用于治疗实体癌症和微生物感染的方法和组合物与流程
本发明涉及用于治疗微生物感染和实体癌症的方法和组合物,并且尤其涉及表达socs4的重组病毒及其使用方法。
背景技术:
hsv-1感染包括呼吸道在内的各种各样的粘膜组织,并且已经报道的是hsv-1诱导的肺炎是由于炎症反应而不是病毒本身的直接细胞病理效应。这种不受控的炎症反应是促炎性细胞因子的过量释放的结果,这被称为“细胞因子风暴”,其最初被用于描述流感诱导的细胞因子在短时间内过量产生,其与不受控的促炎性反应和重要的免疫病理学以及严重疾病的结局相联系。不幸的是,对参与细胞因子风暴的分子、细胞因子对发病机制的贡献、以及预防或减轻症状的治疗策略的了解仍然是不足的。
被基因工程化来选择性地在癌细胞中复制并杀死癌细胞的溶瘤病毒(ov),代表了抗肿瘤治疗的新方法。由于其基于机制的选择性、对于调节肿瘤细胞死亡的潜力以及在肿瘤位点表达额外的治疗性转基因的可能性,所以该方法是尤其具有吸引力的。考虑到溶瘤病毒被设计用于瘤内注射的事实,这种独特的癌症疗法通常与宿主的抗肿瘤免疫(免疫病毒疗法,immunovirotherapy)相结合。基于溶瘤hsv(ohsv)的1型单纯疱疹病毒(hsv-1),talimogenelaherparepvec(t-vec,imlygic)是fda首次批准的ov。如其他ov候选者一样,针对ohsv的宿主反应是复杂的、多方面的、并且通过宿主免疫力和肿瘤微环境进行调节;此外,各种免疫和炎症反应可以既是有利的又是有害的,并且通过ov递送至特定器官(例如肺)诱导的细胞因子风暴是日益被认识到的损害之一。细胞因子风暴伴随的肺损伤的进展随后已经在各种病毒、细菌或真菌感染中被报道。一致的观察表明,细胞因子风暴的概念是更加复杂的,但当前对促进细胞因子风暴的机制的了解仍然有限,并且对控制在适当细胞因子释放和细胞因子过量产生之间的平衡的对策仍然相对未开发。
技术实现要素:
本发明的第一个方面涉及包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段的重组病毒,其中一旦在细胞中复制,该重组病毒表达socs4或该功能性片段。
在一些实施方式中,重组病毒是携带编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段的重组溶瘤病毒,并且该病毒在其中复制的细胞是肿瘤细胞。在一些实施方式中,重组病毒是携带编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段的重组病毒载体,并且该病毒在其中复制的细胞是正常细胞。
本发明的另一方面涉及用于在对象中治疗癌症或者用于在对象中减少或消除溶瘤病毒疗法的副作用的方法,其包含向该对象施用治疗上有效量的重组溶瘤病毒,其中该重组溶瘤病毒包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段,并且一旦在癌细胞复制,该重组溶瘤病毒表达socs4或该功能性片段。
本发明的另外的方面涉及用于在对象中减少或消除微生物感染治疗的副作用的方法,其包含向该对象施用治疗上有效量的重组病毒载体,该载体包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段,其中一旦在正常细胞复制,该重组病毒表达socs4或该功能性片段。
在本发明中,我们已经构建了具有socs4蛋白插入片段(proteininsert)的hsv株(hsv-socs4)以研究其在控制对细胞因子风暴的诱导中的影响。本发明研究了几种起关键作用的代表性细胞因子,包括mcp-1、il-1β、tnf-α、il-6和ifn-γ,并且发现在被hsv-socs4感染的小鼠中的它们的浓度远低于在被hsv-1(f)感染的小鼠中的浓度,并且hsv-socs4小鼠示出轻微肺损伤、体重减轻较少以及100%的存活率。本发明提供了调控由溶瘤病毒疗法诱导的细胞因子风暴的有前景的技术方案。
从下文的本发明的详细阐述中,本发明的其他方面将是容易知道的。
附图说明
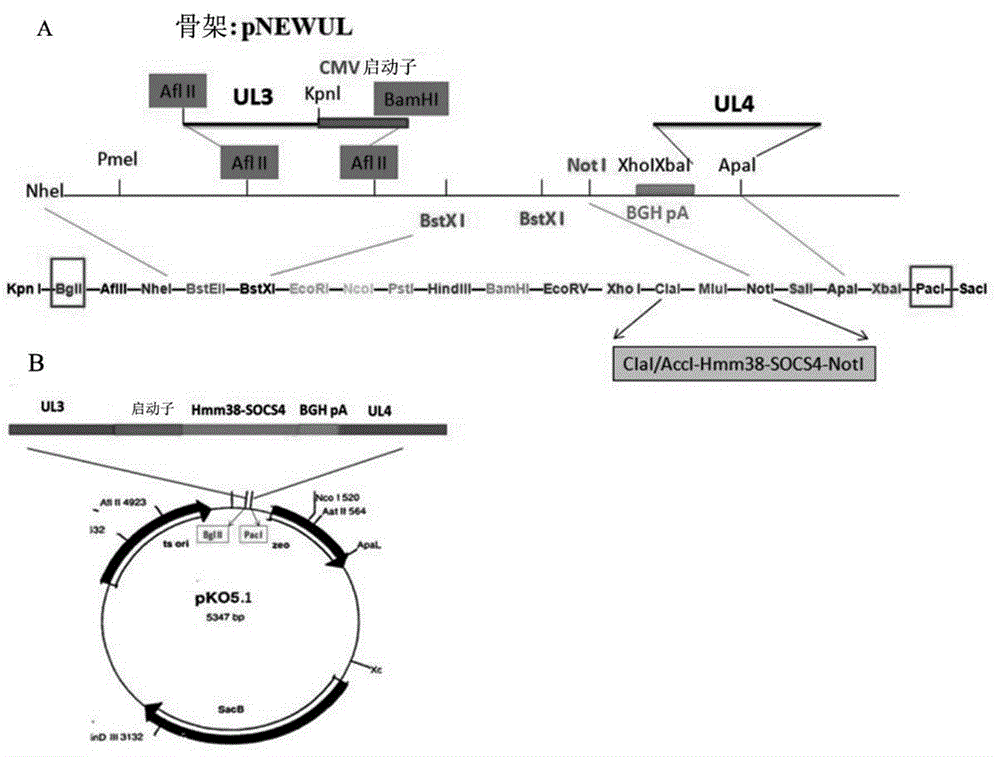
图1:hsv-socs4重构的示意图。(a)将经确认的socs4基因连接到在clai/acci和noti处的pnewul骨架位点。(b)从pnewul中切割在pnewul骨架的bglii和paci之间的序列(包括ul3、ul4和socs4),并在相同位点将其克隆到质粒pko5.1中。
图2:hsv-socs4重构的pcr确认。(a)条带1示出来自preveiver-m02的socs4(1397bp)的pcr产物。(b)从重构病毒中提取dna以进行pcr用于最终的确认,在1000bp至2000bp之间的条带示出了:ul49(1492bp)、ul3(1319bp)和socs4(1397bp)。
图3:在来自被pbs、hsv-1-f或hsv-socs4感染的小鼠的balf中的细胞因子产量。在感染后第1、3和7天收集来自小鼠(n=6)的balf以进行elisa测定。(a)在整个三天的时间里,与从hsv-socs4小鼠中检测到mcp-1的浓度相比,从hsv-1-f小鼠中检测到的显著更高浓度的mcp-1。hsv-1-f小鼠的mcp-1产量在第7天下降,但在hsv-socs4小鼠中仅示出可忽略的差异。(b)与hsv-socs4小鼠相比,在第1天和第3天观察到hsv-1-f小鼠中的il-1β水平升高,但在第7天没有观察到。hsv-1-f小鼠的il-1β的产量示出上升趋势-下降趋势曲线。(c)在hsv-1-f小鼠和hsv-socs4小鼠之间的tnf-α水平差异明显,并且hsv-1-f小鼠中测试到的最高的tnf-α水平是在第1天,然后其下降。(d)在第1、3和7天,与从hsv-1-f小鼠中测试到的il-6水平相比,从hsv-socs4小鼠中测试到明显更低水平的il-6,并且对于两组来说在第7天均增加了产量。(e)hsv-1-f小鼠的ifn-γ产量远高于来自hsv-socs4小鼠的ifn-γ产量,并且其在第3天增加并在第7天保持在高水平。作为阴性对照的模拟小鼠(nc)没有诱导可评估数量的细胞因子。*表示p值<0.05。
图4:在来自被pbs、hsv-1-f或hsv-socs4感染的小鼠血清中的细胞因子的产量。在感染后第1、3和7天收集来自每只小鼠的血清以进行elisa测定。(a)在第1天,与从hsv-socs4小鼠中检测到的mcp-1的浓度相比,从hsv-1-f小鼠中检测到显著更高浓度的mcp-1,并且hsv-1-f小鼠中的其产量连续地下降,但hsv-socs4小鼠中其仅在第7天下降。(b)hsv-1-f和hsv-socs4小鼠的il-1β值在第1天和第3天是相似的,但hsv-1-f小鼠在第7天示出了强烈的上调,因此发现了在两组小鼠之间的极大的差异。(c)hsv-1-f小鼠和hsv-socs4小鼠之间的tnf-α水平差异是明显的,并且在第7天示出hsv-1-f小鼠的tnf-α水平最高。(d)在整个三天时间里,与从hsv-1-f小鼠中测试到的il-6水平相比,从hsv-socs4小鼠中测试到明显更低水平的il-6。hsv-1-f小鼠中检测到il-6产量持续地增加,但hsv-socs4小鼠中仅在第7天示出增加的水平。(e)从hsv-1-f小鼠中测试到更多的ifn-γ产量,并且对两组来说其在第7天均增加。作为阴性对照的模拟小鼠(nc)不会诱导可评估数量的细胞因子。*表示p值<0.05。
图5:来自被hsv-1-f或hsv-socs4感染的小鼠的balf细胞的流式细胞术分析。感染后第1天和第7天收集来自小鼠的balf细胞,用cd11b染色,然后进行流式细胞术分析。(a)示出来自在第1天和第7天每一组小鼠的一个代表性结果。标记了cd11b+细胞的数量。(b)示出小鼠(n=6)的结果。从hsv-1-f小鼠中检测到占优势的cd11b+细胞;以及在两组中,与在第7天染色阳性的细胞相比,在第1天染色阳性的细胞数量更多。*表示p值<0.05。
图6:来自被hsv-1-f或hsv-socs4感染的小鼠的脾脏细胞的流式细胞术分析。在感染后第1天和第7天收集来自小鼠的脾脏细胞,用cd62l、cd8a或cd4染色,然后进行流式细胞术分析。(a)示出在第1天和第7天来自每组小鼠的代表性cd8+且cd62l+细胞的结果。指明双阳性细胞数量。(b)示出小鼠(n=6)的结果。从第1天到第7天,双阳性细胞大大增加,并且在第7天可察觉hsv-1-f小鼠和hsv-socs4小鼠之间的双阳性细胞的数量差异。(c)呈现在第1天和第7天由每组小鼠生成的一种典型的cd4+且cd62l+细胞。记录双阳性细胞的数量。(d)示出小鼠(n=6)的结果。在第7天检测到显著升高的cd4+且cd62l+细胞,并且hsv-1-f和hsv-socs4小鼠之间阳性细胞数量的区别是明显的。*表示p值<0.05。
图7:被感染小鼠中的病毒滴度(viraltitration)和病理分析。在从被感染的小鼠中移除之后,将未被提取balf的小鼠的肺切碎,收集上清液用于在单层vero细胞上的病毒滴度分析。(a)在第1天观察到最大病毒滴度,然后下降,并且在第7天未检测到病毒。在第3天,病毒载量在hsv-1-f小鼠和hsv-socs4小鼠之间展示出明显的差异。(b)没有被采集balf的小鼠的肺被送去进行病理分析,并示出200x代表性的照片。在第1天,hsv-socs4小鼠的肺部几乎未示出病理变化,但在hsv-1-f小鼠肺中示出了明显的细胞浸润,伴随局部毛细血管的轻度至中度扩张和充血。在第7天,在被hsv-socs4感染的小鼠的肺中观察到一些免疫细胞的浸润以及毛细血管的轻微扩张和充血,但是肺泡壁的结构没有破裂;hsv-1-f小鼠的肺中出现增厚和破裂的肺泡壁,伴随周围严重充血,被免疫细胞充斥。*表示p值<0.05。
图8:被pbs、hsv-1-f或hsv-socs4感染后的小鼠的体重和死亡率。通过鼻内途径被感染后,每天两次对小鼠进行监测,持续12天。(a)示出了所有存活小鼠的平均体重(g)。被hsv-1-f感染的小鼠在第2天体重开始逐渐地减轻,在第7天这样的减轻变得急剧,并且最后存活的小鼠在第10天减轻了50%体重。hsv-socs4组小鼠轻微地减重,并且普遍在第12天保持80%的体重。(b)hsv-1-f小鼠的存活率在第7天开始下降,然后小鼠迅速死亡,并且在第11天没有小鼠存活。hsv-socs4小鼠的存活率维持在100%。pbs模拟小鼠没有示出体重的减轻,也没有死亡。*表示p值<0.05。
发明详述
定义
应当注意的是,术语“一”或“一个”实体指该实体中的一个或多个,例如,“癌细胞”被理解为代表一个或多个癌细胞。因此,术语“一个”(或“一”),“一个或多个”和“至少一个”在本文中可以互换地使用。
如本文所用,术语“治疗”指治疗性处理和预防性或防止性措施,其中目的是预防或减缓(减轻)不期望的生理变化或病症,例如癌症的进展。无论是可检测的还是不可检测的,有益或期望的临床结果都包括但不限于:症状的缓解、疾病程度的减轻、疾病状态的稳定(即不恶化)、疾病进展的延迟或减慢,疾病状态的改善或缓和以及缓解(无论是部分还是全部)。“治疗”也可意为与未接受治疗的预期生存期相比使生存期延长。需要治疗的对象包括已患有疾病或病症的那些对象以及易于患疾病或病症的那些对象或要预防疾病或病症的那些对象。
“对象”或“个体”或“动物”或“患者”或“哺乳动物”是指需要诊断、预后或治疗的任意对象,尤其是哺乳动物对象。哺乳动物对象包括人类、家养动物、农场动物以及动物园动物、竞技动物或宠物,例如狗、猫、豚鼠、兔、大鼠、小鼠、马、牛、奶牛等。本文的对象优选为人类。
如本文所用,例如“对于需要治疗的患者”或“需要治疗的对象”的短语包括将从施用例如用于检测、用于诊断程序和/或用于治疗的本公开的重组病毒或组合物中受益的对象,例如哺乳动物对象。
本说明书中所使用的术语“治疗上有效量”或“药学上有效量”是指可以在临床上发挥显著效果的每种活性成分的量。根据例如制剂方法、给药方式、患者年龄、体重、性别、病理状况、饮食、给药时间、给药间隔、给药方式、排泄速度以及反应敏感性的因素,可以以各种方式开具用于单次剂量的重组病毒的药学上有效量。例如,用于单次剂量的重组病毒的药学上有效量可以在0.001至100mg/kg或0.02至10mg/kg的范围内,但不限于此。可以将用于单次剂量的药学上有效量配制成单位剂型的单一制剂或配制成适当分开的剂型,或者可以将其制造成被容纳在多剂量容器中。
本文可互换使用的术语“多聚核苷酸”和“核酸”是指任何长度的核苷酸的聚合形式,即核糖核苷酸或脱氧核糖核苷酸。这些术语包括单链、双链或三链dna,基因组dna,cdna,基因组rna,mrna,dna-rna杂合体或包含嘌呤和嘧啶碱基或其他天然的、化学、生物化学修饰的、非天然或衍生的核苷酸碱基的聚合物。多聚核苷酸的骨架可包含糖和磷酸基团(通常可在rna或dna中发现)或被修饰或被取代的糖或磷酸基团。或者,多聚核苷酸的骨架可包含合成亚基的聚合物,例如氨基磷酸酯,并且因此可以是寡脱氧核苷氨基磷酸酯(p-nh2)或混合的氨基磷酸酯-磷酸二酯的寡聚物。
此外,可以进行核苷酸或氨基酸替换、缺失或插入,从而导致在“非必需”氨基酸区域处的保守性替换或改变。例如,衍生自指定蛋白质的多肽或氨基酸序列可以与起始序列相同,除了一个或多个单独的氨基酸替换、插入或缺失(例如一个、两个、三个、四个、五个、六个、七个、八个、九个、十个、十五个、二十个或更多单独的氨基酸替换、插入或缺失)之外。在某些实施方式中,衍生自指定蛋白质的多肽或氨基酸序列相对于该起始序列具有一个至五个、一个至十个、一个至十五个或一个至二十个单独的氨基酸替换、插入或缺失。
本说明书所使用的术语“重组病毒”或“重构病毒”是指已经被遗传改变的病毒,例如通过将异源性核酸构建体添加或插入到病毒的基因组中和/或通过缺失来自病毒的基因组的内源性多聚核苷酸。如本申请中所使用的,该术语是指携带编码socs4蛋白或其功能性片段的异源性核酸序列片段的重组溶瘤病毒或重组病毒载体。
溶瘤病毒
在本领域中已知并描述了许多溶瘤病毒,可以预期将其中任意一个用于本发明。举例来说,合适的溶瘤病毒包括1型单纯疱疹病毒、2型单纯疱疹病毒、水疱性口炎病毒、溶瘤腺病毒、新城疫病毒、牛痘病毒及这些病毒的突变株。在一个实施方式中,溶瘤病毒是选择复制性的或有复制能力的。在一个实施方式中,溶瘤病毒是无复制能力的。
在一些实施方式中,可用于当前方法和组合物的溶瘤病毒是复制选择性的。可以理解的是,如果将溶瘤病毒的复制置于基因表达的调控因子(例如衍生自白蛋白基因5'侧的增强子/启动子区域)的控制之下,则可使溶瘤病毒是复制选择性的。举例来说,通过将hsv基因可操作地连接到tgf-β启动子,可将hsv的主要转录单位置于肿瘤生长因子-β(tgf-β)启动子的转录控制之下。已知的是,相对于相同类型的非肿瘤细胞,某些肿瘤细胞过度表达tgf-β。因此,其中复制受tgf-β启动子的转录控制的溶瘤病毒是复制选择性的,因为与相同类型的非肿瘤细胞相比其更能在某些肿瘤细胞中复制。可以使用已知在受影响的细胞中选择性地引起过度表达的任何基因表达的调控因子,来制备相似的复制选择性的溶瘤病毒。例如复制选择性溶瘤病毒可以是hsv-1突变体,其中使编码icp34.5的基因突变或缺失。
根据本发明的溶瘤病毒可以在其基因组中进一步包含其他的修饰。例如,它可以包含被插入到ul44基因中的其他dna。该插入可产生ul44基因的功能失活和生成的裂解表型,或者可将其插入已经失活的基因中或替换缺失的基因。
溶瘤病毒还可在其中掺入一个或多个启动子,这些启动子赋予该病毒增强水平的肿瘤细胞特异性。以这种方式,可以使用肿瘤细胞特异性启动子,将溶瘤病毒靶向至具体的肿瘤类型。术语“肿瘤细胞特异性启动子”或“肿瘤细胞特异性转录调节序列”或“肿瘤特异性启动子”或“肿瘤特异性转录调节序列”表明,与正常细胞相比在靶标肿瘤细胞中以较高水平存在的转录调节序列、启动子和/或增强子。例如,用于本发明的溶瘤病毒可以在被外源地加入的调节因子(例如四环素)的控制下。
在一个实施方式中,本发明的溶瘤病毒(例如ohsv)被工程化以将至少一种病毒复制必需的病毒蛋白置于肿瘤特异性启动子的控制下。或者,可以将编码细胞毒性剂的基因(病毒基因或外源性基因)置于肿瘤特异性启动子的控制之下。如本文所用,细胞毒性剂意为引起细胞死亡的任何蛋白质。例如,这将包括蓖麻毒素、白喉毒素或其截短形式。还包括编码前药、细胞因子或趋化因子的基因。这样的溶瘤病毒可以利用来自在被靶向的肿瘤中被高度表达的基因的启动子,例如表皮生长因子受体(egfr)的启动子或碱性成纤维细胞生长因子(bfgf)的启动子、或其他与肿瘤相关的启动子或增强子元件。
用于本发明的一种这样的溶瘤病毒是溶瘤单纯疱疹病毒(ohsv)。ohsv将包含编码本文所述多肽中的一个或多个的一个或多个外源性核酸。生成包含这样外源性核酸的ohsv的方法在本领域中是已知的。将核酸插入到ohsv基因组中的具体位置可以由熟练的技术人员确定。
溶瘤单纯疱疹病毒(ohsv)在本领域中是已知的,并且包括1型单纯疱疹病毒和2型单纯疱疹病毒。在一个实施方式中,用于本发明的方法、组合物和试剂盒的ohsv是复制选择性的或具有复制能力的,例如本文所述的实施例中的一个。在一个实施方式中,ohsv是无复制能力的。
1型单纯疱疹病毒是其中优选的病毒,例如不表达功能性icp34.5的hsv-1病毒的变体,并且因此其展示显著低于对应野生型的神经毒性。这样的变体包括但不限于ohsv-r3616。其他示例性hsv-1病毒包括1716、r3616和r4009。可以使用的其他具有复制选择性的hsv-1病毒株包括:例如r47δ(其中缺失了编码蛋白质icp34.5和icp47的基因)、g207(缺失了编码icp34.5和核糖核苷酸还原酶的基因)、nv1020(缺失了编码ul24、ul56和内部重复的基因)、nv1023(缺失编码ul24、ul56、icp47和内部重复的基因)、3616-ub(缺失了编码icp34.5和尿嘧啶dna糖基化酶的基因)、g92δ(其中白蛋白启动子驱动必需的icp4基因的转录)、hrr3(缺失编码核糖核苷酸还原酶的基因)和r7041(缺失了us3基因)以及不表达功能性icp34.5的hsv株。
用于本文所述的方法和组合物的ohsv不限于本文所述的hsv-1突变株中的一个。可以使用任意单纯疱疹病毒的复制选择性毒株。除了本文所述的hsv-1突变株之外,本领域还描述了其他具有复制选择性的hsv-1突变株。此外,在本文所述的方法和组合物中还可以使用hsv-2突变株,例如hsv-2株2701(缺失rl基因)、deltarr(缺失icp10pk基因)和fuson-h2(缺失icp10pk基因)。
也可以分离hsv的非实验室毒株并且使其适合用于本发明。此外,hsv-2突变株(例如hsv-2株hsv-2701、hsv-2616和hsv-2604)可以用于本发明的方法。
在一个实施方式中,重组溶瘤病毒是重组ohsv-1,其包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段,其中该外源多聚核苷酸片段位于ohsv-1的ul3和ul4基因之间。在一个实施方式中,重组ohsv-1是f株(ohsv-1(f))。
病毒载体
在本发明的一个实施方式中,重组病毒是携带编码socs4多聚核苷酸的重组病毒载体。病毒载体也可以称为载体、载体病毒粒子或载体颗粒。在另一个实施方式中,病毒载体衍生自逆转录病毒、腺病毒、腺相关病毒、单纯疱疹病毒、牛痘病毒或杆状病毒。本发明中所用的术语“病毒载体”是指衍生自病毒的用于将多聚核苷酸(例如编码socs4或其变体)递送至正常细胞并在该细胞中表达该多聚核苷酸的载体。
逆转录病毒载体可衍生自或能够衍生自任意适当的逆转录病毒。已经鉴定出大量不同的逆转录病毒。例子包括:鼠白血病病毒(mlv)、人类t细胞白血病病毒(htlv)、小鼠乳腺肿瘤病毒(mmtv)、劳斯肉瘤病毒(rsv)、藤浪肉瘤病毒(fusv)、莫洛尼鼠白血病病毒(momlv)、fbr鼠骨肉瘤病毒(fbrmsv)、莫洛尼鼠肉瘤病毒(mo-msv)、abelson鼠白血病病毒(a-mlv)、禽骨髓细胞瘤病毒29(mc29)和禽成红细胞增多症病毒(aev)。逆转录病毒可以衍生自泡沫病毒。
慢病毒是更大群体的逆转录病毒的一部分。简而言之,慢病毒可以分为灵长类群体和非灵长类群体。灵长类慢病毒的例子包括但不限于:人类免疫缺陷病毒(hiv),人类自身免疫缺陷综合症(aids)的致病物和猿猴免疫缺陷病毒(siv)。非灵长类慢病毒群体包括原型“慢病毒”维斯纳/梅迪病毒(visna/maedivirus,vmv)以及相关的山羊关节炎-脑炎病毒(caev)、马传染性贫血病毒(eiav)、猫免疫缺陷病毒(fiv)和牛免疫缺陷病毒(biv)。如本文所用的慢病毒载体是包含至少一个可衍生自慢病毒的组成部分的载体。优选地,该组成部分参与通过其载体感染细胞、表达基因或被复制的生物学机制。慢病毒载体可以来源于灵长类慢病毒(例如hiv-1)或非灵长类慢病毒。非灵长类慢病毒的例子可以是慢病毒科家族的任何成员,它不会天然地感染灵长类,并且可以包括猫免疫缺陷病毒(fiv)、牛免疫缺陷病毒(biv)、山羊关节炎脑炎病毒(caev)、梅迪维斯纳病毒(mvv)或马传染性贫血病毒(eiav)。在另一个实施方式中,慢病毒载体衍生自hiv-1、hiv-2、siv、fiv、biv、eiav、caev或维斯纳慢病毒。
在本发明的另一个实施方式中,病毒载体可以是腺病毒载体。腺病毒是双链线性dna病毒,该病毒不会通过rna中间体复制。腺病毒是双链dna无包膜病毒,其能够在体内、离体和体外转导一系列的人类和非人类来源的细胞类型。这些细胞包括呼吸道上皮细胞、肝细胞、肌肉细胞、心肌细胞、滑膜细胞、原代乳腺上皮细胞和有丝分裂后期终末分化的细胞(例如神经元)。腺病毒已被用作用于基因治疗和异源性基因表达的载体。大(36kb)基因组可以容纳多达8kb的外源插入dna,并且能够在互补细胞系中高效地复制以产生非常高的滴度,高达每毫升1012个转导单位。因此,腺病毒是研究在原代非复制细胞中的基因表达的最佳系统之一。来自腺病毒基因组的病毒或外源基因的表达不需要复制细胞。腺病毒载体通过受体介导的内吞作用进入细胞。一旦在细胞之内,腺病毒载体就很少整合到宿主染色体中。相反,它们作为附加体(独立于宿主基因组)以线性基因组形式存在于宿主细胞核中。
腺相关病毒(aav)是本发明中使用的吸引人的载体系统,因为它具有高整合频率并且可以感染非分裂细胞。这使得其可用于将基因递送到哺乳动物细胞中。aav具有广宿主范围的感染性。重组aav载体已成功用于标记基因和参与人类疾病的基因的体外、离体和体内转导。已经开发出某些aav载体,其可以有效地结合大的有效载荷(高达8-9kb)。一种这样的载体具有aav5衣壳和aav2itr。
单纯疱疹病毒(hsv)是一种包膜的双链dna病毒,其可天然地感染神经元。它可以容纳外源dna的大区段,使其成为有吸引力的载体系统,并已被用作至神经元的基因递送的载体。在治疗程序中hsv的使用需要使毒株减毒,从而它们不能建立裂解周期。尤其地,如果将hsv载体用于人类的基因治疗,则多聚核苷酸应优选地被插入到必需基因中。这是因为如果病毒载体遇到野生型病毒,则可以通过重组将异源性基因转移至野生型病毒。然而,如果以防止其复制的方式构建重组病毒,则可以通过将寡核苷酸插入对复制必需的病毒基因中来实现。
本发明的病毒载体可以是牛痘病毒载体,例如mva或nyvac。牛痘载体的替代物包括:禽痘载体(avipoxvector),例如称为alvac的鸡痘或金丝雀痘;以及由其衍生的毒株,其可以在人类细胞中感染并表达重组蛋白但不能复制。应当理解的是,在插入重组基因后,病毒基因组的部分可以保持完整。这意味着病毒载体可以保留感染细胞并随后表达额外的基因的能力的概念,该额外的基因支持其复制并可能促进被感染细胞的裂解和死亡。
和功能性片段和变体
本文所述的溶瘤病毒或病毒载体包含编码socs4或其生物活性片段的核酸序列,其以可表达的形式掺入病毒基因组中。因此,该病毒用作将socs4递送至被感染细胞的载体。本发明预期了各种形式的socs4(例如本文所述的那些形式的socs4)的使用,各种形式的socs4包括但不限于socs4的功能性结构域或治疗性socs4结构域或治疗性socs4变体,以及它们的片段、变体和衍生物,以及包含这些形式的socs4(例如本文所述)中的一个的融合蛋白。
如本说明书中所使用的术语“socs4”是指细胞因子信号传导抑制因子4,其是含有sh2结构域和socsbox结构域的蛋白质。因此,该蛋白质属于细胞因子信号转导抑制因子(socs),也称为stat诱导的stat抑制剂(ssi)蛋白质家族。已知通过抑制信号转导及转录激活因子3(stat3),socs4负责抑制免疫信号传导,而缺乏功能性socs4的小鼠对甲型流感病毒的原发性感染高度易感,展现出在肺部中的促炎性细胞因子产生失调,病毒清除延迟,以及流感特异性cd8+t细胞至感染部位的运输受损。socs4的功能性片段/变体/衍生物是指与天然socs4基本上同源的多肽,但是由于一个或多个缺失、插入或替换,所以其具有不同于天然socs4的氨基酸序列。socs4的例子是来自智人,其具有genbank登记号nc_000014.9。
还预期了用于本发明的保留socs4的期望生物学活性的天然socs4蛋白的片段、变体和衍生物通过溶瘤病毒或病毒载体进行递送。在一个实施方式中,socs4的片段、变体或衍生物的生物学活性基本上等于相应的天然socs4蛋白的生物学活性。在一个实施方式中,用于确定活性的生物学活性是抑制细胞因子信号传导的活性。在一个实施方式中,通过片段、变体或衍生物保留了100%的活性。在一个实施方式中,与全长的天然socs4相比保留了少于100%的活性(例如95%、90%、85%、80%、75%、70%、65%、60%、55%、50%、45%、40%)。
例如,可以通过天然socs4核苷酸序列的一个或多个添加、缺失、突变或替换来获得socs4变体。变体氨基酸或dna序列与天然socs4序列优选为至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或更多相同。例如可以通过使用在万维网上通常用于此目的的免费使用的计算机程序来比较两个序列,以确定天然序列和突变序列之间的同源性程度或同一性百分比。
天然氨基酸序列的改变可以通过本领域技术人员已知的许多已知技术中的任意一种来完成。例如,可以通过合成含有突变序列的寡核苷酸,在特定的基因座处引入突变,所述寡核苷酸的两侧是能够与天然序列的片段连接的限制性位点。在连接后,生成的重建序列编码具有所期望的氨基酸插入、替换或缺失的类似物。或者,可采用寡核苷酸定向的位点特异性突变形成方法,以提供改变的核苷酸序列,该改变的核苷酸序列具有根据所需要的替换、缺失或插入而改变的特定密码子。
在一些实施方式中,socs4变体可以包含被保守替换的序列,意味着天然socs4多肽的一个或多个氨基酸残基由不同的残基替换,并且被保守替换的socs4多肽保留了所期望的生物学活性,该生物学活性基本上等同于天然socs4多肽的生物学活性。保守替换的例子包括不改变socs4的二级和/或三级结构的氨基酸的替换。
其他核酸
包含socs4核酸的重组溶瘤病毒或病毒载体可以进一步包含额外的异源性核酸序列(例如以可表达的形式),在本文中称为第二异源性核酸序列、第三异源性核酸序列等。或者,重组溶瘤病毒或病毒载体可不含有额外的异源性核酸序列。
可以插入任何所期望的dna,包括编码可选择标记的dna,或优选编码治疗性、生物活性蛋白(例如干扰素、细胞因子、趋化因子)的基因,或更优选编码前药转化酶(包括胸苷激酶、胞嘧啶脱氨酶、cyp450以及其他)的dna。在一个实施方式中,核酸编码抑制肿瘤生长的蛋白质(例如化学治疗剂、生长调节剂)或修饰免疫应答的蛋白质。化学治疗剂的例子是丝裂霉素c。在一个实施方式中,核酸编码生长调节分子(例如在肿瘤的肿瘤发生中已经丢失的一种)。这样分子的例子不限于来自半胱天冬酶家族的蛋白,例如半胱天冬酶-9(p55211(casp9_human);hgnc:15111;entrezgene:8422;ensembl:ensg000001329067;omim:6022345;uniprotkb:p552113),半胱天冬酶-8(q14790(casp8_human);9606[ncbi]),半胱天冬酶-7(p55210(casp7_human);9606[ncbi])和半胱天冬酶-3(hcgn:1504;ensembl:ensg00000164305;hprd:02799;mim:600636;vega:otthumg00000133681);促凋亡蛋白,例如bax(hgnc:9591;entrezgene:5812;ensembl:ensg000000870887;omim:6000405;uniprotkb:q078123),bid(hgnc:10501;entrezgene:6372;ensembl:ensg000000154757;omim:6019975;uniprotkb:p559573),bad(hgnc:9361;entrezgene:5722;ensembl:ensg000000023307;omim:6031675;uniprotkb:q92934),bak(hgnc:9491;entrezgene:5782;ensembl:ensg000000301107;omim:6005165;uniprotkb:q166113),bcl2l11(hgnc:9941;entrezgene:100182;ensembl:ensg000001530947;omim:6038275;uniprotkb:0435213),p53(hgnc:119981;entrezgene:71572;ensembl:ensg000001415107;omim:1911705;uniprotkb:p046373),puma(hgnc:178681;entrezgene:271132;ensembl:ensg000001053277;omim:6058545;uniprotkb:q96pg83;uniprotkb:q9bxh13),diablo/smac(hgnc:215281;entrezgene:566162;ensembl:ensg000001840477;omim:6052195;uniprotkb:q9nr283)。在一个实施方案中,核酸编码免疫调节剂(例如,免疫刺激性转基因),包括但不限于:flt-3配体、hmbg1、钙网蛋白、gitr配体、白介素-12、白介素-15、白介素18或ccl17。
本领域技术人员可以将外源性核酸插入到溶瘤病毒中。在一个实施方式中,溶瘤病毒是hsv,并且将外源性核酸插入到病毒基因组的胸苷激酶(tk)基因中或替换缺失的tk基因。当溶瘤病毒包含第二外源性核酸时,该核酸优选编码抗癌基因或溶瘤基因产物。基因产物可以是仅抑制被病毒感染的细胞的生长或复制的一种产物(例如反义寡核苷酸),或者其可以是在裂解被病毒感染的细胞时发挥显著的旁观者效应的一种产物(例如胸苷激酶)。
组合物
本发明的另一方面提供了药物组合物,其包含治疗上有效量的重组病毒和药学上可接受的载体。该药物组合物旨在用于在对象中治疗肿瘤或消除或减少溶瘤肿瘤疗法或抗病毒治疗的副作用。重组病毒可以在合适的药学上可接受的载体或赋形剂中制备。在正常的储存和使用的条件下,这些制剂可包含防腐剂以防止微生物的生长。适用于可注射用途的药物形式包括无菌水溶液或分散液和用于临时制备无菌注射溶液或分散液的无菌粉末。在所有情况下,该形式都必须是无菌的,并且必须是流体以便容易注射。在生产和储存条件下它必须是稳定的,并且必须使其不受微生物(如细菌和真菌)的污染。
载体可以是含有例如水、乙醇、多元醇(例如甘油、丙二醇和液体聚乙二醇等)、其合适的混合物和/或植物油的溶剂或分散介质。可以例如通过使用例如卵磷脂的包衣,在分散液的情况下通过维持所需的粒径,以及通过使用表面活性剂,来维持适当的流动性。可以通过各种抗菌剂和抗真菌剂,例如对羟基苯甲酸酯、氯丁醇、苯酚、山梨酸、硫柳汞等来预防微生物的作用。在许多情况下,优选包括等渗剂,例如糖或氯化钠。通过在组合物中使用延迟吸收的试剂(例如单硬脂酸铝和明胶),可以使可注射组合物的吸收延长。
对于在水溶液中的肠胃外给药,例如溶液应适当地缓冲溶液(如果需要),并且首先用足够的盐水或葡萄糖使液体稀释剂等渗。这些特定的水溶液特别适合于静脉内、肌内、皮下、肿瘤内和腹膜内给药。就此而言,根据本公开,可以采用的无菌水性介质是本领域技术人员已知的。例如,可以将一个剂量溶解在1ml等渗的nacl溶液中,或者加入到1000ml皮下灌注液中,或者在所建议的输注部位注射。根据所治疗的对象的状况剂量必然会发生一些变化。在任何情况下,负责给药的人员均应确定个体对象的适合剂量。此外,对于人体给药,制剂应符合fda生物制品标准所要求的无菌性、热原性、一般安全性和纯度标准。
通过将活性化合物以所需数量与上述列举的各种其他成分掺入适当的溶剂中,然后过滤灭菌,从而制备无菌可注射溶液。通常,通过将各种灭菌的活性成分掺入无菌载体中来制备分散体,所述无菌载体含有基本分散介质和来自上述列举的那些的所需的其他成分。在用于制备无菌注射溶液的无菌粉末的情况下,优选的制备方法是真空干燥和冷冻干燥技术,其从其先前的无菌过滤溶液中生产活性成分和任何其他所需成分的粉末。
如本文所用,“载体”包括任何和所有溶剂、分散介质、载体、包衣、稀释剂、抗菌剂和抗真菌剂、等渗剂和吸收延迟剂、缓冲剂、载体溶液、悬浮液、胶体等。这样的介质和试剂用于药物活性物质的用途是本领域已知的。除非任何常规介质或试剂与活性成分是不相容的,否则预期其在治疗组合物中的用途。补充的活性成分也可以掺入组合物中。
短语“药学上可接受的”是指当施用至人类时不会产生过敏或类似不良反应的分子实体和成分。含有蛋白质作为活性成分的水性组合物的制备在本领域中是充分理解的。通常,将这类组合物制备成可注射剂,或是液体溶液或是悬浮液;也可以制备适合于在注射之前溶解或悬浮在液体中的固体形式。
治疗的方法
本发明的另一方面涉及在对象中治疗增殖性病症的方法。该方法包括向对象施用包含本文所述的socs4核酸序列的重组溶瘤病毒,从而使展现出非期望的增殖的细胞与有效量的重组溶瘤病毒进行接触。
在一个实施方式中,该增殖性疾病是肿瘤,并且本发明的方法涉及抑制肿瘤进展的方法。使有效量的重组溶瘤病毒与肿瘤进行接触,从而将该病毒递送至肿瘤细胞。术语“肿瘤”或“癌症”是指经历不受控制的增殖的组织块或组织类型或细胞类型。如本文所用,术语“肿瘤”是指包含不受控制地生长(即,过度增殖性疾病)的被转化细胞的恶性组织。肿瘤包括白血病、淋巴瘤、骨髓瘤、浆细胞瘤等;以及实体肿瘤。可以根据本发明治疗的实体肿瘤的例子包括但不限于肉瘤和癌,例如黑素瘤、纤维肉瘤、粘液肉瘤、脂肪肉瘤、软骨肉瘤、成骨肉瘤、脊索瘤、血管肉瘤、内皮肉瘤、淋巴管腺肉瘤、淋巴管内皮肉瘤(lymphangioendotheliosarcoma)、滑膜瘤、间皮瘤、尤因氏肿瘤(ewing'stumor)、平滑肌肉瘤、横纹肌肉瘤、结肠癌、胰腺癌、乳腺癌、卵巢癌、前列腺癌、鳞状细胞癌、基底细胞癌、腺癌、汗腺癌、皮脂腺癌、乳头状癌、乳头状腺癌、囊状腺癌、髓样癌、支气管癌、肾细胞癌、肝癌、胆管癌、绒毛膜癌、精原细胞瘤、胚胎性癌、威尔姆斯肿瘤(wilms'tumor)、宫颈癌、睾丸肿瘤、肺癌、小细胞肺癌、膀胱癌、上皮癌、神经胶质瘤、星形细胞瘤、成神经管细胞瘤、颅咽管瘤、室管膜瘤、松果体瘤、成血管细胞瘤、听神经瘤、少突神经胶质瘤、脑膜瘤、成神经细胞瘤和视网膜母细胞瘤。
消除或减少副作用的方法
本发明的另一个方面涉及用于在对象中减少或消除溶瘤病毒疗法的副作用的方法,该方法包含向该对象施用治疗上有效量的重组溶瘤病毒,其中该重组溶瘤病毒包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段,并且一旦在癌细胞中复制,就表达socs4或该功能性片段。
在一些实施方式中,副作用是细胞因子过量产生,在本文中也称为细胞因子风暴。在一些实施方式中,细胞因子是mcp-1、il-1β、il-6、tnf-α和ifn-γ中的任意一个或多个。在一些实施方式中,细胞因子过量产生的临床综合征或结果是肺组织损伤。在一些实施方式中,示出肺组织损伤为以下特征中的任意一个或多个:细胞浸润,轻度至中度扩张,局部毛细血管充血,肺泡壁增厚,肺泡壁破裂,肺泡壁周围充血以及充斥的免疫细胞。
本发明的另一方面涉及在对象中减少或消除微生物感染治疗的副作用的方法,该方法包括向该对象施用治疗上有效量的重组病毒,该重组病毒包含编码细胞因子信号传导抑制因子4(socs4)或其功能性片段的外源性多聚核苷酸片段,其中一旦在细胞中复制该重组病毒表达socs4或该功能性片段。在一些实施方式中,微生物感染是细菌感染、病毒感染或真菌感染中的任意一种。
在一些实施方式中,副作用是细胞因子过量产生,在本文中也称为细胞因子风暴。在一些实施方式中,细胞因子是mcp-1、il-1β、il-6、tnf-α和ifn-γ中的任意一个或多个。在一些实施方式中,细胞因子过量产生的临床综合征或结果是肺组织损伤。在一些实施方式中,示出肺组织损伤为以下特征中的任意一个或多个:细胞浸润,轻度至中度扩张,局部毛细血管充血,肺泡壁增厚,肺泡壁破裂,肺泡壁周围充血以及充斥的免疫细胞。
实施例
材料和方法
动物:6周龄的雌性balb/c小鼠购自广东省实验动物中心,并在无微生物病原体情况下在广州医科大学动物中心饲养。所有涉及小鼠的程序均经过广州医科大学动物保护与使用委员会的批准。
细胞和病毒株:vero细胞获得自美国典型培养物保藏中心,并在dmem(高糖)中培养,其中dmem(高糖)分别补充了5%(vol/vol)胎牛血清(fbs)或5%(vol/vol)新生牛血清(nbcs)。在我们实验室中使用的原型hsv-1毒株hsv-1(f),在vero细胞上繁殖并滴定。具有智人的细胞因子信号传导抑制因子4(socs4)mrna的preveiver-m02购自genecopoeiainc.。
具有表达socs4的hsv重组病毒的构建:根据智人的socs4基因序列设计了两个寡核苷酸引物:正向,5'-gtcgacatgtggtggcgcctgtggtggctctgctgctgtggcccatggtgtgggccgcagaaaataatgaaaatattag-3’(带有acci位点以及下划线的信号肽hmm38);反向,5’-gcggccgcctagcattgctgttctggtgcatc-3’(带有noti位点)。在在50µl总反应体积中进行pcr,30次循环,该循环由在95°c进行1分钟的变性步骤,在58°c进行30秒的引物退火步骤和在72°c进行2分钟的引物延伸步骤组成。将socs4的pcr产物连接到t-easy质粒中,然后转化到xl-1blue细胞中并送去进行测序确认。将已确认的socs4基因从t-easy质粒上切下,并在clai/acci和noti处的位点处连接到载体pnewul骨架(图1a),然后从pnewul中将bglii与paci之间的序列(包括ul3、ul4和socs4)切割,并在相同位点将其克隆到质粒pko5.1中(图1b)。将重组质粒转化成bac,并在43°c在具有氯霉素和博莱霉素(zeocin)的lb平板上培养过夜,然后,挑选细菌克隆并于30°c在具有氯霉素和蔗糖的lb平板上富集过夜以切除质粒pko5.1。在通过socs4基因的pcr鉴定后,培养阳性重组bac使其增殖,然后根据制造商(lifetechnologiescorporation)的说明书使用lipoectamine®ltx和plus将其转化为vero细胞。孵育细胞直到示出病毒细胞病变效应。将病毒蚀斑(viralplaque)收集到1ml牛奶中,然后经过冻融(-80°c和37°c)3次以释放病毒。将病毒接种并培养于25cmvero细胞(pd)中,提取dna以进行pcr用于最终确认(包括ul3、ul4和socs4基因)。将所期望的病毒命名为hsv-socs4,收获并储存在-80°c用于进一步的使用。
小鼠感染:将balb/c小鼠随机分为3组:分别地,一组用hsv-1(f)进行感染,一组用hsv-socs4,以及一组用pbs作为模拟感染。详细地,将每只小鼠轻度麻醉,然后通过鼻内途径用30μl106pfu的hsv-1(f)或hsv-socs4的pbs溶液或仅30μlpbs进行感染。在感染后,每天称重小鼠并监测其发病率和死亡率,持续12天。
血清和支气管肺泡灌洗液样品的收集和双夹心elisa:感染后第1天、第3天和第7天收集来自每只小鼠的眼眶血,并分离血清,然后保存于-20°c用于进一步的elisa测定。出血后,处死小鼠,并通过被插入到气管中的钝针用1mlpbs冲洗肺3次,以收集支气管肺泡灌洗液(balf)。离心样品,然后移除上清液用于elisa测定,并且合并来自同一组的两只小鼠的细胞,从而获得足够的细胞来进行流式细胞术分析。对于balf和血清样品,通过双夹心elisa评估了一组细胞因子,包括mcp-1、il-1β、il-6、tnf-α和ifn-γ。根据制造商(dakewebiothechcompanylimited)的说明书,相对于标准曲线确定了每种细胞因子的浓度。
细胞分离和流式细胞术分析:将从balf收获的细胞用tris-nh4cl处理以裂解红细胞,然后洗涤两次并重悬于冷rpmi1640培养基中。将来自被处死的小鼠的脾脏切除,充分研磨,并通过无菌金属丝网冲洗组织。收集脾脏细胞悬浮液并裂解红细胞,然后洗涤细胞并重悬。计数balf细胞和脾脏细胞,并将其调节至2×106个细胞/ml。洗涤细胞并用表面标记物染色:用于脾脏细胞的apc-cd4(apc抗小鼠cd4mab,克隆gk1.5)、fitc-cd8a(fitc抗小鼠cd8mab,克隆53-6.7)和pe-cd62l(pe抗小鼠cd62lmab,克隆mel-14);以及用于balf细胞的pb-cd11b(pb抗小鼠cd11bmab,克隆m1/70)。将所有细胞染色10分钟,洗涤并重悬于2%甲醛-pbs中,用于在cytoflex流式细胞仪(beckmancoulterinc.)和cytexpert2.0软件上的流式细胞术分析。
肺样品用于病毒滴度分析和病理分析:取血并处死后,直接移除来自每组的两只小鼠的肺,用细胞培养基完全切碎,然后收集组织匀浆,离心并收集上清液用于病毒滴度分析。vero细胞以2×105个/孔在6孔板上生长,直到细胞形成90%的单层,然后加入上清液样品。孵育24小时后,小心移除细胞,离心,将细胞团块重悬于1ml牛奶中,并保存于-80°c。在上述的3次冻融处理后,用1%nbcs将被释放的病毒样品分别以1:100、1:1000和1:10000稀释。将100μl的每种稀释液加入到在6孔板上的vero单层细胞中并培养2小时,然后将培养基替换为新的培养基(bme+1%nbcs+0.5%igg),并将细胞孵育额外的72小时。之后,收集细胞并用0.03%的亚甲基蓝染色以量化噬菌斑。由广州医科大学病理学部门对来自被hsv-1(f)感染、被hsv-socs4感染小鼠的小鼠肺(n=2,未收集balf)进行病理分析。
结果和讨论
hsv-socs4重组病毒的构建
我们用socs4基因插入片段重构了新的hsv-1株,其被命名为hsv-socs4,并且成功证明了表达socs4蛋白,及我们将它用于鼻内感染的小鼠以评估其对细胞因子风暴的作用。我们已经将socs4基因插入bac中,以重构hsv-socs4病毒。socs4基因的pcr产物和测序鉴定均证实了该重组体。图2a示出了来自preveiver-m02的socs4(1397bp)的pcr产物;如图2b所示,最后的pcr确认包括ul2(1492bp)、ul3(1319bp)和socs4(1397bp)的片段。
病毒感染后balf和血清中的细胞因子产量
因为被灌注到肺的细胞因子可以进入血流,所以提供了局部和系统反应之间的直接交流;因此我们在感染后的第1天、第3天和第7天收集了balf和血清样品,以检测处于细胞因子风暴的核心的几种细胞因子。
为了分析socs4蛋白在细胞因子分泌中的作用,我们分析了来自用pbs或hsv-1(f)或hsv-socs4病毒感染的小鼠在感染后第1天、第3天和第7天的balf和血清中的主要的炎症细胞因子(例如mcp-1、il-1β、il-6、tnf-α和ifn-γ)图谱。在我们检查的每个时间点,模拟小鼠都没有在balf或血清中诱导可评估数量的细胞因子。balf样品中的细胞因子产量的结果如图3所示。在第1天、第3天、第7天,与来自hsv-socs4小鼠的细胞因子的水平相比,除了两组小鼠之间仅示出可忽略的差异的在第7天的il-1β产量外,我们观察到在来自hsv-1(f)感染小鼠的所有五种细胞因子中显著更高水平。发现了hsv-1(f)感染的小鼠中的il-1β产量的上升-下降趋势曲线,并且在第7天迅速地下降至约50%(图3b)。hsv-1(f)小鼠的il-6和ifn-γ产量的趋势是增加的,而前者在第7天增加,后者在第3天增加。在第1天和第3天检测到hsv-1(f)小鼠的最高的mcp-1水平,然后其在第7天下降。在第1天也观察到最高的tnf-α水平,但在第3天下降。有趣的是发现,在被分析的时间点,除了在第7天il-6的明显增加之外,来自hsv-socs4感染小鼠的balf的细胞因子分泌是稳定的,差异很小。
为了剖析socs4蛋白对系统循环中的细胞因子产量的影响,我们收集了小鼠血清用于elisa,并且结果如图4所示。在第1天,与hsv-socs4小鼠中检测到的mcp-1的浓度相比,hsv-1(f)小鼠中检测到显著更高浓度的mcp-1,并且hsv-1(f)小鼠中的其产量连续地下降,但hsv-socs4小鼠中其仅在第7天下降;此外,在两组中的每一天的时间点,balf中的mcp-1水平都远高于血清中的mcp-1水平。hsv-1(f)和hsv-socs4小鼠的il-1β值在第1天和第3天是相似的,但在第7天检测到hsv-1(f)小鼠中的强烈上调;因此,这两组之间存在很大差异。tnf-α产生中也观察到了类似的扩增模式,除了在第1天和第3天hsv-1(f)小鼠的该值还高于hsv-socs4小鼠的值的事实之外。在第1天、第3天和第7天,从hsv-1(f)小鼠中检测到il-6的水平持续地升高,但仅在第7天发现hsv-socs4小鼠的水平增加,并且在我们测试的每一天hsv-1(f)小鼠的il-6水平都远高于hsv-socs4小鼠的il-6水平。结果表明,hsv-1(f)小鼠和hsv-socs4小鼠中在第1天和第3天ifn-γ的差异化的增加是轻微的,但在第7天变得明显。在第7天血清中ifn-γ的浓度高于balf中的ifn-γ的浓度,这证实了被活化的t细胞成为ifn-γ产生的主要来源。为了进一步评估hsv-socs4小鼠的血清中的细胞因子产量,我们在第12天测试了细胞因子水平,未观察到显著的差异(数据未示出)。简而言之,balf中被hsv-socs4感染的小鼠的细胞因子产量在各个时间点几乎是一致的,但是在血清中却示出多样性。
il-1β主要由dc和巨噬细胞产生,是驱动促炎症活性的关键细胞因子。当在感染早期出现时,它促进恢复;但在感染的后期出现时,则与导致严重的发病机理和死亡率的破坏性炎症反应相关。我们发现,在balf样品中il-1β在早期(第1天和第3天)保持在高水平,然后在第7天示出降解;相反地,在第7天hsv-1(f)小鼠中其血清中的产量增加。但是在hsv-socs4小鼠的所有时间点内,balf和血清的il-1β均维持在相当的低水平。这些数据表明,socs4蛋白可抑制早期在balf中的il-1β产量以及后期血清中的产量。
il-1和il-6的作用是协同的:il-1主要在感染的早期表达,随后il-6的表达增加。在hsv感染的部位处balf中的il-6的产生已经被报道,并且我们的结果是一致的:hsv-1(f)小鼠的balf中的il-6水平的强烈上调是明显的,随后持续的增加,尤其是在第7天,并且在具有相对较低浓度的血清中也展示了相同的增加。hsv-socs4小鼠中il-6在balf和血清中的水平均明显受到限制,伴随着在第7天的上升。在某种意义上,il-6后期的大量产生是与病毒的存在无关的,并且可能涉及促进th2反应。
另一种重要的急性反应细胞因子肿瘤坏死因子α(tnf-α)主要由巨噬细胞、肺上皮细胞和辅助t细胞产生,并可在感染后早期出现。tnf-α促进在h5n1病毒感染后的严重疾病的症状,并代表了细胞因子风暴的典型特征,并且其还参与了与hsv感染相关的免疫病理学。在感染后的早期通过病毒与巨噬细胞以及nk细胞的相互作用,tnf-α从先天免疫系统中被释放(如我们测试中的在第一天balf中的显著的高水平);以及通过激活病毒特异性cd4+或cd8+t细胞,tnf-α从适应性免疫系统中释放,如观察到在第7天的来自hsv-1(f)小鼠的血清中tnf-α水平增加。在balf和血清样品中,在每天的时间点都不能区分出被hsv-socs4感染小鼠的tnf-α水平的变化。据报道,抗tnf治疗可以减少h3n2病毒攻击后的体重减轻和疾病的严重性,表明它可能是有前途的治疗性靶标,并且我们推测受抑制的tnf产生也可以减轻由hvs感染引起的症状。
单核细胞趋化蛋白-1(mcp-1)由多种细胞类型快速产生,主要是在炎症刺激和组织损伤后的单核细胞、巨噬细胞、上皮细胞和内皮细胞。它招募单核细胞、记忆性t细胞、nk细胞和树突状细胞至组织损伤和感染的部位,并且其通常在炎症期间内在组织中被表达。我们从hsv-1(f)小鼠中观察到在早期balf中的明显更高水平的mcp-1,这说明在hsv-1(f)小鼠的balf样品中发现了更多的cd11b+细胞。
ifn-γ是具有多种功能的强效细胞因子,该功能包括:促进dc和巨噬细胞的活化,增强nk细胞的细胞毒性,以及诱导b细胞的抗体产生。在被hsv-1(f)感染的小鼠中,在第3天示出的balf中增加的ifn-γ水平可能主要是由感染早期的nk细胞产生的,并且其有助于控制病毒的复制,并且在来自两组小鼠的血清中的后期(第7天)水平升高是因为t细胞成为ifn-γ的主要来源,但在后期反应中ifn-γ的某些活性已与炎症和肺损伤有关。此外,当在肺组织中未发现病毒时,在第12天在被hsv-socs4感染的小鼠的血清样品中观察到了延长的且无差别的ifn-γ产生,这证实了通过其他细胞因子或免疫应答的特征可以在下游诱导ifn-γ,并且socs4蛋白可以抑制该血清中的这种延长的产生。促炎性细胞因子负责细胞活化和组织损伤,此外,一种细胞因子的释放可诱导新的细胞因子产生,这将反过来进一步引起细胞和器官的坏死。与被hsv-1(f)感染的小鼠相比,hsv-socs4小鼠的balf和血清中的促炎性细胞因子产量的稳定的低水平被认为与小鼠肺损伤的减轻相关。
用病毒鼻内感染后的balf和脾脏的细胞分析
为了确定balf和血清中不同的细胞因子水平是否与免疫细胞的数量有关,我们从balf中收集了细胞,并从被感染的小鼠中收集脾脏,以进行流式细胞术分析。因为在第1天和第7天细胞因子产量的差异变得明显,所以我们从第1天和第7天收集细胞以进行比较。考虑到包括巨噬细胞、嗜中性粒细胞和nk细胞的cd11b阳性细胞构成了balf中存在的主要细胞群体,我们决定分析被hsv-1(f)感染的小鼠和被hsv-socs4感染的小鼠之间cd11b+细胞的数量变化。结果表明,来自hsv-1(f)小鼠的cd11b+细胞胜过来自hsv-socs4小鼠的cd11b+细胞,并且两组中在第1天被染色的阳性细胞多于在第7天被染色的阳性细胞(图5)。来自脾脏的cd4+和cd8+细胞均被染色,并且cd62l被用作活化标记。如图6所示,两组小鼠中在第1天几乎没有双阳性细胞,但是在第7天检测到明显增加的cd8+且cd62l+细胞,并且hsv-1(f)和hsv-socs4小鼠之间的阳性细胞数量的区别是明显的。cd4+且cd62l+细胞中也观察到了相同的模式(图6)。
在第1天,hsv-1(f)小鼠的balf中发现了优势数量的cd11b+细胞(包括巨噬细胞、单核细胞、中性粒细胞和nk细胞),这是balf中较高的mcp-1水平的结果。据报道,通过迅速地分泌促炎性细胞因子的初级波,巨噬细胞在肺内对hsv的第一道防线中起着至关重要的作用,并且这解释了在第1天hsv-1(f)小鼠的balf中tnf-α、il-1β和il-6的产量增加,因为在初始反应时巨噬细胞和nk细胞是这些细胞因子的主要来源。巨噬细胞到肺和肺泡间隙中的募集是初始免疫应答的标志,脾脏中的cd4和cd8t细胞可反映适应性免疫应答的活性。cd62l通常用作t细胞的激活标记物,并且其在将淋巴细胞引导至感染和炎症的位点中起主要作用。在第1天来自两组的脾脏中均未发现活化的t细胞,但在第7天明显极大地激活细胞,hsv-1(f)小鼠的cd62l+cd4+t细胞和cd62l+cd8+t细胞数量均两倍高于hsv-socs4小鼠,这是与在第7天hsv-1(f)小鼠的血清中升高的tnf-α、il-1β、il-6和ifn-γ的水平密切相关。通过细胞因子的产生和/或被感染细胞的直接裂解,受影响的th细胞和ctl对于病毒感染的有效解决是至关重要的,然而这些相同的机制也促成肺损伤。几篇报道表明,在被流感感染小鼠中急性肺损伤(ali)与肺泡环境中的细胞因子风暴是直接相关的。
病毒滴度和肺部的病理学改变
为了评估细胞因子与病毒复制/清除的相关性,对来自被感染小鼠肺的病毒滴度进行了定量。在第1天观察到最大病毒滴度,此后其大幅下降,并且在第7天未检测到病毒,此外,在第3天hsv-1(f)小鼠和hsv-socs4小鼠之间的病毒清除率示出明显的差异(图7a)。对未收集balf的小鼠的肺进行组织病理学分析,典型的200倍照片如图7b所示。第1天,hsv-socs4小鼠的肺几乎没有病理变化。但是hsv-1(f)小鼠的肺部中展示出明显的细胞浸润,伴随局部毛细血管的轻微扩张和充血。在第7天,被hsv-socs4感染的小鼠肺中观察到一些免疫细胞的浸润以及毛细血管的轻度至中度扩张和充血,但肺泡壁的结构未破裂;hsv-1(f)小鼠的肺部中出现了严重的病理变化:肺泡壁增厚并破裂,周围严重充血,伴随充斥的免疫细胞。
结果表明,在hsv-1(f)和hsv-socs4小鼠之间在第1天肺中的病毒滴度是相似的,但在第3天下降并且检测到在两组之间的明显的差异。这种快速的病毒清除与先天免疫机制是一致的,并且由于大量的免疫细胞的激活(如在第1天通过流式细胞术和病理分析所观察到的),所以从hsv-1(f)小鼠肺中检测到病毒载量降低。通常,裂解性感染在第7天停止,因此没有测试到病毒。在被hsv-1(f)感染后,在第7天观察到小鼠肺的严重病理变化,这与小鼠的细胞因子水平和t细胞增加相一致。
在用hsv-1(f)或hsv-socs4鼻内感染后的小鼠体重及死亡率
在通过鼻内途径被感染后,每天两次对所有小鼠进行检测,持续12天,以确定体重和发病死亡率(onsetofmortality)。被hsv-1(f)感染的小鼠在第2天开始逐渐减轻其体重,并且在第7天这种减轻变得急剧,以及最后存活的小鼠在第10天减轻50%的体重(图8a),并且一致地,存活率的百分比在第7天开始下降(图8b),然后小鼠迅速死亡,而在第11天来自hsv-1(f)组的小鼠没有存活。hsv-socs4组小鼠轻微地减轻体重,在第12天普遍保持80%的体重。hsv-socs4小鼠的存活率维持在100%,这与被hsv-1(f)感染组的存活率是显著不同的。模拟小鼠没有体重减轻,也没有死亡。
由于肺损伤(可能还涉及其他器官损伤),在第7天hsv-1(f)小鼠体重开始过度减轻,第8天死亡率达到75%,第11天死亡率达到100%。另一方面,hsv-socs4小鼠仅在第12天示出轻微的体重减轻并且没有死亡。这些结果证明,控制细胞因子风暴过度释放可以维持被感染小鼠的体重、健康和存活。我们的结果示出,带有socs4蛋白插入片段的hsv抑制了细胞因子的过度产生、免疫细胞过度浸润,减轻了肺部病理损伤并降低了小鼠的死亡率。
因为对病毒感染的免疫力是多方面的且高度复杂的,并且细胞因子的诱导是一系列具有冗余和放大级联的蔓延的网络,因此干预策略应针对多种细胞因子通路。我们重组hsv-1变体的hsv-socs4株的尝试为抑制细胞因子风暴及其灾难性后果提供了有价值的工具,其可改善ohsv的临床治疗。
应当理解的是,尽管已经通过优选实施方式和可选特征具体地公开了本发明,但是本领域技术人员可以对本文所公开的本发明进行修改、改进和变形,并且这样的修改、改进和变形被认为在本公开的范围内。这里提供的材料、方法和实施例是优选的实施方式的代表,是示例性的,并且不旨在作为对本公开的范围的限制。
- 还没有人留言评论。精彩留言会获得点赞!