一种百合萜类花香基因LoTPS3及其应用的制作方法
本发明涉及基因工程技术领域,更具体地,涉及一种百合萜类花香基因lotps3及其应用。
背景技术:
花香在植物繁殖过程中发挥了重要的作用,不仅可以提高作物的产量和质量,还可以增加观赏植物和切花的审美特性。大多数植物的花香是由一系列低分子量的挥发物混合在一起形成,其化学成分主要是萜烯类化合物、苯丙酸类化合物/苯环型化合物和脂肪酸衍生物(picherskyetal.,2006;dudarevaetal.,2004),而萜烯类化合物种类繁多,是植物次生代谢中种类最多的一类(picherskyanddudareva,2007)。萜类化合物是一类由数个异戊二烯(isoprene,c5)结构单位构成的化合物的统称,根据其数目不同可分为单萜(monoterpene,c10)、倍半萜(sesquiterpene,c15)和二萜(diterpene,c20)等。萜类不仅是组成花香的主要成分,还有其它众多的生理生态作用。如,吸引授粉者;调节植物的生长和发育;调节植物耐热性;抵御光氧化胁迫;直接和间接的植物防御。除此之外,萜类还广泛应用于香料、化妆品、食物、药物和杀虫剂等行业,有非常大的商业价值(tholl,2006;yuandutsumi,2009)。
萜类化合物如沉香醇,罗勒烯等是组成‘西伯利亚’百合花香的主要成分之一,且释放具有昼夜节律性(范燕萍,2008)。萜类合成酶(terpenesynthase,tps)是萜类合成的关键酶,按形成产物的不同,可分为单萜合成酶、倍半萜合成酶和二萜合成酶等。萜类合成酶以异戊二烯焦磷酸(gpp,fpp和ggpp)为底物合成大量结构、功能多样的萜类化合物,由于大多数萜类化合物为环状,因此萜类合成酶也被称为环化酶(liangetal.,2002)。萜类合成酶以金属离子为辅因子,使异戊二烯焦磷酸发生亲电反应,首先,底物通过焦磷酸基团断裂或质子化的离子化过程,形成与酶结合的异戊二烯碳正离子中间体,由于酶活性位点的限制,碳正离子发生电子重排,然后经过一系列的异构化、环化或重排,最后经过去离子化或加水形成环状或非环状的萜类化合物(degenhardtetal.,2009;zulakandbohlmann,2010)。
萜类合成酶蛋白一般是由550~850个氨基酸残基组成,相对分子质量为50~100kda(bohlmannetal.,1998)。研究表明,萜类合成酶在三维结构上有很高的相似性,由α-螺旋、短的连接环和拐角等立体结构组成,并含有两个明显的结构区域,即c-末端结构区域和n-末端结构区域。酶的活性中心是c-末端的一个疏水区域,由6个α-螺旋组成,这个疏水区域利于底物疏水烃基部分进入和结合。n-末端没有特殊的功能元件,突变分析显示,这个区域起到方便c-末端催化活性区域正确折叠的脚手架作用(degenhardtetal.,2009;starksetal.,1997)。几乎所有的萜类合成酶都含有一个天冬氨酸富集基序(ddxxd),这个基序被认为起到结合金属离子的作用(sacchettiniandpoulter,1997),它定位在活性位点的入口处,在引导底物催化时发挥重要的作用,若基序发生突变则会导致酶催化活性的下降或产生异常的产物。根据萜类合成酶氨基酸序列的相关性,将tps家族分为七个亚家族,从tpsa—tpsg,同一亚家族的不同成员之间有最小40%的同源性(bohlmannetal.,1998;dudarevaetal.,2003)。
萜类合成酶是萜类化合物生物合成的关键酶,因此也成了萜类生物合成过程中研究最多和最深入的酶类,自1992年在烟草中克隆了两个倍半萜合成酶基因(facchiniandchappell,1992)以来,科学家已在40多种植物中克隆了200多个单萜和倍半萜合成酶基因(degenhardtetal.,2009),涉及农作物(chenetal.,1995;kollneretal.,2008;yuanetal.,2008)、针叶类植物(bohlmannetal.,1999;martinetal.,2004)、药用植物(deguerryetal.,2006)、香料植物和观赏植物以及拟南芥(chenetal.,2003)等模式植物。
dudareva等(1996)在仙女扇中克隆了一个花器官特异表达并受植物发育调控的单萜合成酶基因-沉香醇合成酶基因,这是第一次克隆到特异表达的花香合成酶基因。在拟南芥基因组中也发现了三个在花中特异表达的萜类合成酶基因,并且这三个酶催化产生了拟南芥花中主要的挥发性物质(chenetal.,2003)。
挥发性萜类物质经常在特定的时期或发育阶段,从特定的植物组织中释放出来,用来吸引授粉昆虫。在金鱼草中(dudarevaetal.,2003),两个主要的香气成分(月桂烯和罗勒烯)在花的上唇瓣和下唇瓣部位释放,并且受发育和昼夜节律的调控,这种昼夜节律可能与相应萜类合成酶基因在一天中的表达量不同有关。在烟草花瓣和柱头中的许多萜类物质在夜间释放,主要由于1,8-桉油醇合成酶基因在转录时受昼夜节律的调控(roederetal.,2007)。在矮牵牛,猕猴桃等花香释放具有昼夜节律的植物中,相关的萜类合成酶基因的表达也具有明显的昼夜节律(nieuwenhuizenetal.,2009;kongetal.,2012)。许多植物萜类花香物质释放都遵循昼夜节律,萜类释放的这种特异改变可能与授粉昆虫的出现时间有关系。例如,由夜间活动昆虫授粉的花卉可能在傍晚时候香气释放量达到最大,使其与夜间活动昆虫的最大传粉时期相一致(yuandutsumi,2009),这都是植物为适应外界环境而逐渐进化来的。
近年来,植物萜类生物合成的基因操作已成为一个热门的研究领域。lucker等(2004b)通过基因工程手段将三个不同的柠檬单萜合成酶转入到萜类挥发量很少的烟草中,结果在转基因烟草的叶和花中,单萜混合物((+)-柠檬烯、γ-萜品烯,β-蒎烯)的含量得到了明显的提高,组分得到了改变,并且人类的嗅觉可以感知到转基因烟草释放的单萜挥发性物质。这为采用基因工程技术培育香花品种提供了一条有效的途径。
技术实现要素:
目前,国内对萜类的研究主要集中在药用、香料提取等方面,而对百合萜类花香基因却鲜有报道。克隆花香基因是对百合花香形成机理研究的前提,揭示百合花香和诱导防御反应的分子机理可以增加花香和增强其抗性。这为采用基因工程的方法培育具浓郁花香植物新品种提供了一条崭新的途径。本发明的目的是分离克隆‘西伯利亚’百合中携带的一个控制萜类花香成分罗勒烯和橙花叔醇的双功能酶基因lotps3。
本发明另一目的在于提供上述lotps3基因编码的蛋白质。
本发明另一目的在于提供含有上述基因的表达载体。
本发明还有一个目的在于提供上述基因在产生分子标记及其在选育对花香品种中的应用,以及上述基因编码的蛋白质在制备香精和药物中的应用。
本发明通过以下技术方案予以实现上述目的:
一种百合萜类花香基因lotps3,所述基因的全长cdna序列如seqidno:1所示,所述基因可赋予百合花朵沉香醇香味。本发明所示百合萜类花香基因lotps3仅在有香味的百合花组织表达,在无香味的百合品种中不表达,并且其表达与花发育进程有关,另外该基因的表达具有明显的昼夜节律性。lotps3基因cdna全长为1693bp,如seqidno:1所示;基因编码区(cds)共1617bp,如seqidno:2所示;推测编码538个氨基酸,其氨基酸序列如seqidno:3所示,推测蛋白分子量为62.7kda。该基因序列包含ddxxd保守序列,对其进行基因系统进化树分析表明,属于植物萜类合成酶基因家族tps-g亚族。
本发明还保护将百合花香基因lotps3的全长cdna序列连接到原核表达载体上,经过体外诱导得到高纯度的体外重组蛋白。通过给予不同反应底物gpp和fpp分别对上述体外重组蛋白进行体外酶活实验。结果显示lotps3蛋白催化gpp生成单萜类物质沉香醇,占反应总产物的100%;催化fpp生成倍半萜类物质橙花叔醇,占反应总产物的74.6%。沉香醇和橙花叔醇均为百合花香组成成分之一。表明本发明的lotps3蛋白不同于其他具有单一催化功能的单萜合成酶和倍半萜合成酶,可以同时催化不同类型的底物,从而生成种类多样的萜烯类化合物,是一个双功能酶基因。
根据本发明提供的lotps3基因序列信息,本领域技术人员可以通过以下方法容易地获得与lotps3等同的基因:(1)通过数据库检索获得;(2)以lotps3基因片段为探针筛选百合或其它植物的基因组文库或cdna文库获得;(3)根据lotps3基因序列信息设计寡核苷酸引物,用pcr扩增的方法从百合或其它植物的基因组、mrna和cdna中获取;(4)在lotps3基因序列的基础上用基因工程方法改造获得;(5)用化学合成的方法获得该基因。
同时,本发明还提供一种用于扩增百合萜类花香基因lotps3的引物对,引物序列如seqidno:4~5所示。
同时还提供一种重组载体,所述载体包含上述百合萜类花香基因lotps3。
一种包含上述重组载体的重组菌。
一种包含上述重组载体的细胞系。
本发明提供的百合花香基因lotps3具有重要的应用价值。应用之一是将所述的lotps3基因序列连接到任何一种植物转化载体,用任何一种转化方法将lotps3花香基因导入百合或其他植物细胞,可获得表达所述基因的转基因香花植株,从而应用于生产。本发明所述的基因构建到植物转化载体中时,可以对所述基因或其调控序列适当修饰,也可以在其转录起始密码子前用其它启动子取代所述基因原有的启动子,从而拓宽和增强植物产生花香和增强抗性的能力。
本发明提供的花香基因lotps3的另一个应用是根据所述基因序列信息产生特异性的分子标记,包括但不限于snp(单核苷酸多态)、ssr(简单序列重复多态)、rflp(限制性内切酶长度多态)、cap(切割扩增片段多态)。用这些标记可鉴定百合或其它植物的花香基因型,用于分子标记辅助选择育种,从而提高育种的选择效率。
与现有技术相比,本发明具有以下有益效果:
本发明提供了一种新的花香萜类物质合成酶基因lotps3,所述基因不仅可以催化gpp形成单萜类化合物沉香醇,同时可以催化fpp形成倍半萜类化合物橙花叔醇,在提高植物观赏特性和抗性方面都发挥着重要作用。将本发明的基因片段构建在植物表达载体上,可以外源转化其他植物材料,从而获得含有花香合成酶基因的转基因材料,对培育香花植物提供了一种有效地方法。花香基因的克隆是克服传统育种中不能在植物种间转移花香基因的问题的前提。另外,本发明能够进一步提供或应用利用上述dna片段获得的花香的转基因植株和相应的种子,以及用本发明的基因或基于该基因的重组体转化的植株或由这类植株获得的种子。可以用有性杂交的方式将本发明的基因转入其他的植株。
附图说明
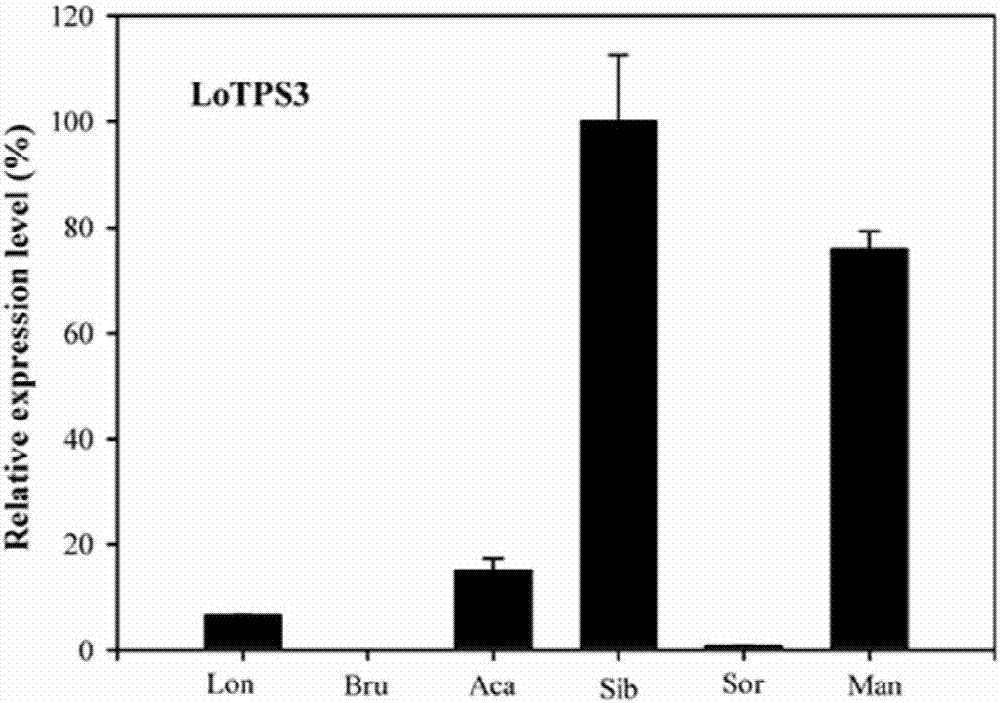
图1为本发明lotps3基因在不同百合品种中的表达情况;lon:铁炮;bru:布鲁诺;aca:元帅;sib:西伯利亚;sor:索邦;man:黄天霸。
图2为本发明lotps3基因在百合不同组织中表达特异性情况。
图3为本发明lotps3基因在百合花器官不同发育时期的表达情况。
图4为本发明lotps3基因在百合花器官中的昼夜节律表达规律。
图5为本发明lotps3重组蛋白以gpp为底物体外酶催化反应。
图6为本发明lotps3重组蛋白以fpp为底物体外酶催化反应。
具体实施方式
下面结合附图和具体实施例进一步详细说明本发明。除非特别说明,实施例中采用的试剂和方法均为本领域常规使用的试剂和方法。
实施例1lotps3基因cdna全长的获得
s1.百合花瓣rna的提取:称取0.2g百合盛开期花瓣,加液氮迅速研磨成粉末状,快速转入4°c存放的加有0.5ml2%ctab(含0.1%的巯基乙醇)提取液的2ml离心管中,振荡至彻底混匀;室温放置5min,平放离心管,使表面积最大;4°c12000rpm离心1min,上清转入新的无rnase离心管。加入0.1ml5mnacl,温和混匀;加入0.3ml氯仿,上下颠倒混匀;4°c12000rpm离心10min,取上层水相转入新的无rnase离心管。加入与所得水相等体积的预冷的异丙醇,颠倒混匀,室温放置10min,平放离心管,使表面积最大;4°c12000rpm离心10min,弃掉上清,注意不要倒出沉淀,加1ml75%乙醇(depc水配制)洗涤rna沉淀,颠倒混匀;4°c5000rpm离心3min,倒出液体,注意不要倒出沉淀。剩余的少量液体短暂离心,然后用枪头吸出,室温晾干2~3min;加入30µldepch2o,反复吹打,混匀,充分溶解rna。放于-80°c保存备用。
s2.以百合盛开期的花瓣总rna作为模板,用生工m-mulvfirstcdnasynthesiskit合成第一链cdna。据百合转录组数据库相关注释基因序列,设计引物,上游引物f1:5’-tgactcctctttccttctgcatgc-3’(如seqidno:4所示)。下游引物r1:5’-ttgtaatctaccatcgtaatgtc-3’(如seqidno:5所示)。并交由上海生物工程公司合成。以上述合成的cdna为模板采用takarapcramplificationkit进行pcr扩增反应,具体方法按照说明书进行。pcr程序为:94°c预变性4min;94°c变性30s、56°c复性30s、72°c延伸2min,35个循环;然后72°c延伸10min。在-20°c下保存备用。pcr反应结束后,用1.0%琼脂糖凝胶电泳初步检测pcr产物中是否含有目的片段条带。pcr扩增产物经1%琼脂糖凝胶电泳检测后,用手术刀在紫外灯下切出含有目的片段的胶块,用dna凝胶回收试剂盒(agarosegeldnapurificationkit,takara)进行回收,回收方法基本参照试剂盒说明书。之后要对回收产物进行1%琼脂糖电泳检测,看其回收效果及大致浓度,以便保证后续试验的进行。根据回收的目的片段大小及其有效浓度,取适量回收纯化的产物与克隆载体连接,载体选用takarapmd19-t载体,目的dna与克隆载体的摩尔比控制在3:1左右,具体操作按说明书进行。在16°c下恒温连接3~6h,连接时间的长短视目的片段的长度而定。提前将感受态细胞dh5α(takara)从-80℃冰箱取出,并置于冰盒中待其自然融化,将10µl连接液全量加入感受态细胞的离心管中,冰浴30min后,42°c水浴热激50s,迅速冰上放置2~5min,然后加入37°c预温的soc液体培养基890µl补足到1ml,混匀后37°c下180rpm振荡培养1h。在含100µg/ml氨苄青霉素的lb固体培养基平板表面上涂30µl的x-gal(20mg/ml)和30µliptg(20mg/ml),然后涂布适量转化液,待转化液完全被吸收后倒置于37°c恒温箱中过夜培养,约16h后观察结果,通过x-gal/iptg蓝白斑筛选白色菌落,并初步鉴定重组质粒,平板置于4°c保存。通过蓝白斑初步筛选后,通常选6个菌斑摇菌后提取质粒,用于进一步鉴定。用灭菌的牙签从lb平板培养基上挑取白色单菌落接种于含有100µg/ml氨苄青霉素的lb液体培养基中,于控温震荡水浴摇床上37°c、240rpm振荡过夜培养,用质粒dna小抽试剂盒(上海博亚生物有限公司)提取质粒,步骤按说明书上方法进行。对所提质粒进行1.0%琼脂糖凝胶电泳检测,比较质粒大小并将明显滞后的质粒进行双酶切(ecori/hindiii,takara)分析。在37°c下酶切1h后,对酶切产物进行1.0%琼脂糖凝胶电泳检测。随意选送酶切鉴定后含目的片段的重组质粒进行dna序列测定。测序工作交由上海生工生物技术有限公司完成,采用美国abi377序列分析仪。所得的序列和转录组原有序列信息进行比对,在ncbi进行比对和同源性分析,确定获得的基因序列为tps家族的完整全长序列,同时根据cdna序列推测出其蛋白序列。
结果所示lotps3基因的全长cdna序列如seqidno:1所示,cdna全长为1693bp;基因编码区(cds)如seqidno:2所示,共1617bp;推测其氨基酸序列如seqidno:3所示。
实施例2lotps3基因的表达分析
1、选取不同品种百合花瓣,西伯利亚百合不同发育时期花瓣及西伯利亚百合不同组织部位提取rna,其rna提取均使用trizol法(takara),荧光定量pcr采用sybrgreen(taraka)法,染料法的具体原理见说明书。利用primerpremier5.0软件设计实时荧光定量pcr引物,按荧光定量pcr引物设计原则,用primerpremier5.0分别设计引物,通过荧光定量pcr检测其是否有错配或引物二聚体及其扩增效率,从中选择一对最佳引物,p1:5’-atctagtggatgcatctttgaatagtg-3’(如seqidno:6所示)。p2:5’-ctaccatcgtaatgtccaatggc-3’(如seqidno:7所示)。内参基因gapdh根据real-timepcr引物的设计原则,用primerpremier5.0设计引物,gapdh-p1:5’-ggtattgtcgagggtttgatg-3’(如seqidno:8所示),gapdh-p2:5’-gctgttggcaaagttctccct-3’(如seqidno:9所示)。通过real-timepcr进行检测,并制作标准曲线,以检测其扩增效率(e)是否在90~110%范围内进行筛选。以各样品的cdna为模板,在abi荧光定量pcr仪上进行荧光定量pcr反应。每个样品设3个重复,以ddh2o为阴性对照。反应体系为sybrpremixextaq(takara)10.0µl,上游引物(10µm)0.4µl,下游引物(10µm)0.4µl,cdna2.0µl,ddh2o7.2µl。反应程序为94°c,30s;94°c,15s;55°c,30s;72°c,30min;40个循环,94°c,15s;72°c,30s;0.4°c/s融解曲线分析。反应结束后确认扩增曲线和融解曲线,采用2-△△ct法(livaketal.,2001)进行数据分析,计算百合lotps3在不同样品中的表达情况。
2、结果
基因表达分析结果见图1,2,3所示。从图1中可以看出:lotps3仅在有香味的百合品种花瓣中表达,在没有香味或淡香的百合释放的品种中不表达或少量表达;图2表明:lotps3仅在百合花器官中表达,在根、茎和叶等营养器官不表达,具有花器官特异表达性。图3表明:该基因的表达量受花发育的调控,随着花朵的发育表达量逐渐升高,在盛开期达到最高,在衰败期表达量下降,与百合花香释放量的变化趋势完全一致,说明lotps3是参与调控百合花香萜类物质合成的关键基因。
实施例3lotps3基因的昼夜节律表达分析
以早上8:00完全盛开的百合作为第一个时间点,每间隔8h取一次样,连续取样4天,提取不同时间点的rna。提取使用trizol法(takara),荧光定量pcr采用sybrgreen(taraka)法,染料法的具体原理见说明书。参考实施例2的方法,使用同样的荧光引物,进行基因表达分析;结果如图4所示:lotps3具有连续的昼夜节律性表达,在早上8:00表达量最高,24:00表达量最低。
实施例4lotps3基因原核表达
s1.载体构建:根据所得到lotps3基因的编码区,用包含kpni和notⅰ酶切位点的特异引物f:5’-ggtaccatgcctacgttcaagctctc-3’(如seqidno:10所示);r:5’-gcggccgcacaatgtctgtaagagtaataat-3’(如seqidno:11)所示。进行pcr扩增。pcr产物用takara回收试剂盒回收,回收产物直接用kpni和noti限制性内切酶进行双酶切,1%琼脂糖凝胶回收目的片段。pet-28a原核表达载体用kpni和noti限制酶进行双酶切1%琼脂糖凝胶回收大片段。于16℃连接过夜,将连接产物转化大肠杆菌(e.coli)dh5α感受态细胞;提取质粒经酶切和测序鉴定后,获得重组原核表达载体。
s2.重组蛋白表达:用经鉴定的重组质粒dna转化rosetta(de3)感受态细胞,挑取单菌落接种于5mllb(含25mg/lkan,34mg/lchl)培养基中,37°c震荡培养过夜。转接100µl种子液于新鲜的100ml(含25mg/lkan,34mg/lchl)的lb培养基中,37°c180rpm培养4~6h,测od值至0.4~0.6后用一定量的iptg(0.1~0.2mm)在14~18°c诱导14~16h。同时取另一对照组其中未加iptg诱导。离心收集菌体,用5ml裂解buffer(50mm磷酸bufferph8.0)悬浮细胞,将菌体冷却,置于冰上超声波破碎细胞。12000rpm,4°c离心10min,将上清移至新的离心管中,用双蒸水清洗沉淀一次,再用5ml裂解buffer悬浮沉淀。分别取上清和沉淀各50µl,保存与-20°c,进行sds-page电泳分析。配制12.5%sds聚丙烯酰胺凝胶,按照顺序上样。分别用60v和120v的电压对浓缩胶和分离胶的进行电泳。电泳完毕,考马斯亮蓝染色30min,再用脱色液脱色1~8h,观察并记录试验结果。
s3.重组蛋白的纯化:挑取单菌落接种于5ml的液体lb培养基中,37°c,180rpm,培养过夜,后全部转接至500ml的新鲜液体lb培养集中,37°c,180rpm,培养4h,用iptg(0.1~0.2mm),18°c条件下诱导16h后,收集菌体。取200µl菌体于离心管中,4°c保存,进行sds-page电泳分析。用5ml的裂解buffer悬浮细胞并转移至离心管内,置于冰上使菌体始终保持冷却,超声波破碎细胞。10,000g,4°c离心20~30min,收集上清液。取20µl的上清于-20°c保存,进行sds-page分析。向5ml细胞裂解液中加入1~2ml的ni-ntaresin(镍--次氮基三乙酸树脂填料)混匀,并低速在4°c摇床上结合60min。将结合完全的细胞裂解液和树脂混合物装入层析柱中,移走底部盖帽收集流出液部分(flow-throughfraction,f),取20µl的流出液,于-20°c保存,进行sds-page分析。用4ml的洗涤buffer洗脱层析柱两次,收集每部分的洗脱液(washfraction,w),各取20µl保存于-20°c冰箱中作sds-page分析。用0.5ml的洗脱buffer洗柱四次,分别用不同的收集管依次收集每部分的洗脱液(elutionfraction,e),分别标记为e1,e2,e3,e4,各取20µl进行sds-page分析。根据sds-page的检测结果,集中含有目标蛋白洗脱部分于一个离心管中,用移液器转移至透析袋中,4°c透析过夜。收集透析后的溶液,加入甘油使蛋白溶液中甘油总含量为20%,并且以200µl/管的量进行分装,取微量进行sds-page蛋白浓度检测,其余的放于-70°c超低温冰箱进行保存备用。
s4.酶催化功能鉴定:将30mmhepes,100ulph7.5,5mmdtt,20mmmgcl2各100ul,以及蛋白提取液100ul,2ulgpp或者fpp,加ddh2o588µl,使终体积为1ml,加入样品瓶中密封,30°c水浴1h,将75µm聚二甲基氧烷(pmds)萃取纤维头插入玻璃瓶中,顶空固相微萃取1h,反应结束后将萃取纤维头放至气相色谱-质谱连用仪中进行分析,气相色谱条件为:色谱柱为hp-1nnowax柱(30m×0.25mm);载气为高纯氦气,分流比20:1,柱前压50pa,流量1ml/min;取样时间2min;程序升温:柱起始温度45°c,保持2min,以5°c/min的速度升至80°c保持1min,然后以10°c/min的速度升至250°c保持5min。质谱条件为:gc-ms的接口温度220°c,电子轰击源ei、350v;离子源温度170°c;电子能量70ev;扫描质量范围35~335aum,采集到的质谱图用willey/mainlib库进行分析。
lotps3基因原核表达结果见图5,6。图5所示:以gpp为底物时,pet-28a-lotps3体外酶催化反应产物经质谱鉴定为单萜类物质沉香醇。图6所示,以fpp为反应底物时生成产物经质谱鉴定为倍半萜类物质橙花叔醇,说明lotps3基因编码生成的酶是一种同时可以催化gpp和fpp的双功能酶基因。
序列表
<110>华南农业大学
<120>一种百合萜类花香基因lotps3及其应用
<130>1713510zbsh042
<141>2017-10-17
<160>11
<170>siposequencelisting1.0
<210>1
<211>1693
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>1
tgactcctctttccttctgcatgcctacgttcaagctctcggttcccttcatgccagtgg60
ccaggatattcacaactaaaacccaacgatgctccaaagccagtagttcggaggagtttt120
ctgacattaaacatttaaataacctagcaaaagtcagaagtatactctcaaaagaaagtg180
ataaaatgcagttgttaaagataattgatttacttcaaagaatggctattgattaccact240
tcaaggaagatattgaagatattctgaaatctcattatgagaacataggagacatgaagg300
gtgatgaaactaataatttattcaatatctcaattcaatttcgattattgagacaaggaa360
ggtacaatatatcacatgatgtttttaaaagattcattgataaggaaggtagatttgacc420
tatccttgagaaaagatacacggggattgcttagtttatttgatgcttcacaactaactg480
gaggcgaggaacttcttcataatgcatgtgaattttctcggaaacatcttatgtcttctc540
tgagttctttggatcctgattcacaaagatttattattcatacaatagagtatccttgtt600
tcacgactcttcaaagatataaagctcgacactatctcggtcattgtggaacaaattatg660
gattggaaggatttctacagcaagttgcaagatttgacttcaatcaagttcaatatttat720
accaaaaggaactaaaagaaattttcagttggtggaaaaacttgggcttgttacaagaat780
taagtttaattcgagatgaccctatgaaatggtatctattggttatgtgcattctccctg840
atccactattatcgaggcaaaggactgagcttgtcaaagtggtagcattaatttatatta900
ttgatgatatatttgacatctatgggtcacctgatgaactatgccttctaacagaggcta960
ttcagaaatgggatgtttcagcatcagataaacttccaaagtacatgagagtttgcttta1020
caaccttgtacaacattaccactgaaatttcggatattgtagtaaatgaaagtggttggg1080
atcctattaacagtctacaaaaaatgtggatgaaactgttcaatgcgttcttaacggaat1140
caaaatggtttgcatcaaaatatgttccgcccactgaagagtacttgaaaaatggaatta1200
ttagctccggtgtacatattctgtttgtacacatattttttctactgggacaagacatta1260
caaaggagactgtgaaaaaaatagagagtattcctactctgatttccttaccagcaacac1320
ttcttcgtctatgggatgatctagaaattgccaatgataagaagcaagaggaatttgacg1380
gttcctatttagactgctacattaaagaacatccaggatgctctactgaagtcgcaaggg1440
aaaaagtgatgaatctgatcttctgtacatcacaatacctcaacaaggagtgtttctcag1500
acaattctttctctccgtatctagtggatgcatctttgaatagtgcaaggatttttaaag1560
tgatgcacagttatgaccaagacaatcttccaattttggaggactatacaaaattattac1620
tcttacagacattgtaataatttgttaaaaccattgtctgatagccattggacattacga1680
tggtagattacaa1693
<210>2
<211>1617
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>2
atgcctacgttcaagctctcggttcccttcatgccagtggccaggatattcacaactaaa60
acccaacgatgctccaaagccagtagttcggaggagttttctgacattaaacatttaaat120
aacctagcaaaagtcagaagtatactctcaaaagaaagtgataaaatgcagttgttaaag180
ataattgatttacttcaaagaatggctattgattaccacttcaaggaagatattgaagat240
attctgaaatctcattatgagaacataggagacatgaagggtgatgaaactaataattta300
ttcaatatctcaattcaatttcgattattgagacaaggaaggtacaatatatcacatgat360
gtttttaaaagattcattgataaggaaggtagatttgacctatccttgagaaaagataca420
cggggattgcttagtttatttgatgcttcacaactaactggaggcgaggaacttcttcat480
aatgcatgtgaattttctcggaaacatcttatgtcttctctgagttctttggatcctgat540
tcacaaagatttattattcatacaatagagtatccttgtttcacgactcttcaaagatat600
aaagctcgacactatctcggtcattgtggaacaaattatggattggaaggatttctacag660
caagttgcaagatttgacttcaatcaagttcaatatttataccaaaaggaactaaaagaa720
attttcagttggtggaaaaacttgggcttgttacaagaattaagtttaattcgagatgac780
cctatgaaatggtatctattggttatgtgcattctccctgatccactattatcgaggcaa840
aggactgagcttgtcaaagtggtagcattaatttatattattgatgatatatttgacatc900
tatgggtcacctgatgaactatgccttctaacagaggctattcagaaatgggatgtttca960
gcatcagataaacttccaaagtacatgagagtttgctttacaaccttgtacaacattacc1020
actgaaatttcggatattgtagtaaatgaaagtggttgggatcctattaacagtctacaa1080
aaaatgtggatgaaactgttcaatgcgttcttaacggaatcaaaatggtttgcatcaaaa1140
tatgttccgcccactgaagagtacttgaaaaatggaattattagctccggtgtacatatt1200
ctgtttgtacacatattttttctactgggacaagacattacaaaggagactgtgaaaaaa1260
atagagagtattcctactctgatttccttaccagcaacacttcttcgtctatgggatgat1320
ctagaaattgccaatgataagaagcaagaggaatttgacggttcctatttagactgctac1380
attaaagaacatccaggatgctctactgaagtcgcaagggaaaaagtgatgaatctgatc1440
ttctgtacatcacaatacctcaacaaggagtgtttctcagacaattctttctctccgtat1500
ctagtggatgcatctttgaatagtgcaaggatttttaaagtgatgcacagttatgaccaa1560
gacaatcttccaattttggaggactatacaaaattattactcttacagacattgtaa1617
<210>3
<211>538
<212>prt
<213>百合(liliumbrowniivar.viridulum)
<400>3
metprothrphelysleuservalprophemetprovalalaargile
151015
phethrthrlysthrglnargcysserlysalasersersergluglu
202530
pheseraspilelyshisleuasnasnleualalysvalargserile
354045
leuserlysgluserasplysmetglnleuleulysileileaspleu
505560
leuglnargmetalaileasptyrhisphelysgluaspilegluasp
65707580
ileleulysserhistyrgluasnileglyaspmetlysglyaspglu
859095
thrasnasnleupheasnileserileglnpheargleuleuarggln
100105110
glyargtyrasnileserhisaspvalphelysargpheileasplys
115120125
gluglyargpheaspleuserleuarglysaspthrargglyleuleu
130135140
serleupheaspalaserglnleuthrglyglyglugluleuleuhis
145150155160
asnalacysglupheserarglyshisleumetserserleuserser
165170175
leuaspproaspserglnargpheileilehisthrileglutyrpro
180185190
cysphethrthrleuglnargtyrlysalaarghistyrleuglyhis
195200205
cysglythrasntyrglyleugluglypheleuglnglnvalalaarg
210215220
pheasppheasnglnvalglntyrleutyrglnlysgluleulysglu
225230235240
ilephesertrptrplysasnleuglyleuleuglngluleuserleu
245250255
ileargaspaspprometlystrptyrleuleuvalmetcysileleu
260265270
proaspproleuleuserargglnargthrgluleuvallysvalval
275280285
alaleuiletyrileileaspaspilepheaspiletyrglyserpro
290295300
aspgluleucysleuleuthrglualaileglnlystrpaspvalser
305310315320
alaserasplysleuprolystyrmetargvalcysphethrthrleu
325330335
tyrasnilethrthrgluileseraspilevalvalasnglusergly
340345350
trpaspproileasnserleuglnlysmettrpmetlysleupheasn
355360365
alapheleuthrgluserlystrpphealaserlystyrvalpropro
370375380
thrgluglutyrleulysasnglyileileserserglyvalhisile
385390395400
leuphevalhisilephepheleuleuglyglnaspilethrlysglu
405410415
thrvallyslysilegluserileprothrleuileserleuproala
420425430
thrleuleuargleutrpaspaspleugluilealaasnasplyslys
435440445
glngluglupheaspglysertyrleuaspcystyrilelysgluhis
450455460
proglycysserthrgluvalalaargglulysvalmetasnleuile
465470475480
phecysthrserglntyrleuasnlysglucyspheseraspasnser
485490495
pheserprotyrleuvalaspalaserleuasnseralaargilephe
500505510
lysvalmethissertyraspglnaspasnleuproileleugluasp
515520525
tyrthrlysleuleuleuleuglnthrleu
530535
<210>4
<211>24
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>4
tgactcctctttccttctgcatgc24
<210>5
<211>23
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>5
ttgtaatctaccatcgtaatgtc23
<210>6
<211>27
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>6
atctagtggatgcatctttgaatagtg27
<210>7
<211>23
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>7
ctaccatcgtaatgtccaatggc23
<210>8
<211>21
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>8
ggtattgtcgagggtttgatg21
<210>9
<211>21
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>9
gctgttggcaaagttctccct21
<210>10
<211>26
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>10
ggtaccatgcctacgttcaagctctc26
<210>11
<211>31
<212>dna
<213>百合(liliumbrowniivar.viridulum)
<400>11
gcggccgcacaatgtctgtaagagtaataat31
- 还没有人留言评论。精彩留言会获得点赞!