人先天性淋巴细胞前体:鉴定、表征、应用的制作方法
背景技术:
先天性淋巴细胞(ilc)是在早期免疫应答中发挥重要作用的淋巴效应细胞的新家族,由细胞毒性细胞(nk细胞)和‘辅助样’ilc组成。后者的特征在于表达白细胞介素-7受体(il-7rα/cd127),并基于它们的转录因子(tf)和特征性的与t辅助(th)细胞类似的细胞因子产生而分成三个不同的组。第1组ilc(ilc1)表达t-bet/tbx21,并产生th1相关细胞因子ifn-γ和tnf-α。第2组ilc(ilc2)通过gata-3和rorα依赖性途径分泌th2相关的细胞因子il-5和il-13。第3组ilc(ilc3)利用相关的孤儿受体c(编码rorγt的rorc)来驱动th17相关的细胞因子il-17和/或il-22的产生(serafini等,2015;spits等,2013)。这些不同的ilc亚组存在于不同的淋巴组织和非淋巴组织中,并且在粘膜位点富集,它们在所述粘膜位点的屏障功能和先天免疫防御中发挥重要作用(artis和spits,2015;eberl等,2015)。
首先在次级淋巴组织中鉴定出多个人ilc亚组,随后在几个非淋巴组织位点(肠、肺、肝、皮肤)也有报道(综述于(juelke和romagnani,2016))。
第1组ilc可以产生1型细胞因子(例如,ifnγ和tnf),并且包含自然杀伤(nk)细胞和ilc1(维基百科)。ilc1是与ilc3密切相关的弱细胞毒性细胞(同上)。nk细胞是细胞毒性先天效应细胞(同上)。它们遍及血液、器官和淋巴组织分布,占外周血淋巴细胞的15%左右(同上)。nk细胞在肿瘤监测和病毒感染细胞的快速消除中发挥作用(同上)。
已经描述了两种不同的产生ifn-γ的ilc1群。已经在扁桃体和发炎的肠道中鉴定了表达高水平cd127(称为cd127+ilc1)和cd161,但缺乏其他特异性表面标志物的t-bet+细胞(bernink等,2013)。相反,表达nkp44和cd103但不表达cd127的上皮内ilc1存在于粘膜位点(fuchs等,2013)。这两种ilc1均响应il-12产生ifn-γ,并且可通过最小的eomesodermin(eomes)表达由nk细胞分化。
第2组ilc可产生2型细胞因子(例如il-4、il-5、il-9、il-13)(维基百科)。ilc2(也称为天然辅助细胞、核细胞或先天辅助细胞2)在响应蠕虫感染时发挥分泌2型细胞因子的关键作用(同上)。它们还与过敏性肺部炎症的发展有关(同上)。它们表达趋化因子的特征性表面标志物和受体,其参与淋巴细胞在特定器官位点的分布(同上)。它们需要il-7用于其发育,所述il-7激活两种转录因子(这些细胞都需要)-rorα和gata3。ilc2对于肺中局部th2抗原的初级应答是至关重要的,但对于全身递送的th2抗原的应答是不必要的(同上)。
人gata-3+ilc2表达化学引诱物受体crth2、il-25r和il-33r(
第3组ilc通过它们产生细胞因子il-17a和/或il-22的能力来定义(维基百科)。它们包含ilc3和淋巴组织诱导(lti)细胞(同上)。ilc3是淋巴细胞群,其可以产生il-22并表达nkp46(nk细胞活化受体)(同上)。然而,ilc3与nk细胞不同,因为它们依赖于转录因子rorγt,它们缺乏细胞毒性效应物(穿孔素、颗粒酶和死亡受体)并且它们不产生ifnγ或tnf(同上)。它们主要存在于粘膜组织中,特别是在肠道中(同上)。
淋巴组织诱导(‘lti’)细胞是表达淋巴组织发育所需分子的ilc亚组(同上)。它们对于胚胎发育期间淋巴器官的发育至关重要,并在出生后调节淋巴组织的结构(同上)。它们还与t细胞记忆的维持有关(同上)。
第3组ilc包括胎儿淋巴组织诱导(lti)细胞以及表达转录因子rorγt并产生细胞因子il-17a和/或il-22的成年谱系-cd127+cd117+细胞(综述于(montaldo等,2015)。已经在胎儿肠系膜淋巴结和脾(cupedo等,2009)和成人扁桃体、肠、脾、皮肤、肺、子宫内膜和蜕膜中鉴定了ilc3。ilc3的一个亚组表达天然细胞毒性受体(ncr,包括nkp30、nkp44和nkp46),并在产生il-22的细胞中富集(cella等,2009)。
小鼠成熟ilc通过共同的淋巴祖细胞(clp)从造血干细胞(hsc)分化,在胎儿的肝(fl)和成人的骨髓(bm)中产生不同的id2+tcf-1+plzf+ilc前体(ilcp)(constantinides等,2014;yang等,2015)。在小鼠中,不同的tf和信号转导通路调节这一过程(serafini等,2015);相比之下,人ilc的发育尚未很好地表征(在(juelke和romagnani,2016)中综述)。已经在fl、bm、脐带血(cb)和成人扁桃体中鉴定了产生细胞毒性cd56+nk细胞的nk前体(nkp)(renoux等,2015),而定型ilc3前体(ilc3p)则在扁桃体和肠固有层中发现,但在外周血(pb)、胸腺或bm中未发现,所述ilc3p前体在体外产生生产il22的ncr+ilc3(montaldo等,2014)。最近的一项研究鉴定了表达rorγt的扁桃体人ilcp,其可以发育为成熟的细胞毒性和辅助性ilc(scoville等,2016)。有趣的是,这些人nkp、ilc3p和ilcp是cd34+,并且在次级淋巴组织中富集,但是在循环中很少或不存在。尚不清楚这种ilcp是否与组织中发现的成熟ilc亚组在发育上相关。
先天性淋巴细胞在先天免疫反应的发展中非常重要,并且在保护性免疫以及调节体内平衡和炎症中起重要作用(维基百科)。因此,它们的失调可导致免疫病理学,例如过敏、支气管哮喘和自身免疫疾病(同上)。为了提供ilc的来源,本领域需要开发用于分离ilc的前体细胞的组合物和方法。本发明满足了这种需要。
发明简述
本发明涵盖包含先天性淋巴细胞前体(ilcp)的组合物、该组合物的用途、以及制备和使用这些组合物的方法。
在各种实施方案中,组合物包含纯化的先天性淋巴细胞前体(ilcp)群,其中所述群中至少75%,优选至少90%的细胞具有表型cd127+cd117+cd3-crth2-,并且任选地具有表型cd7+、nkp44-、cd94-和/或lin-,和/或任选的cd26+和/或cd62l+。在一些实施方案中,所述群中至少75%,优选至少90%的细胞具有表型cd127+cd117+lin-crth2-(其中lin-包含cd3-),并且任选地具有表型cd7+、nkp44-和/或cd94-,和/或任选的cd26+和/或cd62l+。优选地,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
在一个实施方案中,本发明涵盖制备纯化的先天性淋巴细胞前体(ilcp)群的方法,包括提供人细胞样品,并在细胞样品中选择具有表型cd127+cd117+cd3-crth2-,任选地具有表型cd7+、nkp44-、cd94-和/或lin-,和/或任选的cd26+和/或cd62l+的细胞以提供细胞群,其中所述群中至少75%的细胞具有表型cd127+cd117+cd3-crth2-,并且任选地具有表型cd7+、nkp44-、cd94-和/或lin,和/或任选的cd26+和/或cd62l+。在一些实施方案中,该方法包括在细胞样品中选择具有表型cd127+cd117+lin-crth2-(其中lin-包含cd3-),并且任选地具有表型cd7+、nkp44-和/或cd94-,和/或任选的cd26+和/或cd62l+的细胞。在该方法的一些实施方案中,所述群中至少75%的细胞具有表型cd127+cd117+lin-crth2-(其中lin-包含cd3-),并且任选地具有表型cd7+、nkp44-和/或cd94-,和/或任选的cd26+和/或cd62l+。优选地,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
在一个实施方案中,本发明涵盖制备选自ilc1、ilc2、ilc3和nk细胞的细胞类型的方法,包括提供先天性淋巴细胞前体(ilcp)群,使细胞群经受外部刺激,并检测选自ilc1、ilc2、ilc3和nk细胞的细胞类型的增加。
优选地,所述群中至少90%的细胞具有表型cd127+cd117+cd3-crth2-,任选地具有表型cd7+、nkp44-、cd94-和/或lin-,和/或任选的cd26+和/或cd62l+。在该方法的一些实施方案中,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-(其中lin-包含cd3-),并且任选地具有表型cd7+、nkp44-和/或cd94-,和/或任选的cd26+和/或cd62l+。优选地,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
在一个实施方案中,方法在体内进行。
在一个实施方案中,本发明涵盖治疗人患者的方法,包括向所述患者施用纯化的ilcp群,其中所述群中至少90%的细胞具有表型cd127+cd117+cd3-crth2-,任选地具有表型cd7+、nkp44-、cd94-和/或lin-,和/或任选的cd26+和/或cd62l+。在该方法的一些实施方案中,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-(其中lin-包含cd3-),并且任选地具有表型cd7+、nkp44-和/或cd94-,和/或任选的cd26+和/或cd62l+。优选地,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
在一个实施方案中,本发明涵盖筛选影响ilc发育的化合物的方法,包括提供先天性淋巴细胞前体(ilcp)群,使细胞群与测试化合物接触,并检测细胞群中的细胞的表型变化。
在一个实施方案中,测试化合物引起ilcp分化减少。在一个实施方案中,测试化合物引起ilcp分化增加。
在一个实施方案中,该方法包括向小鼠输注先天性淋巴细胞前体(ilcp)群,并向小鼠施用测试化合物。
在各种实施方案中,细胞扩增而没有可塑性。
在一个实施方案中,本发明涵盖扩增ilcp的方法,包括在包含il-1β和il-2的培养基中培养根据本发明的纯化的ilcp群。
在各种实施方案中,在包含il-1β和il-2的培养基中培养ilcp。
在一个实施方案中,本发明涵盖以最小可塑性扩增ilc3细胞的方法,其中培养基包含il-1β、il-2和il-7。
附图说明
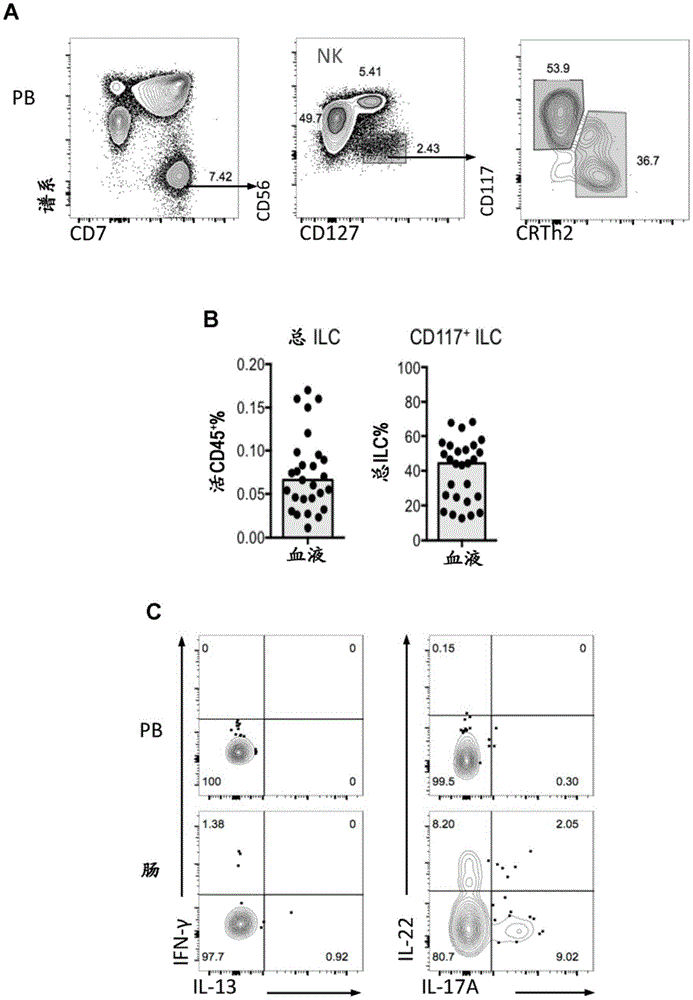
图1a-d描绘了外周血cd117+ilc的表征。(a)用于人pbilc的facs分析的门控策略。在活的cd45+lin-(cd3-cd4-cd5-tcrαβ-tcrγδ-cd14-cd19-)cd7+cd127+细胞(红色)上门控总ilc。通过cd56dim鉴定nk细胞(灰色),通过crth2+细胞标记ilc2(绿色),并且在crth2-cd117+群体上门控cd117+ilc(蓝色)。(b)pb中来自健康成年供体的总ilc的活cd45+和cd117+ilc的总ilc的百分比。结果来自27个健康个体(中值)。(c)pbcd117+ilc和肠cd117+nkp44+/-ilc的表面表型(nkp44、il1r1和cd69)和胞内转录因子(eomes、t-bet、gata-3和rorγt)谱的表达。(d)响应于3小时pma/iono刺激,pbcd117+ilc和肠cd117+nkp44+/-ilc的功能谱(ifn-γ、il-13、il-22和il-17a)。代表性数据来自至少3个独立实验分析的至少3个个体。
图2a-e描绘了cd117+ilc的转录特征和染色质图谱。(a)通过facs从健康供体的外周血中新鲜分离的cd117+ilc和cd34+hsc以进行批量rnaseq和chipseq的示意图和门控策略。(b)描绘标准化chipm-seq信号的热图,其显示36449个高信度增强区的h3k4me2强度,从cd117+ilc特异性鉴定了1989个区。维恩图显示了三类h3kme2+区域(‘cd34+hsc特异性’;‘cd117+ilc特异性’和‘共有’)。如果h3k4me2+富集水平差异>2倍,则认为该区域是细胞类型特异性的。(c)从cd117+ilc(蓝色)和cd34+hsc(橙色)独特鉴定的位于gre附近的基因的分子途径富集的热图。(d)显示与通过rnaseq鉴定的cd34+hsc相比,由cd117+ilc差异性表达的1540个基因的聚集的热图。(e)由来自rnaseq的cd117+ilc高度表达的1540个基因和从‘cd117+ilc特异性’h3kme2+gre鉴定的2283个基因的重叠。
图3a-e描绘了克隆揭示了体外cd117+ilc的多ilc谱系潜力。(a)在含有细胞因子(每种细胞因子20ng/ml)的无基质细胞条件下扩增大批培养的cd117+ilc(10天)。结果来自四个独立供体;ns,p>0.05;**,p<0.01;****,p<0.0001,使用配对studentt检验(中值)。(b)用pma/离子霉素刺激3小时后,facs分析大批培养的cd117+ilc的表面表型、胞内eomes和细胞因子表达,以鉴定nk细胞(eomes+细胞)、ilc1(ifn-γ+细胞)、ilc2(il-13+)和ilc3(il-22+和/或il-17a+)。(c)cd117+ilc-op9基质共培养系统的示意图和形态。(d)将单个pbcd117+ilc进行facs指数分选,并在op9或op9-dl4基质细胞上培养14-18天。在分析表面和细胞因子谱之前,用pma/iono刺激细胞3h。当通过facs检测到至少100个活的人cd45+细胞时,认为克隆是阳性的。当在总的活cd45细胞中检测到超过5%的相应细胞因子时,对ilc亚组的存在进行评分。(e)饼图描绘了检测到的所有可能的ilc组合。总的阳性孔中每个单个或多个ilc分化的频率。数据来自四个独立实验,其中每个实验一个供体。平均而言,op9的克隆效率为40%,op9-dl4基质细胞的克隆效率为26%。
图4a-c描绘了cd117+ilc有效地在体内产生多ilc谱系。(a)体内转移实验的示意图。(b-c)将通过facs从健康个体的pb新鲜分离的1-3×105个cd117+ilc或cd34+hsc肝内转移到新生brgs小鼠。注射后4周分析这些群体的后代。(b)facs分析来自用cd117+ilc或对照cd34+hsc转移的brgs小鼠的骨髓和肠的活的人cd45+细胞门控的淋巴细胞和骨髓表面标志物。(c)facs分析用cd117+ilc转移的brgs小鼠的肺、肠、肝和脾中的不同ilc亚组的表面nkp44和cd117表达、胞内eomes和胞内细胞因子(ifn-γ、il-13、il-22和il-17a)的产生。代表性数据来自3个独立实验,每组至少4只小鼠。
图5a-e描绘了人ilcp在人免疫系统(his)小鼠中累积。(a)制备his小鼠的示意图。将从人胎儿的肝中分离的1.5-2×105个cd117+ilc或cd34+cd38-hsc肝内转移到新生brgs小鼠中。移植后8-9周分析小鼠。(b)his小鼠的脾、bm、肺和肝中人ilcp(lin-cd7+cd127+cd117+)的代表性facs分析。(c)his小鼠的脾、bm、肺和肝中总的人cd45+的ilcp百分比(中值)。(d)来自his小鼠脾脏的ilcp和nk细胞的表面表型和转录因子谱的facs分析。(e)培养前和在具有il-2、-7、-1β和-23的op9-dl4上培养10天后的his小鼠的脾cd117+ilc的细胞因子产生。在pma/iono刺激3小时后分析细胞因子的产生。代表性数据来自至少3次独立实验的8只小鼠。
图6a-n描绘了来自淋巴和非淋巴器官的cd117+ilc的体外批量和克隆测定。将来自不同器官的大批(100-300个细胞)或单个cd117+nkp44+/-cd117+ilc用facs分选到预先接种有op9或op9-dl4且补充有il-2、-7、-1β和-23(各20ng/ml)的96孔圆底板中。在大批培养8-10天(a、d、g、j、m)或单细胞培养14-18天(bc、ef、hi、kl、no)后,对响应pma/iono刺激3小时的细胞因子产生进行胞内facs分析,以鉴定ilc1(ifn-γ+)、ilc2(il-13+)和ilc3(il-22+和/或il-17a+)。代表性facs分析来自从(a)fl、(d)cb、(g)肺、(j)扁桃体分离的大批cd117+nkp44-ilc和从(m)扁桃体分离的cd117+nkp44+的后代。饼图描绘了来自(b-c)fl、(e-f)cb、(h-i)肺、(k-l)扁桃体的cd117+nkp44-ilc和来自(m-n)扁桃体的cd117+nkp44+在op9和op9-dl4上克隆扩增后的所有可能的ilc组合。另见图3图例。数据总结自至少2次独立实验,其中每次实验一个供体。
图7a-c描绘了来自rorc-/-患者的ilcp的发育潜力
(a)来自健康和rorc-/-患者样品的外周血ilc亚组的facs分析。(b)来自健康和rorc-/-患者的pb的总ilc的活cd45+细胞、ilc、ilc2和ilcp的nkdim和nkbr的百分比。结果来自22个健康供体和2名rorc-/-患者。ns,p>0.05;*,p<0.05;**,p<0.01使用配对studentt检验(中值)。(c)将来自健康供体或rorc-/-患者的ilcp进行facs分选,并在含有il-2、-7、-1β和-23的op9-dl4上培养8天。用pma/iono刺激3小时后,分析表面表型、胞内eomes表达和细胞因子产生谱。
图8a-f描绘了ilcp标志物的流式细胞术分析。
使用各种标志物对来自几个正常健康供体的细胞进行facs分析,并测定所示细胞的百分比。
图9a-g描绘了ilcp标志物的流式细胞术分析。
使用各种标志物对来自几个正常健康供体的细胞进行facs分析,并测定所示细胞的百分比。
图10描绘了cd62l和cd26人ilcp亚组的分析。
显示了用于谱系耗尽的人外周血细胞的facs分析的门控策略。在lin-cd7+cd127+cd117+门(人ilcp)内,存在cd45ra的同源表达以及cd62l和cd26的可变表达。
图11描绘了人ilcp克隆扩增。
使用补充有人il-1b、il-2、il-7和il-23的op9基质细胞(表达或不表达dll4)进行单细胞分选后,从指定组织克隆人ilcp(lin-cd7+cd127+cd117+)。然后用pma/离子霉素刺激3小时后,分析克隆的细胞因子产生(ifn-α、il-13、il-17a、il-22)。推定的ilcp被鉴定为细胞因子非生产者。显示了单个ilcp克隆中的绝对细胞数。
图12描绘了人ilcp克隆表型。将衍生自成人外周血的人ilcp克隆的指定细胞表面标志物进行染色。
发明详述
发明人已经表征了外周血cd117+ilc的表型、分子和功能属性。虽然以前提出具有这种表型的细胞代表人ilc3(hazenberg和spits,2014),但是出乎意料地发现这些细胞在多能和单能ilc前体(ilcp)中大量富集,所述多能和单能ilc前体可以在体外和体内产生所有已知的ilc亚组,包括eomes+nk细胞。cd117+ilcp不仅存在于循环中,而且还存在于其中它们保持ilc多能性的组织中。系统分布的ilcp的鉴定提出了一种模型,其中循环ilcp为组织中响应于感染、炎症和细胞转化的ilc分化提供细胞底物。
在该报道中,发明人鉴定并表征人ilc前体(ilcp)作为脐带和成人血液以及胎肝和几种成人组织中的lin-cd7+cd127+cd117+细胞的亚组。人ilcp产生所有成熟的ilc亚组,其在体外培养后或在体内转移至免疫缺陷小鼠后能够产生一系列细胞因子(ifn-α、il-13、il-17a、il-22)。人ilcp还产生eomes+nk细胞,证明它们具有细胞因子产生以及细胞毒性ilc的潜力。这是在任何物种中循环ilcp的第一个证据,并进一步证明ilcp在人淋巴和非淋巴组织(包括粘膜位点)中的广泛系统性分布。
使用标志物cd127+cd117+细胞分选产生约20%的ilcp和60-80%的cd3+t细胞(图8a或9a)。排除谱系细胞(包括cd3)强烈富集了ilcp中的cd127+cd117+细胞(图8e-f;图9e-g)。排除lin-cd127+cd117+细胞中的crth2ilc2提供了进一步的ilcp富集,产生至少75%的ilcp(图8e-f;图9e、9f和9g)。使用标志物cd127+cd117+cd7+crth2-lin-分选产生大约90%的ilcp(图8e和9f)。通过排除cd56+nk细胞(这是表达cd94的唯一lin细胞),可以鉴定出纯的ilcp群体。因此,使用标志物cd127+cd117+crth2-lin-cd94-分选产生大约100%的ilcp(图8f和9g)。在lin+细胞内,cd3+t细胞基本上是表达cd127或cd117的唯一细胞。因此,当比较cd127+cd117+crth2-cd3-与cd127+cd117+crth2-lin-时,获得的ilcp百分比基本上没有差异。
由于基于op9基质细胞的稳定分析可以在单细胞水平评估ilc潜力,因此可以鉴定人ilcp。使用该方法,发明人鉴定了可以产生ifn-α+ilc1、il-13+ilc2或il-17a+和/或il-22+ilc3的单能ilcp以及可以产生两个或更多ilc亚组的多能人ilcp。发明人证明,人cd34+hsc可在体内发育成携带具有多谱系ilc潜能的ilcp的cd117+细胞。总之,本发明人将提出用于人ilcp开发的模型,其中全能(pluripotent)cd34+hsc将逐渐分化成可产生三个主要ilc组(包括eomes+nk细胞)的多能ilcp(具有cd34-cd7+cd127+cd117+cd45ra+表型)。cd34+hsc和多能ilcp均存在于胎肝中,表明该组织允许这种转变。知道cd117+ilcp是否存在于人bm中将是有趣的。先前描述的人扁桃体ilcp(scoville等,2016)可代表该通路中的中间体。bm以及成人和脐带血中缺乏cd34+cd117+cd45ra+ilcp(scoville等,2016)表明这些ilcp在局部产生。本发明人在本文描述的循环和组织驻留的人ilcp还包含具有更受限制的单能ilc的细胞。尽管发明人尚未鉴定能够区分多能和单能ilcp的标志物,但发明人认为它们保持前体-产物关系。
循环人ilcp的转录组学和表观基因组学分析揭示了与ilc谱系的部分规格一致的特征。已知对小鼠ilc发育至关重要(包括tcf7、tox、id2和gata3;(klose等,2014;seehus等,2015;yagi等,2014;yang等,2015))的tf与循环hsc相比,在ilcp中明显上调。相反,早期b和t淋巴细胞生成的特征不明显,这与这些细胞在体外或体内不能适应淋巴细胞一致。ilc组定义的tf(bcl2、tbx21、eomes、rorc)不存在或以低水平表达,表明定向分化(commitment)为ilc1、ilc2或ilc3尚未完成。有趣的是,编码这些因子的基因座仍然“稳定”,这可以通过丰富的h3k4me2修饰来证明。这种染色质图谱可能有助于在细胞因子驱动的扩增后快速产生分化的ilc亚组(zook等,2016),这与初始t细胞中的情况形成对比,在初始t细胞中特征性细胞因子和tf基因座保持无活性,主要具有h3k27甲基化(koues等,2016;shih等,2016)。
虽然在每个测试的人体组织样本中都发现了单能和多能ilcp,但单能和多能ilcp的相对比例明显不同。因此,每个组织可能具有由环境信号调节的独特ilcp“库(repertoire)”。这些可包括调节ilc分化的后期阶段的相同生长因子和细胞因子(在(diefenbach等人,2014)中综述),其将作用于ilcp以诱导特定ilc亚组的发育。或者,细胞因子受体的随机表达可以提供一部分能够进一步分化的ilcp。更好地理解在不同组织环境中调节ilcp反应性的机制对于在人类疾病中的潜在治疗应用至关重要。
本发明人的研究强调了notch信号在调节从单能和多能ilcp分化人ilc中的重要作用。来自组织和血液的ilcp在notch信号(op9-dl4培养系统)存在下显示更大的多能性。这可能表明多能ilcp对notch依赖性存活和增殖信号的依赖性更高(chea等,2016b)。或者,就其体内平衡而言,特定的ilc亚组可能更依赖于notch。特别是,小鼠中的ncr+ilc3亚组是notch依赖性的(chea等,2016a),尽管其作用机制仍不清楚。op9-dl4培养物中产生il-17a和il-22的细胞在批量和克隆水平上的频率增加可能反映了人体系统中的类似需求。
本发明人对人胎肝的分析为妊娠期间多能ilcp和ilc3限制性祖细胞提供了第一个证据。值得注意的是,在该组织中很少检测到其他单能ilcp,表明在胎儿发育的这个阶段,肝脏微环境可以递送使ilcp向ilc3强烈极化的信号。在小鼠中,已经报道了类似的发现(cherrier等,2012)。已经提出notch信号在指导小鼠胎肝中的淋巴细胞命运决定中起作用,促进t-谱系引发的前体的发育,但也改变ilcp的体内平衡(chea等,2016b;dallas等,2005)。由于ilcp表达几种细胞因子受体(il-1r、il-2r、il-18r),使其能够感知组织炎症和压力,因此也可能涉及可溶性因子。
tf表达的调节决定了ilc的命运和功能。已经鉴定了用于ilc亚组的特征性tf,其在表面表型和效应物输出水平上“固定”它们的分化,尤其是对于细胞因子(在(serafini等人,2015)中综述)。tfrorc有助于定义ilc3亚组,并且是小鼠中ilc3(但不是ilc1、ilc2或nk细胞)的发育和维持所必需的(luci等人,2009;sawa等人,2010)。正如所料,rorc由人ilc3和定型ilc3p表达(montaldo等,2014)。最近报道所有人ilc亚组表达rorc(scoville等,2016),表明该tf在人ilc分化中具有更广泛的作用。通过分析来自rorc缺陷患者的血液,发明人可以证明rorc不是人类全身ilc分化所必需的,但对il-17+ilc3亚组的分化至关重要。rorc缺陷患者中的ilcp保留了产生其他ilc和nk细胞亚组的能力。有趣的是,il-22+ilc3以独立于rorc的方式发育,表明这些细胞在人体中具有代偿途径。
已经表明使用op9基质使人ilc2的可塑性最小化(lim等,2016),并且本发明人表明,绝大多数nkp44+ilc3克隆保留其功能属性并且在该培养系统中显示出向ilc1表型很小的可塑性。此外,先前的报道提出,ilc1克隆在il-1b存在下迅速分化为ilc3命运(bernink等,2015),而在发明人的op9培养系统(含有il-1β)中的ilc1克隆保留了它们的ifn-γ特征。因此,发明人的培养系统似乎可用于评估促进来自ilcp的“初级”ilc命运的信号。
最后,发明人对循环和组织驻留的人ilcp的鉴定提出了“按需生产ilc(ilc-poiesisondemand)”的概念,其中ilc分化可以在任何组织和任何年龄发生。最近在小鼠中使用异种共生(parabiosis)的研究提出,ilc是长寿命的组织驻留细胞,在稳态和某些炎症病症下不会再循环(gasteiger等,2015)。相比之下,其他报道表明,几个粘膜ilc亚组的半衰期大约为数周,表明这些细胞必须更新(sawascience)。循环ilcp的发现提供了补充组织ilc以响应稳态损失以及在感染和炎症情况下的机制。本发明涵盖包含ilcp的组合物,以及制备和使用ilcp的方法。
包含ilcp的组合物
本发明涵盖包含如本文所述的先天性淋巴细胞前体(ilcp)的组合物。本文(例如,在实施例中)使用的所有标志物在任何和所有组合中特别预期用作ilcp的标志物,并且可以用于本发明的各种实施方案中。
在一个实施方案中,本发明涵盖纯化的先天性淋巴细胞前体(ilcp)群。优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有表型cd34-cd7+cd127+cd117+cd45ra+。优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞缺少nkp44和/或rorγt表达。优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是il-1r1+和/或cd69-。更优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有表型cd34-cd7+cd127+cd117+cd45ra+,缺少nkp44和/或rorγt表达,并且是il-1r1+和/或cd69-。更优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有表型cd34-cd7+cd127+cd117+cd45ra+,缺少nkp44和/或rorγt表达,并且是il-1r1+和/或cd69-,且任选地还表达cd62l和/或cd26。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd7+cd127+cd117+。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd7+cd127+cd117+crth2-。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117。
更优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+cd3-crth2-。在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞进一步是cd7+、nkp44-、cd94-,和/或lin-,和/或进一步是cd26+,和/或cd62l+。在该方法的一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+,和/或cd62l+。优选地,所述群中至少75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。在一些优选的实施方案中,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。优选地,在肠cd117+细胞产生il-17a或il-22的条件下,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞在刺激后不产生这些细胞因子。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞不表达t-bet、eomes和gata-3hi。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞不表达cd94、cd244和crth2。
优选地,在用药理活化剂刺激后,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞不表达il-13或ifn-γ。
优选地,细胞群包含至少104、105、106、107、108、109或1010个ilcp细胞。
制备ilcp的方法
本发明涵盖用于制备纯化的本发明的先天性淋巴细胞前体(ilcp)群的方法。在一个实施方案中,该方法包括提供人细胞样品,并选择样品中的ilcp。可以用本文(例如实施例)所示的标志物的任何组合选择ilcp。在一个实施方案中,选择细胞的以下标志物的任何组合:lin-、cd34-cd7+cd127+cd117+cd45ra+、nkp44-、rorγt-、il-1r1+、cd69-、cd62l+、cd26+;特别是lin-、cd34-cd7+cd127+cd117+cd45ra+、nkp44-、il-1r1+、cd69-、cd62l+、cd26+。lin-指谱系阴性细胞;lin-包括并且指cd3-、cd4-、cd5-、tcrαβ-、tcrαβ-、cd14-和cd19-。可以通过本领域的常规技术进行选择,例如通过facs分析和细胞分选,例如如实施例中所述。
在一些实施方案中,在肠cd117+细胞产生il-17a或il-22的条件下选择在刺激后不产生这些细胞因子的细胞。
在一些实施方案中,选择不表达t-bet、eomes和gata-3hi的细胞。
在一些实施方案中,选择不表达cd94、cd224和crth2的细胞。
在一些实施方案中,选择在用药理活化剂刺激后不产生il-13或ifn-γ的细胞,例如如实施例中所述。
在一些实施方案中,样品是血液或组织样品。样品可以是成人或胎儿样品。在一些实施方案中,样品是血液、扁桃体、肠、胎肝或肺样品。
在一些实施方案中,例如通过细胞分选选择细胞样品中的lin-cd7+cd127+cd117+crth2-细胞。在一些实施方案中,选择细胞样品中的lin-cd94-crth2-cd127+cd117+细胞。在一些实施方案中,选择细胞样品中的cd127+cd117+cd3-crth2-细胞。在一些实施方案中,选择细胞样品中的cd127+cd117+cd3-crth2-细胞,和进一步的cd7+、nkp44-、cd94-和/或lin-细胞,和/或进一步的cd26+和/或cd62l+细胞。在一些优选的实施方案中,选择细胞样品中的cd127+cd117+lin-crth2-细胞(其中lin-包括cd3+),和任选的进一步的cd7+、nkp44-、cd94-和/或lin-细胞,和/或任选的进一步的cd26+和/或cd62l+细胞。优选地,选择细胞样品中的cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-细胞,和任选的进一步的cd26+和/或cd62l+细胞;更优选地,选择细胞样品中的cd127+cd117+lin-crth2-cd94-细胞,和任选的进一步的cd26+和/或cd62l+细胞。本领域的常规技术,例如实施例中列出的技术,可用于选择这些细胞。在优选的实施方案中,使用本领域熟知的方法分选细胞。在一些实施方案中,使用facs或
扩增ilcp的方法
本发明涵盖添加ilcp增殖的方法。当它们与il-1β一起培养时,观察到强烈的细胞增殖。对于ilcp的增殖,培养基包含il-1β,优选il-1β和il-2。此外,培养基可任选地包含其他细胞因子,例如il-7。
可以如实施例中所示(例如,实施例3)或通过其他类似技术生长细胞。例如,可以在yssel培养基中培养ilc,其中可以使用人ab血清、基质细胞、il-7、il-2和il-1β。或者,可以使用其他培养基,例如dmem、imdm或rpmi-1640。如本领域已知,用于细胞培养的培养基和/或培养基补充剂可以变化。还预期用哺乳动物血清补充细胞培养基。
培养基优选含有选自以下的血清:牛血清、小牛血清、胎牛血清、新生小牛血清、山羊血清、马血清、人血清、鸡血清、猪血清、绵羊血清、兔血清和大鼠血清,或血清替代物或胚胎液。可以在培养基中添加额外的补充剂,例如氨基酸。也可以在培养基中添加抗生素和抗真菌剂。
制备ilc1、ilc2、ilc3和nk细胞的方法
本发明涵盖制备ilc1、ilc2、ilc3和nk细胞的方法。可以通过本领域的常规技术从本发明的ilcp产生ilc1、ilc2、ilc3和nk细胞。例如,可以使用实施例中公开的特定技术产生ilc1、ilc2、ilc3和nk细胞。在一个实施方案中,可以使用细胞系统(例如,mohtashami等(2010)中公开的op9基质细胞系统)从本发明的ilcp产生ilc1、ilc2、ilc3和nk细胞。最优选地,细胞扩增而表型或功能没有变化,也称为“可塑性”。
在各种实施方案中,用各种细胞因子处理ilcp以促进分化成ilc1、ilc2、ilc3和nk细胞。这些细胞因子包括il-1β、il-12、il-18、il-25、il-33、il-23、il-2和il-7的任何和所有组合。
在各种实施例中,可以使用基于基质细胞的方法来扩增ilc亚组。虽然其他人也已经表明成熟的ilc可以在体外扩增,但发明人的结果是不同的,因为在那种情况下细胞扩增而没有“可塑性”(效应物功能改变,特别是细胞因子产生,例如ifn-γ)。这种可塑性可以由特定的人细胞因子(il-12)驱动,如发明人在ilc2的早期出版物中所示(lim等,jexpmed2016)。使用相同的方法,可以以最小的可塑性在体外扩增ilc3亚组。该方法可用于生长大量成熟的ilc2或ilc3而不改变其功能特性。
可以从以下扩增ilc亚组:分离的ilcp产生的ilc亚组或直接从患者样本中分离的ilc亚组。
可以如实施例中所述或通过其他类似技术生长细胞。例如,可以在yssel培养基中培养ilc,其中可以使用人ab血清、基质细胞、il-7、il-2和il-1β。或者,可以使用其他培养基,例如dmem、imdm或rpmi-1640。如本领域已知,用于细胞培养的培养基和/或培养基补充剂可以变化。还考虑用哺乳动物血清补充细胞培养基。
培养基优选含有选自以下的血清:牛血清、小牛血清、胎牛血清、新生小牛血清、山羊血清、马血清、人血清、鸡血清、猪血清、绵羊血清、兔血清和大鼠血清,或血清替代物或胚胎液。可以在培养基中添加额外的补充剂,例如氨基酸。也可以在培养基中添加抗生素和抗真菌剂。
最优选地,使用含有il-7、il-2和il-1β的培养基。
在没有基质细胞的情况下,使用细胞因子在体外扩增人ilcp。其他细胞系可用于培养ilcp。
ilcp的优选细胞来源是外周血,但也可包括骨髓、扁桃体、淋巴结、皮肤、脂肪组织、肠、肝和肺。可以在体外培养来自所有这些不同组织的ilcp,并产生成熟的ilc亚组。
可以将特定的生长因子组合添加到培养基中以将ilcp分化成特定的亚组。例如,可以添加il-12和il-18以产生ilc1亚组。可以添加il-25和il-33以产生ilc2亚组。可以添加il-23以产生ilc3亚组。
可以将特定的生长因子组合添加到培养基中以抑制ilcp分化成特定的亚组。例如,改变tbet或tbx21表达的小分子、化学试剂或遗传修饰可用于抑制分化成ilc1亚组。改变bcl11b表达的小分子、化学试剂或遗传修饰可用于抑制分化成ilc2亚组。改变rorγt(小鼠中的rorc)表达的小分子、化学试剂或遗传修饰可用于抑制分化成ilc3亚组。
op9细胞系可通过atcc(开放获取)获得。先前已经使用op9细胞来开发早期人t细胞前体,例如在us8,772,028和us9,533,009中。
以最小可塑性扩增ilc3的方法不同于lim等,jexpmed2016中所述的用于ilc2的方法。对于ilc2,用il-2、il-7、il-25和il-33(在op9上)培养成熟ilc2(从血液分离)。对于ilc3,用il-2、il-7和il-1β培养成熟ilc3。(参见实施例3,其中成熟ilc3分离自扁桃体,并用il-2、il-7和il-1β在op9-dl4上培养)。
除扁桃体外,产生ilc3的方法已成功用于胎肝、脐带血、成人外周血、肺、脂肪和肠样品。
也可以用crispr、zfn或talen或其他基因组编辑技术修饰ilcp或ilc1、ilc2或ilc3细胞,以添加或消除所需的基因组序列。也可以使用载体(包括逆转录病毒、aav和慢病毒载体)修饰这些细胞。在各种实施方案中,使选自rorc或rortγt、bcl11b、tbet和tbx21的基因失活。
还可以修饰细胞以包含嵌合抗原受体(car)。这些car通常包含例如来自单克隆抗体或纳米抗体的单链结合结构域,其与跨膜结构域和内结构域融合,导致响应于结合结构域与其靶标的结合而传递信号。car的实例在本领域中是众所周知的。这种基因工程化受体可用于将单克隆抗体的特异性移植到成熟ilc上。表达car的ilc可用于某些自身免疫疾病,因为一些ilc亚组(例如ilc2)通过髓样细胞抑制免疫应答。
在各种实施方案中,在体内施用ilcp以促进分化成ilc1、ilc2、ilc3和nk细胞。
在各种实施方案中,该方法包括提供先天性淋巴细胞前体(ilcp)群,使细胞群经受外部刺激,并检测选自ilc1、ilc2、ilc3和nk细胞的至少一种细胞类型的增加。在各种实施方案中,分离、纯化和/或收获ilc1、ilc2、ilc3和/或nk细胞。
可以使细胞群在体内或体外经受外部刺激。在一些实施方案中,在人源化小鼠模型中使细胞群经受外部刺激。在各种实施方案中,外部刺激是病毒、寄生虫、微生物或细菌生物(例如hiv或疟疾)或其组分(例如dna或蛋白质)。在各种实施方案中,外部刺激是细胞因子或细胞因子的混合物。在各种实施方案中,外部刺激物是测试化合物。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有cd34-cd7+cd127+cd117+cd45ra+表型,缺少nkp44和rorγt表达,和/或是il-1r1+和cd69-,并且任选地进一步表达cd62l和/或cd26。优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有lin-cd34-cd7+cd127+cd117+cd45ra+表型,缺少nkp44和rorγt表达,和/或是il-1r1+和cd69-,并且任选地进一步表达cd62l和/或cd26。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd7+cd127+cd117+crth2-。优选地,所述群中至少75%、80%、85%、90%、95%或99%的细胞是cd7+cd127+cd117+crth2-;更优选地,所述群中至少90%、95%或99%的细胞是lin-cd7+cd127+cd117+crth2-。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117+。优选地,所述群中至少75%、80%、85%、90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117+;更优选地,群中至少90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117+;还更优选地,所述群中至少99%或100%的细胞是lin-cd94-crth2-cd127+cd117+。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+cd3-crth2-。更优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+cd3-crth2-,且进一步是cd7+、nkp44-、cd94-和/或lin-,和/或进一步是cd26+,和/或cd62l+。在该方法的一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。优选地,所述群中至少75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。在一些优选的实施方案中,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
用于治疗的方法和组合物
本发明涵盖包含先天性淋巴细胞前体(ilcp)的组合物用于治疗需要先天免疫系统调节的患者的用途。因此,本发明涵盖这些化合物用于治疗患者的用途和治疗他们的方法。
在各种实施方案中,患者具有蠕虫感染、肠道病原体感染、肿瘤、病毒感染、过敏、哮喘、炎症或自身免疫疾病(例如,多发性硬化、系统性红斑狼疮或i型糖尿病)。
在各种实施方案中,患者可以是免疫缺陷、免疫受损或免疫抑制的。在各种实施方案中,患者是癌症患者或患有慢性疾病(例如克罗恩病、ibd)。
在各种实施方案中,本发明包括治疗人患者的方法,包括向所述患者施用纯的ilcp群,其中所述群中至少90%的细胞具有表型cd34-cd7+cd127+cd117+cd45ra+,缺乏nkp44和rorγt表达,和/或是il-1r1+和cd69-,和/或任选的cd26+和/或cd62l+。
在各种实施方案中,本发明包括治疗人患者的方法,包括向所述患者施用纯的ilcp群,其中所述群中至少90%的细胞具有表型lin-cd34-cd7+cd127+cd117+cd45ra+,缺乏nkp44和rorγt表达,和/或是il-1r1+和cd69-,和/或任选的cd26+和/或cd62l+。在一些实施方案中,所述群中至少90%、95%或99%的细胞是lin-cd7+cd127+cd117+crth2-。在一些实施方案中,所述群中至少90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117+。
优选地,所述群中至少90%、95%或99%的细胞是cd127+cd117+cd3-crth2-。更优选地,所述群中至少90%、95%或99%的细胞是cd127+cd117+cd3-crth2-,且进一步是cd7+、nkp44-、cd94-和/或lin-,和/或进一步是cd26+,和/或cd62l+。在一些优选的实施方案中,所述群中至少90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。优选地,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
在各种实施方案中,取决于待治疗的疾病,单能和多能ilcp,尤其是多能ilcp,可以与适当的因子组合以使它们在体外或体内分化成特定类型(ilc1、ilc2或ilc3)。在一些实施方案中,取决于疾病,增加或抑制ilcp的分化或抑制分化成特定类型可能是有益的(参见例如wo2016/138590、us2016/0145344和us2016/0304574,其在此通过引用并入)。
ilc亚组涉及各种疾病和细胞过程,包括感染、癌症炎症、组织修复和体内稳态。taitwojne等,2016通过引用并入本文。由于ilc3促进肠中的galt形成、炎症、免疫和体内稳态(同上),因此通过本发明的方法产生的ilc3可用于治疗涉及这些过程的疾病。由于ilc2通过与造血细胞和非造血细胞的相互作用影响炎症、免疫、组织修复和体内稳态(同上),因此通过本发明的方法产生的ilc2可用于治疗涉及这些过程的疾病。由于ilc1表达t-bet和ifn-γ并且有助于1型炎症(同上),因此通过本发明的方法产生的ilc1可用于治疗涉及这些过程的疾病。
可以通过本领域的常规技术向患者施用细胞。优选地,向患者施用104、105、106、107、108、109或1010个ilcp细胞。
可通过直接注射到组织或血液等向受试者,优选人施用自体、同种异体或异种ilc或ilcp。优选地,将细胞与药学上可接受的载体组合施用。可以注射单次或注射至少2、3、4、5次等施用细胞。可以对细胞进行遗传修饰以改变其免疫识别。
使用ilcp的筛选方法
本发明涵盖用于筛选调节(即,抑制或增强)ilcp向ilc1、ilc2、ilc3和/或nk细胞分化的化合物的方法。在各种实施方案中,将本发明的ilcp群在体内或体外与测试化合物接触,并评估化合物对分化的影响。这种影响可以通过检测细胞群中细胞表型的变化观察。
测试化合物可以是天然化合物或合成化合物。在各种实施方案中,测试化合物是病毒、寄生虫、微生物或细菌生物(例如hiv或疟疾)或其组分(例如dna或蛋白质)。在各种实施方案中,测试化合物是细胞因子或细胞因子的混合物。
在一些实施方案中,通过测量与测试化合物接触之前和之后细胞群中ilcp、ilc1、ilc2、ilc3和/或nk细胞的水平来检测细胞群中细胞表型的变化。可以如实施例中所公开的和通过本领域技术人员已知的类似技术检测细胞的表型。在一些实施方案中,将与测试化合物接触后的ilcp、ilc1、ilc2、ilc3和/或nk细胞的水平与未处理的ilcp对照进行比较。
在一个实施方案中,本发明包括用于筛选影响ilc发育的化合物的方法,包括提供先天性淋巴细胞前体(ilcp)群,使细胞群与测试化合物接触,并检测细胞群中细胞表型的变化。
在一些实施方案中,测试化合物导致ilcp分化减少。在一些实施方案中,测试化合物导致ilcp分化增加。在一些实施方案中,测试化合物导致分化为特定ilc亚组的减少。在一些实施方案中,测试化合物导致分化为特定ilc亚组的增加。
在一些实施方案中,该方法包括将ilcp与能够分化它们的刺激物(例如,op9-dl4培养系统)组合,并使细胞群与测试化合物接触以测定化合物对分化的影响。在其他实施方案中,在不存在这种刺激物和/或添加其他化合物或刺激物(例如细胞因子)的情况下,测定化合物的影响。
在一些实施方案中,该方法包括向小鼠输注先天性淋巴细胞前体(ilcp)群,并向小鼠施用测试化合物。优选地,小鼠是人源化小鼠。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有表型cd34-cd7+cd127+cd117+cd45ra+,缺乏nkp44和rorγt表达,是il-1r1+和cd69-,和/或任选的cd26+和/或cd62l+。
优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞具有表型lin-cd34-cd7+cd127+cd117+cd45ra+,缺乏nkp44和rorγt表达,是il-1r1+和cd69-,和/或任选的cd26+和/或cd62l+。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd7+cd127+cd117+crth2-。
在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是lin-cd94-crth2-cd127+cd117+。更优选地,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+cd3-crth2-。在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞进一步是cd7+、nkp44-、cd94-和/或lin-,和/或进一步是cd26+,和/或cd62l+。在一些实施方案中,所述群中至少25%、35%、50%、60%、75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。优选地,所述群中至少75%、80%、85%、90%、95%或99%的细胞是cd127+cd117+llin-crth2-(其中lin-包含cd3-),且任选地进一步是cd7+、nkp44-和/或cd94-,和/或任选地进一步是cd26+和/或cd62l+。在一些优选的实施方案中,所述群中至少90%的细胞具有表型cd127+cd117+lin-crth2-cd7+或cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+;更优选地,所述群中至少99%或100%的细胞具有表型cd127+cd117+lin-crth2-cd94-,和任选的cd26+和/或cd62l+。
具体实施方式
材料和方法
人血液和组织样品
健康供体的血液样品来自与巴斯德研究所签署了协议的establissemet
人免疫系统(his)小鼠模型
已经描述了balb/crag2-/-il2rg-/-sirpanod(brgs)小鼠,其被保持在巴斯德研究所的隔离器中。使用facsaria从健康供体的外周血中分选cd34+hsc或cd117+ilc。使用cd34microbeadkit(miltenyi)分离胎肝cd34+hsc。对于体内转移实验,将1-3×105个cd117+ilc或cd34+hsc与0.3μgil-2和-7(miltenyi)一起肝内注射到经亚致死辐射(3gy)的新生(3-7天龄)brgs小鼠中。小鼠每周通过腹膜内注射接受il-2、-7、-1β、-23、-25和-33(各0.3μg),并在移植后4周分析。为了制备his小鼠,将源自胎肝的cd34+hsc肝内注射到经亚致死辐射(3gy)的新生(3-7天龄)brgs小鼠中。注射后8-9周处死小鼠。实验由巴斯德研究所的伦理委员会批准,并由法国教育和研究部验证。
从血液、扁桃体、肠、胎肝和肺分离细胞
通过ficoll-paque(gehealthcare)密度梯度离心从cb和pb分离人外周血单核细胞(pbmc)。用机械破碎通过70-μm过滤器获得胎肝和扁桃体的单细胞悬浮液。将肺和肠样品切碎,并在37℃振荡培养箱中用liberasetl(25μg/ml;roche)和dnasei(50μg/ml;sigma-aldrich)消化45分钟。将消化的组织通过70-μm过滤器。通过ficoll-paque密度梯度离心从肝、肺和肠分离淋巴细胞。
facs分析和细胞分选
对于facs分析,首先用flexibleviabilitydyeefluor506(ebioscience)将细胞染色10分钟,然后在冰上用brilliantstainedbuffer(bd)进行表面抗体染色20分钟。对于涉及胞内tf染色的实验,使用foxp3/转录因子染色缓冲试剂盒(ebioscience)将细胞固定、透化并染色。对于胞内的细胞因子染色,在golgiplug(bd)存在下用pma(10ng/ml;sigma)加离子霉素(1μg/ml;sigma)刺激细胞3小时。将细胞固定、透化并用cytofix/cytoperm试剂盒(bd)染色。在lsrfortessa(bd)上获得样品,并通过flowj10(treestar)分析。
对于来自健康pb的细胞分选,根据制造商的说明,首先通过用生物素偶联的抗cd3、抗cd4、抗cd19、抗cd14、抗cd123标记,然后通过抗生物素微珠(miltenyi)耗尽pbmc中的t细胞、b细胞、pdc和单核细胞。用谱系消耗进行cb和组织的分选。将批量群体分类至纯度≥99%,或作为单细胞指数分选(均使用facsariaii;bd)。
批量rna分离、文库构建、测序和分析
将来自每个群体的103个细胞直接facs分选到50μl裂解/结合缓冲液(lifetechnologies)中。用dynabeadsoligo(dt)(lifetechnologies)捕获mrna,洗涤,并在70℃下用10μl10mmtris-cl(ph7.5)洗脱。使用如所述的针对单细胞rna-seq开发的mars-seq衍生版(24531970)来制备表达文库,其中每个群体最少两个重复。对平均每个文库4百万个读段(read)进行测序,并使用默认参数的tophatv2.0.10与人参考基因组(ncbi)比对(19289445)。使用homer软件(homer.salk.edu)计算表达水平,并将每个样品归一化至总读段数。关注高度表达的基因,其在任何两个亚型的平均值之间相对于噪声(8个读段)具有2倍差异。使用david进行kegg分析(12734009)。
染色质免疫沉淀,并用chipmentation进行测序(chip-seq)
在室温下,将facs分选的细胞(20-50k)在含有1%甲醛(sigma)的pbs中立即交联10分钟,用于chip-seq分析。通过加入甘氨酸(0.125m终浓度),然后在室温下孵育5分钟淬灭交联。将细胞置于冰上,用pbs洗涤,并快速冷冻以在-80℃下储存。平行处理沉淀以使技术变化最小化。将细胞重悬于100μl超声缓冲液(1%sds、10mmedta、50mmtris-hclph8和1x无edta的完全蛋白酶抑制剂;roche),并转移至0.65ml的bioruptor超声处理管(diagenode)。在冰上孵育15分钟后,使用bioruptorpico超声波仪(diagenode)将细胞超声处理30个循环(30秒on-30秒off),以将染色质剪切至±250bp片段。通过加入900μl的10xchip稀释缓冲液(0.01%sds、1.1%tritonx-100、1.2mmedta、16.7mmtris-hclph8、167mmnacl)平衡染色质,并在4℃下与1μl的h3k4me2特异性抗体(ab32356,abcam)或作为阴性对照的正常兔igg(sc-2027,santacruz)孵育过夜。另外,通过在4℃孵育过夜,将每个ip20μl的蛋白质adynabeads(thermofisherscientific)在含有0.1%bsa(sigma)的pbs中封闭。第二天,将初始体积的珠子重悬于chip稀释缓冲液中,并加入染色质提取物中。在4℃孵育2小时后,收集珠子,并用低盐缓冲液(0.1%sds、1%tritonx-100、2mmedta、20mmtris-hclph8、150mmnacl)、高盐缓冲液(0.1%sds、1%tritonx-100、2mmedta、20mmtris-hclph8、500mmnacl)和licl缓冲液(10mmtris-hclph8、1mmedta、250mmlicl、0.5%np-40、0.5%脱氧胆酸)洗涤。然后如最近所述(schmidl等,2015),将固定在磁珠上的染色质-抗体进行标记(tagmentation)。使用minelute离心柱(qiagen)纯化洗脱的dna,并使用nexterapcr引物扩增8-12个循环。将文库用双(0.5x-2.0x)spriampurexp珠(beckmancoulter)纯化,合并(最多每次测序运行10个),并在运行单读75bp程序的nextseq500(illumina)上测序。
chip-seq数据处理、分析和可视化
使用basespace(illumina)将读段解编,并使用标准设置的bowtie(langmead,b.&salzberg,s.l.fastgapped-readalignmentwithbowtie2.natmethods9,357-359,doi:10.1038/nmeth.1923(2012))将其比对至小鼠基因组(mm10build),除去无法唯一映射的读段。将索引且分选的bam文件解析为homer(heinz,s.等.simplecombinationsoflineage-determiningtranscriptionfactorsprimecis-regulatoryelementsrequiredformacrophageandbcellidentities.molcell38,576-589,doi:10.1016/j.molcel.2010.05.004(2010)),用于进一步分析。为每个样品生成标记目录,去除重复读段(-tbp1选项)。使用makeucscfilehomer脚本生成显示标准化计数(每百万读段)的bedgraph文件,以便在ucscgenomebrowser(https://genome.ucsc.edu/)中直接可视化。使用具有-region-size1000-mindist2500选项的homerfindpeaks鉴定富含h3k4me2的区域。使用bedtools的交叉功能或需要最小重叠1bp的homermergepeaks脚本(-d给定选项)鉴定两个样品之间的重叠和非重叠区域(quinlan,a.r.&hall,i.m.bedtools:aflexiblesuiteofutilitiesforcomparinggenomicfeatures.bioinformatics26,841-842,doi:10.1093/bioinformatics/btq033(2010))。使用javatreeview将一组细胞类型特异性的h3k4me2+区域可视化为热图(saldanha,a.j.javatreeview--extensiblevisualizationofmicroarraydata.bioinformatics20,3246-3248,doi:10.1093/bioinformatics/bth349(2004))。将区域/峰指定为推定的靶基因great(mclean,c.y.等.greatimprovesfunctionalinterpretationofcis-regulatoryregions.natbiotechnol28,495-501,doi:10.1038/nbt.1630(2010))。随后,使用全基因组作为背景,将great用于计算已知通路特征的这些基因的富集。
大批和单个细胞培养
所有体外培养实验均在补充有2%人ab血清(efs)的yssel培养基(18432890)中进行。在培养前一晚,将2-3×103个基质细胞预先接种在96孔圆底板中。使用imdm(invitrogen)加0.25%(w/v)牛血清白蛋白(sigma)、1.8μg/l2-氨基乙醇、40μg/lapo-转铁蛋白、5μg/l胰岛素和青霉素/链霉素自己制备yssel培养基。对于大批培养,将100-300个facs分选的细胞铺板在基质细胞上。对于克隆实验,将细胞直接索引分选到预先接种有基质细胞的96孔板中。如所示的以各种组合提供细胞因子il-2、-7(各20ng/ml,miltenyi)、il-12、-18、-25、-33、-1β、-23(各20ng/ml,r&d)。对于大批培养,每5天补充新鲜细胞因子和培养基,并在扩增10天后进行分析。对于克隆实验,每7天补充细胞因子和培养基,并在培养14-18天后进行分析。
实施例1:表征人外周血cd117+ilc
除非另有说明,否则数据表示为中值。每个实验的样品大小和重复的实验数量均包含在图例中。可以在健康个体以及患有多种临床综合征的患者的谱系cd7+cd56-cd12+外周血(pb)细胞中将循环ilc鉴定为低频率群体(<总cd45+细胞的0.2%)(hazenberg和spits,2014;munneke等,2014;图1a和1b)。使用鉴定胎儿组织和扁桃体中ilc亚组的表型标志物,包括cd161、crth2、cd117和nkp44,进一步将pbilc分级为ilc1、ilc2和ilc3(spits等,2013)。因此,之前的报道已经在人血液中鉴定了循环ilc2(cd161+crth2+gata-3+细胞)和ilc1(cd161+crth2-cd117-t-bet+细胞)(
pbcd117+ilc不表达表征其他已知ilc亚组的特征性tf(即:t-bet、eomes、gata-3hi)(图1c)。因此,pbcd117+ilc不能表达与nk细胞、ilc1和ilc2相关的标志物,例如cd94、cd244、crth2(图1a),并且在用药理学活化剂刺激后不产生ifn-γ或il-13(图1d)。总之,这些结果表明pbcd117+ilc不代表任何经典的ilc子亚组。
实施例2:cd117+ilc的转录和染色质图谱揭示ilc前体谱
由于cd117在血液淋巴样祖细胞上高度表达(ikuta和weissman,1992;kikushige等,2008),因此假设pbcd117+ilc可能包括未定型的淋巴前体。为了进一步了解pbcd117+ilc的特性,对高纯度循环cd117+ilc的转录组和表观遗传图谱进行分析,并与cd34+hsc进行比较(图2a);后者代表具有多谱系潜力的未成熟造血祖细胞(baum等,1992;mohtashami等,2010)。
进行染色质免疫沉淀,然后使用转座酶介导的标记(chipm-seq)进行高通量测序,其允许发明人直接分析少量纯化细胞的表观基因组。为了暴露cd34+hsc和cd117+ilc的普通和独特的表观遗传特征,定位组蛋白h3赖氨酸4二甲基化(h3kme2),因为它标记活性和平衡基因调控元件(gre)的精确度高于其他组蛋白修饰(zhang等.2012cell,koche等,cellstemcell2011)。在cd117+ilc和cd34+hsc中分别鉴定了约18,000和35,000个gre(图2b),其中大部分位于内含子和基因间区域(补充图x)。两种细胞类型共有大量的h3k4me2+gre:在cd117+ilc中鉴定的89%的gre在hsc中显示相似的富集,并且与13159个基因相关,其中许多编码管家功能。然而,在cd117+ilc中检测到的11%的h3k4me2+gre在cd34+hsc中不存在,其可能调节2283个基因。这些基因的通路分析显示免疫系统和淋巴细胞相关过程的强烈富集(图2c)。例如,对淋巴发育和功能至关重要的细胞因子/趋化因子信号传导基因,如il1r1、il7r、il2ra/b与cd117+ilc特异性gre相关。相反,仅在cd34+hsc中有活性的gre(cd34+hsc中所有gre的54%)位于涉及对造血作用(包括止血、血小板活化和notch信号传导通路)重要的更一般的通路基因附近(图2c)。
为了比较cd117+ilc和cd34+hsc的转录组,进行rna测序(rna-seq)。出现了基因表达谱的明显差异,其中一大簇1540个基因在cd117+ilc中以显著更高的水平表达(图2d)。其中存在许多与淋巴谱系密切相关的基因,包括ikzf1、cd2、cd7和il7r(图2d)。相反,cd34+hsc细胞高度表达参与多种造血谱系广泛发育的基因,包括id1、gata1、gata2和myb(图2d)以及髓样谱系的细胞因子受体(csf3r、csf2rb、flt3)。与hsc相比,cd117+ilc表达高水平的已被证明对小鼠ilc发育至关重要的tf,包括id2、gata3、tox和tcf7。在cd117+ilc中未检测到t和b细胞发育特征性的转录物,例如rag1、rag2、ebf1、cd3e、bcl11a或lmo2,尽管hsc表达这些基因中的一些。
由于cd117+ilc的转录组学和表观遗传学分析均鉴定了强烈的淋巴特征,因此将这些数据集相交以深入了解cd117+ilc的发育状态。在cd117+ilc中最高表达的相当大比例(26%)的基因直接位于cd117+ilc特异性gre的附近(图2e)。令人惊讶的是,这些包括许多先前涉及小鼠ilc发育的转录因子,包括id2、gata3、ets1、tox、tcf7、rora和notch1(图2e)-这与cd117+ilc定型为先天性淋巴命运一致。相反,未检测到任何成熟ilctf(eomes、tbx21、rorc)、细胞因子受体(ccr6、il1rl1、il23r)或特征性细胞因子(ifng、il13、il5、il22、il17a)的显著表达水平。然而,这些成熟的ilc特征基因中的一些已经被h3k4me2标记,证明它们以平衡状态驻留(图2e)。相反,b和t细胞发育的关键调节基因(rag1、rag2、ebf1、bcl11a、hes1、lmo2)未选择性地用h3k4me2标记。总之,这些分析表明,cd117+ilc代表携带tf表达谱的淋巴样偏向祖细胞,其类似于具有表观遗传平衡状态的关键成熟ilc特征基因的多能ilc前体(ilcp)。
实施例3:外周血cd117+ilc包括多能ilc前体(ilcp)
为了评估循环cd117+ilc的造血潜力,将这些细胞在各种细胞因子的存在下进行大批培养。当cd117+ilc表达cd25、cd127和cd121a(il-1r1)(图1a,1c)时,在这些培养物中加入il-2、il-7和/或il-1β。尽管大批培养物在il-2和il-7存在下扩增最小,但当细胞在il-1β中培养时观察到强烈的增殖(图3a)。可以驱动ilc1/nk(il-12、-18)、ilc2(il-25、-33)或ilc3(il-23)发育的细胞因子的额外存在不会比用il-1β获得的进一步增加细胞产量(图3a)。大批培养的细胞不含b(cd19+)或t(cd3+cd5+)细胞,但包含纯的cd7+细胞群,其是cd16+并表达各种水平的cd117和cd25(图3b)。
值得注意的是,扩增的细胞包括一些eomes+cd94+nk细胞以及代表三个经典ilc组的细胞:ifn-γ+ilc1、il-13+ilc2和nkp44+il-17a+il-22+ilc3(图3b)。虽然补充il-25和il-33没有明显改变这些培养物中ilc亚组的分布,但il-12的加入明显促进了产生eomes+cd94+ifn-γ的nk细胞的发育,且il-23对于产生il-17a的ilc3至关重要(图3b)。这些结果不仅定义了支持多谱系ilc和nk细胞生成的细胞因子“混合物”(il-2、il-7、il-1β、il-23),而且表明pbcd117+ilc含有多谱系ilc前体(ilcp)。
使用改良的基于基质细胞的培养系统进一步表征循环cd117+ilc的多谱系潜力,所述培养系统允许b细胞、t细胞和骨髓细胞发育(图3c;mohtashami等,2010)。此外,该系统可以在克隆水平上以最小可塑性广泛扩增人nk细胞和ilc亚组(lim等,2016)。分析在op9和op9-dl4上培养的单个pbcd117+ilc的后代,以鉴定产生ifn-γ、il-13、il-17a和/或il-22的eomes+nk细胞和ilc亚组(图3d)。op9-dl4基质表达强notch配体,允许发明人评估触发该通路的影响(mohtashami等,2010)。发明人对超过340种克隆培养物的分析允许证明几个点。首先,pbcd117+ilc代表单能和多能ilc前体(ilcp)的异质群体。大约一半来自单个cd117+ilc的培养物产生单个ilc亚组(仅ilc1、ilc2或ilc3),因此代表谱系限制的ilcp,而其余的是多能ilcp,其可以产生2个或更多个单独的lin-cd7+ilc谱系(图3e)。未观察到b细胞和t细胞潜能。其次,在多能ilcp群中,大部分(9-17%)能够产生所有三个ilc亚组,并且可能代表未成熟的未定型ilcp。此外,克隆ifn-γ+培养物还包含eomes+nk细胞,证明一些pbilcp具有在单细胞水平产生“辅助”和“细胞毒性”ilc谱系的潜力。第三,lin-cd7+ilc克隆的亚组未能产生任何测试的细胞因子(图3e)。由于这些克隆保持高水平的cd7和cd117但缺乏其他ilc标志物,它们可能代表没有进一步分化的ilcp。第四,notch信号明显影响cd117+ilcp的细胞命运潜力,因为含有ilc3的细胞的多能性和发育在op9-dl4上得到增强(图3e)。总之,这些数据将pbcd117+ilc鉴定为定型ilc祖细胞的循环池。大批和克隆测定的比较清楚地证明了单细胞方法在定义cd117+ilc细胞命运潜力的异质性和建立功能多能性方面的重要性。
实施例4:循环cd117+ilcp在人源化小鼠体内具有多ilc潜力
接下来评估pbcd117+ilcp的体内潜力。之前的研究已经证明,用人cd34+造血干细胞(hsc)祖细胞移植的严重免疫缺陷小鼠品系能够产生人淋巴细胞(b、t、nk)和骨髓(dc、巨噬细胞、中性粒细胞)细胞亚组(综述于(shultz等,2012))。使用balb/crag2-/-il2rg-/-sirpanod(brgs)小鼠,其允许稳健的多谱系人造血细胞移植(legrand等,2011)。将来自相同供体的人pbcd34+hsc和cd117+ilcp过继转移至新生brgs小鼠;提供细胞因子补充(il-2、il-7、il-1β、il-23),并在4周后分析小鼠(图4a)。植入人cd34+hsc的brgs小鼠在骨髓中发育cd19+b细胞和cd14/cd33+骨髓细胞,而在肠中检测到cd3/cd5+t细胞和lin-cd7+nk/ilc(图4b)。相反,接受pbcd117+ilcp的brgs小鼠发育lin-cd7+细胞,但没有发育髓样细胞、b细胞或t细胞。在多个器官中检测到来自转移的cd117+ilcp的人cd45+后代,包括脾、肺、肠和肝(图4c)。在这些组织位点中的每一个,可以鉴定eomes+nk细胞以及不同的cd127+ilc亚组,其一经刺激离体产生ifn-γ、il-13、il-17a和/或il-22(图4c)。这些结果表明,pbcd117+ilcp能够在体内产生所有已知的ilc亚组和nk细胞。由于pbcd117+ilcp缺乏骨髓、b和t细胞潜力,总结这些细胞包含定型的ilc祖细胞。
实施例5:在体内从cd34+hsc发育人cd117+ilcp
接下来研究cd34+hsc和cd117+ilcp之间的发育关系。用纯化的cd34+hsc移植免疫缺陷的新生brgs小鼠,并在8-9周后处死(图5a)。在骨髓、肺、肝和脾中分析人cd45+细胞。如所预期的(legrand等,2011),这些不同的组织位点包含人cd45+细胞,包括多种谱系+t、b和骨髓细胞(图5b,数据未显示)。此外,在lin-cd7+细胞亚组内,可以在缺乏t-bet和eomes表达的多个组织中辨别出明确界定的cd127+cd117+细胞亚群(图5b)。这些包括cd127+cd117+细胞,其表达低水平的gata-3和rorγt,并且是nkp44-(图5d),因此类似于pbcd117+ilcp。离体刺激不能引发cd127+cd117+细胞产生细胞因子(图5e)。分选这些细胞,并在il-2、il-7、il-1β和il-23存在下进行大批培养。扩增的细胞含有能够产生ifn-γ、il-13、il-17a和il-22的亚组(图5e),从而证实了人ilcp的存在。这些结果表明,cd34+hsc可在体内产生cd117+ilcp。
实施例6:人cd117+ilcp存在于胎肝、脐带血和成人肺
接下来评估出现人cd117+ilcp的发育阶段。首先研究胎肝(fl)中的人ilc亚组,因为已经表明该器官携带几个未成熟的造血前体群(rollini等,2007),并且该器官被提出作为小鼠中淋巴组织诱导细胞发育的视角(cherrier等,2012)。fl中的lin-cd127+ilc主要含有cd117+亚组。有趣的是,这些细胞表达的rorγt水平超过其外周血对应物(图1c),而且表达ccr6、neuropilin-1(nrp-1)但不表达nkp44。尽管存在这些差异,但flcd117+ilc在刺激后未产生大量的il-17a或il-22,表明它们不是完全成熟的ilc3。然而,当在体外扩增flcd117+ilc时,产生大量的产il-17a的ilc3。此外,il17a+ilc3在缺乏dl4的基质细胞上发育,表明该过程不需要额外的notch参与(图6a)。有趣的是,flcd117+ilc的大批培养物也含有可检测的产il-13和ifn-γ的细胞,尽管频率较低。克隆分析显示,如预期的,flcd117+ilc含有高比例的ilc3定型祖细胞。尽管如此,该群体的大部分携带在notch配体存在下显示更清楚的多能ilcp(图6b、c)。这些结果证明,人fl携带可以产生所有已知的ilc亚组的cd34-cd127+cd117+多能ilcp。ilc3定型的祖细胞在该组织位点中富集,表明在此期间环境信号可指导多能ilcp进一步规格化为ilc3命运。
接下来表征来自人脐带血(cb)的cd117+ilc。与它们的pb对应物一样,cbcd117+ilc缺乏nkp44表达以及ccr6和nrp-1表达,并且是cd45ra+。此外,cbcd117+ilc不表达rorγt和t-bet,但是为gata-3lo,因此类似于pbilcp。与pbcd117+ilc一样,cbcd117+ilc在刺激后离体不产生细胞因子(ifn-γ、il-13、il-17a或il-22)。然而,在il-2、il-7、il-1β和il-23中大批培养cbcd117+ilc产生了多种产细胞因子的ilc亚组,包括ifn-γ+ilc1,il-13+ilc2和il-17a+或il-22+ilc3(图6d)。在cbcd117+ilc培养物中未检测到t、b或髓样细胞(数据未显示)。进一步的克隆分析显示,cbcd117+ilc携带单能和多能ilcp的各种混合物(图6e和6f)。与flcd117+ilc不同,cbcd117+ilc不偏向于ilc3定型祖细胞,而是与pbcd117+ilcp更接近。对于来自pb或fl的ilcp,notch刺激导致多能ilcp(特别是具有il-17a+和il-22+ilc3潜力的那些)的频率增加,并且降低细胞因子-ilc克隆的频率,表明该通路促进了特定ilc亚组的定向发育。
还检查了来自成人肺组织的cd117+ilc的表型和潜力。肺cd117+ilc携带nkp44+和rorγt+ilc的离散群体,但主要是cd45a-。大批培养肺cd117+ilc产生多种产细胞因子的ilc亚组和eomes+nk细胞(图6g);使用克隆测定的进一步分析将肺cd117+ilc的nkp44-部分定义为单能和多能ilcp的混合物(图6h和6i)。这些结果表明,在人粘膜组织中存在多种ilcp,包括多能祖细胞。
实施例7:ilc前体驻留于次级淋巴组织
人次级淋巴组织(淋巴结、扁桃体)携带不同的ilc亚组及其前体(bernink等,2013;cella等,2009;fehniger等,2003;
为了更好地理解nkp44-和nkp44+cd117+ilc之间的关系,制备来自两个亚组的克隆,并分析它们的细胞因子产生潜力。观察到显著的差异。衍生自nkp44+cd117+ilc的克隆是高度富集的ilc3,其产生il-17a和/或il-22(图6n和6o)。一部分共表达ifn-γ(14%)的克隆可能代表“塑性”ilc3,其可以如先前所示上调t-bet(bernink等,2015)。相反,来自nkp44-cd117+ilc的克隆非常异质,其中细胞不仅产生il-22和/或il-17a,而且还产生丰富的单个ifn-γ+克隆以及单个il-13+克隆(图6k和6l),这在nkp44+cd117+ilc中没有检测到(图6n和6o)。观察到ifn-γ+ilc1克隆这一事实出乎意料,因为之前有报道称扁桃体cd127+ilc1在il-2、il-23和il-1β存在下分化为产生il-22的ilc3(bernink等,2015)。最后,仅在nkp44-cd117+ilc中发现了产生三种ilc亚组的多能ilcp。总之,这些结果表明,扁桃体cd117+ilc非常异质性,包含nkp44-ilcp和nkp44+ilc3。此外,本发明人使用克隆测定明显允许界定在大批培养水平上不可见的ilcp库。
实施例8:rorc缺陷患者携带ilcp但未能产生il-17a+ilc3
已经表明,人次级淋巴组织中具有cd34+cd45ra+cd117+表型的定型ilcp高度表达tfrorc(scoville等,2016)。由于cd117+ilcp在发育上位于cd34+hsc下游(图5),因此先前描述的cd34+cd45ra+cd117+ilcp亚组可能是该通路的中间体。为了解决是否需要rorc来产生人cd117+ilcp,对rorc缺陷患者进行了研究。人的rorc缺乏与皮肤粘膜念珠菌病有关,之前的研究表明,这种tf对预防真菌病原体的th17细胞的分化至关重要(okada等,2015)。研究了2名rorc缺陷患者的外周血细胞的ilc亚组(图7a)。在对照和rorc缺陷患者中,lin-cd+细胞均含有主要的cd56+nk细胞群(cd56bright和cd56dim),并且还清楚地检测到cd127+ilc的离散亚群。如前所述(okada等,2015),在rorc缺陷患者中注意到表达cd117的ilc的频率降低,尽管该群体仍然明显存在(图7a和7b)。相反,在没有rorc的情况下,存在ilc1,并且来自总ilc的ilc2的百分比显著增加(图7b)。如上所述,将来自对照和rorc缺陷患者的分选的cd117+ilc进行大批培养。观察到lin-cd7+细胞的稳健生长,且wt和rorc缺陷细胞之间没有显著差异。在这些培养物中鉴定了多种产生细胞因子的细胞,包括产生ifn-γ、il-13或il-22的细胞,然而,不存在产生il-17a的细胞(图7c)。eomes+ifn-γ+nk细胞的发育不受rorc缺失的影响。这些结果表明,rorc不是nk细胞、ilc1、ilc2或il-22+ilc3发育所必需的,但对于从人ilcp产生il-17a+ilc3是必需的。
实施例9:分析血液中的ilcp标志物
虽然ilcp表达cd127和cd117,但它们并不具有特异性。为了深入了解最少的必需标志物,分析了可用于高度富集ilcp的标志物组合。因此,使用不同标志物的facs来确定不同细胞类型的百分比,以从成人外周血中分离细胞。结果表示使用多参数facs分析外周血中人ilcp的富集的分析。使用不同的门控方案,可以估计ilcp以及可能存在的其他细胞类型(t细胞、nk细胞、ilc2)的富集。结果如图8a-f和图9a-g所示。
使用标志物cd127+cd117+细胞分选产生约20%的lcp和60-80%的t细胞(图8a和9a)。包括cd7,不包括crth2或两者的组合不足以富集cd127+cd117+细胞中的ilcp,因为污染t细胞占优势(图8a-d:图9a-d)。排除谱系细胞(包括cd3)强烈富集cd127+cd117+细胞中的ilcp(图8e-f;图9e-g)。在lin-cd127+cd117+细胞中排除crth2ilc2提供ilcp的进一步富集,产生至少75%的ilcp(图9e)。此效果不需要cd7(图8e和9f)。排除cd94允许从lin-cd127+cd117+crth2-细胞中分离纯ilcp(图8f和9g)。因此,使用标志物cd127+cd117+cd7+crth2-lin-的分选产生大约90%的ilcp(图8e和9f),而使用标志物cd127+cd117+crth2-lin-cd94-的分选产生大约100%的ilcp(图8f和9g)。
实施例10:其他ilcp标志物
另外筛选人ilcp(定义为lin-cd127+cd117+crth2-)上表达的可以作为分离这些细胞的有用替代物的其他细胞表面标志物。在人ilcp上鉴定了各种cd62l和cd26表达,其中大多数细胞是cd62l+,并且大部分细胞表达cd26(图10)。
克隆分析表明,所有这些亚组都携带多能ilcp(表1)。
表1:cd62l和cd26人ilcp亚组的克隆分析
将所示人ilcp亚组分选为单个细胞,并在补充有人il-1b、il-2、il-7和il-23的op9基质上培养。用pma/离子霉素刺激3小时后,分析克隆的细胞因子产生(ifng、il-13、il-17a、il-22)。示出了单能和多能ilcp的频率。推定的ilcp鉴定为“无细胞因子”。
这些结果鉴定了可用于分离人ilcp亚组的其他“任选的”标志物(cd62l、cd62)。
人ilcp扩增:分析人ilcp克隆(使用op9基质细胞和il-2、il-7、il-1β和il-23的组合)鉴定了未能表达任何测试细胞因子(ifn-g、il-13、il-17a、il-22)的细胞。这些ilcp“克隆”在数量上扩增了100-1000倍(图11)。分析这些细胞的表型发现其表达cd117、cd45ra和cd26,但是cd62l-(图12)。再克隆实验表明,这些细胞持续具有对所有ilc亚组的多能性。结论是,可以使用基质细胞和细胞因子的组合来扩增人ilcp,并使其保留功能特性。
参考文献
artis,d.,andspits,h.(2015).thebiologyofinnatelymphoidcells.nature517,293-301.
baum,c.m.,weissman,i.l.,tsukamoto,a.s.,buckle,a.m.,andpeault,b.(1992).isolationofacandidatehumanhematopoieticstem-cellpopulation.procnatlacadsciusa89,2804-2808.
bernink,j.h.,krabbendam,l.,germar,k.,dejong,e.,gronke,k.,kofoed-nielsen,m.,munneke,j.m.,hazenberg,m.d.,villaudy,j.,buskens,c.j.,etal.(2015).interleukin-12and-23controlplasticityofcd127(+)group1andgroup3innatelymphoidcellsintheintestinallaminapropria.immunity43,146-160.
bernink,j.h.,peters,c.p.,munneke,m.,tevelde,a.a.,meijer,s.l.,weijer,k.,hreggvidsdottir,h.s.,heinsbroek,s.e.,legrand,n.,buskens,c.j.,etal.(2013).humantype1innatelymphoidcellsaccumulateininflamedmucosaltissues.natimmunol14,221-229.
cella,m.,fuchs,a.,vermi,w.,facchetti,f.,otero,k.,lennerz,j.k.,doherty,j.m.,mills,j.c.,andcolonna,m.(2009).ahumannaturalkillercellsubsetprovidesaninnatesourceofil-22formucosalimmunity.nature457,722-725.
cella,m.,otero,k.,andcolonna,m.(2010).expansionofhumannk-22cellswithil-7,il-2,andil-1betarevealsintrinsicfunctionalplasticity.procnatlacadsciusa107,10961-10966.
chea,s.,perchet,t.,petit,m.,verrier,t.,guy-grand,d.,banchi,e.g.,vosshenrich,c.a.,disanto,j.p.,cumano,a.,andgolub,r.(2016a).notchsignalingingroup3innatelymphoidcellsmodulatestheirplasticity.scisignal9,ra45.
chea,s.,schmutz,s.,berthault,c.,perchet,t.,petit,m.,burlen-defranoux,o.,goldrath,a.w.,rodewald,h.r.,cumano,a.,andgolub,r.(2016b).single-cellgeneexpressionanalysesrevealheterogeneousresponsivenessoffetalinnatelymphoidprogenitorstonotchsignaling.cellrep14,1500-1516.
cherrier,m.,sawa,s.,andeberl,g.(2012).notch,id2,androrγtsequentiallyorchestratethefetaldevelopmentoflymphoidtissueinducercells.jexpmed209,729-740.
constantinides,m.g.,mcdonald,b.d.,verhoef,p.a.,andbendelac,a.(2014).acommittedprecursortoinnatelymphoidcells.nature508,397-401.
cupedo,t.,crellin,n.k.,papazian,n.,rombouts,e.j.,weijer,k.,grogan,j.l.,fibbe,w.e.,cornelissen,j.j.,andspits,h.(2009).humanfetallymphoidtissue-inducercellsareinterleukin17-producingprecursorstororc+cd127+naturalkiller-likecells.natimmunol10,66-74.
dallas,m.h.,varnum-finney,b.,delaney,c.,kato,k.,andbernstein,i.d.(2005).densityofthenotchliganddelta1determinesgenerationofbandtcellprecursorsfromhematopoieticstemcells.jexpmed201,1361-1366.
diefenbach,a.,colonna,m.,andkoyasu,s.(2014).development,differentiation,anddiversityofinnatelymphoidcells.immunity41,354-365.
eberl,g.,colonna,m.,disanto,j.p.,andmckenzie,a.n.(2015).innatelymphoidcells.innatelymphoidcells:anewparadigminimmunology.science348,aaa6566.
fuchs,a.,vermi,w.,lee,j.s.,lonardi,s.,gilfillan,s.,newberry,r.d.,cella,m.,andcolonna,m.(2013).intraepithelialtype1innatelymphoidcellsareauniquesubsetofil-12-andil-15-responsiveifn-γ-producingcells.immunity38,769-781.
gasteiger,g.,fan,x.,dikiy,s.,lee,s.y.,andrudensky,a.y.(2015).tissueresidencyofinnatelymphoidcellsinlymphoidandnonlymphoidorgans.science350,981-985.
hazenberg,m.d.,andspits,h.(2014).humaninnatelymphoidcells.blood124,700-709.
hoorweg,k.,peters,c.p.,cornelissen,f.,aparicio-domingo,p.,papazian,n.,kazemier,g.,
ikuta,k.,andweissman,i.l.(1992).evidencethathematopoieticstemcellsexpressmousec-kitbutdonotdependonsteelfactorfortheirgeneration.procnatlacadsciusa89,1502-1506.
juelke,k.,andromagnani,c.(2016).differentiationofhumaninnatelymphoidcells(ilcs).curropinimmunol38,75-85.
kikushige,y.,yoshimoto,g.,miyamoto,t.,iino,t.,mori,y.,iwasaki,h.,niiro,h.,takenaka,k.,nagafuji,k.,harada,m.,etal.(2008).humanflt3isexpressedatthehematopoieticstemcellandthegranulocyte/macrophageprogenitorstagestomaintaincellsurvival.jimmunol180,7358-7367.
kim,b.s.,andartis,d.(2015).group2innatelymphoidcellsinhealthanddisease.coldspringharbperspectbiol7.
klose,c.s.,flach,m.,
koues,o.i.,collins,p.l.,cella,m.,robinette,m.l.,porter,s.i.,pyfrom,s.c.,payton,j.e.,colonna,m.,andoltz,e.m.(2016).distinctgeneregulatorypathwaysforhumaninnateversusadaptivelymphoidcells.cell165,1134-1146.
langmead,b.,andsalzberg,s.l.(2012).fastgapped-readalignmentwithbowtie2.natmethods9,357-359.
legrand,n.,huntington,n.d.,nagasawa,m.,bakker,a.q.,schotte,r.,strick-marchand,h.,degeus,s.j.,pouw,s.m.,
lim,a.i.,menegatti,s.,bustamante,j.,lebourhis,l.,allez,m.,rogge,l.,casanova,j.l.,yssel,h.,anddisanto,j.p.(2016).il-12drivesfunctionalplasticityofhumangroup2innatelymphoidcells.jexpmed213,569-583.
luci,c.,reynders,a.,ivanov,i.i.,cognet,c.,chiche,l.,chasson,l.,hardwigsen,j.,anguiano,e.,banchereau,j.,chaussabel,d.,etal.(2009).influenceofthetranscriptionfactorrorgammatonthedevelopmentofnkp46+cellpopulationsingutandskin.natimmunol10,75-82.
mohtashami,m.,shah,d.k.,nakase,h.,kianizad,k.,petrie,h.t.,and
montaldo,e.,juelke,k.,andromagnani,c.(2015).group3innatelymphoidcells(ilc3s):origin,differentiation,andplasticityinhumansandmice.eurjimmunol45,2171-2182.
montaldo,e.,teixeira-alves,l.g.,glatzer,t.,durek,p.,stervbo,u.,hamann,w.,babic,m.,paclik,d.,
munneke,j.m.,
okada,s.,markle,j.g.,deenick,e.k.,mele,f.,averbuch,d.,lagos,m.,alzahrani,m.,al-muhsen,s.,halwani,r.,ma,c.s.,etal.(2015).immunodeficiencies.impairmentofimmunitytocandidaandmycobacteriuminhumanswithbi-allelicrorcmutations.science349,606-613.
rollini,p.,faes-van'thull,e.,kaiser,s.,kapp,u.,andleyvraz,s.(2007).phenotypicandfunctionalanalysisofhumanfetalliverhematopoieticstemcellsinculture.stemcellsdev16,281-296.
sawa,s.,cherrier,m.,lochner,m.,satoh-takayama,n.,fehling,h.j.,langa,f.,disanto,j.p.,andeberl,g.(2010).lineagerelationshipanalysisofrorgammat+innatelymphoidcells.science330,665-669.
schmidl,c.,rendeiro,a.f.,sheffield,n.c.,andbock,c.(2015).chipmentation:fast,robust,low-inputchip-seqforhistonesandtranscriptionfactors.natmethods12,963-965.
scoville,s.d.,mundy-bosse,b.l.,zhang,m.h.,chen,l.,zhang,x.,keller,k.a.,hughes,t.,cheng,s.,bergin,s.m.,mao,h.c.,etal.(2016).aprogenitorcellexpressingtranscriptionfactorrorγtgeneratesallhumaninnatelymphoidcellsubsets.immunity44,1140-1150.
seehus,c.r.,aliahmad,p.,delatorre,b.,iliev,i.d.,spurka,l.,funari,v.a.,andkaye,j.(2015).thedevelopmentofinnatelymphoidcellsrequirestox-dependentgenerationofacommoninnatelymphoidcellprogenitor.natimmunol16,599-608.
serafini,n.,vosshenrich,c.a.,anddisanto,j.p.(2015).transcriptionalregulationofinnatelymphoidcellfate.natrevimmunol15,415-428.
shih,h.y.,sciumè,g.,mikami,y.,guo,l.,sun,h.w.,brooks,s.r.,urban,j.f.,davis,f.p.,kanno,y.,ando'shea,j.j.(2016).developmentalacquisitionofregulomesunderliesinnatelymphoidcellfunctionality.cell165,1120-1133.
shultz,l.d.,brehm,m.a.,garcia-martinez,j.v.,andgreiner,d.l.(2012).humanizedmiceforimmunesysteminvestigation:progress,promiseandchallenges.natrevimmunol12,786-798.
spits,h.,artis,d.,colonna,m.,diefenbach,a.,disanto,j.p.,eberl,g.,koyasu,s.,locksley,r.m.,mckenzie,a.n.,mebius,r.e.,etal.(2013).innatelymphoidcells--aproposalforuniformnomenclature.natrevimmunol13,145-149.
vély,f.,barlogis,v.,vallentin,b.,neven,b.,piperoglou,c.,ebbo,m.,perchet,t.,petit,m.,yessaad,n.,touzot,f.,etal.(2016).evidenceofinnatelymphoidcellredundancyinhumans.natimmunol17,1291-1299.
yagi,r.,zhong,c.,northrup,d.l.,yu,f.,bouladoux,n.,spencer,s.,hu,g.,barron,l.,sharma,s.,nakayama,t.,etal.(2014).thetranscriptionfactorgata3iscriticalforthedevelopmentofallil-7rα-expressinginnatelymphoidcells.immunity40,378-388.
yang,q.,li,f.,harly,c.,xing,s.,ye,l.,xia,x.,wang,h.,wang,x.,yu,s.,zhou,x.,etal.(2015).tcf-1upregulationidentifiesearlyinnatelymphoidprogenitorsinthebonemarrow.natimmunol16,1044-1050.
zook,e.c.,ramirez,k.,guo,x.,vandervoort,g.,sigvardsson,m.,svensson,e.c.,fu,y.x.,andkee,b.l.(2016).theets1transcriptionfactorisrequiredforthedevelopmentandcytokine-inducedexpansionofilc2.jexpmed213,687-696.
- 还没有人留言评论。精彩留言会获得点赞!