病毒复制抑制剂及其应用
1.本发明涉及分子免疫学和病毒学技术领域,具体涉及一种病毒复制抑制剂及其应用。
背景技术:
2.马立克氏病(marek’s disease,md)是由马立克氏病病毒(marek’s disease virus,mdv)引起的一种鸡的恶性淋巴肿瘤性疾病,同时还能引起被感染鸡只的免疫抑制。1907年匈牙利病理学家jozef marek首先发现了因单核细胞浸润于外周神经及脊柱神经根下而导致公鸡的坐骨神经肿大和瘫痪。1961年,在英国科学家biggs的倡议下,开始使用马立克氏病这一名称。1967年,英国学者分离获得马立克氏病病毒。上世纪70年代以来,尽管家禽养殖广泛使用了各种mdv疫苗。但在全世界范围内本病仍是养禽业健康发展的主要危胁。1984年,仅美国因mdv直接死亡和养殖业主放弃的病鸡损失达1200万美元,若加上疫苗及其接种的费用和产蛋量的下降,损失就高达1.69亿美元,全世界因马立克氏病的损失高达9.43亿美元。随着各类mdv疫苗制剂的广泛使用,马立克氏病毒出现了野毒株与疫苗株重组,疫苗株毒力增强,随之频频出现mdv疫苗失败,感染受污染的疫苗株及新型野毒株的鸡群生产性能严重下降问题日益凸显。疫苗无法对新型mdv产生保护性等诸多问题,马立克氏病的防控出现了严峻的挑战。生产中对预防mdv感染并降低mdv对鸡群生产性能影响的生物制剂需求迫切。
3.先天免疫反应是宿主抵抗病原微生物入侵的首要屏障,当病原微生物入侵机体的时候,其所包含的物理屏障,急性反应期蛋白,特异性免疫反应会依次发挥作用抵抗病原微生物入侵。当病原微生物侵入时,许多急性反应期蛋白都会迅速大量升高并发挥各种直接或间接的抗病毒作用,例如直接与病毒结合抑制病毒附着或复制,增强宿主机体免疫能力,创造不利于病原微生物生存的为生态环境等来增强机体的抗病能力。同时,先天免疫蛋白是宿主本身产生的,宿主不会对其产生较强的排异反应,先天免疫蛋白在抗病毒领域有着巨大的开发应用前景。
4.研发预防和抑制mdv感染,降低mdv对生产性能造成影响的先天免疫蛋白,对于md的防控有着重要而紧迫的现实意义。
技术实现要素:
5.针对上述现有技术,本发明的目的是提供一种病毒复制抑制剂及其应用。本发明发现,血清淀粉样蛋白a(serum amyloid a,saa)可以作为病毒复制抑制剂,能直接、显著的抑制mdv复制,且对细胞无毒副作用;可开发成mdv的治疗药物,具有广阔的应用前景。
6.为实现上述目的,本发明采用如下技术方案:
7.本发明的第一方面,提供血清淀粉样蛋白a在如下(1)或(2)中的应用:
8.(1)制备预防和/或治疗马立克氏病病毒所致疾病的药物;
9.(2)制备抑制马立克氏病病毒复制的药物;
10.所述血清淀粉样蛋白a是如下任一所示的蛋白质:
11.a1)氨基酸序列为seq id no.1的蛋白质;
12.a2)在seq id no.1所示的氨基酸序列的n端或/和c端连接标签得到的融合蛋白。
13.其中,a1)和a2)所述的蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
14.上述蛋白中,蛋白标签是指利用dna体外重组技术,与目的蛋白一起融合表达的一种多肽或者蛋白,以便于目的蛋白的表达、检测、示踪和/或纯化。其中,为了使(a1)中的蛋白质便于纯化,可在(a1)的蛋白质的氨基末端或羧基末端连接上标签。所述标签可以为poly-arg(通常为6个rrrrr),poly-his(通常为6个hhhhhh),flag(dykddddk),strep-tag ii(wshpqfek)或c-myc(eqkliseedl)。
15.本发明的第二方面,提供编码血清淀粉样蛋白a的基因在如下(1)或(2)中的应用:
16.(1)制备预防和/或治疗马立克氏病病毒所致疾病的药物;
17.(2)制备抑制马立克氏病病毒复制的药物。
18.上述应用中,所述编码血清淀粉样蛋白a的基因为如下i)或ii)或iii)所示的核酸分子:
19.i)核苷酸序列是seq id no.2所示的核酸分子;
20.ii)与i)的核苷酸序列具有90%或90%以上同一性且表达相同功能蛋白质的核酸分子;
21.iii)除i)以外的编码seq id no.1所示氨基酸序列的核酸分子。
22.所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
23.这里使用的术语“同一性”指与天然核酸序列的序列相似性。同一性可以用计算机软件进行评价,例如可采用blast算法测定(altschul et al.1990.journal of molecular biology 215:403-410;karlin and altschul.1993.proceedings of the national academy of sciences 90:5873-5877)。
24.上述核酸分子中,所述90%或90%以上同一性可以为至少90%、91%、92%、95%、96%、98%或99%的同一性。
25.本发明的第三方面,提供与血清淀粉样蛋白a相关的生物材料在如下(1)或(2)中的应用:
26.(1)制备预防和/或治疗马立克氏病病毒所致疾病的药物;
27.(2)制备抑制马立克氏病病毒复制的药物。
28.上述应用中,所述与血清淀粉样蛋白a相关的生物材料为如下任一所示:
29.b1)含有血清淀粉样蛋白a编码基因的重组表达载体;
30.b2)含有血清淀粉样蛋白a编码基因的重组微生物;
31.b3)含有血清淀粉样蛋白a编码基因的转基因细胞系;
32.所述血清淀粉样蛋白a编码基因的核苷酸序列如seq id no.2所示。
33.上述应用中,可以用现有的表达载体构建含有血清淀粉样蛋白a编码基因的重组表达载体。现有的表达载体可以为真核表达载体,例如pex-3载体(pgcmv/mcs/neo);也可以为原核表达载体,例如pet-22b(+)载体质粒。使用本发明的血清淀粉样蛋白a编码基因构建
重组表达载体时,还可使用增强子,包括翻译增强子或转录增强子等。
34.上述应用中,重组微生物具体可以为病毒(例如慢病毒)、酵母、细菌、藻类或真菌等。
35.本发明的第四方面,提供一种抑制马立克氏病病毒复制的药物,所述药物以有效量的血清淀粉样蛋白a为活性成分。
36.优选的,所述药物中,血清淀粉样蛋白a的浓度为5μg/ml-10μg/ml。
37.进一步的,所述药物中还可以含有:增强或提高血清淀粉样蛋白a活性的物质、提高血清淀粉样蛋白a含量的物质、提高血清淀粉样蛋白a编码基因表达量的物质中的一种或多种。
38.本发明的有益效果:
39.本发明首次研究发现血清淀粉样蛋白a可以作为病毒复制抑制剂,能直接地、显著地抑制mdv复制,收取病毒时与对照组相比病毒载量降低了3倍以上,并且对细胞无毒副作用。
40.本发明的血清淀粉样蛋白a可作为抗md药物广泛用于家禽业防御mdv的生产实践中,具有重大的经济价值及研究意义。
附图说明
41.图1:mdv激活saa的表达相关示意图;
42.a:感染mdv后不同时间段mdv病毒滴度图;b:感染mdv后不同时间段mdv meq蛋白表达量图;c:感染mdv后不同时间段saa的mrna表达水平图。
43.图2:过表达saa显著增加mdv载量相关示意图,
44.a:转染空载质粒和saa过表达质粒后,感染mdv维持36h后saa的mrna表达量;b、c、d:转染空载质粒和saa过表达质粒后,感染mdv维持36h后mdv的病毒滴度(c),mdv meq蛋白的mrna表达量(b)以及蛋白表达量(d)。
45.图3:干扰saa明显促进mdv的复制相关示意图;
46.e:转染无序干扰rna和saa干扰rna后,感染mdv维持36h后saa的mrna表达量;f、g和h:转染无序干扰rna和saa干扰rna后,感染mdv维持36h后mdv的病毒滴度(g),mdv meq蛋白的mrna表达量(f)以及蛋白表达量(h)。
47.图4为本发明构建的pet22b-his-saa重组蛋白表达质粒模式图。
48.图5为本发明重组saa纯化后sds-page检测图,如图5所示,重组saa纯化效果良好,仅有单一条带。
49.图6为经bca蛋白定量检测试剂盒绘制标准曲线。
50.图7为本发明复性后重组saa对细胞毒性cck-8检测图,如图不同浓度的saa对cef细胞均未产生较大的细胞毒性,
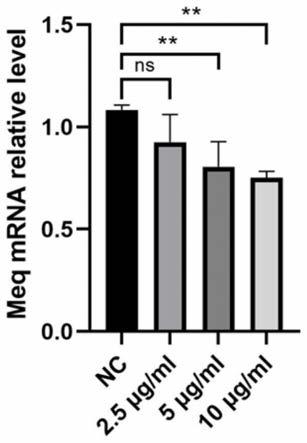
51.图8为本发明24h、复性后不同浓度的重组saa在cef细胞上对mdv转录的影响实时荧光定量pcr检测结果图,如图8所示,24h时,重组saa可以抑制mdv转录,且具有浓度依赖性。
具体实施方式
52.应该指出,以下详细说明都是例示性的,旨在对本技术提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本技术所属技术领域的普通技术人员通常理解的相同含义。
53.如前所述,md是养鸡业所面临的最重要的疾病之一。目前防治md采用的主要方法为疫苗免疫。但随着mdv毒力的不断增强,mdv疫苗失效,感染受污染的疫苗株以及新型野毒株的鸡群生产性能严重下降的问题日益凸显。
54.基于此,为解决当前养鸡业在防治md中所面临的问题,发明人从先天免疫蛋白入手,开发了一种能够抑制mdv复制的抑制剂,为md的防治提供了新的药物来源和防治手段。
55.本发明的抑制mdv复制的抑制剂是以血清淀粉样蛋白a(serum amyloid a,saa)为活性成分。saa蛋白的相对分子量约13kda,由127个氨基酸构成外分泌型糖蛋白,其氨基酸序列如seq id no.1所示,具体如下:
56.mrlcicfvll sivlyasadn pfssgikfvr daaggardmw rayrdmrean yigadkyfha rgnydaarrg pggawaakvi sdaregwqsr vsgrgaedtr ldqeanewgr rggdpnrfrp aglpsky。
57.与目前已知的蛋白同源性极低,saa蛋白在物种之间比较保守。saa是细胞核内被转录,被激活后分泌到细胞浆中,最后到达细胞外基质中。在现有技术中并没有血清淀粉样蛋白a与mdv相关的报道出现,两者之间关系的研究处于空白状态。
58.基于发明人的诸多努力,通过大量实验证明了saa具有抵抗mdv复制的作用,并且建立了该蛋白可以作为mdv复制抑制剂的制备和使用方法。为后续生产实际中利用其达到预防,抵抗mdv病毒感染,降低mdv对生产性能的影响提供了理论和研究基础。
59.本发明具体思路如下:
60.通过荧光定量pcr试验,对saa的表达量进行体外验证,结果表明:mdv激活了saa的表达;为验证saa是否可以抑制mdv复制依赖性,发明人构建了saa过表达载体。cef细胞过表达saa重组质粒后,接种mdv,实验结果显示:过表达saa可以极显著降低细胞中病毒载量;相反,敲低saa后,mdv的病毒载量显著上升;我们进一步原核表达了saa蛋白,通过细胞毒性实验证明了原核表达的saa蛋白对于细胞没有毒性作用,之后将其加入到cef细胞中发现mdv的复制被抑制,且具有浓度依赖性。结果显示:saa具有抑制mdv复制的作用。
61.为了使得本领域技术人员能够更加清楚地了解本技术的技术方案,以下将结合具体的实施例详细说明本技术的技术方案。
62.本发明实施例中所用的未进行具体说明试验材料均为本领域常规的试验材料,均可通过商业渠道购买得到。本发明实施例中未注明具体实验条件和方法的,通常按照常规条件,如j.萨姆布鲁克等主编,科学出版社,2002,分子克隆实验指南(第三版);d.l.斯佩克特等主编,科学出版社,2001,细胞实验指南;或者按照制造厂商建议的条件。
63.本发明实施例中所使用的mdv为马立克病病毒强毒株md5,来自山东农业大学,公众自申请日起20年内从申请人处获得该毒株以用于重复本发明。
64.实施例1:mdv激活saa的表达
65.为了确证mdv对saa的激活作用,本实施例建立了mdv感染的体外细胞模型。通过qpcr检测mdv感染后saa表达量的变化。
66.(1)mdv病毒滴度测定
67.收集mdv md5毒株接毒后不同时间点的细胞,将不同时间点的样品进行1000倍稀释,接种到铺满鸡胚成纤维细胞(cef)单层的六孔板中,每个稀释度做3个重复,接毒96h后,分别计数不同稀释度孔内的蚀斑数,取其平均数,即为该时间点的病毒滴度。
68.结果如图1a所示;(a)提取md5感染不同时间点的cef细胞,在将其接种到正常cef细胞上,计算不用时间点md5的病毒滴度。兙
69.俥图1a作为辅助数据说明,本实施例接毒的可靠性。
70.(2)western blot
71.将md5感染6h、12h、24h、36h、48h、72h的细胞收获,将放射免疫沉淀法缓冲液(ripa)与苯甲基磺酰氟(pmsf)按照100:1的体积比混合裂解细胞;调整各蛋白样品为均一浓度,加入蛋白上样缓冲液于100℃水中加热5min。样品加入电泳仪,浓缩胶在80v状态下稳压进行,分离胶在110v状态下稳压进行。电泳完成后,将蛋白胶上的蛋白转移到0.22μm pvdf膜上,使用5%脱脂奶粉37℃封闭2h,tbst洗涤后加入鼠源抗mdv meq蛋白抗体后37℃孵育1h,tbst洗涤后,加入羊抗鼠抗体37℃孵育1h,最后使用western blot显影仪进行观察。
72.如图1b所示,(b)western blot检测mdv感染细胞和正常细胞中mdv meq蛋白表达量。通过图1b能够说明,本实施例接毒的可靠性。
73.(3)荧光定量pcr/qpcr
74.设计qpcr特异性引物:禽宿主蛋白saa qpcr引物序列为:
75.f:5
’‑
ttctccagcggtatcaagtt-3’,(seq id no.3)
76.r:5
’‑
ccgagcagcatcgtaatt-3’;(seq id no.4)
77.禽内参基因gpadh的引物序列为:
78.f:5
’‑
gaacatcatcccagcgtcca-3’,(seq id no.5)
79.r:5
’‑
cggcaggtcaggtcaacaac-3’。(seq id no.6)
80.meq荧光定量引物序列为:
81.f:5
′‑
ttgtcatgagccagtttgccctat-3
′
,(seq id no.7)
82.r:5
′‑
tgagggaggtggaggagtgcaaat-3
′
。(seq id no.8)
83.引物由青岛华大基因生物技术有限公司合成。
84.将md5感染6h、12h、24h、36h、48h、72h的cef细胞收获,使用细胞rna总提试剂盒(qiagen,america)提取细胞总rna,然后将rna逆转录为cdna,以此为模板通过qpcr检测saamrna的表达变化。
85.反应体系为:在20μl反应体系中进行:10μl sybr premix ex taqtm(2
×
),0.4μl上游引物,0.4μl下游引物,1μl cdna,8.2μl ddh2o。
86.反应条件为:采用2步法pcr反应条件:95℃预变性30s;95℃变性5s;64℃退火34s;95℃变性10s,65℃退火60s,97℃延伸1s,变性-退火-延伸重复38个循环。
87.如图1c所示,(c)提取md5感染cef细胞和正常cef细胞rna。通过qpcr检测saa mrna表达变化。
88.图1c能够说明mdv md5强毒株感染cef细胞早期可以激活cef细胞中saa的表达。
89.实施例2:过表达saa显著抑制mdv强毒株md5病毒复制
90.为了明确激活saa能够抑制md5复制,本实施例构建了saa过表达质粒,通过过表达
saa,验证saa对md5病毒复制的抑制作用。
91.1.过表达saa检测对mdv复制的影响
92.(1)进行真核表达载体的构建。saa真核表达质粒的构建交由吉玛基因公司采用现有技术完成,本发明选用pex-3载体(pgcmv/mcs/neo)真核表达质粒表达saa。
93.(2)然后将构建的saa真核表达质粒转染cef细胞;
94.1)转染前细胞培养于12孔板,确保细胞保持最佳浓度和状态;
95.2)试剂复温:x-tremegene hp dna transfection reagent、pex-saa真核表达质粒或pex-3空载体和稀释液升温至20℃左右,短暂漩涡混匀x-tremegene hp dna transfection reagent;
96.3)溶液配制:使用opti-mem培养基作为稀释液,将saa过表达质粒稀释至终浓度为1μg/100μl培养基,轻柔混匀。将含有1μg saa过表达质粒的100μl稀释液分别加入无菌离心管中。x-tremegene hp dna transfection reagent直接添加到含有稀释dna的培养基中,质粒dna和转染试剂的比例为1:3,在此过程中,枪头切勿接触离心管管壁;使用的稀释剂体积多于100μl;
97.4)孵育:转染复合物在20℃左右环境中孵育15min;
98.5)转染:培养箱中取出的细胞,无需弃掉原有培养基,转染复合物直接滴加到细胞中;
99.6)mdv感染细胞:当cef细胞瞬时转染saa过表达质粒6h后,弃掉原有培养基,使用pbs洗涤3次,接种mdv,每孔加入1ml带有md5株的dmem培养基,37℃维持36h,收获细胞。提取细胞rna、裂解细胞蛋白、分别通过qpcr检测saa表达水平,通过virus titer、qpcr、western blot检测mdv复制水平。
100.结果如图2a-d所示。(a)cef细胞转染pex-saa或对照质粒pex-3后接种md5,维持36h后提取细胞rna。通过qpcr检测saa mrna水平,结果显示,过表达saa组细胞内saamrna的表达量显著高于对照组。(b)cef细胞转染pex-saa或pex-3并接种md5 36h后通过qpcr检测其meq mrna水平,结果显示,过表达saa组细胞mdv meq蛋白mrna的表达量显著低于对照组。(c)cef细胞转染pex-saa或对照质粒pex-3后接种mdv,维持36h。通过病毒滴度实验检测不同组cef细胞中mdv的病毒滴度,结果显示,过表达saa组mdv病毒滴度显著低于对照组。(d)cef细胞转染pex-saa或对照质粒pex-3后接种mdv,维持36h。通过western blot检测细胞内mdv meq蛋白的表达量,结果显示,过表达saa组细胞内meq蛋白的表达量显著低于对照组。
101.通过图2a-d证明,与对照组相比,过表达saa能够抑制mdv强毒株md5病毒复制。通过以上结果说明,过表达saa能够显著抑制强毒株md5复制。
102.实施例3:干扰saa显促进mdv的复制
103.为了明确saa的表达量对mdv载量的重要性,本实施例构建了saa干扰rna,通过敲低saa,验证saa对宿主细胞抵抗mdv复制的重要性。
104.1.saa干扰质粒的构建
105.(1)对saa抑制mdv病毒复制的结果通过sirna进行反向验证,由吉玛公司构建了3条saa干扰rna,其核苷酸序列如下所示;
106.sirna-1:
107.正义链:ccagcgguau caaguuugut t;(seq id no.9)
108.反义链:acaaacuuga uaccgcuggt t。(seq id no.10)
109.sirna-4:
110.正义链:gugcugacaa guauuuccat t;(seq id no.11)
111.反义链:uggaaauacu ugucagcact t。(seq id no.12)
112.sirna-nc:
113.正义链:uucuccgaac gugucacgut t;(seq id no.13)
114.反义链:acgugacacg uucggagaat t。(seq id no.14)
115.2.干扰saa检测对mdv复制的影响
116.(1)转染前细胞培养于12孔板,确保细胞保持最佳浓度和状态;
117.(2)试剂复温:x-tremegene hp dna transfection reagent、三种saa干扰sirna和稀释液升温至20℃左右,短暂漩涡混匀x-tremegene hp dna transfection reagent;
118.(3)溶液配制:使用opti-mem培养基作为稀释液,将含有1μg saa干扰质粒的100μl稀释液分别加入无菌离心管中。x-tremegene hp dna transfection reagent直接添加到含有稀释dna的培养基中,质粒dna和转染试剂的体积比例为1:3,在此过程中,枪头切勿接触离心管管壁;使用的稀释剂体积多于100μl;
119.(4)孵育:转染复合物在20℃左右环境中孵育15min;
120.(5)转染:培养箱中取出的细胞,无需弃掉原有培养基,转染复合物直接滴加到细胞中;
121.(6)mdv感染细胞:细胞瞬时转染6h后弃掉原有培养基,使用pbs洗涤3次,接种mdv强毒株md5毒株,37℃维持36h。
122.(7)检测mdv载量:收获细胞,分别通过qpcr、virus titer、western blot检测mdv病毒复制量。
123.结果如图3e-h所示。(e)用sirna(sirna-1,sirna-4或sirna-nc)转染cef细胞随后接种mdv,通过qpcr检测saa表达量,结果显示,转染sirna-4组的细胞内saa表达量低于对照组。(f)sirna转染cef细胞(sirna-1,sirna-4或sirna-nc)后,mdv感染细胞并提取细胞rna。通过qpcr检测mdv meq mrna表达水平,结果显示,转染sirna-4组的细胞内mdv meq mrna表达量高于对照组,而转染sirna-1组的细胞内mdv meq mrna表达量与对照组相比没有差异显著性。(g)cef细胞转染sirna-1,sirna-4或sirna-nc后接种mdv,维持36h。通过病毒滴度实验检测不同组cef细胞中mdv的病毒滴度,结果显示,sirna-4组mdv病毒滴度显著高于对照组。(h)sirna-1,sirna-4或sirna-nc转染细胞,然后mdv感染细胞,通过wb检测mdvmeq蛋白表达量,结果显示,转染sirna-4组的细胞mdv meq蛋白表达量显著高于对照组。图3e-h证明,敲低saa能够促进mdv复制。通过以上结果说明,saa具有抑制mdv强毒株md5复制的能力。
124.实施例4:重组质粒的获得和转化
125.发明人基于saa的氨基酸序列,如seq id no.1所示,获得了编码该氨基酸的核苷酸序列,如seq id no.2所示;在此基础上,发明人通过艾伯森公司构建了获得了pet22b-his-saa重组质粒(pet22b-his-saa重组质粒的结构示意图如图4所示)。
126.将上述pet22b-his-saa重组质粒转化到大肠杆菌dh5α感受态细胞中,通过含有氨苄霉素的lb固体培养基中筛选出具有阳性质粒的大肠杆菌,通过测序筛选出含有saa重组表达质粒的大肠杆菌,具体转化步骤如下所示:
127.将大肠杆菌dh5α感受态细胞置于冰上融化,向100μl大肠杆菌dh5α感受态细胞中加入5μl pet22b-his-saa重组质粒,轻轻旋转离心管以混匀内容物,冰浴放置30min。将离心管置于42℃水浴中放置60~90s,然后快速转移到冰浴中放置2~3min,注意不要摇动离心管;向离心管中加入500μl无菌无抗的lb培养基,37℃摇床培养1h。取适量已转化的感受态细胞涂布到含有amp抗性的lb平板中,37℃倒置培养12~16小时,挑取单一菌落进行大量培养。
128.再从上述含有saa表达质粒的大肠杆菌中提取质粒,将其转化到大肠杆菌bl21感受态细胞中,转化步骤参照上述步骤进行即可。通过含有100μg/ml氨苄霉素的lb固体培养基中筛选出具有阳性质粒的大肠杆菌,筛选出能够表达saa的大肠杆菌,剩余菌液保存至-80℃冰箱备用。
129.实施例5:saa的表达,纯化及复性
130.1.saa的表达
131.将能够表达saa的大肠杆菌的菌液(阳性菌液),按照amp:阳性菌液:lb液体培养基=1:10:1000(体积比)的比例混合,于37℃摇床上扩繁并表达saa。
132.制备具体过程如下:
133.取500ml锥形瓶分别加入100ml lb液体培养基、100μl amp(氨苄霉素)、1ml实施例4构建的能够表达saa的大肠杆菌于37℃220r/min的摇床上培养6h得到菌液;取沉淀,用pbs重悬沉淀后用超声破碎仪破碎,破碎3s,间隔4s,重复99次为一个循环,破碎菌体至溶液透光即可。4℃、12000r/min离心15min,弃去上清,加pbs重悬沉淀后使用相同方法离心,清洗沉淀。
134.2.saa的纯化
135.用镍离子亲saa和层析柱纯化蛋白,加入足量的le buffer重悬沉淀,置于4℃冰箱中过夜溶解包涵体并使蛋白变性,将过夜溶解后的包涵体4℃、12000r/min离心5min,弃掉沉淀,将溶液加入层析柱,控制流速载3~5s/滴,收集流出液,如有必要,可以使流出液多次上样,提高收率。
136.上样结束后使用2倍柱体积的洗涤缓冲液(其中含有较低浓度的imidazole可以将杂蛋白洗掉从而只保留目的蛋白)洗涤层析柱,使用2ml的洗脱缓冲液加入层析柱中,将流出的洗脱液反复上样5-10次,最终得到的洗脱液中含有纯化后的saa,可使用sds-page检测纯化效率。
137.上述过程中采用的各种试剂如下:
138.lysis(le buffer):100mm na2hpo4、10mm tris
·
cl、8m urea、ph=8.0;
139.洗涤缓冲液:100mm na2hpo4、10mm tris
·
cl、10mm imidazole、8m urea、ph=8.0;
140.洗脱缓冲液:100mm na2hpo4、10mm tris
·
cl、250mm imidazole、8m urea、ph=8.0;
141.3.纯化后saa的检测(sds-page)
142.sds-page检测纯化后saa,步骤如下:
143.(1)首先配制12%分离胶,用底座的夹子固定洗干净的玻璃板(垂直放置),先加入蒸馏水检查固定板是否完全固定。然后倒掉蒸馏水后将按照比例配好的凝胶用1ml的移液枪快速注入到两个玻璃板之间,在分离胶的上端加入一定量的蒸馏水进行封闭,以确保分
离胶平整凝固。等35min待分离胶完全凝固之后,倒掉分离胶上层的水,残留的水分用滤纸尽量吸干。然后加入配置好的上层浓缩胶,最后插入孔径大小适合的梳子(1mm),这个过程中注意防止气泡的产生,等待其完全凝固后即可使用(约30min)。
144.(2)取纯化后saa加入1/5体积的5x蛋白上样缓冲液于pe管中,蒸馏水煮沸5min。
145.(3)拔掉浓缩胶的梳子,向电泳槽中加入提前配好的1xtris-甘氨酸蛋白电泳缓冲液,使电泳缓冲液完全淹没加样孔,用10μl移液器上样,20μl/孔加样。将电泳槽正负极和电泳仪正确连接,打开电源。先用80v电压进行电泳,待样品在浓缩胶上浓缩成一条直线到达浓缩胶底部快进入分离胶时,调高电压至100v,样品至分离胶的底部时停止电泳。
146.(4)染色:电泳完毕,用刀片小心切下凝胶,放到加有考马斯亮蓝染色液的培养皿中,染色40min。
147.(5)脱色:染色结束以后,将凝胶置于脱色液中,在摇床上缓慢摇动,定时更换2次以上脱色液至到凝胶蓝色背景完全脱去后,放入蒸馏水中终止脱色并拍照保存。
148.将纯化后的saa用sds-page检测,结果如图5所示,蛋白纯化效果良好。
149.4.saa的复性
150.采用不同浓度的尿素对变性的蛋白进行透析复性,具体步骤如下:
151.使用分子截流量合适的(分子量约为5kda)透析袋装取纯化后的saa分别置于复性液1-4(按照复性液1-4的顺序)中,每种复性液中停留3-4h,最后置于pbs中过夜,使变性的蛋白重新折叠,恢复其生物活性,从而表达出具有生物活性的saa。经bca蛋白定量检测试剂盒绘制标准曲线,如图6所示,检测小肽浓度为1029μg/ml,经计算每升菌液可以最终得到2-3mg saa小肽。
152.所采用的复性液组成如下:
153.复性液1:500mm tris
·
cl、100mm nacl、4m urea、ph=8.0
154.复性液2:500mm tris
·
cl、100mm nacl、3m urea、ph=8.0
155.复性液3:500mm tris
·
cl、100mm nacl、2m urea、ph=8.0
156.复性液4:500mm tris
·
cl、100mm nacl、1m urea、ph=8.0。
157.实施例6:重组saa体外抗马立克氏病毒实验
158.1.重组saa对细胞的增殖-毒性检测
159.在细胞上做重组saa抗mdv实验之前,先通过cell counting kie-8(cck-8)检测一个saa对cef细胞毒性影响。
160.ccck-8细胞增殖-毒性检测实验步骤:
161.在96孔板里配制100μl的df-1细胞悬液,将96孔板在细胞培养箱里培养24h;向96孔板孔里依次加入10μl不同浓度的saa蛋白(蛋白浓度依次是2.5μg/ml;5μg/ml;10μg/ml);将96孔板在细胞培养箱里孵育24h;向每孔中加入10μl cck-8溶液;将96孔板在细胞培养箱里孵育4h;用酶标仪测定在450nm处的吸光度,如图7所示,saa施加对cef细胞不会产生明显毒性。
162.为了确定saa能否体外抑制mdv感染,分别对这3个不同的浓度进行检测,分别是mdv接毒组、mdv+2.5μg/ml saa、mdv+5μg/ml saa、mdv+10μg/ml saa。
163.2.mdv病毒实时荧光定量pcr引物设计和合成
164.根据已经发表的mdv基因序列,合成实时荧光定量pcr引物,由华大公司合成。
165.禽内参基因gpadh的引物序列为:
166.f:5
’‑
gaacatcatcccagcgtcca-3’,(seq id no.5)
167.r:5
’‑
cggcaggtcaggtcaacaac-3’。(seq id no.6)
168.meq荧光定量引物序列为:
169.f:5
′‑
ttgtcatgagccagtttgccctat-3
′
,(seq id no.7)
170.r:5
′‑
tgagggaggtggaggagtgcaaat-3
′
。(seq id no.8)
171.3.重组saa体外抗mdv实时荧光定量pcr检测
172.将纯化复性后的重组saa进行cef细胞实验,将cef细胞传至12孔板中,在细胞密度达到待50%~60%时加入saa,以加入pbs的组别为阴性对照(nc),当细胞密度70%~80%时所有组接入mdv(md5),维持24h后分别收集细胞。使用细胞rna总提试剂盒(qiagen,america)提取细胞总rna,然后将rna逆转录为cdna。进行实时荧光定量pcr检测,如图8所示。
173.实时荧光定量pcr检测结果显示,加入saa浓度为10μg/ml时,重组saa体外抗mdv具有明显的效果,同时也可以确定重组saa体外抗mdv的最低浓度为5μg/ml。
174.以上所述仅为本技术的优选实施例而已,并不用于限制本技术,对于本领域的技术人员来说,本技术可以有各种更改和变化。凡在本技术的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本技术的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1