一种构建Serac1基因敲除小鼠模型的方法
本发明涉及生物技术领域,涉及一种构建serac1基因敲除小鼠模型的方法,具体涉及一种基于crispr-cas9基因敲除技术构建serac1基因敲除小鼠模型的方法。
背景技术:
线粒体病是一类由于线粒体功能异常所导致的疾病。线粒体功能异常与癌症、糖尿病、阿尔兹海默综合征等退行性疾病的发生密切相关。线粒体疾病分为线粒体常见病和线粒体罕见病,线粒体罕见病通常包括线粒体脑病伴乳酸酸中毒及卒中样发作综合征(melas),肌阵挛癫痫伴肌肉破碎红纤维综合征(merrf),亚急性坏死性脑病(ls)等疾病。统计表明,由线粒体氧化磷酸化(oxphos)系统异常所致的线粒体罕见病在新生儿中的发病率约为1/5000左右。目前线粒体病的诊断需要依据患者的临床症状、生化检测、影像检查、病理结果以及相关的线粒体功能研究及基因检测等。
目前线粒体病是临床医学中的一大挑战。线粒体的主要功能是为细胞的生存提供能量,是细胞进行有氧呼吸的重要场所,除了提供能量,线粒体还可以参与细胞的分化、信息传递、凋亡、调控细胞的生长周期等过程。而线粒体的能量生成主要依赖线粒体氧化磷酸化系统(oxphos),所以氧化磷酸化系统发生障碍后线粒体功能也会出现相应异常,从而导致疾病的发生。
megdel综合征是3-甲基戊烯二酸尿症(3-methylglutaconicaciduria,3-mga-uria)伴耳聋、脑病和leigh综合征,有新生儿低血糖和无病原体感染的类似于败血症的临床特点。虽然目前对该疾病的诊断已经成熟,也检测出较多的serac1基因位点的突变,为后续的诊断和治疗提供了很大的帮助,但是文献中对serac1基因的致病机制没有研究,只是在2篇文献报道中对serac1突变患者的磷脂构成进行了检测,发现serac1突变后磷脂酰甘油34:1和36:1的比例构成发生了变化,从而导致心磷脂的代谢发生异常,同时在病人的成纤维细胞中发现有胆固醇的聚集。serac1是核基因编码的蛋白,在它的n末端信号序列,在c端有一个丝氨酸脂酶域并包含一个gxsxg的脂肪酶模块,这可能与其调节线粒体磷脂酰甘油的重组和细胞内胆固醇的转运相关,而作者也认为线粒体功能损伤和胆固醇稳态的变化被认为是导致megdel的内在原因。然而这一结论仍然存在较大的争议,因此,后续很多相关的研究出现,但是总体上看这些报道仅限于对于疾病临床表型的分析,对于serac1是如何影响线粒体氧化磷酸化复合体酶的活性、如何影响磷脂的代谢以及为何对肝脏存在较严重的影响,这些机制均不明确,这也为疾病的治疗造成一定的限制;同时仅有少数的研究采用了成纤维细胞的模型,通过染色发现细胞中胆固醇聚集,以及通过干扰serac1的表达后检测磷脂的构成发生变化,对机制也没有明确研究。目前对于线粒体megdel综合征的研究主要集中于病例研究,没有动物模型研究,而目前动物模型是很好的基因突变研究模型,因此,设计动物模型对其临床特征进行分析研究研究其致病的分子机制具有重要意义。
技术实现要素:
本发明的目的是提供一种构建serac1基因敲除小鼠模型的方法,基于crispr-cas9基因敲除技术设计了serac1-/-小鼠模型,以此对研究对象建立megdel疾病模型并充分了解疾病的相关临床表型,明确serac1导致线粒体功能异常的分子机制,以期为megdel综合征的诊断和治疗提供新的依据。
为实现上述目的,本发明提供了如下方案:
本发明提供了一种构建serac1基因敲除小鼠模型的sgrna,所述sgrna作用位点位于serac1基因的intron2和intron7的非保守区,所述sgrna的dna序列如seqidno:1和seqidno:2所示。
进一步地,intron2区设计的sgrna的dna序列为:5’-tcagcagtccaggattggatagg-3’;
intron7区设计的sgrna的dna序列为:5’-gcatataccacatgtaggctggg-3’。
本发明还提供一种构建serac1基因敲除小鼠模型的方法,所述方法基于crispr-cas9基因敲除技术,所述方法包括以下步骤:
步骤1:设计靶向小鼠待敲除serac1基因的sgrna;
步骤2:sgrna与cas9核酸酶的mrna经体外转录后,一起注射到小鼠受精卵中,然后将受精卵移植到假孕母鼠体内,获得嵌合体的f0代;对f0代进行pcr鉴定,将阳性f0代小鼠与野生型小鼠交配获得具有稳定基因型的f1代小鼠,筛选serac1基因敲除的纯合子代,作为serac1基因敲除小鼠模型;
其中,所述sgrna作用位点位于serac1基因的intron2和intron7的非保守区,所述sgrna的dna序列如seqidno:1和seqidno:2所示。
优选的是,步骤2中所述的pcr鉴定用引物如下所示:
wt-f:5’-agagtcacagtcagcaactctgtgc-3’;
wt-r:5’-gttatgcaaccaggatcaccacgg-3’;
mut-f:5’-gagtcacagtcagcaactctgtgc-3’;
mut-r:5’-agttctacgcttcaacccctgcta-3’。
本发明还提供一种所述的sgrna在构建serac1基因敲除小鼠模型中的应用。
本发明公开了以下技术效果:
本发明采用cas9/sgrna注射受精卵的方法构建基因敲除小鼠,sgrnas分别设计在serac1基因的intron2和intron7的非保守区,获得嵌合体的f0代小鼠后进行其基因型鉴定,阳性小鼠再与野生型小鼠交配获得具有稳定基因型的f1代小鼠。待确定f1代小鼠的基因型后,f1代纯合子小鼠为动物模型,对其临床特征进行分析研究,发现serac1突变以后,小鼠的肌肉活力减弱,纯合基因型小鼠出生率略低,肝脏的线粒体复合体表达水平以及酶活性均有不同程度的降低,小鼠尿液3-mga增高,核磁代谢发现小鼠脑部的乳酸有轻微的增高、naa有轻度下降,小鼠的肝脏组织中胆固醇有聚集等。这些结果也提示我们sera1突变以后,会引起megdel综合征的相关临床表现。因此,本发明构建的serac1基因敲除的小鼠模型对于megdel综合征的诊断和治疗提供协助作用,并且对于serac1基因致病机理的研究也提供了一定的线索和动物模型。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动性的前提下,还可以根据这些附图获得其他的附图。
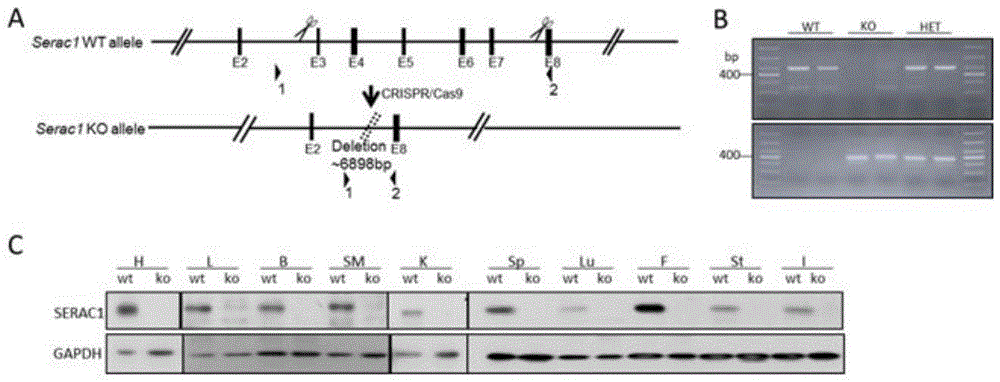
图1为serac1-/-小鼠模型的设计及鉴定;a.serac1基因sgrna设计示意图;b.琼脂糖凝胶电泳鉴定小鼠尾巴基因型,分别为野生型、纯合型和杂合型;c.westernblot分析小鼠各个组织中serac1基因的ko效果;
图2为serac1的mrna水平、蛋白水平以及serac1-/-小鼠生育能力检测;a.q-pcr技术检测野生型小鼠各个组织中serac1的mrna表达水平,n=4;b.westernblot分析野生型小鼠各个组织中serac1的蛋白表达水平,n=3;c.杂合小鼠交配后各基因型小鼠的出生数目统计;d.野生型小鼠和纯合型小鼠各自交配后,生育胎数及每胎小鼠数目统计;
图3为serac1-/-小鼠的体重和肌肉活力检测;a.雌性小鼠的体重变化,n=6;b.雄性小鼠的体重变化,n=9/10;c.雌性小鼠抓力检测结果,n=6;d.雄性小鼠抓力检测结果,n=6;e.3m和6m雌性小鼠悬吊得分检测结果;f.3m和6m雄性小鼠悬吊得分检测结果;g.3m和6m雌性小鼠平衡线上悬挂维持时间检测结果;h.3m和6m雄性小鼠平衡线上悬挂维持时间检测结果;n=3,*,p<0.05;**,p<0.01;***,p<0.001;
图4为serac1-/-小鼠的听力和视网膜电图检测;a.短声刺激下3m小鼠的左右耳听力检测结果,n=6;b.短声刺激下12m小鼠的左右耳听力检测结果,n=4/6;c.暗适应下3m小鼠在不同光强度刺激下的a波的波幅强度;d.暗适应下3m小鼠在不同光强度刺激下的b波的波幅强度;e.光适应下3m小鼠在光照刺激下的a、b波的潜伏期;f.光适应下3m小鼠在光照刺激下的a、b波的波幅;g.暗适应下12m小鼠在不同光强度刺激下的a波的波幅强度;h.暗适应下12m小鼠在不同光强度刺激下的b波的波幅强度;i.光适应下12m小鼠在光照刺激下的a、b波的潜伏期;j.光适应下12m小鼠在光照刺激下的a、b波的波幅,n=8/9;*,p<0.05;***,p<0.001;
图5为serac1-/-小鼠的mri和3-mga检测;a.mri检测3m和12m小鼠脑部基底节区结构;b.3m小鼠脑部代谢物naa、lac、glx、cho的含量变化;c.12m小鼠脑部代谢物naa、lac、glx、cho的含量变化,n=5;d.gc-ms法检测小鼠尿液中3-mga的含量;*,p<0.05;
图6为serac1-/-小鼠模型心脏组织线粒体复合体表达水平和酶活性;a.bn-page检测3m小鼠线粒体呼吸链复合体的表达水平;b.bn-page检测6m小鼠线粒体呼吸链复合体的表达水平;c.提取心脏组织线粒体,采用分光光度计法检测3m小鼠线粒体呼吸链复合体的酶活性;d.提取心脏组织线粒体,采用分光光度计法检测6m小鼠线粒体呼吸链复合体的酶活性;*,p<0.05;***,p<0.001;
图7为serac1-/-小鼠模型脑组织线粒体复合体表达水平和酶活性;a.bn-page检测3m小鼠线粒体呼吸链复合体的表达水平;b.bn-page检测6m小鼠线粒体呼吸链复合体的表达水平;c.提取脑组织线粒体,采用分光光度计法检测3m小鼠线粒体呼吸链复合体的酶活性;d.提取脑组织线粒体,采用分光光度计法检测6m小鼠线粒体呼吸链复合体的酶活性;*,p<0.05;***,p<0.001;
图8为serac1-/-小鼠模型肌肉组织线粒体复合体表达水平和酶活性;a.bn-page检测3m小鼠线粒体呼吸链复合体的表达水平;b.bn-page检测6m小鼠线粒体呼吸链复合体的表达水平;c.提取肌肉组织线粒体,采用分光光度计法检测3m小鼠线粒体呼吸链复合体的酶活性;d.提取肌肉组织线粒体,采用分光光度计法检测6m小鼠线粒体呼吸链复合体的酶活性;*,p<0.05;
图9为serac1-/-小鼠模型肾脏组织线粒体复合体表达水平和酶活性;a.bn-page检测3m小鼠线粒体呼吸链复合体的表达水平;b.bn-page检测6m小鼠线粒体呼吸链复合体的表达水平;c.提取肾脏组织线粒体,采用分光光度计法检测3m小鼠线粒体呼吸链复合体的酶活性;d.提取肾脏组织线粒体,采用分光光度计法检测6m小鼠线粒体呼吸链复合体的酶活性;**,p<0.01;
图10为serac1-/-小鼠模型肝脏组织线粒体复合体表达水平和酶活性;a.bn-page检测1m小鼠线粒体呼吸链复合体的表达水平;b.bn-page检测3m小鼠线粒体呼吸链复合体的表达水平;c.bn-page检测6m小鼠线粒体呼吸链复合体的表达水平;d.bn-page检测12m小鼠线粒体呼吸链复合体的表达水平;e.提取肝脏组织线粒体,采用分光光度计法检测3m小鼠线粒体呼吸链复合体的酶活性;f.提取肝脏组织线粒体,采用分光光度计法检测6m小鼠线粒体呼吸链复合体的酶活性;*,p<0.05;**,p<0.01;***,p<0.001;
图11为serac1-/-小鼠模型肝脏组织线粒体复合体单体表达水平及aav-serac1病毒回复实验;a.bn-page检测1m小鼠线粒体呼吸链复合体单体的表达水平;b.bn-page检测3m小鼠线粒体呼吸链复合体单体的表达水平;c.bn-page检测6m小鼠线粒体呼吸链复合体单体的表达水平;d.bn-page检测12m小鼠线粒体呼吸链复合体单体的表达水平;e.1mko小鼠注射aav-serac1肝脏特异性过表达的病毒液,westernblot分析serac1的蛋白表达水平;f.bn-page检测1m小鼠注射aav-serac1肝脏特异性过表达的病毒液的肝脏线粒体呼吸链复合体的表达水平;*,p<0.05;**,p<0.01;***,p<0.001;
图12为serac1-/-小鼠模型睾丸、脂肪和皮肤组织线粒体复合体表达水平;a.bn-page检测3m小鼠睾丸线粒体呼吸链复合体的表达水平;b.bn-page检测3m小鼠脂肪组织线粒体呼吸链复合体的表达水平;c.bn-page检测3m小鼠皮肤组织线粒体呼吸链复合体的表达水平;d.bn-page检测12m小鼠皮肤组织线粒体呼吸链复合体的表达水平;*,p<0.05;**,p<0.01。
具体实施方式
现以实施例方式对本发明技术方案进行具体说明,但其不应该被认为是对本发明的限制,而应理解为是对本发明的某些方面、特性和实施方案的更详细的描述。
下述实施例中所使用的试验方法如无特殊说明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂和材料。
实施例1serac1基因敲除小鼠模型的构建
1、通过对小鼠serac1基因的转录本、结构域和调控区域等基因结构进行分析,发现serac1基因的exon3-7可被大片段敲除,从而达到基因敲除的目的。故将sgrna设计在intron2和intron7的非保守区(图1a)。sgrna序列如下所示:
sgrna1:
sgrna2:
2、c57bl/6j雌性小鼠(3周龄,平均体重20g),46h后注射hcg,与雄性小鼠合笼交配,次日取受精卵进行显微注射,将上述的sgrna与cas9的核酸酶的mrna经体外转录后,一起注射到受精卵中,再将注射后的存活受精卵移植到假孕母鼠体内,产出f0代小鼠。
提取f0代小鼠尾部dna,pcr鉴定是否为嵌合体。
将阳性的f0代小鼠与野生型小鼠交配获得杂合子小鼠f1代,小鼠出生后20天进行pcr鉴定,筛选纯合子f1代小鼠,作为serac1基因敲除动物模型。
本发明利用上述构建的serac1基因敲除动物模型进行后续的试验。
一、小鼠基因型鉴定:
1、pcr反应体系:
表1
2、pcr反应程序:
表2
3、扩增引物:
表3
判断小鼠基因型:
使用设计好的针对小鼠尾巴基因型鉴定的引物,将模板dna进行pcr,再采用琼脂糖凝胶电泳鉴定仔鼠的基因型(图1b),根据发明人引物设计的原理,用第一对引物扩增出来的序列中只有野生型小鼠和杂合型的小鼠有条带;由于在同样的pcr反应条件下,第二对引物扩增的产物中,野生型小鼠约7000bp长度的产物无法扩增,所以第二对引物扩增的产物中只有杂合型和纯合型的小鼠有目的条带。根据此原理将新生的仔鼠进行基因型分类,有野生型(wt,+/+)、杂合基因型(het,+/-)和纯合基因型(ko,-/-)小鼠。
为了明确serac1-/-小鼠组织中serac1蛋白的ko效果,发明人分离小鼠的心脏、肝脏、脑、肌肉、肾脏、脾脏、肺、脂肪、胃、肠,提取相应组织的蛋白质,采用蛋白质免疫印迹(wb)的方法检测serac1蛋白在各个组织中的表达(图1c),结果显示在ko小鼠的这十个组织中serac1蛋白均有敲除效果。
4、利用实时荧光定量pcr技术分别用于检测小鼠正常各组织中serac1基因的mrna水平(如图2a所示)、基因敲除小鼠各组织及细胞中线粒体的转录水平和拷贝数水平。
按照表中各试剂比例配制所需要的母液,最后加入底物;
表4
反应条件设置:预变性,95℃,30s;循环反应,95℃,10sec;60℃,30sec,循环40次;融解曲线,95℃,15sec;60℃,60sec,95℃,15sec;
该实验所用到的引物如下表5所示:
表5
如图2b所示,与野生型小鼠各组织相比,发现serac1-/-小鼠各组织的serac1均有ko效果;并且发现野生型小鼠各组织serac1-mrna水平和serac1蛋白表达水平存在差异,其中心脏、脑子和肌肉含量较高。
如图2c-d所示,统计杂合小鼠之间交配,幼鼠数量和基因型鉴定发现,ko小鼠的生育能力与wt小鼠相比没有明显差异。
二、serac1-/-小鼠模型的表型分析
1、serac1-/-小鼠的体重和肌肉活力检测
分别对雌鼠和雄鼠进行体重检测,记录从第三周开始的每周小鼠体重的变化(图3a-b),发现雌鼠在出生以及后期的生长过程中ko与wt小鼠没有明显的体重差异,而野生型雄鼠在出生时体重就略高于ko雄鼠,在后期生长过程中体重也均高于ko小鼠,并且从23周开始到26周,相较于前期的体重,ko雄鼠体重有轻微的降低现象,而wt雄鼠体重仍在稳定的增加。可见serac1敲除对雄性小鼠的体重影响比较大。
对小鼠进行抓力检测、双前肢悬吊得分检测以及小鼠在平横线上悬挂维持时间检测。抓力检测发现雌鼠在开始的5周、10周和15周时,抓力有明显的差异,但从20周开始,抓力没有明显的变化;雄鼠在整个抓力检测的过程中均有明显的差异,ko小鼠的抓力与wt小鼠相比有一定的减弱(图3c-d)。小鼠双前肢的悬吊得分检测发现ko雌鼠的悬吊得分明显下降,ko雄鼠悬吊得分在3月和6月均有明显的降低,并且小鼠随着年龄的增大,悬吊得分也有降低(图3e-f)。小鼠在平衡线上维持的时间统计,发现雌鼠在平衡线上维持的时间相似,而雄性ko小鼠在两次检测中,维持时间均有所降低,并且6月大的小鼠坚持的时间与3月小鼠相比也有一定的减少(图3g-h),这可能是年龄增长对小鼠的体力有一定影响,但总体上ko小鼠在抓力、悬吊时间和平衡维持时间上要弱于wt小鼠,表明serac1-/-小鼠的肌肉活力受到一定的影响。
2、serac1-/-小鼠的听力和视网膜电图检测
1)采用短声刺激诱发听性脑干反应(abr)听力的检测方法,对serac1-/-小鼠模型进行听力的检测(图4a-b)。
结果发现:3月大wt小鼠的左右耳听阈维持在40-50db之间,ko小鼠的听阈维持在60-70db之间,ko小鼠听力受到一定的影响;在12月的小鼠听力检测结果中发现ko小鼠的听阈已经超过100db,wt小鼠的听阈上升到70-90db之间,由于小鼠听力受年龄的影响,wt小鼠的听力也有下降,但是ko小鼠的听力下降更明显,几乎只能听到相对于人来说的噪音水平,并且对声音刺激的反应比较迟钝。听力检测结果表明ko小鼠的听力随着年龄的增大,听力有明显的受损。
2)采用retiport刺激器系统,按照国际临床视网膜电生理标准化方案(iscev)进行全视野的scoterg和photoerg检测,刺激强度设置为dark0.01、dark3.0、dark10.0。
结果发现:随着光照刺激强度的增加,3月大ko小鼠和wt小鼠的erg-a、b波波幅没有差异(图4c-d);光适应下对小鼠a、b波潜伏时间和波幅进行检测(图4e-f),发现ko小鼠和wt小鼠之间无明显的差别;随后为了明确小鼠的年龄对视网膜电图是否存在影响,我们检测了12月大的小鼠erg(图4g-j),我们发现随着年龄的增长ko小鼠与wt小鼠之间在暗适应和光适应下erg-a、b波波幅及潜伏时间也没有变化,但是12月大小鼠的a、b波的波幅与3月小鼠相比有轻微的降低。以上数据表明,serac1-/-小鼠的视网膜电生理的没有发生明显的变化。
3、serac1-/-小鼠的mri和3-mga检测
1)小鼠mri检测是使用中国科学院药物研究所的磁共振仪器完成检测的。先将小鼠用气体麻醉药进行麻醉,待小鼠完全麻醉后,固定到检测台上,然后推入检测仪内,根据小鼠的麻醉状态调节气体麻醉药的剂量,确保小鼠处于完全麻醉的状态。
分别对小鼠3m和12m的ko小鼠与wt小鼠进行核磁检测(图5a),结果发现:小鼠还未出现明显的双侧基底节病变;对小鼠脑部进行活体的代谢物如n-乙酰门冬氨酸(naa)、胆碱类化合物(cho)、谷氨酸和谷氨酰胺(glx)、乳酸(lac)进行分析,以肌酐(cr)作为内参(图5b),发现在3m小鼠的检测结果中,ko小鼠naa、glx和cho有轻微的下降、lac有明显增高;12m小鼠的检测结果发现类似的现象,ko小鼠naa明显的降低,而lac水平也有升高的趋势,表明ko小鼠神经元的数量和功能可能发生了一定的变化,在葡萄糖的代谢上ko小鼠可能进行糖酵解增加;但serac1-/-小鼠没有出现明显结构改变,可能小鼠的脑部结构的病变与生化水平变化相比,出现的时间稍晚未检测出来,这需要我们对更年长的小鼠进行核磁检测。
2)收集小鼠的尿液保存在无酶的离心管中,然后离心,将上清的尿液取出放入到新的离心管中,冻存于-80℃冰箱中,由于小鼠尿液有机酸的检测需要小鼠的尿液较多,最少需要500ml,所以,需要将小鼠几次收集的尿液先保存起来。
检测前,先将小鼠的尿液在室温解冻,然后将尿液转移到检测的尿检测试纸上,检测试纸在使用前再打开,尿液均匀的扩散到试纸片上,然后室温下晾干,标记好检测的小鼠的基因型和信息,室温送检。
收集了1m和3m小鼠的尿液,采用gc-ms的方法对小鼠尿液的有机酸进行检测(图5c),在1m小鼠中检测到3-mga,并且ko小鼠的含量高于wt小鼠;在3m的wt小鼠中未检测出3-mga,ko小鼠的3-mga含量增高,并且比1m的ko小鼠含量高,可能随着年龄的增长,3-mga的含量也有所增高。表明serac1-/-小鼠尿液有机酸中出现3-mga聚集现象。
三、serac1-/-小鼠模型的病理学分析
对小鼠肝脏组织进行组化分析,分别采用常规染色方法:he染色、糖原染色、油红o染色,以及filipin染色,同时也采用透射电镜观察serac1-/-小鼠肝脏组织线粒体形态及数量的变化。
he染色的结果发现serac1-/-小鼠肝脏组织中出现炎性细胞浸润,有些肝脏组织细胞肿胀,细胞膜破裂;从糖原染色的结果发现ko小鼠肝脏组织中糖原增加,同时可见细胞空泡状变化;油红o染色结果发现ko小鼠与wt小鼠肝脏组织相比脂肪滴增加;在胆固醇染色结果中发现随着年龄的增长,ko小鼠肝脏组织中有胆固醇聚集。以上数据表明,同megdel综合征患者的病理学结果类似,我们发现随着年龄的增长serac1-/-小鼠肝脏组织中糖原和脂肪滴有轻微聚集,并且肝脏组织细胞中有胆固醇的聚集。采用透射电镜观察肝脏组织的线粒体形态及数量,发现ko小鼠肝脏组织细胞中线粒体形态改变,线粒体的嵴数量减少,并且呈环形分布。
总上所述,serac1-/-小鼠肝脏组织的细胞形态出现一定的损伤;serac1-/-小鼠肝脏组织细胞内糖原、脂肪滴和胆固醇发生聚集;serac1-/-小鼠肝脏组织中线粒体的形态部分发生肿胀,线粒体嵴有轻微减少,呈环形分布。
四、serac1-/-小鼠模型的线粒体复合体表达水平和酶活性
1)线粒体复合体酶活性检测
i、取出-80℃冰箱中的不同组织线粒体样本放置于冰上使其溶解,利用bca蛋白浓度测定试剂盒测定线粒体的浓度,计算所需要的线粒体体积,然后取相应的线粒体重悬液进行液氮反复冻融三次,每次一分钟;
ii、将反复冻融后的线粒体重新采用bca蛋白浓度测定试剂盒测定线粒体的浓度,计算上样体积:复合体i上样10μg,复合体ii/iii/iv/v上样5μg,柠檬酸合成酶上样2μg;
iii、复合体i活性测定反应体系的配制:
表6
复合体ii活性测定反应体系的配制:
表7
复合体iii活性测定反应体系的配制:
表8
复合体iv活性测定反应体系的配制:
表9
复合体v活性测定反应体系的配制:
表10
柠檬酸合成酶活性测定反应体系的配制:
表11
每一步均先配置好相应的体系后再分别加入每个检测孔,设置3个复孔;检测完成后导出并分析数据,并且复合体i/ii/iii/iv/v的功能均通过柠檬酸合成酶活性矫正后才能相互比较。
结果1:对小鼠的心脏组织进行了复合体的检测,发现3m小鼠心脏线粒体复合体i、iv的超级复合体水平有降低趋势,复合体iii的超级复合体水平有轻微增高,而复合体ii和v表达量也有轻微升高,但均无统计学差异(图6a);6m小鼠的心脏bng的结果中,我们发现各个复合体表达量均有下降,其中复合体i和iii的超级复合体表达量明显降低,复合体v也有明显降低,而ii和iv虽有降低但无统计学差异(图6b)。心脏组织酶活性检测发现,3mserac1-/-小鼠心脏组织的复合体活性均有降低的趋势,但无统计学差异(图6c);6mserac1-/-小鼠心脏组织的复合体iii的酶活性明显降低,其他复合体酶活性没有明显变化(图6d)。表明在心脏组织中,复合体iii的复合体表达量及酶活性均有明显的降低。
结果2:serac1-/-小鼠脑组织的线粒体复合体表达水平检测发现3月ko小鼠的复合体超级复合体表达水平与wt小鼠相比均有降低,除复合体v之外其他复合体表达均无统计学差异(图7a);6月大ko小鼠除了复合体iv的超级复合体有轻微降低外,其他几个复合体表达量均明显下降(图7b);线粒体复合体酶活性检测结果发现3月ko小鼠的复合体i、iii、iv的酶活性均明显的下降,ii和v的酶活性也有下降的趋势(图7c);6月大小鼠的复合体酶活性检测中,只有复合体iii明显下降,其他几个复合体均有下降趋势(图7d)。以上数据表明,在serac1-/-小鼠脑组织中,复合体表达水平变化及酶活性的变化也主要是复合体iii。
结果3:serac1-/-小鼠肌肉组织的线粒体复合体表达水平检测,发现3月ko小鼠的复合体表达水平与wt小鼠相比复合体i和iv超级复合体表达明显下降,复合体iii的超级复合体表达有下降趋势,但无统计学差异(图8a)。
6月ko小鼠除了复合体v的超级复合体没有变化外,其他几个复合体表达量均有不同水平的下降(图8b);线粒体复合体酶活性检测结果发现,3月ko小鼠的复合体i、iii、iv、v的酶活性均有下降趋势,ii的酶活性有上升的趋势,但均无统计学差异(图8c);6月ko小鼠的复合体酶活性检测中,只有复合体ii和iii明显下降,复合体iv、v有下降趋势,复合体i没有明显变化(图8d)。以上数据表明,在serac1-/-小鼠脑组织中,复合体表达水平变化及酶活性的变化也主要是复合体iii。
结果4:serac1-/-小鼠肾脏组织的线粒体复合体表达水平检测,发现3月ko小鼠的复合体表达水平与wt小鼠相比复合体没有明显的变化(图9a);6月ko小鼠除了复合体超级复合体均有上升趋势,但无统计学差异(图9b);线粒体复合体酶活性检测结果发现,3月ko小鼠的复合体iii、iv酶活性均有下降趋势,其他复合体的酶活性没有明显的变化,但均无统计学差异(图9c);6月ko小鼠的复合体酶活性检测中,只有复合体i有明显下降,其他复合体均无明显变化(图9d)。以上数据表明,在serac1-/-小鼠肾脏组织中,复合体表达水平没有明显变化。
结果5:1月ko小鼠的复合体表达水平与wt小鼠相比复合体i、iii、iv的超级复合体表达水平明显降低,ii和v有下降趋势(图10a);3月ko小鼠复合体的超级复合体表达量均有明显下降(图10b);6月ko小鼠的复合体i、iii、iv表达水平明显降低,ii和v有下降趋势(图10c);12月ko小鼠的复合体表达量均明显降低(图10d)。线粒体复合体酶活性检测结果发现,3月ko小鼠的复合体iii、iv、v酶活性明显下降,复合体i、ii的活性有下降趋势(图10e);6月ko小鼠的复合体酶活性检测结果与3月相似(图10f)。以上数据表明,在serac1-/-小鼠肝脏组织中,复合体表达量均有明显变化,酶活性检测主要是复合体iii、iv、v活性下降。
结果6:serac1-/-小鼠肝脏组织的线粒体复合体单体表达量检测,发现与wt小鼠相比1月ko小鼠的复合体i、iii、iv、v表达量明显降低,ii有下降趋势(图11a);3月ko小鼠复合体单体表达量均有明显下降(图11b);6月ko小鼠的复合体i、ii、iii、iv表达水平明显降低,复合体v的表达量有下降趋势(图11c);12月ko小鼠的复合体i、iii、v表达量明显降低,ii和iv有下降趋势(图11d)。以上数据表明,在serac1-/-小鼠肝脏组织中,复合体单体表达量均有明显变化,主要是iii、iv、v的表达量明显下降,提示serac1-/-小鼠肝脏组织的氧化磷酸化系统有明显的受损,这也可能是megdel综合征患者早期出现严重的新生儿肝衰竭症状的原因。
ko小鼠注射病毒液5周时,serac1在ko小鼠肝脏组织中表达(图11e),并且线粒体复合体的表达水平也与野生型小鼠一致,均高于ko小鼠(图11f)。表明,serac1大片段缺失确实能够导致oxphos功能异常,也确实导致了megdel综合征。
结果7:对3月小鼠的睾丸进行复合体表达量的检测。统计结果发现复合体i、ii、iv的表达量均有明显下降,复合体iii、v的表达水平仅有下降的趋势(图12a)。表明3个月大的serac1-/-小鼠的睾丸线粒体功能有一定的受损。
提取脂肪组织的线粒体蛋白,bn-page检测线粒体复合体的表达水平,发现serac1-/-小鼠的脂肪组织线粒体复合体i、iii、iv表达量下降,复合体ii和v表达量有上升趋势(图12b)。表明ko小鼠脂肪组织的线粒体复合体表达受到一定的影响。
综上所述:serac1-/-小鼠组织中,线粒体复合体的表达水平出现不同程度的变化,主要集中在复合体i、iii、iv上,并且肝脏组织的复合体表达水平明显下降,同时肝脏复合体单体水平也有明显的下降;注射aav-serac1过表达病毒的ko小鼠肝脏组织线粒体复合体表达水平回复;serac1-/-小鼠组织中,线粒体部分复合体的酶活性降低比较明显,主要是复合体iii和iv的活性受损。
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利要求书确定的保护范围内。
序列表
<110>温州医科大学
<120>一种构建serac1基因敲除小鼠模型的方法
<160>6
<170>siposequencelisting1.0
<210>1
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>1
tcagcagtccaggattggatagg23
<210>2
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>2
gcatataccacatgtaggctggg23
<210>3
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>3
agagtcacagtcagcaactctgtgc25
<210>4
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>4
agttatgcaaccaggatcaccacgg25
<210>5
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>5
agagtcacagtcagcaactctgtgc25
<210>6
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>6
tagttctacgcttcaacccctgcta25
- 还没有人留言评论。精彩留言会获得点赞!