能显著提高棉花纤维长度的小G蛋白Rab基因的制作方法
本发明涉及一种棉花的小G蛋白Rab基因,该基因是通过cDNA差异显示技术发现,在陆地棉(G.hirsutum)李氏超短纤维突变体Li1及其纤维发育正常的野生型材料开花后10天的纤维/胚珠细胞中差异表达,进一步通过PCR技术在陆地棉遗传标准系TM-1中获得该基因全长ORF序列及基因组序列,属于生物技术应用领域。
背景技术:
棉花是世界性重要的经济作物,异源四倍体陆地棉占全球棉花种植面积的90%以上。棉花纤维是植物领域所知的最长单细胞,它由位于胚珠表皮的表皮毛原基分化发育而来。纤维生长以及形态建成由四个主要的发育时期组成:纤维起始期,伸长期,次生壁合成期以及脱水成熟期(Basra and Malik,1984,Wilkins and Arpat,2005)。不同纤维发育时期的转录组分析显示,在二倍体栽培种中,参与棉纤维发育的转录本大约涉及18 000个基因,而在四倍体栽培种中,参与棉花纤维发育的转录本约36 000个基因,包含来自A亚组和D亚组的部分同源基因(Wilkins and Arpat,2005)。因此,发掘与功能鉴定更多与纤维发育相关基因将有助于提高我们对棉花纤维发育机理的理解,同时,也有利于改良棉花纤维的产量和品质(郭旺珍等,2003)。
Rab家族属于小G蛋白Ras超家族(包括Ras、Rho/Rac、Ran、Arf、Rab五大类)中的一类。第一个Rab基因(YPT1)是1983年在酵母中发现(Gallwitz et al.,1983)。四年之后,A.Salminen和P.Novick第一次证明Ras蛋白(Sec4)参与囊泡运输过程(Salminen and Novick,1987)。同年,A.Tavitian和他的同事们第一次通过大鼠大脑的cDNA文库克隆到了与SEC4/YPT基因功能相似的同源基因,并命名为Rab(ras-likeinratbrain)(Martinez and Goud,1998)。
Rab蛋白家族成员大约有200个氨基酸,序列都比较保守,它们相互之间具有很高的序列相似性。所有的Rab蛋白都具有五个典型的保守结构域,包括G1、G3、G4、G5四个鸟嘌呤核苷酸结合结构域和G2一个效应因子结合结构域(Takai et al.,2001,Agarwaletal.,2009)。通常,Rab蛋白的C端氨基酸序列高度可变,但一般以两个保守的半胱氨酸残基(CC)结尾(Rutherford and Moore,2002),这两个非常保守的半胱氨酸残基在膜定位以及蛋白功能方面具有非常重要的作用。
Rab蛋白家族成员的主要功能是负责细胞内的蛋白运输,它们是细胞内囊泡运输途径必不可少的调节者(Novick and Zerial,1997;Brennwald,2000)。新合成的分泌蛋白一般会通过囊泡从细胞器的一个膜室运输到另一个膜室(Guakan et al.,2005),它们首先会被运输到内质网,然后通过高尔基体运输到质膜(Heo et al.,2005),有的还会送达前溶酶体。即使某些大分子被质膜吸收,它还可以通过囊泡运输到内吞体(endosmome)和溶酶体中。一些包括受体的细胞表面蛋白还可以通过内体循环再送回到质膜中,所以胞吞和胞吐的循环(Battey et al.,1999)都是通过细胞内的囊泡运输来实现的,而所有的这一切都是为了应答各种上游信号(如激素)的刺激(Koh et al.,2009)。一般而言,囊泡运输为细胞内各种生理功能所需,如细胞极性伸长、胞质分裂、细胞板的形成等(Barr,2009)。囊泡运输至少由4个步骤组成:首先是囊泡从供体膜上出芽,然后是囊泡向受体运动,随着囊泡停泊在受体膜上,囊泡与受体膜融合,上述过程都需要有Rab蛋白参与(Tuvim et al.,2001,Yang,2002)。
已有研究表明,小G蛋白的Rab家族对于花粉管极性生长十分重要。在烟草中,Rab11b的活化态及非活化态的过量表达,都会抑制花粉管的伸长,同时,也会影响花粉管生长的方向性(de Graaf et al.,2005)。在拟南芥中,RabA4d对于花粉管极性生长的正确调控十分必要,该基因在花粉中特异表达。亚细胞定位表明RabA4d标记的增强型黄色荧光蛋白定位于生长中花粉管的顶端,RabA4d的缺失会导致极性生长的破坏以及改变细胞壁的结构,说明RabA4d在调控花粉管极性生长中发挥重要作用(Szumlanski and Nielsen,2009)。
技术实现要素:
本发明的目的是提供一种棉花的小G蛋白Rab基因(GhRab11d3),并提供该基因的cDNAORF和基因组序列。以此基因为靶基因,通过基因工程的方法进行转基因功能验证,以便用于培育新种质并在生产上应用。
本发明的另一目的在于提供上述棉花的小G蛋白Rab基因所编码的蛋白。
本发明的又一目的在于提供上述棉花的小G蛋白Rab基因的应用。
本发明的目的通过以下技术方案实现:
一种棉花小G蛋白Rab基因(GhRab11d3),该基因的cDNA ORF序列如SEQ ID NO.1所示;基因组序列如SEQ ID NO.2所示。
上述的Rab基因编码的蛋白;其氨基酸序列如SEQ ID NO.3所示。
含有上述基因的重组载体、表达盒、转基因细胞系或重组菌。
上述的小G蛋白Rab基因在提高棉纤维长度、改善棉纤维品质或培育棉花新种质中的应用。
上述的重组载体、表达盒、转基因细胞系或重组菌在高棉纤维长度、改善棉纤维品质和培育棉花新种质中的应用。
本发明的优点表现在:
(1)本发明所克隆的基因编码小G蛋白,该Rab蛋白与棉纤维伸长时期的囊泡运输密切相关,是囊泡运输途径中的关键基因。在囊泡运输过程中,GhRab11d3会沿着细胞骨架蛋白Actin从内质网、高尔基体运输大分子多糖物质到细胞膜,最后通过外排作用将大分子多糖物质排出到细胞壁。在此过程中,一方面会影响到细胞中糖类物质积累量,另一方面也会影响到细胞骨架肌动蛋白F-actin微丝在纤维细胞中的数量和形态。过表达该基因可以使纤维细胞中糖类物质(果胶)积累增多,同时提高纤维细胞中细胞骨架肌动蛋白F-actin微丝的数量,从而促进棉纤维伸长。由此推断,以本发明棉花小G蛋白Rab基因作为靶基因进行转基因研究,有望改善棉花纤维品质。
(2)本发明所克隆的基因从结构上来看更类似于拟南芥的小G蛋白Rab基因,在棉花中前人还未研究过其功能。
(3)实时荧光定量PCR结果表明该基因在陆地棉遗传标准系TM-1纤维伸长期优势表达,该基因在不同纤维发育时期表达量有差异,在纤维起始时期即开始表达,在纤维快速伸长时期(开花后5、10天)表达量最高,在次生壁增厚时期(开花后15、20和25天)表达量逐渐下降,这一结果表明该基因与纤维发育伸长过程密切相关。
(4)构建植物正反义表达载体,以陆地棉材料W0为受体进行转基因研究,对转基因材料表达分析发现:与野生型相比,过表达株系中GhRab11d3表达量显著提高,转基因植株表现纤维变长的表型,抑制表达株系中GhRab11d的表达被显著抑制,转基因植株出现纤维变短的表型,表明该基因表达水平影响棉花纤维发育。
(5)对GhRab11d3转基因株系及非转基因野生型对照材料,选取开花后10天纤维进行肌动蛋白F-actin荧光染色分析,结果表明,过表达株系肌动蛋白F-actin微丝的含量显著增加,相反,抑制表达株系的肌动蛋白F-actin微丝的含量显著减少;同时,细胞骨架肌动蛋白F-actin微丝的形态结构也发生了变化,抑制表达株系纤维中的肌动蛋白F-actin微丝结构受损断裂,表明该基因影响细胞骨架肌动蛋白结构,进而影响纤维发育。
(6)对GhRab11d3转基因株系及非转基因野生型对照材料,选取开花当天胚珠及开花后10天纤维,利用FM4-64荧光染料进行染色发现,与野生型相比,在GhRab11d3过表达转基因材料中,无论是开花当天胚珠表面突起中,还是开花后10天的纤维顶端均聚集更多的囊泡,最终运输的糖类物质果胶含量显著增加。相反,在GhRab11d3抑制表达株系中,其相同发育时期囊泡数量明显减少,最终运输的糖类物质果胶含量显著降低,表明GhRab11d3在囊泡运输中发挥重要作用。
附图说明
图1 GhRab11d3和来源其它物种的Rab基因聚类分析。
利用GhRab11d3和其它物种的Rab基因其氨基酸序列比对和分析,说明不同物种间的Rab蛋白十分保守。
TcRabA4c(EOX96740.1),EgRabA4c(XP_010067891.1),PtRab(XP_002298447.1),PeRabA4d(XP_011025048.1),VvRGP1(XP_002264293.1),NtRGP1(XP_016459335.1),AtRabA4d(NP_187823.1),VvRabA4d(XP_002282798.1),TcRabA4d(EOY09425.1),AtRabA4c(NP_199607.1),GhRab11a(AF165095.1),GhRab11b(AF165096.1)
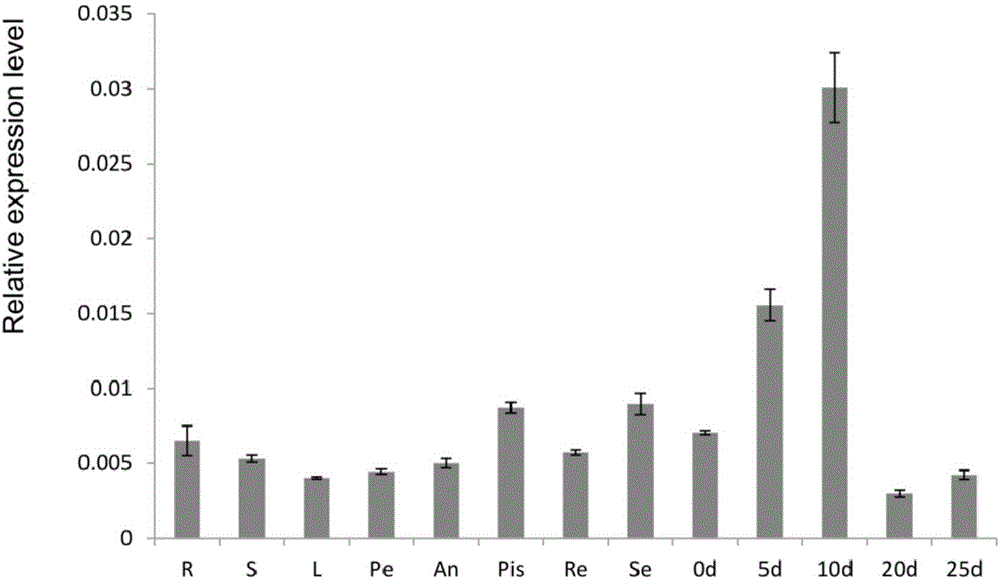
图2 GhRab11d3在棉花不同组织表达特征分析。
实时荧光定量PCR(qRT-PCR)检测棉花小G蛋白Rab基因(GhRab11d3)在棉花不同组织表达特点。R:根;S:茎;L:叶;Pe:花瓣;An:花药;Pis:雌蕊;Re:花托;Se:花萼;0d:0DPA胚珠;5d:5DPA纤维;10d:10DPA纤维;20d:20DPA纤维;25d:25DPA纤维。
图3 过量和抑制GhRab11d3表达的转基因棉花材料创制。
转基因植株PCR检测,利用启动子+目的基因的检测表明该基因正反义载体均已导入棉花。M、P、C分别为Marker、阳性对照和阴性对照;1-9为转基因植株目标基因PCR检测结果。
图4 GhRab11d3在转基因棉花中的表达分析。
定量qPCR分析GhRab11d3基因在3个不同表达载体(35S-SC、RDL-SC、RDL-ASC)转基因株系开花后5天、10天以及15天纤维中的表达。W0为受体对照,35S-SC和RDL-SC示目标基因过量表达载体,RDL-ASC示目标基因抑制表达载体,35S代表组成型表达启动子,RDL代表纤维发育优势表达启动子。28、347、372和303为35S-SC转基因材料;56、210和73为RDL-SC转基因材料;98、217和111为RDL-ASC转基因材料。均为T3代转基因纯合株系。
图5 转基因植株和受体W0的纤维长度比较。
在转基因纯系和W0中,开花后10、15、20天以及成熟时期纤维长度的表型分析。表明过表达植株的纤维长度显著变长,抑制表达植株的纤维长度显著变短。
图6 Rab11d3蛋白和四个Actin蛋白的互作。
Rab11d3与ACTa、ACTb、ACT2、ACT4在酵母中的互作验证。
图7转基因植株和受体W0开花后10天纤维中肌动蛋白F-actin微丝荧光染色分析。
表明过表达株系纤维中肌动蛋白F-actin微丝的含量显著增加,相反,抑制表达株系纤维中肌动蛋白F-actin微丝的含量显著减少;同时,细胞骨架肌动蛋白F-actin微丝的形态结构也发生了变化,抑制表达株系纤维中的肌动蛋白微丝结构受损断裂。
图8转基因株系和野生型开花当天和开花后10天纤维中囊泡荧光染色分析。
利用FM4-64染色,与野生型相比,在GhRab11d3过表达材料中无论是开花当天胚珠表面突起中还是开花后10天纤维顶端均聚集更多的囊泡,相反,在GhRab11d3抑制表达株系中囊泡数量明显减少。标尺=50μm。
图9转基因株系和野生型在开花后5天和10天纤维中果胶含量测定分析
与野生型相比,在GhRab11d3过表达材料中,无论是开花后5天还是开花后10天,果胶含量明显增加,相反,在GhRab11d3抑制表达株系中,果胶含量明显减少。
具体实施方式
(一)棉花小G蛋白基因GhRab11d3全长序列获得:
本实验利用cDNA差异显示技术,在陆地棉(G.hirsutum)李氏超短纤维突变体Li1及其纤维发育正常的野生型材料开花后10DPA的胚珠纤维复合体样品比较分析中,发现了一个在Li1野生型中,比超短纤维突变体中极显著高表达的EST序列,进一步克隆获得全长基因。通过同源比对,其编码Rab蛋白,进一步在Rab家族基因中查找,将其命名为GhRab11d3(如图1所示)。具体命名规则是:拟南芥中一共含有57个Rab蛋白基因家族成员。这57个Rab蛋白家族成员被分为八大类,分别命名为RabA、RabB、RabC、RabD、RabE、RabF、RabG以及RabH。在这八大类中,每一大类又分为若干小类,其中,RabA分为RabA1-RabA6六小类,RabB分为RabB1一小类,RabC分为RabC1和RabC2两小类,RabD分为RabD1和RabD2两小类,RabE分为RabE1一小类,RabF分为RabF1和RabF2两小类,RabG分为RabG1-RabG3三小类,RabH分为RabH1一小类,这样,拟南芥的RabA-RabH八大类又分为十八小类。相比拟南芥,雷蒙德氏棉数据库中有87个Rab蛋白基因家族成员,也被精确地分为八大类(依据最初Rab蛋白命名规则,分别对应命名为Rab11、Rab2、Rab18、Rab1、Rab8、Rab5、Rab7和Rab6八大家族),但是只有十七小类,唯一比拟南芥缺少的一小类是拟南芥中RabG的RabG1小类。
进一步研究发现,拟南芥中的RabA、RabB、RabC、RabD、RabE、RabF、RabG和RabH八大家族分别对应雷蒙德氏棉中的Rab11、Rab2、Rab18、Rab1、Rab8、Rab5、Rab7和Rab6八大家族。通过雷蒙德氏棉的Rab蛋白和拟南芥聚类分析,明确十七小类在8大类中的归属,在阿拉伯数字之后加小写英文字母加以区别,所以命名为Rab11a-Rab11f,Rab2a、Rab18a-Rab18b、Rab1a-Rab1b、Rab8a、Rab5a-Rab5b、Rab7b-Rab7c以及Rab6a等十七小类,每一小类的不同成员之间再按照染色体从小到大、从上到下的位置顺序排列,分别在小写英文字母后面按顺序添加阿拉伯数字,完成雷蒙德氏棉的所有87个Rab蛋白基因家族成员的命名。
在本研究中,我们的目的基因与拟南芥的RabA4类中基因同源性最高,与棉花Rab11d类对应,进一步分析发现该基因在这个小类里位于所有13条染色体的第三个位置,最终命名为GhRab11d3。
根据已有序列,设计PCR扩增引物,从陆地棉TM-1中分别扩增该基因的ORF全长和基因组序列。扩增所用引物见表1。
表1:扩增所用引物
(二)棉花小G蛋白基因GhRab11d3的生物信息学分析
BLAST分析(http://www.ncbi.nlm.nih.gov/blast)发现,GhRab11d3编码的氨基酸序列与拟南芥小G蛋白Rab基因(AtRabA4d)具有很高同源性(77%)。在NCBI网站CD Search中(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi),对该基因的保守结构域预测发现,GhRab11d3具有完整的小G蛋白Rab基因的结构域,4个鸟嘌呤核苷酸结合结构域以及一个效应因子结合结构域,以及C末端的两个半胱氨酸残基。
(三)棉花小G蛋白基因GhRab11d3的定量RT-PCR分析
设计GhRab11d3的特异引物:
F:5’-CCAATGATGAACAAGAAGAAT-3’
R:5’-CTATGGCTATGATGATGACG-3’
在不同组织器官中,进行定量RT-PCR检测。结果表明该基因在根,茎,叶和花器官中都有所表达,在纤维发育各个不同时期差异表达,在纤维起始(开花前3天)就开始表达,随着纤维发育表达量逐渐提高,在纤维快速伸长期(开花后5和10天)表达量最高,后续表达量逐渐下降(如图2所示)。
(四)棉花小G蛋白基因GhRab11d3的转基因功能验证
pBI121是一种传统的植物双元表达载体,其启动子为35S启动子。以此为基础构建了35S启动子的正义表达载体,正义片段的插入位置为35S-P(启动子)与Nos-T(终止子)之间。
改造过的pCAMBIA2301是一种传统的植物双元表达载体,其启动子为RDL启动子。以此为基础构建了RDL启动子的正反义双元载体,正反义片段的插入位置为RDL-P(启动子)与Nos-T(终止子)之间。
以本发明基因SEQ NO ID.1序列全长片段构建正反义植物表达载体,构建完成后,通过农杆菌介导法转化棉花进行功能验证,获得35S启动子正义转基因植株以及RDL启动子正反义转基因植株(如图3所示)。以上三个载体转基因植株均已通过PCR鉴定(如图4所示)。表达分析发现在正义载体转基因植株中,该基因的表达被显著提高,纤维长度变长,而反义载体转基因植株中,该基因的表达被显著抑制,纤维长度变短(如图5所示)。因此使该基因与棉纤维发育相关的功能得到直接验证。
表2:转基因目标基因在基因组和表达水平上检测所用引物
(五)棉花小G蛋白基因GhRab11d3的互作蛋白挖掘
构建好诱饵载体GhRab11d3-pGBKT7后,我们从棉纤维酵母表达文库中筛选到一个与细胞骨架相关的Actin序列。在棉花基因组序列中,我们发现有5个Actin基因在纤维发育期优势表达。而在这五个Actin基因中,有两个翻译成氨基酸序列以后完全一致,所以,我们只选择其中四个Actin基因作为后续实验验证的基因,分别命名为ACTa、ACTb、ACT2和ACT4。为了验证GhACT的全长基因是否和诱饵GhRab11d3-pGBKT7存在相互间作用,我们将GhACTa、GhACTb、GhACT2和GhACT4基因分别克隆到pGADT7载体上,最后成功地构建成载体GhACTa-pGADT7、GhACTb-pGADT7、GhACT2-pGADT7和GhACT4-pGADT7。
进一步将载体GhRab11d3-pGBKT7分别和GhACTa-pGADT7、GhACTb-pGADT7、GhACT2-pGADT7、GhACT4-pGADT7共转化到菌株AH109中,第一次筛选在添加有20mM的3-AT的三缺培养基SD/-His/-Leu/-Trp上进行,同时,在二缺培养基SD/-His/-Leu上也做同样的转化实验,以此检查转化效率。将第一次筛选到的阳性克隆在涂有X-α-Gal上的四缺培养基SD/-Ade/-His/-Leu/-Trp上划线,并且进一步确认二者是否为真正的阳性克隆。阳性和阴性对照则首先在二缺培养基SD/-His/-Leu上转化生长,之后,同时在添加有20mM的3-AT的三缺培养基和涂有X-α-Gal的四缺培养基上点种繁殖。
通过酵母双杂实验证明,Rab11d3蛋白与四个在纤维伸长期优势表达的Actin蛋白存在互作(如图6所示),表明Rab11d3蛋白功能与纤维骨架蛋白相关。
(六)棉花小G蛋白基因GhRab11d3转基因株系及野生型开花后10天纤维中的细胞骨架肌动蛋白F-actin微丝的染色
在大田中取开花后10天棉花纤维,立即用4%多聚甲醛进行固定10-20min,用PBS(PH=7.0)缓冲液冲洗2-3次,每次10分钟,将棉花纤维细胞置于含1U of Alexaphalloidin的PBS(PH=7.0)缓冲液中在室温染色60-90分钟,用PBS(PH=7.0)缓冲液冲洗2-3次将纤维细胞表面残留的Alexaphalloidin洗净。将洗净的棉花胚珠及纤维置于激光共聚焦(LSM710,Zeiss)下观察。对绿色荧光信号分析发现:过表达株系纤维中肌动蛋白F-actin微丝的含量显著增加,而抑制表达株系纤维中的肌动蛋白F-actin微丝的含量显著减少(如图7所示)。同时,细胞骨架蛋白微丝的形态结构也发生了变化,抑制表达株系其纤维中的肌动蛋白F-actin微丝结构受损断裂。
(七)棉花小G蛋白基因GhRab11d3转基因株系及野生型开花当天胚珠及开花后10天纤维中囊泡荧光染色
在大田取开花当天胚珠及开花后10天棉花纤维细胞,立刻置于7.5mg/L FM4-64荧光染料中进行染色,室温染色2小时,用DMSO冲洗2-3遍,每次5分钟,将胚珠及纤维表面残留FM4-64染液洗净。将洗净的棉花胚珠及纤维置于激光共聚焦(LSM710,Zeiss)下观察。对红色荧光信号分析发现,与野生型相比,在GhRab11d3过表达材料中无论是开花当天胚珠表面突起中还是开花后10天纤维顶端均聚集有更多的囊泡,相反,在GhRab11d3抑制表达株系中囊泡数量明显减少(如图8所示)。囊泡运输能力的强弱,最终导致囊泡运输多糖类物质到细胞壁上的果胶含量产生变化。选取转基因株系及对照W0在5DPA以及10DPA的纤维样品,使用苏州科铭生物技术有限公司的果胶含量试剂提取试剂盒(微量法,100管/48样),具体方法参考其果胶含量试剂盒说明书。通过测量,我们发现:在5DPA,转35S-SC的303系其果胶含量比W0显著提高,转RDL-SC的210系其果胶含量比W0极显著增加,而转RDL-ASC的98系的果胶含量则比W0极显著降低。在10DPA,转35S-SC的303系和转RDL-SC的210系其果胶含量与W0相比,都是极显著增加,而转RDL-ASC的98系其果胶含量与W0相比,则是极显著降低(如图9所示)。
<110> 南京农业大学
<120> 能显著提高棉花纤维长度的小G蛋白Rab基因
<160> 3
<210> 1
<211> 690
<212> DNA
<213> 陆地棉(Gossypium hirsutum)
<220>
<221> 棉花小G蛋白Rab基因ORF序列
<400> 1
atgtctagtt tgcaaagggg tttcgatcaa aagatagatt acgtgttcaa agtggtgcta 60
ataggagact ctgctgtggg gaaatcgcag ctcttatccc gcttcacaag aaacgaattc 120
aatatcgatt ccaaggccac catcggcgtc gaattccaaa ctaaaacttt gcttatcgat 180
cataaatccg tcaaggctca gatttgggat actgctggtc aagaaaggta ccgggcggta 240
acgagtgcat actacagagg agcagtgggg gcaatgctgg tgtatgacat aactaagcgt 300
cagtcatttg atcatgttgc caagtggtta gaggaattgc gtggacatgc tgataaaaac 360
attgtgatta tgcttgtggg caacaagtct gaccttgcct cccttagagc tgttcccata 420
gaggatgcta aggaattcgc ccaaagggaa agcctcttct tcatggagac ttcagccctt 480
gaggccacta atgtcgaatc ggctttcctt actgttgtaa cagagattta ccgagtcatc 540
agcaagaaaa acctggtagc caatgatgaa caagaagaat ccgggggtaa tgcatcactt 600
ctcaagggaa ctaccattgt tgtccctggt ccccaaccac aatctggttc tggaggaaag 660
agtttcagtt gctgtgcgtc atcatcatag 690
<210> 2
<211> 958
<212> DNA
<213> 陆地棉(Gossypium hirsutum)
<220>
<221> 棉花小G蛋白Rab基因基因组序列
<400> 2
atgtctagtt tgcaaagggg tttcgatcaa aagatagatt acgtgttcaa agtggtgcta 60
ataggagact ctgctgtggg gaaatcgcag ctcttatccc gcttcacaag aaacgaattc 120
aatatcgatt ccaaggccac catcggcgtc gaattccaaa ctaaaacttt gcttatcgat 180
cataaatccg tcaaggctca gatttgggat actgctggtc aagaaaggta tataaacttt 240
atacatacaa acccttttcc tattactcat tttactgcta tttgtttatg ggaattggga 300
cgatgccctt tatcaaccat taaacatatc tatgttgcta aaaagcttga gactttggat 360
tagaggattg gtttctcttt gatttctgac tgtatttggt cagtcataat tgagcttgat 420
aataggtgaa actatgtata tatatatatg tacggtatgt atgatatgtt gataatggtg 480
gttgatggaa cacaggtacc gggcggtaac gagtgcatac tacagaggag cagtgggggc 540
aatgctggtg tatgacataa ctaagcgtca gtcatttgat catgttgcca agtggttaga 600
ggaattgcgt ggacatgctg ataaaaacat tgtgattatg cttgtgggca acaagtctga 660
ccttgcctcc cttagagctg ttcccataga ggatgctaag gaattcgccc aaagggaaag 720
cctcttcttc atggagactt cagcccttga ggccactaat gtcgaatcgg ctttccttac 780
tgttgtaaca gagatttacc gagtcatcag caagaaaaac ctggtagcca atgatgaaca 840
agaagaatcc gggggtaatg catcacttct caagggaact accattgttg tccctggtcc 900
ccaaccacaa tctggttctg gaggaaagag tttcagttgc tgtgcgtcat catcatag 958
<210> 3
<211> 229
<212> PRT
<213> 陆地棉(Gossypium hirsutum)
<220>
<221> 棉花小G蛋白氨基酸序列
<400> 3
Met Ser Ser Leu Gln Arg Gly Phe Asp Gln Lys Ile Asp Tyr Val Phe
1 5 10 15
Lys Val Val Leu Ile Gly Asp Ser Ala Val Gly Lys Ser Gln Leu Leu
20 25 30
Ser Arg Phe Thr Arg Asn Glu Phe Asn Ile Asp Ser Lys Ala Thr Ile
35 40 45
Gly Val Glu Phe Gln Thr Lys Thr Leu Leu Ile Asp His Lys Ser Val
50 55 60
Lys Ala Gln Ile Trp Asp Thr Ala Gly Gln Glu Arg Tyr Arg Ala Val
65 70 75 80
Thr Ser Ala Tyr Tyr Arg Gly Ala Val Gly Ala Met Leu Val Tyr Asp
85 90 95
Ile Thr Lys Arg Gln Ser Phe Asp His Val Ala Lys Trp Leu Glu Glu
100 105 110
Leu Arg Gly His Ala Asp Lys Asn Ile Val Ile Met Leu Val Gly Asn
115 120 125
Lys Ser Asp Leu Ala Ser Leu Arg Ala Val Pro Ile Glu Asp Ala Lys
130 135 140
Glu Phe Ala Gln Arg Glu Ser Leu Phe Phe Met Glu Thr Ser Ala Leu
145 150 155 160
Glu Ala Thr Asn Val Glu Ser Ala Phe Leu Thr Val Val Thr Glu Ile
165 170 175
Tyr Arg Val Ile Ser Lys Lys Asn Leu Val Ala Asn Asp Glu Gln Glu
180 185 190
Glu Ser Gly Gly Asn Ala Ser Leu Leu Lys Gly Thr Thr Ile Val Val
195 200 205
Pro Gly Pro Gln Pro Gln Ser Gly Ser Gly Gly Lys Ser Phe Ser Cys
210 215 220
Cys Ala Ser Ser Ser
225
- 还没有人留言评论。精彩留言会获得点赞!