基因工程菌及其应用和以葡萄糖为原料生产阿洛酮糖的方法与流程
1.本发明涉及生物工程领域,具体涉及一株基因工程菌,所述基因工程菌在以葡萄糖为原料生产阿洛酮糖中的应用,以及一种以葡萄糖为原料生产阿洛酮糖的方法。
背景技术:
2.d-阿洛酮糖(d-psicose)是d-果糖(d-fructose)c-3的差向异构体,其甜度相当于蔗糖的70%,热量相当于蔗糖0.3%,是一种具有特殊保健功能的新型功能性单糖。d-阿洛酮糖具有与蔗糖相近的口感及容积特性,并与蔗糖一样可与食物中的氨基酸或蛋白质发生美拉德反应,可作为食品中蔗糖理想的替代品。动物实验显示,d-阿洛酮糖具有控制2型糖尿病患者血糖的功效,同时对肥胖等相关疾病也一定的预防作用。动物与人体实验证明,d-阿洛酮糖没有任何不良副作用。2011年8月,美国食品与药物管理局(fda)确定d-阿洛酮糖为公认安全使用物质(gras),并可作为食品或食品添加剂的组成成分。d-阿洛酮糖在膳食、保健、医药等领域拥有广阔的应用前景。
3.目前生产d-阿洛酮糖的主要方法是酶法转化,以d-果糖为底物,在d-阿洛酮糖-3-差向异构酶的催化下生成d-阿洛酮糖。d-阿洛酮糖-3-差向异构酶的特点在于具有反应平衡常数,一般在28-33%之间。d-果糖是以葡萄糖为底物,在葡萄糖异构酶的催化下转化而来。葡萄糖异构酶也具有反应平衡常数,一般在42-45%之间。d-阿洛酮糖-3-差向异构酶和葡萄糖异构酶的最适反应温度一般都在50-60℃之间,并且都具有金属离子依赖性。因此,在葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶共同存在的情况下,以葡萄糖为底物一步转化生成d-阿洛酮糖,从而减少中间产物d-果糖的纯化步骤,对于减少d-阿洛酮糖的生产成本具有重要的生产意义,而采用能够共表达葡萄糖异构酶和阿洛酮糖差向异构酶的菌株是能够实现一步法转化的途径。
技术实现要素:
4.本发明的目的是为了提供一株基因工程菌,该基因工程菌能够共表达葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶,用于一步法将葡萄糖转化为阿洛酮糖,减少中间产物d-果糖的纯化步骤,对于减少d-阿洛酮糖的生产成本具有重要的生产意义。
5.为了实现上述目的,本发明第一方面提供一株基因工程菌,所述基因工程菌能够共表达葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶。
6.优选地,所述葡萄糖异构酶活性为具有seq id no.2所示氨基酸序列的酶。
7.优选地,编码所述葡萄糖异构酶的基因具有seq id no.1所示的核苷酸序列。
8.优选地,所述d-阿洛酮糖-3-差向异构酶为具有seq id no.4所示氨基酸序列的酶。
9.优选地,编码所述d-阿洛酮糖-3-差向异构酶的基因具有seq id no.3所示的核苷酸序列。
10.优选地,所述葡萄糖异构酶以葡萄糖异构酶活性聚集体的形式存在,所述葡萄糖
异构酶活性聚集体包含葡萄糖异构酶和自聚集短肽。
11.优选地,所述d-阿洛酮糖-3-差向异构酶以d-阿洛酮糖-3-差向异构酶活性聚集体的形式存在,所述d-阿洛酮糖-3-差向异构酶活性聚集体包含d-阿洛酮糖-3-差向异构酶和自聚集短肽。
12.本发明第二方面提供如上所述的基因工程菌在以葡萄糖为原料生产阿洛酮糖中的应用。
13.本发明第三方面提供一种以葡萄糖为原料生产阿洛酮糖的方法,该方法包括在如上所述的基因工程菌,或所述基因工程菌表达的葡萄糖异构酶或葡萄糖异构酶活性聚集体以及d-阿洛酮糖-3-差向异构酶或d-阿洛酮糖-3-差向异构酶的存在下,将葡萄糖转化为阿洛酮糖。
14.本发明所述的基因工程菌能够共表达葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶,用于一步法将葡萄糖转化为阿洛酮糖,减少中间产物d-果糖的纯化步骤,对于减少d-阿洛酮糖的生产成本具有重要的生产意义。
15.在优选的条件下,比如优选的序列和菌株种类以及优选的酶的形态,能够进一步提高利用葡萄糖生产阿洛酮糖的产量以及葡萄糖的转化率。
附图说明
16.图1为实施例1中的pet30-rum55-gslinker-surgl质粒图谱;
17.图2是实施例1中葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的电泳图;
18.图3为实施例2中的pma5-rum55-gslinker-surgl质粒图谱;
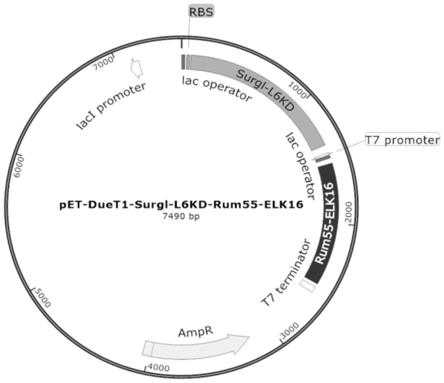
19.图4为实施例3中的pet-duet1-surgl-l6kd-rum55-elk16质粒图谱;
20.图5是实施例3中surgl-l6kd和rum55-elk16活性聚集体共表达蛋白电泳图。
具体实施方式
21.在本文中所披露的范围的端点和任何值都不限于该精确的范围或值,这些范围或值应当理解为包含接近这些范围或值的值。对于数值范围来说,各个范围的端点值之间、各个范围的端点值和单独的点值之间,以及单独的点值之间可以彼此组合而得到一个或多个新的数值范围,这些数值范围应被视为在本文中具体公开。
22.本发明第一方面提供一株基因工程菌,所述基因工程菌能够共表达葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶。
23.在本发明中,所述葡萄糖异构酶的种类可以不受特别的限制,只要具有葡萄糖异构酶活性即可。
24.所述葡萄糖异构酶的来源菌株可以不受特别的限制,只要该菌株能够产生葡萄糖异构酶即可;优选地,所述葡萄糖异构酶来源于链霉菌(streptomyces)、热杆菌(fervidobacteriumsp.)和酸性嗜热菌(acidothermuscellulolyticus)中的至少一种。
25.优选地,所述葡萄糖异构酶来源于热杆菌,更优选来源于冈瓦红热杆菌(fervidobacteriumgondwanense)。
26.在本发明的一种优选的实施方式中,所述葡萄糖异构酶为(a)-(e)中任意一项所述的酶:
id no.4所示氨基酸序列的酶。
46.本发明中的d-阿洛酮糖-3-差向异构酶氨基酸序列可以是相比于seq id no.4所示氨基酸序列在如下位点突变获得的序列:第23位、第67位、第74位、第112位、第164位、第177位、第180位、第182位、第183位、第190位、第194位、第195位、第213位、第215位、第218位、第221位、第239位、第253位和第263位中的至少一位;优选具有如下突变:l183m、n190t、v67i、t164e、g253c、w263s、g213s、a112f、c180v/i182v/l183m、d74e、s215c、g213s/l183m、g213s/s215c、c221g、r218m、i23l/g213s、i23l、d194c、d239c、s177c和s195c中的至少一种。
47.为了方便纯化,还可以采用本领域常见的标签对氨基酸序列进行添加修饰,举例来说,可以通过在氨基酸序列的氨基末端和/或羧基末端连接有标签的氨基酸序列(如poly-arg、poly-his、flag、strep-tagⅱ和c-myc中的至少一种)而获得。所述标签不会影响酶的活性,在实际应用过程中,可以根据需求选择是否添加标签。
48.在本发明中,所述葡萄糖异构酶或所述d-阿洛酮糖-3-差向异构酶的氨基末端还可以连接信号序列。所述信号序列可以来自地衣芽孢杆菌、解淀粉芽孢杆菌和枯草芽孢杆菌,但不限于此。
49.在本发明中,仍具有酶活力是指在相同的测定条件下,由(a)衍生的酶仍具有酶活力,其酶活力与(a)的酶活力之间的百分比(相对活性)不低于60%,优选不低于70%(或80%,或90%,或95%,或96%,或97%,或98%,或99%,或100%)。
50.上述的葡萄糖异构酶或d-阿洛酮糖-3-差向异构酶可以通过人工合成得到,也可以先合成其编码基因,再通过生物表达获得。
51.优选地,编码所述d-阿洛酮糖-3-差向异构酶的基因具有seq id no.3所示的核苷酸序列或seq id no.3所示的核苷酸序列经过取代、缺失或添加一个或几个核苷酸残基且仍能够表达具有d-阿洛酮糖-3-差向异构酶活性的酶的核苷酸序列。
52.在本发明的一种优选的实施方式中,编码所述d-阿洛酮糖-3-差向异构酶的基因具有seq id no.3所示的核苷酸序列。
53.如上所述,相应地,核苷酸序列的5'端和/或3'端还可以连接有标签的编码序列。
54.本领域技术人员可用公知的人工化学合成的方法来合成有关氨基酸或核苷酸序列。
55.优选地,所述葡萄糖异构酶以葡萄糖异构酶活性聚集体的形式存在,所述葡萄糖异构酶活性聚集体包含葡萄糖异构酶和自聚集短肽。
56.在本发明中,所述自聚集短肽可以是本领域现有的自聚集短肽,包括但不限于口蹄疫病毒衣壳蛋白vp1(foot-and-mouth disease virus capsid protein vp1)、人β-淀粉样蛋白aβ42(humanβ-amyloid peptide aβ42)、麦芽糖结合蛋白male(maltose-binding protein)、麦芽糖结合蛋白突变体male31、纤维素结合域(cellulose-binding domain,cbd
clos
)、类弹性蛋白多肽(elastin-like polypeptide,elp)、elk16、l6kd和18a。
57.优选地,所述葡萄糖异构酶活性聚集体中葡萄糖异构酶和自聚集短肽中间还插入有连接肽。
58.所述连接肽可以是本领域现有的连接肽,优选地,所述连接肽选自pt-linker、gslinker、(gly)6、(gly)8和(eaaak)n中的至少一种;其中,n=1-3。
59.所述的连接肽与自聚集短肽的拼接顺序可以是连接肽-自聚集短肽或者自聚集短
肽-连接肽,相应的自聚集短肽与葡萄糖异构酶或d-阿洛酮糖-3-差向异构酶融合表达,自聚集短肽可位于葡萄糖异构酶或d-阿洛酮糖-3-差向异构酶的c端或者n端。
60.应当理解的是,当所述基因工程菌表达的是葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶时,两个酶也可以通过自连接肽连接。
61.优选地,所述d-阿洛酮糖-3-差向异构酶以d-阿洛酮糖-3-差向异构酶活性聚集体的形式存在,所述d-阿洛酮糖-3-差向异构酶活性聚集体包含d-阿洛酮糖-3-差向异构酶和自聚集短肽。
62.优选地,所述d-阿洛酮糖-3-差向异构酶活性聚集体中d-阿洛酮糖-3-差向异构酶和自聚集短肽中间还插入有连接肽。
63.优选地,所述葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体中的自聚集短肽和连接肽可以相同或不同。
64.优选地,所述基因工程菌为大肠杆菌或枯草芽孢杆菌,更优选为枯草芽孢杆菌。
65.所述基因工程菌的制备方法可以为本领域常规的制备方法,比如可以先通过载体拼接编码基因,构建表达载体,然后将表达载体转化到受体菌中,得到目标基因工程菌。
66.在本发明中,可以在本领域已知的各种载体中完成拼接,构建表达载体,如市售的各种质粒、粘粒、噬菌体及反转录病毒等。所述载体优选为可在受体菌中过量表达的诱导型载体,其具有诱导目的蛋白在受体菌体内自聚集为活性聚集体的能力。
67.当所述受体菌为大肠杆菌时,载体优选为pet30a、pet28a、pet24a、pet21a、pet22b、pet32a、pet14a、pbad和pcold i中的至少一种。
68.当所述受体菌为枯草芽孢杆菌时,载体优选为pma5、phy300plk、paxo1、phthisq和phthisr中的至少一种。
69.表达载体构建可采用能够在载体多克隆位点具有切割位点的各种核酸内切酶进行酶切获得线性载体,与采用相同核酸内切酶切割的基因片段连接,获得表达载体。
70.可以通过本领域常规的方法将所述表达载体转化、转导或者转染到受体菌中,如热激法、氯化钙法和电转化法等。
71.当所述受体菌为大肠杆菌时,优选地,所述受体菌选自e.colidh5α、e.colibl21、e.coli bl21(de3)、e.coli bl21(de3)/plyss、e.coli rosetta、e.coli rosetta(de3)、e.coli jm109、e.coli jm110和e.coli top10中的至少一种。
72.当所述受体菌为枯草芽孢杆菌时,优选地,所述受体菌为枯草芽孢杆菌bs1012、bs168、wb600n和wb800n中的至少一种。
73.在本发明的一种优选的实施方式中,基因工程菌的制备方法包括以下步骤:(1)将葡萄糖异构酶的编码基因、d-阿洛酮糖-3-差向异构酶的编码基因通过连接肽的编码基因进行拼接,构建表达载体;(2)将步骤(1)得到的表达载体转化到受体菌中,得到基因工程菌;其中,连接肽的编码基因位于葡萄糖异构酶的编码基因和d-阿洛酮糖-3-差向异构酶的编码基因之间;其中,所述连接肽选自gslinker、(gly)6和(gly)8、(eaaak)n中的至少一种;其中,n=1-3。
74.在本发明的一种优选的实施方式中,基因工程菌的制备方法包括以下步骤:(1)将葡萄糖异构酶的编码基因与第一自聚集短肽的编码基因进行拼接,将d-阿洛酮糖-3-差向异构酶的编码基因与第二自聚集短肽的编码基因进行拼接,将两个拼接基因构建至含有两
surgl接种在含有50μg/ml卡那霉素抗性的lb液体培养基中,培养至od
600
达到0.4-0.6,加入终浓度为0.5mm iptg,在25℃、200rpm条件下诱导18小时。离心收集菌体,用ph=8.0的tris-hcl(50mm)缓冲液重悬,采用高压均质或超声破碎法进行菌体破碎,破碎后10000rpm离心10min,所得样品即为含葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的粗酶液。蛋白电泳结果如图2所示,箭头标记为rum55-surgl融合蛋白(seq id no.11)。
108.(5)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的活性测定
109.1ml反应体系如下:rum55-surgl粗酶液、50mm tris-hcl缓冲液(ph=8.0)、40mg/ml葡萄糖、和0.1mm cocl2,50℃条件下反应,每隔1h取样,hplc检测葡萄糖、果糖和阿洛酮糖的含量,结果如表2所示。rum55-surgl融合蛋白兼具葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶活性。
110.表2
[0111][0112][0113]
实施例2
[0114]
本实施例用于说明葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶共表达枯草芽孢杆菌工程菌的构建。
[0115]
(1)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合片段扩增
[0116]
将d-阿洛酮糖-3-差向异构酶(rum55)基因与葡萄糖异构酶(surgl)基因通过gslinker进行扩增连接。引物如表3所示,下划线碱基为限制性内切酶识别位点。扩增步骤如下:
[0117]
①
以pet30-rum55质粒为模板,以pma5-bamhi-rum55-f和gslinker-rum55-r为引物进行pcr扩增,得到片段4;
[0118]
②
以pet30-surgl质粒为模板,以gslinker-surgl-f和pma5-mlui-surgl-r为引物进行pcr扩增,得到片段5;
[0119]
③
以片段4和片段5为模板,以pma5-bamhi-rum55-f和pma5-mlui-surgl-r为引物进行overlap pcr扩增,得到含有pma5载体序列的rum55-gslinker-surgl融合片段6(seq id no.12)。
[0120]
表3
[0121][0122]
(2)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合表达载体构建
[0123]
将pma5空载体用限制性内切酶bamhi和mlui进行双酶切,并对酶切后的载体片段进行切胶回收。使用peasy-uni seamless cloning and assembly kit(北京全式金生物技术有限公司),将回收的载体片段与步骤(1)中获得的片段6进行混合,反应15min后,转化至trans-t1感受态细胞(北京全式金生物技术有限公司),通过菌落pcr、酶切及测序分析鉴定阳性转化子,得到质粒pma5-rum55-gslinker-surgl,如图3所示。
[0124]
(3)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合表达工程菌构建
[0125]
将步骤(2)中构建的质粒转化至bs168感受态细胞,通过菌落pcr、酶切及测序分析鉴定阳性转化子,得到bs168/pma5-rum55-gslinker-surgl。
[0126]
(4)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的表达
[0127]
将步骤(3)中构建的工程菌bs168/pma5-rum55-gslinker-surgl用lb培养基在37℃下过夜活化,然后按1:100比例将活化菌液转化至新鲜的lb培养基中,37℃培养,待od600达到0.5,将菌液转接至kb培养基中,30℃发酵24h,收集菌体,用ph=8.0的tris-hcl(50mm)缓冲液重悬,采用高压均质或超声破碎法进行菌体破碎,破碎后10000rpm离心10min,所得样品即为含葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的粗酶液。
[0128]
(5)葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶融合蛋白的活性测定
[0129]
1ml反应体系如下:rum55-surgl粗酶液、50mm tris-hcl缓冲液(ph=8.0)、40mg/ml葡萄糖、和0.1mm cocl2,50℃条件下反应,每隔1h取样,hplc检测葡萄糖、果糖和阿洛酮糖的含量,结果如表4所示。rum55-surgl融合蛋白兼具葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶活性。
[0130]
表4
[0131]
反应时间h葡萄糖浓度g/l果糖浓度g/l阿洛酮糖浓度g/l040.07——137.711.780.51233.295.221.49331.106.931.98428.269.132.61524.8011.823.38621.9914.014.00719.2116.174.62
[0132]
实施例3
[0133]
本实施例用于说明葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体共表达大肠杆菌工程菌的构建。
[0134]
(1)pet-duet1-surgl-l6kd载体构建
[0135]
将surgl-ptlinker-l6kd(以下简称“surgl-l6kd”)基因构建至pet-duet1的mcs1上,引物如表5所示,下划线碱基为限制性内切酶识别位点。步骤如下:
[0136]
①
以实验室前期构建的pet30-surgl-l6kd质粒为模板,以pet-duet1-ncoi-surgl-f和pet-duet1-noti-l6kd-r为引物进行pcr扩增,得到含有pet-duet1-mcs1载体序列的surgl-l6kd片段7(seq id no.15);
[0137]
②
将pet-duet1空载体用限制性内切酶ncoi和noti进行双酶切,并对酶切后的载体片段进行切胶回收。使用peasy-uni seamless cloning and assembly kit(北京全式金生物技术有限公司),将回收的载体片段与步骤
①
中获得的片段7进行混合,反应15min后,转化至trans-t1感受态细胞(北京全式金生物技术有限公司),通过菌落pcr、酶切及测序分析鉴定阳性转化子,得到质粒pet-duet1-surgl-l6kd。
[0138]
表5
[0139][0140][0141]
(2)pet-duet1-surgl-l6kd-rum55-elk16载体构建
[0142]
将rum55-ptlinker-elk16(以下简称“rum55-elk16”)基因构建至pet-duet1-surgl-l6kd的mcs2上,引物如表6所示,下划线碱基为限制性内切酶识别位点。步骤如下:
[0143]
①
以实验室前期构建的pet30-rum55-elk16质粒为模板,以pet-duet1-ndei-rum55-f和pet-duet1-avrii-elk16-r为引物进行pcr扩增,得到含有pet-duet1-mcs2载体序列的rum55-elk16片段8(seq id no.18);
[0144]
②
将步骤(1)构建的pet-duet1-surgl-l6kd载体用限制性内切酶ndei和avrii进行双酶切,并对酶切后的载体片段进行切胶回收。使用peasy-uni seamless cloning and assembly kit(北京全式金生物技术有限公司),将回收的载体片段与步骤
①
中获得的片段8进行混合,反应15min后,转化至trans-t1感受态细胞(北京全式金生物技术有限公司),通过菌落pcr、酶切及测序分析鉴定阳性转化子,得到质粒pet-duet1-surgl-l6kd-rum55-elk16,如图4示。
[0145]
表6
[0146][0147]
(3)葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体共表达工程菌构建
[0148]
将步骤(2)中构建的质粒转化至e.colibl21(de3)感受态细胞,通过菌落pcr、酶切
及测序分析鉴定阳性转化子,得到e.colibl21(de3)/pet-duet1-surgl-l6kd-rum55-elk16。
[0149]
(4)葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体的共表达
[0150]
将步骤(3)中构建的工程菌e.coli bl21(de3)/pet-duet1-surgl-l6kd-rum55-elk16接种在含有50μg/ml卡那霉素抗性的lb液体培养基中,培养至od600达到0.4-0.6,加入终浓度为0.5mm iptg,在25℃、200rpm条件下诱导18小时。离心收集菌体,用ph=8.0的tris-hcl(50mm)缓冲液重悬,采用高压均质或超声破碎法进行菌体破碎,破碎后10000rpm离心10min,收集沉淀,所得样品即为含葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体的混合物。蛋白电泳结果如图5示,箭头标记为surgl-l6kd(seq id no.21)和rum55-elk16(seq id no.22)。
[0151]
(5)葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体的活性测定
[0152]
1ml反应体系如下:葡萄糖异构酶活性聚集体和d-阿洛酮糖-3-差向异构酶活性聚集体共表达沉淀重悬液、50mm tris-hcl缓冲液(ph=8.0)、4mg/ml葡萄糖、和0.1mm cocl2,50℃条件下反应,每隔1h取样,hplc检测葡萄糖、果糖和阿洛酮糖的含量,结果如表7所示。沉淀兼具葡萄糖异构酶和d-阿洛酮糖-3-差向异构酶活性。
[0153]
表7
[0154]
反应时间h葡萄糖浓度g/l果糖浓度g/l阿洛酮糖浓度g/l040.07——138.581.190.23235.863.480.66334.494.620.89432.676.091.24530.537.881.59628.779.341.89726.9710.782.25
[0155]
以上详细描述了本发明的优选实施方式,但是,本发明并不限于此。在本发明的技术构思范围内,可以对本发明的技术方案进行多种简单变型,包括各个技术特征以任何其它的合适方式进行组合,这些简单变型和组合同样应当视为本发明所公开的内容,均属于本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1