NMN衍生物的合成方法和NMN及其衍生物的医学应用
nmn衍生物的合成方法和nmn及其衍生物的医学应用
发明领域
1.本发明涉及药物化学技术领域,特别地,涉及烟酰胺单核苷酸在制备用于预防或减轻受试者的肝损伤或肝纤维化的药物中的用途,以及用于预防受试者的肝损伤或肝纤维化的方法。本发明还涉及新的nad前体(命名为二氢烟酰胺单核苷酸(nmnh))的合成方法,以及其突出的nad促进作用(boosting effect)和其他有益的生物学功能。
2.发明背景
3.以细胞外基质(ecm)的沉积为特征的肝纤维化是由急性肝损伤或慢性肝损伤触发的可逆创伤
‑
愈合反应(hernandez
‑
gea和friedman,2011)。更近期的研究已经揭示,酒精性肝病(ald)、非酒精性脂肪性肝病(nafld)和病毒性肝炎是全世界肝损伤的主要原因(brunt等人,2015;seitz等人,2018)。在早期阶段,肝损伤可以诱导肝细胞死亡,并且是肝炎症、反应性氧物质(ros)生成和肝纤维化的重要触发因素(wick等人,2013)。在晚期阶段,持续的肝损害将促进肝纤维化进展为肝硬化和肝细胞癌(hcc)(affo等人,2017;ringelhan等人,2018)。肝损伤在肝纤维化的发展和进展中起到至关重要的作用。因此,预防肝损伤已经被认为是肝纤维化的治疗策略(wattacheril等人,2018)。
4.先前的研究已经表明,在肝损伤期间观察到增加的ros水平(paik等人,2014)。此外,降低ros的含量改善肝损伤和肝纤维化(aoyama等人,2012;bettaieb等人,2015;de minicis等人,2010;paik等人,2011)。ros,包括超氧化物(o2‑
)、过氧化氢(h2o2)和羟基自由基(oh
·
),是主要由线粒体电子传递链(etc)和nadph氧化酶(nox)产生的高反应性分子(apel和hirt,2004;bedard和krause,2007;sies等人,2017;zorov等人,2014)。然而,当ros的产生增加或ros的消除减少时,高反应性ros将导致dna、蛋白质和脂质损害(d'autreaux和toledano,2007;reczek和chandel,2017)。在ros消除的处理期间,gsh和还原的trx是gpx和prx到还原的h2o2的主要电子供体(lei等人,2007;shadel和horvath,2015)。此外,gsh可以经由谷胱甘肽
‑
s
‑
转移酶(gst)使许多反应性亲电化合物脱氧,这也防止dna,蛋白质和脂质损害(hayes等人,2005;perkins等人,2015)。
5.尽管ros在生理过程和病理过程中具有双重作用,但过度的氧化剂挑战导致肝中生物分子的损害(reczek和chandel,2017;sena和chandel,2012)。因此,探索针对肝中ros调节的新机制在开发用于肝损伤和肝纤维化的新疗法中起重要作用。烟酰胺腺嘌呤二核苷酸(nad
+
),超过500种酶促反应所需的一种重要的辅酶,因其在氧化和还原中的作用而众所周知(ansari和raghava,2010;rajman等人,2018;stein和imai,2012)。越来越多的研究指示,增加nad
+
当量明显改善多器官功能,包括肝功能、肾功能、心功能和骨骼肌功能(canto等人,2012;mills等人,2016;rajman等人,2018)。nad
+
可以在从头生物合成途径(de novo biosynthesis pathway)中使用色氨酸,在preiss
‑
handler途径中使用烟酸(na),以及在补救途径(salvage pathway)中使用烟酰胺(nam)、烟酰胺核糖苷(nr)和烟酰胺单核苷酸(nmn)来合成(canto等人,2015;chiarugi等人,2012;johnson和imai,2018)。特别地,作为关键nad
+
中间体,nam、nr和nmn已经因为它们在许多小鼠疾病模型中的潜在治疗作用而被广泛地研究(mills等人,2016)。尽管通过补充中间体来提高nad
+
水平有助于健康并延长寿
命(fang等人,2016),但它们在不同疾病状况下在不同器官中的药代动力学和代谢归宿(fate)仍在研究中。此外,尚不清楚补充nad
+
前体是否能够通过维持氧化还原稳态来预防肝损伤或肝纤维化。
6.像nmn这样的关键nad中间体(通常被称为“nad补充剂”)已经引起了研究人员、投资者和消费者对其与疾病和老化相关的显著作用的关注(gariani等人,2016;guan等人,2017;mills等人,2016;mukherjee等人,2017)。传统的nad补充剂如nmn是氧化形式nad前体。这里我们引入新的还原形式nad前体—二氢烟酰胺单核苷酸(nmnh),它具有比nmn更好的nad促进作用以及其他的生物学功能如增加细胞抗氧化能力、减少脂肪积累、减少炎性反应和抑制肿瘤细胞生长。我们提出,nmnh是一种具有显著商业潜力的健康促进试剂。
7.发明概述
8.在一些实施方案中,本发明涉及烟酰胺单核苷酸在制备用于预防或减轻受试者的肝损伤或肝纤维化的药物中的用途。特别地,烟酰胺单核苷酸通过增加肝中gsh的水平来预防或减轻肝损伤或肝纤维化。在一些实施方案中,本发明涉及用于预防受试者的肝损伤或肝纤维化的方法。特别地,该方法包括以对于增加肝中gsh的水平有效的量向受试者施用烟酰胺单核苷酸。
9.在一些实施方案中,本发明涉及用于生产二氢烟酰胺单核苷酸(nmnh)的方法和nmnh的用途。
10.以上概述并非意图描述本发明的每种公开的实施方案或每种实施方式。当参考以下详细描述时,本发明的这些和其他方面对于本领域普通技术人员将更容易明显。
附图说明
11.图1 nmn预防ccl4处理的小鼠的肝纤维化。
12.(a)小鼠组和ccl4处理的小鼠的实验设置的示意图。
13.(b)来自ccl4处理的小鼠(n=5只/组)的肝的h&e染色和masson染色。h&e染色中的箭头指示肝窦周间隙的组织病理学损害和炎性细胞浸润。masson染色中的箭头指示肝窦周间隙的胶原沉积。h&e染色照片和masson染色照片中黑线的长度是2mm。
14.(c)在研究进程中的小鼠体重轨迹。
15.(d)和(e).通过rt
‑
qpcr检查ccl4处理的小鼠中两个标记物基因col1a1和tgfb1的表达。
16.图2 小鼠肝中nmn介导的代谢变化的代谢组学分析。
17.(a)代谢组学测定的工作流程的示意图。
18.(b)nmn处理的小鼠和pbs处理的小鼠之间代谢物丰度的比较。火山图中的红点指示代谢物水平增加(倍数变化>1.2,并且p<0.05)。火山图中的绿点指示代谢物水平降低(倍数变化<0.83,并且p<0.05)。火山图中的蓝点指示代谢物没有显著变化。
19.(c)nmn处理的小鼠和pbs处理的小鼠之间改变的代谢物的代谢物集富集分析。
20.(d)通过使用nmn处理的小鼠与pbs处理的小鼠相比的改变的代谢物进行的富集分析的前5项的代谢物
‑
途径网络分析。
21.(e)nmn+ccl4处理的小鼠和ccl4处理的小鼠之间代谢物丰度的比较。火山图中的红点指示代谢物水平增加(倍数变化>1.2,并且p<0.05)。火山图中的绿点指示代谢物水平降
低(倍数变化<0.83,并且p<0.05)。火山图中的蓝点指示代谢物没有发生显著变化。
22.(f)nmn+ccl4处理的小鼠和ccl4处理的小鼠之间改变的代谢物的代谢物集富集分析。
23.(g)通过使用nmn+ccl4处理的小鼠与ccl4处理的小鼠相比的改变的代谢物进行的富集分析的前5项的代谢物
‑
途径网络分析。
24.图3 nmn介导的肝nad
+
代谢的变化
25.(a)nad
+
代谢途径的示意图。
26.(b)至(e)本研究中的nad
+
代谢途径中中间代谢物的变化。中间代谢物包括nmn、nad
+
、nadp
+
和甲基
‑
nam。n=5只/组。
27.图4 nmn提高ccl4处理的小鼠的肝gsh水平。
28.(a)gsh代谢途径的示意图。
29.(b)至(e)本研究中的gsh代谢途径中中间代谢物水平的变化。中间代谢物包括gsh、gssg和glu
‑
cys。n=5只/组。
30.图5 nmn改善ccl4处理的小鼠和taa处理的小鼠两者的肝损伤。
31.(a)至(d)ccl4处理的小鼠中四种主要肝损伤标记物(alt、ast、tp和alb)水平的变化。本研究中通过使用小鼠血清进行的血清生化测定来分析四种标记物的水平。n=5只/组。
32.(e)至(h)taa处理的小鼠中四种主要肝损伤标记物(alt、ast、tp和alb)水平的变化。n=5只/组。
33.(i)拟建模型表明,nmn经由增强(boost)肝抗氧化剂来预防肝损伤和肝纤维化。
34.图6 nmn预防taa处理的小鼠的肝纤维化。
35.(a)小鼠组和taa处理的小鼠的实验设置的示意图。
36.(b)taa处理的小鼠的小鼠肝的h&e染色和masson染色,n=5只/组。h&e染色中的箭头指示肝窦周间隙的组织病理学损害和炎性细胞浸润。masson染色中的箭头指示肝窦周间隙的胶原沉积。
37.(c)和(d).通过rt
‑
qpcr检查taa处理的小鼠中两个标记物基因tgfb1和col1a1的表达。
38.图7 nmn补充的小鼠的肝代谢组学剖析(liver metabolomics profiling)的数据处理和热图分析。
39.(a)来自相同qc样品的三次技术重复的变化性频率。绿色柱表示从5%至100%的变化性频率。红色折线表示累积的变化频率。
40.(b)通过z评分计算进行归一化之前和之后样品的箱线图和密度图。
41.(c)通过z评分计算进行归一化之前和之后50个特征的箱线图和密度图。
42.(d)前50种代谢物的热图聚类分析。在数据处理(缺失值估计和z评分归一化)之后,通过单向anova分析计算全部改变的代谢物的前50种。
43.图8 ccl4处理的小鼠中nad
+
代谢的其他中间代谢物的nmn介导的变化。
44.(a)至(e)以下四组中na、nam、nr、namn和色氨酸的水平的变化:pbs+油、pbs+ccl4、nmn+油和nmn+ccl4。
45.图9 tdo衍生的nmnh的ms1光谱。
46.图10 nudc表达载体(左)和纯化的nudc的考马斯亮蓝染色凝胶图(右)。
47.图11 nudc反应混合物的ms1光谱。
48.图12 tdo衍生的nmnh(上)和nudc衍生的nmnh(下)之间的ms2光谱比较。
49.图13 在786
‑
o细胞中nmn/nmnh处理持续6h之后细胞总nad
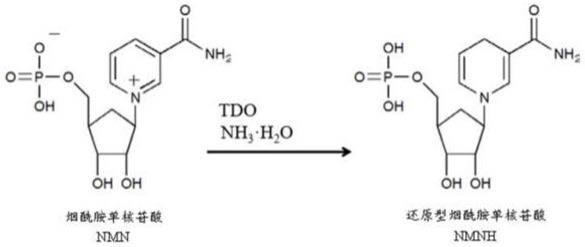
+
浓度倍数变化。
50.图14 在786
‑
o细胞中nmnh处理持续24h之后细胞gsh浓度倍数变化。
51.图15 在nmnh处理持续24h之后3t3细胞中的相对tag丰度。
52.图16 不同处理下的il
‑
6mrna水平。
53.图17 在nmnh处理下786
‑
o细胞和hk
‑
2细胞的72h生长速率。
54.发明详述
55.本发明的发明人发现,在ccl4诱导的小鼠肝纤维化模型和taa诱导的小鼠肝纤维化模型两者中,nmn预防肝纤维化,nmn补充提高了小鼠肝中gsh的水平,并且通过补充nmn显著减小了肝损伤。本发明提出了通过补充nmn的用于肝损伤和肝纤维化的新治疗策略。
56.在第一方面中,本发明提供了烟酰胺单核苷酸在制备用于预防或减轻受试者的肝损伤或肝纤维化的药物中的用途。在一些实施方案中,烟酰胺单核苷酸通过增加肝中gsh的水平来预防或减轻肝损伤或肝纤维化。在一些优选的实施方案中,药物适合于腹膜内注射。在一些优选的实施方案中,受试者是哺乳动物,诸如灵长类动物、小鼠、狗、猫、马和牛,并且优选地是人类。
57.在第二方面中,本发明提供了用于预防受试者的肝损伤或肝纤维化的方法,包括向受试者施用烟酰胺单核苷酸或包含烟酰胺单核苷酸的组合物。特别地,烟酰胺单核苷酸或包含烟酰胺单核苷酸的组合物以对于增加肝中gsh的水平有效的量施用至受试者。在一些实施方案中,通过腹膜内注射将烟酰胺单核苷酸或包含烟酰胺单核苷酸的组合物施用至受试者。在一些优选的实施方案中,烟酰胺单核苷酸的量在400mg/kg受试者体重至600mg/kg受试者体重的范围内。在一些优选的实施方案中,烟酰胺单核苷酸的量是500mg/kg受试者体重。在一些优选的实施方案中,受试者是哺乳动物,诸如灵长类动物、小鼠、狗、猫、马和牛,并且优选地是人类。
58.在第三方面中,本发明提供了药物试剂盒,该药物试剂盒包括:容器、在容器内的包含烟酰胺单核苷酸的组合物、以及容器上的或与容器相关的标签,该标签指示该组合物用于预防或减轻肝损伤或肝纤维化。
59.在第四方面中,本发明提供了用于增加受试者的肝中gsh的水平的方法,包括向受试者施用烟酰胺单核苷酸或包含烟酰胺单核苷酸的组合物。在一些实施方案中,通过腹膜内注射将烟酰胺单核苷酸或包含烟酰胺单核苷酸的组合物施用至受试者。在一些优选的实施方案中,烟酰胺单核苷酸的量在400mg/kg受试者体重至600mg/kg受试者体重的范围内。在一些优选的实施方案中,烟酰胺单核苷酸的量是500mg/kg受试者体重。在一些实施方案中,受试者处于或将处于氧化应激下。在一些优选的实施方案中,受试者是哺乳动物,诸如灵长类动物、小鼠、狗、猫、马和牛,并且优选地是人类。
60.在第五方面中,本发明提供了烟酰胺单核苷酸,用于预防或减轻肝损伤或肝纤维化。在另外的方面中,本发明提供了包含烟酰胺单核苷酸的药物组合物,用于预防或减轻肝损伤或肝纤维化。
61.在第六方面中,本发明提供了用于通过用还原剂化学地还原nmn来生产nmnh的方
法。在一些实施方案中,还原剂是二氧化硫脲(tdo)。在一些实施方案中,用tdo的还原反应在20℃
‑
60℃在5%
‑
50%(wt)氨溶液中进行。
62.在其他方面中,本发明提供了nmnh或二氢烟酸单核苷酸(namnh)的用途。在一些实施方案中,提供了用于增加细胞中总nad
+
/nadh浓度的方法,包括使细胞与有效量的nmnh或namnh接触。在一些实施方案中,提供了用于增加受试者的总nad
+
/nadh水平的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了用于增加细胞中谷胱甘肽(gsh)浓度的方法,包括使细胞与有效量的nmnh或namnh接触。在一些实施方案中,提供了用于增加受试者的gsh水平的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了用于在有相应需要的受试者中减轻体重或减少体重增加的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了用于减轻体重或减少体重增加的组合物,该组合物包含有效量的nmnh或namnh。在一些实施方案中,提供了用于治疗有相应需要的受试者的炎性紊乱的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了用于降低有相应需要的受试者的il
‑
6水平的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了抗炎组合物,该抗炎组合物包含有效量的nmnh或namnh。在一些实施方案中,提供了用于治疗有相应需要的受试者的肿瘤的方法,包括向受试者施用有效量的nmnh或namnh。在一些实施方案中,提供了抗肿瘤组合物,该抗肿瘤组合物包含有效量的nmnh或namnh。在一些实施方案中,提供了nmnh或namnh在制备用于治疗受试者的肿瘤的药物中的用途。在一些实施方案中,肿瘤是肾肿瘤。
63.除非另外解释,否则本文使用的所有技术术语和科技术语均具有如本公开内容所属的领域中的普通技术人员通常所理解的相同含义。为了便于回顾本发明的各种实施方案,具体术语的解释提供如下。
64.除非上下文另外清楚地指示,否则单数术语“一(a)”、“一(an)”和“该(the)”包括复数指示物。
65.如本文使用的术语“受试者”包括人类和非人类哺乳动物两者,非人类哺乳动物诸如小鼠、狗、猫、马和牛。
66.如本文使用的术语“减轻”包括减少或改善疾病或状况(例如,肝损伤或肝纤维化)的至少一种症状,抑制疾病或状况,例如,阻止疾病或状况的发展,缓解疾病或状况,引起疾病或状况的消退,缓解由疾病或状况引起的状况,或停止疾病或状况的症状。
67.如本文使用的术语“预防肝损伤或肝纤维化”指的是抑制肝损伤或肝纤维化的部分或全部的发展,例如,在已知具有肝损伤或肝纤维化倾向的受试者中。
68.如本文使用的术语“总nad
+
/nadh浓度”指的是细胞中nad
+
和nadh的总浓度。类似地,如本文使用的术语“总nad
+
/nadh水平”指的是受试者的nad
+
和nadh的总水平,其可以被方便地评估,例如,通过确定取自受试者的血样中的nad
+
和nadh浓度。
69.化合物诸如烟酰胺单核苷酸的术语“有效量”意指当施用至细胞或受试者时,足以在细胞或受试者中实现期望效果的量。对于受试者,有效量将取决于所施用的化合物、受试者、痛苦的严重程度和类型以及化合物的施用途径。
70.尽管可以在本公开内容的实践或测试中使用类似于或等效于本文描述的那些方法和材料的方法和材料,但合适的方法和材料在下文被描述。此外,这些材料、方法和实例仅是说明性的并且不意图是限制性的。
实施例
71.实验模型和受试者
72.6至8周大的雄性c57bl/6j小鼠被饲养在清华大学实验动物研究中心的特定的无病原体条件下。为了建立肝纤维化的四氯化碳(ccl4)小鼠模型,c57bl/6j小鼠每两天一次以2μl/g体重腹膜内地注射在玉米油或媒介物(玉米油)中3:10稀释的ccl4,持续32天的时间。在ccl4注射的同时,通过ip施用烟酰胺单核苷酸(500mg/kg体重)。为了建立肝纤维化的taa小鼠模型,c57bl/6j小鼠每两天一次以200mg/kg体重腹膜内地注射溶解在pbs或媒介物(pbs)中的taa,持续40天的时间。在taa注射的同时,腹膜内地注射烟酰胺单核苷酸(500mg/kg体重)。在最后的ccl4或taa注射之后24小时处死小鼠,并且然后收集血液和肝样品用于后续研究,后续研究包括组织学分析、生物化学分析和代谢组学分析。
73.肝组织学和成像
74.将小鼠肝分离并且分成小块,并且在4%多聚甲醛溶液中固定持续12小时。然后,将组织包埋在石蜡中并且切成切片(5μm)。将所有样品用苏木精
‑
伊红(h&e)和masson三色染色,用于病理学评价和使用标准程序的胶原检测。
75.定量rt
‑
pcr测定
76.对于定量rt
‑
pcr,使用trizol提取从细胞裂解物中分离总rna。根据制造商的方案,用逆转录系统进行互补dna合成。通过使用sybr green试剂进行定量rt
‑
pcr。相对标准曲线法被用于定量,并且2
(
‑
δδct)
法被用于基因表达计算。研究中使用的引物的序列如下:tgf
‑
β1(正向引物,seq id no:1):ctttgtacaacagcacccgc;tgf
‑
β1(反向引物,seq id no:2):tgcttcccgaatgtctgacg;col1a1(正向引物,seq id no:3):ggagagagcatgaccgatgg;col1a1(反向引物,seq id no:4):aagttccggtgtgactcgtg。
77.用于代谢组学测定的样品制备
78.为了提取组织代谢物,将在
‑
80℃预冷冻的1ml的80%(v/v)hplc级甲醇(甲醇:水=80:20)添加至2ml离心管中的100mg肝组织。组织研磨机被用于粉碎在管中的干冰上的肝组织。将管中粉碎的肝组织涡旋持续少于1min,并且在
‑
80℃孵育持续1小时。在孵育之后,将管在4℃以14000g离心持续20min,并且将上清液转移至新的离心管。在通过使用真空离心机(speedvac)将新离心管中的上清液干燥之后,将干燥的样品重新悬浮在150μl溶解缓冲液中。样品在24小时内通过联接至质谱仪的液相色谱法(lc
‑
ms/ms)进行分析。
79.基于lc
‑
ms/ms的代谢组学测定
80.基于lc
‑
ms/ms的非靶向定量代谢组学测定是在联接至具有beh酰胺柱(waters)的ultimate 3000的q exactive轨道阱质谱仪(thermo
‑
fisher scientific)上进行的。在正离子模式下,代谢物以洗脱缓冲液通过30min梯度洗脱进行分离,该洗脱缓冲液包含流动相a(95%乙腈中的10mm乙酸铵)和流动相b(95%乙腈中的10mm乙酸铵)。施加甲酸以将流动相a和流动相b的ph调节至3.0。在负离子模式下,使用氢氧化铵溶液将流动相a和流动相b的ph调节至9.0。在分析中进行数据依赖型采集模式,并且将前体和碎片的质量分辨率设置为70,000和17,500。在正负离子切换模式下,具有ultimate 3000的tsq quantiva质谱仪(thermo
‑
fisher scientific)被用于进行靶向定量代谢物测定。对于lc分离,在该分析中使用了含水流动相(在h2o中的10mm三甲胺和15mm乙酸)和有机相(100%乙腈)。嵌入由librarymanager创建的内部数据库(2.0版,thermo
‑
fisher scientific)的tracefinder软
件(3.2版,thermo
‑
fisher scientific)被应用于lc
‑
ms/ms数据的识别和定量。
81.血清生化测定
82.通过自动生化分析仪(advia 2400,siemens,德国)对天冬氨酸转氨酶(ast)、丙氨酸转氨酶(alt)、白蛋白(alb)和总蛋白(tp)进行分光光度法分析。
83.热图聚类分析
84.单向anova被用于分析通过使用metaboaanlyst软件包(chong等人,2018)归一化(z评分算法)的代谢组学数据。通过使用complexheatmap软件包(ding等人,2016)选择全部改变的代谢物的前50种(p<0.05)进行热图聚类分析。
85.代谢组学的途径富集和代谢物
‑
途径分析
86.代谢物的增加的水平(倍数变化>1.2,并且p<0.05)和代谢物的降低的水平(倍数变化<0.83,并且p<0.05)被选择用于通过使用metaboaanlyst4.0(chong等人,2018)的代谢物集富集分析(kgee)。igraph包被应用于代谢物
‑
途径分析。
87.实施例1 nmn预防肝纤维化
88.为了研究nmn是否能够体内抑制肝纤维化,通过每两天一次腹膜内注射以0.4ml/kg体重的剂量的四氯化碳(ccl4)持续32天的时段,诱导经典的肝纤维化小鼠模型(图1a)。然后,进行h&e染色和masson染色以评估肝纤维化的水平,并且结果揭示,与ccl4处理的小鼠相比,在nmn+ccl4处理的小鼠中胶原沉积减少(图1b)。与先前的研究(mills等人,2016)一致,还观察到nmn+ccl4处理的小鼠的平均体重大于ccl4处理的小鼠的平均体重(图1c)。此外,两个重要的纤维化标记物基因tgfb1和col1a1通过rt
‑
qpcr确定,并且结果示出,与ccl4处理的小鼠相比,标记物基因的表达在nmn+ccl4处理的小鼠中显著减少(图1d和图1e)。数据清楚地指示,nmn在预防肝纤维化方面具有至关重要的作用。
89.为了确定nmn是否在另一种肝纤维化小鼠模型中具有相同作用,使用硫代乙酰胺(taa)诱导的肝纤维化模型来验证我们从ccl4诱导的小鼠模型中的发现(图6a)。如预期的,masson染色结果呈现,nmn+taa处理的小鼠的胶原沉积也少于taa处理的小鼠的胶原沉积(图6b)。此外,与taa处理的小鼠相比,在nmn+taa处理的小鼠中标记物基因tgfb1和col1a1的表达也被抑制(图6c和图6d)。综上所述,数据证明nmn预防小鼠的肝纤维化。
90.实施例2 nmn补充的小鼠的肝代谢组学剖析
91.nmn作为重要的nad
+
前体,被用于通过体内补救途径合成nad
+
,并且nad
+
在氧化还原反应和代谢调节中具有重要作用(rajman等人,2018)。为了研究nmn补充的小鼠的代谢适应性,代谢物通过使用80%甲醇(v/v)从肝组织中提取,并且通过基于非靶向和靶向的液相色谱法
‑
串联质谱法液相色谱法(lc
‑
ms/ms)的代谢组学分析进行分析(图2a)。随后,发明人鉴定了总共387种离子物质,这些离子物质通过将它们的测量质量映射到人类代谢物推定地注释。为了评价检测方法的变化性,通过使用来自相同质量控制(qc)样品的三次技术重复的数据来计算变化性的频率,并且发现90%的代谢物的变化小于20%(图7a)。此外,全部代谢物的前50种通过数据归一化之后的单向anova分析进行(图7b和图7c),并且前50种代谢物的热图聚类分析示出,ccl4处理的小鼠、nmn处理的小鼠和nmn+ccl4处理的小鼠在代谢物变化模式方面彼此不同(图7d)。因此,进一步提供了全面的代谢组变化资源,以便从代谢组学的角度研究nmn补充如何减轻肝纤维化。
92.实施例3 nmn介导的小鼠肝代谢组的变化
93.如上文提及的,nmn处理的小鼠和nmn+ccl4处理的小鼠的总体蛋白质组变化是不同的。为了进一步研究nmn处理的小鼠和nmn+ccl4处理的小鼠的总体代谢物变化,首先通过与pbs处理的小鼠进行比较来分析nmn处理的小鼠的代谢物变化,并且发现与pbs处理的小鼠相比,nmn处理的小鼠中19种代谢物的水平增加(倍数变化>1.2,并且p<0.05)并且19种代谢物的水平降低(倍数变化<0.83,并且p<0.05)(图2b)。然后,代谢物集富集分析(msea)揭示,改变的代谢物主要富集在nad
+
代谢、氨基糖和核苷酸糖代谢以及氨基酸代谢中(图2c)。为了进一步研究nmn处理的小鼠中代谢物和代谢途径之间的关系,进行了代谢物
‑
途径网络分析以揭示改变的代谢物和富集的代谢途径之间的复杂关系(图2d)。此外,通过与ccl4处理的小鼠进行比较来评估nmn+ccl4处理的小鼠中改变的代谢物,并且发现与ccl4处理的小鼠相比,nmn+ccl4处理的小鼠中15种代谢物水平增加(倍数变化>1.2,并且p<0.05)并且13种代谢物水平降低(倍数变化<0.83,并且p<0.05)(图2e)。然后,富集分析揭示,改变的代谢物主要富集在谷胱甘肽代谢、nad
+
代谢和鞘脂代谢中(图2f)。此外,代谢物
‑
途径网络分析示出,与ccl4处理的小鼠相比,在nmn+ccl4处理的小鼠中改变的代谢物和富集的代谢途径之间的复杂关系(图2g)。结果揭示,在氧化应激下或不在氧化应激下,通过补充nmn诱导的代谢模式是不同的,并且表明nmn补充可以通过在氧化应激下提高gsh的水平来改善肝损伤。
94.实施例4 nmn介导的肝nad
+
代谢的变化
95.从头合成途径和从前体(nam、nr和nmn)的补救途径是促成nad
+
合成的两个主要途径,并且cd38、sirt、parp和nadk是哺乳动物中nad
+
的主要消耗物(canto等人,2015)(图3a)。为了研究在补充nmn之后nad
+
代谢途径中的代谢物变化,发明人分析了代谢组学数据,并且发现与pbs处理的小鼠相比,nmn处理的小鼠中nmn、nad
+
、nadp
+
和namn的水平增加,而色氨酸、na、nr和nam的水平没有显著地不同。然而,与nmn处理的小鼠相比,nmn+ccl4处理的小鼠中nmn、nad
+
、nadp
+
、nr、namn和甲基
‑
nam的水平降低(图3b
‑
图3e;图8a
‑
图8e)。这些数据揭示,nmn补充增加了肝中nmn、nad
+
和nadp
+
的水平,并且增加的nad
+
可能被消耗以抵抗氧化损伤。
96.实施例5 nmn在氧化应激下增加肝gsh水平
97.如上文描述的,在nmn+ccl4处理的小鼠和ccl4处理的小鼠之间的途径富集分析的最突出的途径是谷胱甘肽代谢,其包括合成、降解、再循环和解毒过程(图2f和图4a)。随后,通过评估代谢组学数据来研究gsh途径的代谢调节。非常有趣的是,数据揭示,与ccl4处理的小鼠相比,nmn+ccl4处理的小鼠中gssg的水平没有显著差异,而gsh的水平和gsh/gssg的比率显著增加。然而,在pbs处理的小鼠和nmn处理的小鼠之间,gsh的水平和gsh/gssg的比率没有显著差异(图4b
‑
图4d)。结果示出,在氧化应激下,nmn补充改善了小鼠肝中的gsh水平。此外,观察到与ccl4处理的小鼠相比,在nmn+ccl4处理的小鼠中glu
‑
cys的水平也增加,指示nmn可能在氧化应激下增强gsh的合成(图4e)。
98.实施例6 nmn改善肝损伤
99.先前的研究已经示出,gsh在改善肝损伤中ros诱导的氧化损害中具有至关重要的作用(kennon j.heard,2008;whyte等人,2007)。基于来自肝代谢组学的上述发现,本发明人提出nmn可能改善ccl4处理的小鼠的肝损伤。为了探索这种可能性,四种主要的肝损伤标记物(alt、ast、tp和alb)通过血清生化测定进行了测试。如预期的,在ccl4处理的小鼠中发现了四种标记物的增加的水平,而与ccl4处理的小鼠相比,nmn+ccl4处理的小鼠呈现出显著
的降低(图5a
‑
图5d)。为了确定nmn在taa诱导的肝纤维化小鼠模型中是否具有相同的作用,通过血清生化测定示出,与taa处理的小鼠相比,taa+nmn处理的小鼠中ast和alt的水平显著降低,并且tp和alb的水平也略微降低(图5e
‑
图5h)。这些数据证明,nmn补充改善了小鼠肝损伤。
100.总之,这表明nmn通过增加gsh的水平和nad(p)
+
依赖性氧化还原酶的丰度改善了肝损伤,这防止了肝纤维化(图5e)。
101.实施例7 用于生产nmnh的方法和nmnh的用途
102.烟酰胺腺嘌呤二核苷酸(nad
+
)是与诸如dna修复、基因表达调节和代谢的多种必需生物学过程相关的关键辅因子(magni等人,2012;canto等人,2015),其于1906年由harden和young在酵母发酵中发现。最近的研究指示,nad
+
代谢参与老化和健康调节,这使得它成为研究和商业中流行的明星分子(yoshino等人,2018)。nad
+
在哺乳动物中通过三种途径合成,包括由色氨酸的从头途径、由烟酸(na)的preiss
‑
handler途径和由烟酰胺(nam)的补救途径(yaku等人,2018)。此外,还可以由烟酰胺核糖苷(nr)和烟酰胺单核苷酸(nmn)合成nad
+
(bieganowski等人,2018)。
103.像nmn这样的关键nad
+
中间体(通常被称为“nad
+
补充剂”)已经引起了研究人员、投资者和消费者对其与疾病和老化相关的显著作用的关注(guan等人,2017;mills等人,2016;gariani等人,2016;mukherjee等人,2017)。传统的nad
+
补充剂如nmn是氧化形式nad
+
前体。这里我们介绍新的还原形式nad
+
前体—二氢烟酰胺单核苷酸(nmnh),它具有比nmn更好的nad
+
促进作用和其他生物学功能,如增加细胞抗氧化能力,减少脂肪积累,减少炎性反应和抑制肿瘤细胞生长。我们提出,nmnh是一种具有显著商业潜力的健康促进试剂。
104.7.1 nmnh合成和鉴定
105.7.1.1 nmnh合成
106.二氢烟酰胺单核苷酸(nmnh)是通过使用二氧化硫脲(tdo)作为还原剂还原烟酰胺单核苷酸(nmn)生产的。tdo是一种比具有自燃倾向的连二亚硫酸钠安全得多的还原剂,其用于二氢烟酰胺核糖苷(nrh)生产(yang等人,2019)。与nrh生产程序相比,我们的nmnh生产方法要求简单和温和得多的反应条件,具有更低的成本,并且更易于生产规模扩大(yang等人,2019)。
107.反应程序在下文示出。对于1毫升规模的反应,将100mg
‑
500mg nmn和50mg
‑
300mg tdo溶解在5%
‑
50%的氨溶液中。然后将反应混合物在20℃
‑
60℃水浴中孵育持续0.1小时
‑
10小时,并且然后使用酰胺柱通过hplc纯化nmnh。
[0108][0109]
使用酰胺柱(waters,xbridge,10
×
250mm)纯化nmnh。洗脱梯度在表1中示出。流动相a是h2o(ph 10),并且流动相b是乙腈(ph 10)。
[0110]
表1用于nmnh纯化的洗脱梯度
[0111][0112]
7.1.2 nmnh鉴定
[0113]
质谱分析被用于正确的产物鉴定。如图9中所示,获得的nmnh在负模式下具有335.06的m/z(ms1),指示nmnh比nmn多2个氢原子。nmnh的元素组成是c11 h17 n2 o8 p1,这与ms结果一致。
[0114]
先前的研究指示,一种来自大肠杆菌(e.coli)的被称为nudc的酶能够将nadh分解成nmnh和amp(friedlos等人,1992)。为了进一步证实我们的nmnh产物的正确性,我们使用大肠杆菌表达nudc,以通过分解nadh产生nmnh。表达质粒和纯化的nudc的考马斯亮蓝染色凝胶图在图10中示出。
[0115]
对于1毫升规模的反应,将20mg nadh和13μg nudc溶解在含有500mm乙酸铵和3mm氯化镁的水中。然后将反应混合物在48℃水浴中孵育持续1小时。由nudc催化的nadh分解的反应程序在下文示出。
[0116][0117]
图11示出了nudc反应混合物的负模式ms1光谱。如先前研究所表明的,nadh被催化以形成nmnh和amp(friedlos等人,1992)。
[0118]
我们比较了tdo衍生的nmnh和nudc衍生的nmnh之间的ms2特征,并且发现它们是相同的(如图12中所示)。因此我们验证了我们已经成功地通过tdo还原反应生产nmnh。
[0119]
我们比较了tdo还原法和nudc催化法的nmnh生产,并且揭示tdo法比nudc法更有效,因为产物浓度高、反应条件温和以及不含金属离子和蛋白质。
[0120]
7.2 nmnh生物学功能
[0121]
7.2.1 nmnh是比nmn更好的nad
+
促进剂
[0122]
nmnh是nad
+
/nadh的前体,因此我们验证了nmnh的nad促进作用。
[0123]
按照用户的使用说明,使用nad
+
/nadh检测试剂盒(beyotime,s0175)测量细胞内总nad浓度。简言之,将786
‑
o细胞在6孔板中培养直到90%覆盖(coverage),并且然后用500μm nmnh或nmn处理持续6小时。然后移除培养基,并且用1ml pbs洗涤细胞。向细胞中添加200μl冷nad
+
/nadh提取缓冲液进行细胞裂解。在离心(12,000g,10min,4℃)之后,收集上清液用于总nad测量。将20μl样品上清液添加至96孔板中,并且与90μl乙醇脱氢酶工作液混合,并且在37℃孵育持续10min。然后向混合物中添加10μl显色试剂,并且在37℃孵育之后测量450nm的吸光度。根据标准曲线计算总nad浓度。
[0124]
如图13中所示,在6h处理之后,在786
‑
o细胞中,与未处理组相比,nmnh使细胞总nad
+
浓度增加了3倍。在另一方面,在786
‑
o细胞中,nmn仅使细胞nad
+
浓度增加了1.2倍。结果清楚地表明,nmnh是比nmn更好的nad
+
促进剂。我们提出,nmnh是比广泛使用的nmn更好的nad
+
补充剂。
[0125]
7.2.2 nmnh增加细胞gsh浓度,gsh是一种关键的抗氧化分子
[0126]
我们还测量了nmnh处理的786
‑
o细胞的gsh浓度。
[0127]
按照用户的使用说明,使用gsh和gssg检测试剂盒(beyotime,s0053)测量细胞gsh浓度。简言之,将786
‑
o细胞在6
‑
cm盘中培养直到80%覆盖,并且然后用500μm nmnh处理持续24小时。然后移除培养基,并且用1ml pbs洗涤细胞。将细胞收集并且重新悬浮在溶液m中,并在液氮和水中冷冻和解冻。在离心之后,将10μl细胞裂解物与150μl总gsh检测溶液和50μl 0.5mg/ml nadph混合。在孵育之后测量412nm的吸光度。根据标准曲线计算总gsh浓度。通过从总gsh浓度减去gssg浓度来计算gsh浓度。
[0128]
我们发现,在0.5mm nmnh处理持续24h之后,gsh浓度显著增加,如图14中所示,这指示nmnh能够通过促进gsh合成来增加细胞抗氧化能力。
[0129]
7.2.3 nmnh是潜在的体重减轻试剂
[0130]
接下来,我们验证了nmnh是否能够影响脂肪代谢。
[0131]
按照用户的使用说明,使用游离甘油试剂(sigma,f6428)和甘油三酯试剂(sigma,t2449)测量细胞tag积累。简言之,将3t3细胞在6孔板中培养,直到90%覆盖,并且然后用200μm油酸处理,以同时在具有或不具有0.5mm nmnh的情况下诱导脂肪积累持续24小时。接下来,移除培养基,并且用1ml pbs洗涤细胞。然后将细胞重新悬浮在rapa缓冲液中,并且通过超声裂解。将20μl细胞裂解物与100μl游离甘油缓冲液或甘油三酯缓冲液混合。在37℃孵育持续15min之后,测量540nm的吸光度并且计算细胞tag浓度。
[0132]
我们用200μm油酸处理前脂肪细胞3t3细胞以诱导脂肪积累。我们发现,在0.5mm nmnh处理持续24h之后,3t3细胞中三酰甘油(triacylglycerol,tag)积累显著减少,如图15中所示。这指示nmnh是有效的体重减轻试剂。
[0133]
7.2.4 nmnh具有抗炎作用
[0134]
然后我们检查了nmnh的抗炎作用。
[0135]
我们用100ng/ml佛波醇
‑
12
‑
肉豆蔻酸酯
‑
13
‑
乙酸酯(pma)处理单核细胞thp
‑
1细胞持续24h以诱导巨噬细胞形成,然后用100ng/ml脂多糖(lps)处理以诱导炎症。在0.5mm nmnh处理持续24h之后,提取mrna并且逆转录成cdna(cwbio,cw2019)。il
‑
6表达水平通过qpcr测量(cwbio,cw0957)。
[0136]
我们发现,在nmnh处理组中il
‑
6的表达水平显著降低(如图16中所示),这指示nmnh是炎症抑制试剂。
[0137]
7.2.5 nmnh抑制肿瘤细胞的生长
[0138]
我们发现,nmnh能够抑制特定种类的肿瘤细胞,诸如786
‑
o细胞。
[0139]
实验程序如下:将细胞在96孔板中培养,并且用不同浓度的nmnh处理。在孵育持续3天之后,使用10%cck8(dongren,ck04)测量生长速率。
[0140]
如图17中所示,nmnh处理抑制786
‑
o的生长,786
‑
o是肾肿瘤细胞系。在另一方面,nmnh对处于相同浓度的hk
‑
2细胞没有明显作用,hk
‑
2细胞是肾正常细胞系(ryan等人,1994)。结果指示,nmnh对癌细胞而不是正常细胞具有优先的抑制作用,这使其成为潜在的抗肿瘤试剂。
[0141]
讨论
[0142]
健康维持和长寿对人类是重要的。nad
+
,作为一种重要的辅酶,在氧化和还原中具有重要作用(rajman等人,2018)。大量研究已经表明,老化过程期间nad
+
的水平降低,并且增加nad
+
当量可以明显改善多器官功能和寿命(gomes等人,2013;massudi等人,2012;mills等人,2016;zhao等人,2015)。作为主要的nad
+
中间体,nmn可以被用于经由补救途径合成nad
+
,并且越来越多的研究指示,nmn补充通过提高小鼠中nad
+
的水平来改善健康维持(canto等人,2015;mills等人,2016)。虽然nmn补充可以改善肝功能(amano等人,2019;rajman等人,2018),但尚不清楚它是否可以通过维持氧化还原稳态来预防肝损伤和肝纤维化。
[0143]
在这项研究中,发现nmn补充通过减轻肝损伤来改善肝纤维化。代谢组学分析指示,在氧化应激下,nmn补充提高了gsh的水平和gsh/gssg的比率,这被认为是nmn改善肝损伤的主要原因。作为重要的抗氧化剂,gsh可以经由gpx将h2o2还原成h2o,并且经由gst使反应性亲电化合物脱氧(hayes等人,2005;lei等人,2007)。此外,先前的研究已经表明,通过补充gsh前体(n
‑
乙酰半胱氨酸)来提高gsh的水平可以为对乙酰氨基酚(apap)诱导的急性肝损伤解毒(kennon j.heard,2008)。然而,另外的研究指示,11.1%的用静脉内n
‑
乙酰半胱氨酸处理的患者示出了不良药物反应和类过敏反应(whyte等人,2007;yang等人,2009)。许多研究表明,在小鼠中在nmn补充之后没有明显的不良反应(yoshino等人,2018),本文提出nmn可以被用作更安全的药物,以提高肝中gsh的水平,用于肝损伤治疗。
[0144]
总之,本发明人提出,nmn补充通过提高肝中gsh的水平来预防肝损伤,这进而改善肝纤维化。此外,本技术呈现了在小鼠肝中进行nmn补充之后代谢组变化的全景,并且提出了通过补充nmn的用于肝损伤和肝纤维化的新治疗策略。
[0145]
鉴于我们的公开内容的原理可以被应用的许多可能的实施方案,应认识到,例证的实施方案仅仅是公开内容的特定示例,并且不应被视为对公开内容的范围的限制。
[0146]
我们还已经合成了一种新的nad
+
前体—nmnh,它具有显著的nad促进作用和其他健康促进作用。此处我们介绍了nmnh的生产和纯化过程,并且验证了其正确性,并且同时我
unfolded protein response by nicotinamide adenine dinucleotide repletion reverses fatty liver disease in mice[j].hepatology,63(4):1190
‑
204
[0169]
gomes,a.p.,price.n.l.,ling,a.j.,moslehi,j.j.,montgomery,m.k.,rajman,l.,white,j.p.,teodoro,j.s.,wrann,c.d.,hubbard,b.p.,et al.(2013).declining nad(+)induces a pseudohypoxic state disrupting nuclear
‑
mitochondrial communication during aging.cell 155,1624
‑
1638.
[0170]
guan y,wang s r,huang x z,et al.(2017).nicotinamide mononucleotide,an nad(+)precursor,rescues age
‑
associated susceptibility to aki in a sirtuin 1
‑
dependent manner[j].j am soc nephrol,28(8):2337
‑
52.
[0171]
hayes,j.d.,flanagan,j.u.,and jowsey,i.r.(2005).glutathione transferases.annu rev pharmacol toxicol 45,51
‑
88.
[0172]
hemandez
‑
gea,v.,and friedman,s.l.(2011).pathogenesis of liver fibrosis.annu rev pathol 6,425
‑
456.
[0173]
hong,s.,moreno
‑
navarrete,j.m.,wei,x.,kikukawa,y.,tzameli,i.,prasad,d.,lee,y.,asara,j.m.,fernandez
‑
real,j.m.,maratos
‑
flier,e.,et al.(2015).nicotinamide n
‑
methyltransferase regulates hepatic nutrient metabolism through sirtl protein stabilization.nat med 21,887
‑
894.
[0174]
johnson.s.,and imai,s.i.(2018).nad(+)biosynthesis,aging,and disease.f1000res7,132.
[0175]
kennon j.heard,m.d.(2008).acetylcysteine for acetaminophen poisoning.n engl jmed 359,285
‑
292.
[0176]
lei,x.g.,cheng,w.h.,and mcclung,j.p.(2007).metabolic regulation and function of glutathione peroxidase
‑
1.annu rev nutr 27,41
‑
61.
[0177]
liu,l.,su,x.,quinn,w.j.,3rd,hui,s.,krukenberg,k.,frederick,d.w.,redpath,p.,zhan,l.,chellappa,k.,white,e.,et al.(2018).quantitative analysis of nad synthesis
‑
breakdown fluxes.cell metab 27,1067
‑
1080e1065.
[0178]
magni g,amici a,emanuelli m,et al.(2004).enzymology of nad plus homeostasis in man[j].cell mol life sci,61(1):19
‑
34.
[0179]
massudi,h.,grant,r.,braidy,n.,guest,j.,farnsworth,b.,and guillcmin,g.j.(2012).age
‑
associated changes in oxidative stress and nad+metabolism in human tissue.plos one 7,e42357.
[0180]
mills,k.f.,yoshida,s.,stein,l.r.,grozio,a.,kubota,s.,sasaki,y.,redpath,p.,migaud,m.e.,apte,r.s.,uchida,k.,et al.(2016).long
‑
term administration of nicotinamide mononucleotide mitigates age
‑
associated physiological decline in mice.cell metab 24,795
‑
806.
[0181]
mukherjee s,chellappa k,moffitt a,ct al.(2017).nicotinamide adenine dinucleotide biosynthesis promotes liver regeneration[j].hepatology,65(2):616
‑
30.
[0182]
paik,y.h.,iwaisako,k.,seki,e.,inokuchi,s.,schnabl,b.,osterreicher,
c.h.,kisseleva,t.,and brenner,d.a.(2011).the nicotinamide adenine dinucleotide phosphate oxidase(nox)homologues nox1 and nox2/gp91(phox)mediate hepatic fibrosis in mice.hepatology 53,1730
‑
1741.
[0183]
paik.y.h,,kim,j.,aoyama,t.,de minicis,s.,bataller,r.,and brenner,d.a.(2014).role of nadph oxidases in liver fibrosis.antioxid redox signal 20,2854
‑
2872.
[0184]
perkins,a.,nelson,k.j.,parsonage,d.,poole,l.b.,and karplus,p.a.(2015).peroxiredoxins:guardians against oxidative stress and modulators of peroxide signaling.trends biochem sci 40,435
‑
445.
[0185]
rajman,l.,chwalek,k.,and sinclair,d.a.(2018).therapeutic potential of nad
‑
boosting molecules:the in vivo evidence.cell metab 27,529
‑
547.
[0186]
reczek,c.r.,and chandel,n.s.(2017).the two faces of reactive oxygen species in cancer.annual review of cancer biology1,79
‑
98.
[0187]
ringelhan,m.,pfister,d.,o
′
connor.t.,pikarsky,e.,and heikenwalder,m.(2018).the immunology of hepatocellular carcinoma.nat immunol19,222
‑
232.
[0188]
ryan m j,johnson g,kirk j,et al.(1994).hk
‑
2:an immortalized proximal tubule epithelial cell linefrom normal adult human kidney[j].kidney international,45(1):48
‑
57.
[0189]
seitz,h.k.,bataller,r.,cortez
‑
pinto,h.,gao,b.,gual,a..lackner.c..mathurin,p.,mueller,s.,szabo.g.,and tsukamoto,h.(2018).alcoholic liver disease.nat rev dis primers 4,16.
[0190]
sena,l.a.,and chandel,n.s.(2012).physiological roles of mitochondrial reactive oxygen species.mol cell 48,158
‑
167.
[0191]
shadel,g.s.,and horvath,t.l.(2015).mitochondrial ros signaling in organismal homeostasis.cell 163,560
‑
569.
[0192]
sies,h.,berndt,c.,and jones,d.p.(2017).oxidative stress.annu rev biochem 86,715
‑
748.
[0193]
stein,l.r.,and imai,s.(2012).the dynamic regulation of nad metabolism in mitochondria.trends endocrinol metab 23,420
‑
428.
[0194]
tai,h.h.,chi,x.,and tong,m.(2011).regu1ation of 15
‑
hydroxyprostaglandin dehydrogenase(15
‑
pgdh)by non
‑
steroidal anti
‑
inflammatory drugs(nsaids).prostaglandins other lipid mediat 96,37
‑
40.
[0195]
wattacheril,j.,issa,d.,and sanyal,a.(2018).nonalcoholic steatohepatitis(nash)and hepatic fibrosis:emerging therapies.annu rev pharmacol toxicol 58,649
‑
662.
[0196]
whyte,i.m.,francis,b.,and dawson,a.h.(2007).safety and efficacy of intravenous n
‑
acetylcysteine for acetaminophen overdose:analysis of the hunter area toxicology service(hats)database.curr med res opin 23,2359
‑
2368.
[0197]
wick,g.,grundtman,c.,mayerl.c.,wimpissinger,t.f.,feichtinger,j.,
zelger,b.,sgonc,r.,and wolfram,d.(2013).the immunology of fibrosis.annu rev immunol 31,107
‑
135.
[0198]
yaku k,okabe k,nakagawa t.nad(2018)metabolism:implications in aging and longevity[j].ageing res rev,47∶1
‑
17.
[0199]
yang,r.,miki,k.,he,x.,killeen,m.e.,and fink,m.p.(2009).prolonged treatment with n
‑
acetylcystine delays liver recovery from acetaminophen hepatotoxicity.crit care 13,r55.
[0200]
yang y,mohammed fs,zhang n,et al.(2019).dihydronicotinamide riboside is a potent nad(+)concentration enhancer in vitro and in vivo[j].journal of biological chemistry,294(23):9295
‑
307.
[0201]
yao,l.,chen,w.,song,k.,han,c.,gandhi,c.r.,lim,k.,and wu,t.(2017).15
‑
hydroxyprostaglandin dehydrogenase(15
‑
pgdh)prevents iipopolysaccharide (lps)
‑
induced acute liver injury.plos one 12,e0176106.
[0202]
yoshino,j.,baur,j.a.,and imai,s.i.(2018).nad(+)intermediates:the biology and therapeutic potential of nmn and nr.cell metab.27:513
‑
28.
[0203]
yu,g.,wang,l.g.,han,y.,and he,q.y.(2012).clusterprofiler:an r package for comparing biological themes among gene clusters.omics 16,284
‑
287.
[0204]
zhao,y.,hu,q.,cheng,f.,su,n.,wang,a.,zou,y.,hu,h.,chen,x.,zhou,h.m.,huang,x.,et al.(2015).sonar,a highly responsive nad+/nadh sensor,allows high
‑
throughput metabolic screening of anti
‑
tumor agents.cell metab 21,777
‑
789.
[0205]
zorov,d.b.,juhaszova,m.,and sollott,s.j.(2014).mitochondrial reactive oxygen species(ros)and ros
‑
induced ros release.physiol rev 94.909
‑
950.。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1