具有溶血磷脂酶及磷脂酶活力的蛋白的制作方法
1.本技术涉及磷脂酶领域,具体地,本技术涉及一种具有溶血磷脂酶及磷脂酶活力的蛋白。
背景技术:
2.在黑曲霉中已报道了有两种脂肪酶,称为lipasea和lipaseb,或者称为lipase1和lipase2。其中lipaseb的性质比较特殊,zhu shu-sen从黑曲霉a733中克隆表达了lipaseb,发现lipaseb的最适温度为15℃,最适ph在3.5-4.0,不能耐受超过40℃以上的温度,能够水解pnpc4-pnpc18链长的底物,其中pnpc12为最适底物。但lipb的比酶活极低,经过纯化后的lipb比酶活仅为6.8u/mg。
3.jiangke yang等人从黑曲霉cicc 4009中克隆表达了lipas2,虽然这个lipase2和zhu shu-sen克隆的lipaseb高度同源,仅有两个氨基酸的差别,但是性质有一定程度的差异,最适底物为pnpc8和pnpc10,最适ph小于6.5,最适温度为50℃,不能耐受超过40℃以上的温度。
4.从上所述,黑曲霉来源的liapse2或者lipaseb的实用性不强,首先比酶活极低,温度耐受性不高,适用的最适底物短链脂肪酸的甘油三脂。不如用途较为广泛的脂肪酶tl和rml,比酶活能够达到12000或8000u/mg,tl能够耐受60℃的温度保持20小时不失活,rml能水解各种长链脂肪酸的甘油三脂。
5.为了解决上述问题,需要一种新的蛋白序列,其具有溶血磷脂酶及磷脂酶活力。
6.发明概述
7.本发明在第一方面提供了一种具有溶血磷脂酶和/或磷脂酶活力的蛋白。
8.本发明提供的蛋白具有seq id no:3、4、5、6或19所示序列,或与seq id no:3、4、5、6或19所示序列具有至少90%同源性,且所述蛋白源自黑曲霉。
9.在本发明的一些具体实施方案中,所述蛋白具有与seq id no:3、4、5、6或19所示序列至少95%,至少96%,至少97%,至少98%,或至少99%的同源性。
10.第二方面,本发明还提供了一种多核苷酸序列。
11.本发明提供的多核苷酸序列选自:(1)编码第一方面的多肽的多核苷酸;(2)与(1)所述的多核苷酸序列互补的多核苷酸;和(3)(1)或(2)所述的多核苷酸的长10-40个碱基的片段。在本发明的一些具体实施方案中,所述多核苷酸的序列如seq id no:7、8、9、10或20所示。
12.第三方面,本发明还提供了一种多核苷酸构建物。
13.本发明提供的多核苷酸构建物含有第二方面的多核苷酸序列。
14.在本发明的一些具体实施方案中,所述多核苷酸构建物是表达载体或克隆载体。
15.第四方面,本发明还提供了一种宿主细胞。
16.本发明提供的宿主细胞:(1)表达第一方面的多肽;和/或(2)含有第二方面的多核苷酸序列或第三方面的多核苷酸构建物。
17.在本发明的一些具体实施方案中,所述宿主细胞选自原核或真核微生物。
18.在本发明的一些具体实施方案中,所述宿主细胞优选选自黑曲霉、毕赤酵母、大肠杆菌、芽孢杆菌属、里氏木霉和/或米曲霉。
19.第五方面,本发明还提供了一种包含第一方面的蛋白的组合物。
20.在本发明的一些具体实施方案中,所述组合物为酶组合物。在本发明的一些具体实施方案中,所述酶组合物还包括磷脂酶a1、磷脂酶a2、磷脂酶b、磷脂酶c、淀粉酶、脂肪酶、蛋白酶、纤维素酶中的一种或多种。
21.在本发明的一些具体实施方案中,所述组合物为第四方面的宿主细胞的发酵表达物,例如,发酵液、发酵浓缩液、发酵上清液、发酵液制备的酶制剂。
22.第六方面,本发明还提供了一种组合物,其含有第一方面的蛋白和任选的辅料。
23.在本发明的一些具体实施方案中,所述辅料为选自活性炭、氧化铝、硅藻土、多孔陶瓷、多孔玻璃的吸附材料。
24.第七方面,本发明提供了制备第一方面的蛋白的方法,包括发酵第四方面的宿主细胞的步骤。
25.第八方面,本发明还提供了第一方面的多肽、第二方面的多核苷酸序列、第三方面的核酸构建体、第四方面的宿主细胞、第五方面的组合物或第六方面的组合物在植物油脱胶、面包面团改性、淀粉水解产物处理、溶血磷脂酶制备和/或l-α-甘油磷酸胆碱中的应用。
附图说明
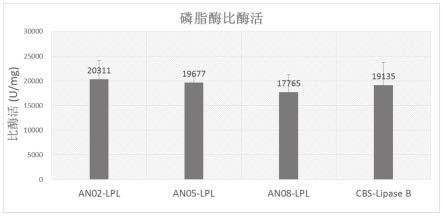
26.图1显示了an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb磷脂酶a1比酶活。
27.图2显示了an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb溶血磷脂酶比酶活。
28.图3显示了an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb脂肪酶比酶活。
29.图4显示了an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb脱胶测试结果。
30.发明详细描述
31.虽然本技术含有许多细节,但这些不应被解释为对发明或要求保护的任何范围的限制,而应被解释为可对特定发明的特定实施方案具有特异性的特征的描述。在本技术的单独实施方案中描述的某些特征也可以在单个实施方案中组合实现。相反地,在单个实施方案中描述的各种特征也可以在多个实施方案中单独地或以任何合适的子组合来实现。此外,尽管特征可以在上文中被描述为以某些组合起作用并且甚至最初如此被要求保护,但是来自所要求保护的组合的一个或多个特征可以在一些情况下从组合中删除,并且所要求保护的组合可以涉及子组合或子组合的变型。
32.除非另外说明,本文中的术语的含义与本领域技术人员通常理解的含义相同,例如,涉及原料和产物、操作步骤、工艺参数、使用设备和工具以及数值单位中的术语。
33.在本文中,词语“包括”和“包含”表示开放式,也可以是封闭式。例如,所述“包括”或“包含”可以表示还可以包括或包含没有列出的其他组分或步骤或其他要素,也可以仅包括或包含列出的组分或步骤或其他要素。
34.在本文中,术语“约”(例如,在组分含量和处理参数中)以本领域技术人员通常能够理解的含义来解释。一般情况下,术语“约”可以理解为给定数值的正负5%范围内的任意数值,例如,约x可以代表95%x至105%x的范围中的任意数值。
35.还应当理解,本文中给出的具体数值(例如,在组分含量、温度和处理时间中)不仅可作为单独的数值理解,还应当认为提供了某一范围的端点值,并且可以相互组合提供其他范围。例如,当公开了处理可以进行30分钟或180分钟的时候,也相应地公开了处理可以进行30分钟至180分钟。此外,本文给出的具体数值还可以理解为在所有情况下被术语“约”修饰。因此,除非有相反规定,本技术所记载的数值是可以根据要求改变的近似值。例如,处理的时间为30分钟可以被理解为处理的时间为约30分钟,处理的时间为30分钟至180分钟可以被理解为处理的时间为约30分钟至约180分钟或约30分钟至180分钟。
36.第一方面,本发明提供了一种蛋白,其具有溶血磷脂酶和/或磷脂酶活力,因此,可单独作为磷脂酶和/或溶血磷脂酶使用,而且,还可以用于同时需要磷脂酶和溶血磷脂酶的应用,例如,可以应用于l-α-甘油磷酸胆碱的制备。
37.本发明提供的蛋白具有seq id no:3、4、5、6或19所示序列,或与seq id no:3、4、5、6或19所示序列具有至少90%同源性,且所述蛋白源自黑曲霉。
38.在本发明的一些具体实施方案中,所述蛋白具有与seq id no:3、4、5、6或19所示序列至少91%、92%、93%、94%、95%的同源性。
39.在本发明的一些具体实施方案中,所述蛋白具有与seq id no:3、4、5、6或19所示序列至少95%,至少96%,至少97%,至少98%,或至少99%的同源性。
40.在本发明的一些具体实施方案中,部分蛋白具有较高的热稳定性,在本发明的这些实施方案中,使用的蛋白为具有seq id no:19所示序列,或与seq id no:19所示序列具有至少90%同源性,且所述蛋白源自黑曲霉。
41.本发明中,所述蛋白可以是“分离的”蛋白。本文中,“分离的”意指在自然界中不存在的一种形式或物质。分离的物质的非限制性例子包括任何非天然存在的物质和至少部分地从与其在自然界中相关联的一种或多种或全部天然存在的组分中除去的任何物质,包括但不限于任何酶、变体、核酸、蛋白质、肽或辅因子。
42.本发明还包括在seq id no:3、4、5、6或19的基础上具有一个或多个(通常为1-10个,例如1、2、3、4、5、6、7、8、9或10个)氨基酸突变(缺失、插入和/或取代)、同时保留了seq id no:3、4、5、6或19所示氨基酸序列的酶活性的多肽。在某些实施方案中,所述氨基酸突变是在seq id no:3、4、5、6或19的c末端和/或n末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为8以内)氨基酸的多肽。
43.本领域技术人员公知,在基因克隆操作中,常常需要设计合适的酶切位点,这势必在所表达的蛋白末端引入了一个或多个不相干的残基,而这并不影响目的蛋白的活性。又如为了构建融合蛋白、促进重组蛋白的表达、获得自动分泌到宿主细胞外的重组蛋白、或利于重组蛋白的纯化,常常需要将一些氨基酸序列添加至重组蛋白的n末端、c末端或该蛋白内的其它合适区域内,这类氨基酸序列包括但不限于接头肽、信号肽、前导肽、末端延伸、谷胱甘肽s-转移酶(gst)、麦芽糖e结合蛋白、蛋白a、标签(如6his或flag)、或合适的蛋白水解酶位点等。应理解,这些氨基酸序列的存在不会影响到所得多肽的活性。因此,本发明也包括在本发明多肽的c末端和/或n末端或其蛋白内合适的区域内具有一个或数个利于该多肽表达载体的构建、该多肽的表达和/或纯化的氨基酸的多肽,这些多肽仍具有本文所述的酶活性。
44.本发明中,序列同源性用于描述两个氨基酸序列之间或两个核苷酸序列之间的相
关性。可使用本领域周知的方法来计算序列相同性。例如,可使用emboss包(emboss:欧洲分子生物学开放软件套件,赖斯等,2000,遗传学趋势,16:276-277)的尼德尔(needle)程序中所实施的尼德尔曼-翁施(needleman-wunsch)算法(needleman和wunsch,1970,分子生物学杂志,48:443-453)来确定两个氨基酸序列之间的序列同源性,或者,可使用ncbi上的blastp来计算两条氨基酸序列之间的序列同源性。
45.第二方面,本发明还提供了一种多核苷酸序列。
46.本发明提供的多核苷酸序列选自:(1)编码第一方面的多肽的多核苷酸;(2)与(1)所述的多核苷酸序列互补的多核苷酸;和(3)(1)或(2)所述的多核苷酸的长10-40个碱基的片段。
47.在本发明的一些具体实施方案中,所述多核苷酸的序列如seq id no:7、8、9、10或20所示。
48.在本发明的一些实施例中,本领域的技术人员可以根据所使用的表达质粒和/或宿主,对多核苷酸序列进行密码子偏爱性优化。
49.本发明中,编码本发明多肽的序列包括:只编码成熟多肽的编码序列;成熟多肽的编码序列和各种附加编码序列;成熟多肽的编码序列(和任选的附加编码序列)以及非编码序列。应当理解,由于真核生物本身所具有的性质,在本发明的多核苷酸序列中加入内含子序列,也是可以的,只要真核生物表达的酶属于本发明的第一方面的酶。
50.第三方面,本发明还提供了一种多核苷酸构建物。
51.本发明提供的多核苷酸构建物含有第二方面的多核苷酸序列。
52.本发明也涉及包括与指导编码序列在合适宿主细胞中,在与该调控序列相适合的条件下表达的一个或多个调控序列可操作连接的本发明的分离多核苷酸的核酸构建体。术语“可操作连接”指调控序列位于适当的位置从而能控制、指导感兴趣的多核苷酸序列的表达。编码本发明多肽的多核苷酸可以多种方式被操作以保证该多肽的表达。
53.调控序列可以是合适的启动子序列,为用于表达编码本发明多肽的多核苷酸的宿主细胞识别的核苷酸序列。启动子序列包含接到多肽表达的转录调控序列。启动子可以是在所选择的宿主细胞中显示转录活性的任何核苷酸序列,包括突变的、截短的和杂合启动子,并且可以从编码与该宿主细胞同源或异源的胞外或胞内多肽的基因获得。
54.用于指导本发明的核酸构建体,特别是在细菌宿主细胞中转录的合适启动子的实例是从噬菌体t7启动子、大肠杆菌lac操纵子、天蓝色链霉菌(streptomyces coelicolor)琼脂糖酶基因、枯草芽孢杆菌果聚糖蔗糖酶基因、地衣芽孢杆菌α-淀粉酶基因、解淀粉芽孢杆菌α-淀粉酶基因、地衣芽孢杆菌青霉素酶基因等中获得的启动子序列。
55.用于指导本发明的核酸构建体在丝状真菌宿主细胞正转录的合适启动子的实例是从米曲霉taka淀粉酶、米黑根毛霉(rhizomucor miehei)天冬氨酸蛋白酶、黑曲霉中性α-淀粉酶、黑曲霉酸稳定的α-淀粉酶、黑曲霉或泡盛曲霉糖化酶(glaa)、里氏木霉纤维二糖水解酶i、氏木霉纤维二糖水解酶ii、米曲霉碱性蛋白酶、米曲霉磷酸丙糖异构酶、里氏木霉葡聚糖内切酶等基因获得的启动子及其突变、截短和杂合(hybrid)启动子。
56.在酵母宿主中,有用的启动子可获得自酿酒酵母烯醇酶(eno-1)、酿酒酵母半乳糖激酶(gal1)、酿酒酵母乙醇脱氢酶、3-磷酸甘油醛脱氢酶、酿酒酵母磷酸丙糖异构酶、酿酒酵母3-磷酸甘油酸激酶的基因、巴斯德毕赤酵母醇氧化酶基因。用于酵母宿主细胞的其它
有用启动子由romanos et al.,1992,yeast8:423-488描述。
57.调控序列也可以是合适的转录终止子序列,为宿主细胞识别以终止转录的序列。终止子序列与编码该多肽的核苷酸序列的3
′
末端可操作连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。
58.用于细菌宿主的优选终止子可以是来自t7噬菌体的终止子。
59.用于丝状真菌宿主细胞的优选终止子获得自米曲霉taka淀粉酶、黑曲霉葡萄糖淀粉酶、构巢曲霉邻氨基苯甲酸合酶、黑曲霉α-葡萄糖苷酶的基因。
60.用于酵母宿主细胞的优选终止子获得自酿酒酵母烯醇酶、酿酒酵母细胞色素c、酿酒酵母甘油醛-3-磷酸脱氢酶、巴斯德毕赤酵母醇氧化酶基因等。
61.调控序列也可以是合适的前导序列,对宿主细胞翻译重要的mrna的非翻译区。签到序列与编码该多肽的核苷酸序列的5
′
末端可操作连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。
62.调控序列也可以是编码与多肽的氨基酸末端连接的氨基酸序列并且指导该编码的多肽进入细胞分泌途径的信号肽编码区。核苷酸序列编码序列的5
′
端可固有地包含天然连接具有编码分泌多肽的编码区节段的翻译阅读框的信号肽编码区。备选地,编码序列的5
′
端可包含与该编码区外源的信号肽编码区。当编码序列非天然地包含信号肽编码区时,可能需要外来的信号肽编码区。备选地,外来的信号肽编码区可简单地替换天然的信号肽编码区以增强多肽的分泌。然而,指导表达的多肽进入选择的宿主细胞的分泌途径的任何信号肽编码区,即分泌进入培养基,都可用于本发明。
63.在某些实施方案中,本发明的核酸构建体为表达框。术语“表达框”是指表达一个基因所需的完整元件,包括启动子、基因编码序列、polya加尾信号序列。
64.在本发明的一些具体实施方案中,所述多核苷酸构建物是表达载体或克隆载体。
65.表达载体可以是能够方便地经受重组dna方法并且可导致感兴趣的核苷酸序列表达的任何载体(如质粒或病毒)。克隆载体通常在引入到宿主细胞中后能在宿主细胞中大量繁殖。
66.载体的选择一般取决于载体与其中被导入该载体的宿主细胞的相容性。该载体可以是线性或闭合的环形质粒。
67.载体可以是自主复制的载体,即作为染色体外实体存在,其复制不依赖于染色体复制的载体,例如质粒、染色体外元件、微型染色体或人工染色体。载体可包含用于保证自我复制的任何方式。备选地,载体可以是当被导入宿主细胞时,整合到基因组中并且与其已经被整合进入的染色体一起复制的载体。此外,可使用一起包含将被导入宿主细胞基因组的总dna的单个载体或质粒或两个或多个载体或质粒,或转座子。
68.本发明的载体优选包含一个或多个容许容易选择转化、转染、转导等细胞的可选择标记。可选择的标记是基因,其产物提供对抗生素或病毒的抗性、对重金属的抗性、原养型至营养缺陷型等。
69.本发明的载体优选包含容许该载体整合进入宿主细胞基因组或该载体在细胞中独立于基因组自主个复制的元件。
70.一个以上拷贝的本发明的多核苷酸可被插入宿主细胞以增加该基因产物的产量。多核苷酸拷贝数的增加可通过将至少一个附加拷贝的序列整合进入宿主细胞基因组或通
过包括可扩增的选择标记基因和该多核苷酸来获得,其中包含扩增拷贝的选择标记基因并且由此包含附加拷贝多核苷酸的细胞可通过在存在适当的选择剂时培养该细胞来筛选。
71.本发明的载体优选包含人工合成的一段序列,含有多个限制内切酶识别位点,能为外源dna提供多种可插入的位置或插入方案。
72.第四方面,本发明还提供了一种宿主细胞。
73.本发明提供的宿主细胞:(1)表达第一方面的多肽;和/或(2)含有第二方面的多核苷酸序列或第三方面的多核苷酸构建物。
74.在本发明的一些具体实施方案中,所述宿主细胞选自原核或真核微生物。
75.宿主细胞可以是单细胞微生物或非单细胞微生物。单细胞微生物例如革兰氏阳性细菌,包括但不限于芽孢杆菌细胞,例如,嗜碱芽孢杆菌、解淀粉芽孢杆菌、短芽孢杆菌、巨大芽孢杆菌、枯草芽孢杆菌、地衣芽孢杆菌、凝结芽孢杆菌、嗜热脂肪芽孢杆菌和苏云金芽孢杆菌等;或链霉菌细胞,例如钱青紫链霉菌;或革兰氏阴性细菌,例如大肠杆菌和假单胞菌属。在优选的方面,细菌宿主是枯草芽孢杆菌、大肠杆菌、地衣芽孢杆菌、嗜热脂肪芽孢杆菌和大肠杆菌细胞。
76.宿主细胞也可以是真核生物,例如哺乳动物、昆虫、植物、酵母或真菌细胞。在优选的方面,宿主细胞是真核细胞,如在此使用的“真核”包括子囊菌门(ascomycota)、担子菌门(basidiomycota)、壶菌门(chytridiomycota)、接合菌门(zygomycota)以及卵菌门等。
77.在更优选的方面,宿主细胞是子囊菌门的细胞如酵母属(saccharomyces)、毕赤酵母属(pichia)、耶氏酵母属(yarrowia)、假丝酵母属(candida)以及komagataella属等。
78.在本发明的一些具体实施方案中,所述宿主细胞选自黑曲霉、毕赤酵母、大肠杆菌、芽孢杆菌属、里氏木霉和/或米曲霉。
79.第五方面,本发明还提供了一种包含第一方面的蛋白的组合物。
80.在本发明的一些具体实施方案中,所述组合物为酶组合物。在本发明的一些具体实施方案中,所述酶组合物还包括磷脂酶a1、磷脂酶a2、磷脂酶b、磷脂酶c、淀粉酶、脂肪酶、蛋白酶、纤维素酶中的一种或多种。
81.在本发明的一些具体实施方案中,所述组合物为第四方面的宿主细胞的发酵表达物,例如,发酵液、发酵浓缩液、发酵上清液、发酵液制备的酶制剂。
82.第六方面,本发明还提供了一种组合物,其含有第一方面的蛋白和任选的辅料。
83.在本发明的一些具体实施方案中,所述辅料为选自活性炭、氧化铝、硅藻土、多孔陶瓷、多孔玻璃的吸附材料。
84.第七方面,本发明提供了制备第一方面的蛋白的方法,包括发酵第四方面的宿主细胞的步骤。
85.第八方面,本发明还提供了第一方面的多肽、第二方面的多核苷酸序列、第三方面的核酸构建体、第四方面的宿主细胞、第五方面的组合物或第六方面的组合物在植物油脱胶、面包面团改性、淀粉水解产物处理、溶血磷脂酶制备和/或l-α-甘油磷酸胆碱中的应用。
86.本发明磷脂酶可以被用于降低食用油的磷脂含量。这一方法可以被用于纯化任何含有磷脂的食用油,如植物油(如大豆油、菜籽油、向日葵籽油)。通常,在开始用磷脂酶处理之时,油中含有50-1000ppm以磷脂形式的磷,处理可以将磷值降到低于5-10ppm。磷脂酶处理是通过散布磷脂酶的水性溶液来完成,优选的为平均直径低于10um的小滴形式。优选的,
水量占油重量的0.5-5%。可以加入乳化剂。可以使用机械搅动来保持乳化。磷脂酶处理可以在约3.5到5的ph范围进行,以最大化酶的性能,或者可以使用约1.5到3(如2-3)的ph范围,以抑制甘油三脂的碱水解(皂化)。可以通过加入柠檬酸、柠檬酸缓冲液或盐酸来调节pho适合的温度通常是30-70℃(尤其是30-45℃,如35-40℃)反应时间通常是1-12个时(如2-6小时)。合适的酶剂量通常是0.1-10mg每升(如0.5-5mg每升)。磷脂酶处理可以分批进行(如在一搅拌的容器中),或者也可以连续进行(如在一系列搅拌的反应容器内)。磷脂酶处理之后是对水相和油相的分离。此分离可以通过传统的方式完成(如离心)。水相中含有磷脂酶,并且这些酶可以被重复利用以提高此过程的经济性。可以使用本领域已知的各种方法完成此处理。
87.烘焙产品是由生面团制成,生面团通常由基本成分面粉、水和可选的盐制成。依赖于烘焙产品的不同,其它选择性的成分包括糖、调味品等等。对于发酵产品,面包酵母主要在化学发酵系统(如酸(产生酸的化合物)和重碳酸盐的组合)之后被使用。为了提高生面团的操作特性和烘焙产品的最终性质,一直在努力发展有助于提高性质的手段。需要被提高的生面团性质包括可加工性、气体保留能力等等。可以被提高的烘焙产品的性质包括:面包体积、面包皮脆度、面包心质地和柔软性、味道和香味以及保质期。目前具有的加工辅助手段可以被分为两组:化学添加剂和酶。
88.改善性质的化学添加剂包括氧化剂(如抗坏血酸、澳酸盐、和偶氮碳酸盐)、还原剂(如l-半胱氨酸和谷胱甘肽)、作为生面团调节物的乳化剂(如二乙酰酒石酸单/双甘油酯(datem)、硬脂酰乳酸钠(ssl)或硬脂酰乳酸钙(csl)),或作为面包心软化剂的乳化剂(如单硬脂酸甘油脂(gms)等)、脂肪材料(如甘油三脂(脂肪)或卵磷脂及其它)。
89.目前趋向于用酶来代替化学添加剂。后者被认为是更天然的化合物,因此更加被消费者接受。合适的酶可以选自淀粉酶、阿拉伯糖木聚糖及其它半纤维素降解酶、纤维素降解酶、氧化酶、脂肪解酶和蛋白质酶。
90.本发明也涉及制备生面团或烘焙产品的方法,该方法包括向生面团中整合入有效量的本发明磷脂酶,相对于没有加入多肽的生面团或烘焙产品而言,该磷脂酶改善生面团或来自此生面团的烘焙产品的一种或多种性质。
91.本发明磷脂酶可以被用于水性糖溶液或浆状物的脱胶,以提高其过滤性,尤其是淀粉水解产物,特别是难于过滤并产生浑浊滤液的小麦淀粉水解产物。此处理可以使用本领域所熟知的方法完成。
92.本发明磷脂酶可以被用于任何需要水解磷脂或获得它的特异切割产物的应用。例如应用本发明磷脂酶可以产生溶血磷脂、二酰甘油、胆碱-或乙醇胺磷酸、溶血磷脂酰胆碱、溶血磷脂酰乙醇胺和各种磷脂酸。本发明磷脂酶被优选用于最佳活力ph。
实施例
93.下面结合具体实施例,进一步阐述本技术。应理解,这些实施例仅用于说明本技术而不用于限制本技术的范围。下述实施例中未注明具体条件的实验方法,通常按照常规条件或按照制造商建议的条件。除非另外说明,否则百分比按质量计算。除非另行定义,文中所使用的所有专业术语或科学用语与本领域熟练人员所熟悉的意义相同。此外,任何与所记载内容相似或均等的方法及材料皆可应用于本技术中。本文中所述的较佳的实施方法与
材料仅作示范之用。
94.本技术的实施例中,使用的
95.黑曲霉菌株gim 3.24(an02),gim 3.150(an03),gim 3.488(an04),gim 3.452(an07),gim3.564(an05)购自广东省微生物菌种保藏中心,
96.cicc40273(an08)购自中国工业微生物菌种保藏管理中心;
97.黑曲霉as3.795菌株购自中科院微生物所。
98.paop-eno载体由发明人参考《分子克隆实验指南》(第三版,纽约,冷泉港实验室出版社,new york:cold spring harbor laboratory press,1989)的方法构建,具体过程如下:
99.用sphi和hindiii酶切位点将从生工生物工程(上海)股份有限公司全基因合成获得的rml基因(seq id no:23,带米曲霉α-淀粉酶信号肽(ncbi序列号:xm_001821384.2,1-63bp序列))插入到以米曲霉烯醇化酶启动子(ncbi序列号:d63941.1,215-734bp;含12拷贝增强子序列(seq id no:24))和黑曲霉糖化酶终止子(ncbi序列号:af214480.1,其中终止子序列部分)的表达框中,将整个表达框以bglii和xhoi插入到克隆载体psp72的多克隆位点,最后将米曲霉来源的pyrg表达基因(ncbi序列号:ab017705.1)以xhoi酶切位点插入到载体上,从而构建好载体paop-eno。
100.mini beadbeater购自美国biospec;
101.dna polymerase购自takara,货号:r010a ;
102.mighty ta-cloning reagent set for试剂盒购自takara,货号:6019;
103.本发明中,所使用的,
104.黑曲霉发酵培养基配方为:
105.2%葡萄糖,10%麦芽糖,7%柠檬酸钠,1.5%硫酸铵,4%tryptic soy broth,0.1%磷酸二氢钠,0.1%硫酸镁,0.07%tween 80,微量元素(ki 0.83g/l,h3bo
3 6.2g/l,mnso4·
4h2o 22.3g/l,znso4·
7h2o 8.6g/l,na2moo4·
2h2o 0.25g/l,cuso4·
5h2o 0.025g/l,cocl2·
6h2o 0.025g/l按1/1000比例添加;feso4·
7h2o 2.78g/l,na2·
edta 3.73g/l按1/100比例添加)。
106.lysis buffer配方为:100mm tris-hcl ph 8.0;50mm na
·
edta;1%sds。
107.bmmy-大豆磷脂培养基:
108.组分a ,bmmy固体培养基:1%yeast extract,2%peptone,100mm柠檬酸-柠檬酸钠缓冲液,ph 6.6,1.34%ynb,4x 10-5%biotin(倒平板前加),2%methanol(倒平板前加),2%琼脂溶于250ml去离子水中。
109.组分b,大豆磷脂底物溶液250ml:配4%大豆磷脂,用高速匀浆机8000rpm乳化3min,暂停1min后再乳化3min,制备底物溶液。
110.灭菌后将a和b组分混合,加入10ml甲醇,倒平板。
111.glass beads购自:美国biospec。
112.实施例1:黑曲霉lipaseb基因克隆
113.将黑曲霉菌株gim 3.24(an02),gim 3.150(an03),gim 3.452(an07),gim3.488(an04),gim3.564(an05),cicc40273(an08)用黑曲霉发酵培养基30℃培养24小时。取发酵
培养物,4000rpm离心5min,取菌体。用700ul lysis buffer重悬,转移至冻存管中,加入300ul glass beads,上mini beadbeater振动40s,12000rpm离心10min,取600ul上清加入275ul 7m醋酸铵,65℃水浴10min,冰浴5min,加入等体积的酚氯仿异戊醇(体积比,酚:氯仿:异戊醇=24:25:1),充分涡旋,12000rpm离心5min,取上清加入等体积的氯仿,充分涡旋,12000rpm离心5min,取上清加入2倍体积的无水乙醇,-80℃放置20min,12000rpm离心10min,得到白色dna沉淀,用70%乙醇洗2次,待乙醇充分挥发后加入无菌水溶解dna,分别获得gim 3.24(an02),gim 3.150(an03),gim 3.452(an07),gim 3.488(an04),gim3.564(an05),cicc40273(an08)基因组。
114.根据ncbi中的黑曲霉cbs513.88的lipaseb的基因序列设计引物lpl-1/lpl-2,序列如下:
115.lpl-1:5
’‑
atgtttctccgcagggaatt-3’(seq id no:1);
116.lpl-2:5
’‑
ctacgagcattcactaatgt-3’(seq id no:2)。
117.以lpl-1/lpl-2为引物对,利用dna polymerase克隆黑曲霉菌株gim 3.24(an02),gim 3.150(an03),gim 3.452(an07),gim 3.488(an04),gim3.564(an05),cicc40273(an08)的lpl dna,用mighty ta-cloning reagent set for试剂盒进行ta克隆后转化dh5a大肠杆菌,送生工生物工程有限公司进行dna测序,最终得到gim 3.24(an02),gim 3.150(an03),gim 3.452(an07),gim 3.488(an04),gim3.564(an05),cicc40273(an08)的lpl dna序列,结果显示,gim 3.24(an02)lpl、gim 3.150(an03)lpl、gim 3.452(an07)lpl序列完全相同,其序列如seq id no:3所示,gim3.564(an05)lpl序列如seq id no:4所示,cicc40273(an08)lpl序列如seq id no:5所示。gim 3.488(an04)的lpl dna序列与cbs513.88的lipaseb的dna序列相同,其序列如seq id no:6所示。
118.根据cbs513.88的lipaseb的dna序列的内含子序列,找出gim 3.24(an02),gim 3.150(an03),gim 3.452(an07),gim3.564(an05),cicc40273(an08)的内含子序列,去除后翻译成氨基酸序列情况如下:
119.gim 3.24(an02)lpl、gim 3.150(an03)lpl、gim 3.452(an07)lpl序列如seq id no:7所示,gim3.564(an05)lpl序列如seq id no:8所示,cicc40273(an08)lpl序列如seq id no:9所示,gim 3.488(an04)lpl即cbs513.88的lipaseb氨基酸序列如seq id no:10所示。
120.实施例2:an02-lpl,an05-lpl,an08-lpl和cbs-lipaseb的磷脂酶a1活力
121.设计并合成引物lpl-11c、lpl-12c和lpl-2c:
122.lpl-11c:
123.cccaagctttttccaactcaatttacctctatccacacttctcttccttcctcaatcctctatatacacaactggggatccttcaccatgatggtcgcgtggtggtct(seq id no:11);
124.lpl-12c:
125.atgatggtcgcgtggtggtctctatttctgtacggccttcaggtcgcggcacctgctttggctgctcccgcacctgctccgat(seq id no:12);
126.lpl-2c:acatgcatgcctaggagcactcggag(seq id no:13)。
127.以an02,an04,an05,an08的基因组为模板,先以lpl-12c和lpl-2c为引物对进行
pcr,再以此pcr产物作为模板,以lpl-11c和lpl-2c进行pcr,获得an02-lpl,an04lpl(即cbs-lipaseb),an05-lpl和an08-lpl的dna序列(在基因序列的5’端均增加了tpi 5
’‑
utr(seq id no:14)序列)和,amylase signal peptide(seq id no:15)序列pcr产物用hindiii和sphi进行酶切,通过hindiii和sphi酶切位点连入paop-eno载体中,测序正确的克隆,转化黑曲霉as3.795菌株,得到的菌株分别命名为an02l,an04l,an05-l和an08-l。
128.转化方法如下:
129.将培养在pda固体培养基(购自bd公司,货号bd 213400)的黑曲霉菌株孢子用孢子洗液洗脱,将洗脱下来的孢子涡旋震荡1min后,通过mircloth过滤,制备成均一的孢子悬液;接种1
×
107个孢子悬液至发酵培养基(2%葡萄糖,6%麦芽糖,7%柠檬酸钠,1.5%硫酸铵,4%tryptic soy broth,0.1%磷酸二氢钠,0.1%硫酸镁,0.07%tween 80,微量元素(ki 0.83g/l,h3bo
3 6.2g/l,mnso4.4h2o 22.3g/l,znso4.7h2o 8.6g/l,na2moo4.2h2o 0.25g/l,cuso4.5h2o 0.025g/l,cocl2.6h2o 0.025g/l按1/1000比例添加;feso4.7h2o 2.78g/l,na2.edta 3.73g/l按1/100比例添加))中,28℃,200rpm,培养42-48h;用灭菌mircloth(购自milliproe)过滤收集长出的菌丝;将收集的菌丝体用灭菌的渗透压稳定剂(0.6mol/l mgso4)冲洗三次,压干;菌丝转移至100ml三角瓶中,每0.8g菌丝体重悬在10ml的酶解液(1%纤维素酶(购自sigma,货号c1184-25ku)、1%溶壁酶(购自sigema,货号l1412-25g)、0.1%蜗牛酶(购自生工,货号a600870-0005))中,分散;30℃,60rpm,60-90min(30min后每隔10min观察一次);用酶酶解的菌丝用mircloth过滤,再用0.6mol/l mgso4冲洗,收集滤液;4℃,1000g离心10min,弃上清;5ml预冷1.0mol/l山梨醇溶液重悬原生质体沉淀,800g,4℃离心10min,弃上清;原生质体重悬于1ml预冷1.0mol/l山梨醇,冰浴待用。取原生质体离心,用预冷的stc(1.0m sorbitol,50mm cacl2,50mm tris-hcl,ph=7.5)溶液调整至1
×
107个/ml;向200μl原生质体悬液中,加入5μg dna和50μl ptc(40%peg4000,50mm cacl2,50mm tris-hcl,ph=7.5)溶液,轻轻拍打混匀,冰浴30min;加0.2ml ptc溶液,混匀,再加入0.8ml ptc溶液,混匀,室温30min;将上述混合液加入到5ml再生培养基(0.2%kh2po4,0.1%kcl,0.05%mgso4·
7h2o,0.005%feso4·
7h2o,1m蔗糖,10mm乙酰胺,20mm氯化铯,0.6%琼脂)中,混匀;铺于再生培养基(成分同上,1.5%琼脂)上,28℃,培养3天以上。
130.将再生培养基上长出的阳性克隆涂布于3%的pda固体培养基(购自bd公司,货号bd 213400)上,28℃培养3天左右带有大量孢子形成,将洗脱下来的孢子涡旋震荡1min后,通过mircloth过滤,制备成均一的孢子悬液;接种1
×
107个孢子悬液至发酵培养基(2%葡萄糖,10%麦芽糖,7%柠檬酸钠,1.5%硫酸铵,4%tryptic soy broth,0.1%磷酸二氢钠,0.1%硫酸镁,0.07%tween 80,微量元素)中,28℃,200rpm,培养8天,进行酶活测定。
131.酶活测定方法如下:
132.9ml底物:5ml 1%大豆磷脂,1ml 20%triton x-100,2.5ml 0.1m柠檬酸-柠檬酸钠缓冲液。
133.10ul稀释的酶液+90ul底物50℃反应10min,95℃5min灭活,7000rpm离心5min,取1ul上清加nefa试剂盒中的试剂a 80ul,37℃反应10min,加试剂b 160ul反应10min。测定550nm吸光值。
134.an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的磷脂酶a1比酶活如图1所示。
135.图1结果显示:an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的磷脂酶a1比酶活为
20311u/mg,19677u/mg,17765u/mg和19135u/mg。
136.经检测,an02-lpl dna序列seq id no:7,an05-lpl dna序列seq id no:8,an08-lpl dna序列seq id no:9,cbs-lipase b dna序列seq id no:10;an02-lpl氨基酸序列seq id no:3,an05-lpl氨基酸序列seq id no:4,an08-lpl氨基酸序列seq id no:5,cbs-lipase b氨基酸序列seq id no:6。
137.实施例3:an02-lpl,an05-lpl,an08-lpl和cbs-lipaseb的溶血磷脂酶活力
138.同样取an02-lpl,an04lpl(即cbs-lipaseb),an05-lpl和an08-lpl的摇瓶发酵培养液,以1-棕榈酸甘油酰磷酸胆碱(溶血磷脂)作为底物测定溶血磷脂酶活力,酶活测定方法如下:
139.9ml底物:5ml 1%溶血磷脂,1ml 20%triton x-100,2.5ml 0.1m柠檬酸-柠檬酸钠缓冲液。
140.10ul稀释的酶液+90ul底物50℃反应10min,95℃5min灭活,7000rpm离心5min,取1ul上清加nefa试剂盒中的试剂a 80ul,37℃反应10min,加试剂b 160ul反应10min。测定550nm吸光值。
141.an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的溶血磷脂酶比酶活如图2所示。
142.图2结果显示:an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的比酶活分别为22547u/mg,22134u/mg,21413u/mg,22263u/mg。
143.实施例4:an02-lpl,an05-lpl,an08-lpl和cbs-lipaseb的脂肪酶活力
144.同样取an02-lpl,an05-lpl和an08-lpl的摇瓶发酵培养液,以橄榄油作为底物测定脂肪酶活力,脂肪酶活力酸碱滴定法测定如下:
145.量取4%pva溶液150ml,加橄榄油50ml,用高速匀浆机8000rpm乳化3min,暂停1min后再乳化3min,制备底物溶液(此溶液需现配现用)。
146.酶活定义及计算公式
147.脂肪酶酶活力单位定义为:每分钟催化底物释放出1μmol脂肪酸的酶量为1个脂肪酶活力单位(u)。
148.酶活计算公式:
[0149][0150]
式中:v:滴定样液所消耗的naoh溶液体积(ml)
[0151]v0
:滴定空白样所消耗的naoh溶液体积(ml)
[0152]
t:反应时间(min)
[0153]
n:酶液体积(ml)
[0154]
m:滴定用的naoh溶液的浓度(mmol/l)
[0155]
an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的脂肪酶比酶活如图3所示。
[0156]
根据图3结果,an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的脂肪酶比酶活极低。
[0157]
根据图1-3的结果,相比于溶血磷脂酶比酶活及磷脂酶a1比酶活,an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的脂肪酶比酶活有200倍的差距。作为脂肪酶an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb明显不具备实用性。
[0158]
实施例5:an02-lpl,an05-lpl和an08-lpl的脱胶测试
[0159]
取毛油60g摇匀,毛油搅拌加热,温度稳定在55℃;加入50%ca.h20 500ppm,于20000rpm剪切1.5min;55℃搅拌反应1h;将an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb与水混合和后加入,剪切200000 1.5min;升温至55℃开始计时,反应4h反应结束后,升温至85℃灭酶,至少保持>8min,离心得到脱胶油后取样送分析测试中心进行磷含量测试。
[0160]
各组分添加量:
[0161]
60g毛油;3%h2o 1.8ml;50ppm 1%的an02-lpl,an05-lpl,an08-lpl,或cbs-lipaseb加0.3ml;500ppm 50%ca.h2o 0.03ml 5%ca.h2o 0.3ml;h2o1.2ml
[0162]
an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb的脱胶测试结果如图4所示。根据图4结果,an02-lpl,an05-lpl,an08-lpl,cbs-lipaseb均可以将毛油的含磷量降低至10ppm以下,实现酶法脱胶,可以符合进行下一步物理精炼的标准。
[0163]
实施例5:an02-lpl在毕赤酵母中表达及随机突变
[0164]
选取an02-lpl的成熟肽,如seq id no:16所示,送生工生物工程有限公司合成基因序列(由生工生物工程有限公司根据毕赤酵母密码子偏爱性优化),克隆至pet-paom-7-9plc(上海生工合成)载体中,得到pic-an02质粒。用bglii线性化后,取500ng线性化的dna,用电转化法将载体转化至毕赤酵母gs115菌株(购自invitrogen)的感受态细胞中。将转化物接种于mgys平板(1.34%酵母氮源碱(ynb)含硫酸铵不含氨基酸,1%甘油,1m山梨醇,4
×
10-5
%d-生物素,2%琼脂)上,30℃培养3天,pic-an02的毕赤酵母转化子。挑取平板上的单克隆,至bmm-大豆磷脂筛选平板上,选取白色沉淀圈大的克隆,分别命名为pic-an02-lpl。
[0165]
以pic-an02-lpl作为模板,使用takara taq酶和引物对plpl-1/plpl-2进行易错pcr(在pcr时额外添加0.3mm的mncl2),得到大小为约1000bp的突变扩增子片段集合。通过sac-ii和ecori酶切位点将得到的片段克隆至pic-an02质粒,并将得到的载体转化入大肠杆菌dh5α菌株,其中,plpl-1/plpl-2序列如下所示:
[0166]
plpl-1:tccccgcggcgaaacgatgagatttccttc(seq id no:17),
[0167]
plpl-2:ccggaattcttaagaacactcagaaatg(seq id no:18)。
[0168]
将含pic-an02突变体的平板用2ml的无菌水进行洗涤,抽提质粒,用sali进行线性化,回收约8.5kb的片段。取500ng载体,用电转化法将载体转化至毕赤酵母gs115菌株的感受态细胞中。将转化物接种于bmm-大豆磷脂筛选平板上,30℃培养3天,得到pic-an02-lpl的毕赤酵母突变体文库。
[0169]
挑取46个在bmm-大豆磷脂筛选培养基平板上有白色沉淀圈的突变体克隆,及2个pic-an02-lpl的野生型克隆至2个24孔深孔板中。深孔板中装有2ml bmgy培养基。培养24h后离心,去除液体培养基,加入2ml bmmy培养基进行诱导培养。经过72h诱导培养后。离心取发酵上清。上清用0.1m ph4.0的柠檬酸缓冲液稀释10倍后,在60℃的水浴中保温30min;然后再稀释20倍(总计稀释200倍)后取4ul进行酶活测定。在4℃条件下的样品直接稀释200倍后取4ul测定酶活作为对照,记为100%,计算热处理后的残留酶活。测定方法如下:
[0170]
9.6ml底物:5ml 1%溶血磷脂,1ml 20%triton x-100,2.5ml 0.1m柠檬酸-柠檬酸钠缓冲液,补水至9.6ml。
[0171]
4ul稀释的酶液+96ul底物50℃反应10min,95℃5min灭活,7000rpm离心5min,取4ul上清加nefa试剂盒中的试剂a 80ul,37℃反应10min,加试剂b 160ul反应10min。测定550nm吸光值。
[0172]
48个样样品经过热处理之后的残留酶活结果如表1所示,其中6c和6d为野生型pic-an02-lpl,取两者的平均残留酶活为27.6%。而突变体1c的残留酶活为66.3%,是野生型的2.4倍,提高了140%。
[0173]
表1
[0174]
123456 36.6%31.3%27.8%17.5%34.9%30.0%a35.4%31.1%30.1%17.7%2.9%22.6%b66.3%28.0%30.2%14.3%17.1%26.8%c30.1%30.6%33.9%29.4%22.7%28.4%d9.9%23.9%8.5%18.4%2.4%8.2%e12.3%0.0%26.5%27.1%26.0%36.5%f6.4%7.3%37.2%21.3%33.3%38.0%g9.1%0.0%41.4%11.9%28.1%39.5%h
[0175]
将1c菌株接种于3ml ypd液体培养基中,30℃培养过夜,抽提基因组dna。以pic-an0m1菌株的基因组dna为模板,使用dna聚合酶和引物对aox-5/3
’‑
aox1(其中,aox-5:gactggttccaattgacaacg(seq id no:21),3
’‑
aox1:gcaaatggcattctgacatcc(seq id no:22))进行pcr扩增,得到pic-an02m1菌株中an02突变体的dna序列。将得到的序列送往上海生工生物工程公司,用引物对aox-5/3
’‑
aox1进行测序。1c菌株的an02突变体的氨基酸测序结果,如seq id no:19所示,突变位点为t255i,其核苷酸序列如seq id no:20所示。
[0176]
本发明中的序列同时具有磷脂酶和溶血磷脂酶的活力,在工业中具有广泛的用途。
[0177]
作为磷脂酶,首先可以被用在植物油脱胶过程中。在酶法脱胶过程中,植物油(如大豆油、菜子油、亚麻子油、葵花籽油等)中的磷脂被磷脂酶作用后转化为更加容易进入水相的溶血磷脂从而被去除,并且本发明的序列在ph4.0的条件下具有良好的磷脂酶活力,从而可以在酶法脱胶过程中不添加碱调节ph,减少了皂的生成。作为磷脂酶的另一个工业用途是在烘焙中应用,可以改善面团和面包的特性,比如面团的粘性和延展性,面包的结构和颜色。作为磷脂酶还可以用于具有特殊乳化性能的溶血磷脂。
[0178]
作为溶血磷脂酶可以改善水解小麦淀粉糖浆过滤性。溶血磷脂具有乳化特性,这通常导致水解淀粉糖浆过滤困难,小麦来源的糖浆尤其严重。溶血磷脂酶能够分解溶血磷脂,形成游离脂肪酸和水溶性甘油磷酸酯。溶血磷脂去除后可以提高水解淀粉糖浆的过滤性和澄清度。溶血磷脂酶也可提高玉米淀粉糖浆的过滤速率。
[0179]
由于同时具有磷脂酶和溶血磷脂酶的活力,可以应用于l-α-甘油磷酸胆碱的制备。在磷脂酶活力的催化下,当底物为卵磷脂时,产物为l-β-溶血磷脂酰胆碱和脂肪酸,在一定的反应体系和催化条件下l-β-溶血磷脂酰胆碱中sn-2位酰基会转移到sn-1位,再被该酶水解,生成l-α-甘油磷酸胆碱和脂肪酸。l-α-甘油磷酸胆碱在医药和保健品领域有广泛用途。
[0180]
最后,应理解的是,虽然本说明书的各个方面描述了具体实施方案,但本领域技术人员容易理解,这些公开的实施方案仅是本文公开的主题的原理的说明。因此,应该理解的
是,除非明确说明,所公开的主题不限于本文所述的具体组合、方法和/或配方等。此外,本领域普通技术人员将认识到,在不脱离本说明书的精神的情况下,根据本文的教导可以做出某些改变、修改、置换、变更、增加、减少和亚组合。因此,本技术所附的权利要求旨在被解释为包括在其真实精神和范围内的所有这些改变、修改、置换、变更、增加、减少和亚组合。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1