrtrp
245250255
leugluasnglyasnvalserargthrgluthralaserthrvalthr
260265270
gluasnlysaspglythrtyrasntrpmetsertrpleuleuvalasn
275280285
valseralahisargaspaspvallysleuthrcysglnvalgluhis
290295300
aspglyglnproalavalserlysserhisaspleulysvalserthr
305310315320
arghishishishishishis
325
<210>71
<211>328
<212>prt
<213>智人
<400>71
gluglugluleuglnmetileglnproglulysleuleuleuvalthr
151015
valglylysthralathrleuhiscysthrvalthrserleuleupro
202530
valglyprovalleutrppheargglyvalglyproglyarggluleu
354045
iletyrasnglnlysgluglyhispheproargvalthrthrvalser
505560
aspleuthrlysargasnasnmetasppheserileargileserser
65707580
ilethrproalaaspvalglythrtyrtyrcysvallysphearglys
859095
glyserprogluasnvalgluphelysserglyproglythrglumet
100105110
alaleuglyalalysproseralaprovalvalleuglyproalaala
115120125
argthrthrprogluhisthrvalserphethrcysgluserhisgly
130135140
pheserproargaspilethrleulystrpphelysasnglyasnglu
145150155160
leuserasppheglnthrasnvalaspprothrglyglnservalala
165170175
tyrserileargserthralaargvalvalleuaspprotrpaspval
180185190
argserglnvalilecysgluvalalahisvalthrleuglnglyasp
195200205
proleuargglythralaasnleuserglualaileargvalpropro
210215220
thrleugluvalthrglnglnprometargvalglyasnglnvalasn
225230235240
valthrcysglnvalarglysphetyrproglnserleuglnleuthr
245250255
trpsergluasnglyasnvalcysglnarggluthralaserthrleu
260265270
thrgluasnlysaspglythrtyrasntrpthrsertrppheleuval
275280285
asnileseraspglnargaspaspvalvalleuthrcysglnvallys
290295300
hisaspglyglnleualavalserlysargleualaleugluvalser
305310315320
thrarghishishishishishis
325
<210>72
<211>214
<212>prt
<213>智人
<400>72
aspilevalilethrglnserprolysphemetserthrservalgly
151015
aspargvalserilethrcyslysalaserglnaspvalserthrala
202530
valalatrppheglnglnlysproglyglnserprolysleuleuile
354045
tyrseralasertyrargtyrthrglyvalproaspargphethrgly
505560
serglyserglythraspphethrphethrileserservalglnala
65707580
gluaspleualavaltyrtyrcysglnglnhistyrserthrprotrp
859095
thrpheglyglyglythrlysleugluilelysargalaaspalaala
100105110
prothrvalserilepheproprosersergluglnleuthrsergly
115120125
glyalaservalvalcyspheleuasnasnphetyrprolysaspile
130135140
asnvallystrplysileaspglysergluargglnasnglyvalleu
145150155160
asnsertrpthraspglnaspserlysaspserthrtyrsermetser
165170175
serthrleuthrleuthrlysaspglutyrgluarghisasnsertyr
180185190
thrcysglualathrhislysthrserthrserproilevallysser
195200205
pheasnargasnglucys
210
<210>73
<211>325
<212>prt
<213>智人
<400>73
alaserthrlysglyproservalpheproleualaproserserlys
51015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
lysvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
proalaprogluleuleuglyglyproservalpheleupheleupro
115120125
prolysprolysaspthrleumetileserargthrprogluvalthr
130135140
cysvalvalvalaspvalserhisgluaspprogluvallyspheasn
145150155160
trptyrvalaspglyvalgluvalhisasnalalysthrlysproarg
165170175
glugluglntyrasnserthrtyrargvalvalservalleuhisgln
180185190
asptrpleuasnglylysglutyrlyscyslysvalserasnlysala
195200205
leuproalaproileglulysthrileserlysalalysglyglnpro
210215220
argaspproglnvaltyrthrleuproproserargaspgluleuthr
225230235240
lysasngluvalserleuthrcysleuvallysglyphetyrproser
245250255
aspilealavalglutrpglutyrmetprometgluasnasntyrlys
260265270
thrthrproprovalleuaspseraspglyserphepheleutyrser
275280285
lysleuthrvalasplysserargtrpglnglnglyasnvalpheser
290295300
cysservalmethisglualaleuhisasnhistyrthrglnlysser
305310315320
leuserleuprogly
325
- 还没有人留言评论。精彩留言会获得点赞!