一种活细胞内原位加速的DNA马达及其制备方法和应用与流程
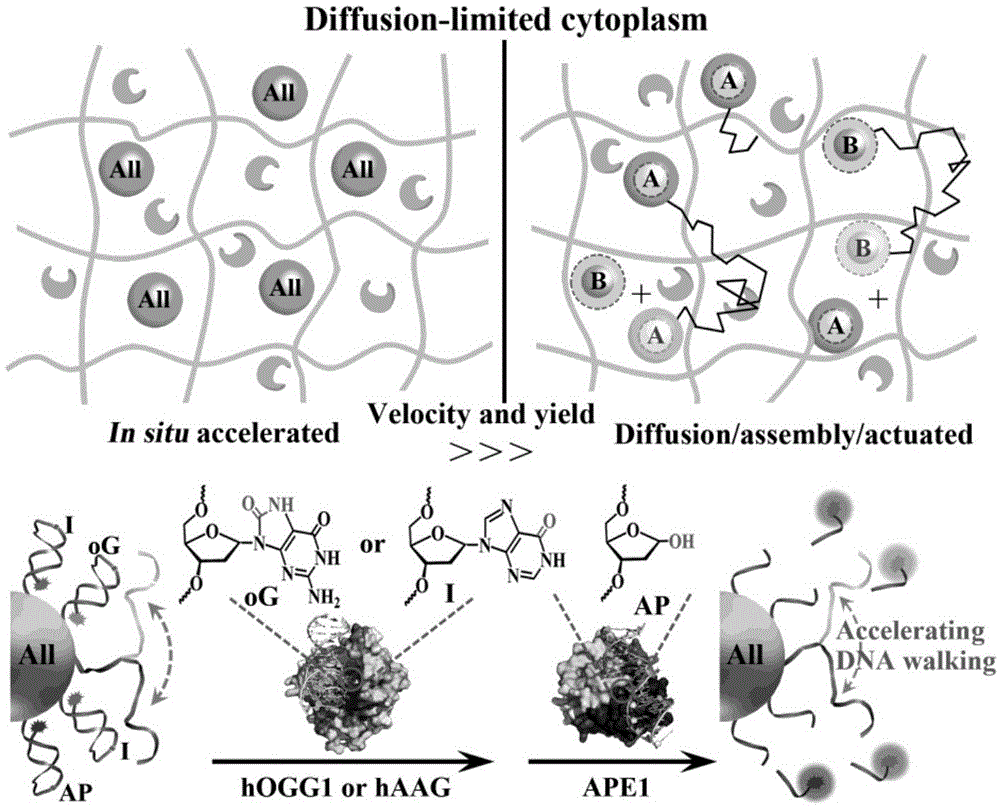
本发明属于活细胞成像技术领域,涉及一种活细胞内原位加速的dna马达及其制备方法和应用。
背景技术:
活细胞成像可实时观察细胞分子、结构和过程,对阐明分子作用机制、生物功能以及疾病发生发展规律具有重要意义。当前,荧光成像是主要的活细胞研究手段之一,结合dna信号放大技术,如链杂交反应、催化发夹组装等,已被广泛用于胞内蛋白质、酶和核酸等分子标志物以及相应生物过程的监测。上述成像方法主要基于一个三步反应的循环过程(dna杂交-切除或置换-游离释放),过程耗时,信号放大速率不足,灵敏度仍待提高。
另一种基于dna步移(walking)的活细胞成像方法,相比于经典的游离dna探针,步移探针可直接、快速结合界面荧光探针,信号放大效率更强,灵敏度更高。但是,它们依旧需要供给细胞额外的能量物质或核酸探针来介导信号放大反应,体系复杂,极易引起由试剂进胞效率不同导致的信号差异,干扰准确成像、产生误判。
同时,现有活细胞信号放大成像方法缺少内参信号,没有信号校准功能,抗胞内复杂环境干扰能力差,成像结果不准确,更不能研究细胞异质性,限制单细胞研究应用。
技术实现要素:
本发明的目的在于提供一种活细胞内原位加速的dna马达及其制备方法和应用,以克服现有技术中多模块结构胞浆内扩散限制导致灵敏度不足、反应动力学慢、缺少内参校准信号等缺点。
为了达到上述目的,本发明采用以下技术方案予以实现:
本发明公开的一种活细胞内原位加速的dna马达,以金纳米颗粒aunp作为载体,并在其上负载三种dna链;所述三种dna链分别为:
第一dna链,为发生运动过程的dw链,其3’端的巯基可共价连接金纳米颗粒aunp;
第二dna链,为带有化学标记的dt链,其为dna茎环结构,在该dna茎环结构的环状部分含有一个8-氧-7-氢脱氧鸟嘌呤og或次黄嘌呤i损伤碱基位点,茎环结构茎部3’端的巯基可共价连接金纳米颗粒aunp,5’端修饰荧光分子tamra或fam;
第三dna链,为带有化学标记的r链,其为dna茎环结构,在该dna茎环结构环状部分含有一个脱嘌呤/脱嘧啶ap位点,其茎部3’端的巯基可共价连接金纳米颗粒aunp,5’端修饰荧光分子cy5。
优选地,第一dna链即dw链的5’端的14个碱基与第二dna链即dt链的环状部分的14个碱基互补。
优选地,第一dna链即dw链的5’端的14个碱基与第二dna链即dt链的环状部分互补成双链dna;
其中,dt链上的损伤碱基位点og或i能被人类8-羟基鸟嘌呤dna糖基化酶hogg1或人类烷基腺嘌呤dna糖基化酶haag作用成为脱嘌呤/脱嘧啶ap位点,再由人源脱嘌呤/脱嘧啶ap核酸内切酶ape1酶切后,释放出荧光。
只有互补杂交成双链dna后,dt链上的损伤碱基位点og或i才能被人类8-羟基鸟嘌呤dna糖基化酶hogg1或人类烷基腺嘌呤dna糖基化酶haag作用成为脱嘌呤/脱嘧啶ap位点,单链上的损伤碱基位点不能被酶作用。
优选地,r链上的脱嘌呤/脱嘧啶ap位点,直接由人源脱嘌呤/脱嘧啶ap核酸内切酶ape1酶切后,释放出荧光。
本发明还公开了上述的活细胞内原位加速的dna马达的制备方法,包括以下步骤:
1)将待组装的dw链、dt链和r链分别加入终浓度为3mm的tcep溶液中,静置1h;
2)将步骤1)配制的含有dw链的tcep溶液加入金纳米颗粒aunp溶液中,振荡处理1h;
3)然后向步骤2)获得的含有dw链的金纳米颗粒aunp溶液中加入步骤1)制得的含有dt链的tcep溶液和含有r链的tcep溶液,室温孵育过夜;
4)向步骤3)制得的溶液体系中加入nacl缓冲液直至体系中nacl浓度为0.3m,室温孵育12h;反应结束后,反应溶液经离心、洗涤处理,制得活细胞内原位加速的dna马达,即负载三种dna链的金纳米颗粒aunp。
优选地,dw链、dt链和r链的摩尔比为1:9:1.5;dw链和金纳米颗粒aunp的摩尔比为30:1。
本发明还公开了上述的活细胞内原位加速的dna马达在活细胞成像监测中的应用。
本发明还公开了上述的活细胞内原位加速的dna马达作为活细胞內源酶驱动的自校准dna步移器的应用。
优选地,使用时,操作如下步骤:
1)将所述的活细胞内原位加速的dna马达导入活细胞内;
2)活细胞内原位加速的dna马达在内源酶的酶切作用下释放荧光;
3)通过激光共聚焦荧光显微镜进行观察,实现细胞内损伤碱基修复酶的原位成像和作用通路监测。
与现有技术相比,本发明具有以下有益效果:
本发明创新地设计并制备了活细胞内原位加速的dna马达,其以金纳米颗粒aunp作为载体,并在其上负载三种dna链,构成一体化结构,该一体化结构集成所有反应模块,能够避免各反应模块受到粘性胞浆扩散的限制,且活细胞内源损伤碱基修复酶驱动、不需要给细胞额外提供任何物质,能避免细胞差异和胞内复杂环境造成的干扰,实现高灵敏、动力学加速的活细胞成像及检测,从而更好的应用于发展活细胞內源酶驱动的自校准dna步移机器,实现单细胞低丰度靶标的高灵敏精准成像分析。具体优势如下:
1)本发明的活细胞内原位加速的dna马达在扩散运动受到限制的粘性活细胞胞浆内,反应速率明显高于传统组装反应模式(游离-扩散-组装-酶切),进一步提高检测及成像灵敏度;
2)本发明的活细胞内原位加速的dna马达在反应过程由细胞内源性酶作用,不需额外加入能量物质或dna链,反应体系简单,不受进胞效率差异等因素影响,提高成像准确度;
3)将自校准功能一体化地加入设计中,能够指示不同反应环境中工具酶的差异,避免细胞差异和胞内复杂环境造成的干扰,实现准确成像,并可用于细胞异质性及单细胞的研究应用;
4)本发明活细胞内原位加速的dna马达制备方法简便,细胞毒性低,通过与细胞进行温育,即可实现细胞内碱基损伤修复酶的原位荧光成像和相关作用通路的监测。
附图说明
图1为本发明的原理图。
图2为扩散限制条件下dna马达的作用机制与动力学;其中,a为利用sted(受激发射损耗)荧光显微镜对dna马达的单颗粒荧光分析结果;b为几种粘性扩散限制环境中实验研究对比两种dna马达的反应速率;c为从原理上对一体化设计提高局部反应浓度、加速反应的分析结果;
图3为原位加速dna马达的反应动力学研究;其中,a为不同dw密度对dna马达反应动力学的影响;b为一体化双轨道(双dt)dna马达同时对hogg1/ape1andhaag/ape1两条修复通路进行监测的信号变化;c为对自由扩散的dw链在两种金纳米颗粒(分别修饰og-dt和i-dt)之间的移动过程进行研究分析。
图4为一体化dna马达自校准内参的性能分析;
图5为一体化原位加速dna马达监测活细胞(hela)内dna损伤碱基修复通路;
图6为活细胞内(hela)自校准一体化dna马达对两种损伤碱基修复酶的监测成像图;其中,a为三种不同ape1活性的细胞样本中,利用dna马达对hogg1监测的结果;b为自校准双轨道(双dt)dna马达在同一细胞样本中,对hogg1和haag同时监测的成像结果。
具体实施方式
为了使本技术领域的人员更好地理解本发明方案,下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分的实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都应当属于本发明保护的范围。
需要说明的是,本发明的说明书和权利要求书及上述附图中的术语“第一”、“第二”等是用于区别类似的对象,而不必用于描述特定的顺序或先后次序。应该理解这样使用的数据在适当情况下可以互换,以便这里描述的本发明的实施例能够以除了在这里图示或描述的那些以外的顺序实施。此外,术语“包括”和“具有”以及他们的任何变形,意图在于覆盖不排他的包含,例如,包含了一系列步骤或单元的过程、方法、系统、产品或设备不必限于清楚地列出的那些步骤或单元,而是可包括没有清楚地列出的或对于这些过程、方法、产品或设备固有的其它步骤或单元。
下面结合附图对本发明做进一步详细描述:
参见图1,为本发明的原理图,图中比较了一体化原位加速dna马达和常规扩散-组装-反应的dna马达在扩散限制的活细胞胞浆内的反应,并具体描述了dna马达被dna损伤碱基修复酶作用的原理及化学过程。
通过扩散限制条件下dna马达的作用机制与动力学研究,说明本申请设计的dna马达的加速功能,结果参见图2。图2中,a为利用sted(受激发射损耗)荧光显微镜对dna马达的单颗粒荧光分析结果,展示了sted原理、一体化原位加速dna马达和常规扩散-组装-反应的dna马达不同的反应速率,证明本申请设计的dna马达的加速功能。图2中b为在几种粘性扩散限制环境中实验研究对比两种dna马达的反应速率,图2中c则是从原理上对一体化设计提高局部反应浓度、加速反应进行分析,说明了本发明一体化dna马达的优势。
图3为原位加速dna马达的反应动力学研究,其中a图是不同dw密度对dna马达反应动力学的影响,d代表距离,不同dw密度导致相邻dw距离发生变化,影响马达反应动力学。图中b为一体化双轨道(双dt)dna马达同时对hogg1/ape1andhaag/ape1两条修复通路进行监测的信号变化。图中c是对自由扩散的dw链在两种金纳米颗粒(分别修饰og-dt和i-dt)之间的移动过程进行研究分析。
图4为一体化dna马达自校准内参的性能分析,结果显示,内参信号cy5能够指示工具酶ape1的活性变化,为实际应用中的信号分析工作提供可靠的信息。
本发明公开的活细胞内原位加速的dna马达的方法,操作如下:
dw、dt、r链中分别加入终浓度3mm的tcep溶液(用来还原巯基,需要新鲜配制),室温下放置1小时;将dw链加入aunp溶液(摩尔比30:1),振荡1小时;再加入上述dt链和r链,dw、dt和r链的摩尔比为1:9:1.5,室温过夜孵育;缓慢逐次加入nacl缓冲液(2.0m)至溶液中nacl浓度为0.3m,室温继续孵育12小时;反应结束后,溶液离心清洗3次,完成制备过程。
制备得到的活细胞内原位加速的dna马达,以金纳米颗粒aunp作为载体,并在其上负载三种dna链;
第一dna链,为发生运动过程的dw链,其3’端的巯基可共价连接aunp
第二dna链,为带有特定化学标记位点的dna茎环结构(dt链),该dna茎环结构环状部分含有一个8-氧-7-氢脱氧鸟嘌呤(guanineoxidation,og)或次黄嘌呤(adenosinedeamination,i)损伤碱基位点,其茎部3’端的巯基可共价连接aunp,5’端修饰荧光分子tamra或fam;
第三dna链,为带有特定化学标记位点的dna茎环结构(r链),该dna茎环结构环状部分含有一个脱嘌呤/脱嘧啶ap位点,其茎部3’端的巯基可共价连接aunp,5’端修饰荧光分子cy5。
上述活细胞内原位加速的dna马达在活细胞成像监测时,包括以下操作:
1)将上述dna马达导入活细胞内;
2)上述dna马达上的dw链5’端14个碱基与dt链环状部分互补成双链dna,其中dt链上的损伤碱基位点og或i与细胞内的人类8-羟基鸟嘌呤dna糖基化酶(hogg1)或人类烷基腺嘌呤dna糖基化酶(haag)作用成为脱嘌呤/脱嘧啶ap位点,再由人源脱嘌呤/脱嘧啶(ap)核酸内切酶ape1(ape1)酶切后,释放出荧光信号tamra或fam。r链上的脱嘌呤/脱嘧啶ap位点,直接由人源脱嘌呤/脱嘧啶(ap)核酸内切酶ape1(ape1)酶切后,释放出cy5荧光。
3)通过激光共聚焦荧光显微镜进行观察,实现细胞内损伤碱基修复酶的原位成像和作用通路监测。
下面以hela宫颈癌细胞的成像观测作为实例,利用原位加速的dna马达进行活细胞内dna损伤碱基修复酶的成像及相关作用通路监测,对本发明的效果进行说明。
以hela宫颈癌细胞系为基本模型,将hela细胞(0.5ml,1×106ml-1)种在八孔共聚焦培养皿中,培养24小时。然后将20μl探针加入培养皿并37℃温育4小时,接着利用激光共聚焦荧光显微镜进行观察。在dna损伤碱基修复酶阳性的细胞中,dw链5’端14个碱基与dt链环状部分互补成双链dna,其中dt链上的损伤碱基位点og或i能被人类8-羟基鸟嘌呤dna糖基化酶(hogg1)或人类烷基腺嘌呤dna糖基化酶(haag)作用成为脱嘌呤/脱嘧啶ap位点,再由人源脱嘌呤/脱嘧啶(ap)核酸内切酶ape1(ape1)酶切后,释放出tarma或fam荧光。r链上的脱嘌呤/脱嘧啶ap位点,直接由人源脱嘌呤/脱嘧啶(ap)核酸内切酶ape1(ape1)酶切后,释放出cy5荧光,用于细胞内“工具酶”ape1的分析研究。
结果参见图5和图6,图5为利用设计的dna马达分别监测活细胞内两种dna损伤修复过程(g氧化成og、a脱氨成i)的荧光共聚焦成像图及对应流式细胞图。可以看到,两种阴性对照dna马达(无dw链、随机dw链)在细胞中没有产生信号或信号非常弱,包含特异性dw链的dna马达产生很高的荧光信号。这些实验结果说明,本发明中的dna马达可实现活细胞内dna损伤修复通路的实时监测,且具有同时检测相关通路中各种酶活性的功能。
本发明中dna马达所用序列:
[a]redcharactersrepresentthednadamagebases.
图6为活细胞内(hela)自校准一体化dna马达对两种损伤碱基修复酶的监测成像图。图6中,a为三种不同ape1活性的细胞样本中,利用dna马达对hogg1监测的结果。原始荧光成像图中,每个像素点的荧光强度信息(0-256)由matlab程序提取获得,对应的荧光强度分布点图列于右侧。其中,越高的分布强度表明越高的荧光信号;内参信号(紫色)能够指示并调节工具酶ape1在不同细胞样本中的差异。b图为自校准双轨道(双dt)dna马达在同一细胞样本中,对hogg1和haag同时监测的成像结果,ape1触发的内参信号稳定清晰。
以上内容仅为说明本发明的技术思想,不能以此限定本发明的保护范围,凡是按照本发明提出的技术思想,在技术方案基础上所做的任何改动,均落入本发明权利要求书的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!