CNPY3蛋白作为治疗登革热靶点的用途
cnpy3蛋白作为治疗登革热靶点的用途
技术领域
1.本发明属于抗病毒药物技术领域,具体涉及cnpy3蛋白作为治疗登革热靶点的用途。
背景技术:
2.登革热是登革热病毒(dengue virus,dv)引起的一种急性传染病。每年约有0.5~1亿人感染登革病毒。登革热病毒感染可导致从登革热(df)到登革出血热(dhf)或登革休克综合征(dhs)等临床症状。dhf或dhs以严重出血和重要器官损伤为特征,病死率高,但其发病机制尚不清楚。由于登革热病毒感染的独特发病机制,目前还没有可以广泛用于预防或治疗登革热的疫苗和抗病毒药物。由赛诺菲巴斯德公司开发的dengvaxia(也被称为cyd-tdv)是唯一一种获准在亚洲、拉丁美洲、大洋洲和欧洲大约20个登革热流行国家使用的疫苗。这种疫苗适用于生活在登革热流行国家的9至45岁的人,该疫苗对1型和2型血清的保护低于3型和4型血清,并且最容易出现严重登革热相关症状的人群无法接种。因此基于登革疫苗开发受阻,针对登革病毒的靶点药物开发尤为重要。
3.从宿主-病原体相互作用的角度来看,大多数denv感染者的病毒血症在早期由先天免疫系统和适应性免疫系统控制。先天抗病毒免疫在抵御病毒病原体和建立适应性免疫反应中起着重要作用。模式识别受体(prr)是先天免疫中免疫受体的代表,可识别病原体相关分子模式(pamp)。当模式识别受体检测到病原相关分子时,这些检测信号就可激活各种转录因子,促进i型和iii型干扰素(ifn)等抗病毒蛋白的产生。toll样受体(toll-like receptor,tlrs)是prr的一个重要家族,调节先天免疫。tlrs能够识别多种pamps并聚合到mavs中,进而刺激irf3、nf-κb和i型ifn的产生。多项研究表明登革病毒进化后可以隐藏和掩盖其外来分子特征,并在多个水平上调节tlr信号通路,抑制抗病毒反应,从而促进病毒的复制和传播。我们发现cnpy3蛋白与登革病毒的发病机制和免疫逃逸有关,cnpy3蛋白与tlrs的成员结合作为伴侣帮助这些蛋白折叠和输出。此外,通过体外感染和小鼠模型,发现cnpy3具有抗登革病毒的功能,是denv感染过程中宿主先天免疫系统的关键调控因子,也是潜在的治疗靶点。
技术实现要素:
4.本发明要解决的技术问题是为登革热感染的治疗提供一种新选择。
5.本发明的技术方案是cnpy3蛋白作为治疗登革热靶点的用途。
6.本发明还提供了上调cnpy3蛋白的表达在抗登革热病毒中的用途。
7.具体的,所述cnpy3蛋白的氨基酸序列如seq id no.1所示。
8.进一步的,所述cnpy3蛋白的编码基因的核苷酸序列如seq id no.2或seq id no.45所示。
9.本发明还提供了抗登革热病毒的药物,其主要成分为cnpy3蛋白的mrna、cnpy3蛋白、表达cnpy3蛋白的载体或表达cnpy3蛋白的宿主细胞。
10.具体的,所述cnpy3蛋白的氨基酸序列如seq id no.1所示。
11.进一步的,所述cnpy3蛋白的编码基因的核苷酸序列如seq id no.2或seq id no.45所示。
12.其中,所述cnpy3蛋白的mrna的核苷酸序列如seq id no.2或seq id no.45所示。
13.进一步的,将cnpy3蛋白的mrna用lnp进行包裹。
14.具体的,将cnpy3蛋白的mrna克隆到t/a载体上,线性化后体外转录,对转录得到mrna进行加帽和加尾,采用lnp进行包裹。
15.seq id no.1:cnpy3蛋白的氨基酸序列
16.mdsmpepasrcllllplllllllllpapelgpsqagaeendwvrlpskcevckyvavelksafeetgktkevigtgygildqkasgvkytksdlrlievtetickrlldyslhkertgsnrfakgmsetfetlhnlvhkgvkvvmdipyelwnetsaevadlkkqcdvlveefeeviedwyrnhqeedlteflcanhvlkgkdtsclaeqwsgkkgdtaalggkkskkkssrakaaggrsssskqrkelgglegdpspeedegiqkasplthsppdel.
17.进一步的,所述cnpy3蛋白的编码基因的核苷酸序列如seq id no.2所示。
18.seq id no.2:cnpy3蛋白的编码基因的核苷酸序列
19.atggattcaatgcctgagcccgcgtcccgctgtcttctgcttcttcccttgctgctgctgctgctgctgctgctgccggccccggagctgggcccgagccaggccggagctgaggagaacgactgggttcgcctgcccagcaaatgcgaagtgtgtaaatatgttgctgtggagctgaagtcagcctttgaggaaaccggcaagaccaaggaggtgattggcacgggctatggcatcctggaccagaaggcctctggagtcaaatacaccaagtcggacttgcggttaatcgaagtcactgagaccatttgcaagaggctcctggattatagcctgcacaaggagaggaccggcagcaatcgatttgccaagggcatgtcagagacctttgagacattacacaacctggtacacaaaggggtcaaggtggtgatggacatcccctatgagctgtggaacgagacttctgcagaggtggctgacctcaagaagcagtgtgatgtgctggtggaagagtttgaggaggtgatcgaggactggtacaggaaccaccaggaggaagacctgactgaattcctctgcgccaaccacgtgctgaagggaaaagacaccagttgcctggcagagcagtggtccggcaagaagggagacacagctgccctgggagggaagaagtccaagaagaagagcagcagggccaaggcagcaggcggcaggagtagcagcagcaaacaaaggaaggagctgggtggccttgagggagaccccagccccgaggaggatgagggcatccagaaggcatcccctctcacacacagcccccctgatgagctctga
20.本发明的有益效果:为了识别与denv发病机制有关的基因,并发现一些新的治疗靶点,本发明使用公开的数据集分析了与登革病毒感染有关的基因并鉴定了一个重要的宿主蛋白cnpy3。该宿主基因在登革病人血液以及在登革病毒感染的树突状细胞和thp-1细胞中显著下调。其表达与登革疾病进展呈负相关,与大多数toll样受体表达呈正相关。进一步研究表明在thp-1细胞中下调该基因抑制了ifn-β的产生和isgs基因的表达,促进denv-2的感染。而在vero和hek 293t细胞中上调该基因的表达,则可以抑制denv-2的感染。这些结果说明cnpy3参与了固有免疫应答信号途径,具有抗登革病毒的作用,是登革热潜在的治疗靶点。
附图说明
21.图1、登革病毒感染人单核细胞来源的树突状细胞和登革患者的转录组分析。a:维恩图表示两个数据集共有的差异基因的交集。b:差异基因功能富集分析。热图代表了排在前的基因本体(go)生物过程的热图。离散色标表示统计意义。c:热图显示了在本研究中使
用的两个数据集中表达一致的基因。d:转录组分析差异基因在denv-2感染thp-1后表达验证。结果以标准差表示(n=3)。星号表示显著差异(t-test,*,p《0.05;**,p《0.01;***,p《0.001);relative expression:相对表达量,mock:空白;denv-1、denv-2、denv-3、denv-4:分别表示1、2、3、4型登革病毒。
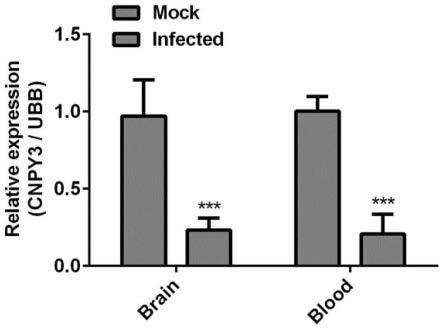
22.图2、denv-2感染乳鼠后cnpy3的表达情况。3日龄balb/c乳鼠组用3μldenv-2病毒鼻内接种感染(病毒滴度8
×
106pfu/ml),mock组用3μl pbs鼻内感染。感染后第7天取小鼠脑和全血提取rna进行检测。mock:未感染denv-2病毒的对照组;infected:denv-2病毒感染7d的组织。结果以标准差表示(n=3)。星号表示显著差异(t-test,*,p《0.05;**,p《0.01;***,p《0.001)。infected:感染,brain:脑,blood:血液。
23.图3、cnpy3表达与denv疾病严重程度呈负相关。a:cnpy3在denv-2感染乳鼠后2d和4d的表达情况。infected:用20μl(60pfu)病毒颅内注射3日龄balb/c乳鼠组;mock:用20μl pbs颅内注射。感染后2d和4d取乳鼠全血测定cnpy3的表达情况。结果以标准差表示(n=3)。b:cnpy3在登革病毒感染人单核细胞来源的树突状细胞不同时相点的表达情况。原始数据来源于公开数据集gse58278。uninfected为对照组。c和d:cnpy3不同疾病进展的登革病人中的表达情况。原始数据来源于公开数据集gse18090和gse51808。星号表示显著差异(t-test,*,p《0.05;**,p《0.01;***,p《0.001)。df:登革热,dhf:登革出血热,convalesent:康复者。
24.图4、利用hpa数据库分析cnpy3在人体组织和血液细胞中的表达特异性。a:cnpy3在人体各组织中rna和蛋白表达水平分析。b:cnpy3在血液细胞中的表达分析。c:血液中单细胞水平上cnpy3的表达。单细胞数据包括来自不同外周血单核细胞类型(pbmc)和内皮细胞簇的scrna-seq数据。plasmacytoid dc:浆细胞样树突状细胞;neutrophil:中性粒细胞;classical monocyte:经典单核细胞;myeloid dc:髓系树突状细胞;intermediate monocyte:中间型单核细胞;non-classical monocyte;memory b-cell:记忆b细胞;naive b-cell幼稚b细胞;eosinophil:嗜酸性粒细胞;total pbmc:总外周血单核细胞;basophil:嗜碱性粒细胞;nk-cell:自然杀伤细胞;memory cd8 t-cell:记忆性cd8 t细胞;naive cd8 t-cell:幼稚cd8 t细胞t-reg调节性t细胞;dendritic cells c-11:树突状细胞c-11;endothelial cells c-11:内皮细胞c-11;monocytes c-0:单核细胞c-0;monocytes c-7:单核细胞c-7;monocytes c-9:单核细胞c-9;b-cells c-8:b细胞c-8;nk-cells c-5:自然杀伤细胞c-5。
25.图5、健康人血液中cnpy3与toll样受体表达相关性分析。tlr1、tlr2、tlr3、tlr4、tlr5、tlr6、tlr7、tlr8、tlr9和tlr10与cnpy3表达相关性分析。
26.图6、toll样受体在登革病人中的表达分析;control:对照组。
27.图7、thp-1细胞中下调cnpy3抑制ifn-β的产生和干扰素刺激基因(isgs)的表达。a:cnpy3的mrna和(b)蛋白表达水平分析。使用靶向cnpy3的sirna或阴性对照sirna转染thp-1细胞,36h后收集细胞测定cnpy3的相对表达量和蛋白含量。c:ifn-β的含量测定。d:干扰素刺激基因(isgs)的表达量分析。β-actin作为内参基因。nc:阴性对照sirna转染的thp-1细胞,sirna:靶向cnpy3的sirna转染thp-1细胞。使用nc-sirna中cnpy3的表达量为对照,结果以标准差表示(n=3),学生t检验,*,p《0.05;***,p《0.001。mx1、oas1、oas2、isg15、irf7、usp18均为基因名。
28.图8、下调cnpy3促进登革病毒的复制。a:持续培养18h后检测denv-2的包膜(e)蛋白、非结构蛋白ns3、ns5的相对表达量。b:持续培养2d、4d、6d检测细胞上清中的病毒拷贝数。c:培养6d检测细胞上清中e蛋白含量。β-actin作为内参基因。nc:阴性对照sirna转染的thp-1细胞,sirna:靶向cnpy3的sirna转染thp-1细胞。结果以标准差表示(n=3),t-test,*,p《0.05。
29.图9、上调cnpy3抑制登革病毒的复制。a:荧光显微镜观察转染pcdna3.1(载体对照)或pcdna3.1-cnpy3后denv-2感染vero细胞的数量。将这两种质粒以相同浓度转染vero细胞。36h后用denv-2(moi=2)孵育2h,弃去病毒液,持续培养18h,对标本进行固定、渗透、染色和成像。dapi:4',6-二脒基-2-苯基吲哚,anti-dengue:抗登革抗体;uninfected cells:未感染细胞。b:denv-2感染vero细胞数量的定量分析。使用image j软件在8个不同区域对每个实验条件的感染细胞进行计数。c:e蛋白、ns3和ns5在denv-2感染vero细胞中的相对表达量分析,β-actin作为内参基因。d和e:cnpy3 mrna和蛋白表达水平分析。用pcdna3.1(载体对照)或pcdna3.1-cnpy3转染hek 293t细胞。36h后,采用rt-pcr和western blot检测cnpy3的表达。f:e蛋白、ns3和ns5在denv-2感染hek 293t细胞中的相对表达量分析,β-actin作为内参基因。g:过表达cnpy3后denv-2感染hek 293t细胞6d后上清的拷贝数检测。h:免疫荧光观察病毒感染vero细胞的情况。pcdna3.1(载体对照)或pcdna3.1-cnpy3转染hek 293t细胞36h,denv-2(moi=2)孵育2h,弃去病毒液,持续培养6d后,收集细胞上清中的病毒,用以感染vero细胞,18h后进行免疫荧光观察。结果以标准差表示(n=3),t-test,*,p《0.05;**,p《0.01;***,p《0.001。
30.图10、生存率统计。取10μl denv-2(60pfu)与10μl的空载lnp和cnpy3制剂混匀后同时打颅内注射3日龄乳鼠,每日统计死亡率。days,天数。percent survival:存活百分比。
具体实施方式
31.下面结合附图对本发明进行说明。
32.实施例1denv病毒感染过程中宿主差异基因的筛选
33.下载geo公共数据库的数据集gse58278 denv感染树突状细胞的芯片数据集(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse58278),利用ncbi在线差异表达分析工具geo2r对该数据集denv 2感染细胞后24h同未感染的细胞的差异基因进行分析,筛选出p≤0.05,log2fc≥1的差异基因,其中387个基因上调,295个基因下调。
34.另联合登革病人转录组数据集gse51808(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi),以9个健康人的全血作为对照组,10个登革出血热患者的全血为感染组,利用ncbi在线差异表达分析工具geo2r筛选出p≤0.05,log2fc≥1的差异基因。健康人样本编号为gsm1253075~gsm1253083,出血热患者样本编号为gsm1253032、gsm1253034、gsm1253037、gsm1253039、gsm1253040 gsm1253041、gsm1253046、gsm1253048、gsm1253049、gsm1253052。最终获得1528个上调基因,1184个下调基因。对denv感染树突状细胞的芯片数据集与登革病人转录组数据集的共同差异基因进行维恩分析,结果显示,共有46个共同上调的基因,32个共同下调的基因(图1a)。共有的degs生物学功能富集分析结果表明,上调的基因如usp18、mix1、osa1、osa2、isg20等主要集中在ⅰ型干扰素信号通路,众多报道已证明ⅰ型干扰素通路是机体清除病原体的主要天然免疫信号通路,登革病毒感染后ⅰ型干扰素信
号通路相关基因的上调有助于登革病毒的清除(图1b和c)。值得注意的是,fcer1a、ifngr1、cd300lf、clec7a、clec4a、cnpy3、tlr5、tgm2这些在细胞水平和人体水平共同下调的基因参与了正向调节防御反应的生物学功能途径,因此这些基因的下调有可能降低机体对登革病毒感染的防御(图1b和c)。
35.进一步通过实时定量pcr(rt-qpcr)验证了3个参与干扰素信号通路的上调基因(ifi27、mx1和oas1)和3个参与正调控防御反应的下调基因(tlr5、cnpy3和fcer1a)在denv感染thp-1单核细胞细胞系中的表达水平。在thp-1细胞感染denv1 denv-2,denv3和denv4 24h后检测这些基因的相对表达量。结果显示与未感染denv的对照组相比,ifi27、mx1和oas1在denv感染后显著上调,tlr5、cnpy3和fcer1a显著下调(图1d)。这些结果与数据集分析结果一致,验证了denv感染过程中的差异表达基因。
36.实施例2cnpy3表达与denv疾病严重程度呈负相关
37.在实施例1分析得到的这些基因中,cnpy3在denv感染中的作用的研究很少。因此,采用乳鼠denv-2感染模型进行了进一步研究。
38.本发明使用的denv-1(毒株名称thd1-0102-01)、denv-2(毒株名称new guinea)、denv-3(毒株名称80-2)和denv-4(毒株名称gd07-78)病毒株由中国广州军区疾病预防控制中心提供。denv-1、denv-2、denv-3和denv-4在vero细胞中繁殖。
39.登革病毒感染乳鼠实验:使用new guinea denv 2颅内注射3日龄bablc乳鼠(雌性balb/c孕鼠购买于陆军军医大学实验动物中心),从乳鼠脑部分离denv-2病毒(病毒滴度8
×
106pfu/ml),取3μl对3日龄乳鼠进行滴鼻,感染后7日取小鼠的脑和血液提取rna,对照组取3μl健康小鼠的脑部组织上清液对3日乳鼠滴鼻,并在7日后取同样的部位提取rna。收集的细胞和乳鼠组织的rna提取使用组织/细胞快速提取试剂盒(boer,china),严格按照说明书进行提取。随后使用primescripttm rt reagent kit(takara,japan)进行逆转录。使用tbpremix ex taq
tm
ii(takara,japan)和96system(roche,usa)进行荧光定量pcr检测。扩增反应体系如表1所示。
40.表1荧光定量pcr扩增反应体系
41.名称体积tb green premix ex taq ii(tli rnaseh plus)(2
×
)5μl上游引物(10μm)0.3μl下游引物(10μm)0.3μl模板1μl无核酸酶的ddh2oup to 10μl
42.反应条件:step1:预变性95℃30s;变性95℃5s;0个循环;60℃30s,step 3,溶解曲线反应条件:逐渐升温至95℃。pcr反应结束后,检查溶解曲线与扩增曲线是否异常,导出实验数据,目的基因的表达水平使用-δδct方法计算。目的基因检测所使用的引物见表2。
43.表2实施例中所用相关引物
[0044][0045][0046]
rt-qpcr结果显示,在denv-2感染后7d,脑和血液中cnpy3的表达受到抑制(图2)。
[0047]
进一步对乳鼠血液中cnpy3的表达情况进行检测发现,随着denv-2感染的时间延长,cnpy3在血液中的表达受到抑制的情况更为明显(图3a)。同样的在数据集gse58278中(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse58278)),cnpy3的表达随着感染时间的延长而下调,尤其是在12、18、24h(图3b)。因此,推测cnpy3同denv感染后疾病严重程度相关。denv介导的疾病严重程度从无症状感染和轻度登革热(df)到登革出血热(dhf)和休克综合征(dss)。利用公开可用的转录组数据集gse18090(https://
www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse18090),评估了cnpy3在对照组(发热患者确认未感染登革热)、轻度登革热(df)和登革出血热(dhf)患者外周血单个核细胞中的表达。数据显示cnpy3表达在重症登革患者的外周血单个核细胞的表达明显下调。进一步利用gse51808数据集,分析了28例登革热患者(df n=18,dhf n=10)、19例恢复期患者(df n=13,dhf n=6)和9例健康、非感染献血者的全血cnpy3表达。数据显示cnpy3表达同登革热患者疾病严重程度成负相关(图3c和d)。这些数据说明cnpy3在denv疾病进展中起重要作用。
[0048]
实施例3cnpy3在toll样受体依赖的免疫应答中发挥重要作用
[0049]
denv对细胞和组织的亲嗜性影响了denv感染的结果为了探讨cnpy3在denv感染中的作用,使用hpa数据库(https://www.proteinatlas.org/)研究了cnpy3在健康人体组织中的表达。数据表明cnpy3rna和蛋白在脑、血、骨髓和淋巴组织中表达更丰富(图4a)。免疫系统、肝脏和血液内皮细胞内层器官系统在dhf/dss发病机制中发挥重要作用。进一步使用hpa数据库检测血液中各种免疫细胞中cnpy3的表达。
[0050]
从hpa数据库获得的单细胞测序数据显示,cnpy3在血液中的树突状细胞、内皮细胞和单核细胞中表达最高(图4b和4c)。单核细胞和树突状细胞(dendritic cells,dcs)是denv的主要靶细胞。cnpy3在这些器官和细胞中的高表达说明其在抗登革病毒免疫系统中具有重要作用。
[0051]
tlrs(toll like receptors)是宿主防御的关键蛋白,参与宿主对微生物入侵的固有免疫反应。cnpy3是除tlr3外,gp96正常折叠和表达tlrs所必需的。这说明cnpy3可能通过固有免疫应答信号通路抵御登革病毒感染。利用gepia数据库(http://gepia.cancer-pku.cn/)探讨cnpy3表达与tlrs表达的相关性。gepia数据库获得,在健康人全血中,除tlr3外,tlr1、tlr2、tlr4、tlr5、tlr6、tlr7、tlr8、tlr9、tlr10等其他tlr的表达均与cnpy3的表达呈正相关(图5)。
[0052]
此外,基于gse51808数据集,除tlr3和tlr7外,大多数tlrs表达与登革热病情严重程度呈负相关(图6),这个结果同cnpy3在登革病人的表达情况一致。
[0053]
tlrs信号途径促进i型干扰素、炎症因子和趋化因子的产生,并建立抗病毒免疫(6)。为进一步探索cnpy3在抗病毒免疫应答中发挥的作用,使用sirna抑制thp-1细胞中cnpy3的表达,并检测cnpy3下调后,固有免疫信号通路下游抗病毒感染的i型干扰素的含量以及抗病毒相关分子的表达。靶向cnpy3的sirna瞬时转染thp-1细胞采用zeta life advanced dna rna转染试剂(zeta life,usa),按照试剂盒说明书进行。将thp-1细胞用含血清的完全培养基重悬并计数,以3
×
105/ml接种于6孔细胞培养板,每孔接种2ml。sirna(由生工生物工程(上海)股份有限公司合成,序列:sense(seq id no.41)5
’‑3’
gagcuguggaacgagacuucuutt,antisense(seq id no.42)5
’‑3’
agaagucucguuccacagcuctt)与转染试剂的体积为1︰1,将合成的nc和sicnpy3用无酶水溶解至终浓度为20μm,将8μl的nc/sicnpy3与8μl的转染试剂轻轻混匀,室温孵育15min后加入细胞培养板,转染36h后收集细胞进行检测。
[0054]
结果显示,利用靶向cnpy3的sirna可明显降低thp-1细胞中cnpy3 mrna和蛋白的表达水平(图7a和7b)。利用human ifn-βelisakit试剂盒(联科生物,中国)检测cnpy3sirna转染的thp-1细胞上清中i型干扰素ifn-β的含量,发现cnpy3下调导致i型干扰
素ifn-β的产生受到抑制(图7c),相关的干扰素刺激基因(isgs)包括mx1、oas1、oas2、isg15、irf7和usp18下调(图7d)。因此cnpy3下调会影响机体的抗病毒免疫响应。
[0055]
实施例4cnpy3抑制denv-2的复制
[0056]
考虑到cnpy3敲除对toll样受体依赖免疫应答的影响,推测cnpy3敲除可能增加denv感染。为了验证这一假设,用靶向cnpy3的sirna转染thp-1细胞,36小时后用denv-2感染这些细胞。18h后检测thp-1细胞denv-2包膜(e)蛋白、非结构蛋白ns3、ns5的表达。正如预期的那样,相对于对照组,cnpy3的蛋白的下调增强了denv-2感染,denv-2包膜蛋白与非结构蛋白ns3、ns5基因在cnpy3下调的thp-1细胞中的表达均显著高于对照组(图8a)。随后,检测denv-2感染2d、4d、6d后细胞上清中病毒含量也表明,denv-2在cnpy3下调的thp-1细胞中病毒拷贝数明显高于对照组(图8b),western blot的结果同样显示denv-2包膜蛋白在denv-2感染6d的cnpy3干扰的thp-1细胞上清中更多(图8c)。
[0057]
cnpy3在denv感染过程中表达下调,提示denv可能通过抑制cnpy3的表达,抑制toll-like固有免疫系统下游抗病毒信号通路,从而逃过机体固有免疫系统的监视以利于病毒的复制。推测提高cnpy3的表达会抑制dnev病毒的复制。因此,构建空载对照(pcdna3.1)或cnpy3过表达质粒(pcdna3.1-cnpy3)。然后进行瞬时转染,采用3000试剂(invitrogen,usa)。按照试剂盒说明书进行。将细胞以3
×
105/ml接种于6孔细胞培养板,每孔接种2ml,置于含体积分数5%co2的37℃培养箱培养过夜,待细胞密度为70%时进行转染。将2μg cnpy3/pcdna3.1质粒用125μl无血清的高糖dmem培养基(含5μl p3000 reagent)稀释,用同样体积的无血清的高糖dmem培养基稀释7.5μl lipofectamine 3000reagent转染试剂。将稀释后的质粒加入转染试剂中,轻轻混匀室温放置15min。将细胞培养板中的完全培养基换成无血清的dmem高糖培养基,然后将混合液加入到细胞培养板中。24h后更换为新鲜完全培养基,转染36h后收集细胞进行检测。
[0058]
瞬时转染vero细胞36小时,然后用denv-2(moi=2)感染细胞,18小时后用免疫荧光检测病毒含量。具体实施方法如下:vero细胞接种于24孔组织培养板上,细胞密度50%时转染pcdna3.1-cnpy3或pcdna3.1空载质粒。转染36h后的vero细胞用denv-2(moi=5)感染18小时。随后,用4%多聚甲醛溶液室温固定细胞15min,再使用0.5%triton x-100溶液(预温到37℃)处理细胞10min。pbs洗涤2次后加入300μl 10%山羊血清进行封闭,室温孵育30min后用滤纸吸去封闭液,滴加200μl(1:100稀释)抗dengue virus 1+2+3+4antibody(abcam,england,ab26837),放入湿盒,4℃孵育过夜。pbst洗涤2次后用滤纸吸干水分,滴加200μl(1:500稀释)alexa488标记羊抗兔igg(h+l)二抗(abcam,england,ab150077),37℃孵育1h。随后用pbst洗涤2次后用滤纸吸干水分,滴加200μl dapi(碧云天,中国),室温孵育5min,用pbs洗涤2次后在荧光显微镜(奥林巴斯ix53,德国)下观察并拍照。使用image j(nih)软件统计特定区域denv感染细胞的数量。利用荧光显微镜观察denv-2感染vero细胞的总百分比,实时荧光定量pcr检测denv-2包膜(e)蛋白、非结构蛋白ns3和ns5的表达。结果显示,与空载对照相比,过表达cnpy3蛋白的vero细胞中denv-2感染受到显著抑制(图9a、9b和9c)。由于vero细胞为干扰素缺陷型细胞,进一步在hek293t细胞中验证了cnpy3的抗登革病毒的作用。与vero细胞数据一致,在hek 293t细胞中过表达cnpy3蛋白同样导致denv-2感染显著降低(图9d、9e和9f)。
[0059]
pcdna3.1-cnpy3或pcdna3.1空载质粒瞬时转染hek 293t细胞,或靶向cnpy3的
sirna瞬时转染thp-1细胞36h后,以denv-2(moi=2)进行感染,感染2d,4d和6d后,收集200μl的细胞培养上清,使用minibest viral rna/dna extraction kit(takara,japan)提取病毒rna。rna用30μl无rna酶的水洗脱。随后取7μl rna使用primescripttm rt reagent kit(takara,japan)进行逆转录,总共获得20ul cdna。取1ul cdna为模板进行定量pcr。定量pcr引物为dv2-f:5
’‑
gcagaatgccccaacacaaa-3’(seq id no.43)和dv2-r:5
’‑
acaaaatacatcctgcctttctct-3’(seq id no.44)。pcr扩增程序如下:预变性95℃3min;变性95℃10s;退火57℃30s;延伸72℃30s;总共42个循环。利用含denv-2ns1基因的puc57阳性质粒(上海金斯瑞公司合成)进行rt-pcr建立标准曲线,参考lee et al.(2006)的方法进行denv-2拷贝数换算。根据标准曲线从定量pcr cq值计算样本每200ul病毒的拷贝数。结果如如图9g所示,在293t细胞超量表达cnpy3后,病毒含量较空载对照明显降低。
[0060]
在hek 293t细胞中过表达cnpy3蛋白后,收集感染denv-2(moi=2)6d后的细胞上清用免疫荧光检测病毒含量。结果如图9h所示,pcdna3.1-cnpy3转染的vero细胞中的病毒数量明显低于pcdna3.1空载质粒转染的vero细胞,说明cnpy3具有明显的抗病毒功能。
[0061]
实施例5动物实验
[0062]
(1)制备cnpy3 mrna:合成puc57-mcnpy3质粒(上海,金斯瑞公司合成)。mcnpy3序列如下:
[0063]
seq id no.45:
[0064]
gcagtcccggaagcggccgggggaagctgctccgcgcgcgctgccggaggaagcgccgccgggtccgctctgctctgggtccggctgggccgccaccatggagtccatgtctgagctcgcgccccgctgcctcttatttcctttgctgctgctgcttccgctgctgctccttcctgccccgaagctaggcccgagtcccgccggggctgaggagaccgactgggtgcgattgcccagcaaatgcgaagtgtgcaagtatgttgctgtggagctgaagtcggcttttgaggaaacgggaaagaccaaggaagtgattgacaccggctatggcatcctggacgggaagggctctggagtcaagtacaccaagtcggacttacggttaattgaagtcactgagaccatttgcaagaggcttctggactacagcctgcacaaggagaggactggcagcaaccggtttgccaagggtatgtcggagacctttgagacgctgcacaacctagtccacaaaggggtcaaggtggtgatggatatcccctatgagctgtggaacgagacctcagcagaggtggctgacctcaagaagcagtgtgacgtgctggtggaagagtttgaagaggtgattgaggactggtacaggaaccaccaggaggaagacctgactgaattcctctgtgccaaccacgtgctgaagggaaaggacacgagttgcctagcagagcggtggtctggcaagaagggggacatagcctccctgggagggaagaaatccaagaagaagcgcagcggagtcaagggctcctccagtggcagcagcaagcagaggaaggaactggggggcctgggggaggatgccaacgccgaggaggaggagggtgtgcagaaggcatcgcccctcccacacagcccccctgatgagctgtgattgtgtatgcgttaataaaaagaaggaactcgta
[0065]
使用sal酶(takara,日本)进行线性化质粒,随后使用近岸公司体外转录试剂盒(t7 high yield rna transcription kit,货号:e131-01a)进行体外转录,再使用近岸公司加帽试剂盒(mrna cap 2
′‑
o-methyltransferase,货号:m072)加尾试剂盒(e.coli poly(a)polymerase,货号:m012)进行加帽和加尾。随后使用lnp(lipid nanoparticle,脂质纳米粒)进行包裹。
[0066]
(2)颅内攻毒时取10μl denv-2(60pfu)分别与10μl的空lnp和1μg mcnpy3 lnp制剂混匀后同时打入5只或6只3日龄乳鼠。然后统计每日存活率。结果表明,cnpy3对感染denv-2的乳鼠具有保护效率(图10)。
[0067]
以上所有结果表明,denv通过抑制cnpy3的表达,进而扰乱toll样受体依赖的免疫
应答,促进denv的侵袭和传播;cnpy3具有抑制denv复制的作用。在小鼠体内mcnpy3做成mrna制剂与病毒同时进行乳鼠颅内攻毒,对乳鼠具有保护效果。
[0068]
本发明使用公开的数据集分析了与登革病毒感染有关的基因并鉴定了一个重要的宿主蛋白cnpy3。该宿主基因在登革病人血液以及在登革病毒感染的树突状细胞和thp-1细胞中显著下调。其表达与登革疾病进展呈负相关,与大多数toll样受体表达呈正相关。进一步研究表明在thp-1细胞中下调该基因抑制了ifn-β的产生和isgs基因的表达,促进denv-2的感染。而在vero和hek 293t细胞中上调该基因的表达,则可以抑制denv-2的感染。这些结果说明cnpy3参与了固有免疫应答信号途径,具有抗登革病毒的作用,是登革热潜在的治疗靶点。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1