一种金鱼Tgf2转座酶及其制备与保存方法与流程
本发明属于生物技术领域,涉及具体地说,是一种表达条件温和及高活性的金鱼tgf2转座酶的制备方法与保存方法。
背景技术
转座子是一类可以在基因组中发生位置移动的一段dna序列,被称为“跳跃因子”,转座子的应用在鱼类遗传育种、转基因、生物基因组的基因修饰等方面发挥着重要的作用。目前,国内外已经开展了大量关于dna转座子的研究,结果表明具有自主转座活性的转座子所编码的转座酶不仅能够提高转座效率而且可以促进养殖鱼类的转基因以及启动子捕获研究。tol2和tgf2被称为dna转座子中的“双鱼座”,其中tol2是日本学者koga等在白化的青鳉中发现的第一类具有自主转座活性的转座子,金鱼tgf2是继青鳉之后的第二类具有自主转座活性的脊椎动物转座子,它们均属于hat(hobo-ac-tam3)超家族转座子,金鱼tgf2转座酶是金鱼tgf2转座子所编码的活性转座酶。但有关金鱼tgf2密码子优化表达及其保存方法的研究还未见报道。
金鱼tgf2转座酶(tgf2-tpase)编码区全长2061bp,其序列包括4个开放阅读框(ofr),同时具有中间反向重复序列和末端反向重复序列等与转座相关的结构域。已经证明,用tgf2-tpase蛋白来代替tgf2-tpasemrna,与供体质粒共同直接注射到胚胎中有助于提高养殖鱼类中的插入效率。之前的研究表明在大肠杆菌rosetta1(de3)菌株中生产重组tgf2-tpase蛋白需要低温(22℃)、生长早期(od600=0.3-0.4)的这种严苛的发酵条件。菌种在生长早期易染杂菌,不利于大量发酵。本发明中,将重组蛋白的发酵温度提高到了30℃,od600提高到了0.5-0.6,使其发酵条件变得温和,同时不易染杂菌。
技术实现要素:
tgf2-tpase中存在一些稀有密码子,必要地优化表达不仅可以提高生产条件,而且有可能还会增加tgf2-tpase活性。
本发明涉及一种表达条件温和及高活性的金鱼tgf2转座酶的制备及保存方法,根据大肠杆菌对密码子的简并性和偏好性对tgf2-tpase中83个稀有密码子进行同义替换,并命名为tgf2-tpase83,在bl21(de3)中进行表达。经过histraptmhp亲和柱纯化后用maldi-(tof)/tof串联质谱进行鉴定。比较分析了-80℃下冻干酶粉、20%甘油、8%蔗糖和4%甘露醇四种不同保存方法的dnase酶活性,并用体积排阻色谱法鉴定了tgf2-tpase83dna结合活性及其特异性。结果显示:(1)在相同的原核表达系统中,可溶性的重组tgf2-tpase83在30℃,od600=0.5–0.6,1mm的iptg诱导6h时可大量表达;(2)经过亲和层析纯化后,与tgf2-tpase比较,优化的tgf2-tpase83有较高的酶活性;(3)对比四种不同的保存方法发现,将tgf2-tpase83保存在终浓度为20%的甘油中有助于保持其dnase消化活性;(4)体积排阻色谱结果表明纯化的tgf2-tpase83能够识别并特异性地结合含有tgf2转座子末端反向重复(tir)和亚末端重复(str)序列的dna探针。通过优化tgf2转座酶中的83个密码子不仅使其发酵条件变得温和,而且酶活性也得到了提高,可溶性的活性tgf2-tpase83蛋白生产为鱼类转基因提供了一种选择性的工具。
本发明的第一个目的是针对稀有密码子难以表达的不足,提供一种发酵条件温和的表达方法。
本发明的第二个目的是针对表达出来的蛋白杂蛋白多的不足,提供一种纯化及其鉴定的方法。
本发明的第三个目的是针对金鱼tgf2转座酶保存方法中的不足,提供了一种可保存其活性的方案及检测方法。
本发明的第四个目的是针对的体外dna结合活性不足,提供了一种鉴别金鱼tgf2转座酶体外dna结合活性和特异性的方法。
为实现上述第一个目的,本发明采取的技术方案是:
一种发酵条件温和的tgf2-tpase83的表达方法,包括如下步骤:
(1)根据编码金鱼tgf2转座酶的cdna序列(jn886586),参考大肠杆菌对密码子的简并性和偏好性对其中的83个密码子进行优化,命名为tgf2-tpase83;
(2)将优化后tgf2基因序列克隆至pet28a(+)载体,由大肠杆菌dh5α克隆,进而转化至表达菌bl21(de3)中扩大培养;
(3)表达菌在bl21(de3)中扩大培养至od600=0.5-0.6时加入1mm的iptg,30℃诱导6h。
为实现第二个目的,本发明采取的技术方案是:
一种tgf2-tpase83的纯化方法,包括如下步骤:
(1)表达后的tgf2-tpase83在37℃、-80℃下反复冻融三次;
(2)菌体超声破碎15min(功率100w,破碎3s,间隔6s),破碎三次,共45min;
(3)离心(12000g,20min,4℃),取上清,用0.22μm膜过滤,得到的上清液用histraptmhp亲和柱纯化;
(4)纯化后的tgf2-tpase83用maldi-(tof)/tof串联质谱进行鉴定。
为实现第三个目的,本发明采取的技术方案是:
一种保存tgf2-tpase83活性的方案及其检测方法,包括如下步骤:
对经过histraptmhp亲和柱纯化后的tgf2-tpase83做如下处理:
(1)直接冷冻干燥;
(2)加入8%蔗糖,进行冷冻干燥;
(3)加入4%甘露醇,进行冷冻干燥;
(4)加入20%乙醇;
将上述处理后的tgf2-tpase83在-80℃下保存。
(5)不同保存方法下的tgf2-tpase83与质粒ptgf2-ef1-egfp混合溶于反应缓冲液中,置于30℃中分别孵育0h、1h、2h、3h、4h、5h,加入终止缓冲液终止反应,最终的反应液用1.2%琼脂糖凝胶电泳检测。tgf2-tpase与质粒ptgf2-ef1-egfp的反应作为对照。
为实现第四个目的,本发明采取的技术方案是:
一种鉴别tgf2-tpase83体外dna结合活性和特异性的方法,包括如下步骤:
(1)在金鱼tgf2转座子左臂末端设计了4种含有不同重复序列数目的探针;
(2)纯化后的tgf2-tpase83与各探针在室温下共同孵育20min,用葡聚糖凝胶层析检测其dna结合活性,单独孵育的tgf2-tpase83作为对照;
(3)蛋清溶菌酶与探针在室温下共同孵育20min,用葡聚糖凝胶层析检测其dna结合特异性,单独孵育的蛋清溶菌酶作为对照。
较为具体地,本发明第一方面提供了一种金鱼tgf2转座酶,所述金鱼tgf2转座酶的核酸编码序列如seqidno.6所示。
本发明第二方面提供了一种金鱼tgf2转座酶的制备方法,包括如下步骤:
(1)金鱼tgf2转座酶密码子的优化,根据表达宿主对密码子的偏爱性和简并性把金鱼tgf2转座酶中稀有密码子进行同义替换;
(2)重组蛋白的表达;
(3)重组蛋白的纯化。
在一些实施方案中,所述宿主是大肠杆菌。
在一些实施方案中,根据大肠杆菌对密码子的偏爱性和简并性把金鱼tgf2转座酶中30-100个,优选50-90个,更优选83个稀有密码子进行同义替换。
在一些实施方案中,金鱼tgf2转座酶的编码序列参照genebank登录号:jn886586。
在一些实施方案中,所述优化后的金鱼tgf2转座酶的核酸编码序列如seqidno.6所示。
在一些实施方案中,在菌体生长至od600=0.5-0.6时加入1mm的iptg在30℃下诱导6h。
在一些实施方案中,通过亲和柱中的ni2+对重组蛋白中的his标签进行特异性结合纯化所述金鱼tgf2转座酶。
本发明第三方面提供了一种金鱼tgf2转座酶的保存方法,所述金鱼tgf2转座酶是如本发明第一方面所述的金鱼tgf2转座酶或本发明第二方面所述的方法制备的金鱼tgf2转座酶,对所述金鱼tgf2转座酶冷冻干燥。
在一些实施方案中,在所述冷冻干燥过程中加入保护剂,所述保护剂选自蔗糖、甘露醇、乙醇。
附图说明
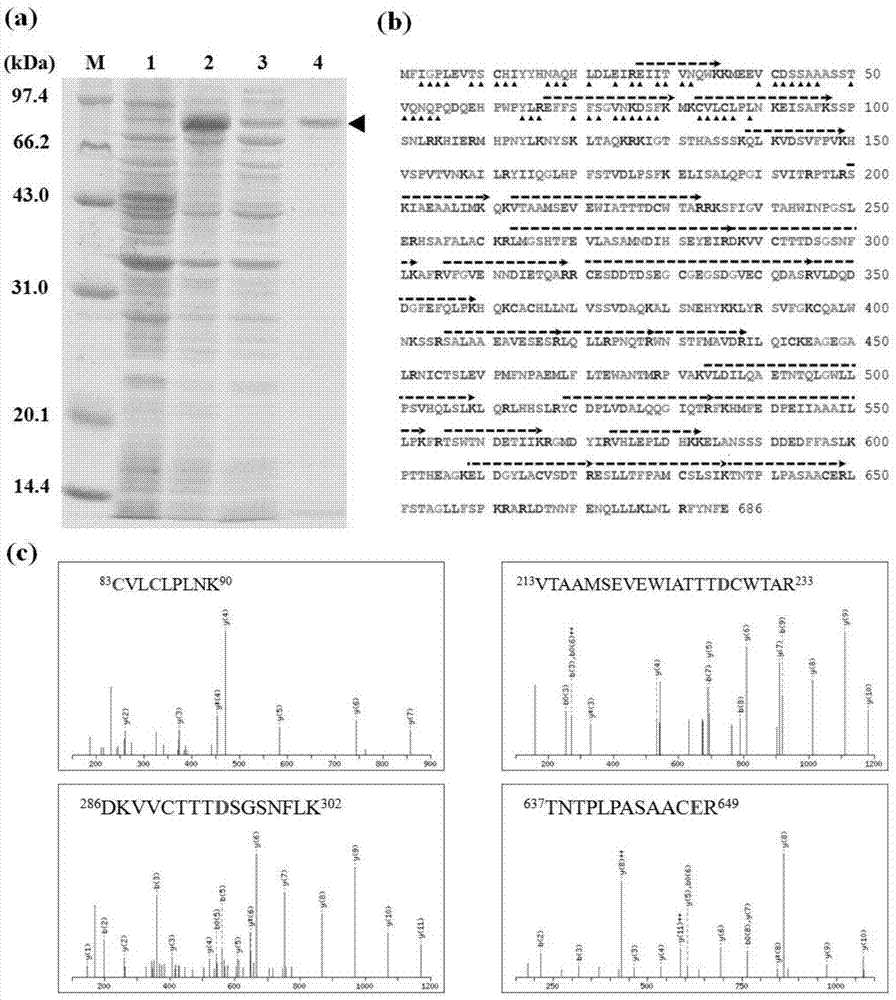
图1示出了密码子优化后的重组蛋白tgf2-tpase83的纯化和鉴定。
其中,(a)tgf2-tpase83的sds-page分析。m,标准蛋白分子量;1,未加诱导剂iptg;2,加诱导剂iptg;3,超声破碎后的上清液;4,ni2+亲和柱纯化后的洗脱峰。(b-c)tgf2-tpase83的maldi-tof/tof串联质谱鉴定。(b)tgf2-tpase83的氨基酸序列。黑色虚线,通过ms/ms检测到的氨基酸序列。箭头,优化的氨基酸。(c)部分多肽片段的质谱。c,d,e分别是dna结合域和催化域。
图2示出了不同保存条件下的tgf2-tpase83dnase活性分析。
其中,125ngptgf2-ef1-egfp质粒与6ng酶溶液在30℃孵育不同的时间,反应后的产物用琼脂糖凝胶电泳检测。1泳道均为标准蛋白分子量。oc:开环产物;l:线性体;sc:超螺旋体。(a)新鲜的tgf2-tpase酶液;(b)新鲜的tgf2-tpase83酶液;(c-f):20%甘油保存(c),8%蔗糖(d),直接冻干(e),4%甘露醇(f)。
图3示出了四种不同的探针在tgf2转座子上的位置。
其中,ph:高重复序列(含有6个重复序列);pm1,pm2:中重复序列(含有3个重复序列);pl:低重复序列(无重复序列)。
图4示出了体积排阻色谱法的tgf2-tpase83dna结合活性和特异性分析。
其中,(a,f)双链dna探针ph的洗脱曲线;(b)tgf2-tpase83单独孵育洗脱曲线;(c)tgf2-tpase83与ph混合孵育后的洗脱曲线;(d)tgf2-tpase83和pm1,pm2混合孵育后的洗脱曲线;(e)tgf2-tpase83与pl混合孵育后的洗脱曲线;(g)蛋清溶菌酶单独孵育后的洗脱曲线;(h)蛋清溶菌酶与ph混合孵育后的洗脱曲线。根据出峰时间,峰i为tgf2-tpase83,峰ii为探针ph,峰iii为蛋清溶菌酶。
图5示出了体积排阻色谱法的tgf2-tpasedna结合活性和特异性分析。
其中,(a)tgf2-tpase单独孵育洗脱曲线;(b)tgf2-tpase与ph混合孵育后的洗脱曲线;(c)tgf2-tpase和pm1,pm2混合孵育后的洗脱曲线;(d)tgf2-tpase与pl混合孵育后的洗脱曲线。根据出峰时间,峰i为tgf2-tpase,峰ii为探针ph。
图6示出了tgf2-tpase的sds-page分析结果。
其中:m,标准蛋白分子量;1,未加诱导剂iptg;2,加诱导剂iptg;3,ni2+亲和柱纯化后的洗脱峰。
具体实施方式
为了更好的解释本发明的技术方案,下面结合附图详细介绍本发明的实施例。以下实施例用于进一步说明本发明,但不应理解为对本发明的固定或限制。若未特别指明,实施例中所用的技术特征可以替换为具有在不背离发明构思前提下等同或相似功能或效果的其他本领域已知的技术特征。
1材料和方法
1.1材料
材料:质粒pet28a(+)、大肠杆菌dh5α、bl21(de3),均由上海海洋大学保存。异丙基-β-d-硫代半乳糖苷(iptg)、硫酸卡那霉素、甘油、蔗糖、甘露醇等试剂均购于国药集团化学试剂有限公司。
从tgf2(genebank登录号:hm146132,4720bp)(如图3)左臂末端设计了四种50bp探针进行dna结合分析。ph
(5’-cagaggtgtaaaagtacttaagtaattttacttgattactgtacttaagt-3’,
5’-acttaagtacagtaatcaagtaaaattacttaagtacttttacacctctg-3’),两条链互补,其第一条链的序列如seqidno.1所示;
pm1(5’-tttttactttacttgagtacaattaaaaatcaatacttttatttttactt-3’,5’aagtaaaaataaaagtattgatttttaattgtactcaagtaaagtaaaaa-3’),两条链互补,其第一条链的序列如seqidno.2所示;
pm2(5’-tagaaaaaaagtactttttactccttacaattttatttacagtcaaaaag-3’,5’ctttttgactgtaaataaaattgtaaggagtaaaaagtactttttttcta-3’),两条链互补,其第一条链的序列如seqidno.3所示;
pl(5’-ccttgctattaccaaaccaattgaattgcgctgatgcccagtttaattta-3’,5’taaattaaactgggcatcagcgcaattcaattggtttggtaatagcaagg-3’),两条链互补,其第一条链的序列如seqidno.4所示。
上述四种探针在使用中是双链状态,上面seqidno.1-4显示的是该基因编码链对应的核酸序列。
1.2tgf2-tpase密码子优化
tgf2是转座子,tgf2-tpase是tgf2编码的转座酶。活性的转座酶可以识别相应的转座子两端的特异性序列,能把转座子原位置中脱离出来,再插入到新的靶位点上,实现转座子的转座。
tgf2-tpase基因(genebank登录号:jn886586,2061bp)编码686个氨基酸,包含有四个开放阅读框。经过金瑞斯稀有密码子分析工具(https://www.genscript.com/tools/rare-codon-analysis),我们根据大肠杆菌对密码子的简并性和偏好性,对83个稀有密码子进行同义替换。新的cdna命名为tgf2-tpase83,优化后的密码子适应指数为0.86,gc含量为52.96%,密码子分布频率为0。利用稀有密码子分析软件genscriptrarecodonanalysistool对本实验中tgf2转座酶的686个氨基酸的密码子进行分析,其中有83个稀有密码子。利用大肠杆菌对密码子的简并性和偏好性,对这83个稀有密码子同义替换,从而使tgf2转座酶能够在大肠杆菌bl21(de3)中易表达。优化前的tgf2-tpase编码序列如seqidno.5所示;优化后的tgf2-tpase83编码序列如seqidno.6所示。
1.3tgf2-tpase和tgf2-tpase83原核表达
分别将tgf2-tpase、tgf2-tpase83基因序列克隆至pet28a(+)中,重组的质粒pet28a(+)-tgf2-tpase和pet28a(+)-tgf2-tpase83转化到bl21(de3)感受态细胞中。阳性的转化菌株进行pcr检测并测序。测序成功的bl21(de3)-pet28a(+)-tgf2-tpase和bl21(de3)-pet28a(+)-tgf2-tpase83在30℃、od600=0.5-0.6条件下加入1.0mmiptg诱导6h。诱导后的菌液离心(12000g,15min,4℃)收集并通过sds-page凝胶进行分析。
1.4tgf2-tpase和tgf2-tpase83纯化
表达后的重组蛋白超声破碎45min(功率100w,破碎3s,间隔6s)后离心(12000g,20min)取上清,0.22μm膜过滤。得到的上清液用histraptmhp亲和柱纯化。平衡缓冲液:20mmol/l磷酸盐、0.5mol/lnacl、20mmol/l咪唑、ph7.4;结合缓冲液:20mmol/l磷酸盐、0.5mol/lnacl、40mmol/l咪唑、ph7.4;洗脱缓冲液:20mmol/l磷酸盐、0.5mol/lnacl、500mmol/l咪唑、ph7.4。根据紫外吸收值(od280),收集目的蛋白组分进行sds-page检测。
1.5tgf2-tpase83鉴定
从sds-page凝胶上选择合适的条带切割,用abi4800plusmaldi-(tof)/tof串联质谱进行鉴定。maldi-(tof)/tof串联质谱是通过随机打断氨基酸序列,根据其飞行时间和质量间的关系对重组tgf2-tpase83转座酶蛋白的氨基酸片段序列进行鉴定。
maldi-(tof)/tof串联质谱的基本流程:向样品中加入水重复清洗两次后将水移除,加入胶内消化脱色液直至胶粒完全变干,加入dtt在56℃中孵育1h,后移除多余的液体,加入iam,暗室孵育45min,后移除多余的液体,加入nh4hco3重复清洗后移除,加入脱色液进行脱色。加入胰蛋白酶在37℃中消化过夜,加入0.1%的fa终止消化,取样上机,使用质谱仪检测,从而得到检测结果。
1.6tgf2-tpase83dnase活性测定
125ngptgf2-ef1α-egfp质粒中(该质粒参见文献:吴芳,叶星,邹曙明,等.tgf2转座系统在转rfp基因斑马鱼上的应用[j].中国水产科学,2014,21(4):647-654)加入6ng复溶后的酶液,混合溶于20μl反应缓冲液(25mmhepes、50mmnacl、1mmmncl2、2mmdtt、2%甘油、0.1mg/mlbsa,ph为7.6)中,置于30℃中分别孵育0h、1h、2h、3h、4h、5h,加入终止缓冲液(1%sds、20mmedta)终止反应,最终的反应液用1.2%琼脂糖凝胶电泳检测。
1.7不同保存方法对tgf2-tpase83活性的影响
多元醇及糖类能够有效地提高酶蛋白的稳定性,故采用以下4种不同的保存方法来研究重组tgf2-tpase83转座酶酶活性的变化情况,以新鲜酶液的活性作为对照。(1)直接将新鲜的酶液进行冷冻干燥;(2)向酶液中加入20%甘油;(3)向酶液中加入4%甘露醇,进行冷冻干燥;(4)向酶液中加入8%蔗糖,进行冷冻干燥。将经过上述不同方法处理的样品保存于-80℃。
1.8tgf2-tpase和tgf2-tpase83dna结合活性和特异性分析
tgf2的转座过程主要包含四个阶段:(1)转座酶通过二聚或寡聚化配对;(2)转座酶与转座子的末端序列特异性结合;(3)在转座子的末端反向重复序列处切割dna链;(4)捕获新的基因与靶dna进行整合。tgf2转座的发生得益于转座酶的活性,转座酶的活性影响着转座效率的高低,体外dna结合活性是研究其体内转座效率的前提。
重组tgf2-tpase和tgf2-tpase83的dna结合活性采用superdex20010/300gl葡聚糖层析,利用分子排阻色谱法。我们从tgf2左臂中设计了四种不同的50bpdna双链探针。ph,含有6个重复序列,称为高重复序列;pm1,含有3个重复序列,称为中重复序列;pm2,含有3个重复序列,称为中重复序列;pl,没有重复序列,称为低重复序列;具体序列参见1.1材料部分的记载。
tgf2-tpase和tgf2-tpase83分别与探针ph,pm1,pm2和pl混合,25℃下孵育20min后上样。根据紫外吸收值(od280,od260)的变化,判断tgf2-tpase和tgf2-tpase83转座酶是否具有体外dna结合活性以及密码子优化前后dna结合活性的比较,以新鲜酶液为空白对照,以蛋清溶菌酶与探针的结合活性为阴性对照,判断其特异性。洗脱液为0.5mol/lnacl磷酸盐缓冲液,流速为0.5ml/min。
2结果
2.1tgf2-tpase83表达、纯化和鉴定
重组蛋白tgf2-tpase83的诱导表达,离心的上清液和经过亲和层析后的纯化样品结果见图1a。sds-page的灰度扫描结果表明诱导后的重组蛋白占总蛋白的6.3%(图1a,2泳道),纯化后的重组蛋白纯度>85%(图1a,4泳道)。重组蛋白tgf2-tpase的诱导表达,离心的上清液和经过亲和层析后的纯化样品结果见图6。tgf2-tpasesds-page的灰度扫描结果表明诱导后的重组蛋白占总蛋白的4.7%(图6,2泳道),纯化后的重组蛋白纯度>70%(图6,3泳道)。二者结果相比,tgf2-tpase83的表达量和纯度得到了同时提高。
从sds-page胶上切割下来的重组蛋白进行maldi(tof)/tof串联质谱分析。用mascot进行ms/ms离子质谱后,至少有21个肽片段被检测出与推测的金鱼tgf2转座酶片段相重合(genebank登录号:afc96942.1),如图1b。三处寡肽片段的ms/ms如图1c所示,肽片段(83-90氨基酸残基)包含了位于n-末端bed锌指结构域的两个保守的半胱氨酸(c83和c86),其涉及到tgf2转座酶的dna结合。催化三联体,位于c-末端的rnase-h域中的dde的残基(d228,d295和e648),其在(213-233,286-302,637-649氨基酸残基)中都有被检测到。因此,可溶性的重组蛋白在bl21(de3)中成功表达,而且质谱结果显示重组表达的蛋白即为金鱼tgf2转座酶。
2.2tgf2-tpase83dnase活性
tgf2-tpase和tgf2-tpase83酶液与质粒ptgf2-ef1-egfp的dnase活性结果见图2。随着时间的增加,tgf2-tpase83对质粒的裂解在60min之内就可完成,将近100%的dna,其中包括开环、线性和超螺旋三种形式在40min之内被完全消解(图2b)。然而,tgf2-tpase对质粒的裂解在5h之内才完成,将近100%的三种形式的dna质粒完全被消解需要4h(图2a)。与tgf2-tpase相比,优化后的tgf2-tpase83有较高的酶活性。
用四种不同的方法对tgf2-tpase83进行保存,dnase结果如图2c-f所示。20%甘油保存时,将近100%的dna包括超螺旋和线性均在3h以内被消化,而开环形式的在4h之后才被消解(图2c)。与新鲜酶液相比,这种保存方法有助于tgf2-tpase83长时间的保存其活性。而其他三种保存方法(8%蔗糖,4%甘露醇和冻干酶粉)的dna消化均有一定程度的下降(图2d-f),与20%甘油相比,它们没有明显的dnase消化活性。所以,上述结果表明给重组蛋白中加入20%甘油在-80℃下保存有助于提高蛋白的稳定性。
2.3tgf2-tpase和tgf2-tpase83对tgf2转座子的dna结合活性和特异性
体积排阻色谱法又称葡聚糖凝胶层析法,其原理为:转座酶和dna结合后会使蛋白峰处的吸收值增加,从而判断转座酶是否有dna结合活性。与单独孵育的蛋白洗脱曲线相比较,若蛋白与dna共同孵育后,在相同的蛋白出峰位置处,若od280和od260的吸收值均有增加,且od260>od280,则说明有dna结合活性。同理,与单独孵育的蛋清溶菌酶洗脱曲线相比较,若蛋清溶菌酶与相同的dna共同孵育后,在相同的蛋白出峰位置处,od280和od260的吸收值均没有增加,则说明有dna结合活性是具有特异性的。
四种不同的探针ph,pm1,pm2,pl在tgf2转座子上的位置见图3。利用体积排阻色谱法对tgf2-tpase83体外的dna结合活性及其特异性研究结果见图4。核酸洗脱曲线(峰ii,~15min)od260比od280高(图4a,f),而tgf2-tpase83所呈现的蛋白峰(峰i,~9min)od260和od280相当(图4b)。
tgf2-tpase83和ph在25℃下孵育20min后用体积排阻色谱法洗脱,如图4c中峰i由蛋白形式转变到了核酸形式。当tgf2-tpase83分别与pm1,pm2和pl混合后,结果与ph类似(图4d,e)。结果表明tgf2-tpase83对tgf2左臂末端不同的重复序列具有dna结合活性。而且对于不同的探针,tgf2-tpase83也表现出了不同的dna结合能力。n-末端的tir和str对于转座酶的dna结合是必不可少的,而且重复序列多的比重复序列少的有更高的结合能力,可能是由于探针中含有更多的重复片段或者特殊的信息包含在其中(图3)。所以,n-末端中的重复序列越多,tgf2-tpase83的dna结合能力就越强。
为了研究金鱼tgf2-tpase83的特异性,用蛋清溶菌酶(~14.4kda)作为阴性对照。探针ph和蛋清溶菌酶按照文中的方法进行孵育。与图4f中的峰ii和图4g中的峰iii(~25min,od280>od260)相比,混合物的洗脱曲线中峰ii和峰iii并没有变化(图4h)。结果表明蛋清溶菌酶对探针ph并没有dna结合活性。所以,重组的tgf2-tpase83对tgf2左臂末端具有dna结合活性和特异性。
利用体积排阻色谱法对密码子优化之前tgf2-tpase体外的dna结合活性及其特异性研究结果见图5。
根据图5可以看出:tgf2-tpase所呈现的蛋白峰(峰i,~9min)od260和od280相当(图5a)。
图a为tgf2-tpase的洗脱图,出峰(峰i)时间~9min;图b-d为tgf2-tpase与不同探针共同孵育后的洗脱图。从图中可以看出,当tgf2-tpase与探针ph(图b)、pm1,pm2(图c)和pl(图d)共同孵育后,蛋白峰处的吸收值均有增加。结果表明tgf2-tpase对tgf2左臂末端不同的重复序列具有dna结合活性。
tgf2-tpase和ph在25℃下孵育20min后用体积排阻色谱法洗脱,如图5b中峰i由蛋白形式转变到了核酸形式。当tgf2-tpase分别与pm1,pm2和pl混合后,结果与ph类似(图5c,d)。结果表明tgf2-tpase对tgf2左臂末端不同的重复序列也具有dna结合活性。
比较图4和图5可以看出:tgf2-tpase和tgf2-tpase83对不同重复序列的探针均具有dna结合活性。比较图4c(峰ⅰ)和图5b(峰ⅰ)可以看出,tgf2-tpase83与探针结合后的od吸收值比tgf2-tpase与探针结合后的od吸收值高,且tgf2-tpase和tgf2-tpase83与探针pm1,pm2和pl的dna结合活性比较结果与ph相似,故密码子优化后的金鱼tgf2转座酶比优化前的酶活性高。利用此结果,可以对金鱼tgf2转座子末端序列通过基因敲除或基因编辑等方法进行改造,从而可以构建更高活性的转座子系统。
比较图4和图5可以看出,图4中tgf2-tpase83与各探针共同孵育后蛋白峰处的od吸收值(od280、od260)增加程度均比tgf2-tpase与各探针共同孵育后蛋白峰处的od吸收值(od280、od260)增加程度高,所以密码子优化后的金鱼tgf2转座酶比优化前的酶活性高。
以上各个实施例只是用于进一步说明本发明,并不是用来限制本发明的保护范围,凡是基于本发明的构思所作出的等同变换及对本发明的各个技术方案显而易见的改进,均落入本发明的保护范围。
序列表
<110>上海海洋大学
<120>一种金鱼tgf2转座酶及其制备与保存方法
<141>2018-07-05
<160>6
<170>siposequencelisting1.0
<210>1
<211>50
<212>dna
<213>金鱼(carassiusauratus)
<400>1
cagaggtgtaaaagtacttaagtaattttacttgattactgtacttaagt50
<210>2
<211>50
<212>dna
<213>金鱼(carassiusauratus)
<400>2
tttttactttacttgagtacaattaaaaatcaatacttttatttttactt50
<210>3
<211>50
<212>dna
<213>金鱼(carassiusauratus)
<400>3
tagaaaaaaagtactttttactccttacaattttatttacagtcaaaaag50
<210>4
<211>50
<212>dna
<213>金鱼(carassiusauratus)
<400>4
ccttgctattaccaaaccaattgaattgcgctgatgcccagtttaattta50
<210>5
<211>2061
<212>dna
<213>金鱼(carassiusauratus)
<400>5
atgttcattggtcctttggaagtgacgtcatgtcacatctattaccacaatgcacagcac60
cttgacctggaaattagagaaattataacagtcaatcagtggaagaaaatggaggaagta120
tgtgattcatcagcagctgcgagcagcacagtccaaaatcagccacaggatcaagagcac180
ccgtggccgtatcttcgcgaattcttttcttttagtggtgtaaataaagattcattcaag240
atgaaatgtgtcctctgtctcccgcttaataaagaaatatcggccttcaaaagttcgcca300
tcaaacctaaggaagcatattgagagaatgcacccgaactacctgaaaaactactctaaa360
ctgaccgcacagaaacgcaaaaacggcacctcaacgcacgcctcatcgtccaagcagctg420
aaagttgactcagtctttccggtgaagcatgtttcgccggtcaccgtgaacaaagccatt480
ctgcgctatattatccagggtctgcacccgttttccaccgtggatctgccgtcattcaaa540
gaactgatctcgacgctgcaaccgggcattagcgttatcacccgtccgacgctgcgctct600
aagattgcagaagcggccctgatcatgaaacagaaggtcaccgcagctatgagtgaagtg660
gaatggattgcgaccacgaccgattgctggaccgcccgtcgcaaatcctttattggtgtg720
acggcgcattggatcaatccgggctctctggaacgccacagtgcttttgcgctggcctgt780
aaacgtctgatgggtagccataccttcgaagttctggcaagcgctatgaacgatattcac840
tctgaatacgaaatccgcgacaaagtggtttgcacgaccacggattctggtagtaatttc900
ctgaaggcctttcgtgtcttcggcgtggaaaacaatgacattgaaacccaggcacgtcgc960
tgtgaatccgatgacacggattcagaaggctgcggtgaaggcagtgatggtgttgaatgt1020
caggacgcttcccgcgtcctggatcaagatgacggctttgaattccagctgccgaaacat1080
caaaagtgcgcgtgtcacctgctgaacccggttagctctgtcgatgcgcagaaagccctg1140
agcaatgaacattataaaaagctgtaccgctctgtctttggcaaatgccaagcgctgtgg1200
aacaagagttcccgtagcgccctggccgcggaagcggtggaatccgaatcacgtctgcag1260
ctgctgcgcccgaaccaaacccgttggaattctacgttcatggccgtggaccgcattctg1320
cagatctgcaaagaagcaggtgaaggcgctctgcgtaacatttgtaccagcctggaagtt1380
ccgatgtttaatccggccgaaatgctgttcctgaccgaatgggcaaacacgatgcgtccg1440
gtggctaaagttctggatatcctgcaggcagaaaccaatacgcaactgggttggctgctg1500
ccgtcggttcatcagctgagcctgaaactgcaacgcctgcatcacagtctgcgttattgt1560
gatccgctggtcgacgccctgcagcaaggcattcagacccgctttaaacacatgttcgaa1620
gatccggaaattatcgcagctgcgatcctgctgccgaaatttcgtaccagctggacgaac1680
gacgaaaccattatcaagcgcggtatggattatattcgtgtgcatctggaaccgctggac1740
cacaaaaaggaactggcgaattcatcgagcgatgacgaagatttctttgcatcactgaaa1800
ccgaccacgcatgaagcttcgaaggaactggacggttacctggcgtgcgtttctgatacc1860
cgcgaaagtctgctgacgtttccggctatgtgttcgctgagcatcaaaaccaatacgccg1920
ctgccggccagcgcggcatgtgaacgtctgtttagtaccgcgggtctgctgttctccccg1980
gaacgtgcacgcctggataccaactcctttgaaaaccaactgctgctgaaactgaacctg2040
cgcttctacaactttgaatga2061
<210>6
<211>2061
<212>dna
<213>金鱼(carassiusauratus)
<400>6
atgttcatcggcccgctggaagtgaccagctgccatatttattaccacaacgcgcaacac60
ctggacctggagattcgtgagattattaccgtgaaccagtggaagaaaatggaggaagtt120
tgcgacagcagcgcggcggcgagcagcaccgtgcagaaccaaccgcaggatcaagagcac180
ccgtggccgtacctgcgtgaattctttagcttcagcggtgtgaacaaggacagctttaag240
atgaaatgcgttctgtgcctgccgctgaataaagaaatcagcgcgttcaaaagcagcccg300
agcaatctgcgtaagcacatcgagcgtatgcacccgaactacctgaaaaactactctaaa360
ctgaccgcacagaaacgcaaaaacggcacctcaacgcacgcctcatcgtccaagcagctg420
aaagttgactcagtctttccggtgaagcatgtttcgccggtcaccgtgaacaaagccatt480
ctgcgctatattatccagggtctgcacccgttttccaccgtggatctgccgtcattcaaa540
gaactgatctcgacgctgcaaccgggcattagcgttatcacccgtccgacgctgcgctct600
aagattgcagaagcggccctgatcatgaaacagaaggtcaccgcagctatgagtgaagtg660
gaatggattgcgaccacgaccgattgctggaccgcccgtcgcaaatcctttattggtgtg720
acggcgcattggatcaatccgggctctctggaacgccacagtgcttttgcgctggcctgt780
aaacgtctgatgggtagccataccttcgaagttctggcaagcgctatgaacgatattcac840
tctgaatacgaaatccgcgacaaagtggtttgcacgaccacggattctggtagtaatttc900
ctgaaggcctttcgtgtcttcggcgtggaaaacaatgacattgaaacccaggcacgtcgc960
tgtgaatccgatgacacggattcagaaggctgcggtgaaggcagtgatggtgttgaatgt1020
caggacgcttcccgcgtcctggatcaagatgacggctttgaattccagctgccgaaacat1080
caaaagtgcgcgtgtcacctgctgaacccggttagctctgtcgatgcgcagaaagccctg1140
agcaatgaacattataaaaagctgtaccgctctgtctttggcaaatgccaagcgctgtgg1200
aacaagagttcccgtagcgccctggccgcggaagcggtggaatccgaatcacgtctgcag1260
ctgctgcgcccgaaccaaacccgttggaattctacgttcatggccgtggaccgcattctg1320
cagatctgcaaagaagcaggtgaaggcgctctgcgtaacatttgtaccagcctggaagtt1380
ccgatgtttaatccggccgaaatgctgttcctgaccgaatgggcaaacacgatgcgtccg1440
gtggctaaagttctggatatcctgcaggcagaaaccaatacgcaactgggttggctgctg1500
ccgtcggttcatcagctgagcctgaaactgcaacgcctgcatcacagtctgcgttattgt1560
gatccgctggtcgacgccctgcagcaaggcattcagacccgctttaaacacatgttcgaa1620
gatccggaaattatcgcagctgcgatcctgctgccgaaatttcgtaccagctggacgaac1680
gacgaaaccattatcaagcgcggtatggattatattcgtgtgcatctggaaccgctggac1740
cacaaaaaggaactggcgaattcatcgagcgatgacgaagatttctttgcatcactgaaa1800
ccgaccacgcatgaagcttcgaaggaactggacggttacctggcgtgcgtttctgatacc1860
cgcgaaagtctgctgacgtttccggctatgtgttcgctgagcatcaaaaccaatacgccg1920
ctgccggccagcgcggcatgtgaacgtctgtttagtaccgcgggtctgctgttctccccg1980
gaacgtgcacgcctggataccaactcctttgaaaaccaactgctgctgaaactgaacctg2040
cgcttctacaactttgaatga2061
- 还没有人留言评论。精彩留言会获得点赞!