诱导间歇性禁食和调节自噬的方法
诱导间歇性禁食和调节自噬的方法
1.相关申请的交叉引用
2.本技术要求2019年12月2日提交的美国临时专利申请号62/942,354的优先权,该申请的全部内容通过引用并入本文。
技术领域
3.本公开涉及用于通过定期施用精氨酸消耗剂在受试者中诱导间歇性禁食和调节细胞或器官中的自噬的方法。
背景技术:
4.间歇性禁食(间歇性能量限制)已被证明可以带来许多健康益处。它可以帮助预防和治疗多种疾病。例如,间歇性禁食可以预防糖尿病、癌症、心脏病和神经退化。它还可以帮助减少肥胖、高血压、哮喘和类风湿性关节炎。它可以促进多系统再生,增强认知能力和健康寿命[brandhorst等人cell metab.2015;22(1):86-99]并延迟衰老。
[0005]
间歇性禁食是在给定时期内在自愿禁食(或减少卡路里摄入)和非禁食之间循环的各种喂食模式的总称。存在不同的间歇性禁食方法,主要通过食物剥夺和/或消耗卡路里限制饮食来实现。例如,一种方法是隔日禁食,包括在受试者摄入少于通常能量需求的25%的24小时禁食日和随后的24小时非禁食盛宴日之间交替。另一种方法是定期禁食,包括任何超过24小时的连续禁食期,例如5:2饮食,其中每周禁食2天。在禁食期间,受试者的定期每日卡路里摄入量非常低或约为25%。另一种方法是限时禁食,包括每天只在一定小时数内进食,例如16:8饮食(禁食16小时和非禁食8小时进行循环)。一些研究通过连续几天喂食模拟禁食的饮食(禁食模拟饮食“fmd”)实现间歇性禁食,并以规律的时间间隔重复fmd循环[brandhorst等人cell metab.2015;22(1):86-99]。虽然这些喂食模式和/或饮食可以带来健康益处,但可能难以实现长期依从性。许多药剂还可以通过减少食物摄入来发挥健康益处。例如,雷公藤红素(celastrol)诱导的体重减轻是由厌食症(hypophagia)引起的[pfuhlmann等人diabetes.2018;67(11):2456-2465]。然而,许多这些药剂的半衰期短,需要每天给药才能达到治疗效果。
[0006]
细胞可以发生巨自噬(简称自噬)。在自噬过程中,细胞以一种受调节的方式消耗自身的部分,这包括将细胞成分递送到溶酶体以通过双膜结合结构进行降解。自噬以低水平组成性地发生,以平衡生物分子的不断合成。它通过降解长寿命的细胞内蛋白质和受损的细胞器,并将它们的成分再循环成代谢前体来维持细胞的完整性。自噬被认为是维持细胞稳态以及应对压力的关键过程。当细胞受到压力时,例如营养缺乏或禁食,自噬被强烈上调。这种上调增加了大量的多种底物的隔离和降解,包括整个细胞器的降解,从而将大分子释放回胞质溶胶以提供必要的代谢反应并产生能量。最近的研究表明,自噬还可以清除多种细胞内病原体。自噬受损与许多类型的疾病有关,包括癌症、传染性疾病、神经退行性疾病、炎症、年龄相关疾病和代谢疾病。有了关于间歇性禁食、自噬和疾病之间关系的新发现、知识和信息,就有可能设计出有效的治疗方法和策略。可以调节自噬的药剂是治疗许多疾
病的潜在疗法[levine等人,j clin invest.2015;125(1):14-24]。
[0007]
自噬是一个受到严格调节的分解代谢过程,其中受损的蛋白质和细胞器被递送到溶酶体并降解以释放游离氨基酸到细胞质中。自噬通过哺乳动物雷帕霉素靶蛋白(mtor)复合物1(mtorc1)响应氨基酸饥饿而被特异性活化,mtorc1是细胞的中心代谢传感器。mtorc1是协调氨基酸可用性和自噬的关键枢纽[carroll等人,amino acids.2015;47(10):2065-2088]。当材料丰富时,它会抑制自噬诱导。当细胞缺乏这些营养物质时,mtorc1会失活,从而促进自噬的增加。
[0008]
肥胖诱导的糖尿病以高血糖、胰岛素抵抗和进行性β细胞衰竭为特征。在患有肥胖诱导的糖尿病的小鼠的胰岛中,liu等人[autophagy.2017;13(11):1952-1968]观察到β细胞死亡增加和自噬潮受损。他们发现间歇性禁食会刺激自噬潮以改善肥胖诱导的糖尿病。他们表明,尽管持续摄入高脂肪,但是间歇性禁食通过增强葡萄糖刺激的胰岛素分泌、β细胞存活和neurog3(一种胰腺再生标志物)的核表达来恢复胰岛中的自噬潮并改善葡萄糖耐量。他们发现间歇性禁食不能挽救β细胞死亡或诱导肥胖小鼠的neurog3表达,这些小鼠患有溶酶体功能障碍,继发于溶酶体膜蛋白lamp2的缺乏或becn1/beclin 1的单倍体不足,becn1/beclin1是一种对自噬体形成至关重要的蛋白质。因此,间歇性禁食可以通过自噬-溶酶体通路保持细胞器质量,从而提高β细胞存活率,并且可以刺激肥胖诱导的糖尿病的再生标志物。
[0009]
因此,需要开发用于诱导间歇性禁食状态和/或调节自噬受试者的新方法,以克服上述缺点中的至少一些。
技术实现要素:
[0010]
本文公开了使用长效精氨酸消耗剂的定期给药来诱导间歇性禁食和自噬的周期性发生以给受试者带来健康益处。精氨酸消耗剂可以是精氨酸酶、精氨酸脱亚氨酶或精氨酸脱羧酶。这些酶的循环半衰期可以通过使用本领域已知的任何常规方法来延长,例如通过聚乙二醇化、与白蛋白结合结构域或人血清白蛋白或人igg fc结构域进行融合。精氨酸消耗剂可以单独施用,或与其他方法或药剂联合施用以增强间歇性禁食和自噬,例如二甲双胍及其类似物、类视黄醇及其衍生物、绿茶多酚(-)-表没食子儿茶素-3-没食子酸酯(egcg)及其衍生物、雷帕霉素及其类似物。
[0011]
在第一方面,本文提供了一种在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法,包括向受试者施用治疗有效量的精氨酸消耗剂的步骤。
[0012]
在某些实施方案中,在受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬导致至少一种自噬相关或间歇性禁食相关疾病或健康状况的治疗,所述治疗选自由以下组成的组:增加受试者寿命,预防衰老的症状或预防与年龄有关的疾病,和促进细胞再生。
[0013]
在某些实施方案中,受试者血清中的精氨酸浓度维持在低于50μm、低于25μm、低于20μm、低于10μm或低于5μm。
[0014]
在某些实施方案中,精氨酸消耗剂是精氨酸酶蛋白、精氨酸脱亚氨酶蛋白或精氨酸脱羧酶蛋白。
[0015]
在某些实施方案中,精氨酸酶蛋白、精氨酸脱亚氨酶蛋白或精氨酸脱羧酶蛋白还包含一个或多个聚乙二醇(peg)基团。
[0016]
在某些实施方案中,精氨酸酶蛋白包含具有seq id no:101、seq id no:102、seq id no:103或seq id no:104的多肽。
[0017]
在某些实施方案中,精氨酸酶蛋白、精氨酸脱亚氨酶蛋白或精氨酸脱羧酶蛋白还包含白蛋白结合结构域或人血清白蛋白,或人igg fc结构域。
[0018]
在某些实施方案中,精氨酸消耗剂是包含abd多肽和精氨酸酶多肽、abd多肽和精氨酸脱亚氨酶多肽、或abd多肽和精氨酸脱羧酶多肽的融合蛋白。
[0019]
在某些实施方案中,精氨酸消耗剂包含与seq id no:49、seq id no:50、seq id no:51、seq id no:52、seq id no:53、seq id no:54、seq id no:55、seq id no:56、seq id no:75、seq id no:107或seq id no:76具有至少98%序列同源性的多肽。
[0020]
在某些实施方案中,精氨酸消耗剂与治疗有效量的自噬诱导剂共同施用。
[0021]
在某些实施方案中,自噬诱导剂选自由类视黄醇衍生物、(-)-表没食子儿茶素-3-没食子酸酯(egcg)衍生物、绿茶儿茶素和雷帕霉素衍生物组成的组。
[0022]
在某些实施方案中,自噬诱导剂选自由卡马西平、可乐定、锂、二甲双胍、雷帕霉素(和雷帕霉素类似物)、利美尼定、丙戊酸钠、维拉帕米、三氟拉嗪、他汀类药物、酪氨酸激酶抑制剂、bh3模拟物、咖啡因、ω-3多不饱和脂肪酸、白藜芦醇、亚精胺、维生素d、海藻糖、多酚(-)-表没食子儿茶素-3-没食子酸酯及其组合组成的组。
[0023]
在某些实施方案中,精氨酸消耗剂与治疗有效量的葡萄糖降低剂共同施用。
[0024]
在某些实施方案中,葡萄糖降低剂是α-葡萄糖苷酶抑制剂、双胍、胆汁酸螯合剂、多巴胺-2激动剂、二肽基肽酶4(dpp-4)抑制剂、美格列奈、钠-葡萄糖转运蛋白2(sglt2)抑制剂、磺酰脲、噻唑烷二酮或其组合。
[0025]
在某些实施方案中,双胍是二甲双胍;α-葡萄糖苷酶抑制剂是阿卡波糖或米格列醇;胆汁酸螯合剂是考来维仑;多巴胺-2激动剂是溴隐亭;dpp-4抑制剂是阿格列汀、利格列汀、沙格列汀或西格列汀;美格列奈是那格列奈或瑞格列奈;sglt2抑制剂是卡格列净、达格列净或恩格列净;磺酰脲是氯磺丙脲、格列美脲、格列吡嗪或格列本脲;并且噻唑烷二酮是罗格列酮或吡格列酮。
[0026]
在某些实施方案中,精氨酸消耗剂与治疗有效量的类视黄醇衍生物共同施用。
[0027]
在某些实施方案中,类视黄醇衍生物是阿曲汀(acitretin)、阿利维a酸贝沙罗汀(alitretinoin bexarotene)、异维a酸(isotretinoin)、视黄醇、视黄酸或其药学上可接受的盐。
[0028]
在某些实施方案中,类视黄醇衍生物是视黄酸。
[0029]
在某些实施方案中,精氨酸消耗剂与治疗有效量的(-)-表没食子儿茶素-3-没食子酸酯(egcg)衍生物、绿茶儿茶素或其药学上可接受的盐或产物共同施用。
[0030]
在某些实施方案中,egcg衍生物是egcg或其药学上可接受的盐或egcg过乙酸盐。
[0031]
在某些实施方案中,精氨酸消耗剂与治疗有效量的雷帕霉素衍生物或其药学上可接受的盐共同施用。
附图说明
[0032]
当结合附图时,根据以下具体实施方式,本公开的以上和其他目标和特征将变得显而易见,其中:
[0033]
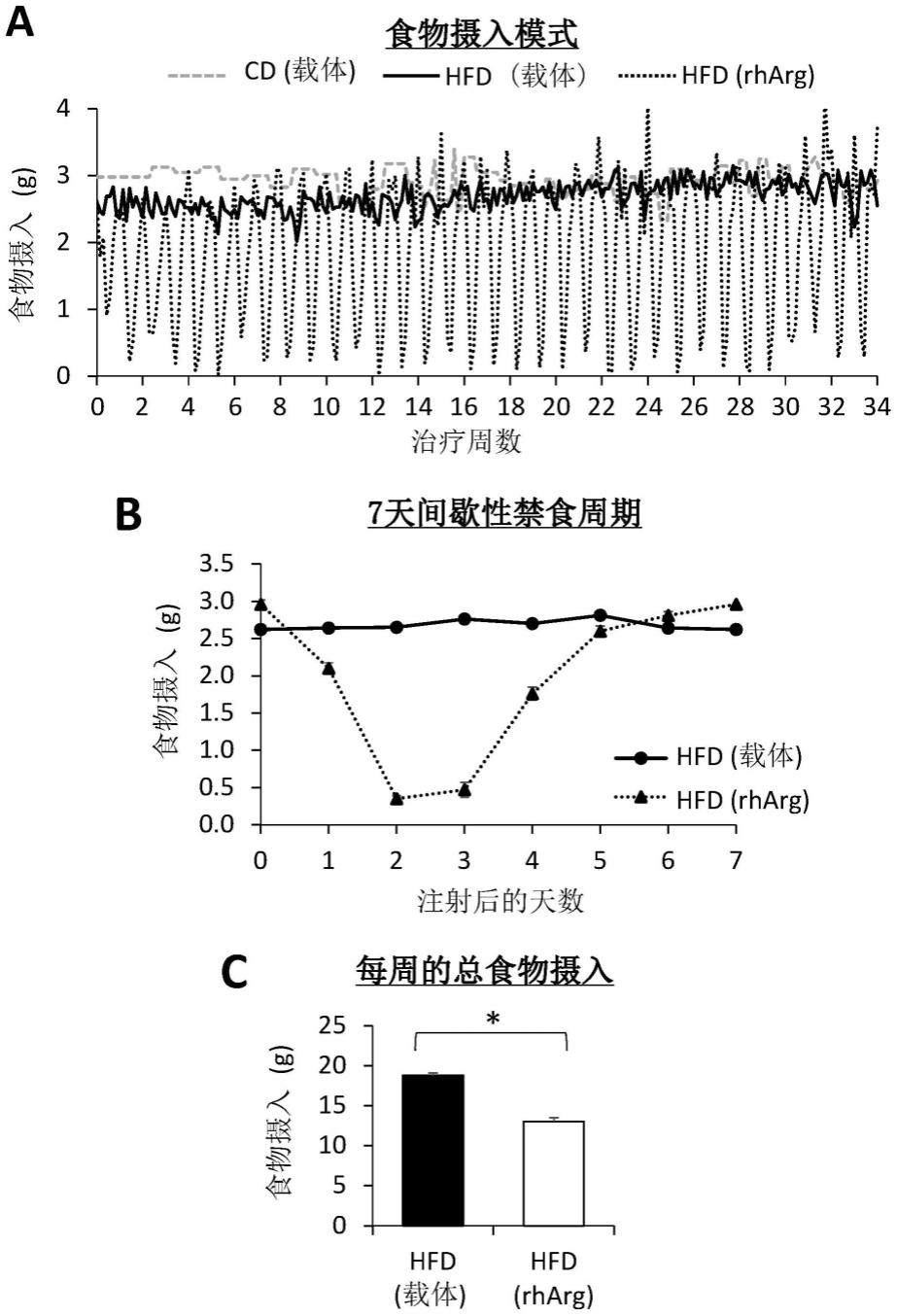
图1说明了当每周一次施用约600u n-abd094-rharg(seq id no:50)(rharg)持续34周时,预先存在通过从5周大开始喂食高脂肪饮食(hfd)持续12周而诱导的肥胖症的c57bl/6j雄性小鼠(称为饮食诱导肥胖(dio)小鼠)表现出由禁食期和再喂食期组成的重复7天间歇性禁食周期。(a)3组小鼠的食物摄入模式:喂食hfd并注射rharg的dio小鼠[hfd(rharg)组];喂食hfd并注射生理盐水(载体)的dio小鼠[hfd(载体)组];喂食普通饲料(cd)并注射载体的小鼠[cd(载体)组]作为瘦对照组。(b)7天间歇性禁食周期中每天的平均食物摄入。第0天代表rharg注射当天,即前一个周期的第7天。第7天代表第0天注射rharg后的第7天,并且是下一个周期的第0天。(c)hfd(rharg)组的每周总食物摄入比hfd(载体)组减少约30%。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=8-12。
[0034]
图2说明向图1中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可以在治疗的6-7周内诱导显著的体重减轻,并且在剩余的治疗期中,体重保持在30g左右相对恒定。(a)34周治疗期间的体重变化。(b)治疗期结束时3组小鼠的代表性图像。
[0035]
图3说明当每周一次施用约600u n-abd094-rharg(seq id no:50)(rharg)持续49周时,预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠表现出重复7天间歇性禁食周期。(a)3组小鼠的食物摄入模式:喂食hfd并注射rharg的dio小鼠[hfd(rharg)组];喂食hfd并注射生理盐水(载体)的dio小鼠[hfd(载体)组];喂食普通饲料(cd)并注射载体的小鼠[cd(载体)组]作为瘦对照组。(b)7天间歇性禁食周期中每天的平均食物摄入。(c)hfd(rharg)组的每周总食物摄入比hfd(载体)组减少约29%。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,对于cd(载体)组,n=5;对于hfd(载体)和hfd(rharg)组,n=9。
[0036]
图4说明向图3中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)持续49周可以在6-7周内诱导显著的体重减轻,并且在剩余的治疗期中,体重保持在30g左右相对恒定。
[0037]
图5说明图1中hfd(rharg)组小鼠的血清中检测到的抗rharg抗体不具有中和活性。(a,b)rharg治疗后5周(a)和23周(b)时从小鼠获取的血清中的抗rharg抗体滴度是相似的。(c)rharg治疗后23周时获取的血清与rharg一起孵育,并且对中和rharg的酶活性没有任何影响。
[0038]
图6说明向图1中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)持续34周可有效减少脂肪量。(a)主要内脏(肾周)和皮下(腹股沟)白色脂肪组织(wat)仓库和主要(肩胛间)棕色脂肪组织(bat)仓库的新鲜解剖脂肪垫的代表性图像。(b)与用载体治疗的dio小鼠[hfd(载体)组]相比,用rharg治疗的dio小鼠[hfd(rharg)组]的肾周和腹股沟wat和肩胛间bat的质量显著降低。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0039]
图7说明向图1中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)持续34周可以有效减少肝脏质量,并降低一些常用的肝损伤生物标志物的血清浓度,达到与瘦对照小鼠[cd(载体)组]相似的水平。(a)3组小鼠的新鲜全肝的代表性
图像。(b)肝脏质量。(c)丙氨酸转氨酶(alt)和天冬氨酸转氨酶(ast)的血清浓度。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0040]
图8说明向图1中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)持续34周可以减少肾脏质量,并降低常用的肾损伤生物标志物的尿液浓度,达到与瘦对照小鼠[cd(载体)组]相似的水平。(a)肾脏质量。(b)尿液中白蛋白与肌酐的比率。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0041]
图9说明向图1中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)持续34周可以减少心脏质量,并降低血压和心率,达到与瘦对照小鼠[cd(载体)组]相似的水平。(a)心脏质量。(b-e)rharg治疗后12周(b,c)和27周(d,e),使用无创血压监测系统(coda scientific)通过尾套法分别测量的收缩压和舒张压(b,d)和心率(c,e)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0042]
图10说明向图3中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转胰岛素抵抗。(a-c)在(a)之前和rharg治疗后16周(b)和32周(c)进行胰岛素耐量试验(itt)。itt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0043]
图11说明向图3中的喂食hfd的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转受损的葡萄糖耐量。(a-c)在(a)之前和rharg治疗后15周(b)和31周(c)进行葡萄糖耐量试验(gtt)。gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0044]
图12说明对预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠喂食预定量的hfd以创建人工7天间歇性禁食周期[hfd(人工if)组]持续5周可以有效降低小鼠体重,该人工7天间歇性禁食周期模拟每周一次施用约600u n-abd094-rharg(seq id no:50)的dio小鼠[hfd(rharg)组]的食物摄入模式。(a)5周期间的食物摄入模式。(b)hfd(rharg)组和hfd(人工if)组每周的总食物摄入分别比用载体治疗的dio小鼠[hfd(载体)组]减少26%和34%。(c)5周期间的体重变化。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=5。
[0045]
图13说明图12中的经历使用hfd进行5周的人工7天间歇性禁食喂食周期的dio雄性小鼠[hfd(人工if)组]显示肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪垫质量显著减少,并且与用载体治疗的dio小鼠[hfd(载体)组]相比,还具有肝脏质量的显著减少。脂肪垫和肝脏质量与每周一次施用rharg的dio小鼠[hfd(rharg)组]相当。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0046]
图14说明图12中的经历使用hfd的人工7天间歇性禁食喂食周期的dio雄性小鼠[hfd(人工if)组]显示出葡萄糖耐量的显著改善,但与用载体治疗的dio小鼠[hfd(载体)组]相比,没有显示胰岛素敏感性的改善。相比之下,dio小鼠[hfd(rharg)组]在rharg治疗后2周时表现出胰岛素敏感性的显著改善。(a)在rharg治疗后2周和4周进行的胰岛素耐量试验(itt)。(b)在rharg治疗后3周进行的葡萄糖耐量试验(gtt)。itt和gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0047]
图15说明预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠当hfd的每日食物摄入
减少30%[hfd(减少)组]持续5周时,与每周一次施用约600u n-abd094-rharg(seq id no:50)的dio小鼠[hfd(rharg)组]相比显示出显著更少的体重减轻。(a)随意喂食或喂食固定量(2.0g)的hfd的用载体和rharg治疗的dio小鼠的食物摄入模式。(b)hfd(减少)组每周的总食物摄入比随意喂食hfd的用载体治疗的dio小鼠[hfd(载体)组]低约30%。(c)5周期间的体重变化。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=5。
[0048]
图16说明图15中的接受减少30%的hfd的每日食物摄入持续5周的dio雄性小鼠[hfd(减少)组]与每周一次施用rharg的dio小鼠[hfd(rharg)组]相比,显示肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪垫质量以及肝脏质量的显著更少的减少。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem,每组n=5。
[0049]
图17说明接受减少30%的hfd的每日食物摄入的dio雄性小鼠[hfd(减少)组]显示与每周一次施用约600u n-abd094-rharg(seq id no:50)的dio小鼠[hfd(rharg)组]类似的葡萄糖耐量的显著改善。相比之下,与随意喂食的载体治疗的dio小鼠[hfd(载体)组]相比,只有hfd(rharg)组的小鼠而不是hfd(减少)组的小鼠显示出胰岛素敏感性的改善。(a)在治疗2周后进行的胰岛素耐量试验(itt)。(b)在治疗3周后进行的葡萄糖耐量试验(gtt)。itt和gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem,每组n=5。
[0050]
图18说明向预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠[hfd(peg-rharg)组]每周一次施用约250u聚乙二醇化his-rharg(seq id no:101)持续8周可诱导重复7天间歇性禁食周期。(a)8周治疗期中的食物摄入模式。(b)7天间歇性禁食周期中每天的平均食物摄入。(c)hfd(peg-rharg)组的每周总食物摄入比载体治疗的dio小鼠[hfd(载体)组]减少约28%。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=5。
[0051]
图19说明向图18中的喂食hfd的dio雄性小鼠[hfd(peg-rharg)组]施用聚乙二醇化his-rharg(seq id no:101)可在6-7周内诱导显著的体重减轻,达到与喂食普通饲料的载体治疗的瘦对照小鼠[cd(载体)组]相似的水平。每组n=5。
[0052]
图20说明向图18中的喂食hfd的dio雄性小鼠[hfd(peg-rharg)组]施用聚乙二醇化his-rharg(seq id no:101)可有效逆转胰岛素抵抗并改善葡萄糖耐量。(a)在rharg治疗后7周进行的胰岛素耐量试验(itt)。(b)在rharg治疗后6周进行的葡萄糖耐量试验(gtt)。itt和gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0053]
图21说明向图18中的喂食hfd的dio雄性小鼠[hfd(peg-rharg)组]施用聚乙二醇化his-rharg(seq id no:101)持续8周可有效减少肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪量以及肝脏,达到与瘦对照小鼠[cd(载体)组]相当的重量。与载体治疗的dio小鼠[hfd(载体)组]相比,肾脏和心脏的质量也显著降低。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0054]
图22说明每周一次向预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠施用50u n-abd094-rharg-co
2+
[seq id no:50(钴取代)]持续2周可诱导7天间歇性禁食周期和伴随的体重减轻。(a)在7天间歇性禁食周期中具有禁食期和再喂食期的食物摄入模式。(b)体重变化。n=1。
rharg(seq id no:50)持续56周可显著减少肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪量,达到与喂食饲料的年龄匹配的载体治疗的对照小鼠[cd(载体)组]相当的水平。包括肝脏、肾脏和心脏在内的几个主要器官的质量也显著低于用载体治疗的喂食hfd的dio小鼠[hfd(载体)组]。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0064]
图32说明向图29中的喂食hfd的dio雌性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转胰岛素抵抗。(a-c)在(a)之前和rharg治疗后15周(b)和31周(c)进行胰岛素耐量试验(itt)。itt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0065]
图33说明向图29中的喂食hfd的dio雌性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转受损的葡萄糖耐量。(a-c)在(a)之前和rharg治疗后16周(b)和30周(c)进行葡萄糖耐量试验(gtt)。gtt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0066]
图34说明每周一次向图29中的喂食hfd的dio雌性小鼠[hfd(rharg)组]长期施用n-abd094-rharg(seq id no:50)可以显著改善它们的神经肌肉力量和协调性,达到与喂食饲料的年龄匹配的对照小鼠[cd(载体)组]相当的水平。(a)治疗后54周进行的倒置网格悬挂测试。(b)治疗后55周进行的旋转棒测试。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0067]
图35说明每周一次向图29中的喂食hfd的dio雌性小鼠[hfd(rharg)组]长期施用n-abd094-rharg(seq id no:50)持续56周可有效预防肝细胞癌发生。
[0068]
图36说明当每周一次施用约600u n-abd094-rharg(seq id no:50)(rharg)持续25周时,预先存在通过从5周大开始喂食hfd而诱发的肥胖症的约16个月大(相当于人类50岁中期)的c57bl/6j雄性小鼠表现出重复7天间歇性禁食周期。该反应类似于图1中的c57bl/6j dio雄性小鼠,其在4-5个月大(相当于人类20岁中期)开始用n-abd094-rharg(seq id no:50)治疗。(a)3组小鼠的食物摄入模式:喂食hfd并注射rharg的16个月大的dio小鼠[hfd年老(rharg)组];喂食hfd并注射生理盐水(载体)的16个月大的dio小鼠[hfd年老(载体)组];喂食普通饲料(cd)并注射载体的5个月大的小鼠[cd年轻(载体)组]作为年轻瘦对照组。(b)7天间歇性禁食周期中每天的平均食物摄入。(c)hfd年老(rharg)组的每周总食物摄入比hfd年老(载体)组减少约31%。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem;对于cd年轻(载体)组和hfd年老(载体)组,n=6,对于hfd年老(rharg)组,n=9。
[0069]
图37说明向图36中的喂食hfd的16个月大的dio雄性小鼠[hfd年老(rharg)组]施用n-abd094-rharg(seq id no:50)可以在治疗的10周内诱导显著的体重减轻,并且在剩余的治疗期中,体重保持在30g左右相对恒定。
[0070]
图38说明向图36中的喂食hfd的16个月大的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转胰岛素抵抗。(a,b)在(a)之前和rharg治疗后9周(b)进行胰岛素耐量试验(itt)。itt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0071]
图39说明向图36中的喂食hfd的16个月大的dio雄性小鼠[hfd(rharg)组]施用n-abd094-rharg(seq id no:50)可有效逆转受损的葡萄糖耐量。(a,b)在(a)之前和rharg治疗后12周(b)进行葡萄糖耐量试验(gtt)。gtt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0072]
图40说明当每周一次施用约600u n-abd094-rharg(seq id no:50)(rharg)持续21周时,喂食普通饲料的约17个月大(相当于人类大约50岁中期)的c57bl/6j雄性小鼠表现出重复7天间歇性禁食周期。(a)3组小鼠的食物摄入模式:喂食饲料并注射rharg的17个月大的小鼠[cd年老(rharg)组];喂食饲料并注射生理盐水(载体)的17个月大的小鼠[cd年老(载体)组];喂食饲料并注射载体的5个月大的雄性小鼠(相当于人类大约20岁中期)[cd年轻(载体)组]作为年轻对照组。(b)7天间歇性禁食周期中每天的平均食物摄入。(c)cd年老(rharg)组的每周总食物摄入比cd年老(载体)组减少约11%。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem;对于cd年轻(载体)组和cd年老(载体)组,n=7,对于hfd年老(rharg)组,n=9。
[0073]
图41说明向图40中的喂食饲料的17个月大的雄性小鼠[cd年老(rharg)组]施用n-abd094-rharg(seq id no:50)可以在治疗的8周内诱导体重从40g减轻到30g,并且在剩余的治疗期中,体重保持在该水平相对恒定。
[0074]
图42说明向图40中的喂食饲料的17个月大的雄性小鼠[cd年老(rharg)组]施用n-abd094-rharg(seq id no:50)可有效改善胰岛素敏感性。(a,b)在(a)之前和rharg治疗后13周(b)进行胰岛素耐量试验(itt)。itt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0075]
图43说明向图40中的喂食饲料的17个月大的雄性小鼠[cd年老(rharg)组]施用n-abd094-rharg(seq id no:50)可有效改善葡萄糖耐量。(a,b)在(a)之前和rharg治疗后15周(b)进行葡萄糖耐量试验(gtt)。gtt的结果表示为曲线下面积(auc)。**p《0.05,曼-惠特尼u检验;*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0076]
图44说明在21周治疗结束时,与图40中的喂食饲料并接受载体的年龄匹配的雄性小鼠[cd年老(载体)组]以及图40中的喂食饲料并从5个月大开始接受载体注射的10个月大(相当于人类大约30岁中期)的雄性小鼠[cd年轻(载体)组]相比,图40中的喂食饲料并从17个月大开始接受每周施用n-abd094-rharg(seq id no:50)的22个月大(相当于人类大约60岁中期)的雄性小鼠[cd年老(rharg)组]显示出肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪量的显著减少。cd年老(rharg)组的肝脏质量在所有3组小鼠中最低。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0077]
图45说明当每周一次施用约600u n-abd094-rharg(seq id no:50)(rharg)持续30周时,喂食普通饲料的约25个月大(相当于人类70岁)的c57bl/6j雄性小鼠表现出重复7天间歇性禁食周期。(a)3组小鼠的食物摄入模式:喂食饲料并注射rharg的25个月大的小鼠[cd年老(rharg)组];喂食饲料并注射生理盐水(载体)的25个月大的小鼠[cd非常老(载体)组];喂食饲料并注射载体的8个月大的雄性小鼠(相当于人类30岁出头)[cd中年(载体)组]
51样自噬活化激酶1(ulk1,一种自噬引发剂)的刺激,如通过蛋白质印迹测定的。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem,每组n=4。
[0084]
图52说明在n-abd094-rharg注射后7天间歇性禁食周期的第3天和第7天通过透射电子显微镜检查的肩胛间棕色脂肪组织(ibat)。(a)与用载体治疗的hfd喂食小鼠[hfd(载体)组](b)相比,每周一次施用600u n-abd094-rharg(seq id no:50)持续4周的喂食hfd的雄性小鼠[hfd(rharg)组]的bat显示脂滴大小显著减小,在第3天发生大量的自噬(包括溶酶体、自噬体和自噬性溶酶体的存在)以分解脂质(脂肪吞噬)。(c)在5000x放大倍率下检查,自噬体吞噬了脂滴并分解成微小的颗粒。(d)在第7天,自噬体和自噬性溶酶体数量截止第7天显著减少,但观察到线粒体数量增加。
[0085]
图53说明每周一次向预先存在hfd诱导的肥胖症的c57bl/6j雄性小鼠[hfd(rharg)组]施用600u n-abd094-rharg(seq id no:50)持续12周可有效逆转棕色脂肪组织(bat)变白,其中储存为大的单个小球(白色脂肪细胞的特征)的脂质变成多个小液滴中的脂质储存(棕色脂肪细胞的特征),这与通过tem观察到的诱导脂肪吞噬的发现一致。(a)新鲜肩胛间bat(上图)的代表性图像,显示用载体治疗的喂食hfd的dio小鼠[hfd(载体)组]的肩胛间bat增大,其中许多细胞在用苏木精和曙红染色的石蜡切片中表现出增大的单个脂质样小球(下图)。用rharg治疗的hfd喂食小鼠[hfd(rharg)组]的ibat显示器官大小的减小和组织学外观的恢复,类似于喂食饲料的载体治疗的对照小鼠[cd(载体)]。(b)3组小鼠的ibat的质量与a中所示的大小一致。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem,每组n=5。
[0086]
图54说明n-abd094-rharg(seq id no:50)在小鼠下丘脑神经元的原代培养物中诱导自噬。在用n-abd094-rharg(seq id no:50)治疗1、4、8和24小时(+)或没有治疗(-)的小鼠下丘脑神经元的原代培养物中,(a)相对于β-微管蛋白的lc3ii蛋白水平升高,和(b)相对于β-微管蛋白的p62蛋白水平降低。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,n=3-6。
[0087]
图55说明n-abd094-rharg(seq id no:50)在小鼠下丘脑神经元的原代培养物中诱导eif2α/atf4通路的活化和mtor通路的失活,如p70s6k1的磷酸化降低所证明的。在用n-abd094-rharg(seq id no:50)治疗1、4、8和24小时(+)或没有治疗(-)的小鼠下丘脑神经元的原代培养物中,(a)磷酸化真核翻译起始因子2a(eif2α)与总eif2α蛋白水平的比率,(b)相对于β-微管蛋白的atf4蛋白水平,和(c)磷酸化p70s6k1与总p70s6k1蛋白水平的比率。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,n=4-7。
[0088]
图56说明n-abd094-rharg(seq id no:50)在小鼠下丘脑神经元的原代培养物中诱导阿黑皮质素原pomc(pomc的活性形式)的糖基化。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,n=4-7。
[0089]
图57说明组合n-abd094-rharg(seq id no:50)和二甲双胍对诱导间歇性禁食和减少食物摄入的协同作用。当通过每周一次腹腔注射来施用生理盐水中的300u n-abd094-rharg(seq id no:50)(rharg)联合通过胃饲每日施用300mg/kg二甲双胍水溶液[hfd(rharg+met)组]持续9周时,预先存在通过从5周大开始喂食高脂饮食(hfd)12周诱导的肥胖症的c57bl/6j雄性小鼠表现出显著的重复7天间歇性禁食周期。(a)5组小鼠的食物摄入模式:hfd(rharg+met)组;每周注射一次300u rharg联合每日胃饲水的喂食hfd的dio小鼠
[hfd(rharg)组];每周注射一次生理盐水联合每日胃饲300mg/kg二甲双胍的喂食hfd的dio小鼠[hfd(met)组];每周注射一次生理盐水联合每日胃饲水的喂食hfd的小鼠[hfd(载体)组];喂食饲料并每周注射一次生理盐水联合每天胃饲水的小鼠[cd(载体)组]。(b)在第0天注射rharg或生理盐水后7天内每天的平均食物摄入。hfd(rharg+met)组表现出显著的7天间歇性禁食周期,而hfd(rharg)组也表现出食物摄入减少然后食物摄入增加的7天模式,但幅度小于hfd(rharg+met)组。(c)hfd(rharg+met)组的每周总食物摄入比hfd(载体)组减少31%。单独使用二甲双胍[hfd(met)]也导致每周食物摄入显著减少13%,而单独使用rharg[hfd(rharg)]与hfd(载体)组相比,没有诱导每周总食物摄入的显著变化。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=5。
[0090]
图58说明组合每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍对减轻体重的协同作用。图57中的用rharg和met联合治疗的喂食hfd的dio小鼠[hfd(rharg+met)组]显示在治疗5周内体重从52g显著降低至35g,然后在剩余的治疗期里,它们的体重保持相对恒定。相比之下,对rharg[hfd(rharg)组]或met[hfd(met)组]的单独治疗在9周的治疗期内只能防止体重进一步增加,但不会导致体重减轻。
[0091]
图59说明组合每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍对改善图57中的喂食hfd的dio雄性小鼠[hfd(rharg+met)组]的胰岛素敏感性和葡萄糖耐量的协同作用。(a)胰岛素耐量试验(itt)和(b)葡萄糖耐量试验(gtt)分别在治疗后6周和7周进行。itt和gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0092]
图60说明组合每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍对显著减少图57中的喂食hfd的dio雄性小鼠[hfd(rharg+met)组]的肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪垫质量的协同作用。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0093]
图61说明组合每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍对于在图57中的喂食hfd的dio雄性小鼠[hfd(rharg+met)组]中显著降低肝脏质量并逆转肝脂肪变性的协同作用。(a)肝脏质量。(b)新鲜全肝(上图)和肝脏切片中脂质的油红o染色的代表性图像显示,接受联合治疗的小鼠的肝脏质量显著减少,并且肝脏中的脂质清除是显著的。(c)肝脏中的甘油三酯浓度与油红o染色结果一致。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0094]
图62说明预先存在hfd诱导的肥胖症的喂食hfd的c57bl/6j雄性小鼠在透射电子显微镜下检查的肝细胞超微结构,该小鼠已经接受了每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍的3周联合治疗[hfd(rharg+met)组],或每周一次300u n-abd094-rharg(seq id no:50)[hfd(rharg)组]或每天300mg/kg二甲双胍[hfd(met)组]的单一治疗,或载体[hfd(载体)组]。在7天间歇性禁食周期的第3天(禁食阶段)检查的接受rharg和二甲双胍联合治疗的hfd喂食小鼠的肝细胞显示,与接受载体治疗的小鼠相比,脂滴大小显著减小,并且有大量的自噬体。然而,在7天间歇性禁食周期的第7天(再喂食阶段)检查时,自噬囊泡的数量大大减少。在第3天接受rharg单一治疗或单独二甲双胍治疗的hfd喂食小鼠的肝细胞显示脂滴大小减小,但很少见到自噬体。
[0095]
图63说明在接受联合治疗[hfd(rharg+met)组]的小鼠肝细胞中发生的大规模脂
肪吞噬,其特征在于形成自噬体,这些自噬体隔离大脂滴的多个部分以形成双膜囊泡,将脂滴分解成更小的大小。通过透射电子显微镜在5000x放大倍率下观察。
[0096]
图64说明接受联合治疗[hfd(rharg+met)组]的小鼠肝细胞发生巨自噬,其特征在于含有多种细胞质成分的大型自噬体与溶酶体融合,进一步形成自噬性溶酶体。通过透射电子显微镜在5000x放大倍率下观察。
[0097]
图65说明每周一次300u n-abd094-rharg(seq id no:50)和每天300mg/kg二甲双胍的联合治疗抑制了预先存在hfd诱导的肥胖症的雄性小鼠[hfd(rharg+met)组]的肝脏和肩胛间棕色脂肪组织中mtorc1的磷酸化。mtorc1是自噬的主要调节因子,抑制mtorc1可触发细胞自噬。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem,每组n=5。
[0098]
图66说明组合n-abd094-rharg(seq id no:50)和全反式视黄酸(ra)对诱导间歇性禁食和减少食物摄入的协同作用。当通过每周一次腹腔注射来施用生理盐水中的200u n-abd094-rharg(seq id no:50)(rharg)联合通过胃饲每日施用0.33mg在花生油中的全反式视黄酸(ra)[hfd(rharg+ra)组]持续10周时,预先存在通过从5周大开始喂食高脂饮食(hfd)12周诱导的肥胖症的c57bl/6j雄性小鼠表现出显著的重复7天间歇性禁食周期。(a)5组小鼠的食物摄入模式:hfd(rharg+ra)组;每周注射一次200u rharg联合每日胃饲花生油的喂食hfd的dio小鼠[hfd(rharg)组];每周注射一次生理盐水联合每日胃饲0.33mg ra的喂食hfd的dio小鼠[hfd(ra)组];每周注射一次生理盐水联合每日胃饲花生油的喂食hfd的小鼠[hfd(载体)组];喂食饲料并每周注射一次生理盐水联合每天胃饲花生油的小鼠[cd(载体)组]用作瘦对照组。(b)在第0天注射rharg或生理盐水后7天内每天的平均食物摄入。hfd(rharg+ra)组表现出显著的7天间歇性禁食周期,而hfd(rharg)组也表现出食物摄入减少然后食物摄入增加的7天模式,但幅度小于hfd(rharg+ra)组。(c)hfd(rharg+ra)组的每周总食物摄入比hfd(载体)组减少41%。与hfd(载体)组相比,单独的ra[hfd(ra)组]或单独的rharg[hfd(rharg)组]导致每周总食物摄入略有下降,但不显著。*p《0.05,曼-惠特尼u检验。数据表示为平均值
±
sem,每组n=6。
[0099]
图67说明组合每周一次200u n-abd094-rharg(seq id no:50)和每天0.33mg ra对减轻体重的协同作用。图66中的用rharg和ra联合治疗的喂食hfd的dio小鼠[hfd(rharg+ra)组]显示在治疗7周内体重从53g显著降低至30g。相比之下,对rharg[hfd(rharg)组]或ra[hfd(ra)组]的单独治疗在10周的治疗期内只能防止体重进一步增加,但不会导致体重减轻。
[0100]
图68说明组合每周一次200u n-abd094-rharg(seq id no:50)和每天0.33mg ra对改善图66中的喂食hfd的dio雄性小鼠[hfd(rharg+ra)组]的胰岛素敏感性和葡萄糖耐量的协同作用。(a)胰岛素耐量试验(itt)和(b)葡萄糖耐量试验(gtt)分别在治疗后6周和7周进行。itt和gtt的结果表示为曲线下面积(auc)。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0101]
图69说明组合每周一次200u n-abd094-rharg(seq id no:50)和每天0.33mg ra对显著减少图66中的喂食hfd的dio雄性小鼠[hfd(rharg+ra)组]的肾周(内脏)和腹股沟(皮下)白色脂肪组织(wat)和肩胛间棕色脂肪组织(bat)的脂肪垫质量的协同作用。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0102]
图70说明组合每周一次200u n-abd094-rharg(seq id no:50)和每天0.33mg ra对于在图66中的喂食hfd的dio雄性小鼠[hfd(rharg+ra)组]中显著降低肝脏质量并逆转肝脂肪变性的协同作用。(a)肝脏质量。(b)新鲜全肝(上图)和肝脏切片中脂质的油红o染色的代表性图像显示,接受联合治疗的小鼠的肝脏质量显著减少,并且肝脏中的脂质清除是显著的。(c)肝脏中的甘油三酯浓度与油红o染色结果一致。*p《0.05,单因素方差分析,然后进行bonferroni检验。数据表示为平均值
±
sem。
[0103]
图71说明预先存在hfd诱导的肥胖症的喂食hfd的c57bl/6j雄性小鼠在透射电子显微镜下检查的肝细胞超微结构,该小鼠已经接受了每周一次200u n-abd094-rharg(seq id no:50)和每天0.33mg ra的3周联合治疗[hfd(rharg+ra)组],或每周一次200u n-abd094-rharg(seq id no:50)[hfd(rharg)组]或每天0.33mg ra[hfd(ra)组]的单一治疗,或载体[hfd(载体)组]。大量大液滴积聚在用载体治疗的hfd喂食小鼠[hfd(载体)组]的肝细胞的细胞质中。在7天间歇性禁食周期的第3天(禁食期)检查的接受rharg和ra联合治疗的hfd喂食小鼠的肝细胞显示脂滴大小显著减小,并且有大量的自噬体。然而,在7天间歇性禁食周期的第7天(再喂食阶段)检查时,自噬囊泡的数量大大减少。在第3天接受rharg单一治疗或单独ra治疗的hfd喂食小鼠的肝细胞显示脂滴大小减小,但很少见到自噬体。
[0104]
图72说明在接受联合治疗[hfd(rharg+ra)组]的小鼠肝细胞中发生的大规模脂肪吞噬,其特征在于形成自噬体,这些自噬体隔离大脂滴的多个部分以形成双膜囊泡,将脂滴分解成更小的大小。通过透射电子显微镜在5000x放大倍率下观察。
具体实施方式
[0105]
术语的定义
[0106]
本文使用的术语的定义旨在纳入生物技术领域中每个术语公认的当前最先进的定义。在适当的地方,提供了示例。除非在特定情况下单独或作为更大组的一部分另有限制,否则定义适用于在整个说明书中使用的术语。
[0107]
如本文所用,术语“半衰期”或“1/2-寿命”是指药剂(例如本文所述的融合蛋白或精氨酸消耗剂)的浓度在体外或体内(例如,在注射到哺乳动物体内之后)下降一半所需的时间。在某些情况下,注射后血浆精氨酸的浓度在本文中用作药剂半衰期的替代指标。在这些情况下,术语“治疗持续时间”用于指规定剂量的精氨酸消耗剂能够将精氨酸的血浆浓度维持在观察到所需治疗效果的规定阈值浓度以下的时间长度。在某些实施方案中,血浆精氨酸的阈值浓度低于50μm、低于40μm、低于30μm、低于20μm、低于10μm、低于5μm、低于3μm或处在低于常规分析仪器的检测限的浓度。例如,在注射本文所述的精氨酸分解代谢酶之后7天,血浆精氨酸消耗至低于biochrom 30氨基酸分析仪的检测限(检测限为3μm)的浓度表明治疗持续时间为7天并且半衰期为例如大约7天。
[0108]
如本文所用,术语“连接”或“连接的”是指通过键或非键相互作用连接或结合以将两种或更多种化合物保持在一起,这包括直接连接或间接连接,使得例如第一多肽直接与第二多肽或其他分子结合,以及其中一种或多种中间体化合物(例如,接头),例如多肽,设置在第一多肽和第二多肽或其他分子之间的实施方案。
[0109]
如本文所用,术语“蛋白质”或“多肽”表示由两个或更多个氨基酸单体和/或其类似物构成的有机聚合物。术语“多肽”包括任何长度的氨基酸聚合物,包括全长蛋白质和肽,
以及它们的类似物和片段。三个或更多氨基酸的多肽也称为寡肽。如本文所用,术语“氨基酸”、“氨基酸单体”或“氨基酸残基”是指二十种天然存在的氨基酸中的任一种,包括具有非天然侧链的合成氨基酸并且包括d和l旋光异构体。术语“氨基酸类似物”是指一种氨基酸,其中一个或多个单独的原子被替换为不同的原子、同位素或不同的官能团,但在其他方面与其天然氨基酸类似物相同。
[0110]
如本文所用,术语“非天然氨基酸”是指不是20种常见的天然存在的氨基酸、硒代半胱氨酸或吡咯赖氨酸之一的任何氨基酸、修饰氨基酸和/或氨基酸类似物。
[0111]
如本文所用,术语“融合蛋白”是指包含具有不同来源的蛋白质或功能性蛋白质片段(例如,精氨酸酶或其变体)的嵌合蛋白质,这些蛋白质或功能性蛋白质片段例如通过酰胺、酯、脲、氨基甲酸酯、醚和/或二硫键共价连接。
[0112]
如本文所用,术语“变体”是指与参考核酸或多肽不同但保留其基本特性的多核苷酸或核酸。通常,变体总体上非常相似,并且在许多区域中与参考核酸或多肽相同。
[0113]
例如,变体可以包含具有至少一个保守氨基酸取代的亲本多肽序列的氨基酸序列。或者或另外,变体可以包含具有至少一个非保守氨基酸取代的亲本多肽序列的氨基酸序列。在这种情况下,优选非保守氨基酸取代不干扰或不抑制功能变体的生物活性。非保守氨基酸取代可增强变体的生物活性,使得变体的生物活性与亲本多肽相比有所增加。
[0114]
当关于多肽使用时,术语“功能片段”是指标的多肽的任何部分或部件,该部分或部件保留了它作为其一部分的多肽(亲本多肽)的生物活性。功能片段可以是包含作为其一部分的多肽的连续氨基酸的任何片段,条件是功能片段仍表现出亲本多肽的至少40%、50%、60%、70%、80%、90%、95%或99%的生物活性,或具有与亲本多肽基本相同或甚至更高的生物活性。关于亲本多肽,功能片段可包含例如约10%、20%、25%、30%、40%、50%、60%、70%、80%、90%、95%、98%或更多的亲本多肽。
[0115]
功能片段可以在氨基末端或羧基末端,或在两个末端包含额外的氨基酸,例如在亲本多肽的氨基酸序列中未发现的氨基酸。
[0116]
所述多肽的氨基酸取代可以是保守氨基酸取代。保守氨基酸取代是本领域已知的,并且包括其中具有某些物理和/或化学特性的一种氨基酸被替换为具有相同或相似化学或物理特性的另一种氨基酸的氨基酸取代。例如,保守氨基酸取代可以是用酸性/带负电的极性氨基酸取代另一种酸性/带负电的极性氨基酸(例如,asp或glu),用具有非极性侧链的氨基酸取代另一种具有非极性侧链的氨基酸(例如,ala、gly、val、ile、leu、met、phe、pro、trp、cys、val等),用碱性/带正电的极性氨基酸取代另一种碱性/带正电的极性氨基酸(例如lys、his、arg等),用具有极性侧链的不带电氨基酸取代另一种具有极性侧链的不带电氨基酸(例如asn、gln、ser、thr、tyr等),用具有β支链侧链的氨基酸取代另一种具有β支链侧链的氨基酸(例如,ile、thr和val),用具有芳香侧链的氨基酸取代另一种具有芳香侧链的氨基酸(例如,his、phe、trp和tyr)等。
[0117]
当关于多肽或多核苷酸序列使用时,术语“百分比同源性”和“百分比序列同一性”在本文中可互换使用以指代多核苷酸和多肽之间的比较,并且通过在比较窗口上比较两个最佳比对序列来确定,其中与用于两个序列的最佳比对的参考序列(其不包含添加或缺失)相比,比较窗口中的多核苷酸或多肽序列的部分可以包含添加或缺失(即,空位)。百分比是如下计算的:确定在两个序列中存在相同核酸碱基或氨基酸残基的位置的数目以得到匹配
位置数目,将匹配位置数目除以比较窗口中的总位置数目,和将结果乘以100以得到序列同一性百分比。使用本领域已知的各种序列比较算法和程序中的任何一种来评估同源性。这些算法和程序包括但决不限于tblastn、blastp、fasta、tfasta和clustalw[pearson和lipman,1988,proc.natl.acad.sci.usa85(8):2444-2448;altschul等人,1990,j.mol.biol.215(3):403-410;thompson等人,1994,nucleic acids res.22(2):4673-4680;higgins等人1996,methods enzymol.266:383-402;altschul等人,1990,j.mol.biol.215(3):403-410;altschul等人,1993,nature genetics 3:266-272]。在某些实施方案中,使用本领域众所周知的基本局部比对搜索工具(“blast”)评估蛋白质和核酸序列同源性(参见例如,karlin和altschul,1990,proc.natl.acad.sci.usa 87:2267-2268;altschul等人,1990,j.mol.biol.215:403-410;altschul等人,1993,nature genetics 3:266-272;altschul等人,1997,nuc.acids res.25:3389-3402)。
[0118]
如本文所用,术语“治疗(treat)”、“治疗(treating)”、“治疗(treatment)”等是指减少或改善病症/疾病和/或与其相关的症状。应当理解,尽管不排除,治疗病症或病状不需要完全消除病症、病状或与其相关的症状。在某些实施方案中,治疗包括预防病症或病状,和/或与其相关的症状。如本文所用,术语“预防(prevention)”或“预防(prevent)”是指抑制或至少延迟病症、病状或与其相关的症状的发展的任何作用。预防可以包括一级、二级和三级预防水平,其中:a)一级预防避免了疾病的发展;b)二级预防活动是针对早期疾病治疗,从而增加预防疾病发展和症状出现的干预机会;和c)三级预防通过恢复功能和减少与疾病相关的并发症来减少已经确定的疾病的负面影响。
[0119]
如本文所用,术语“共同施用(co-administration)”和“共同施用(co-administering)”是指同时施用(同时施用两种或更多种治疗剂)和随时间变化的施用(一种或多种治疗剂的施用时间不同于另外一种或多种治疗剂的施用时间)。在某些实施方案中,治疗剂在一定程度上同时存在于患者体内。
[0120]
如本文所用,术语“分解代谢(catabolism)”或“分解代谢(catabolic)”是指分子化学反应成其他分子,例如更小的分子。例如,精氨酸分解代谢酶是指能够与精氨酸反应从而将其转化为其他分子例如鸟氨酸、瓜氨酸和胍丁胺的任何酶。
[0121]
如本文所用,术语“受试者”是指任何动物(例如哺乳动物),包括但不限于人类、非人类灵长类动物、犬科动物、猫科动物和啮齿动物。
[0122]
本文提供了一种在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法,包括向受试者施用治疗有效量的精氨酸消耗剂的步骤。
[0123]
近年来,大量研究致力于研究间歇性禁食和/或调节自噬的影响,以及诱导间歇性禁食和/或调节自噬可能带来的众多健康益处。例如,已经发现间歇性禁食和/或调节自噬可以提高受试者的寿命(adv nutr.2019年11月1日;10(supplement_4):s340-s350.doi:10.1093/advances/nmz079;aging cell.2019年2月;18(1):e12843.doi:10.1111/acel.12843.epub 2018年10月17日);治疗心血管疾病(circ res.2019年3月15日;124(6):952-965.doi:10.1161/circresaha.118.313352.);治疗炎症性肠病(cell rep.2019年3月5日;26(10):2704-2719.e6.doi:10.1016/j.celrep.2019.02.019.);治疗糖尿病(cell.2017年2月23日;168(5):775-788.e12.doi:10.1016/j.cell.2017.01.040.);治疗衰老、癌症和心血管疾病(sci transl med.2017年2月15日;9(377).pii:eaai8700.doi:
10.1126/scitranslmed.aai8700);治疗自身免疫性疾病(mol cell endocrinol.2017年11月5日;455:4-12.doi:10.1016/j.mce.2017.01.042.epub 2017年1月28日);治疗与年龄相关的病症,包括糖尿病、心血管疾病、癌症和神经系统病症,如阿尔茨海默病、帕金森病和中风(ageing res rev.2017年10月;39:46-58.doi:10.1016/j.arr.2016.10.005.epub 2016年10月31日);治疗自身免疫和多发性硬化症症状(cell rep.2016年6月7日;15(10):2136-2146.doi:10.1016/j.celrep.2016.05.009.epub 2016年5月26日);改善认知、表现、健康寿命,降低内脏脂肪,减少癌症发病率和皮肤损伤,恢复免疫系统和延缓骨矿物质密度损失(cell metab.2015年7月7日;22(1):86-99.doi:10.1016/j.cmet.2015.05.012.epub 2015年6月18日);和减少肥胖、高血压、哮喘和类风湿性关节炎(cell metab.2014年2月4日;19(2):181-92.doi:10.1016/j.cmet.2013.12.008.epub 2014年1月16日),所有这些都通过引用并入本文。
[0124]
研究还表明,间歇性禁食可用于治疗癌症(nat rev cancer.2018年11月;18(11):707-719.doi:10.1038/s41568-018-0061-0;recent results cancer res.2016;207:241-66.doi:10.1007/978-3-319-42118-6_12;cancer cell.2016年7月11日;30(1):136-146.doi:10.1016/j.ccell.2016.06.005;mol cell oncol.2015年12月10日;3(3):e1117701.doi:10.1080/23723556.2015.1117701.ecollection 2016年5月;oncotarget.2015年5月20日;6(14):11820-32;plos one.2012;7(9):e44603.doi:10.1371/journal.pone.0044603.epub 2012年9月11日;drug resist updat.2012年2月-4月;15(1-2):114-22.doi:10.1016/j.drup.2012.01.004.epub 2012年3月4日;sci transl med.2012年3月7日;4(124):124ra27.doi:10.1126/scitranslmed.3003293.epub 2012年2月8日;oncogene.2011年7月28日;30(30):3305-16.doi:10.1038/onc.2011.91.epub 2011年4月25日;和cell cycle.2010年11月15日;9(22):4474-6.epub 2010年11月15日),所有这些都通过引用并入本文。
[0125]
在某些实施方案中,本文所述的精氨酸消耗剂可用于治疗诱导间歇性禁食和/或调节自噬对其具有有益效果的任何疾病或健康状况。在某些实施方案中,诱导间歇性禁食和/或调节自噬对其具有有益效果的疾病或健康状况是上述疾病或健康状况中的任何一种或多种。
[0126]
在某些实施方案中,本文提供了一种方法,该方法包括施用治疗有效的精氨酸消耗剂以诱导间歇性禁食和/或调节自噬以提高寿命和/或减轻衰老症状或预防与年龄相关的疾病的步骤。
[0127]
在某些实施方案中,本文提供了一种方法,该方法包括以下步骤:施用治疗有效的精氨酸消耗剂以诱导间歇性禁食和/或调节自噬以促进蛋白质聚集体的清除,并预防和/或治疗神经退行性疾病,例如阿尔茨海默病。
[0128]
在某些实施方案中,本文提供了一种方法,该方法包括施用治疗有效的精氨酸消耗剂以诱导间歇性禁食和/或调节自噬以治疗炎症和包括类风湿性关节炎的相关疾病的步骤。
[0129]
在某些实施方案中,本文提供了一种方法,该方法包括施用治疗有效的精氨酸消耗剂以诱导间歇性禁食和/或调节自噬以治疗与自噬缺陷相关的疾病的步骤。
[0130]
在某些实施方案中,本文提供了一种方法,该方法包括以下步骤:施用治疗有效的
精氨酸消耗剂以诱导间歇性禁食和/或调节自噬以促进细胞内病原体的清除以治疗细菌和病毒感染。
[0131]
精氨酸消耗剂可以是本领域已知的能够降低受试者中精氨酸的血浆和/或细胞水平的任何精氨酸消耗剂。精氨酸消耗剂可以是小分子或蛋白质。
[0132]
蛋白质可以是融合蛋白质和/或化学修饰蛋白质,例如聚乙二醇化蛋白质。示例性蛋白质包括能够催化精氨酸分解代谢成其他产物的蛋白质,例如具有精氨酸酶、精氨酸脱亚氨酶、精氨酸脱羧酶或精氨酸2单加氧酶活性的蛋白质。
[0133]
精氨酸酶可以是本领域已知的任何精氨酸酶,例如由细菌、真菌、鱼、人、牛、猪、兔、啮齿动物、灵长类动物、绵羊和山羊产生的那些。例如,热溶芽孢杆菌(bacillus caldovelox)精氨酸酶、嗜热栖热菌(thermus thermophilus)精氨酸酶、山羊(capra hircus)精氨酸酶i、裸鼹鼠(heterocephalus glaber)精氨酸酶i、牛(bos taurus)精氨酸酶i、野猪(sus scrofa)精氨酸酶i、香鱼(plecoglossus altivelis)精氨酸酶i、大西洋鲑(salmo salar)精氨酸酶i、虹鳟(oncorhynchus mykiss)精氨酸酶i、胡瓜鱼(osmerus mordax)精氨酸酶i、三角帆蚌(hyriopsis cumingii)精氨酸酶i、褐家鼠(rattus norvegicus)精氨酸酶i、小家鼠(mus musculus)精氨酸酶i、智人(homo sapiens)(人)精氨酸酶i、黑猩猩(pan troglodytes)精氨酸酶i、家兔(oryctolagus cuniculus)精氨酸酶i、褐家鼠精氨酸酶ii、小家鼠精氨酸酶ii、智人(人)精氨酸酶ii、牛精氨酸酶ii、裸鼹鼠精氨酸酶ii、黑猩猩精氨酸酶ii、家兔精氨酸酶ii、代尔夫特菌(delftia)精氨酸酶、凝结芽孢杆菌(bacillus coagulans)精氨酸酶、光养赫夫勒氏菌(hoeflea phototrophica)精氨酸酶和卡氏玫瑰弯菌(roseiflexus castenholzii)精氨酸酶。其他实例包括来自甲醇芽孢杆菌(bacillus methanolicus)、芽孢杆菌nrrl b-14911种(bacillus sp.nrrl b-14911)、东海游动球菌(planococcus donghaensis)、树形类芽孢杆菌(paenibacillus dendritiformis)、侧链孢属(desmospora sp.)、苔原沼泽甲基杆菌(methylobacter tundripaludum)、寡养单胞菌属(stenotrophomonas sp.)、产果聚糖微杆菌(microbacterium laevaniformans)、上野卟啉菌(porphyromonas uenonis)、农杆菌属(agrobacterium sp.)、北极十八杆菌(octadecabacter arcticus)、根癌农杆菌(agrobacterium tumefaciens)、好热黄无氧芽孢菌(anoxybacillus flavithermus)、短小芽孢杆菌(bacillus pumilus)、热葡糖苷酶地芽孢杆菌(geobacillus thermoglucosidasius)、热葡糖苷酶地芽孢杆菌(geobacillus thermoglucosidans)、侧孢短芽孢杆菌(brevibacillus laterosporus)、瘤胃脱硫肠状菌(desulfotomaculum ruminis)、嗜热地芽孢杆菌(geobacillus kaustophilus)、嗜热地芽孢杆菌(geobacillus thermoleovorans)、嗜热反硝化地芽孢杆菌(geobacillus thermodenitrificans)、金黄色葡萄球菌(staphylococcus aureus)、嗜盐古菌(halophilic archaeon)dl31、盐惰菌属(halopigerxanaduensis)、马加迪钠白菌(natrialba magadii)、恶性疟原虫(plasmodium falciparum)、幽门螺旋杆菌(helicobacter pylori)等的精氨酸酶。
[0134]
精氨酸脱亚氨酶可以是本领域已知的任何精氨酸脱亚氨酶,例如由支原体、乳球菌属、假单胞菌属、链球菌属、埃希氏菌属、分枝杆菌属或芽孢杆菌属微生物产生的那些。示例性精氨酸脱亚氨酶包括但不限于由人型支原体(mycoplasma hominis)、精氨酸支原体(mycoplasma arginini)、关节炎支原体(mycoplasma arthritidis)、产气荚膜梭菌
(clostridium perfringens)、地衣芽孢杆菌(bacillus licheniformis)、伯氏疏螺旋体(borrelia burgdorferi)、阿氏疏螺旋体(borrelia afzellii)、粪肠球菌(enterococcus faecalis)、乳酸乳球菌(lactococcus lactis)、蜡状芽孢杆菌(bacillus cereus)、化脓性链球菌(streptococcus pyogenes)、肺炎链球菌(steptococcus pneumoniae)、清酒乳杆菌(lactobacillus sake)、肠贾第虫(giardia intestinalis)、结核分枝杆菌(mycobacterium tuberculosis)、杀香鱼假单胞菌(pseudomonas plecoglossicida)、恶臭假单胞菌(pseudomonas putida)、铜绿假单胞菌(pseudomonas aeruginosa)等。
[0135]
精氨酸脱羧酶可以是本领域已知的任何精氨酸脱羧酶,例如由大肠杆菌、鼠伤寒沙门氏菌(salmonella typhimurium)、肺炎衣原体(chlamydophila pneumoniae)、詹氏甲烷球菌(methanocaldococcus jannaschii)、绿草履虫小球藻病毒1(paramecium bursaria chlorella virus 1)、创伤弧菌(vibrio vulnificus)yj016、空肠弯曲菌亚种(campylobacter jejuni subsp.)、克氏锥虫(trypanosoma cruzi)、硫磺矿硫化叶菌(sulfolobus solfataricus)、地衣芽孢杆菌(bacillus licheniformis)、蜡状芽孢杆菌(bacillus cereus)、番木瓜(carica papaya)、烟草(nicotianatobacum)、大豆(glycine max)、百脉根(lotus coniculata)、创伤弧菌、霍乱弧菌(vibrio cholerae)、小家鼠、热袍菌(thermotoga)、褐家鼠、智人、牛、野猪、嗜热栖热菌(thermus thermophiles)、帕瓦蒂河栖热菌(thermus parvatiensis)、水生栖热菌(thermus aquaticus)、嗜热栖热菌(thermus thermophilus)、冰岛栖热菌(thermus islandicus)、拟南芥(arabidopsis thaliana)、燕麦(avena sativa)等产生的那些。
[0136]
精氨酸2-单加氧酶可以是本领域已知的任何精氨酸2-单加氧酶,例如由球形节杆菌(arthrobacter globiformis)ifo 12137、简单节杆菌(arthrobacter simplex)ifo 12069、黄色短杆菌(brevibacterium helvolum)ifo 12073、同性恋螺杆菌(helicobacter cinaedi)ccug 18818、灰色链霉菌(streptomyces griseus)等产生的那些。
[0137]
精氨酸脱羧酶、精氨酸脱亚氨酶、精氨酸2-单加氧酶和精氨酸酶可以是全蛋白质或其功能片段和/或变体。可以修饰精氨酸脱羧酶、精氨酸脱亚氨酶、精氨酸2-单加氧酶和精氨酸酶以改善它们的药代动力学性质,例如通过将蛋白质或其功能片段和/或变体与人血清白蛋白、白蛋白结合结构域、免疫球蛋白的fc区、peg基团或其组合融合。
[0138]
本文所述的精氨酸分解代谢酶可被改造以包括酶上可选择性连接peg的特定位点。选择的聚乙二醇化位点优选位于远离酶活性位点的位点,并且通常暴露于溶剂以允许与聚乙二醇化试剂反应。
[0139]
例如,可以产生cys
45-人精氨酸酶i(hai)和cys
161-热溶芽孢杆菌精氨酸酶(bca)以与硫醇特异性peg分子进行反应。修饰的精氨酸酶的单个游离半胱氨酸残基与连接至peg化合物的马来酰亚胺基团(mal)之间的缀合可形成peg化合物与修饰的精氨酸酶的游离半胱氨酸之间的共价键。seq id no:102和104包括为cys
45
定点聚乙二醇化而设计的突变体(c168s/c303s),因此可以任选地被聚乙二醇化。seq id no:89还包括为cys
161
定点聚乙二醇化而设计的突变体(s161c),因此可以任选地被聚乙二醇化。
[0140]
在某些实施方案中,精氨酸酶可以包含seq id no:101、seq id no:102、seq id no:103或seq id no:104,其中seq id no:102和seq id no:104任选地包含聚乙二醇基团(peg)。
[0141]
本领域已知的任何聚乙二醇化试剂可用于将peg共价连接至本文所述的精氨酸分解代谢酶。示例性的聚乙二醇化试剂包括但不限于mpeg-ald(甲氧基聚乙二醇-丙醛);mpeg-mal(甲氧基聚乙二醇-马来酰亚胺);mpeg-nhs(甲氧基聚乙二醇-n-羟基-琥珀酰亚胺);mpeg-spa(甲氧基聚乙二醇-琥珀酰亚胺丙酸酯);和mpeg-cn(甲氧基聚乙二醇-三聚氯氰)。
[0142]
peg基团可具有约5,000至约20,000amu、约5,000至约15,000amu、约5,000至约12,000amu、约7,000至约12,000amu或约7,000至约10,000amu的分子量。在某些实施方案中,peg基团具有约2,000amu至10,000amu的分子量。在某些实施方案中,peg基团是peg4,000、peg5,000、peg6,000或peg7,000。
[0143]
peg基团可以直接共价连接到精氨酸酶或通过接头连接到精氨酸酶。在某些实施方案中,精氨酸酶通过丙酸接头共价连接到peg。在其他实施方案中,精氨酸酶通过c2-c10、c2-c9、c2-c8、c2-c7、c2-c6、c2-c5或c2-c4直链或支链羧酸接头连接到peg。
[0144]
在某些实施方案中,本文提供的融合蛋白包括精氨酸酶多肽。精氨酸酶多肽可以源自任何表达精氨酸酶的生物体所表达的精氨酸酶蛋白。示例性的精氨酸酶包括由细菌(例如杆菌、农杆菌、蓝细菌和分枝杆菌)以及哺乳动物(例如牛、猪、绵羊、山羊、啮齿动物和人)产生的精氨酸酶。当精氨酸酶多肽源自人精氨酸酶时,它可以是精氨酸酶1型(arg1)或精氨酸酶2型(arg2)。
[0145]
精氨酸酶多肽可以包含全长精氨酸酶多肽或其功能片段和/或变体。
[0146]
精氨酸酶是一种含锰的酶。如实施例32所示,当本文所述的融合蛋白中存在的一个或多个锰离子被一个或多个二价阳离子金属例如co
2+
或ni
2+
取代时,融合蛋白的催化活性会增加。因此,在某些实施方案中,本文所述的融合蛋白包含一种或多种除锰之外的二价金属,例如co
2+
或ni
2+
。在某些实施方案中,融合蛋白包含一种或多种选自co
2+
和ni
2+
的金属。在某些实施方案中,融合蛋白包含两个co
2+
离子或两个ni
2+
离子。在其他实施方案中,融合蛋白包含两个mn
2+
离子。
[0147]
在某些实施方案中,精氨酸酶多肽是野生型人arg1。在某些实施方案中,精氨酸酶多肽包含与seq id no:69具有至少95%序列同源性的序列。例如,精氨酸酶多肽可以包含与seq id no:69具有至少96%、97%、98%、99%、99.1%、99.4%或99.7%同源性的多肽序列。在某些实施方案中,精氨酸酶多肽的序列可以与seq id no:69相差1、2、3、4、5、6、7、8、9、10、15、20、25或30个氨基酸修饰(例如插入、取代、缺失等)。在某些实施方案中,精氨酸酶多肽包含具有保守氨基酸置换、非保守氨基酸置换或其组合的多肽。
[0148]
在某些实施方案中,精氨酸酶多肽是热溶芽孢杆菌精氨酸酶(bca)。在某些实施方案中,精氨酸酶多肽包含与seq id no:70具有至少95%序列同源性的序列。例如,精氨酸酶多肽可以包含与seq id no:70具有至少96%、97%、98%、99%、99.3%或99.7%同源性的多肽序列。在某些实施方案中,精氨酸酶多肽的序列可以与seq id no:70相差1、2、3、4、5、6、7、8、9、10、15、20、25或30个氨基酸修饰(例如插入、取代、缺失等)。在某些实施方案中,精氨酸酶多肽包含具有保守氨基酸置换、非保守氨基酸置换或其组合的多肽。
[0149]
在某些实施方案中,精氨酸酶多肽是bca,其中丝氨酸161被半胱氨酸置换,如seq id no:71和seq id no:72所示。用半胱氨酸取代丝氨酸允许化学部分的定点掺入,这可以进一步改善融合蛋白的性质。例如,半胱氨酸161的侧链可以与适当活化的peg部分反应,从
drug deliv rev.2013;65:1357
–
69]。肽接头的两种主要类型是:(i)柔性接头(例如,(g4s)n,其中n=1-4);(ii)刚性接头,例如α-螺旋接头[a(eaaak)na]x(其中n=2
–
4且x=1或2)和xpn(其中x为a、k或e)。对于柔性接头,一个优点是可能需要柔性以获得分子的生物活性部分相对于其同源受体的正确取向。然而,柔性接头不会在融合伴侣和生物活性蛋白之间留出很大的空间。另一方面,刚性接头提供更多空间但缺乏灵活性。在ifn-α2b-hsa融合蛋白的情况下,与天然ifn-α2b相比,柔性接头产生约39%的活性,而刚性xp接头和α-螺旋接头分别产生天然ifn-α2b的68%和115%的活性[prot exp purif.2008;61:73
–
7]。
[0156]
某些接头可能对融合蛋白特性产生负面影响。例如,粒细胞集落刺激因子(g-csf)与转铁蛋白(tf)融合。使用短亮氨酸-谷氨酸(le)接头仅产生天然g-csf的约10%的活性。与g-csf-le-tf的活性相比,插入(g4s)3或α-螺旋[a(eaaak)na]m(n=2
–
4,m=1或2)接头显著增加了融合蛋白的活性。用接头(a(eaaak)4alea-(eaaak)4a)构建的融合蛋白产生接近天然g-csf的生物活性[pharm res.2006;23:2116
–
21]。在另一个实例中,虽然fc部分的c末端可以通过肽键直接连接到ifn-β部分的n末端,但gillies等人[us 7,670,595 b2]另外通过接头肽连接fc部分和ifn-β部分。接头肽位于fc部分的c末端和成熟ifn-β部分的n末端之间。在这种情况下,接头肽优选由丝氨酸和甘氨酸残基构成,例如氨基酸序列g4sg4sg3sg。所有这些发现都证明了测试接头技术对于融合蛋白研究和开发项目的成功的重要性。
[0157]
使用不同的方法,已经证明了融合位置对活性的重要性[curr pharmaceut biotechnol.2014;15:856
–
63]。脑利钠肽(bnp)以各种形式融合到hsa的n或c末端。结果表明,均与hsa的n末端融合的bnp-hsa、bnp2-hsa(bnp的两个拷贝)和bnp4-hsa均无活性。然而,与hsa的c末端融合的hsa-bnp2与天然bnp一样具有活性且是持久的。
[0158]
这些实例证明了在融合蛋白的先导优化中做出重大努力以通过接头工程、生物活性蛋白或肽相对于半衰期延长模块的位置或这两者来优化活性的重要性。重要的是,本发明使用一种新的abd融合方法将abd和精氨酸酶连接在一起,从而可以保留精氨酸酶活性。成功生成了稳定且可溶的精氨酸酶-abd融合分子,它们能够以ph依赖性方式与fcrn结合,从而实现有效的内体再循环。例如,本公开令人惊讶地发现,对于与abd融合的rharg,该蛋白质在循环中的终末半衰期在小鼠中从几分钟显著增加至4天。总之,多种不同的方法可用于微调精氨酸酶的药代动力学特性,从而确保针对不同疾病状况在循环中的适当停留时间。开发任何治疗性蛋白质的另一个关键问题是尽量减少免疫原性以避免副作用。因此,观察到的人精氨酸酶的低免疫原性和abd(例如abd094)的成功去免疫化是重要的,abd在本公开中也被用作基因融合伴侣以延长体内半衰期。
[0159]
白蛋白结合蛋白是在由革兰氏阳性细菌表达的各种表面蛋白中发现的三螺旋蛋白结构域。源自链球菌蛋白g的白蛋白结合蛋白具有214个氨基酸,并包含三个白蛋白结合结构域(abd1-3),它们用于与人血清白蛋白结合并逃避宿主的免疫系统。abd3对应于46个氨基酸的序列,该序列已被证明与人血清白蛋白结合,并且已经成为人血清白蛋白的许多研究和亲和力成熟的主题,以开发具有不同特性(例如结合亲和力和结合选择性)的abd多肽。此类研究已经产生了大量具有广泛不同特性的abd多肽。
[0160]
在其他细菌中发现白蛋白结合蛋白。例如,天然存在的白蛋白结合蛋白包括来自革兰氏阳性菌的某些表面蛋白,例如链球菌m蛋白(例如m1/emml、m3 emm3、m12/emml2、emml55/emm55、emm49/emml49和蛋白h)、链球菌蛋白g、mag和zag,以及来自某些大芬戈尔德
菌(finegoldia magna)菌株的ppl和pab。
[0161]
在某些实施方案中,本文所述的融合蛋白包含源自链球菌蛋白g白蛋白结合结构域的abd多肽。在某些实施方案中,abd多肽是完全链球菌蛋白g白蛋白结合结构域3或其功能片段和/或变体。
[0162]
在某些实施方案中,融合蛋白包含abd多肽,该多肽包含与seq id no:66具有至少93%序列同源性的多肽序列。例如,abd多肽可以包含与seq id no:66具有至少94%、96%或98%同源性的多肽序列。在某些实施方案中,abd多肽的序列可以与seq id no:66相差1、2、3、4、5、6、7、8、9或10个氨基酸修饰(例如插入、取代、缺失等)。在某些实施方案中,abd多肽包含具有保守氨基酸置换、非保守氨基酸置换或其组合的多肽。
[0163]
在某些实施方案中,融合蛋白包含abd多肽,该多肽包含与seq id no:67具有至少93%序列同源性的多肽序列。例如,abd多肽可以包含与seq id no:67具有至少94%、96%或98%同源性的多肽序列。在某些实施方案中,abd多肽的序列可以与seq id no:67相差1、2、3、4、5、6、7、8、9或10个氨基酸修饰(例如插入、取代、缺失等)。在某些实施方案中,abd多肽包含具有保守氨基酸置换、非保守氨基酸置换或其组合的多肽。
[0164]
在某些实施方案中,融合蛋白包含abd多肽,该多肽包含与seq id no:68具有至少93%序列同源性的多肽序列。例如,abd多肽可以包含与seq id no:68具有至少93%、95%或97%同源性的多肽序列。在某些实施方案中,abd多肽的序列可以与seq id no:68相差1、2、3、4、5、6、7、8、9或10个氨基酸修饰(例如插入、取代、缺失等)。在某些实施方案中,abd多肽包含具有保守氨基酸置换、非保守氨基酸置换或其组合的多肽。
[0165]
abd多肽和精氨酸酶多肽的相对位置可以变化。例如,abd多肽可以在精氨酸酶多肽之前(例如,精氨酸酶多肽可以直接或间接地从abd多肽的c末端连接)或精氨酸酶多肽可以在abd多肽之前(例如,abd多肽可以直接或间接地从精氨酸酶多肽的c末端连接)。
[0166]
在某些实施方案中,融合蛋白可以包含一种或多种精氨酸酶多肽和/或一种或多种abd多肽。例如,融合蛋白可以具有一般结构abd-rharg-abd、abd094-rharg-abd094、abd-bca-abd、abd094-bca-abd094、rharg-abd-rharg、rharg-abd094-rharg、bca-abd-bca或bca-abd094-bca。
[0167]
abd多肽和精氨酸酶多肽可以通过直接共价连接进行连接,或通过肽接头间接连接。
[0168]
肽接头或接头是长度的范围通常为约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20个或更多氨基酸的多肽,该多肽被设计成促进abd多肽和精氨酸酶多肽功能连接成连接的融合蛋白。术语功能连接表示促进多肽正确折叠成三维结构的连接,该三维结构允许连接的融合蛋白表现出其多肽成分所源自的蛋白质的一些或全部功能方面或生物活性。
[0169]
多肽接头可以布置在abd多肽的n末端和精氨酸酶多肽的c末端之间,或可选地布置在精氨酸酶多肽的n末端和abd多肽的c末端之间。
[0170]
肽接头可包含天然存在的氨基酸、非天然氨基酸及其组合。
[0171]
在某些实施方案中,肽接头可包含甘氨酸、丝氨酸、天冬酰胺或其组合。示例性肽接头包括含有聚甘氨酸、(gs)n和(ggs)n的接头,其中n是1-30。另外的示例性肽接头包括柔性接头(例如,(g4s)n,其中n=1-4),或刚性接头(例如,α-螺旋接头[a(eaaak)na]
x
,其中n=
2-4并且x=1或2;和xpn,其中x是a、k或e并且n=1-10)。在某些实施方案中,肽接头是(a(eaaak)4alea-(eaaak)4a)(seq id no:105)、g4sg4sg3sg(seq id no:106)、gs(n)ngsg(其中n=1
–
10)和gs(q)ngsg(其中n=1
–
10)等。在某些实施方案中,肽接头包含与seq id no:73具有至少90%序列同源性的多肽序列。例如,肽接头可包含与seq id no:73具有至少91%、92%、93%、94%、95%、96%、97%、98%或99%序列同源性或与其相同的多肽。
[0172]
在某些实施方案中,肽接头可包含甘氨酸、丝氨酸、天冬酰胺或其组合。在某些实施方案中,肽接头包含与seq id no:74具有至少90%序列同源性的多肽序列。例如,肽接头可包含与seq id no:74具有至少91%、92%、93%、94%、95%、96%、97%、98%或99%序列同源性或与其相同的多肽。
[0173]
纯化标签可用于提高纯化融合蛋白的便利性,例如通过亲和层析。众所周知的纯化标签是六组氨酸(6x his)标签,它是六个组氨酸残基的序列。因此,在某些实施方案中,融合蛋白还包含包含4-8个组氨酸氨基酸的聚组氨酸,例如6x his标签。聚组氨酸可存在于融合蛋白的c末端、融合蛋白的n末端,或安置于abd多肽和精氨酸酶多肽之间。
[0174]
当聚组氨酸安置于abd多肽和精氨酸酶多肽之间时,它可以充当肽接头或可以除了肽接头之外另外被包含。例如,seq id no 75的融合蛋白包含位于300-305位的六组氨酸多肽接头,其用于连接abd多肽和精氨酸酶多肽并且可以有利地用于通过亲和层析纯化融合蛋白。
[0175]
在使用本领域公知的技术完成纯化后,可以任选地去除聚组氨酸标签。例如,外肽酶可用于去除n末端聚组氨酸标签(例如,qiagen tagzyme),并且c末端聚组氨酸标签之前可以是有助于使用内肽酶去除聚组氨酸标签的合适氨基酸序列。因此,不包含n末端和/或c末端聚组氨酸标签的融合蛋白涵盖在本公开的范围内。
[0176]
在某些实施方案中,abd多肽包含与seq id no:66、seq id no:67或seq id no:68具有至少93%序列同源性的多肽序列,并且精氨酸酶多肽包含与seq id no:69、seq id no:70、seq id no:71或seq id no:72具有至少95%同源性的多肽序列。例如,abd多肽包含与seq id no:66、seq id no:67或seq id no:68具有至少94%、95%、96%、97%、98%或99%序列同源性或与其相同的多肽序列,并且精氨酸酶多肽包含与seq id no:69、seq id no:70、seq id no:71或seq id no:72具有至少96%、97%、98%或99%序列同源性或与其相同的多肽序列。
[0177]
在某些实施方案中,abd多肽包含与seq id no:66具有至少93%、94%、95%、96%、97%、98%或99%序列同源性或与其相同的多肽序列,并且精氨酸酶多肽包含与seq id no:69具有至少95%、96%、97%、98%或99%序列同源性或与其相同的多肽序列。
[0178]
在某些实施方案中,融合蛋白还包含肽接头,该肽接头包含与seq id no:73或seq id no:74具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同源性或与其相同的多肽序列。
[0179]
示例性融合蛋白包括与seq id no:49、seq id no:50、seq id no:51、seq id no:52、seq id no:53、seq id no:54、seq id no:55、seq id no:56、seq id no:75和seq id no:76具有至少95%、96%、97%、98%或99%同源性或与它们相同的融合蛋白。
[0180]
融合蛋白对血浆精氨酸浓度的影响的治疗持续时间取决于所施用的融合蛋白的量并且可以是约1天、约2天、约3天、约4天、约5天、约6天、约7天、约8天、约9天、约10天、约11
天、约12天、约13天、约14天或约15天或更长时间。在某些实施方案中,融合蛋白的治疗持续时间为约5天至约20天、约5天至约19天、约5天至约18天、约5天至约17天、约5天至约16天、约5天至约15天、约6天至约15天、约7天至约15天、约7天至约14天、约7天至约13天、约7天至约12天、约7天至约11天、或约8天至约11天。
[0181]
在某些实施方案中,融合蛋白的半衰期为约1天至约10天、约1天至约9天、约1天至约8天、约1天至约7天、约1天至约6天、约1天至约5天、约1天至约4天、约1天至约3天、或约1天至约2天。在其他实施方案中,融合蛋白的半衰期为约6小时至约30小时。
[0182]
本文所述的融合蛋白的精氨酸酶活性可以与其源自的精氨酸酶多肽的活性基本相同、更低或更高。一个精氨酸酶活性单位被定义为在标准测定条件下催化每分钟产生1μmol尿素的融合蛋白[例如,bha(seq id no:75)、bah(seq id no:76)、n-abd-rharg(seq id no:49)或n-abd094-rharg(seq id no:50)]或精氨酸酶[例如,bca(seq id no:70)]的量。酶的比活性表示为每毫克蛋白质的活性单位。在标准二乙酰单肟(damo)测定条件下(37℃,ph7.4),融合蛋白可具有比其掺入的相应精氨酸酶多肽低或高约5%、约10%、约15%、约20%、约25%、约30%、约35%或约40%的比活性。在某些实施方案中,融合蛋白可具有比其掺入的相应精氨酸酶多肽低或高约5%至约40%、约10%至约40%、约10%至约35%、约10%至约30%、约20%至约30%、约20%至约35%、约15%至约30%、约15%至约25%、或约10%至约20%的比活性。
[0183]
在某些实施方案中,融合蛋白的精氨酸酶活性基本上不受hsa存在的影响。这是有利的,因为abd融合蛋白与hsa的结合会对融合蛋白的活性产生有害影响。
[0184]
还提供了编码本文所述的融合蛋白的多核苷酸序列,作为分离的多核苷酸或作为表达载体的部分或作为线性dna序列的部分,包括用于体外转录/翻译的线性dna序列、与组合物的原核或真核表达、分泌和/或展示相容的载体。本文公开了某些示例性多核苷酸,然而,考虑到给定表达系统中遗传密码的简并性或密码子偏好,编码本文所述的融合蛋白的其他多核苷酸也在本公开的范围内。
[0185]
本文所述的多核苷酸可以通过化学合成产生,例如在自动化多核苷酸合成仪上进行固相多核苷酸合成,并组装成完整的单链或双链分子。或者,本发明的多核苷酸可以通过其他技术产生,例如pcr,然后是常规克隆。用于产生或获得给定已知序列的多核苷酸的技术是本领域众所周知的。
[0186]
本文所述的多核苷酸可包含至少一种非编码序列,例如启动子或增强子序列、内含子、多腺苷酸化信号等。多核苷酸序列还可包含编码额外氨基酸的额外序列,这些额外氨基酸编码例如标记或标签序列(例如用于促进蛋白质的纯化或检测的聚组氨酸(6x his)或ha标签)、信号序列、融合蛋白伴侣(例如编码生物活性剂的cdna)等。
[0187]
在另一个实施方案中,本文提供了一种包含至少一种本文所述的多核苷酸的载体。这样的载体可以是质粒载体、病毒载体、用于杆状病毒表达的载体、基于转座子的载体或适合通过任何方式将本发明的多核苷酸引入给定生物体或遗传背景中的任何其他载体。这样的载体可以是包含可以控制、调节、引起或允许由这样的载体编码的多肽的表达的核酸序列元件的表达载体。这样的元件可以包括转录增强子结合位点、rna聚合酶起始位点、核糖体结合位点和其他促进编码多肽在给定表达系统中表达的位点。这样的表达系统可以是本领域熟知的基于细胞的或无细胞的系统。
[0188]
在许多细菌表达系统中,起始密码子通常编码蛋氨酸,因此在这些表达系统中产生以n末端蛋氨酸起始的蛋白质。然而,众所周知,某些细菌酶,例如蛋氨酸氨基肽酶(metap)等,可以催化新合成多肽的n末端蛋氨酸的水解裂解。这通常在下一个氨基酸是gly、ala、ser或thr的情况下观察到[in vivo processing of n-terminal methionine in e.coli,febs lett.1990年6月18日;266(1-2):1-3]。因此,本文所述的融合蛋白的某些实施方案包括其中不存在蛋白质的n末端甲硫氨酸的变体。
[0189]
本文所述的融合蛋白可以使用本领域熟知的用于捕获、固定、分配或沉降的分离程序进行分离,并纯化至商业应用所需的程度。
[0190]
对于治疗用途,本文所述的融合蛋白可以制备为药物组合物,这些药物组合物在药学上可接受的运载体中含有治疗有效量的本文所述的融合蛋白作为活性成分。术语“运载体”是指与活性化合物一起施用的稀释剂、佐剂、赋形剂或载体。这样的载体可以是液体,例如水和油,包括石油来源、动物来源、植物来源或合成来源的油,例如花生油、大豆油、矿物油、芝麻油等。例如,可以使用0.9%的生理盐水和0.3%的甘氨酸。这些溶液是无菌的,并且通常不含颗粒物质。它们可以通过常规的、众所周知的灭菌技术(例如过滤)进行灭菌。组合物可以含有接近生理条件所需的药学上可接受的辅助物质,例如ph调节剂和缓冲剂、稳定剂、增稠剂、润滑剂和着色剂等。这种药物制剂中融合蛋白的浓度可以广泛变化,例如以重量计从小于约0.5%,通常为或至少约1%至多达15%或20%,并且将主要根据所需的剂量、流体体积、粘度等,根据所选的特定给药模式进行选择。合适的载体和制剂描述于例如remington:the science and practice of pharmacy,第21版,troy,d.b.编,lipincott williams and wilkins,philadelphia,pa.2006,第5部分,pharmaceutical manufacturing第691-1092页,特别参见第958-989页。
[0191]
用于本文所述的融合蛋白的治疗用途的给药模式可以是将药剂递送至宿主的任何合适的途径,例如肠胃外给药,例如皮内、肌内、腹膜内、静脉内或皮下、肺;经粘膜(口腔、鼻内、阴道内、直肠);在片剂、胶囊、溶液、悬浮液、粉末、凝胶、颗粒中使用制剂;和包含在注射器、植入装置、渗透泵、药筒、微型泵中;或本领域技术人员理解的其他方式,如本领域公知的。部位特异性给药可以通过例如关节内、支气管内、腹腔内、囊内、软骨内、腔内、体腔内、小脑内、脑室内、结肠内、宫颈内、胃内、肝内、心内、骨内、盆腔内、心包内、腹膜内、胸膜内、前列腺内、肺内、直肠内、肾内、视网膜内、脊柱内、滑膜内、胸内、子宫内、血管内、膀胱内、病灶内、阴道、直肠、颊、舌下、鼻内或经皮递送来实现。
[0192]
在受试者中观察治疗效果所需的血浆精氨酸浓度可基于多种因素而变化,包括受试者的状况和疾病的类型和严重程度和/或医学状况和/或饮食组成。目标血浆精氨酸水平的选择完全在本领域普通技术人员的技能范围内。在某些实施方案中,血浆精氨酸的浓度低于约100μm、约90μm、约80μm、约70μm、约60μm、约50μm、约40μm、约30μm、约20μm、约10μm或约5μm。在某些实施方案中,血浆精氨酸的浓度为约0.1μm至约100μm、约0.1μm至约90μm、约0.1μm至约80μm、约0.1μm至约70μm、约0.1μm至约60μm,约0.1μm至约50μm、约0.1μm至约40μm、约0.1μm至约30μm、约0.1μm至约20μm、或约0.1μm至约10μm。在某些实施方案中,精氨酸水平低于biochrom 30氨基酸分析仪的检测限(例如低于约3μm)和/或低于agilent 6460液相色谱/电喷雾电离三重四极杆质谱仪的检测限(例如,低于约0.3μm)。
[0193]
治疗持续时间(例如,受试者中血浆精氨酸浓度维持在耗尽状态的持续时间)的确
定完全在本领域普通技术人员的技能范围内。在某些实施方案中,治疗持续时间为约1周、约2周、约3周、约4周、约8周、约12周、约16周、约20周、约24周、约28周、约32周、约36周、约40周、约44周、约48周、约52周、约56周或更长时间。
[0194]
在某些实施方案中,在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法包括对受试者共同施用治疗有效量的精氨酸消耗剂和治疗有效量的自噬诱导剂。
[0195]
本领域已知的任何自噬诱导剂均可用于本文所述的方法中。示例性自噬诱导剂包括但不限于卡马西平、可乐定、锂、二甲双胍、雷帕霉素(和雷帕霉素类似物)、利美尼定、丙戊酸钠、维拉帕米、三氟拉嗪、他汀类药物、酪氨酸激酶抑制剂、bh3模拟物、咖啡因、ω-3多不饱和脂肪酸、白藜芦醇、亚精胺、维生素d、海藻糖、多酚(-)-表没食子儿茶素-3-没食子酸酯及其组合。
[0196]
在某些实施方案中,在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法包括对受试者共同施用治疗有效量的精氨酸消耗剂和治疗有效量的葡萄糖降低剂。
[0197]
在某些实施方案中,葡萄糖降低剂是α-葡萄糖苷酶抑制剂、双胍、胆汁酸螯合剂、多巴胺-2激动剂、二肽基肽酶4(dpp-4)抑制剂、美格列奈、钠-葡萄糖转运蛋白2(sglt2)抑制剂、磺酰脲、噻唑烷二酮或其组合。
[0198]
在某些实施方案中,双胍是二甲双胍;α-葡萄糖苷酶抑制剂是阿卡波糖或米格列醇;胆汁酸螯合剂是考来维仑;多巴胺-2激动剂是溴隐亭;dpp-4抑制剂是阿格列汀、利格列汀、沙格列汀或西格列汀;美格列奈是那格列奈或瑞格列奈;sglt2抑制剂是卡格列净、达格列净或恩格列净;磺酰脲是氯磺丙脲、格列美脲、格列吡嗪或格列本脲;并且噻唑烷二酮是罗格列酮或吡格列酮。
[0199]
在某些实施方案中,在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法包括对受试者共同施用治疗有效量的精氨酸消耗剂和治疗有效量的类视黄醇衍生物。
[0200]
在某些实施方案中,类视黄醇衍生物是阿曲汀(acitretin)、阿利维a酸贝沙罗汀(alitretinoin bexarotene)、异维a酸(isotretinoin)、视黄醇、视黄酸或其药学上可接受的盐。在某些实施方案中,类视黄醇衍生物是视黄酸。
[0201]
在某些实施方案中,在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法包括对受试者共同施用治疗有效量的精氨酸消耗剂和治疗有效量的绿茶儿茶素(-)-表没食子儿茶素-3-没食子酸酯(egcg)衍生物或其药学上可接受的盐或产物。在某些实施方案中,egcg衍生物是egcg或其药学上可接受的盐或egcg过乙酸盐。在某些实施方案中,绿茶儿茶素是(-)-表儿茶素(ec)、(-)-表儿茶素-3-没食子酸酯(ecg)、(-)-表没食子儿茶素(egc)及其衍生物。
[0202]
在某些实施方案中,在有需要的受试者中诱导间歇性禁食、调节自噬或诱导间歇性禁食和调节自噬的方法包括对受试者共同施用治疗有效量的精氨酸消耗剂和治疗有效量的雷帕霉素或雷帕霉素衍生物。
[0203]
可以根据本领域熟知的治疗方案施用精氨酸消耗剂。对于本领域技术人员来说显而易见的是,精氨酸消耗剂和自噬诱导剂的施用可以根据所治疗的疾病或健康状况以及自
噬诱导剂对这种疾病或健康状况的已知作用而变化。此外,根据熟练临床医生的知识,治疗方案(例如,剂量和给药时间)可以根据观察到的给药治疗剂(即自噬诱导剂)对受试者的作用,并鉴于观察到的疾病对给药治疗剂的反应而变化。
[0204]
此外,一般来说,精氨酸消耗剂和自噬诱导剂不必在相同的药物组合物中施用,并且可能由于不同的物理和化学特性,必须通过不同的途径施用。例如,精氨酸消耗剂可以静脉内施用以产生和维持良好的血液水平,而自噬诱导剂可以口服施用。在可能的情况下,在相同的药物组合物中确定给药方式和给药的可取性完全在熟练临床医生的知识范围内。初始给药可以根据本领域已知的既定方案进行,然后根据观察到的效果,熟练的临床医生可以修改剂量、给药方式和给药时间。
[0205]
自噬诱导剂的具体选择将取决于主治医师的诊断以及他们对受试者病情的判断以及适当的治疗方案。
[0206]
精氨酸消耗剂和自噬诱导剂可以同时(例如同时、基本上同时或在相同的治疗方案中)或依次给药,这取决于治疗中的疾病或健康状况的性质、受试者的状况,以及将要与精氨酸消耗剂联合给药(即在单一治疗方案中)的自噬诱导剂的实际选择。
[0207]
如果精氨酸消耗剂和自噬诱导剂不是同时或基本上同时施用,那么精氨酸消耗剂和自噬诱导剂的最佳施用顺序可能因不同的疾病或健康状况而不同。因此,在某些情况下,可以先施用精氨酸消耗剂,然后再施用自噬诱导剂;以及在其他情况下,可以先施用自噬诱导剂,然后再施用精氨酸消耗剂。这种交替给药可以在单一治疗方案中重复。在评估了所治疗的疾病或健康状况以及受试者的状况之后,在治疗方案期间每种治疗剂的施用顺序和重复施用次数的确定完全在熟练医师的知识范围内。例如,可以先施用自噬诱导剂,然后再施用精氨酸消耗剂继续治疗,在确定有利的情况下,然后再施用自噬诱导剂,依此类推,直到治疗方案完成。
[0208]
因此,根据经验和知识,执业医师可以根据个体患者的需要,随着治疗的进行,修改用于施用成分(精氨酸消耗剂和自噬诱导剂)的每种方案。
[0209]
实施例
[0210]
实施例1.n-abd094-rharg(seq id no:50)诱导间歇性禁食的重复周期
[0211]
重组人精氨酸酶[n-abd094-rharg(seq id no:50)]是一种新型的工程化长效精氨酸消耗酶。每周一次用n-abd094-rharg(seq id no:50)治疗可以诱导由禁食期和再喂食期构成的重复的7天间歇性禁食周期。
[0212]
间歇性禁食最显著的影响之一是减轻体重。为了证明这种效果,采用了一种饮食诱导的肥胖小鼠模型[wang和liao,2012,methods mol.biol.821:421-433],该模型与人类肥胖具有许多共同特征,并被广泛用于测试潜在的抗肥胖药物[vickers等人,2011,br.j.pharmacol.164(4):1248-1262]。具体来说,本研究中使用了多基因肥胖模型最常用的小鼠品系c57bl/6j。据报道,以高脂肪饮食喂食的c57bl/6j雄性小鼠体重会显著增加,变得肥胖并发展为高胰岛素血症、高血糖、葡萄糖耐量受损和胰岛素抵抗[gallou-kabani等人,2007,obesity15(8):1996-2005]。从5周大开始,通过随意喂食含有60kcal%脂肪(hfd;research diets,d12492)的高脂肪饮食持续12-14周,在c57bl/6j雄性小鼠中诱导肥胖。这些小鼠被称为饮食诱导的肥胖(dio)小鼠。dio小鼠根据体重分为2组。一组接受腹腔(i.p.)注射约600u溶解在生理盐水中的n-abd094-rharg(seq id no:50)(rharg)[hfd(rharg)
组],而另一组每周一次接受载体(生理盐水)[hfd(载体)组],持续34周。两组小鼠在整个治疗期间继续随意喂食hfd。随意喂食普通啮齿动物饲料(cd;labdiet,50if/6f)并注射载体的一组年龄匹配的c57bl/6j雄性小鼠用作瘦对照组[cd(载体)组]。每天测量小鼠的食物摄入和体重。如图1a所示,虽然用载体治疗的dio和瘦对照小鼠的每日食物摄入相对稳定,但用rharg治疗的dio小鼠在整个治疗期间表现出间歇性禁食的重复周期。图1b说明了7天间歇性禁食周期的模式,显示了周期中每一天的平均食物摄入。dio小鼠在第0天注射rharg,这是前一个周期的第7天。类似地,图1b所示周期的第7天是下一个周期的第0天。间歇性禁食周期由禁食期和再喂食期构成。对于图1b中所示的一组小鼠表现出的典型周期,在rharg给药后的第一天,食物摄入略有减少。在接下来的2天内,食物摄入显著减少(禁食),随后食物摄入逐渐增加(再喂食)。平均而言,用rharg治疗的小鼠在一周内的食物摄入减少了约30%(图1c)。伴随着重复间歇性禁食周期的诱导,rharg的给药可以有效地导致体重大幅减轻,从而在治疗的6-7周内,dio小鼠的体重减少了40%(从50g到30g),并且变得与瘦对照小鼠的体重类似(图2a)。在剩余的治疗期间,尽管继续喂食hfd,但施用rharg的小鼠体重仍保持在30g左右(图2b)。相比之下,在34周治疗期间,不仅喂食hfd并施用载体的dio小鼠体重从50g逐渐增加到66g,瘦对照小鼠在34周治疗期间也表现出体重从28g增加到42g。体重增加是与衰老相关的生物标志物之一。总之,这些结果表明rharg具有有效的抗肥胖作用。
[0213]
为了确定rharg对诱导间歇性禁食和调节体重的影响是否可重现,将rharg施用于dio和瘦对照c57bl/6j雄性小鼠的独立队列,并将治疗期延长至49周。如图3a所示,每周一次施用rharg的dio小鼠在整个治疗期间经历了间歇性禁食的重复周期。7天间歇性禁食周期的模式(图3b)和由此导致的每周食物摄入减少约29%(图3c)与上述用rharg治疗34周的小鼠队列中发生的情况非常相似(图1)。类似地,伴随间歇性禁食的重复周期的诱导,这个小鼠队列的体重在rharg施用的6-7周内从50g减少到30g,然后在剩余的治疗期间保持相对恒定(图4)。这些结果表明rharg的作用是可重现的。
[0214]
实施例2.n-abd094-rharg(seq id no:50)不诱导耐药性
[0215]
如图1a所示,dio小鼠在整个34周的治疗期间经历了间歇性禁食周期,这表明小鼠仍然对rharg的作用敏感,而没有产生耐药性。
[0216]
中和抗体的产生是对蛋白质药物产生耐药性的常见原因。使用酶联免疫吸附测定(elisa)检测每周注射后5周和23周时采集的小鼠血清中抗n-abd094-rharg(seq id no:50)的抗体,发现在5周时可检测到抗rharg抗体(图5a),并且抗体滴度在23周时保持相同(图5b)。然而,尽管血清中存在抗rharg抗体,当将固定浓度的rharg(1,000u/ml)与血清一起孵育,然后测量rharg的酶活性时,rharg的酶活性没有降低(图5c),这暗示血清中的抗rharg抗体没有中和作用,因此表明产生的抗药物抗体不会干扰药物分子的活性位点。
[0217]
总之,这些结果表明n-abd094-rharg(seq id no:50)不会诱导耐药性并适合长期使用。
[0218]
实施例3.n-abd094-rharg(seq id no:50)在饮食诱导的肥胖c57bl/6j雄性小鼠中诱导间歇性禁食并逆转肥胖
[0219]
间歇性禁食已被证明可以减少肥胖、高血压并带来代谢益处。如图6所示,伴随间歇性禁食的重复周期的诱导(图1)和显著的体重减轻(图2),在用rharg治疗的dio小鼠中的肾周脂肪垫(内脏白色脂肪组织wat的代表仓库)(图6a)、腹股沟脂肪垫(皮下wat的代表仓
库)(图6b)和肩胛间脂肪垫(棕色脂肪组织bat的代表仓库)(图6c)的质量小于用载体治疗的dio小鼠的该质量的20%。
[0220]
除了wat和bat,肥胖还与肝脏中脂质的过度积累(肝脏脂肪变性)有关。事实上,载体治疗的dio小鼠的肝脏明显增大(图7a),质量(4.68g)约为瘦对照组(1.73g)的3倍(图7b)。然而,在用rharg治疗的dio小鼠中,肝脏质量显著减少至1.39g(图7b)。丙氨酸转氨酶(alt)和天冬氨酸转氨酶(ast)这两种常用的肝损伤生物标志物的血清浓度恢复到与瘦对照小鼠相当的水平。
[0221]
除了肝脏,肥胖还通常与肾脏(肾脏脂肪变性)和心脏(心脏脂肪变性)中的的脂质积累有关,导致这些器官的脂毒性和功能障碍。事实上,用载体治疗的dio小鼠的肾脏和心脏质量有所增加(图8a和图9a)。尿液中白蛋白与肌酐的比率(通常用作肾损伤的生物标志物)显著增加(图8b)。与瘦对照小鼠相比,载体治疗的dio小鼠的收缩压和舒张压(图9b)以及心率(图9c)显著更高。值得注意的是,在每周接受一次rharg治疗的dio小鼠中,它们的肾脏(图8a)和心脏质量(图9a)显著低于载体治疗的dio小鼠。尿液中白蛋白与肌酐的比率(图8b)、在12周(图9b和c)和27周(图9d和e)测量的血压和心率均处于与年龄匹配的瘦对照小鼠相似的水平。
[0222]
总之,这些发现支持rharg可以有效地逆转肥胖并保护包括肝脏、肾脏和心脏在内的主要器官免受与肥胖相关的功能障碍和疾病的影响,并在持续摄入高脂肪饮食的情况下预防高血压、肾脏和肝脏损害。
[0223]
实施例4.用n-abd094-rharg(seq id no:50)治疗可逆转hfd诱导的胰岛素抵抗和葡萄糖耐量受损
[0224]
间歇性禁食已被证明对2型糖尿病有益。以hfd喂食12周的c57bl/6j雄性小鼠会出现外周胰岛素抵抗(图10a)和受损的葡萄糖耐量(图11a),这是前驱糖尿病的特征。然而,dio小鼠在接受每周一次的n-abd094-rharg(seq id no:50)治疗后的第16周(图10b)和第32周(图10c)进行胰岛素耐受试验时,显示出胰岛素抵抗的完全逆转,并表现出胰岛素敏感性的显著增加。事实上,持续喂食hfd的rharg治疗小鼠(图10b和10c)比喂食饲料的年龄匹配的载体治疗小鼠具有更好的胰岛素敏感性。
[0225]
除了胰岛素敏感性,用n-abd094-rharg(seq id no:50)治疗的小鼠在rharg治疗后的15周(图11b)和31周(图11c)测试时表现出葡萄糖耐量显著改善至与瘦对照小鼠相当的水平。
[0226]
总的来说,我们的研究结果证明了n-abd094-rharg(seq id no:50)在增强胰岛素敏感性和葡萄糖耐量方面的功效,并且它具有作为胰岛素增敏剂用于治疗性治疗以胰岛素抵抗、葡萄糖不耐受和其他与胰岛素抵抗相关的病症为特征的前驱糖尿病和2型糖尿病的巨大潜力。
[0227]
实施例5.通过控制喂食而非每日减少食物摄入的人工间歇性禁食可诱导与n-abd094-rharg(seq id no:50)相似的抗肥胖作用
[0228]
施用n-abd094-rharg(seq id no:50)的小鼠每周减少了约30%的食物摄入。为了破译rharg的抗肥胖作用是否是由间歇性禁食和/或食物摄入减少引起的,对c57bl/6j雄性小鼠进行以下喂食方案:
[0229]
c57bl/6j雄性小鼠从5周大开始喂食高脂肪饮食持续12周。然后根据体重将它们
分为3组并单独关在笼子里:(i)一组小鼠接受每周一次腹腔注射600u n-abd094-rharg(seq id no:50)[hfd(rharg)组]并随意喂食hfd 5周;(ii)一组小鼠接受每周一次腹腔注射生理盐水,并每天喂食预定量的hfd,以人为地创建为期7天的间歇性禁食周期,该周期模仿每周一次施用rharg的小鼠的食物摄入模式[hfd(人工if)组)];(iii)一组小鼠每周注射一次生理盐水,并随意喂食hfd[hfd(载体)组]。
[0230]
3组小鼠的食物摄入模式和每周的总食物摄入量分别如图12a和12b所示。结果表明,通过rharg治疗或人工预定喂食方案实现为期7天的间歇性禁食周期的小鼠在5周期间表现出显著的体重减轻。体重变化的模式相似,但在治疗的前3周内,hfd(rharg)组的体重减轻速度快于hfd(人工if)组。在治疗5周结束时,两组小鼠肾周和腹股沟wat以及肩胛间bat的脂肪垫质量均显著减少,其中hfd(rharg)组wat减少的程度更大。两组的肝脏质量显著减少到相同程度。
[0231]
然而,尽管在降低体重和脂肪量方面表现出显著效果,但在治疗后的2周和4周进行的胰岛素耐量试验中,经历人工间歇性禁食周期的小鼠在胰岛素敏感性方面没有显示出显著改善(图14a)。相比之下,接受每周注射rharg的小鼠在治疗后2周时表现出胰岛素抵抗逆转和胰岛素敏感性显著增加,并且在治疗后4周重复itt测试时,这种改善得到了很好的维持。另一方面,两组小鼠在治疗3周后都显示出葡萄糖耐量的显著改善(图14b)。总之,这些结果表明n-abd094-rharg(seq id no:50)对改善葡萄糖耐量的作用可能是由体重减轻和/或间歇性禁食引起的,而其在增加胰岛素敏感性方面的强大效力是通过其他机制介导的。
[0232]
为了进一步破译n-abd094-rharg(seq id no:50)的有益作用是否是由减少食物摄入和/或间歇性禁食带来的,将dio雄性小鼠根据体重分为3组并单独关在笼子里:(i)一组小鼠接受每周一次腹腔注射600u n-abd094-rharg(seq id no:50)[hfd(rharg)组]并随意喂食hfd 5周;(ii)一组小鼠接受每周腹腔注射一次生理盐水,并每天喂食2.0g的hfd(约占dio小鼠平均每日食物摄入量的70%)[hfd(减少)组];(iii)一组小鼠每周注射一次生理盐水,并随意喂食hfd[hfd(载体)组]。
[0233]
3组小鼠的食物摄入模式和每周的总食物摄入量分别如图15a和15b所示。结果表明,尽管hfd(rharg)组和hfd(减少)组的小鼠每周总食物摄入减少了30%,但经过7天间歇性禁食周期的hfd(rharg)小鼠表现出比hfd(减少)组更加突出和更加快速的体重减轻(图15c)。结果,在治疗5周结束时,hfd(rharg)小鼠的体重下降了44%,而hfd(减少)组的体重仅下降了16%。与这一发现一致,这两组小鼠在肾周和腹股沟wat以及肩胛间bat的脂肪垫质量减少程度方面存在显著差异。hfd(rharg)和hfd(减少)组之间的这类差异远大于hfd(rharg)和hfd(人工if)组之间的差异(图13),这意味着间歇性禁食的抗肥胖作用不是仅仅因为食物摄入减少。
[0234]
与先前的发现相似,在该实验中,接受每周注射rharg的小鼠在治疗后2周时显示出胰岛素敏感性显著增加(图17a),并且具有改善的葡萄糖耐量(图17b)。相比之下,每天减少食物摄入的小鼠仅表现出葡萄糖耐量的显著改善,但仍然存在胰岛素抵抗。
[0235]
总的来说,这两个独立实验的结果支持间歇性禁食的诱导在介导n-abd094-rharg(seq id no:50)的抗肥胖作用中起重要作用。
[0236]
实施例6.不同精氨酸消耗剂可诱导喂食hfd的c57bl/6j雄性小鼠的间歇性禁食并
减轻体重
[0237]
为了确定间歇性禁食的诱导是否仅限于n-abd094-rharg[seq id no:50]的作用,检查了其他精氨酸消耗剂。
[0238]
如图18a中所示,通过腹腔注射向预先存在通过从5周大开始喂食hfd持续12周诱导的肥胖症的c57bl/6j雄性小鼠每周一次施用250u聚乙二醇化his-rharg(seq id no:101)持续8周,可以诱导由禁食期和再喂食期构成的重复7天间歇性禁食周期(图18b),同时每周的总食物摄入相比载体治疗的dio小鼠[hfd(载体)组]减少约28%。伴随间歇性禁食的诱导,hfd(peg-rharg)组小鼠在治疗8周内体重从50g显著减轻至30g(图19)。它们显示出胰岛素敏感性显著增加(图20a)并显著改善了葡萄糖耐量(图20b)。在治疗期结束时,肾周和腹股沟wat和肩胛间bat的脂肪垫质量以及肝脏质量显著降低至与喂食饲料的载体治疗的瘦对照小鼠[cd(载体)组]相似的重量(图21)。肾脏和心脏质量也显著减少。
[0239]
总之,这些发现表明聚乙二醇化his-rharg[seq id no:101]可以诱导间歇性禁食,并具有与n-abd094-rharg[seq id no:50]相似的抗肥胖和胰岛素增敏作用。聚乙二醇化his-rharg(seq id no:101)剥夺精氨酸对治疗肥胖症和与胰岛素抵抗相关的疾病具有治疗效果。
[0240]
接下来,预先存在饮食诱导的肥胖症的c57bl/6j雄性小鼠每周接受一次腹腔注射50u n-abd094-rharg-co
2+
[seq id no:50(钴取代)],持续2周,同时连续喂食hfd。小鼠表现出7天的间歇性禁食周期,其中有4天的长禁食期,然后再喂食(图22a)。与较长的禁食期一致,在治疗的2周内体重从45g显著减轻至32g(图22b)。这些结果表明,低得多的n-abd094-rharg-co
2+
药物剂量可以诱导更长的禁食期,这意味着用co
2+
替代mn
2+
显著增加了abd-rharg融合蛋白在诱导间歇性禁食周期和减少体重方面的效力。
[0241]
除了将精氨酸转化为鸟氨酸和尿素的精氨酸酶之外,还检查了另一种将精氨酸转化为瓜氨酸和氨的精氨酸消耗酶精氨酸脱亚氨酶。通过腹腔注射对具有由hfd诱导的预先存在的肥胖症的c57bl/6j雄性小鼠施用5u adi-abd(seq id no:107)。第一天后食物摄入降至最低水平,然后逐渐增加到正常水平(图23a)。禁食然后再喂食的趋势与n-abd094-rharg(seq id no:50)的趋势相似但不相同,n-abd094-rharg在大约第2天至第3天而不是第1天具有最低食物摄入水平。对于体重,在第一个周期中,它在2天内从47g下降到约44g,并在再喂食阶段保持在该水平(图23b)。
[0242]
总之,这些发现支持可以通过除n-abd094-rharg[seq id no:50]之外的精氨酸消耗剂来实现间歇性禁食的重复周期的诱导。abd-rharg融合蛋白或其他形式的具有延长半衰期的精氨酸酶(例如rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如adi-peg、adi-abd、adc-peg或adc-abd)可能具有类似的治疗效果,例如治疗肥胖和与胰岛素抵抗相关的疾病。
[0243]
实施例7.n-abd094-rharg(seq id no:50)可以防止喂食hfd的c57bl/6j雄性小鼠出现认知缺陷
[0244]
间歇性禁食已被证明可以带来许多健康益处并具有抗衰老作用。它可以帮助预防和治疗多种疾病。例如,间歇性禁食可以防止神经退行性变。它可以提高认知能力。
[0245]
为了确定可诱导重复间歇性禁食周期的n-abd094-rharg[seq id no:50]是否对认知能力具有有益影响,进行了十天的巴恩斯迷宫测试以评估空间学习和记忆功能。测试
在一个平坦的圆形平台(直径100cm,厚1.5cm,高出地面40cm)上进行,平台周围有20个均匀分布的孔(直径7cm),并在其中的一个孔下方放置逃生箱。在实验之前,每只小鼠都被引导到位于逃生箱上方的特定孔中并停留180秒。随后,通过将小鼠置于平台中央的小黑室下方3秒,然后将小黑室抬起,让小鼠在180秒内找到逃生孔,从而训练每只小鼠寻找逃生孔。使用台扇作为不利刺激,平台被明亮地照亮和冷却。如果小鼠在允许的时间内无法找到目标孔,则将其引导至逃生箱。每天进行两次试验,连续四天作为训练期,之后分别在第5天和第10天通过移除逃生盒并让小鼠探索平台整整180秒来探测小鼠的短期和长期空间记忆。每个试验都直接在平台上记录。对于第1天到第4天的每次试验,记录找到逃生孔和离开平台所需的时间,以及探索中探索模式的类型(随机、依次或直接)。对于第5天和第10天的每次探测试验,在180秒内记录小鼠研究20个孔中每个孔的次数,其中一个研究实例定义为给定孔内的鼻子定位。
[0246]
空间记忆丧失是2型糖尿病和肥胖症啮齿动物模型的一个突出特征[boitard等人,2014;underwood and thompson,2016]。为了研究精氨酸酶治疗是否可以预防喂食hfd的小鼠的空间记忆丧失,dio c57bl/6j雄性小鼠从17周大开始每周接受n-abd094-rharg(seq id no:50)施用后43周,通过巴恩斯迷宫测试评估空间记忆。
[0247]
从训练阶段的第2天开始,喂食饲料的年龄匹配的载体治疗小鼠[cd(载体)组]和用n-abd094-rharg(seq id no:50)治疗的hfd喂食小鼠[hfd(rharg)组]比用载体治疗的hfd喂食小鼠[hfd(载体)组]明显更早地离开平台(图24)。此时,尽管发现hfd(载体)组90%的小鼠在第1天使用随机型探索模式,而其他两组主要使用依次型探索模式,但所有组的小鼠都表现出相似的探索模式(图25)。当在第5天探测小鼠的短期记忆时,cd(载体)和hfd(rharg)组的小鼠以越来越高的频率探索离目标最近的孔,其中目标孔是探索最多的孔,而hfd(载体)组小鼠以相似的频率探索所有孔(图26a)。当第10天探测小鼠的长期记忆时,这种趋势得以维持(图26b)。
[0248]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可能具有预防或改善认知缺陷的治疗作用。
[0249]
实施例8.n-abd094-rharg(seq id no:50)可以恢复喂食hfd的c57bl/6j雄性小鼠的神经肌肉力量和协调性
[0250]
为了评估小鼠的平衡性、神经肌肉力量和协调性,喂食hfd的dio c57bl/6j雄性小鼠在从17周大开始接受每周施用n-abd094-rharg(seq id no:50)之后分别在42周和30周进行倒置网格悬挂测试和旋转棒测试。
[0251]
四肢倒置网格悬挂测试使用线栅设置以非侵入性地测量小鼠表现出持续肢体张力以抵抗其重力的能力。记录小鼠从网格上掉下来的时间(延迟)。结果表明,与喂食饲料的载体治疗的对照小鼠[cd(载体)组]相比,长期喂食hfd的载体治疗小鼠[hfd(载体)组]表现出神经肌肉强度严重降低,并且耐力降低了90%。相比之下,每周接受n-abd094-rharg(seq id no:50)[hfd(rharg)组]治疗42周的hfd喂食小鼠表现出显著改善的神经肌肉性能,其耐力甚至高于cd(载体)组的小鼠(图27a)。
[0252]
旋转棒测试用于评估啮齿动物的神经肌肉协调和平衡。小鼠必须在300秒内以5到40rpm的加速模式在旋转杆上保持平衡。记录小鼠从棒上掉下来的时间(延迟)。结果表明,
经过长期喂食hfd的载体治疗小鼠[hfd(载体)组]在旋转棒上表现出比喂食饲料的载体治疗对照小鼠[cd(载体)组]更短的延迟。相比之下,每周接受n-abd094-rharg(seq id no:50)[hfd(rharg)组]治疗30周的hfd喂食小鼠在旋转棒测试中表现出显著改善的性能,其延迟期与cd(载体)组的小鼠类似(图18)。
[0253]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可能具有改善神经肌肉力量和协调性的治疗作用。
[0254]
实施例9.n-abd094-rharg(seq id no:50)可以防止喂食hfd的c57bl/6j雄性小鼠出现肝癌
[0255]
间歇性禁食已被证明具有抗衰老作用并可以预防癌症,这是衰老的生物标志物之一。
[0256]
肝脏肿瘤的发展是长期喂食hfd的c57bl/6j雄性小鼠衰老的主要特征之一,这可能与严重的非酒精性脂肪肝疾病的发展有关。用n-abd094-rharg[seq id no:50]治疗可有效逆转肝脂肪变性并降低肝损伤生物标志物alt和ast的血清浓度(图7)。在实验结束时进行的尸检证实,在从5周大开始以hfd喂食46周(12+34周)的约40%的c57bl/6j雄性小鼠[图1中的hfd(载体)组]中存在肝细胞癌(图28a)(图28b)。在以hfd喂食61周(12+49周)[图3中的hfd(载体)组]的小鼠中,发病率甚至增加到100%(图28c)。相比之下,在从17周大开始每周接受n-abd094-rharg注射34周[图1中的hfd(rharg)组]和49周[图3中的hfd(rharg)组]的dio c57 bl/6j雄性小鼠中,没有一只小鼠患上肝细胞癌。这些发现支持rharg对预防肝癌具有有效作用。
[0257]
实施例10.n-abd094-rharg(seq id no:50)在喂食hfd的肥胖icr雌性小鼠中诱导间歇性禁食并降低体重
[0258]
在确认n-abd094-rharg(seq id no:50)在雄性c57bl/6j小鼠中诱导间歇性禁食并伴随体重减轻中的作用后,研究n-abd094-rharg(seq id no:50)对雌性小鼠的作用以确定是否可能存在任何性别差异。我们发现,与其他报道的结果相似,c57bl/6j雌性小鼠不像c57bl/6j雄性小鼠那样容易患上饮食引起的肥胖症。因此,我们使用了icr雌性小鼠,这些小鼠在从5周大开始用hfd喂食12周后出现肥胖症,体重约50g。图29显示了将1200u的n-abd094-rharg(seq id no:50)每周一次通过腹腔注射施用于饮食诱导的肥胖icr雌性小鼠持续56周的结果,这些小鼠继续食用hfd[hfd(rharg)组]。每日监测食物摄入的结果表明,7天的间歇性禁食周期(图29a)在整个56周的治疗期中反复出现在icr雌性小鼠中,该间歇性禁食周期由禁食期和再喂食期组成(图29b),模式类似于c57bl/6j雄性小鼠(图1b)。尽管与喂食hfd的用载体治疗的icr雌性小鼠[hfd(载体)组]相比,每周总食物摄入量平均减少了14%(图29c),但在9-10周内,体重显著下降了30%,达到与喂食饲料的年龄匹配的瘦对照icr雌性小鼠[cd(载体)组]的体重相似的水平(图30)。尽管持续食用hfd,但hfd(rharg)组小鼠的体重可以在剩余的治疗期间保持在该水平。相比之下,在56周的治疗期间,hfd(载体)和cd(载体)组的小鼠体重分别从50g逐渐增加到70g和从32g逐渐增加到45g。
[0259]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可在两种性别中诱导间歇性禁食并具有抗肥胖作用。
[0260]
实施例11.n-abd094-rharg(seq id no:50)预防喂食hfd的icr雌性小鼠的年龄相关疾病。
[0261]
肥胖增加、胰岛素抵抗、葡萄糖耐量受损、神经肌肉力量和运动协调性下降以及癌症风险增加是常见的与年龄相关的变化,肥胖会进一步加速这些变化。进行研究以确定在通过每周注射n-abd094-rharg(seq id no:50)持续56周而喂食hfd的雌性小鼠中诱导间歇性禁食的同时,这些参数是否有任何改善。
[0262]
与体重减轻的结果(图30)一致,在56周治疗期结束时,用n-abd094-rharg(seq id no:50)治疗的hfd喂食小鼠[hfd(rharg)组]的肾周和内脏wat、肩胛间bat的脂肪垫质量以及肝脏、肾脏和心脏质量明显低于用载体治疗的hfd喂食小鼠[hfd(载体)组](图31)。此外,类似于对c57bl/6j雄性小鼠的作用,每周用n-abd094-rharg(seq id no:50)治疗可以逆转预先存在的胰岛素抵抗(图32a)和葡萄糖耐量受损(图33a)。事实上,在15周(图32b)和31周(图32c)进行的胰岛素耐量试验中,hfd(rharg)组的小鼠表现出比cd(载体)组中年龄匹配的小鼠更好的胰岛素敏感性。在接受rharg治疗后16周(图33b)和30周(图33c)进行的葡萄糖耐量测试也证实了葡萄糖耐量的改善。
[0263]
此外,分别在治疗期的54周和55周进行的倒置网格悬挂测试(图34a)和旋转棒测试(图34b)中hfd(rharg)组小鼠的表现明显优于hfd(载体)组的小鼠,并且与年龄匹配的cd(载体)组相当,表明用n-abd094-rharg(seq id no:50)治疗可以防止神经肌肉力量和协调性下降。此外,hfd(载体)组中超过30%的小鼠发展为肝细胞癌(图35),而hfd(rharg)组中没有一只小鼠发展为肝细胞癌,因此证明了n-abd094-rharg(seq id no:50)的有效抗癌作用。
[0264]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可以在两种性别中诱导间歇性禁食,同时预防各种与衰老相关的疾病。
[0265]
实施例12.n-abd094-rharg(seq id no:50)在喂食hfd的中年雄性c57bl/6j小鼠中诱导间歇性禁食并改善代谢健康
[0266]
在先前的研究中,hfd诱导的肥胖雄性(实施例1)和雌性小鼠(实施例10)从大约4-5个月大(这相当于人类大约二十岁中期)开始接受n-abd094-rharg(seq id no:50)的治疗。为了确定中年受试者对精氨酸消耗的敏感性,从5周大开始喂食hfd的肥胖c57bl/6j雄性小鼠从大约16个月大(这相当于人类50岁中期)开始每周接受n-abd094-rharg(seq id no:50)注射25周[hfd年老(rharg)组]。
[0267]
如图37所示,16个月大的hfd喂食小鼠的平均体重在治疗前约为68g。每周一次注射n-abd094-rharg(seq id no:50)可以诱导由禁食期和再喂食期(图36b)构成的重复的7天间歇性禁食周期(图36a)。总体而言,与用载体治疗的hfd喂食小鼠[hfd年老(载体)组]相比,每周的食物摄入减少了31%。这种模式与年轻小鼠对用n-abd094-rharg(seq id no:50)(图1b、3b)和其他精氨酸消耗剂治疗的反应非常相似(图18b)。
[0268]
伴随间歇性禁食的重复周期的诱导,hfd年老(rharg)组的小鼠体重逐渐下降,使得在9-10个月内体重显著下降50%,并在整个剩余治疗期中保持在该水平(图37)。除了诱导大量的体重减轻之外,n-abd094-rharg(seq id no:50)治疗可以显著逆转先前存在的胰岛素抵抗(图38a)和葡萄糖耐量受损(图39a),并将hfd年老(rharg)组小鼠的胰岛素敏感性
(图38b)和葡萄糖耐量(图39b)增加到与大约7-8个月大并喂食饲料的载体治疗小鼠[cd年轻(载体)]相似(如果不是更好的话)的水平。
[0269]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可以在不分年龄的饮食诱导的肥胖小鼠中诱导间歇性禁食,并改善代谢健康。
[0270]
实施例13.n-abd094-rharg(seq id no:50)在喂食普通食物的c57bl/6j雄性小鼠中诱导间歇性禁食和体重减轻
[0271]
体重增加和肥胖是常见的与年龄相关的变化。例如,喂食普通食物的c57bl/6j雄性小鼠在5个月大(相当于人类二十岁中期)时体重为30g,在18个月大(相当于人类五十岁中期)时逐渐增加到40g(图41)。为了确定n-abd094-rharg(seq id no:50)是否对年龄相关性肥胖具有代谢益处,喂食普通食物的18个月大的年老c57bl/6j雄性小鼠每周注射一次n-abd094-rharg(seq id no:50)[cd年老(rharg)组]或载体[cd年老(载体)组],持续5个月。5个月大的c57bl/6j雄性小鼠作为年轻对照[cd年轻(载体)](图41)。
[0272]
如图40a所示,在cd年老(rharg)组的小鼠中,每周一次注射n-abd094-rharg(seq id no:50)可以诱导具有禁食期和再喂食期的重复的7天间歇性禁食周期(图40b),并且与cd年老(载体)组中的小鼠相比,每周总食物摄入减少了11%(图40c)。虽然cd年轻(载体)组的小鼠的体重从5个月大的30g逐渐增加到10个月大的35g,但是cd年老(rharg)组的小鼠的体重在abd094-rharg(seq id no:50)治疗6周内减少到30g,并在整个治疗期的剩余时间内很好地维持在该水平。与体重下降一致的是,肾周和腹股沟wat、肩胛间bat的脂肪垫质量和肝脏质量(图44)存在显著降低,以至于其中一些器官的质量甚至低于cd年轻(载体)组中的小鼠。除了减少肥胖,n-abd094-rharg(seq id no:50)治疗还可以显著改善cd年老(rharg)小鼠的胰岛素敏感性(图42a和b)和葡萄糖耐量(图43a和b),达到比cd年轻(载体)组的小鼠更好的水平,因为cd年轻(载体)组的小鼠的年龄在一直增加。
[0273]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗可以在喂食普通饲料的小鼠中诱导间歇性禁食并改善代谢健康。
[0274]
实施例14.n-abd094-rharg(seq id no:50)延长喂食普通食物的年老c57bl/6j雄性小鼠的寿命
[0275]
秀丽隐杆线虫和果蝇的特异性自噬上调可延长寿命,而例如雷帕霉素的诱导自噬的药物可促进啮齿动物的寿命(hansen等人,nature reviews molecular cell biology.2018;19:579
–
593)。对于c57bl/6j小鼠,超过24个月大可被视为“非常老”,并且存活率显著下降(https://www.jax.org/news-and-insights/jax-blog/2017/november/when-are-mice-considered-old)。为了确定诱导自噬系统性上调的n-abd094-rharg(seq id no:50)是否可以延长寿命,将喂食普通饲料的25个月大(相当于人类的七十岁)的非常老的c57bl/6j雄性小鼠分为2组。一组每周用600u n-abd094-rharg(seq id no:50)治疗一次[cd非常老(rharg)组],而另一组用载体治疗[cd非常老(载体)组]持续30周。用载体治疗的一组7个月大(相当于人类三十岁早期)的c57bl/6j雄性小鼠[cd年轻(载体)组]作为在30周的周期内从成熟成年到中年晚期的渐进性改变的参考。
[0276]
如图45b所示,非常老的小鼠表现出与年龄相关的肥胖,体重在2岁时达到45g。每
周一次用n-abd094-rharg(seq id no:50)治疗非常老的小鼠[cd非常老(rharg)组]诱导显著的重复7天间歇性禁食周期(图45a),同时体重发生减少,在3周内降至30g,这与7个月大的成年小鼠[cd年轻(载体)组]的体重相似。[cd年轻(载体)组]的小鼠在7个月的研究期间随着发展成中年,体重从30g逐渐增加到43g。相比之下,对于非常老的小鼠[cd非常老(载体)组],随着年龄的进一步增长,它们的体重逐渐下降。此外,在30个月大时(研究结束),存活率下降到20%(图45c)。值得注意的是,通过每周注射n-abd094-rharg(seq id no:50),存活率维持在80%,这意味着寿命显著增加。
[0277]
这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如adi-peg、adi-abd、adc-peg或adc-abd)可以延长动物的寿命。
[0278]
实施例15.n-abd094-rharg(seq id no:50)在7天间歇性禁食周期的禁食阶段诱导自噬(脂肪吞噬),以分解喂食hfd的c57bl/6j小鼠的肝脏中的脂质
[0279]
自噬参与细胞生长、存活、发育和死亡。自噬潮受损与多种人类病理生理过程有关,包括神经变性、癌症、肌病、心血管和免疫介导的病症。越来越需要鉴别和量化不同病理条件下自噬潮的状态。自噬是一个高度动态和复杂的过程,在多个步骤中受到调节。在存在和不存在溶酶体降解抑制剂氯喹(cq)的情况下,可以使用蛋白质印迹分析通过lc3-ii转换来检测自噬潮。如果发生自噬潮,则在存在溶酶体降解抑制剂的情况下,lc3-ii的水平会增加,因为lc3-ii通过自噬通路的转运将被阻断。
[0280]
我们的结果显示,n-abd094-rharg(seq id no:50)治疗组的肝脏样品中的lc3-ii/lc3-i比率在cq注射5小时后显示出显著增加(图46)。这表明n-abd094-rharg治疗(第3天)在4周时在dio小鼠的肝脏中诱导自噬。
[0281]
图47显示了喂食饲料(cd)或hfd并施用n-abd094-rharg(seq id no:50)(rharg)或生理盐水(载体)4周的小鼠的肝脏切片的透射电子显微镜图像。在喂食hfd并施用载体的小鼠[hfd(载体)组]中,在肝细胞(肝脏细胞)中观察到大脂滴和溶酶体的积累。在喂食hfd并施用n-abd094-rharg(seq id no:50)的小鼠[hfd(rharg)组]中,在7天间歇性禁食周期的第1天,在肝细胞中发现了自噬体和自噬性溶酶体。然而,在这个阶段还没有观察到脂肪吞噬。在7天间歇性禁食周期的第3天,在肝细胞中观察到广泛的脂肪吞噬。有大量的自噬性溶酶体,它们正在分解肝细胞中的脂质内容物。在7天间歇性禁食周期的第5天,肝细胞中仍可发现自噬体,但自噬性溶酶体和脂肪吞噬变得罕见。这表明自噬的循环活化几乎结束。在7天间歇性禁食周期的第7天,没有自噬体,也没有观察到脂肪吞噬。存在溶酶体积累,这与hfd(载体)组相似,但肝细胞中积累的脂质含量下降。积累的脂质含量与在喂食饲料的小鼠[cd(载体)组]中观察到的相似。
[0282]
大量脂肪吞噬的存在与n-abd094-rharg(seq id no:50)治疗后肝脏中脂质含量降低是一致的。hfd喂食12周诱导c57bl/6j雄性小鼠的肝脂肪变性(图48a)。延长hfd喂食12周后,肝脏进一步扩大,颜色变淡(图48a)。肝脏重量是喂食饲料的瘦对照小鼠[cd(载体)组]的两倍以上(图48b)。如脂质的油红o染色所示,大量脂质积累已扩展到整个肝脏(图48a)。然而,在用n-abd094-rharg(seq id no:50)治疗的dio小鼠[hfd(rharg)组]中,肝脏显示出明显的尺寸缩小(图48a),以至于重量与瘦对照组相似(图48b)。此外,肝脏恢复为微红色,这与脂滴的显著清除密切相关(图48a),同时甘油三酯浓度降低至与瘦对照组相似的
水平(图48c)。
[0283]
总的来说,我们的发现揭示了n-abd094-rharg(seq id no:50)通过自噬(脂肪吞噬)逆转肝脂肪变性的显著效果。
[0284]
实施例16.n-abd094-rharg(seq id no:50)在7天间歇性禁食周期的禁食阶段诱导自噬(脂肪吞噬),以分解喂食hfd的c57bl/6j小鼠的bat中的脂质
[0285]
p62,也称为sqstm1/sequestome 1,充当lc3b和泛素化底物之间的联系,并被自噬有效降解。因此,p62蛋白的水平可用于监测自噬潮。例如,自噬抑制与p62水平升高相关,同样,自噬活化与p62水平降低相关。棕色脂肪组织(bat)上p62的蛋白质印迹显示在第3天和第5天p62水平显著降低,这意味着在通过n-abd094-rharg(seq id no:50)施用诱导的间歇性禁食周期的禁食期间存在自噬潮(图49)。此外,在存在或不存在cq的情况下检测自噬标志物lc3b的表达水平(图50)。lc3b-i向低迁移形式lc3b-ii的转化被用作细胞自噬水平的指标。lc3b-ii/lc3b-i比值越高表明组织中自噬的发生率越高。与第3天降低的p62水平一致,结果显示在7天间歇性禁食周期的第3天收集的n-abd094-rharg(seq id no:50)治疗组[hfd(rharg)组]的bat样品中的lc3-ii/lc3-i比值在cq注射5小时后显示出显著增加(图50)。这表明n-abd094-rharg治疗在第3天诱导了肝脏中的自噬。在用载体治疗的肥胖小鼠[hfd(载体)组]中,施用cq后没有增加,表明该组织处于自噬停滞状态。
[0286]
然而,在施用600u n-abd094-rharg(seq id no:50)的由hfd诱导的肥胖雄性小鼠的bat中核糖体蛋白s6激酶β-1(p70s6k1,一种哺乳动物雷帕霉素靶标mtor的下游蛋白)的抑制和unc-51样自噬活化激酶1(ulk1,自噬的引发剂)的刺激表明,自噬的启动可能是通过抑制mtor通路介导的,该mtor通路是精氨酸剥夺的主要传感器。
[0287]
在长期喂食hfd后,与瘦对照小鼠[cd(载体)组]相比,载体治疗的dio小鼠[hfd(载体)组]的肩胛间bat的大小(图53a)和重量(图53b)进一步增加,这与大量的增大白色样单叶细胞的存在相关(图53a)。如在透射电子显微镜下观察到的,喂食hfd的小鼠[hfd(载体)组]中的棕色脂肪细胞具有大量的增大的脂滴(图52a)。相比之下,用hfd喂食并用n-abd094-rharg(seq id no:50)治疗的小鼠[hfd(rharg)组]中的棕色脂肪细胞在治疗的第3天表现出活跃的脂肪吞噬,这是7天间歇性禁食周期的禁食期。许多自噬体正在主动地形成和吞噬脂滴(图52b)。还发现自噬性溶酶体分解小脂滴,与结构中的大量溶酶体混合(图52c)。与第3天相比,第7天n-abd094-rharg(seq id no:50)治疗组的bat切片显示,持续的主动自噬(脂肪吞噬)正在发生,因为在棕色脂肪细胞的细胞质中存在大量的自噬体和自噬性溶酶体(图52d)。此外,还有大范围的展现出空洞的区域,其中的脂质内容物已完全降解,这表明自噬,尤其是脂肪吞噬,在棕色脂肪细胞的细胞质中持续发生。此外,值得注意的是,与第3天样品相比,第7天rharg治疗样品的棕色脂肪细胞的细胞质中存在相对大量的线粒体。这可能表明,一旦rharg诱导的脂肪吞噬消散脂滴,线粒体生物发生也主动发生,达到一定程度。
[0288]
伴随着自噬的存在,用n-abd094-rharg(seq id no:50)治疗dio小鼠可以显著地将肩胛间bat恢复到与瘦对照小鼠相似的大小(图53a)和重量(图58b),同时白色样单叶细胞几乎完全缓解(图53b)。总的来说,这些发现表明由n-abd094-rharg(seq id no:50)诱导的自噬逆转了棕色脂肪的变白。
[0289]
实施例17.n-abd094-rharg(seq id no:50)在下丘脑pomc神经元中诱导自噬,导
致食欲抑制
[0290]
下丘脑中的弓状核作为能量稳态和食欲的整合者和调节者受到了广泛的关注。这些神经元可以快速感知血液中的代谢波动。其他人最近说明了中枢食欲调节和瘦素敏感性中的自噬(park等人,nature communications.2020;11:1914)。敲低必需的自噬基因自噬相关7(atg7)会导致p62在pomc神经元中积累,从而导致体重和食欲增加。此外,已经表明氨基酸剥夺/失衡可以通过eif2α/atf4通路触发食物摄入的减少[maurin等人,cell rep.2014年2月,6(3):438
–
444]。在我们的研究中,成熟的下丘脑神经元(体外培养14天后)暴露于载体或2u/ml n-abd094-rharg(seq id no:50)1小时、4小时、8小时或24小时。治疗后,从神经元的全细胞裂解物中收获蛋白质并进行蛋白质印迹分析,以检查精氨酸消耗对下丘脑神经元的短期影响。分析了自噬标志物(lc3b和p62)。在图54中,可以看出在rharg治疗1小时和4小时后,lc3b-ii的水平显著增加(图54a),这表明原代下丘脑神经元中自噬体的形成增强。总之,在n-abd094-rharg(seq id no:50)治疗4小时和8小时后p62水平显著降低(图54b),这表明自噬体降解增加。这些发现支持n-abd094-rharg(seq id no:50)在下丘脑神经元中诱导自噬潮。
[0291]
除了自噬,在n-abd094-rharg(seq id no:50)治疗后8小时或24小时观察到eif2α的磷酸化水平显著增加(图55a),这表明精氨酸消耗诱导了下丘脑神经元中eif2α的活化。此外,在治疗后24小时,在rharg治疗的神经元中观察到atf4水平的显著增加(图55b),这与体内氨基酸剥夺的发现一致[maurin等人,cell rep.2014年2月,6(3):438
–
444]。
[0292]
此外,在治疗1小时、4小时、8小时或24小时后,用n-abd094-rharg(seq id no:50)治疗显著降低了神经元中p70s6k1的磷酸化水平(图55c)。该数据表明,精氨酸剥夺通过mtor通路抑制下丘脑神经元中p70s6k1的活性。
[0293]
阿黑皮质素原(proopiomelanocortin,pomc)神经元产生神经肽前体pomc,pomc被裂解形成α-黑素细胞刺激激素(α-msh),最终减少食物摄入。我们的数据显示,n-abd094-rharg(seq id no:50)治疗显著增加了处理24小时后神经元中糖基化pomc的比例(图56)。
[0294]
总之,mtor通路的抑制是对精氨酸消耗的最早响应,其次是eif2α/atf4通路的活化。最后,在自噬潮发生后观察到糖基化pomc的比例的上调。
[0295]
自噬通路的上调可能具有神经保护作用,并且正在投入大量精力开发能够穿过血脑屏障并增加神经元自噬的药物。一种公认的诱导全身性自噬的方法是限制食物,这会上调包括肝脏和神经元细胞在内的许多器官中的自噬(alirezaei等人2010.autophagy 6(6):702-710)。因此,零星禁食可能代表一种简单、安全和廉价的方法来促进这种潜在的治疗性神经元反应。这意味着n-abd094-rharg(seq id no:50)或其他形式的精氨酸酶(例如rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如adi-peg、adi-abd、adc-peg或adc-abd)(此处显示为有效的间歇性禁食诱导剂和/或自噬诱导剂)可用于治疗或预防由自噬破坏引起的神经退行性疾病。
[0296]
实施例18.组合n-abd094-rharg(seq id no:50)和二甲双胍在诱导喂食hfd的c57bl/6j雄性小鼠的间歇性禁食方面的协同作用
[0297]
二甲双胍是治疗2型糖尿病的一线药物。
[0298]
除了单独应用之外,我们发现当与二甲双胍组合时,n-abd094-rharg(seq id no:50)在诱导间歇性禁食和减少肥胖方面表现出协同相互作用(图57)。在不同条件下治疗五
rharg(seq id no:50)和二甲双胍的联合治疗检测到胰岛素敏感性的显著增强,达到与正常瘦对照小鼠相似的敏感性水平。
[0304]
喂食hfd的小鼠也表现出葡萄糖耐量受损。结果显示,治疗7周后,单独300mg/kg二甲双胍治疗同样不能缓解葡萄糖不耐受;单独用300u/周的n-abd-rharg(seq id no:50)治疗对葡萄糖耐量具有显著改善作用(图60b)。类似地,通过n-abd-rharg(seq id no:50)和二甲双胍的联合治疗检测到葡萄糖耐量的显著增强,达到与正常瘦对照小鼠相似的水平。综上所述,对小鼠联合使用精氨酸酶和二甲双胍可显著提高胰岛素敏感性并改善葡萄糖耐量,最终可逆转肥胖小鼠的2型糖尿病。
[0305]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗与二甲双胍组合时,可在减少器官中的脂肪堆积和改善胰岛素敏感性和葡萄糖不耐受方面产生协同作用。
[0306]
实施例20.组合n-abd094-rharg(seq id no:50)和二甲双胍在诱导喂食hfd的c57bl/6j雄性小鼠的肝脏中的自噬方面的协同作用
[0307]
通过tem检查治疗3周后肝脏中自噬体的存在(图62、63和64)。如前所述,在长期喂食hfd后,肝脏大小增加(图61a),这与载体治疗的dio中大脂滴的积累相关。如通过tem观察到的,喂食hfd的小鼠的肝脏呈现出大量增大的脂滴(图61b)。单独用300mg/kg二甲双胍或300u/周的n-abd-rharg(seq id no:50)治疗确实显示出脂滴大小的显著降低,但在两种治疗条件下均未观察到自噬体(图62)。相比之下,喂食hfd并同时用n-abd094-rharg(seq id no:50)和二甲双胍治疗的小鼠的肝脏在治疗的第3天,即间歇性禁食周期的禁食期可以明显清除肝细胞中的大部分脂滴(图63)。此外,由于观察到大量活跃的自噬体,因此在细胞中触发了自噬(图64)。在5000倍放大倍率下,我们发现脂肪吞噬通过产生自噬体而活跃地发生,这些自噬体隔离部分大脂滴以形成双膜囊泡,从而将脂滴分解成更小且更易消化的大小(图64)。然而,还观察到巨自噬,其特征在于含有多种细胞质成分的大型自噬体与溶酶体融合以进一步形成自噬性溶酶体(图64)。
[0308]
治疗11周后肝脏的一般h&e染色(图61)显示了与治疗第三周在tem中的观察平行的结果。n-abd094-rharg(seq id no:50)和二甲双胍的联合治疗呈现出逆转非酒精性脂肪肝的极端积极作用。低剂量n-abd094-rharg(seq id no:50)或正常剂量二甲双胍的单一治疗在不存在自噬的情况下也具有降低肝脏脂质的轻微作用,这表明脂肪吞噬不是降低肝脏脂质含量的唯一机制。综上所述,通过n-abd094-rharg(seq id no:50)和二甲双胍的联合治疗诱导自噬可以极大地帮助去除肝细胞中的脂滴和逆转脂肪肝。
[0309]
自噬的诱导被认为与mtor磷酸化的抑制相关(图65a),它是主要的精氨酸传感器和自噬抑制物。据报道,抑制mtor通路可增强细胞自噬。在bat中也观察到类似的mtor和下游s6k1抑制(图65b),表明自噬也可能在bat中发生。
[0310]
实施例21.组合n-abd094-rharg(seq id no:50)和全反式视黄酸(ra)在诱导喂食hfd的c57bl/6j雄性小鼠的间歇性禁食方面的协同作用除了二甲双胍之外,我们发现当与视黄酸(ra)组合时,n-abd094-rharg(seq id no:50)在诱导间歇性禁食和减少肥胖方面也可以表现出协同相互作用(图66)。在不同条件下治疗五组喂食hfd持续12周的预先存在的c57bl/6j雄性小鼠。对于联合组,每周给小鼠注射一半剂量的n-abd094-rharg(seq id no:
50,200u),同时每天口服一次悬浮在花生油中的0.33mg ra[hfd(rharg+ra)]。对于对照组,每周给c57bl/6j肥胖雄性小鼠注射一半剂量的n-abd094-rharg(seq id no:50),同时每天喂食花生油[hfd(rharg)]或每周注射生理盐水以及每天喂食0.33mg ra[hfd(ra)]。没有n-abd094-rharg(seq id no:50)和ra治疗的肥胖小鼠用作阴性对照[hfd(载体)],而具有饲料[饲料(载体)]的瘦小鼠用作正常对照。
[0311]
在与二甲双胍联合实验类似的每日食物摄入(图66b)方面,载体治疗组或ra治疗组的食物摄入稳定,并且他们每天消耗约2.5至3克食物颗粒。当小鼠每周单独用300u n-abd094-rharg(seq id no:50)治疗时,即使是一半剂量,我们也可以观察到弱的7天间歇性禁食周期,但食欲抑制的程度不如全剂量600u/周那么强。有趣的是,当小鼠接受每周300u n-abd094-rharg(seq id no:50)和每天0.33mg ra的药物治疗时,我们可以观察到非常清晰的间歇性禁食周期,其强度与先前所述的600u n-abd094-rharg(seq id no:50)的全剂量给药一样强(图66b)。与食物消耗平行,我们可以观察到饲料组的平均体重非常稳定,在治疗期间保持在约30克的水平,而hfd组小鼠在连续喂食11周的hfd后,从开始时的约52g持续小幅增加至约60g(图66b)。每周单独用300u n-abd094-rharg(seq id no:50)进行单一药物治疗或每天单独喂食0.33mg ra持续约11周的小鼠也表现出稳定的体重,即使喂食hfd,体重也不会进一步增加(图67)。相比之下,小鼠接受n-abd094-rharg(seq id no:50)和ra联合治疗11周,我们可以观察到小鼠的平均体重在6周内持续从52g下降到30g,然后保持稳定状态直至11周治疗结束(图67)。n-abd094-rharg(seq id no:50)和ra的联合使用对小鼠体重减轻具有优异的协同作用。
[0312]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd,adc-peg或adc-abd)的治疗与ra组合时可对间歇性禁食产生协同作用。
[0313]
实施例22.组合n-abd094-rharg(seq id no:50)和全反式视黄酸(ra)在喂食hfd的c57bl/6j雄性小鼠中减少器官脂肪量、逆转胰岛素抵抗和葡萄糖不耐受方面的协同作用
[0314]
伴随体重减轻,在n-abd094-rharg(seq id no:50)和ra的联合治疗中,内脏(性腺周围、肾周和肠系膜脂肪垫)和皮下(腹股沟脂肪垫)脂肪量显著减少(图69)。单独使用n-abd094-rharg(seq id no:50)或ra不会减少体脂,这伴随着体重不变。如前所述,用载体治疗的dio小鼠的肝脏、肾脏、胰腺和bat的重量显著增加。我们的结果表明,单独的n-abd-rharg(seq id no:50)或ra已经可以减少包括肝脏、胰腺和bat在内的大多数器官的质量,但减少的幅度不如n-abd-rharg(seq id no:50)和ra的组合那么强(图53b)。
[0315]
在逆转先前存在的肥胖小鼠的胰岛素抵抗方面,采用了胰岛素耐量试验(itt)(图68a)。如前所述,喂食hfd的小鼠表现出胰岛素反应受损。结果显示,治疗6周后,单独的300u/周的n-abd-rharg(seq id no:50)或0.33mg/天的ra治疗对改善胰岛素敏感性有边际作用,但没有达到统计学显著性。而相比之下,通过n-abd-rharg(seq id no:50)和ra的联合治疗检测到胰岛素敏感性的显著增强,达到与正常瘦对照小鼠相似的敏感性水平。
[0316]
喂食hfd的小鼠也表现出葡萄糖耐量受损(图68b)。结果显示,在治疗7周后,0.33mg/天的ra或300u/周的n-abd-rharg(seq id no:50)单独治疗依然不能缓解葡萄糖不耐受。另一方面,通过n-abd-rharg(seq id no:50)和ra的联合治疗检测到葡萄糖耐量的显著增强,达到与正常瘦对照小鼠相似的水平。综上所述,对小鼠联合使用精氨酸酶和ra可显
著提高胰岛素敏感性并改善葡萄糖耐量,最终可逆转肥胖小鼠的2型糖尿病。
[0317]
实施例23.组合n-abd094-rharg(seq id no:50)和全反式ra(ra)在诱导喂食hfd的c57bl/6j雄性小鼠的肝脏中的自噬方面的协同作用
[0318]
通过透射电子显微镜检查治疗3周后肝脏中自噬体的存在(图71和72)。如前所述,在长期喂食hfd后,肝脏大小增加(图70),这与载体治疗的dio中大脂滴的积累相关。如通过tem观察到的,喂食hfd的小鼠的肝脏呈现出大量增大的脂滴(图71)。每天0.33mg ra或300u/周的n-abd-rharg(seq id no:50)的单独治疗已经证明脂滴大小显著减小,其中单独使用ra的效果比单独使用n-abd-rharg更显著。然而,在两种治疗条件下均未观察到自噬体(图71)。相比之下,喂食hfd并同时用n-abd094-rharg(seq id no:50)和ra治疗的小鼠的肝脏在治疗的第3天,即间歇性禁食周期的禁食期可以明显清除肝细胞中的大部分脂滴(图72),结果呈现出逆转非酒精性脂肪肝的极端积极作用。此外,由于观察到大量活跃的自噬体,因此在细胞中触发了自噬(图71)。在4000倍放大倍率下,我们发现活跃的脂肪吞噬通过产生自噬体而发生,这些自噬体隔离部分大脂滴以形成双膜囊泡,从而将脂滴分解成更小且更易消化的大小(图71)。
[0319]
治疗11周后肝脏的h&e染色(图70)显示n-abd094-rharg(seq id no:50)和ra的联合治疗。低剂量n-abd094-rharg(seq id no:50)或正常剂量ra的单一治疗也对减少肝脏中的脂质具有显著效果。另一方面,n-abd094-rharg(seq id no:50)和ra联合治疗可以完全去除肝细胞中的脂滴,从而逆转脂肪肝。
[0320]
总之,这些发现支持abd-rharg融合蛋白或其他形式的精氨酸酶(例如,rharg-peg、bca-peg或bca-abd)或精氨酸消耗酶(例如,adi-peg、adi-abd、adc-peg或adc-abd)的治疗与ra组合时,可在减少器官中的脂肪堆积和改善胰岛素敏感性和葡萄糖不耐受方面产生协同作用。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1