一种突变的马达蛋白及其应用、试剂盒的制作方法
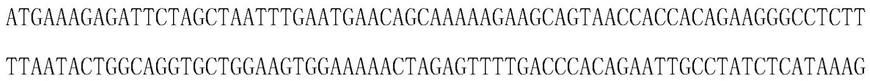
1.本发明属于分子生物学领域,具体涉及一种突变的马达蛋白及其应用和试剂盒。
背景技术:
2.马达蛋白是分布于细胞内部或细胞表面的一类蛋白质,负责细胞内部分物质或整个细胞的宏观运动。生物体内的各种组织、器官乃至整个个体的运动最终都可归结为分子马达在微观尺度上的运动。分子马达将化学键中的能量耦合转化为动能。而化学键中的能量最终来源于细胞膜或线粒体膜内外的电化学梯度。
3.解旋酶是一种常见的马达蛋白。它以单链核酸为轨道沿着核酸链定向移动,并能够解开双链核酸之间的氢键。一般由水解atp供应能量来行使解开氢键的功能。一般在生物体内dna或rna复制过程中起到催化双链dna或rna解旋的作用。解旋酶在生物体内参与多种细胞过程,如dna复制、转录、重组、dna修复等。在细菌中解旋酶的种类很多,都具有atp酶的活性。大部分的移动方向是5’到3’,但也有3’到5’方向移动的。细菌中丰富的解旋酶种类为我们发现、开发高性能酶从而行使各种细胞外功能提供了丰富的研究来源。
4.dna的体外扩增目前最成熟最常用的技术是pcr(聚合酶链式反应)技术。该技术具体描述为在目的扩增区两端设计一对引物,耐热的dna聚合酶在合适的离子、ph条件下通过变性、退火、延伸为一个温度循环使用原料dntp不断产生新的目的扩增片段。pcr扩增技术需要特定的热循环仪器(pcr仪)来进行,因此为了满足不同情境下的需求,科学家们相继开发出了多种不需要热循环仪器的等温扩增技术,如rpa(重组酶聚合酶扩增技术)、lamp(环介导等温扩增技术)、rca(滚环扩增技术)、hda(依赖解旋酶的等温扩增技术)等。它们使用不同的方式来使dna双链解链并使引物结合到模板上。rpa(重组酶聚合酶扩增技术)技术依赖于同源重组酶reca或者t4uvsx与引物形成复合物,复合物在双链dna上进行引物的同源序列搜寻,当搜寻到与引物序列一致的模板序列时即将该部分的双链模板打开从而使引物结合上去,继而进行聚合酶延伸,从而完成扩增。我们在实验测试中惊奇地发现该技术的一个致命缺点,即同源重组酶reca或者t4uvsx不能够对dna和rna的杂合链进行序列搜寻并插入引物。因此对rna的逆转录产物(dna和rna的杂合链)进行扩增时其扩增灵敏度非常低。lamp(环介导等温扩增技术)技术依赖于比较高的反应温度,如60℃
‑
65℃之间。在该比较高的反应温度时双链dna的“呼吸作用”会加强。也就是说在该温度时双链dna会偶尔部分解链成部分单链,过一会儿会再自动变成双链。在单链和双链互相转换的过程中就为引物的结合提供了机会。在该技术方案中dna发生“呼吸作用”的机会本身就不高并且不可控,因此其扩增灵敏度也不高。并且在扩增dna和rna的杂合链方面也同样有劣势。因为dna和rna的杂合链相比于dna和dna形成的双链结合更加稳定,更不容易发生“呼吸作用”。因此,其扩增灵敏度又会变低。另一方面,该技术往往需要四条甚至六条引物参与反应,引物多比较容易造成假阳性扩增现象。rca(滚环扩增技术)技术往往也需要高温变性才可以使双链打开使引物结合上去,其滚环复制扩增的特点也使其应用受诸多限制。hda(依赖解旋酶的等温扩增技术)技术是使用解旋酶使dna双链打开随后引物结合上去并由聚合酶进行扩增的技术。本
发明惊奇地发现在该等温扩增技术中解旋酶和聚合酶的相互配合对扩增起到非常关键的作用。并且经过深入分析其原理后成功筛选并且开发了一种新的突变马达蛋白。在该突变马达蛋白参与下该技术扩增性能远远超过现有的等温扩增技术。
技术实现要素:
5.解旋酶是马达蛋白中的一大类酶。共分为六个家族,三个超级家族sf1、sf2、sf3,两个小家族rho和dnab类似家族,和一个分支家族aaa+家族。本发明中描述的解旋酶结构上属于sf1家族。该家族的酶在结构上普遍分为四个结构域,分别是1a,1b,2a,2b。四个结构域行使不同的功能。经过结构功能分析科学家们普遍认为1b和2b与双链dna的结合关系比较密切,1a和2a与atp水解和dna解旋密切相关。各个结构域相互配合最终行使对双链dna的结合和解旋的功能。
6.该类解旋酶在分子生物学中应用广泛。以hda(依赖解旋酶的等温扩增技术)技术为例,其主要通过两种酶来实现dna的等温扩增。首先就用到解旋酶,其结合到双链dna上,并通过水解atp供能来将双链dna之间的氢键打开。这样使得特异性的引物能够结合到单链dna模板上,然后由dna聚合酶对引物进行延伸,从而完成扩增。在这个体系中,解旋酶和dna聚合酶两者的作用需要有近乎完美的协调能力。首先解旋酶对双链dna进行解旋的速度和能力难以人为把控。以解旋酶tte uvrd为例,其解旋的效率往往远远大于dna聚合酶的聚合效率。当体系中模板dna分子数很少时,解旋酶要想高效的作用于很少的底物双链dna分子,就需要解旋酶的浓度也比较高。解旋酶相对于dna聚合酶处于比较强的工作优势,这样就会容易出现dna的解旋效率进一步大于dna聚合酶的聚合效率,引物结合到dna上去以后还未等dna聚合酶聚合,引物就可能被解旋酶从模板dna上解下来,这样就会造成引物结合不上去导致扩增灵敏度低的问题。
7.为了解决上述技术问题,本发明提供了突变的马达蛋白,来自sf1家族,分为1a、1b、2a、2b四个结构域;采用以下方式中的至少一种突变方式获得:
8.(1)通过1b和/或2b结构域的碱基突变使得马达蛋白提高对dna双链的亲和力;
9.(2)通过1a和/或2a结构域的碱基突变使得马达蛋白降低对dna的解链速度。
10.所述的突变的马达蛋白中野生型马达蛋白包括解旋酶tte uvrd、tth uvrd、pcra、rep中的至少一种。tte uvrd的氨基酸序列如seq id no.2所示。
11.所述的突变的马达蛋白,具体如下:
12.1a和/或2a突变包括:r71s突变、r597k突变、t34i突变、y613i突变中的至少一种。
13.所述的突变的马达蛋白,具体如下:
14.1b和/或2b突变包括:i418g突变、p414k突变、r415i突变、i418l突变、a421k突变中的至少一种。
15.所述的突变的马达蛋白,包括1a和/或2a突变、1b和/或2b突变中的至少两种的任意组合;
16.1a和/或2a突变包括:r71s突变、r597k突变、t34i突变、y613i突变;
17.1b和/或2b突变包括:i418g突变、p414k突变、r415i突变、i418l突变、a421k突变。
18.本发明发现相较于野生型酶30拷贝模板扩增不出的情况,1b/2b结构域中i418g突变酶由于对体系中很少量的dna模板(如低至30拷贝)有更高的亲和抓取的能力,因此已经
能够对更低拷贝数模板进行有效扩增。
19.本发明发现相较于野生型酶30拷贝模板扩增不出的情况,1b/2b结构域中p414k突变、r415i突变、i418l突变、a421k突变由于对体系中很少量的dna模板(如低至30拷贝)有更高的亲和抓取的能力,因此已经能够对更低拷贝数模板进行有效扩增。
20.本发明发现1a/2a结构区域中r71s突变酶相较野生型酶落后3个ct值,即r71s突变一定程度上降低了酶的解旋活性。
21.本发明发现1a/2a结构区域中r597k突变、t34i突变、y613i突变酶ct值大于野生型酶ct值,即r597k突变、t34i突变、y613i突变一定程度上降低了酶的解旋活性。
22.本发明发现1a/2a结构区域中t34i突变和1b/2b结构域中i418l突变的双突变酶能够对低至10拷贝的模板进行等温扩增。
23.本发明发现1b/2b结构域中p414k突变、r415i突变、i418l突变、a421k突变和1a/2a结构区域中r597k突变、t34i突变、y613i突变的七突变酶相比于野生型酶能够对低至2拷贝的模板进行有效等温扩增。
24.进一步地,本发明所述的突变的马达蛋白,还包括相似度不低于95%,能够达到相似功能的氨基酸序列;所述的相似功能包括:提高对dna双链的亲和力和/或降低对dna的解链速度。
25.本发明还提供了表达上述的突变的马达蛋白的核酸序列。
26.本发明还提供了所述的突变的马达蛋白的应用,用于dna扩增体系。
27.进一步地,用于依赖解旋酶的dna等温扩增体系。
28.本发明还提供了一种依赖解旋酶的dna等温扩增试剂盒,包含上述的突变的马达蛋白。
29.本发明突变马达蛋白本质上可以是解旋酶的一种。本发明提出的突变的马达蛋白的应用只是其中的举例,实际突变马达蛋白的应用并不局限于此。
30.本发明提供的经过筛选和优化的突变马达蛋白,解决了野生型马达蛋白的矛盾。马达蛋白主要有四个结构域构成,分别为1a、1b、2a和2b。1a和2a结构域主要有促进atp水解、协调镁离子活动、形成π环堆叠形状最终使双链dna解旋等作用。1b和2b结构域主要行使捕获双链dna并与双链dna牢牢结合的作用。本发明中主要对该马达蛋白进行了两种突变作用。第一种突变主要在1b和2b结构域,其作用是提高了该马达蛋白对dna双链的亲和力,使得很少的马达蛋白的量就可以与比较少的模板进行高效的结合并解链。第二种突变主要在1a和2a结构域,其作用主要是突变本身降低了该马达蛋白对dna的解链速度。最终达到的效果是降低了马达蛋白的量,但是增加了马达蛋白对dna模板底物的亲和力,减慢了马达蛋白的解链速度,从而能够精确调控该马达蛋白与dna聚合酶之间的功能协作,从而使其在核酸扩增中的应用(如基于突变马达蛋白的体外模拟等温扩增技术),尤其是低拷贝dna模板时达到检测灵敏度更高的效果。
附图说明
31.图1为野生型解旋酶tte uvrd及其各结构域;
32.图2为野生型解旋酶tte uvrd和使用商业化试剂盒相比的扩增结果;
33.图3为野生型解旋酶tte uvrd和i418g突变酶的三维结构对比;
34.图4为野生型解旋酶tte uvrd和i418g突变酶30拷贝模板扩增情况;
35.图5为实施例3中11种突变的解旋酶tte uvrd三维结构;
36.图6为实施例3中11种突变的解旋酶tte uvrd30拷贝模板扩增情况;
37.图7为野生型解旋酶tte uvrd和r71s突变酶的三维结构对比;
38.图8为野生型解旋酶tte uvrd和r71s突变酶200拷贝质粒扩增情况;
39.图9为实施例5中9种突变的解旋酶tte uvrd三维结构;
40.图10为实施例5中9种突变的解旋酶tte uvrd200拷贝质粒扩增情况;
41.图11为野生型解旋酶tte uvrd双突变三维结构;
42.图12为野生型解旋酶tte uvrd和双突变酶均使用10拷贝模板进行扩增情况;
43.图13为野生型解旋酶tte uvrd七突变三维结构;
44.图14为野生型解旋酶tte uvrd和七突变酶均分别使用30拷贝、10拷贝、2拷贝模板扩增情况。
具体实施方式
45.以下结合实施例旨在进一步解释和说明本发明,而非限制本发明。
46.实施例1
47.商业化试剂盒和野生型tte uvrd在等温扩增中的应用
48.野生型tte uvrd dna序列如下:
49.seq id no.1:
50.[0051][0052]
野生型tte uvrd氨基酸序列如下:
[0053]
seq id no.2:
[0054]
mkeilanlneqqkeavtttegpllilagagsgktrvlthriaylikekkvspsnilaitftnkaaeemktrvenllgyvgdlwvstfhsacvrilrrdidklgydrnfvifdttdqkalvqeclkeldlsekqypikmvlnaissakdkmvypddyidffgdtyrnrkikeiyklyqhklkkinaldfddiiiktielfkenpeilefyqrkfryimvdeyqdtntpqyyfvnllaqrhrnlcvvgdddqsiygwrgadvrnilnfekdypeakvikleqnyrstkiileaanhvidnnvyrkkkslwtqnkegekivlcelenereeaefviqeiiklkerenrsfkdfailyrtnaqsrpfeealmkvkvpykvvgalrfydrkeikdilaylrlivnpyddisfkrivnvprrgigpatiealekvarekdtslffaiedlknarnkgsllqfkqfildlidkkdamsvsdlikyileqtgyieelkreeseeaegrienlneflnaayefeessedksleaflagitlvsdidmagdigesvvlmtlhsakglefpvvfmvgmeeglfpsyssfeddheleeerrlcyvgitrskerlyltyarqrnlygrsqynsysrfiseiperlivryniptskktgfvsvhtfsdvyersfslgdkvehkiwgigtvvkvegeeitvafpnvgikkldlrfapikais
[0055]
野生型tte uvrd dna序列用限制性内切酶ndei和xhoi克隆到pet28a载体,构成pwy
‑
uvrd。镍柱组氨酸标记纯化。消化组氨酸标签后进一步离子交换柱monos纯化。溶于20mm tris(ph 8),150mm nacl,0.1mm edta,5mm dtt,5%甘油和3%异丙醇中储存。下面实施例中各种突变型酶也用此方法表达纯化。
[0056]
采用人源白细胞基因组dna的braf基因一段区域进行测试检测:
[0057]
检测序列如下:
[0058]
seq id no.3:
[0059]
ttcttaaggggatctcttcctgtatccctctcaggcataaggtaatgtacttagggtgaaacataaggttttctttttctgtttggcttgacttgacttttttactgtttttatcaagaaaacacttggtagacgggactcgagtgatgattgggagattcctgatgggcagattacagtgggacaaagaattggatctggatcatttggggggaacagtctcaagggaaagtggcatggtaagtatgtaatgtggtgacattgtgacaagtcataataggatatgtttaacaacttttattttgtaaaaaatatcatcaaaggaaatattcactgttcgcatcaataaactattttgattagtttcaggactcctccaaaagtttctaacaaaaattatgggaaataaaaactgttcacag
[0060]
对应检测引物:
[0061]
seq id no.4:
[0062]
f:
[0063]
ttatcaagaaaacacttggtag
[0064]
seq id no.5:
[0065]
r:
[0066]
caatgtcaccacattacata
[0067]
对照实验使用isoamp ii universal thda kit(biohelix,#h0110s)。
[0068]
野生型tte uvrd实验反应体系:
[0069]
成分浓度kcl10mmtris
‑
hcl(ph8.8)20mmseq id no.4
‑
f350nmseq id no.5
‑
r350nm硫酸镁8mm
氯化钠80mmdntps0.8mm eachatp6mmwild tte uvrd200ngbst dna聚合酶20u
[0070]
同时以30拷贝、50拷贝、80拷贝、100拷贝、200拷贝为模板进行扩增。
[0071]
实验条件65℃,2小时。
[0072]
实验结果见图2:
[0073]
实施例1结果分析:本实施例说明使用野生型解旋酶tte uvrd和使用商业化试剂盒相比效果相差不大。在80拷贝以上模板时均能稳定检出,在50拷贝模板时偶尔可以检出,在30拷贝以下均较难检出。
[0074]
实施例2
[0075]
1b/2b结构域中i418g突变对马达蛋白抓取双链dna的影响
[0076]
g417、i418、g419及其附近区域构成三维结构图中的圆环部分,该部分是解旋酶结合抓取双链dna的关键区域。图3展示了野生型酶和i418g突变酶的三维结构对比。
[0077]
i418g突变酶的表达实验方法同实施例1中野生型酶的表达方法。等温扩增反应的实验体系、目标区域、引物等也同实施例1。
[0078]
实验结果见图4。
[0079]
实施例2结果分析:相较于野生型酶30拷贝模板扩增不出的情况,i418g突变酶由于对体系中很少量的dna模板(如低至30拷贝)有更高的亲和抓取的能力,因此已经能够对更低拷贝数模板进行有效扩增。
[0080]
实施例3
[0081]
1b/2b结构域中p414k、r415i、r416i、g417e、i418l、i418r、i418t、i418v、p420v、a421k、t422d突变对马达蛋白抓取双链dna的影响
[0082]
图5展示了11种单个点突变酶的三维结构。
[0083]
11种单个突变酶的表达实验方法同实施例1中野生型酶的表达方法。等温扩增反应的实验体系、目标区域、引物等也同实施例1。
[0084]
实验结果见图6。
[0085]
实施例3结果分析:相较于野生型酶30拷贝模板扩增不出的情况,p414k突变、r415i突变、i418l突变、a421k突变由于对体系中很少量的dna模板(如低至30拷贝)有更高的亲和抓取的能力,因此已经能够对更低拷贝数模板进行有效扩增。
[0086]
实施例4:
[0087]
1a/2a结构区域r71s突变对马达蛋白解旋能力的影响
[0088]
图7展示出了野生型和r71s突变型酶的三维结构。
[0089]
采用新冠病毒covid
‑
19序列s基因的一段作为检测序列。其待检测区域序列如下:
[0090]
seq id no.6:
[0091][0092]
引物序列:
[0093]
seq id no.7:
[0094]
f:
[0095]
ataatgaaaatggaacca
[0096]
seq id no.8:
[0097]
r:
[0098]
tggttggactctaaagtta
[0099]
探针序列:
[0100]
seq id no.9:
[0101]
p:
[0102]5’
fam
‑
gtgcacttgaccctct
‑
bhq1 3’[0103]
采用含有seq id no.6序列的质粒作为模板。
[0104]
野生型酶和突变型酶实验反应体系:
[0105]
成分浓度kcl10mmtris
‑
hcl(ph8.8)20mmseq id no.7
‑
f350nmseq id no.8
‑
r350nmseq id no.9
‑
p500nm硫酸镁8mm氯化钠80mmdntps0.8mm eachatp6mm野生型酶/突变型酶200ngbst full length dna聚合酶20u
[0106]
同时以200拷贝质粒为模板进行扩增。
[0107]
实验条件65℃,2小时。每45秒钟收集荧光信号。
[0108]
实验结果见图8。
[0109]
实施例4分析:
[0110]
r71s突变酶相较野生型酶落后3个ct值,即r71s突变一定程度上降低了酶的解旋活性。
[0111]
实施例5:
[0112]
1a/2a结构区域k33t、r283l、r597k、t34i、d219h、e220v、y613g、y613i、y613k突变对马达蛋白解旋能力的影响
[0113]
图9展示了9种单个点突变酶的三维结构。
[0114]
实验方法同实施例4。
[0115]
实验结果见图10。
[0116]
实施例5分析:
[0117]
9种突变中r597k突变、t34i突变、y613i突变酶ct值大于野生型酶ct值,k33t突变、r283l突变、d219h突变、e220v突变、y613g突变、y613k突变酶ct值小于野生型酶ct值,即
r597k突变、t34i突变、y613i突变一定程度上降低了酶的解旋活性。
[0118]
实施例6:
[0119]
1b/2b结构域中i418l突变和1a/2a结构区域中t34i突变对等温扩增的影响
[0120]
图11展示出了双突变(i418l突变和t34i)酶的三维结构。
[0121]
实验方法同实施例4。
[0122]
野生型酶和双突变酶均使用10拷贝模板进行扩增。
[0123]
实验结果见图12。
[0124]
实施例6分析:
[0125]
1a/2a结构区域中t34i突变和1b/2b结构域中i418l突变的双突变酶能够对低至10拷贝的模板进行等温扩增。
[0126]
实施例7:
[0127]
1b/2b结构域中p414k突变、r415i突变、i418l突变、a421k突变和1a/2a结构区域中r597k突变、t34i突变、y613i突变对等温扩增的影响
[0128]
图13展示出了七突变酶的三维结构。
[0129]
实验方法同实施例4。
[0130]
野生型酶和七突变酶均分别使用30拷贝、10拷贝、2拷贝模板进行扩增。
[0131]
实验结果见图14。
[0132]
实施例7分析:
[0133]
1b/2b结构域中p414k突变、r415i突变、i418l突变、a421k突变和1a/2a结构区域中r597k突变、t34i突变、y613i突变的七突变酶相比于野生型酶能够对低至2拷贝的模板进行有效等温扩增。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1