一种β-葡萄糖苷酶Ttbgl3及其应用的制作方法
一种
β-葡萄糖苷酶ttbgl3及其应用
技术领域
[0001]
本发明属于基因工程技术及生物医药领域,具体涉及一种β-葡萄糖苷酶ttbgl3基因的克隆、原核表达、纯化和对苷类化合物的转化。
背景技术:
[0002]
耐热毛栓菌等白腐菌在进化过程中获得了分解难降解木质纤维素的特定酶群,cazy数据库中有包括糖苷水解酶(gh)在内的不同类型的碳水化合物降解酶。糖苷水解酶(gh)有众多的家族参与了木质纤维素的降解。研究表明,丝状真菌在木质纤维素响应过程中,分布在gh1和gh3家族里的β-葡萄糖苷酶基因表达量显著上调,丝状真菌能合成和分泌β-葡萄糖苷酶等胞外酶来同时和选择性降解木质纤维素。β-葡萄糖苷酶可催化在醛糖或葡萄糖的半缩醛-oh基团与另一种非糖化合物(例如氨基醇、芳基醇或伯、仲、叔醇)的-oh基团之间形成的糖苷键的水解;这种水解作用导致的糖苷键的断裂在各种基本生物过程中的重要作用,并引起了人们的极大兴趣;如工业中异黄酮苷的水解,将无效或低效的大豆异黄酮苷转变为有效的大豆异黄酮苷元,增强其生物活性。β-葡萄糖苷酶在食品生物加工、制药和动物饲料的处理方面也有广泛的应用。
技术实现要素:
[0003]
本发明提供了一种β-葡萄糖苷酶ttbgl3,其氨基酸序列如seq id no:1所示,其核苷酸序列如seq id no:2所示,该酶来源于耐热毛栓菌(trametes trogii)s0301,本发明β-葡萄糖苷酶ttbgl3同时具有β-葡萄糖苷键和β-葡萄糖醛酸苷键的水解活性,其在ph 5-10、45-55℃下具有较好的催化活性,其还具有较好的金属离子耐受性,fe
3+
可以使ttbgl3的活性提高到119.6%;本发明β-葡萄糖苷酶ttbgl3对天麻素、七叶苷、大豆苷、黄芩苷等化合物具有较强的转化能力。
[0004]
ttbgl3全长cdna为2547bp,编码849个氨基酸多肽,预测蛋白质大小为97kda;利用ncbi数据库,将ttbgl3氨基酸序列通过blast在线比对;ttbgl3与来源于polyporus arcularius的β-葡萄糖苷酶(pabgl3,id:tfk93148.1)最接近,同源性为81.74%;另外,来源于trametes versicolor的β-葡萄糖苷酶,同源性为80.26%(tvbgl3,id:xp_008036945.1);来源于trametes coccinea的β-葡萄糖苷酶,同源性为79.34%(tcbgl3,id:osd02721.1);来源于gelatoporia subvermispora的β-葡萄糖苷酶,同源性为74.30%(gsbgl3,id:emd34486.1)
[0005]
本发明将β-葡萄糖苷酶ttbgl3基因和酶切后的pet-28b质粒连接得到含有β-葡萄糖苷酶ttbgl3基因的重组质粒;将重组质粒转化表达宿主菌roseeta(de3),采用iptg诱导培养,离心收集菌体,破碎菌体后经ni-nta预装重力柱柱纯化即得β-葡萄糖苷酶ttbgl3;将β-葡萄糖苷酶ttbgl3应用在制备稀有苷元及单糖苷化合物中,实验结果显示β-葡萄糖苷酶ttbgl3具有水解多种糖苷的能力,对天麻素、七叶苷、大豆苷和黄芩苷的具有较强的转化能力;通过同源建模和分子对接,推测β-葡萄糖苷酶ttbgl3中his85和lys467两个残基可能为
含有β-葡萄糖苷键的底物的酶结合位点,glu377和thr424可能为含有β-葡萄糖醛酸苷键的底物的酶结合位点本发明β-葡萄糖苷酶ttbgl3制备简单,具有市场推广应用的价值。
附图说明
[0006]
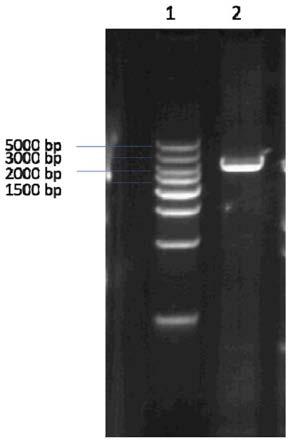
图1为实施例1本发明所述ttbgl3基因特异性的琼脂糖凝胶图;
[0007]
图2为纯化的ttbgl3的纯度鉴定结果图;图a为不同浓度梯度咪唑洗脱液纯化ttbgl3蛋白的sds-page电泳图,其中泳道m为蛋白marker(购自北京擎科新业生物技术有限公司,货号tsp021),泳道1为转化pet28b-ttbgl3重组载体的roseeta(de3)菌株添加最适iptg浓度(0.05mm)产生的蛋白;2为使用含有25mm咪唑的洗脱液洗脱的蛋白;3为使用含有50mm咪唑的洗脱液洗脱的蛋白;4为使用含有100mm咪唑的洗脱液洗脱的蛋白;5为使用含有200mm咪唑的洗脱液洗脱的蛋白;6为使用含有250mm咪唑的洗脱液洗脱的蛋白;图b为200mm咪唑洗脱液中蛋白超滤浓缩检验图,其中泳道m为蛋白marker(购自北京擎科新业生物技术有限公司,货号tsp021),泳道1为转化pet28b载体的roseeta(de3)菌株产生的蛋白;泳道2为转化pet28b-ttbgl3重组载体的roseeta(de3)菌株在不添加iptg诱导剂产生的蛋白;泳道3为转化pet28b-ttbgl3重组载体的roseeta(de3)菌株添加最适iptg浓度(0.05mm)产生的蛋白;泳道4为iptg诱导后经超声破碎收集的上清蛋白;泳道5为iptg诱导后经超声破碎收集的沉淀蛋白;泳道6为上清蛋白经ni-nta预装重力柱(生工,货号c600793)纯化,用200mm咪唑的洗脱液洗脱,并用10kda超滤管(默克,货号ufc9003)浓缩收集至0.16mg/ml的纯酶蛋白;
[0008]
图3为β-葡萄糖苷酶ttbgl3最适反应ph的测定结果图,横坐标为ph,纵坐标为相对酶活力,单位%;
[0009]
图4为β-葡萄糖苷酶ttbgl3最适反应温度的测定结果图,横坐标为温度,单位摄氏度(℃),纵坐标为相对酶活力,单位%;
[0010]
图5为ph稳定性的测定结果图,横坐标为ph,纵坐标为相对酶活力,单位%;
[0011]
图6为温度稳定性的测定结果图,横坐标为保温时间,单位分钟(min),纵坐标为相对酶活力,单位%;
[0012]
图7为糖耐受性的测定结果图,横坐标为各种单糖的浓度,单位(mm),纵坐标为相对酶活力,单位%;
[0013]
图8为实施例3本发明所述β-葡萄糖苷酶ttbgl3的定性测定结果中重组酶ttbgl3以p-npg为底物的动力学参数;
[0014]
图9为β-葡萄糖苷酶ttbgl3转化天麻素的高效液相色谱图,上图为天麻素的色谱图;下图为转化后的色谱图;图中1:天麻素;1a:4-羟基苯甲醇;
[0015]
图10为β-葡萄糖苷酶ttbgl3转化七叶苷的高效液相色谱图,上图为七叶苷的色谱图;下图为转化后的色谱图;图中2:七叶苷;2a:七叶内酯;
[0016]
图11为β-葡萄糖苷酶ttbgl3转化大豆苷的高效液相色谱图,上图为七叶苷的色谱图;下图为转化后的色谱图;图中3:大豆苷;3a:大豆素;
[0017]
图12为β-葡萄糖苷酶ttbgl3转化黄芩苷的高效液相色谱图,上图为黄芩苷的色谱图;下图为转化后的色谱图;图中4:黄芩苷;4a:黄芩素;
[0018]
图13为β-葡萄糖苷酶ttbgl3水解天麻素的转化产物的质谱分析结果图;
[0019]
图14为β-葡萄糖苷酶ttbgl3水解七叶苷的转化产物的质谱分析结果图;
[0020]
图15为β-葡萄糖苷酶ttbgl3水解大豆苷的转化产物的质谱分析结果图;
[0021]
图16是β-葡萄糖苷酶ttbgl3水解黄芩苷的转化产物的质谱分析结果图;
[0022]
图17为不同浓度的ttbgl3酶对天麻素、七叶苷、大豆苷水解效率的影响;柱形图表示用不同浓度的重组酶ttbgl3处理500μg/ml底物天麻素、七叶苷和大豆苷,反应体系中底物的剩余含量;
[0023]
图18为不同浓度ttbgl3酶对天麻素、七叶苷、大豆苷转化产物含量的影响;柱形图表示用不同浓度的重组酶ttbgl3处理500μg/ml底物天麻素、七叶苷和大豆苷,反应体系中转化产物的生成量;
[0024]
图19为不同反应时间对天麻素、七叶苷、大豆苷及其转化产物含量的影响;分别用60u/ml、5u/ml、5u/ml ttbgl3处理500μg/ml底物天麻素、七叶苷和大豆苷在50℃下对酶处理时间进行分析;实线表示底物的剩余量,虚线表示产物的生成量,其中横坐标为反应时间(min),纵坐标为底物与产物的含量,单位(μg/ml);
[0025]
图20为ttbgl3同源建模3d模型;
[0026]
图21为ttbgl3与4种苷类化合物间的分子对接;a.天麻素;b.七叶苷;c.大豆苷;d.黄芩苷。
具体实施方式
[0027]
下面结合说明书附图和实施例对本发明作出更进一步的说明,以便本领域技术人员了解本发明,但本发明的保护内容不局限于以下实施案例。本实施例中方法如无特殊说明的均按常规方法操作,所用试剂如无特殊说明的采用常规试剂或按常规方法配置的试剂。
[0028]
实施例1:β-葡萄糖苷酶ttbgl3基因的克隆及重组质粒pet28b-ttbgl3的构建1、ttbgl3基因的获得及克隆
[0029]
在28℃培养耐热毛栓菌s0301并收集菌丝体;采用trizol法提取总rna,再进行反转录获得cdna;采用常规pcr方法,以cdna为模板,通过特异性引物扩增获得ttbgl3基因片段;并去除终止密码子,pcr扩增后回收2589bp(基因长度2547bp+上下游引物携带的载体同源序列共40bp)的pcr产物;
[0030]
(1)rna的反转录
[0031]
在pcr管中配制如表1所示的反转录体系,并用移液枪混匀,将pcr管放置于pcr仪中,设置程序为50℃,15min;85℃,5s;
[0032]
表1 rna反转录反应体系
[0033]
[0034]
(2)耐热毛栓菌s0301中ttbgl3基因的扩增所用引物序列如下:
[0035]
ttbgl3-y-f:5'-caagcttgcggccgctctagaatgtcgcgcgacttcctcg-3',下划线表示xba i位点;ttbgl3-y-r:5'-gtggtggtggtggtgctcgagcaccccgttccatgtgaatc-3',下划线表示xho i位点;
[0036]
扩增体系如下:
[0037]
表2 pcr扩增反应体系
[0038][0039]
扩增条件如下:
[0040]
将反应体系混匀,先在95℃预变性5min,然后在95℃变性30s,62℃退火30s,72℃延伸3min,35个循环后,72℃延伸10min;反应完后取产物5μl,在1%琼脂糖凝胶中进行电泳分析(见图1)。
[0041]
2、pcr产物的胶回收纯化
[0042]
(1)在电泳胶槽中配制1.0%的琼脂糖凝胶;
[0043]
(2)将pcr产物点样电泳,180v电泳25min;
[0044]
(3)在紫外灯下切下含该目的片断的凝胶,转移到1.5ml的离心管中;
[0045]
(4)用生工胶回收试剂盒进行目的片段的回收,回收方法按说明书操作进行。
[0046]
3、pet28b-ttbgl3重组表达载体的构建
[0047]
(1)使用限制性核酸内切酶xba i和xho i对pet28b载体进行酶切,在37℃下反应4h,回收5317bp的酶切产物,酶切体系如下:
[0048]
表3双酶切反应体系
[0049]
pet28b质粒5μg10
×
m buffer5μlxba i1μlxho i1μl双蒸水补足至50μl;
[0050]
(2)使用非连接酶依赖型单片段快速克隆试剂盒(诺唯赞,货号c112)将双酶切线性化的pet28b载体和pcr产物进行连接;连接体系及温度如表4所示;
[0051]
表4线性化载体与基因片段连接反应体系
[0052]
线性化pet28b质粒100ng5
×
ce ii buffer4μlttbgl3基因片段300ngexnase ii2μl双蒸水补足至20μl;
[0053]
(3)转化大肠杆菌dh5α感受态细胞
[0054]
将连接产物转化大肠杆菌dh5α感受态细胞(购自北京擎科新业生物技术有限公司),筛选阳性克隆,进行序列分析,得到重组质粒pet28b-ttbgl3。
[0055]
实施例2:β-葡萄糖苷酶ttbgl3的制备
[0056]
将重组质粒pet28b-ttbgl3转化大肠杆菌roseeta(de3)菌株,在含有卡那霉素(50μg/ml)的lb平板培养基(胰蛋白胨10g/l、酵母提取物5g/l、nacl5g/l、琼脂15g/l)上经过37℃培养过夜,挑转化子到200ml的lb液体培养基中(50μg/ml卡那霉素)37℃、200rpm振荡培养至od600为0.6-0.8时,加入终浓度为0.05mmol/l异丙基β-d-硫代吡喃半乳糖苷(iptg)诱导剂,18℃培养10h,用高速冷冻离心机将培养液在4℃下,以13,000rpm离心15min,收集菌体;由于重组质粒pet28b-ttbgl3中含有his-tag标签,通过ni-nta预装重力柱(生工,货号c600793)进行纯化,得到纯化的ttbgl3;具体操作过程如下:
[0057]
(1)样品的处理
[0058]
①
将洗涤过的菌体,用1
×
binding buffer 8ml重悬,超声波破壁(150w,超声5s,间歇7s,破碎20min);
[0059]
②
破壁后,13,000g离心30min,取上清即为样品;使用p-npg底物测定破碎上清液中β-葡萄糖苷酶ttbgl3活性,结果发现在上清中有酶活反应,表明已经成功表达ttbgl3蛋白,且具有活性,可以继续后续蛋白纯化;
[0060]
(2)处理柱子
[0061]
①
取1mlni-nta预装重力柱;
[0062]
②
用5ml的pbs缓冲液洗柱子;
[0063]
③
用5ml的1
×
charge buffer洗柱子;
[0064]
④
用3ml的1
×
binding buffer洗柱子。
[0065]
(3)上样
[0066]
①
将样品加入柱子,控制流速约每分钟6滴;
[0067]
②
用3ml 1
×
binding buffer洗柱子,除去未结合的蛋白质;
[0068]
③
用10mmol/l咪唑的洗脱液洗柱子,出去杂蛋白;
[0069]
④
分别用4ml含有不同浓度梯度的咪唑洗脱液洗柱子,将目的蛋白洗脱下来;
[0070]
⑤
用5ml 1
×
strip buffer洗柱子,用含有80%无水乙醇的pbs缓冲液保存填料柱;
[0071]
通过此过程得到纯化的β-葡萄糖苷酶ttbgl3,通过sds-page电泳后染色鉴定β-葡萄糖苷酶ttbgl3的纯度,结果如图2所示;由图2a结果可见,目的蛋白通过his标签纯化后,在95kda处有细微条带,与预测的分子量吻合,使用p-npg底物测定不同浓度梯度的咪唑洗脱液中β-葡萄糖苷酶ttbgl3活性,结果发现均有酶活反应,表明已经成功纯化到ttbgl3蛋
白,纯化效果达到预期;由图2b所示,经过超滤管浓缩后分离了咪唑,并且得到浓度较高且条带单一的ttbgl3蛋白,有利于后续实验。
[0072]
实施例3:β-葡萄糖苷酶ttbgl3的酶学性质及酶活性实验
[0073]
酶活力单位(u)定义为:在测定条件下,每分钟产生1μmol p-npg所需要的酶量为1个酶活力单位;所有测定实验均进行三次平行试验,结果取平均值;
[0074]
1、酶活的测定方法
[0075]
反应体系为100μl,5μl 20mmol/l对硝基苯β-d葡萄糖苷(p-npg)中加入85μl 100mmol/l柠檬酸-磷酸氢二钠缓冲液(ph6.0),先在50℃孵育3min,再加入10μl酶液(稀释到合适的倍数)反应10min,显色后再加入1mol/l的碳酸钠溶液600μl终止反应;在405nm下测定吸光值。
[0076]
2、最适反应ph的测定
[0077]
选择不同ph值的缓冲液,包括:1mol/l柠檬酸磷酸氢二钠缓冲液(ph 2.5-7)、1mol/l tris-hcl缓冲液(ph 7-9)、1mol/l甘氨酸-naoh缓冲液(ph 9-11),50℃分别测定酶活,结果如图3所示;由图3结果可见,本发明所述β-葡萄糖苷酶ttbgl3的最适反应ph为6.0。
[0078]
3、最适反应温度的测定
[0079]
选择最适ph条件下,将酶反应体系分别在30℃、40℃、50℃、55℃、60℃、65℃、70℃、75℃、80℃下进行孵育,测定酶活性,结果如图4所示;由图4结果可见,本发明所述β-葡萄糖苷酶ttbgl3的最适反应温度为50℃。
[0080]
4、ph稳定性的测定
[0081]
将酶液在不同ph的缓冲液中孵育2h,检测残余酶活,以最高酶活作为100%,如图5所示,该酶在ph 5-10范围内酶活力能保持在90%以上。
[0082]
5、温度稳定性的测定
[0083]
将酶液分别在不同的温度(50℃、60℃)下孵育0-300min,检测剩余酶活测定ttbgl3的温度稳定性;如图6所示,在50℃下孵育5h,仍能保持85%以上的酶活性。
[0084]
6、ttbgl3对葡萄糖耐受能力测定
[0085]
使用不同浓度的葡萄糖溶液处理ttbgl3,测定剩余酶活力,以研究葡萄糖浓度对ttbgl3的酶活力影响,如图7所示,随着葡萄糖浓度的增加,ttbgl3的酶活性逐渐降低。当葡萄糖浓度增加到20mm时,酶活性降低到最大酶活性的50%。
[0086]
7、金属离子和有机溶剂对ttbgl3酶活性的影响
[0087]
在50℃、ph6.0条件下,测定了纯化蛋白对1mmol/l不同金属离子(fe
3+
、sr
2+
、nh
4+
、ba
2+
、k
+
、na
+
、co
2+
、mg
2+
、ni
2+
、ca
2+
、mn
2+
、cu
2+
、zn
2+
)和有机溶剂(10%edta、5%dmso、5%tuwen-20、1%triton x-100、10%乙醇、10%sds)的稳定性;以不添加任何金属离子或有机溶解的反应体系为对照。
[0088]
结果如表5所示,加入fecl3后,ttbgl3的活性提高到119.6%;除cuso4和znso4外,其它金属离子对ttbgl3酶活性影响不大,它们的最大酶活性都保持在90%以上;即使在cuso4和znso4的酶的活性也能保持在80%以上;在有机溶剂中,酶活力均被抑制;尤其是在含有强变性剂sds中,酶活力被完全抑制。此外,在edta、dmso、tuween-20、triton-x-100和乙醇中酶活力分别保留96.3%、82.6%、77.6%、66.4%和65.6%;因此除强变性剂sds完全抑制ttbgl3的活性外,ttbgl3在其他有机溶剂中均能保持50%以上的酶活性;
[0089]
表5金属离子和有机溶剂对ttbgl3酶活性的影响
[0090][0091]
8、ttbgl3的酶促动力学研究
[0092]
以p-npg为底物,利用lineweaver-burk双倒数法测定ttbgl3的米氏常数,以1/v为纵坐标,以1/[s]为横坐标做图;如图8所示,以p-npg为底物,在50℃和ph 6.0的最佳条件下进行反应,ttbgl3的k
m
值为1.04m m,v
max
为263.16μm/mg/min。
[0093]
实施例4:β-葡萄糖苷酶ttbgl3对天麻素、七叶苷、大豆苷及黄芩苷的转化实验
[0094]
1、β-葡萄糖苷酶ttbgl3转化苷类化合物的反应体系
[0095]
反应体系(400μl)由磷酸钠缓冲液(终浓度200mmol/l)、底物(终浓度0.5mg/ml天麻素、七叶苷、大豆苷或黄芩苷)和80u/ml纯酶组成;将反应混合物50℃孵育12h后,用400μl甲醇停止反应;以加水的样品作为阴性对照;
[0096]
2、高效液相色谱法测定苷类化合物的转化前后含量及转化效率
[0097]
仪器:高效液相色谱仪(日本岛津公司,包括在线脱气机dgu-20a3r(c)、二元泵lc-20ab、自动进样器sil-20a、柱温箱cto-20a、检测器spd-20a);
[0098]
色谱条件:色谱柱:unitary c18柱(4.6
×
250mm,5m);流动相为0.05%磷酸水溶液(a)-乙腈(b),洗脱程序和检测波长见表6;流速为1ml/min,柱温30℃;
[0099]
表6高效液相色谱法测定苷类化合物的转化的洗脱程序和检测波长
[0100][0101]
通过高效液相色谱分析,如图9-12所示,观察到天麻素、七叶苷、大豆苷和黄芩苷的反应体系在ttbgl3处理后均产生新的组分峰;
[0102]
如表7所示,ttbgl3处理后天麻素、七叶苷和大豆苷3种底物均被完全水解,转化率达100%;黄芩苷水解效率略低,约为50%;当底物浓度为500μg/ml时,转化率最高的是七叶内酯(329μg/ml),其次是对羟基苯甲醇(228.23μg/ml)和大豆素(78.92μg/ml)。可见,ttbgl3对香豆素类化合物具有很强的转化能力;
[0103]
表7 ttbgl3对天麻素、七叶苷、大豆苷及黄芩苷转化
[0104][0105]
3、uhplcesi-q-tof-ms测定苷类化合物转化后的产物组分
[0106]
取80u/ml浓度的β-葡萄糖苷酶ttbgl3在50℃、ph 6.0下处理不同苷类化合物12h后,采用uhplcesi-q-tof-ms鉴定反应产物,并采用高效液相色谱法(hplc)分析其产物含量。
[0107]
如图13-16所示,通过比较各组分的相对分子质量,发现天麻素、七叶苷和大豆苷均失去一个葡萄糖分子,分别转化为4-羟基苯甲醇(4-羟基苯甲醇m/z:125.1065[m+h]+)、七叶内酯(七叶内酯m/z:177.0145[m-h]-)和大豆素(大豆素m/z:253.0438[m-h]-)。此外,黄芩苷失去一个葡萄糖醛酸分子并转化为黄芩素(黄芩素m/z:269.2418[m-h]-)。通过对天
麻素、七叶苷、大豆苷和黄芩苷的化学结构分析,推测ttbgl3作用于天麻素、七叶皂苷和大豆苷的β-葡萄糖苷键,然后分别转化为4-羟基苯甲醇、七叶内酯和大豆素。此外,黄芩苷中β-葡萄糖醛酸键被ttbgl3断裂,生成黄芩素;ttbgl3也可以作用于β-葡萄糖醛酸键。用ttbgl3对黄酮类底物黄芩苷去糖基化,制备出新化合物黄芩素。但ttbgl3对黄芩苷的水解效率较低。当底物浓度为500μg/ml,孵育12h后,高效液相色谱法测定黄芩苷的水解率为49.35%,产物黄芩素的得率为36.22μg/ml。
[0108]
实施例5:β-葡萄糖苷酶ttbgl3水解含β-葡萄糖苷键化合物的优化实验
[0109]
通过lc-ms分析,推测β-葡萄糖苷酶ttbgl3能够水解氧苷键释放一个葡萄糖分子,以产生新的化合物,为了进一步提高ttbgl3的水解能力,选择了三种含有β-葡萄糖苷键的化合物(天麻素、七叶苷和大豆苷)对ttbgl3水解氧苷键的条件进行优化;考虑到长时间酶反应对酶的损伤,将酶反应温度控制在50℃,对酶浓度和酶反应时间进行了优化;
[0110]
采用单因素分析法进一步分析水解过程所需的最适酶浓度和转化时间;在50℃下,向反应体系中加入1u/ml、5u/ml、10u/ml、20u/ml、40u/ml、60u/ml和80u/ml的β-葡萄糖苷酶ttbgl3,确定ttbgl3的最佳用量;ttbgl3的在最适酶浓度下分别水解0.5h、1h、2h、4h、8h、12h,反应结束后加入400μl的甲醇终止反应,确定最佳反应时间;
[0111]
如图17所示,随着酶浓度的增加,底物的剩余含量越来越低,当酶浓度达到5u/ml时,七叶苷和大豆苷几乎降解完全,而当酶浓度达到60u/ml时,天麻素才被全部降解。同样的,在3种底物被水解的同时,新的化合物生成;如图17和18所示,在50℃孵育12h后,当ttbgl3的浓度为60u/ml时,天麻素全部被水解后同时产生最高含量的4-羟基苯甲醇269.15μg/ml,因此转化天麻素的最佳酶浓度为60u/ml。在七叶苷和大豆苷的水解体系中,加入5u/ml ttbgl3后,七叶内酯的生成量趋于稳定,大豆素的生成量达到最高206.99μg/ml;在产物生成的过程中,除4-羟基苯甲醇外,其余的产物七叶内酯和大豆素在超过最适酶浓度后产量反而逐渐下降,因此转化七叶苷和大豆苷的最佳酶浓度为5u/ml。
[0112]
将各底物最佳浓度的ttbgl3加入到同一反应体系中,优化酶反应时间;其中,转化天麻素的最佳酶浓度为60u/ml,转化七叶苷的最佳酶浓度为5u/ml,转化大豆苷的最佳酶浓度为5u/ml。如图19所示,当底物浓度为500μg/ml时,反应2h后,苷类化合物的水解基本完成。同时,随着反应时间的延长,12h后4-羟基苯甲醇的产率最高,达到254.04μg/ml,转化效率为0.17mm/h;1h后,大豆素的产率最高(360.01μg/ml),转化效率为1.47mm/h;2h后,七叶内酯的产率达到最高(342.03μg/ml),转化效率为0.96mm/h。底物的水解速率和产物的产率呈反比,在反应后2小时内效率最高,随后趋于平缓;
[0113]
最终确定了三种天然化合物的最佳转化参数:天麻素500μg/ml、ttbgl3 60u/ml、50℃孵育12h,对羟基苯甲醇转化率为0.17mm/h;七叶苷500μg/ml、ttbgl3 5u/ml、50℃孵育2h,七叶内酯转化率为0.96mm/h;大豆苷500μg/ml、ttbgl3 5u/ml、50℃孵育1h,大豆素转化率为1.47mm/h。
[0114]
实施例6:β-葡萄糖苷酶ttbgl3同源建模与分子对接
[0115]
1、分子对接运算过程
[0116]
(1)配体处理:在pubchem上下载catalpol结构文件,然后用mgltools1.5.6处理,通过加氢,计算电荷,合并非极性氢后保存成pdbqt文件;
[0117]
(2)受体处理:将建模后的受体蛋白用mgltools1.5.6处理蛋白结构,通过加氢、计
算电荷、合并非极性氢后保存成pdb文件,最后保存成pdbqt文件
[0118]
(3)活性位点的确定:将建模结构上传到上传到pocasa(http://altair.sci.hokudai.ac.jp/g6/service/pocasa/)预测其潜在结合位点,根据其潜在结合位点设置grid box坐标和盒子大小;
[0119]
(4)作图:使用autodockvina 1.1.2进行小分子与蛋白对接,共产生9个构象,选取affinity最佳的构象,作为最终的对接构象。并使用meastro 11.9作图分析;
[0120]
2、同源建模与分子对接
[0121]
结构模型如图20所示;采用swiss-model模型数据库在线同源建模建立了ttbgl3的三维结构;四元结构质量评价(qsqs)分数为0.4,序列重匹配度为41.73,这表明该结构模型是可靠的;
[0122]
通过分子对接程序对不同底物与ttbgl3的结合位点进行分析;用autodockvina 1.1.2软件计算蛋白与小分子的结合能量,如表8所示,对接完成后发现天麻素、七叶苷、大豆苷和黄芩苷的结合常数的绝对值分别为7.9,8.2,9.1和9.6;结合常数的绝对值越大,说明化合物结合所需的自由能越少,一般认为其绝对值大于7时,小分子化合物和蛋白两者结合的可能性越大;由图21可以看出,小分子和蛋白表面结合得较为紧密;两者之前具有较好的匹配性,为小分子结合到此处提供了可能。尽管化合物结构各异,但主要通过疏水和氢键作用和酶结合;表8总结了4种天然化合物对接情况,含有β-葡萄糖苷键的3个底物(天麻素、七叶苷和大豆苷)均含有his85、lys467酶催化位点,含有β-葡萄糖醛酸苷键的底物黄芩苷存在glu377、thr424酶催化位点;
[0123]
表8分子对接酶结合位点预测
[0124]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1